

Abstract
Background/Aim: Bufalin, bufadienolide present in Chan Su, has been shown to induce cancer cell apoptosis in many human cancer cells, including human leukemia cells, but its effects on immune responses are unknown. Materials and Methods: This study investigated whether bufalin affected immune responses of mice with WEHI-3 cell-generated leukemia in vivo. BALB/c mice were intraperitoneally injected with WEHI-3 cells to develop leukemia and then were treated with oral treatment with bufalin at different doses (0, 0.1, 0.2 and 0.4 mg/kg) for 2 weeks. At the end of treatment, all mice were weighted and blood was collected; liver and spleen tissues were collected for cell marker, phagocytosis, natural killer (NK) cell activity and T- and B-cell proliferation measurements by using flow cytometric assays. Results: When compared with the leukemia control group, bufalin increased the body weight, but reduced liver and spleen weights, and reduced CD3, CD16 and Mac-3 cell markers at 0.4 mg/kg treatment and increased CD11b marker at 0.1 and 0.2 mg/kg treatment. Furthermore, bufalin at 0.4 mg/kg increased phagocytosis by macrophages isolated from peripheral blood mononuclear cells and at 0.1 mg/kg by those from the peritoneal cavity. Bufalin (0.2 and 0.4 mg/kg) increased NK cell cytotoxic activity at effector:target ratio of 50:1. Bufalin increased B-cell proliferation at 0.1 and 0.2 mg/kg treatment but only increased T-cell proliferation at 0.1 mg/kg. Bufalin increased glutamate oxaloacetate transaminase level at all dose treatments, increased glutamic pyruvic transaminase level only at 0.1 mg/kg treatment, but reduced the level of lactate dehydrogenase at all dose levels in mice with WEHI-3 cell-induced leukemia in vivo. Conclusion: Bufalin increased immune responses by enhancing phagocytosis in mice with leukemia mice.
Keywords: Bufalin, leukemia WEHI-3 cells, macrophage, natural killer cells, phagocytosis
Leukemia is a kind of malignant disease arising from unregulated proliferation of immature blood cells (1). It includes chronic and acute myelomonocytic leukemia (MML) (2). In Western countries, chronic MML remains the most common type of leukemia in adults (3) and is a clonal hematopoietic malignancy which includes two features, myelodysplastic syndrome and myeloproliferative neoplasm (4) which are recognized by the World Health Organization classification of 2008 (5). In about 15%-30% of patients, chronic MML evolves into acute myeloid leukemia (AML) (5,6). AML is also the most common pediatric malignancy (7,8). Currently, chemotherapy is still the main curative treatment in patients with leukemia but it has relatively low efficacy and high toxicity (9). Therefore, numerous studies are focused on the identification and development of novel agents to treat this disease.
Bufalin, a digoxin-like immunoreactive component, is a bufadienolide present in Chan Su and is used as Chinese medicine (10). Bufalin has biological activities including inducing cancer cell apoptosis (11-17) and DNA damage in NCI-H460 human lung cancer cells (18). Recently, we also found that bufalin significantly inhibits cell adhesion, migration and invasion of gefitinib-resistant NCI-H460 cells (19) and induced apoptosis of U-2 OS human osteosarcoma cells (20) and SCC 4 human tongue cancer cells (21).
Although several studies have demonstrated bufalin-induced cytotoxic effects in vitro through the induction of apoptosis in many human cancer cells and also have antitumor activity in vivo, there is no report to show how it affects immune response. Therefore, we investigated the immune response-modulating potential of bufalin on a in vivo model of leukemia.
Materials and Methods
Materials and reagents. Bufalin and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich Corp. (St. Louis, MO, USA). IMDM and RPMI-1640 culture medium, fetal bovine serum (FBS), L-glutamine and penicillin-streptomycin were obtained from Gibco Life Technologies (Carlsbad, CA, USA). Antibodies against CD3 (T-cell marker), CD19 (B-cell marker), CD11b (monocyte marker) and Mac-3 (macrophage marker) were obtained from BD Biosciences Pharmingen Inc. (San Diego, CA, USA). Bufalin was dissolved in DMSO at 1% stock solution and kept at −20˚C in a 50 ml shielded from light before use.
Cell culture. Murine myelomonocytic leukemia cells (WEHI-3 cells) and Yac-1 were purchased from the Food Industry Research and Development Institute (Hsinchu, Taiwan, R.O.C.). About 1×106 cells/ml were maintained in 75 cm2 tissue culture flasks with IMDM or RPMI-1640 medium supplemented with 10% fetal bovine serum, 100 units/ml penicillin, 100 μg/ml streptomycin and 2 mM L-glutamine at 37˚C in 100% humidity, with 5% CO2 as previously described (15).
Male BALB/c mice and bufalin treatment. Forty male BALB/c mice, aged 5 weeks, weighing approximately 22-25 g each, were purchased from the National Laboratory Animal Center (Taipei, Taiwan, R.O.C.). All mice were kept at 25˚C on a 12 h light/dark cycle at the Animal Center of China Medical University (Taichung, Taiwan, R.O.C.). The animal study projects were reviewed and approved by the Institutional Animal Care and Use Committee of China Medical University (approval ID: 104-10-B) and all animals were cared for following the institutional animal ethical guidelines of the China Medical University as described previously (22). The mice were randomly divided into five groups (n=8). Group I mice received a normal diet (Prolab RMH 2500; LabDiet, PMI Nutrition, Brentwood, MO, USA) as control. For the other four groups, the mice were intraperitoneally injected with 1×106 WEHI-3 leukemia cells to generate leukemic mice as described previously (22). Group II mice received a normal diet and treatment with olive oil as leukemia control (positive control). Groups III-V mice received a normal diet and treatment with bufalin (0.1, 0.2 or 0.4 mg/kg, respectively) in olive oil. Bufalin in olive oil was given to mice by oral gavage every 2 days for 14 days. All mice were weighed during the oral treatment every 2 days. At the end of treatment, all mice were weighed and blood was collected before mice were then sacrificed. After sacrifice, liver and spleen were isolated and weighed (22).
Immunofluorescence staining for surface markers in isolated white blood cells. After treatment, blood was collected individually from all mice from each group as described previously (22). Cell markers from isolated leukocytes were measured. A sample of 200 μl blood from each animal was lysed to destroy the red blood cells with 1×Pharm Lyse™ lysing buffer (BD Biosciences) as per guidelines from BD Biosciences and leukocytes were collected. Isolated leukocytes were individually stained by phycoerythrin (PE)-labeled anti-mouse CD3, PE-labeled anti-mouse CD19, fluorescein isothiocyanate (FITC)-labeled anti-mouse CD11b and FITC-labeled anti-mouse Mac-3 for 1 h and washed with phosphate-buffered saline (PBS) three times, and then analyzed by flow cytometry to determine the percentage of cell markers, as previously described (22).
Measurement of macrophage phagocytosis. Macrophages were isolated from peripheral blood mononuclear cells (PBMCs) and the peritoneum of each animal as described previously (22). FITC-conjugated Escherichia coli as target cells (50 μl) were added to the macrophages from mice under each treatment according to PHAGOTEST® kit manufacturer’s instructions (ORPEGEN Pharma Gesellschaft für biotechnologische, Heidelberg, Germany) and mixed well. All samples were analyzed for phagocytosis using flow cytometry and quantified by the CellQuest software (Becton Dickinson), as described previously (22).
Measurement of NK cell cytotoxic activity. Splenocytes isolated from mice were placed in 96-well plate (2.5-5×105 cells/well) with 100 μl of RPMI 1640 medium. Yac-1 cells (1×104 cells) and PKH-67/Dil.C buffer (Sigma-Aldrich Corp.) were added to the cells and mixed thoroughly for 2 min at 25˚C then 2 ml PBS was added to well for 1 min. Next, 4 ml medium was added and cells were incubated for 10 min. After incubation, all cells were centrifuged at 290 × g for 2 min and then NK cell cytotoxic activity was measured by flow cytometry as described elsewhere (22).
Measurement of B-and T-cell proliferation. Splenocytes (1×105 cells/well) in 100 μl of RPMI-1640 medium were placed in 96-well plate. Lipopolysaccharide (LPS, 5 μg/ml) (Sigma Chemical Co., St. Louis, MO, USA) was added to the splenocytes to stimulate for 3 days for measuring B-cell proliferation. Concanavalin A (Con A, 5 μg/ml) (Sigma Chemical Co.) was added to the splenocytes to stimulate them for 5 days for measuring T-cell proliferation. After stimulation, all samples were measured by using CellTiter 96 AQueous One Solution Cell Proliferation Assay kit (Promega, Madison, WI, USA) as previously described (22).
Biochemical measurement of blood glutamic oxaloacetic transaminase (GOT), glutamic pyruvic transaminase (GPT), lactate dehydrogenase (LDH) of BALB/c mice after exposure to bufalin. Isolated blood samples from each animal were assayed for levels of GOT, GPT and LDH using quantification kit as described previously (22). The liquiUV Test (Aspartate Aminotransferase) for GOT, liquiUV Test (Alanine Aminotransferase) for GPT and liquiUV Test (Lactate Dehydrogenase) for LDH were obtained from Human Gesellschaft fur Biochemica und Diagnosica mbH (Wesbaden, Germany).
Statistical analysis. All values are expressed as mean±standard error (S.D.). Comparisons between groups were analyzed by one-way analysis of variance and Tukey test for multiple comparisons (SigmaPlot for Windows version 12.0; Systat Software, Inc., San Jose, CA, USA). A value of p<0.05 was considered to indicate a statistically significant difference.
Results
Bufalin affected the weights of body, liver and spleen from leukemic BALB/c mice. After treatment, blood, liver and spleen were collected from mice; results are presented in Figure 1. The leukemia control group had significantly lower body weight compared with normal control group, while bufalin at 0.4 mg/kg significantly increased body weight compared with the leukemic control group. Representative images of liver and spleen tissues and their weights are shown in Figure 1C and D. These results indicated that bufalin significantly reduced liver (Figure 1C) and spleen (Figure 1D) weights at all doses of bufalin treatment when compared with the leukemic control group.
Figure 1. Bufalin affect the body, liver and spleen weights of leukemia BALB/c mice. Group I was untreated as normal control (NC). Remainingmice were intraperitoneally injected with WEHI-3 cells then were divided into four groups: Group II was treated with normal diet (WC); group IIIwas treated with 0.1 mg/kg of bufalin; group IV was treated with 0.2 mg/kg of bufalin; group V was treated with 0.4 mg/kg of bufalin. Body weights(A), animal appearance (B), liver (C), and spleen (D) weights are presented. The body weights were measured every 2 days. Significantly differentat p<0.05 vs. *normal control group, vs. #leukemia control group.

Bufalin affected cells markers of white blood cells from leukemic BALB/c mice. Blood samples were collected for measuring the levels of cell markers CD3, CD19, CD11b and Mac-3. Results indicated that bufalin reduced CD3 (T-cells) (Figure 2A), CD19 (B-cells) (Figure 2B) and Mac-3 (macrophages) (Figure 2D) at 0.4 mg/kg treatment, however, bufalin promoted CD11b (monocytes) at 0.1 and 0.2 mg/kg treatment (Figure 2C) when compared with the leukemic control group.
Figure 2. Bufalin affected the levels of cell markers in white blood cells from BALB/c mice with leukemia. Blood was collected from all mice andanalyzed for cell markers (A: CD3; B: CD19; C: CD11b and D: Mac-3) by flow cytometry, as described in the Materials and Methods. Significantlydifferent at p<0.05 vs. *normal control group (NC), vs. #leukemia control group (WC).

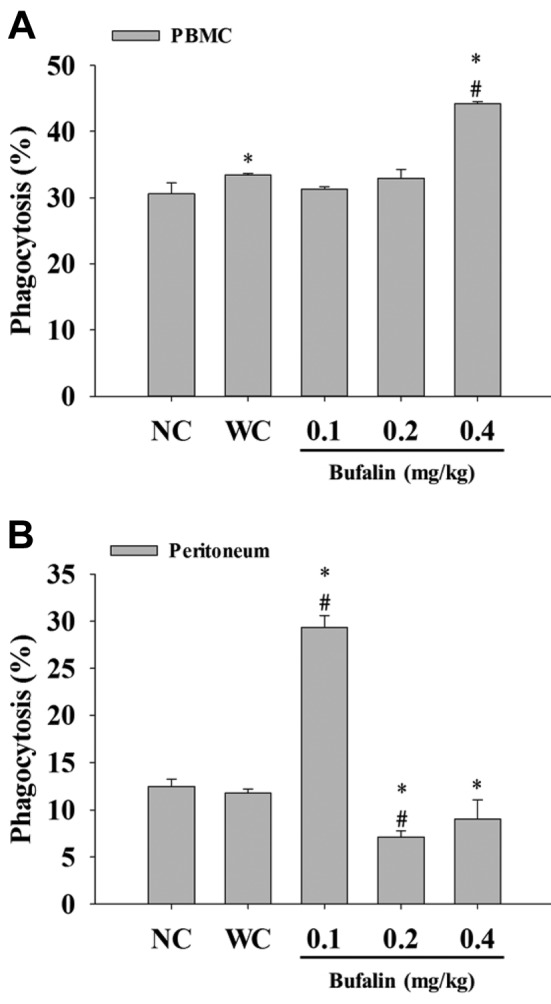
Bufalin affected phagocytosis by macrophages from PBMCs and peritoneal cavity of leukemic BALB/c mice. After treatment, macrophages were isolated from PBMCs and peritoneal cavity in order to measure the level of phagocytosis by flow cytometry. Bufalin treatment at 0.4 mg/kg significantly increased phagocytosis by macrophages isolated from PBMCs (Figure 3A), and at 0.1 mg/kg treatment by those from the peritoneal cavity but it was reduced at 0.2 and 0.4 mg/kg treatment (Figure 3B) when compared with the leukemic control group.
Figure 3. Bufalin promoted macrophage phagocytosis from peripheralblood mononuclear cells (PBMCs) and peritoneal cavity of BALB/cmice with leukemia. Blood samples were collected from mice thenmacrophages were isolated from PBMCs (A) and peritoneum (B) ofeach mouse. Macrophage phagocytosis was measured by flowcytometry, as described in the Materials and Methods. Significantlydifferent at p<0.05 vs. *normal control group (NC), vs. #leukemiacontrol group (WC).
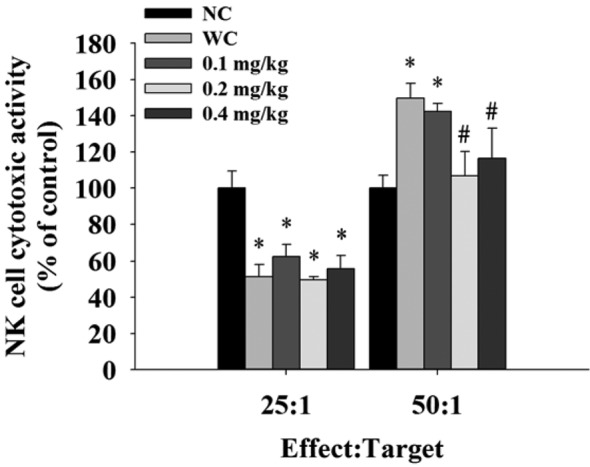
Bufalin affected the cytotoxic activity of NK cells from leukemic BALB/c mice. Yac-1 cells were used as targets for isolated splenocytes and then were assayed by flow cytometry. Results indicated that bufalin did not affect NK cell cytotoxic activity at 25:1 (splenocytes:target cells), but 0.2 and 0.4 mg/kg treatment reduced NK cell cytotoxic activity at 50:1 when compared to the leukemic control group.
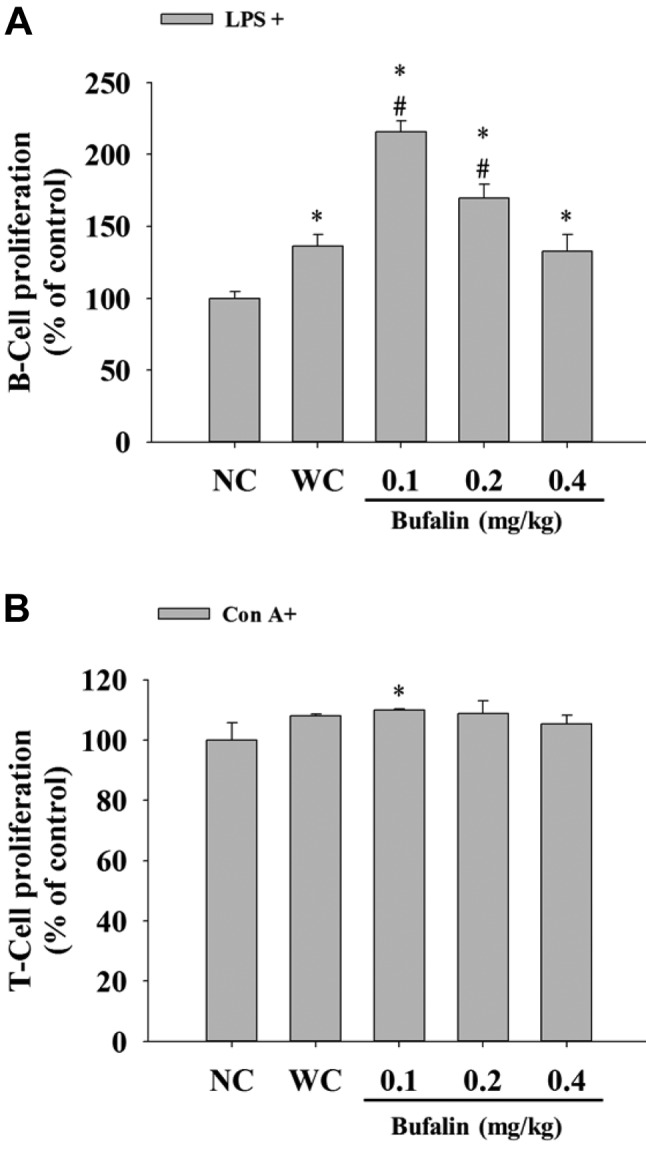
Bufalin affected B- and T-cell proliferation from leukemic BALB/c mice. Isolated splenocytes were assayed for cell proliferation. Results indicated that bufalin at 0.1 and 0.2 mg/kg significantly increased B-cell proliferation (Figure 5A) when compared with the leukemic control group, however, only 0.1 mg/kg treatment of bufalin significantly increased T-cell proliferation (Figure 5B) when compared with the leukemic control group.
Figure 5. Bufalin affected B- and T-cell proliferation in BALB/c micewith leukemia. Isolated B- and T-cells were pretreated withlipopolysaccharide (LPS) and concanavalin A (Con A) for determinationof B-cell (A) and T-cell (B) proliferation, respectively, and then wereanalyzed by flow cytometry as described in the Materials and Methods.Significantly different at p<0.05 vs. *normal control group (NC), vs.#leukemia control group (WC).
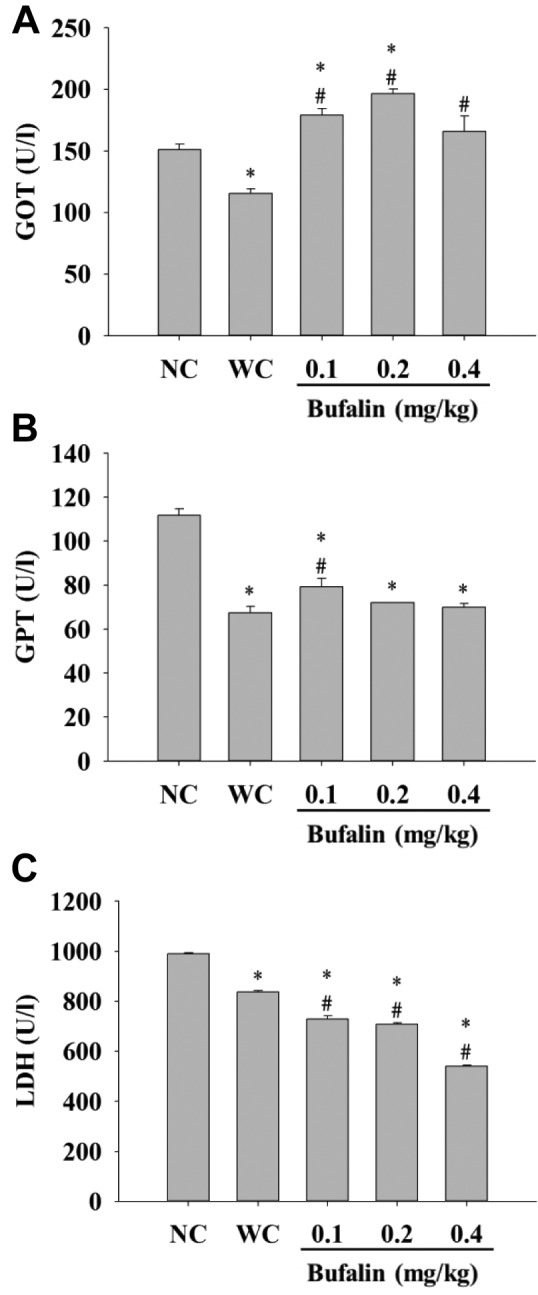
Bufalin affected the blood levels of GOT, GPT, and LDH of BALB/c mice. Blood samples were collected for measurement of GOT, GPT and LDH levels. Results indicated that bufalin significantly increased GOT level at all doses (Figure 6A), increased GPT level at 0.1 mg/kg treatment (Figure 6B), but reduced the level of LDH (Figure 6C) at all doses when compared with the leukemic control group.
Figure 6. Measurement of blood glutamic oxaloacetic transaminase(GOT) (A), glutamic pyruvic transaminase (GPT) (B), lactatedehydrogenase (LDH) (C) activity of BALB/c mice after exposed tobufalin. After treatment, blood samples were collected from each mouseand then the activity of GOT, GTP and LDH was measured, as describedin the Materials and Methods. Significantly different at p<0.05 vs.*normal control group (NC), vs. #leukemia control group (WC).
Discussion
In the present study, we injected murine WEHI-3 cells into normal BALB/c mice to generate an animal model of leukemia and these mice were randomly divided into different groups for oral treatment with bufalin at different doses to examine the effects of bufalin on immune responses in vivo. It is well known that several types of white blood cell interact with each other to produce immune responses in humans in order to act against foreign antigens (23). Many plant-derived bioactive compounds have been used to treat patients with cancer such as paclitaxel from Taxus brevifolia and camptothectin from Camptotheca acuminata (24). Although numerous reports have shown that bufalin induced cytotoxic effects via induction of cell-cycle arrest and apoptosis in many human cancer cell lines in vitro and has antitumor activity in vivo, however, there is no available information or data in literature regarding the effects of bufalin on immune responses in mice with leukemia in vivo. Our results are the first to demonstrate bufalin increases immune responses in leukemia BALB/c mice in vivo.
It is well documented that normal BALB/c mice develop leukemia after injection with WEHI-3 cells (25), which can be seen from their increased spleen weight when compared to normal mice (26). In the present study, we successfully generated leukemia in BALB/c mice using WEHI-3 cells and showed that bufalin significantly increased markers of immune-associated leukocytes in vivo. In particular, bufalin enhanced macrophage phagocytosis in these leukemic mice in vivo. It is well known that macrophages play a key role in innate immune responses to entry of antigens that is necessary to control or eliminate infection (27,28). The exact function of bufalin associated with Mac-3 marker and macrophage and NK cell activities needs further study as to whether they have any effect on survival of such mice with leukemia. Macrophages have great plasticity and can differentiate into several functional states in response to microenvironmental signals (29). Thus, some studies have focused on the roles of immune cell subtypes and their capacity to function in the tumor microenvironment (30). We examined macrophage phagocytosis using FITC-labelled E. coli as a target and assay by flow cytometry, which is a reliable method for this investigation (31). Yac-1 cells were used as targets for NK cell activity and were also assay by flow cytometry (32). The stimulation of NK cell cytotoxicity may increase immune response (33), thus the role of NK associated with bufalin-increased immune responses needs further investigation.
In conclusion, based on these observations, we suggest that bufalin may modulate immune responses not only through increasing monocyte (CD11b) population and T- and B-cell proliferation, but also by increasing macrophage phagocytosis in leukemic mice in vivo. Whether this translates into increased survival needs further investigation.
Acknowledgements
This work was supported by grant 104-47 from Cheng-Hsin General hospital, and SKH-8302-106-NDR-08 from Shin Kong Wu Ho-Su Memorial Hospital, Taipei Taiwan. Experiments and data analysis were performed in part through the use of the Medical Research Core Facilities Center, Office of Research & Development at China medical University, Taichung, Taiwan, R.O.C.
Figure 4. Bufalin affected the cytotoxic activity of natural killer (NK)cells in BALB/c mice with leukemia. Splenocytes were cultured in 96-well plates and target YAC-1 cells were added. NK cell cytotoxic activitywas measured using flow cytometry, as described in the Materials andMethods. Significantly different at p<0.05 vs. *normal control group(NC), vs. #leukemia control group (WC).
References
- 1.Alabsi AM, Ali R, Ideris A, Omar AR, Bejo MH, Yusoff K, Ali AM. Anti-leukemic activity of Newcastle disease virus strains AF2240 and V4-UPM in murine myelomonocytic leukemia in vivo. Leuk Res. 2012;36:634–645. doi: 10.1016/j.leukres.2011.11.001. [DOI] [PubMed] [Google Scholar]
- 2.Vardiman JW, Thiele J, Arber DA, Brunning RD, Borowitz MJ, Porwit A, Harris NL, Le Beau MM, Hellstrom-Lindberg E, Tefferi A, Bloomfield CD. The 2008 revision of the World Health Organization (WHO) Classification of Myeloid Neoplasms and Acute Leukemia: Rationale and Important Changes. Blood. 2009;114:937–951. doi: 10.1182/blood-2009-03-209262. [DOI] [PubMed] [Google Scholar]
- 3.Boelens J, Lust S, Vanhoecke B, Offner F. Chronic lymphocytic leukaemia. Anticancer Res. 2009;29:605–615. [PubMed] [Google Scholar]
- 4.Meggendorfer M, Roller A, Haferlach T, Eder C, Dicker F, Grossmann V, Kohlmann A, Alpermann T, Yoshida K, Ogawa S, Koeffler HP, Kern W, Haferlach C, Schnittger S. SRSF2 mutations in 275 cases with chronic myelomonocytic leukemia (CMML) Blood. 2012;120:3080–3088. doi: 10.1182/blood-2012-01-404863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sabattini E, Bacci F, Sagramoso C, Pileri S. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues in 2008: An overview. Pathologica. 2010;102:83–87. [PubMed] [Google Scholar]
- 6.Germing U, Kundgen A, Gattermann N. Risk assessment in chronic myelomonocytic leukemia (CMML) Leuk Lymphoma. 2004;45:1311–1318. doi: 10.1080/1042819042000207271. [DOI] [PubMed] [Google Scholar]
- 7.Diezi M, Garcia E, Weid N, Beck-Popovic M. No role for cerebrospinal fluid myelin basic protein levels in patients treated for childhood acute lymphoblastic leukemia. J Pediatr Hematol Oncol. 2009;31:393–397. doi: 10.1097/MPH.0b013e31819d1807. [DOI] [PubMed] [Google Scholar]
- 8.Puckett Y, Chan O. Cancer, leukemia, lymphocytic, acute (ALL). In: StatPearls. StatPearls Publishing LLC. 2018 [Google Scholar]
- 9.Marshall GM, Dalla Pozza L, Sutton R, Ng A, de Groot-Kruseman HA, van der Velden VH, Venn NC, van den Berg H, de Bont ES, Maarten Egeler R, Hoogerbrugge PM, Kaspers GJ, Bierings MB, van der Schoot E, van Dongen J, Law T, Cross S, Mueller H, de Haas V, Haber M, Revesz T, Alvaro F, Suppiah R, Norris MD, Pieters R. High-risk childhood acute lymphoblastic leukemia in first remission treated with novel intensive chemotherapy and allogeneic transplantation. Leukemia. 2013;27:1497–1503. doi: 10.1038/leu.2013.44. [DOI] [PubMed] [Google Scholar]
- 10.Takai N, Kira N, Ishii T, Yoshida T, Nishida M, Nishida Y, Nasu K, Narahara H. Bufalin, a traditional oriental medicine, induces apoptosis in human cancer cells. Asian Pac J Cancer Prev. 2012;13:399–402. doi: 10.7314/apjcp.2012.13.1.399. [DOI] [PubMed] [Google Scholar]
- 11.Hsiao YP, Yu CS, Yu CC, Yang JS, Chiang JH, Lu CC, Huang HY, Tang NY, Yang JH, Huang AC, Chung JG. Triggering apoptotic death of human malignant melanoma a375.s2 cells by bufalin: involvement of caspase cascade-dependent and independent mitochondrial signaling pathways. Evid Based Complement Alternat Med. 2012;2012:591241. doi: 10.1155/2012/591241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kang XH, Xu ZY, Gong YB, Wang LF, Wang ZQ, Xu L, Cao F, Liao MJ. Bufalin reverses HGF-induced resistance to EGFR-TKIs in EGFR-mutant lung cancer cells via blockage of MET/PI3K/AKT pathway and induction of apoptosis. Evid Based Complement Alternat Med. 2013;2013:243859. doi: 10.1155/2013/243859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Krenn L, Kopp B. Bufadienolides from animal and plant sources. Phytochemistry. 1998;48:1–29. doi: 10.1016/s0031-9422(97)00426-3. [DOI] [PubMed] [Google Scholar]
- 14.Li D, Qu X, Hou K, Zhang Y, Dong Q, Teng Y, Zhang J, Liu Y. PI3K/AKT is involved in bufalin-induced apoptosis in gastric cancer cells. Anticancer Drugs. 2009;20:59–64. doi: 10.1097/CAD.0b013e3283160fd6. [DOI] [PubMed] [Google Scholar]
- 15.Shen S, Zhang Y, Wang Z, Liu R, Gong X. Bufalin induces the interplay between apoptosis and autophagy in glioma cells through endoplasmic reticulum stress. Int J Biol Sci. 2014;10:212–224. doi: 10.7150/ijbs.8056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Tsai SC, Yang JS, Peng SF, Lu CC, Chiang JH, Chung JG, Lin MW, Lin JK, Amagaya S, Wai-Shan Chung C, Tung TT, Huang WW, Tseng MT. Bufalin increases sensitivity to AKT/mTOR-induced autophagic cell death in SK-HEP-1 human hepatocellular carcinoma cells. Int J Oncol. 2012;41:1431–1442. doi: 10.3892/ijo.2012.1579. [DOI] [PubMed] [Google Scholar]
- 17.Yan S, Qu X, Xu C, Zhu Z, Zhang L, Xu L, Song N, Teng Y, Liu Y. Down-regulation of CBL-B by bufalin results in up-regulation of DR4/DR5 and sensitization of TRAIL-induced apoptosis in breast cancer cells. J Cancer Res Clin Oncol. 2012;138:1279–1289. doi: 10.1007/s00432-012-1204-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wu SH, Wu TY, Hsiao YT, Lin JH, Hsu SC, Hsia TC, Yang ST, Hsu WH, Chung JG. Bufalin induces cell death in human lung cancer cells through disruption of DNA damage-response pathways. Am J Chin Med. 2014;42:729–742. doi: 10.1142/S0192415X14500475. [DOI] [PubMed] [Google Scholar]
- 19.Huang AC, Yang MD, Hsiao YT, Lin TS, Ma YS, Peng SF, Hsia TC, Cheng YD, Kuo CL, Chung JG. Bufalin inhibits gefitinib resistant NCI-H460 human lung cancer cell migration and invasion in vitro. J Ethnopharmacol. 2016;194:1043–1050. doi: 10.1016/j.jep.2016.11.004. [DOI] [PubMed] [Google Scholar]
- 20.Lee CH, Shih YL, Lee MH, Au MK, Chen YL, Lu HF, Chung JG. Bufalin induces apoptosis of human osteosarcoma U-2 OS cells through endoplasmic reticulum stress, caspase-and mitochondria-dependent signaling pathways. Molecules. 2017;22:E437. doi: 10.3390/molecules22030437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Chou HY, Chueh FS, Ma YS, Wu RS, Liao CL, Chu YL, Fan MJ, Huang WW, Chung JG. Bufalin induced apoptosis in SCC4 human tongue cancer cells by decreasing BCL2 and increasing BAX expression via the mitochondriadependent pathway. Mol Med Rep. 2017;16:7959–7966. doi: 10.3892/mmr.2017.7651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lin JJ, Lu KW, Ma YS, Tang NY, Wu PP, Wu CC, Lu HF, Lin JG, Chung JG. Alpha-phellandrene, a natural active monoterpene, influences a murine WEHI-3 leukemia model in vivo by enhancing macrophague phagocytosis and natural killer cell activity. In Vivo. 2014;28:583–588. [PubMed] [Google Scholar]
- 23.Arpinati M, Curti A. Immunotherapy in acute myeloid leukemia. Immunotherapy. 2014;6:95–106. doi: 10.2217/imt.13.152. [DOI] [PubMed] [Google Scholar]
- 24.Kim SH, Kim MJ, Kim YJ, Chang H, Kim JW, Lee JO, Lee KW, Kim JH, Bang SM, Lee JS. Paclitaxel as third-line chemotherapy for small cell lung cancer failing both etoposide-and camptothecin-based chemotherapy. Medicine. 2017;96:e8176. doi: 10.1097/MD.0000000000008176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lin JJ, Hsu SC, Lu KW, Ma YS, Wu CC, Lu HF, Chen JC, Lin JG, Wu PP, Chung JG. Alpha-phellandrene-induced apoptosis in mice leukemia WEHI-3 cells in vitro. Environ Toxicol. 2016;31:1640–1651. doi: 10.1002/tox.22168. [DOI] [PubMed] [Google Scholar]
- 26.Kuo YJ, Liu YJ, Way TD, Chiang SY, Lin JG, Chung JG. Synergistic inhibition of leukemia WEHI-3 cell growth by arsenic trioxide and Hedyotis diffusa Willd extract in vitro and in vivo. Exp Ther Med. 2017;13:3388–3396. doi: 10.3892/etm.2017.4392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Guirado E, Schlesinger LS, Kaplan G. Macrophages in tuberculosis: Friend or foe. Semin Immunopathol. 2013;35:563–583. doi: 10.1007/s00281-013-0388-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Verrall AJ, Netea MG, Alisjahbana B, Hill PC, van CR. Early clearance of Mycobacterium tuberculosis: A new frontier inprevention. Immunology. 2014;141:506–513. doi: 10.1111/imm.12223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sica A, Mantovani A. Macrophage plasticity and polarization: In vivo veritas. J Clin Invest. 2012;122:787–795. doi: 10.1172/JCI59643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Massa C, Seliger B. The tumor microenvironment: Thousand obstacles for effector T-cells. Cell Immunol. 2017 doi: 10.1016/j.cellimm.2017.12.004. [DOI] [PubMed] [Google Scholar]
- 31.Fan MJ, Yeh PH, Lin JP, Huang AC, Lien JC, Lin HY, Chung JG. Anthocyanins from black rice (Oryza sativa) promote immune responses in leukemia through enhancing phagocytosis of macrophages in vivo. Exp Ther Med. 2017;14:59–64. doi: 10.3892/etm.2017.4467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Takeda K, Okumura K. Interferon-gamma-mediated natural killer cell activation by an aqueous panax ginseng extract. Evid Based Complement Alternat Med. 2015;2015:603198. doi: 10.1155/2015/603198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chueh FS, Lin JJ, Lin JH, Weng SW, Huang YP, Chung JG. Crude extract of Polygonum cuspidatum stimulates immune responses in normal mice by increasing the percentage of Mac-3-positive cells and enhancing macrophage phagocytic activity and natural killer cell cytotoxicity. Mol Med Rep. 2015;11:127–132. doi: 10.3892/mmr.2014.2739. [DOI] [PMC free article] [PubMed] [Google Scholar]