

Abstract
Background:
Economy type is an important determinant of reinforcer value. This study investigated the effect of open and closed economies on demand and preference for cocaine and saccharin in rats.
Methods:
In the first phase, rats were trained to lever press for cocaine infusions or saccharin. The number of presses required for each reinforcer increased across sessions. ocaine and saccharin economy type was manipulated over groups by varying post-session availability of these reinforcers. One group of rats had three hours’ post-session access to unlimited cocaine (open economy). A second group had three hours’ post-session access to unlimited saccharin. A third group had no post-session access to either reinforcer (closed economy). In a second phase, rats in the three conditions could make mutually exclusive choices for cocaine or saccharin.
Results:
Post-session access to saccharin caused saccharin demand to become more elastic. Post-session access to cocaine had no effect on demand for cocaine but made demand for saccharin more elastic. Results from the choice phase generally paralleled those from the demand phase, the main finding being that post-session saccharin access caused an increase in cocaine preference.
Conclusions:
These results show that manipulating economy type can affect cocaine and non-drug reinforcers differently. Opening the saccharin economy decreased saccharin’s value. Opening the cocaine economy did not decrease cocaine’s value, but instead led to a devaluation of saccharin. These results suggest that cocaine choice may be determined not only by the reinforcers immediately available, but also by those reinforcers’ broader contexts of availability.
Keywords: Cocaine, economy type, demand, choice, saccharin, rats
1. Introduction
Addiction can be conceptualized as persistent choice of drugs over non-drug alternatives (Ahmed, 2010; Heyman, 2009; Rachlin, 1997). Choice is determined by the value of the reinforcers available (Rachlin, 1992). Understanding why, in some individuals or situations, the value of drug reinforcers surpasses that of non-drug alternatives could help explain addiction and provide clues for potential treatment strategies. The present study investigated, within an animal model, how one potential determinant of reinforcer value – economy type – affected the value of cocaine and a non-drug alternative reinforcer.
Economy type can be an important determinant of a reinforcer’s effect on behavior (Hursh, 1980, 1991). In a closed economy, the experimental session is the only source of the reinforcer. For example, if the only food a subject receives is that earned by lever pressing during a session, the economy for food is closed. In this situation, the total amount of food obtained is completely dependent on the subject’s behavior emitted under that contingency. In an open economy, on the other hand, the subject earns the reinforcer during the experimental session, but also has access to the reinforcer from other sources outside of the experimental session. For example, in studies using food as the reinforcer, the experimenter often provides post-session supplemental food. In such open economies, total food received is less dependent on the subject’s behavioral output under the contingency programmed during the session than it is in a closed economy. The distinction between an open and closed economy is not an absolute one, but rather the openness of an economy can be thought of as being on a continuum (Hursh, 1980). If the experimenter provides a large amount of post-session food, the economy can be described as being more open than when the experimenter provides only a small amount of post-session food.
Hursh et al. (1989) performed an experiment showing how opening the economy for food affected how hard subjects were willing to work for food. Monkeys were trained to press a push-plate for food during four daily “work” sessions. The fixed-ratio (FR) schedule in effect increased over days from 10 to 372. The important manipulation of the experiment was the amount of “free” food that monkeys could obtain after the work sessions. In the closed-economy condition, no free food was available. In a second condition, there was a 20-minute period at the end of the four work sessions during which monkeys could obtain additional (free) food on an FR-1 schedule. In a third condition, food was available on an FR-1 schedule for five minutes at the end of each of the four work sessions. The main finding was that the provision of free food made subjects less willing to work for food, especially when relatively large FRs were operative during the work sessions. Stated in the language of behavioral economics, opening the economy for food made demand for it more elastic.
Little work has been done on investigating the effect of economy type on drug reinforcers. Carroll et al. (2000) found that post-session provision of ethanol reduced monkeys’ progressive-ratio schedule breakpoints for ethanol during the work session. In contrast, post-session provision of phencyclidine (PCP) had no effect on PCP breakpoints. In humans, post-session provision of cigarettes made demand for cigarettes more elastic during a work session where subjects mouse-clicked on ratio schedules for cigarettes (Mitchell et al., 1994, 1998). The same effect was observed in a similar study where coffee was the reinforcer instead of cigarettes (Mitchell et al., 1995). To date, no study has investigated the effect of manipulating the economy type of cocaine.
The primary goal of the present study was to determine the effect of economy type on the elasticity of demand for cocaine and a non-drug alternative, saccharin. This was accomplished by allowing rats to work for cocaine and saccharin reinforcers on an ascending series of FR schedules during a three-hour work session. This work session was followed by a second three-hour session where different groups of rats had unlimited FR-1 access to cocaine (the OpenCoc group), saccharin (the OpenSac group), or neither (the Closed group). It was hypothesized that opening the economy for saccharin would make demand for it more elastic (i.e., rats should be less willing to work for it as price increases). From a behavioral economic perspective, demand for cocaine would also be expected to be more elastic in an open economy for cocaine. However, previous studies have shown that increased access to cocaine can increase its reinforcing strength, as measured by demand analyses (Christensen et al., 2008b), progressive-ratio breakpoints (PatersonandMarkou,2003), and resistance to punishment (Deroche-Gamonet et al., 2004; Pelloux et al., 2007). Because opening the economy for cocaine entails increased cocaine exposure, it is possible that demand for cocaine might not become more elastic when the cocaine economy is opened.
A second goal of the present study was to determine the effect of manipulating the cocaine and saccharin economy types on choice between these reinforcers. The same basic strategy as that described above was used, except that the work session was replaced by a discrete-trials choice session where rats made mutually exclusive choices between the two reinforcers (Lenoir et al., 2007). When Nader and Woolverton (1992) opened the economy for food, they found that monkeys increased their choice of cocaine over food. It was expected that opening the economy for saccharin would also increase choice of cocaine here. However, it is not known how opening the cocaine economy affects choice. Nader and Woolverton did not manipulate the cocaine economy in their study. From a behavioral economic perspective, opening the cocaine economy would be expected to increase choice of saccharin. But, as described above, opening the cocaine economy entails increased access to cocaine, which previous studies have shown can result in an increase in cocaine’s reinforcing strength.
The effects of economy type could be relevant for understanding addiction. Bickel et al. (2011) have attributed addiction to a reinforcer pathology, where drug reinforcers are overvalued and non-drug reinforcers are undervalued (see also Rachlin, 1992, 1997). Economy type could be a factor that contributes to this situation. For example, if an individual is faced with the choice between a drug and a non-drug alternative, that individual may be more likely to choose the drug when it is relatively scarce (as in a closed economy) and the non-drug alternatives that are available can be easily obtained from multiple sources (as in an open economy). Conversely, they may be more inclined to choose the non -drug alternative when it is relatively scarce and the drug is readily available from different sources (e.g., as part of replacement therapy). The present study was designed to model these dynamics and could therefore provide some insight into addiction.
2. Materials and methods
2.1. Subjects
Twenty-seven adult male Long-Evans rats (Charles River Laboratories, Frederick, MD), weighing approximately 400 g at the beginning of experimental sessions, served as subjects. Rats were individually housed in plastic cages with wood-chip bedding and had unlimited access to rat chow and water in their home cages. The colony room where the rats were housed had a 12- hour light:dark cycle with lights on at 08:00 h. Training sessions were conducted five days per week during the light phase of the light:dark cycle. Throughout the experiment, rats were treated in accordance with the Guide for the Care and Use of Laboratory Animals (National Academy of Sciences, 2011) and all procedures were approved by American University’s Institutional Animal Care and Use Committee.
2.2. Apparatus
Training took place in 20 Med Associates (St. Albans, VT) operant test chambers. Each chamber measured 30.5 × 24 × 29 cm and had aluminum front and rear walls with clear polycarbonate side walls. On the front wall of the chamber, two Med-Associates retractable levers were located on either side of a Med Associates small cup liquid receptacle. Saccharin reinforcers (0.3 ml of 0.2% saccharin solution) were delivered to this liquid receptacle. A 100-mA cue light was located above each lever. speaker located in the center of the front wall provided a tone stimulus (4500 Hz and 80 dB). Cocaine (provided by the Drug Supply Program, National Institute on Drug Abuse, Bethesda, MD) in a saline solution at a concentration of 2.56 mg/ml was infused at a rate of 3.19 ml/min by 10-ml syringes driven by Med-Associates (St. Albans, VT) syringe pumps. ygon tubing extended from the 10-ml syringes to a 22-gauge rodent single-channel fluid swivel (Instech Laboratories, Plymouth Meeting, PA) and tether apparatus (Plastics One, Roanoke, VA) that descended through the ceiling of the chamber. Cocaine was delivered to the subject through Tygon tubing that passed through the metal spring of the tether apparatus. For half of the chambers, the 100-mA houselight was located at the top of the front wall of the chamber. For the other half, this houselight was located just outside of the clear left side wall of the chamber and near the floor.
2.3. Surgery
Before beginning operant training, all rats were surgically prepared with chronic indwelling jugular vein catheters, using procedures described in detail elsewhere (Thomsen and Caine, 2005; Tunstall and Kearns, 2014). In brief, approximately 3.5 cm of Silastic tubing was inserted into the right jugular vein. From this insertion site, an additional 8 cm of Silastic tubing passed under the skin to the midscapular region where it connected to the 22-guage stainless steel tubing of a backmount catheter port (Plastics One, Roanoke, VA) that was implanted subcutaneously. The spring tether in the chamber was attached to the threaded plastic cylindrical shaft of the port that protruded through an opening in the skin. All surgery was conducted under ketamine (60 mg/kg) and xylazine (10 mg/kg) anesthesia. Rats were given 7–10 days to recover from surgery. Catheters were flushed daily with 0.1 ml of a saline solution containing 1.25 /ml heparin and 0.08 mg/ml gentamicin.
2.4. Procedure
2.4.1. Overview.
See Figure 1 for a procedural schematic. Rats were randomly assigned to groups prior to beginning training. Rats received two three-hour sessions per day separated by an approximately 15-to 20-minute transition period where rats were returned to their homecages while the chambers were prepared for the second session. The first session of the day was the “work” session. During this session, rats in all groups were trained on a demand procedure (Phase 1) or a choice procedure (Phase 2) where they lever pressed for cocaine and saccharin. During the second session, rats in the OpenCoc group received three hours of access to cocaine only on an FR-1 schedule, rats in the OpenSac group received three hours of access to saccharin only on an FR-1 schedule, and rats in the Closed group were placed in the chamber for three hours without access to either reinforcer. Rats experienced each of the two sessions in a different chamber and for each rat the position of the houselight (near ceiling of front wall vs. near floor of side wall) differed over the two chambers to enhance discriminability. Separate chambers were used for the two daily sessions in an effort to make rats in the open economy groups learn that, in one situation they had to pay an increasing price for each reinforcer (or had limited opportunities to choose that reinforcer) whereas in another situation, one of the reinforcers was always almost freely available.
Figure 1.
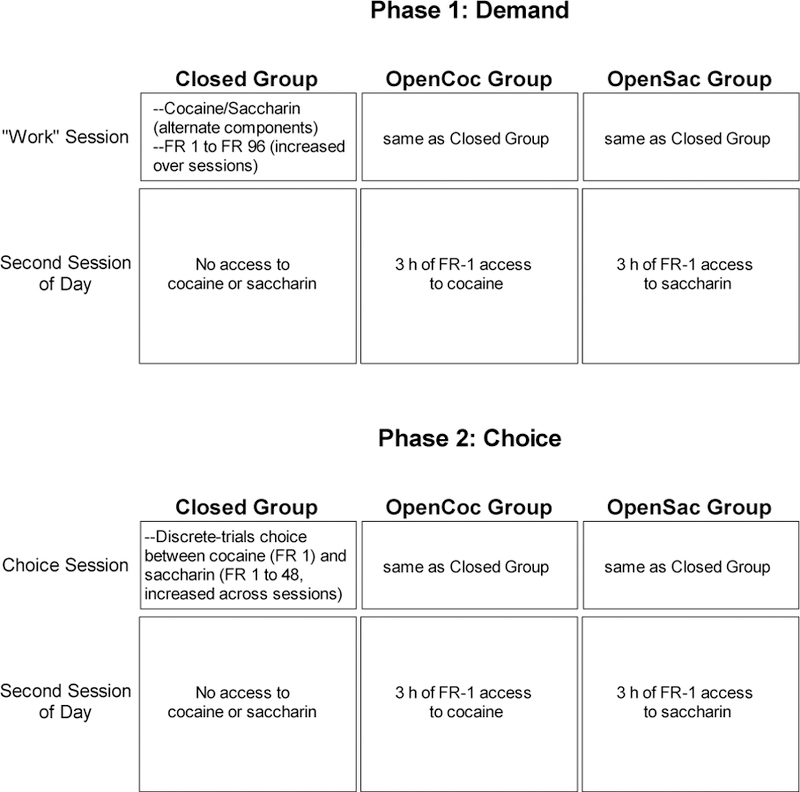
Schematic diagram of procedures used in Phase 1 and Phase 2. In both phases, the procedure used in the first session of the day was the same for all groups. The groups differed in their second session experience.
During the “work” session of the demand phase, rats in all groups had a total of one hour of access to cocaine (in four 15-minute components) and one hour of access to saccharin (in four 15-minute components) for five days per week. The OpenCoc group had an additional three hours of access to cocaine following each work session. During the choice phase, rats in all groups could make up to 14 choices between a single cocaine infusion and a single saccharin reinforcer during sessions that occurred five days per week. Again, the OpenCoc group had an additional three hours of access to cocaine following each choice session. As described below in more detail, across the experiment rats had a total of approximately 40–45 days of access to cocaine spread over sessions conducted five days per week.
2.4.2. Phase 1: Demand for cocaine and saccharin.
The demand procedure used during the work session was similar to that in Christensen et al. (2008a). Rats were first trained to lever press for cocaine and for saccharin an FR-1 schedule. During each session, there were eight, 15-minute components where one of the two retractable levers was inserted. There were four presentations each of the cocaine and saccharin levers. Cocaine and saccharin components randomly alternated, with the restriction that there were no more than two consecutive components of the same type. Each component was followed by a 7.5-minute period where both levers were retracted. Thus, over the course of the three hour work session, rats had access to each of the levers for a total of 60 minutes. The position (left vs. right) of the cocaine and saccharin levers was counterbalanced over rats. During cocaine-lever components, a lever press resulted in a 0.75-mg/kg cocaine infusion, simultaneous illumination of the cue light above the lever, and a 10-seecond tone presentation. Lever presses were recorded, but had no consequences during this 10-second period. During saccharin-lever components, pressing the lever resulted in the delivery of 0.3 ml of saccharin solution in the cup, illumination of a 100-mA bulb located inside the liquid receptacle for 1.66 seconds (the time it took to fill the cup), and initiation of a 10-second timeout that was signaled by illumination of the cue light above the lever. If during lever-press acquisition a rat reliably pressed one of the levers but not the other, the rat was given one or more sessions where only the non-pressed lever was available for the entire three hour session. Once the rat began pressing this lever, the rat was returned to the procedure described above where the two levers alternated for 15-minute components separated by 7.5-minute periods where both levers were retracted.
Rats were trained on the alternating lever procedure with an FR-1 schedule in effect for a minimum of 12 sessions and until the consumption of each reinforcer stabilized. Stability was defined as three consecutive sessions where the total number of reinforcers earned of each type did not vary from the rolling three-session mean by more than 25%. Once this stability criterion was reached, the FR increased over blocks of two sessions according the following sequence: 3, 6, 12, 24, 48, 96. The same price was in effect for both cocaine and saccharin for the duration of a session. Prices increased across sessions.
Upon completion of the work session, rats were removed from the operant chamber and put back in their homecages while the second session was prepared (programs loaded, syringes refilled, etc.). This typically took 15–20 minutes. For rats in the OpenCoc group, the left lever was inserted for the entire three-hour second session. Each lever press resulted in delivery of 0.75 mg/kg cocaine and initiated a 10-second timeout period during which the cue light above the lever was illuminated and the tone sounded. For rats in the OpenSac group, the right lever was inserted into the chamber. Each lever press resulted in the delivery of 0.3 ml saccharin, illumination of the light bulb inside the receptacle for 1.66 seconds, and initiation a 10-second timeout period signaled by illumination of the cue light. Rats in the Closed group also had a three-hour second session each day where the houselight was illuminated, but no levers were inserted and no cocaine or saccharin reinforcers were delivered.
2.4.3. Choice between cocaine and saccharin.
After the demand phase, rats were trained on the discrete-trials choice procedure introduced by Ahmed and colleagues (Lenoir et al., 2007; Cantin et al., 2010) for the first of the two daily sessions. Each choice session began with four forced-choice trials. There were two forced-choice trials each with the cocaine and saccharin levers, with trial order randomized within blocks of two. These forced-choice trials ensured that rats sampled each lever twice at the beginning of each session. On a cocaine trial, the cocaine lever was inserted into the chamber and a single lever press resulted in delivery of a cocaine infusion (0.75 mg/kg/infusion), presentation of the associated 10-second audiovisual cue, and retraction of the lever. On a saccharin trial, the saccharin lever was inserted and a single lever press resulted delivery of 0.3 ml of the saccharin solution, retraction of the lever, and illumination of both the cue light above the lever and the light inside the liquid receptacle. Each trial was followed by a 10-minute intertrial interval (ITI). Following the four forced-choice trials, there were up to 14 free-choice trials. Now, both levers were inserted simultaneously. A single press on the selected lever resulted in delivery of the designated reinforcer and retraction of both levers. (That is, an FR-1 schedule was in effect on both levers.) A 10-minute ITI followed each free-choice trial. During free-choice trials, a limited hold was in effect whereby if no choice was made within 120 seconds, both levers retracted, no reinforcers were delivered, and the ITI began. Such trials were recorded as omissions. Sessions lasted for a maximum of 14 free-choice trials or 195 minutes, whichever occurred first. Rats were trained on this procedure for at least five sessions and until preference stabilized. The stability criterion was three consecutive sessions in which choices for cocaine did not differ from the rolling three-session mean by more than 20%. Once this criterion was reached, rats were tested over an additional five sessions on a choice procedure where the FR in effect on the saccharin lever increased according to the sequence 3, 6, 12, 24, 48. Cocaine was always available on an FR-1 schedule. This was done in case strong preference for saccharin obscured potential group differences. All other aspects of the choice procedure were the same as described above.
2.5. Data analysis
For the demand data, the number of cocaine infusions self-administered and the number of saccharin reinforcers earned were averaged over the training sessions at each FR (for FR 1, the mean of the last two sessions was used). Individual and group mean consumption data were fit by Hursh and Silberberg’s (2008) exponential-demand equation:
| (1) |
where Q is quantity consumed, Q0 is consumption as price approaches 0, k is a constant defining the consumption range in log units (k = 2.5 here), α determines the rate of decline in consumption, and is cost (FR size).
The primary measure of interest from the demand phase was essential value (EV), which is inversely related to α and reflects inelasticity of demand. EVs of cocaine and saccharin were calculated for each subject according the formula given by Hursh (2014):
| (2) |
For the choice phase, the primary measure was percent choice of cocaine. Interpolated indifference points were also calculated (Anderson and Woolverton, 2005). This was the saccharin FR value estimated to produce 50% cocaine/saccharin choice.
For all statistical tests, α was set to 0.05. For the demand phase data, one-way ANOVAs were performed on numbers of sessions required to meet criterion. Non-parametric Kruskall-Wallis and Mann-Whitney tests were used to compare groups on cocaine and saccharin EV. Non-parametric tests were used here due to non-normal distribution of EV. Spearman correlation coefficients were used to evaluate the strength of the association between EV and number of reinforcers consumed in the second of the two daily sessions for the two open economy groups. For the choice data, ANOVAs followed by Tukey’s posthoc tests, where appropriate, were performed on the indifference point measure and numbers of omissions. Spearman coefficients were used to evaluate the relationship between choice indifference point and number of reinforcers consumed in the second session for the two open economy groups.
3. Results
The groups’ mean bodyweights did not differ over the course of the experiment. At the start of Phase 1, the Closed, OpenCoc, and OpenSac groups’ mean weights were 404.1 (± 9.5 SEM), 399.2 (± 7.1), and 421.7 (± 15.7) g, respectively. At the beginning of Phase 2, their mean weights were 457.3 (± 15.3), 455.9 (± 11.9), and 471.7 (± 16.6). At the end of the experiment, their mean weights were 504.4 (± 17.7), 471.6 (± 15.5), and 504.6 (± 20.5) g, respectively. A repeated measures ANOVA indicated there was a main effect of Time (F[2,42] = 69.6, p < 0.001) on weight, but no effect of Group (F < 1) or Group x Time interaction (F[4,42] = 1.1 p > 0.35).
3.1. Phase 1: Demand
During acquisition of lever pressing on the alternating levers procedure, some rats in the two open economy groups regularly pressed one lever but not the other during the work session. To correct this, six of the rats in the OpenCoc group required sessions (mean = 5.2, SEM = 1.5) where only the saccharin lever was available. Another two rats in that group required sessions (mean = 1.5, SEM= 0.5) with only the cocaine lever available. Three rats in the OpenSac group required sessions (mean = 3.7, SEM = 0.7) where only the cocaine lever was available and one rat required five sessions where only the saccharin lever was available. Once these rats reliably pressed both levers, they were put back on the alternating levers procedure. In total, rats in the Closed, OpenCoc, and OpenSac groups required means of 15.1 (± 1.3 EM), 22.3 (± 3.0), and 16.8 (± 1.3) sessions, respectively, on the FR-1 alternating levers procedure to meet the acquisition criterion. These means did not significantly differ (one-way ANOVA, F[2,24] = 3.4, p > 0.05).
Figure 2 shows group mean consumption of cocaine and saccharin during work sessions across the range of FRs tested as well as fits of the exponential demand model. The model fit the data well, with median R2 values (based on model fits to individual subjects’ data) of 0.86 (± 0.14 inter-quartile range), 0.93 (± 0.14), and 0.94 (± 0.10) for the cocaine consumption data in the Closed, OpenCoc, and OpenSac groups, respectively. For the saccharin consumption data, these values were 0.96 (± 0.05), 0.88 (± 0.24), and 0.97 (± 0.05), respectively. As can be seen in the left panel, demand curves for cocaine were highly similar across groups. In contrast, demand for saccharin (right panel) decreased more quickly as price increased in the two open economy groups than it did in the Closed group.
Figure 2.
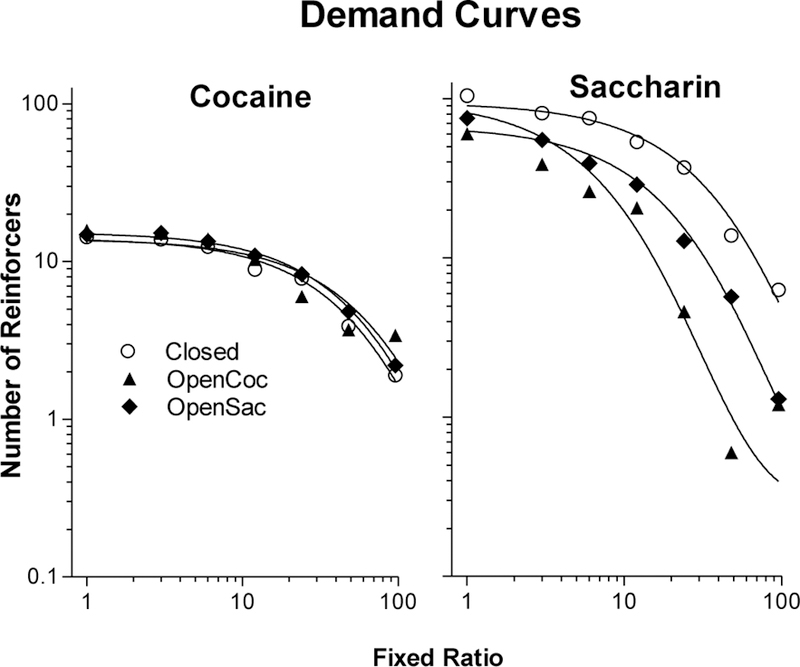
Group mean numbers of cocaine infusions (left panel) and saccharin reinforcers (right panel) earned by the Closed group (open circles), OpenCoc group (triangles), and OpenSac group (diamonds) at each FR during the demand phase as well as fits of the exponential demand model to this consumption data.
Figure 3 presents group mean EVs for cocaine and saccharin based on fits of the exponential demand model to individual subjects’ consumption data. These results largely parallel those presented in Figure 2, where the model was fit to group mean consumption. As the left panel of Figure 3 shows, there was no difference over groups in cocaine EV (Kruskall-Wallis H[2] = 2.2, p > 0.3). However, there was a group difference for saccharin EV (H[2] = 12.9, p < 0.005). Mann-Whitney U tests confirmed that the Closed group had significantly higher saccharin EV than the OpenCoc group (U[8,9] =3.0,p=0.001)or the OpenSac group (U[8,10] = 9.0, p < 0.005), but the two latter groups did not differ from each other (U[9,10] = 27.5, p >0.15).
Figure 3.
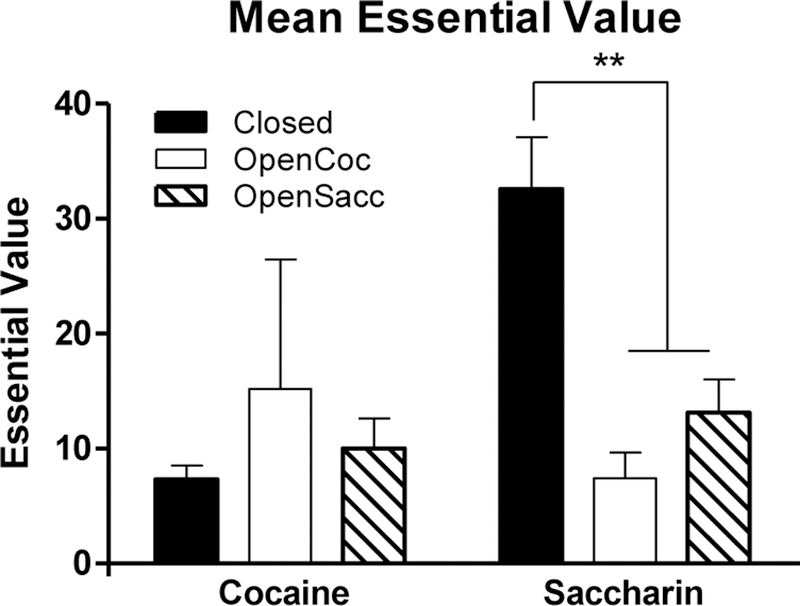
Mean (± SEM) essential value of cocaine (left) and saccharin (right) based on fits of the exponential model to individual subjects’ data for the Closed (black bars), OpenCoc (white bars), and OpenSac (striped bars) groups. ** indicates p < 0.01.
Figure 4 presents consumption of cocaine or saccharin in the Open oc and OpenSac groups, respectively, during the second daily three-hour session for those groups when the FR was always 1. Rats in both groups consumed approximately 40 reinforcers per session throughout the demand phase. A 2 × 7 (Group x FR) ANOVA performed on the numbers of reinforcers earned during the second session in the open economy groups indicated that there was no effect of Group or FR and no Group x FR interaction (all Fs < 1). There was a significant correlation in the Open Sac group between EV of saccharin and number of saccharin reinforcers consumed during the post-demand session (rs[10] = 0.28, p > 0.4).
Figure 4.
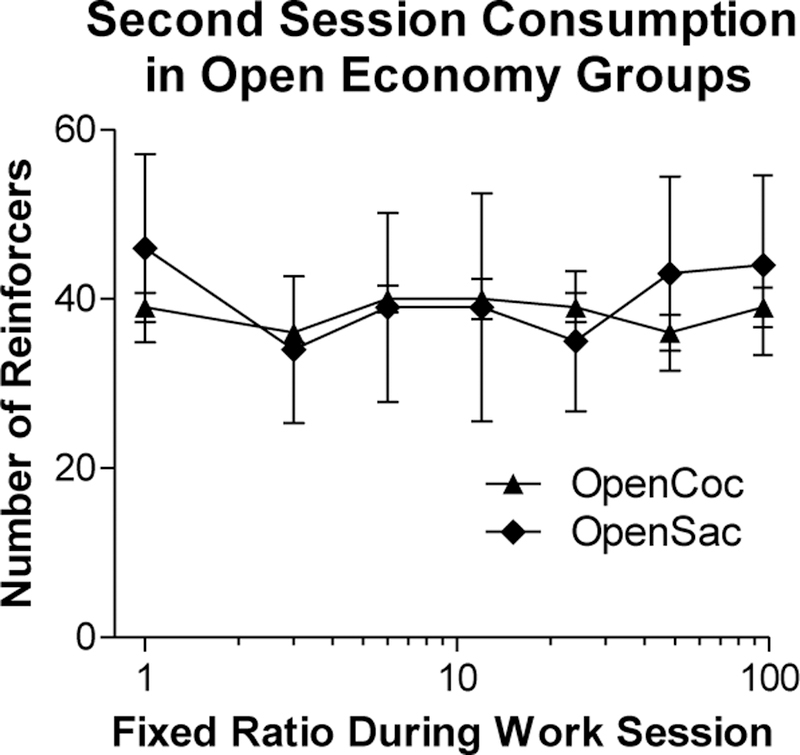
Mean (± SEM) numbers of cocaine infusions or saccharin reinforcers obtained on an FR-1 schedule by the OpenCoc (triangles) and OpenSac (diamonds) groups, respectively, in Phase 1 during their second session of the day (i.e., during the 3-h post-work-session access period). The FR shown on the x-axis corresponds to the ratio in effect during the work session.
3.2. Phase 2: Choice
Two rats in the OpenCoc group and one rat in the OpenSac group became ill and died before completing the choice phase, reducing the ns for these groups to 7 and 9, respectively, for this phase. he Closed, OpenCoc, and OpenSac groups required means of 5.9 (± 0.4 SEM), 5.5 (± 0.4), and 5.1 (± 0.1) sessions, respectively, to meet the stability criterion on the choice procedure (no group difference, F[2,22] = 1.5, p > 0.2). Numbers of omissions remained relatively low throughout the choice phase. Across saccharin FRs, the mean numbers of omissions ranged from 0 to 1.0 in the Closed group, from 0.4 to 1.9 in the OpenCoc group, and from 0.4 to 1.8 in the OpenSac group. There was no significant effect of Group (F < 1), FR (F[5, 105] = 1.4, p > 0.2), or interaction of Group with FR (F[10,105] = 1.2, p > 0.25).
The left panel of Figure 5 shows the mean percentage of choices for cocaine for each of the groups across the range of saccharin FRs tested (the cocaine FR was always 1). The dashed line in the figure represents indifference (50% of choices for cocaine and for saccharin). As can be seen in the figure, all groups increased cocaine choice as the saccharin FR was raised, though the Closed group was overall less inclined to choose cocaine than the other two groups. The right panel of Figure 5 presents mean choice indifference points (based on individual subjects’ indifference points). This figure shows that rats in the Closed group did not become indifferent between cocaine and saccharin until the saccharin FR was raised to a mean of about 22, whereas the two open economy groups were indifferent at a saccharin FR of approximately 7–8. one-way ANOVA performed on the indifference points indicated that the groups significantly differed (F[2,23] = 4.6, p < 0.05). Subsequent Tukey posthoc tests confirmed that the Closed group’s mean indifference point was significantly higher than that of the other two groups (both ps < 0.05), which did not differ from each other (p > 0.95).
Figure 5.
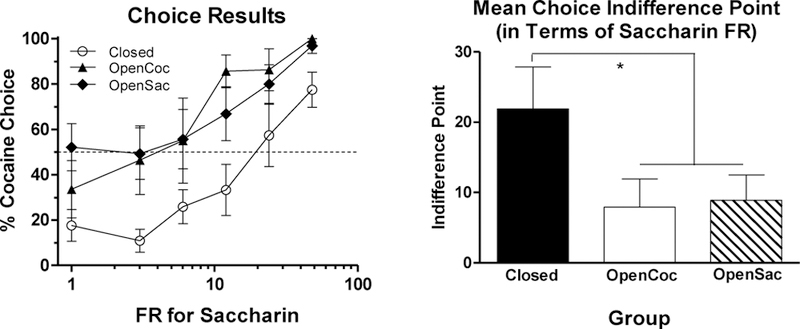
The left panel presents mean (± SEM) percent choice of cocaine at each saccharin FR value for the Closed (open circles), OpenCoc (triangles), and OpenSac (diamonds) groups. The FR for cocaine was always 1 throughout the choice phase. The horizontal dashed line represents indifference (50% choice of cocaine or saccharin). The right panel shows mean (± SEM) indifference points for each of the groups. The indifference point is the estimated saccharin FR at which rats would choose cocaine and saccharin equally. * indicates p < 0.05.
Figure 6 shows the mean numbers of cocaine infusions and saccharin reinforcers obtained by the Open oc and OpenSac groups, respectively, during the three-hour session following the choice session. As during the demand phase, there was no significant effect of Group, FR, or the Group x FR interaction (all Fs < 1.8, all ps > 0.2). There was no significant correlation between choice indifference point and number of reinforcers consumed during the post-choice session in either the OpenCoc group (rs[7] = −0.67, p > 0.10) or the OpenSac group (rs[9] = .10, p > 0.8).
Figure 6.
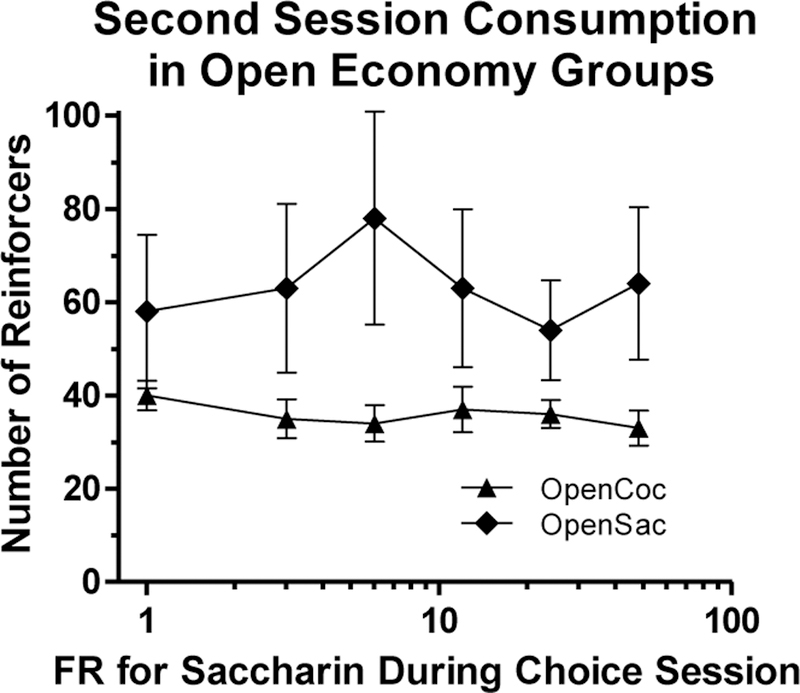
Mean (± SEM) numbers of cocaine infusions or saccharin reinforcers obtained on an FR-1 schedule by the OpenCoc (triangles) and OpenSac (diamonds) groups, respectively, in Phase 2 during their second session of the day (i.e., during the 3-h post-choice-session access period). The FR shown on the x-axis corresponds to the saccharin ratio in effect during the choice session
4. Discussion
Opening the economy for saccharin made demand for it more elastic. That is, rats were less willing to work for saccharin as its price increased when they had post-session access to it. This outcome is consistent with previous studies finding that demand became more elastic when rats were given post-session access to food (Hursh et al., 1989), when monkeys were given post-session access to ethanol (Carroll et al., 2000), and when humans were given post-session access to cigarettes (Mitchell et al., 1995, 1998) or coffee (Mitchell et al., 1995). In the present experiment, post-session access to saccharin also increased rats’ preference for cocaine over saccharin. This result is similar to Nader and Woolverton’s (1992) finding that post-session access to food increased monkeys’ choice of cocaine over food.
Post-session access to cocaine, on the other hand, had no effect on its elasticity of demand. This was observed even though the number of cocaine infusions obtained by the OpenCoc group after the work session was similar to the number of saccharin reinforcers that the OpenSac group obtained after the work session. Therefore, it does not appear that the OpenCoc group received too few extra cocaine infusions to make demand for it more elastic.
It may be thought that different satiation time courses for saccharin and cocaine could explain the different results of opening the economies for these two reinforcers. According to this account, post-session saccharin received on one day may have caused rats in the OpenSac group to be partially sated at the start of the work session the following day. In contrast, due to the short half-life of i.v. cocaine in rats (~18 minutes; Barbieri et al., 1992), post-session cocaine would have been eliminated from OpenCoc rats well before the start of the next day’s work session. It should be noted, however, that the half-life of saccharin in rats is also short enough (~30 minutes; Sweatman and Renwick, 1980; Renwick, 1985) so that saccharin consumed on one day would have been eliminated prior to the start of the next day’s work session. Further, saccharin satiety in rats is controlled by immediate oral sensory stimulation rather than by any longer-term post-ingestive consequences such as fullness of the stomach or overhydration (Mook et al., 1981). Nonetheless, the neurobiological mechanisms involved in saccharin satiety are poorly understood and it is possible that residual satiety-like effects of prior saccharin exposure could have been responsible for the OpenSac group’s results.
An alternative possibility is that differences in rates of delay discounting of cocaine and saccharin could explain the different effects of opening the economies for these two reinforcers. Post-session access to the two reinforcers was necessarily delayed. If rats more steeply discounted delayed cocaine than delayed saccharin, this could explain why there was no effect of opening the cocaine economy on cocaine’s EV while opening the saccharin economy was effective in decreasing saccharin’s EV. While studies comparing discount rates of cocaine and saccharin have not been performed in rats, monkeys discounted delayed cocaine less steeply than they did delayed saccharin (Freeman et al., 2009) or delayed food (Huskinson et al., 2016). If rats discount cocaine and saccharin similarly to the way that monkeys do, the delay discounting explanation described above could not account for the present results.
Post-session access to cocaine caused rats to work less hard to defend saccharin consumption as saccharin’s price increased. The OpenCoc group worked for saccharin (and cocaine) during one session and then self-administered cocaine for three hours in a second session. Saccharin followed by delayed cocaine has been shown in previous studies to result in the devaluation of saccharin as measured by taste reactivity (Green et al., 2015; Grigson, 1997, 2008; Grigson and Twining, 2002; Wheeler et al., 2008, 2011). For example, Green et al. first infused a drop (0.2 ml) of saccharin into rats’ mouths once per minute for 45 minutes. Then, rats self-administered cocaine for two hours. Initially, rats made more appetitive than aversive mouth movements in response to the saccharin, but after 14 sessions they made more aversive than appetitive mouth movements. This devaluation of saccharin has been attributed to the development of a conditioned compensatory response (i.e., a cocaine-opposing response) elicited by the saccharin, which was predictive of delayed cocaine reinforcement (Grigson, 2008; Wheeler et al., 2011). Rats in the OpenCoc group here may have also come to devalue cocaine through similar conditioning processes. However, though it may have been devalued, saccharin still functioned as a reinforcer. For example, at FR 12, rats in the Open oc group continued to work for saccharin, making approximately 250 lever presses per session for it (vs. about 100 for cocaine), even though they were not fluid deprived at any time. Similarly, rats in the OpenCoc group often chose saccharin over cocaine during the choice phase when the price of saccharin was low.
The results of the present experiment are consistent with and extend results of a growing number of studies investigating choice between cocaine and non-drug alternatives in rats (for reviews, see Ahmed, 2010, 2017). The treatment of the Closed group in the present experiment was most like that experienced by rats in other cocaine vs. saccharin choice studies (e.g., Lenoir et al., 2007). At equal prices for cocaine and saccharin, rats in the Closed group showed a high degree of preference (> 80% of choices) for saccharin – a result like that observed by Lenoir et al (2007). As saccharin price increased, however, preference switched to cocaine – an outcome consistent with that reported by Cantin et al. (2010). Though most rats show saccharin preference over cocaine at equal lever press requirements, a common finding in such choice studies is that a subset of rats prefer cocaine over saccharin. In the Closed group reported here, one out of eight subjects preferred cocaine over saccharin (when the FR was the same for both options). This is consistent with the observation from other studies that approximately 10–15% of rats prefer cocaine over saccharin (Ahmed, 2010, 2017). The present study adds to this literature by showing that the broader context of availability of the two choice alternatives can influence preference. For example, the OpenSac group showed greater preference for cocaine than the Closed group. Instead of observing 10–15% cocaine preferers in the OpenSac group, four out of nine subjects were cocaine preferers.
In the present experiment, price was manipulated by increasing the number of responses required on a ratio schedule. There are various other ways that price could have been manipulated. For example, Schwartz et al. (2016) arranged different prices by varying the amount of weight that subjects had to lift. Peck and Byrne (2016) used lever hold-down duration as the cost that was manipulated. In both studies, the exponential demand model described the resulting data just as well as it describes data from more typical studies using number of lever presses as the cost variable. It should be noted that on FR schedules, as number of lever presses required increases, so too does delay to reinforcer.A study investigating this question found that the delay to reinforcement inherent in ratio schedules is a key component of cost (Baumann, 1991). Though we could not compare delays to cocaine and saccharin reinforcement on ratio schedules here, as we did not record inter-reinforcer intervals, this could be an interesting question for future research.
The finding here and in previous cocaine vs. saccharin choice studies (e.g., Cantin et al., 2010) that rats increased their choice of cocaine when the price of saccharin was increased is consistent with the notion that cocaine is an economic substitute for saccharin in rats. The present results further suggest that economy type can influence substitutability between these reinforcers. Specifically, when saccharin was available after the choice session (in the OpenSac group), cocaine appeared to substitute more readily for saccharin (i.e., at lower saccharin prices) than when saccharin was not available after the choice session (i.e., in the Closed group). This suggests that for a non-drug alternative reinforcer to maintain its power to compete with cocaine for the allocation of behavior, the price of that non-drug alternative needs to be lower when it is available in an open economy than when it is available in a closed economy. (The results of the OpenCoc group suggest the same may be true when cocaine is available in an open economy.) Though the results described above are consistent with substitution effects, it should be noted that the discrete-trials choice procedure used here was probably not ideal for studying substitution or cross-price elasticity. In more typical studies of these phenomena (e.g., Carroll et al., 1991; Petry and Heyman, 1995), the two alternatives are available on concurrent ratio schedules for the whole session and subjects can freely respond on both schedules throughout the session. Our procedure only allowed rats to make one mutually exclusive choice between the two options every 10 minutes. This constraint may have caused rats to behave differently from how they would behave if they had access to both levers throughout the whole session as in the typical study of cross-price elasticity.
It has often been stated that drug use is driven by lack of access to alternative reinforcers. The results of the present study suggest a more nuanced picture. The main finding here was that rats were less willing to work to defend consumption of the non-drug alternative as its price increased when its economy was opened and it became more available. This translated into higher choice of cocaine. This suggests that, in the human situation, cocaine choice could be more likely when the available non-drug alternatives are abundant and easy to obtain than when they are scarcer. The results of the present study also suggest that, in contrast to the effect of making non-drug alternatives abundant, increased access to cocaine may not make demand for it more elastic. Such increased cocaine access may even cause a devaluation of non-drug reinforcers under certain conditions. Understanding drug-taking behavior will require consideration not only of which reinforcers are available to an individual but also the broader contexts (economy types) in which those reinforcers are available.
Highlights.
Opening the saccharin economy decreased saccharin’s essential value in rats.
This also increased preference of cocaine over saccharin in rats.
Opening the cocaine economy had no effect on demand for cocaine.
Saccharin had lower essential value when available in an open cocaine economy.
Economy type manipulations differently affect drug vs. non-drug reinforcers.
Acknowledgments
Role of the Funding Source
This research was supported by Award Number R01DA037269 from the National Institute on Drug Abuse (NIDA). The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institute on Drug Abuse or the National Institutes of Health. NIDA did not play a role in study design; in the collection, analysis and interpretation of data; in the writing of the report; or in the decision to submit the paper for publication.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Conflict of Interest
All authors declare that they have no conflicts of interest.
References
- Ahmed SH, Koob GF, 1998. Transition from moderate to excessive drug intake: Change in hedonic set point. Science 282, 298–300. [DOI] [PubMed] [Google Scholar]
- Ahmed SH, 2010. Validation crisis in animal models of drug addiction: Beyond non-disordered drug use toward drug addiction. Neurosci. Biobehav. Rev 35, 172–184. [DOI] [PubMed] [Google Scholar]
- Ahmed SH, 2017. Trying to make sense of rodents’ drug choice behavior. Prog. Neuropsychopharmacol. Biol. Psychiatry 87, 3–10. [DOI] [PubMed] [Google Scholar]
- Anderson KG, Woolverton WL, 2005. Effects of clomipramine on self-control choice in Lewis and Fischer 344 rats. Pharmacol. Biochem. Behav 80, 387–393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barbieri EJ, Ferko AP, DiGregorio GJ, Ruch EK, 1992. The presence of cocaine and benzoylecgonine in rat cerebrospinal fluid after the intravenous administration of cocaine. Life Sci 51, 1739–1746. [DOI] [PubMed] [Google Scholar]
- Bauman R, 1991. An experimental analysis of the cost of food in a closed economy. J. Exp. Anal.Behav 56,33–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bickel WK, Jarmolowicz DP, Mueller ET, Gatchalian KM, 2011. The behavioral economics and neuroeconomics of reinforcer pathologies: implications for etiology and treatment of addiction. Curr. Psych. Rep 13, 406–415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cantin L, Lenoir M, Augier E, Vanhille N, Dubreucq S, Serre F, Vouillac C, Ahmed SH, 2010. Cocaine is low on the value ladder of rats: Possible evidence for resilience to addiction. PLoS ONE 5, e11592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carroll ME, Carmona GG and May SA, 1991. Modifying drug‐reinforced behavior by altering the economic conditions of the drug and a nondrug reinforcer. J. Exp. Anal. Behav 56, 361–376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carroll ME, Cosgrove KP, Campbell UC, Morgan AD, Mickelberg JL, 2000. Reductions in ethanol, phencyclidine, and food-maintained behavior by naltrexone pretreatment in monkeys is enhanced by open economic conditions. Psychopharmacology 148, 412–422. [DOI] [PubMed] [Google Scholar]
- Christensen CJ, Silberberg A, Hursh SR,Huntsberry ME,Riley AL,2008aEssentialvalue of cocaine and food in rats: Tests of the exponential model of demand. Psychopharmacology 198, 221–229. [DOI] [PubMed] [Google Scholar]
- Christensen CJ, SilbeSrberg A, Hursh SR, Roma G, Riley AL, 2008b. Demand for cocaine and food over time. Pharmacol. Biochem. Behav 91, 209–216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deroche-Gamonet V, Belin D, Piazza PV, 2004. Evidence for addiction-like behavior in the rat. Science 305, 1014–1017. [DOI] [PubMed] [Google Scholar]
- Freeman KB, Green L, Myerson J, Woolverton WL, 2009. Delay discounting of saccharin in rhesus monkeys. Behav. Proc 82, 214–218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green JL, Dykstra LA, Carelli RM, 2015. Examination of cocaine dose in a preclinical model of natural reward devaluation by cocaine. Behav.Pharmacol 26, 398–402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grigson PS, 1997. Conditioned taste aversions and drugs of abuse: A reinterpretation. Behav. Neurosci 111, 129–136. [PubMed] [Google Scholar]
- Grigson PS, 2008. Reward Comparison: The Achilles’ heel and hope for addiction. Drug Discov. Today Dis. Models 5, 227–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grigson PS, Twining RC, 2002. Cocaine-induced suppression of saccharin intake: A model of drug-induced devaluation of natural rewards. Behav. Neurosci 116, 321–333. [PubMed] [Google Scholar]
- Heyman GM, 2009. Addiction: A disorder of choice Harvard University Press, Cambridge,MA. [Google Scholar]
- Hursh SR, 1980. Economic concepts for the analysis of behavior. J. Exp. Anal Behav 34, 219–238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hursh SR, 1991. Behavioral economics of drug self-administration and drug abuse policy. J. Exp. Anal. Behav 56, 377–393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hursh SR, 2014. Behavioral economics and the analysis of consumption and choice. In: McSweeney FK, Murphy ES (Eds.), The Wiley Blackwell Handbook of Operant and Classical Conditioning Wiley, Hoboken, NJ, pp. 275–305. [Google Scholar]
- Hursh SR, Raslear TG, Bauman R, Black H, 1989. The quantitative analysis of economic behavior with laboratory animals. In Grunert KG and Olander F (Eds.), Understanding Economic Behavior Kluwer Academic Publishers, Dordrecht, Netherlands, pp. 393–407. [Google Scholar]
- Hursh SR, Silberberg A, 2008. Economic demand and essential value. Psych. Rev 115, 186– 198. [DOI] [PubMed] [Google Scholar]
- Huskinson SL, Myerson J, Green L, Rowlett JK, Woolverton WL, Freeman KB, 2016. Shallow discounting of delayed cocaine by male rhesus monkeys when immediate food is the choice alternative. xp. Clinical Psychopharmacol 24, 456–463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kearns DN, Kim JS, unstall BJ, Silberberg A, 2017. Essential values of cocaine and non-drug alternatives predict the choice between them. Addict. Biol 22, 1501–1514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lenoir M, Serre F, Cantin L, Ahmed SH, 2007. Intense sweetness surpasses cocaine reward. PloS one 2, p. e698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitchell SH, De Wit H, Zacny J ., 1994. Effects of varying the” openness” of an economy on responding for cigarettes. Behav. Pharmacol 5, 159–166. [DOI] [PubMed] [Google Scholar]
- Mitchell SH, De Wit H, Zacny JP, 1998. The impact of three economic factors on cigarette procurement and consumption. Hum. Psychopharmacol 13, 259–266. [Google Scholar]
- Mitchell SH, Laurent CL, De Wit H, Zacny JP, 1995. Effects of price ‘openness’ of the economy and magnitude of the alternative reinforcer on responding for caffeinated coffee. Hum. Psychopharmacol 10, 39–46. [Google Scholar]
- Mook DG, Kushner BD, Kushner LR, 1981. Release of feeding by the sweet taste in rats: The specificity of oral satiety. Appetite 2, 267–280. [DOI] [PubMed] [Google Scholar]
- Nader MA, Woolverton WL, 1992. Choice between cocaine and food by rhesus monkeys: Effects of conditions of food availability. Behav. Pharmacol 3, 635–638. [PubMed] [Google Scholar]
- National Academy of Sciences, 2011. Guide for the Care and Use of Laboratory Animals National Academy Press, Washington, DC. [Google Scholar]
- Paterson NE, Markou A, 2003. Increased motivation for self-administered cocaine after escalated cocaine intake. Neuroreport 14, 2229–2232. [DOI] [PubMed] [Google Scholar]
- Peck S, Byrne T, 2016. Demand in rats responding under duration-based schedules of reinforcement. Behav. Proc 128, 47–52. [DOI] [PubMed] [Google Scholar]
- Petry NM, Heyman GM, 1995Behavioral economics of concurrent ethanol-sucrose and sucrose reinforcement in the rat: Effects of altering variable-ratio requirements. J. Exp. Anal. Behav 64, 331–359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pelloux Y, Everitt BJ, Dickinson A, 2007. Compulsive drug seeking by rats under punishment: ffects of drug taking history. Psychopharmacology 194, 127–137. [DOI] [PubMed] [Google Scholar]
- Rachlin H, 1992. Diminishing marginal value as delay discounting. J. Exp. Anal. Behav 57, 407–415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rachlin H, 1997. Four teleological theories of addiction. Psych. Bull. Rev 4, 462–473. [Google Scholar]
- Renwick AG, 1985. The disposition of saccharin in animals and man—a review. Food Chem.Toxicol 23, 429–435. [DOI] [PubMed] [Google Scholar]
- Schwartz LP, Kim JS, Silberberg A, Kearns DN, 2017. Heroin and saccharin demand and preference in rats. Drug Alcohol Depend 178, 87–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz LP, Silberberg A, Casey AH, Paukner A, Suomi SJ, 2016. Scaling reward value with demand curves versus preference tests. Anim. Cogn 19, 631–641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sweatman TW, Renwick AG, 1980. The tissue distribution and pharmacokinetics of saccharin in the rat. Toxicol. Appl. Pharmacol 55, 18–31. [DOI] [PubMed] [Google Scholar]
- Thomsen M, Caine SB, 2005. Chronic intravenous drug self-administration in rats and mice. Curr. Protocol Neurosci 9, 20. [DOI] [PubMed] [Google Scholar]
- Tunstall BJ, Kearns DN, 2014. Cocaine can generate a stronger conditioned reinforcer than food despite being a weaker primary reinforcer. Addict. Biol 21, 282–293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wee S, Specio SE, Koob GF, 2007. Effects of dose and session duration on cocaine self-administration in rats. J. Pharmacol. Exp. Ther 320, 1134–1143. [DOI] [PubMed] [Google Scholar]
- Wheeler RA, Aragona BJ, Fuhrmann KA, Jones JL, Day JJ, Cacciapaglia F, Wightman RM, Carelli RM,2011. Cocaine cues drive opposing context-dependent shifts in reward processing and emotional state. Biol. Psych 69, 1067–1074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheeler RA, Twining RC, Jones JL, Slater JM, Grigson PS, Carelli RM, 2008. Behavioral and electrophysiological indices of negative affect predict cocaine self-administration. Neuron 57, 774–785. [DOI] [PubMed] [Google Scholar]