

Bovine babesiosis is a serious threat to the cattle industry. We prepared blood DNA samples from 13 cattle with clinical babesiosis from the Badulla (n = 8), Jaffna (n = 3), and Kilinochchi (n = 2) districts in Sri Lanka.
KEYWORDS: Babesia, cattle, clinical babesiosis, Sri Lanka
ABSTRACT
Bovine babesiosis is a serious threat to the cattle industry. We prepared blood DNA samples from 13 cattle with clinical babesiosis from the Badulla (n = 8), Jaffna (n = 3), and Kilinochchi (n = 2) districts in Sri Lanka. These DNA samples tested positive in PCR assays specific for Babesia bovis (n = 9), Babesia bigemina (n = 9), and Babesia ovata (n = 1). Twelve cattle were positive for B. bovis and/or B. bigemina. One cow was negative for the tested Babesia species but was positive for Babesia on microscopic examination; the phylogenetic positions of 18S rRNA and cytochrome oxidase subunit III gene sequences suggested that the cow was infected with Babesia sp. Mymensingh, which was recently reported from a healthy cow in Bangladesh. We then developed a novel Babesia sp. Mymensingh-specific PCR assay and obtained positive results for one other sample. Analysis of gene sequences from the cow with positive B. ovata-specific PCR results demonstrated that the animal was infected not with B. ovata but with Babesia sp. Hue-1, which was recently reported from asymptomatic cattle in Vietnam. The virulence of Babesia sp. Hue-1 is unclear, as the cow was coinfected with B. bovis and B. bigemina. However, Babesia sp. Mymensingh probably causes severe clinical babesiosis, as it was the sole Babesia species detected in a clinical case. The present study revealed the presence of two bovine Babesia species not previously reported in Sri Lanka, plus the first case of severe bovine babesiosis caused by a Babesia species other than B. bovis, B. bigemina, and Babesia divergens.
INTRODUCTION
Clinical bovine babesiosis is mainly caused by Babesia bovis and Babesia bigemina in the tropics and subtropics (1), while Babesia divergens causes clinical bovine babesiosis in Europe (2). Babesia sporozoites injected by infected tick vectors directly invade the host's red blood cells (RBCs) and develop into merozoites (3, 4). Merozoites egress from the infected RBCs, causing massive intravascular hemolysis that leads to clinical signs such as fever, anemia, hemoglobinuria, and jaundice (1). Additionally, bovine babesiosis caused by B. bovis is characterized by neurological and respiratory syndromes because of the cytoadherence of infected RBCs in capillary beds of vital organs such as the brain and lungs (1, 5). Early treatment with antibabesial agents is essential for recovery, and a lack of or delay in treatment may result in severe babesiosis that causes death (6, 7). Thus, bovine babesiosis results in huge economic losses to the cattle industry because of treatment and tick control costs, production losses, and animal mortality (8). Moreover, babesiosis disrupts international cattle trade, as World Organisation for Animal Health (OIE) regulations stipulate rules for exporting cattle from countries in which bovine babesiosis is endemic (9).
Disease development in infected cattle is affected by multiple factors, including age, management practices, immunity, and breed (1). Animals less than 9 months old are usually resistant to clinical babesiosis, while adults are not (10, 11). The immunity acquired by young animals protects them from developing clinical babesiosis when they become infected as adults. In addition, Bos indicus is relatively resistant to clinical babesiosis compared with Bos taurus (12).
Cattle in several countries of endemicity are immunized against B. bovis and B. bigemina via live attenuated vaccines (13). The attenuation of B. bovis and B. bigemina is performed by serially passaging the organisms in splenectomized calves (1). However, the global use of live vaccines is limited due to the expense and time required to produce the vaccines, vaccine breakthrough due to strain variations, and risk of contamination with other bovine blood pathogens (14, 15).
In addition to B. bovis, B. bigemina, and B. divergens, several other species of Babesia are known to infect cattle, such as B. ovata (16), B. occultans (17), B. major (18), and several unclassified species, including Babesia sp. Oshima (19), Babesia sp. Kashi (20), Babesia sp. Hue-1 (21), Babesia sp. Mymensingh (22), and Babesia species isolated in South Africa (23, 24). Most of these Babesia species have low pathogenicity; however, the virulence of the remaining species is unknown, as they have been reported only from apparently healthy animals (21, 22). To determine the virulence of the Babesia species with unknown pathogenicity, there is a need for experimental infections or investigations to identify the Babesia species present in cattle with clinical babesiosis.
Clinical babesiosis is common among cattle in the tropical country of Sri Lanka. A few decades ago, clinical cases of bovine babesiosis were concentrated mainly in the wet zone of Sri Lanka, whereas the disease was uncommon in the dry zone (25). This discrepancy was due to the variation in cattle breeds and management practices between the wet and dry zones (12, 26); most cattle in the wet zone were temperate breeds (Bos taurus) that were managed intensively, while the dry zone contained mostly extensively managed local cattle (Bos indicus) (27). However, farmers in the dry zone have recently started to maintain temperate breeds and their crosses and to adopt an intensive management system, leading to an increased incidence of clinical babesiosis. Hence, clinical babesiosis in cattle is now common in both the wet and dry zones (28). Although live vaccines against both B. bovis and B. bigemina are available in Sri Lanka, only a few hundred calves are vaccinated annually (29, 30).
A recent series of molecular epidemiological surveys was conducted in Sri Lanka to detect and genetically characterize hemoprotozoan parasites in apparently healthy cattle (31–35). In contrast, genetic techniques have never been used to investigate clinical cases of bovine babesiosis in Sri Lanka. Therefore, in the present study, we employed molecular tools to identify the Babesia species present in cattle with clinical babesiosis in Sri Lanka.
MATERIALS AND METHODS
Animals and blood samples.
Between June and December 2017, approximately 2-ml samples of whole blood were obtained from the jugular veins of 13 cattle with high fever and hemoglobinuria in the Badulla (n = 8), Jaffna (n = 3), and Kilinochchi (n = 2) districts of Sri Lanka (see Fig. S1 in the supplemental material) using Vacutainer tubes containing EDTA (NIPRO, Osaka, Japan) (Table 1). The Jaffna and Kilinochchi districts are located in the dry zone, while the sampling locations within the Badulla district were located in the wet zone. The affected animals were either Friesians, Jerseys, or Jersey and Sahiwal crosses, and their ages ranged from 3 to 6 years. None of the affected animals had been vaccinated against B. bovis and/or B. bigemina. Immediately after sampling, the animals were treated with diminazene aceturate and long-acting oxytetracycline. Blood samples were analyzed in a commercial laboratory to determine the hemoglobin concentration (HGB), hematocrit (HCT), and RBC count. DNA samples were extracted from blood samples using a commercial kit (QIAamp DNA Blood minikit; Qiagen, Hilden, Germany) in accordance with the manufacturer's instructions and stored at −30°C until use. All animal procedures were approved by the Animal Care and Use Committee of Obihiro University of Agriculture and Veterinary Medicine, Japan (approval number 29-53).
TABLE 1.
Clinical presentation of bovine babesiosis in Sri Lanka
| Cow no. | District | Animal IDa | Breed | Age (yrs) | Clinical signs |
RBC indicesb |
|||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Temp (°C) | Anemia | Hemoglobinuria | Nervous signs | HGB | HCT | RBC | |||||
| 1 | Badulla | I | Jersey | 4 | 40.6 | Y | Y | Y | NT | NT | NT |
| 2 | Badulla | L1 | Friesian | 3 | 41.4 | N | Y | N | 8.4 | 22.6 | 5.65 |
| 3 | Badulla | L2 | Jersey | 4 | 41.1 | N | Y | N | 9.6 | 27.6 | 5.94 |
| 4 | Badulla | R | Friesian | 4 | 41.1 | Y | Y | N | 6.8 | 19.6 | 3.94 |
| 5 | Badulla | Sc | Jersey | 5 | 39.4 | Y | Y | Y | 2.8 | 7.9 | 1.38 |
| 6 | Badulla | T1 | Jersey | 4 | 41.1 | Y | Y | Y | 8.7 | 24 | 5.59 |
| 7 | Badulla | T2 | Jersey | 4 | 41.1 | N | Y | N | 9.4 | 25.7 | 5.51 |
| 8 | Badulla | V | Jersey | 5 | 42.2 | Y | Y | N | NT | NT | NT |
| 9 | Jaffna | C2 | Jersey | 4 | 40.6 | Y | Y | N | 6.8 | 21.9 | 3.88 |
| 10 | Jaffna | C5 | Jersey | 5 | 40 | Y | Y | Y | 6.7 | 21.5 | 3.5 |
| 11 | Kilinochchi | C6c | Jersey × Sahiwal | 6 | 39.4 | Y | Y | N | 2.83 | 10.6 | 2.22 |
| 12 | Kilinochchi | C7c | Jersey | 3 | 41.1 | Y | Y | Y | 4.8 | 13.3 | 3.03 |
| 13 | Jaffna | C8 | Jersey × Sahiwal | 6 | 40 | Y | Y | N | NT | NT | NT |
All animals were female. ID, identification; RBC, red blood cell; HGB, hemoglobin; HCT, hematocrit; F, female; Y, yes (observed); N, no (not observed); NT, not tested.
RBC indices, including the HGB concentration (grams per deciliter), HCT (percent), and RBC count (×106 per microliter), were measured for 10 of the 13 animals.
This animal died despite treatment with diminazene aceturate.
Diagnostic PCR assays.
A previously described PCR assay based on the rhoptry-associated protein 1 gene (36) was used to detect B. bovis, while apical membrane antigen 1 (ama-1) gene-based PCR assays were employed to detect B. bigemina (37) and B. ovata (38). All parasite species were detected by single-step PCR, although B. bovis- and B. bigemina-specific PCR assays were originally described as nested PCR assays. The list of primer sequences used in the present study is provided in Table S1.
Cloning and sequencing.
An amplicon from the ama-1 PCR assay targeting B. ovata was cloned and sequenced as previously described (21), as this Babesia species has not been reported in Sri Lanka. In addition, the 18S rRNA and cytochrome oxidase subunit III (cox3) gene sequences of babesial origin from a DNA sample that was PCR positive for B. ovata and from a sample that was PCR negative for all three Babesia species tested in the present study were amplified by PCR using sets of common primers. Briefly, a 25-μl reaction mixture containing 1 μl of DNA, 1× PCR buffer (Toyobo, Osaka, Japan), 400 μM each deoxynucleoside triphosphate (dNTP; Toyobo), 0.4 μM each forward primer (18S rRNA, 5′-CATTACAACAGTTATAGTTTCTTTGG-3′ [21]; cox3, 5′-TCAACAAAATGCCAATATGTTCCAA-3′) and reverse primer (18S rRNA, 5′-CTAGGCATTCCTCGTTCATGATTTAG-3′; cox3, 5′-TACAAAGTGCATCTTTGGGAGAAG-3′), 0.5 μl of 1-U/μl KOD FX Neo DNA polymerase (Toyobo), and 4 μl of distilled water was subjected to an initial predenaturation step at 94°C for 2 min and then to 35 cycles of denaturation at 94°C for 10 s, annealing at 58°C (18S rRNA) or 64°C (cox3) for 30 s, and extension at 68°C for 90 s. The PCR amplicons were cloned into a PCR 2.1 plasmid vector (TOPO, Invitrogen, Carlsbad, CA) and then sequenced as previously described (37).
Development of a PCR assay specific to Babesia sp. Mymensingh.
One animal that produced negative results in the PCR assays for B. bovis, B. bigemina, and B. ovata was determined to be infected with Babesia sp. Mymensingh (22) based on an 18S rRNA sequence. A pair of common forward (5′-TGGACCAGGTACATGATCAAGT-3′) and reverse (5′-AATCATCGTGCTGACGACCCTTC-3′) PCR primers (37) was used to amplify the 1,372-bp ama-1 gene fragment from Babesia sp. Mymensingh, as described for 18S rRNA or cox3 amplification, except that the annealing temperature was changed to 62°C. The amplicon was cloned and sequenced. The newly generated ama-1 gene sequence and those already available in the NCBI GenBank database were subjected to multiple alignment, and a set of forward (5′-TGGCGCCGACTTCCTGGAGCCCATCTCCAA-3′) and reverse (5′-AGCTGGGGCCCTCCTTCGATGAACCGTCGG-3′) primers specific to Babesia sp. Mymensingh was designed. A 10-μl PCR mixture containing 1 μl of DNA, 1× PCR buffer (Applied Biosystems, Branchburg, NJ), 200 μM each dNTP (Applied Biosystems), 0.5 μM each forward and reverse primers, 0.1 μl of 5-U/μl AmpliTaq Gold DNA polymerase (Applied Biosystems), and 5.9 μl of distilled water was subjected to initial enzyme activation at 95°C for 5 min, followed by 40 cycles of denaturing at 95°C for 30 s, annealing at 64°C for 30 s, and extension at 72°C for 30 s. After a final elongation step at 72°C for 7 min, the PCR products were resolved by gel electrophoresis and then visualized under UV illumination. Detection of an approximately 371-bp band was considered positive. The specificity of the PCR assay was evaluated using a panel of DNA samples derived from several bovine blood pathogens and uninfected cattle (38, 39). The Babesia sp. Mymensingh-specific PCR assay was then used to screen all 13 cattle DNA samples analyzed in the present study.
Sequencing and phylogenetic analyses.
The identity scores among gene sequences were determined by the EMBOSS NEEDLE online program (http://www.bioinformatics.nl/cgi-bin/emboss/needle). The 18S rRNA, cox3, and ama-1 gene sequences determined in the present study and those retrieved from GenBank were aligned using the MAFFT software program (40). MEGA software (41) was then used to construct three separate maximum likelihood phylogenetic trees based on Tamura-Nei (18S rRNA and cox3) (42) or General Time Reversible (ama-1) (43) substitution models.
Accession number(s).
The gene sequences determined in the present study were registered with GenBank under the accession numbers LC385886 to LC385894.
RESULTS
A total of 13 cattle with high fever and hemoglobinuria were sampled in three districts: Badulla, Kilinochchi, and Jaffna. Clinical examination determined that five of the eight animals sampled in the Badulla district were anemic, whereas all five of the animals sampled in the Kilinochchi and Jaffna districts were anemic (Table 1). Five of the affected animals also showed nervous system abnormalities, such as incoordination. In the anemic animals, except cow T1, all three RBC indices, including HGB concentration, HCT, and RBC count, were below the lower limits of the reference ranges for these parameters in healthy cattle (8 g/dl, 24%, and 5 × 106/μl, respectively) (44). Two animals in the Kilinochchi district (C6 and C7) and one animal in the Badulla district (S) with very low HGB concentrations, HCT, and RBC counts died despite treatment with diminazene aceturate and oxytetracycline, while the remaining animals recovered following treatment. PCR assays revealed that 12 of the 13 DNA samples were positive for B. bovis and/or B. bigemina (Table 2). B. bovis and B. bigemina were each detected in nine animals, while six animals were positive for both Babesia species. The result of the B. ovata-specific ama-1 PCR assay was positive for only one animal (C8 in the Jaffna district).
TABLE 2.
PCR detection of B. bovis, B. bigemina, and B. ovata in clinical cases of bovine babesiosis in Sri Lanka
| Cow no. | Animal ID | Babesia bovis | Babesia bigemina | Babesia ovata |
|---|---|---|---|---|
| 1 | I | + | − | − |
| 2 | L1 | − | + | − |
| 3 | L2 | − | + | − |
| 4 | Ra | − | − | − |
| 5 | S | + | + | − |
| 6 | T1 | + | + | − |
| 7 | T2 | + | + | − |
| 8 | V | + | + | − |
| 9 | C2 | − | + | − |
| 10 | C5 | + | − | − |
| 11 | C6 | + | − | − |
| 12 | C7 | + | + | − |
| 13 | C8 | + | + | + |
| Total | 9 | 9 | 1 |
Animal R was negative for all three Babesia species tested in the present study.
Cow R from the Badulla district was PCR negative for all three Babesia species tested, but microscopic examination revealed Babesia piroplasms in a thin blood smear prepared from this animal (Fig. 1). The length and width of the paired pyriforms were 2.25 to 3.04 μm and 1.58 to 2.20 μm, respectively, whereas the ring forms were 1.52 to 1.97 μm in diameter. The paired pyriforms formed an obtuse angle that reached 180° in some cases. Single forms were often elongated or irregularly shaped. 18S rRNA is the most commonly used molecular marker for species identification of eukaryotes, including Babesia (18, 20, 45–47). Therefore, in an attempt to identify the species of Babesia detected in cow R, a 1,385-bp 18S rRNA sequence was isolated from cow R (GenBank accession number LC385886) and compared with the Babesia sequences in the GenBank database. The analyzed gene sequence shared 99.6% identity with the Babesia sp. Mymensingh sequence (GenBank accession number MF576177) that was recently reported from an asymptomatic cow in Bangladesh (22), followed by 97.8% identity with a B. bigemina sequence (GenBank accession number FJ426361) (48); this confirmed that the Babesia species detected in cow R was Babesia sp. Mymensingh. In the 18S rRNA phylogeny based on a 1,310-bp alignment with no gaps, the Babesia sp. Mymensingh sequence determined in the present study clustered together with that previously reported in Bangladesh (22) and formed a sister clade to B. bigemina (Fig. 2).
FIG 1.

Morphology of the Babesia parasites in cow R. A Giemsa-stained thin blood smear prepared from cow R was observed under a light microscope. Paired pyriforms (A), ring forms (B), and elongated and irregularly shaped single forms (C) of Babesia parasites were detected in cow R.
FIG 2.

Phylogeny of 18S rRNA. A maximum likelihood phylogeny was constructed using 18S rRNA sequences determined in the present study and those retrieved from GenBank. The gene sequences determined in the present study are indicated by boldface type. Note that the Babesia sp. Mymensingh sequence determined in the present study and that previously reported from Bangladesh clustered together and formed a sister clade to B. bigemina. Additionally, Babesia sp. Hue-1 occurred separately, forming a sister clade to the common ancestor of Babesia sp. Mymensingh and B. bigemina.
As B. ovata has not been previously reported in Sri Lanka, the amplicon of the PCR assay targeting B. ovata from cow C8 was cloned and sequenced. The resultant ama-1 gene fragment (GenBank accession number LC385892) shared only 93.3% identity with the B. ovata sequence (GenBank accession number AB634843) (37), while the sequence shared 97.4% to 98.4% identity scores with ama-1 sequences (GenBank accession numbers LC125412 to LC125415) from recently reported Babesia sp. Hue-1 in Vietnam (21). The 18S rRNA amplified from cow C8 included B. bigemina sequences (data not shown), as well as a 1,381-bp sequence (GenBank accession number LC385887) that shared a high identity score (99.0%) with a Babesia sp. Hue-1 sequence (GenBank accession number LC125456) (21). These findings suggest that cow C8 was infected with Babesia sp. Hue-1, as the identity scores shared between the 18S rRNA and ama-1 gene sequences from cow C8 and Babesia sp. Hue-1 are comparable to the identity scores shared among these sequences from different isolates of other bovine Babesia species (49–52). Phylogenetically, Babesia sp. Hue-1 from cow C8 formed a sister clade to the common ancestor of the B. bigemina clade and the Babesia sp. Mymensingh clade (Fig. 2). Moreover, in a phylogeny based on a short alignment (635 bp) of 18S rRNA, Babesia sp. Hue-1 sequences from cow C8 in Sri Lanka and that reported in Vietnam occurred together and formed a sister clade to the clade formed by sequences from Babesia species reported in China (GenBank accession number AY603403) (18) and Korea (GenBank accession number AY081192) (53) (Fig. S2).
In addition to 18S rRNA, phylogenies based on mitochondrial genes are widely used for the identification of species, including Babesia species (54–57). Therefore, to further investigate the phylogenetic positions of Babesia sp. Mymensingh and Babesia sp. Hue-1, cox3 gene sequences (556 bp) were amplified from DNA samples that were positive for these Babesia species (from cows R and C8, respectively). The cox3 sequence from Babesia sp. Mymensingh (GenBank accession number LC385889) shared 93.5% identity with B. bigemina (GenBank accession number LK054939) (58) and B. ovata (GenBank accession number LC146482) (52) sequences and formed a phylogenetic sister clade to B. bigemina (Fig. 3). Only the cox3 sequence from Babesia sp. Hue-1 (GenBank accession number LC385890) was isolated from cow C8. The cox3 sequence from Babesia sp. Hue-1 shared 94.9%, 94.1%, and 93.5% identity with sequences from an unidentified Babesia sp. (Wenchuan, China) (GenBank accession number JN859545) (56), B. ovata (GenBank accession number LC146482), and B. bigemina (GenBank accession number LK054939), respectively, and formed a sister clade to the clade formed by Babesia species reported in China (Wenchuan) and Korea (Fig. 3).
FIG 3.

Phylogeny of cox3. A maximum likelihood phylogeny was constructed using cox3 sequences from Babesia sp. Mymensingh and Babesia sp. Hue-1 and those available in the GenBank database. The gene sequences determined in the present study are indicated by boldface type. Note that Babesia sp. Mymensingh formed a sister clade to B. bigemina, whereas Babesia sp. Hue-1 formed a sister clade to Babesia sp. (Wenchuan, China)/Babesia sp. (Lushi, China).
The ama-1 gene is an attractive target for the development of species-specific PCR assays for detecting Babesia parasites, as the gene is conserved within a given Babesia species but is diverse between species (37, 59–61). Therefore, a 1,372-bp ama-1 gene fragment (GenBank accession number LC385893) was isolated from Babesia sp. Mymensingh to develop a specific PCR assay to detect this Babesia species in the DNA samples. The gene sequence shared 83.9% and 83.3% identity with B. bigemina (GenBank accession number AB481200) (62) and B. ovata (GenBank accession number AB634843) (37) sequences, respectively. The PCR assay developed based on the ama-1 gene amplified only Babesia sp. Mymensingh, while no amplicons were observed in DNA samples from several other bovine blood pathogens and cattle DNA, confirming its specificity (Fig. 4A). When this PCR assay was employed to screen all 13 cattle DNA samples, Babesia sp. Mymensingh was detected not only in cow R but also in cow V (Fig. 4B). PCR amplicons from cows R and V were cloned and sequenced. The 371-bp ama-1 sequences from cows R and V (GenBank accession number LC385894) were identical to each other and to the long ama-1 gene fragment initially isolated from cow R (GenBank accession number LC385893). Phylogenetic trees based on the ama-1 gene were used to investigate the evolutionary relationships of apicomplexan parasites, such as species of Plasmodium and Babesia (61, 63–65). In a phylogenetic construction used to investigate the positions of Babesia sp. Mymensingh and Babesia sp. Hue-1, the ama-1 sequences of Babesia sp. Mymensingh occurred distant to the sequences of B. bigemina (Fig. 5). In addition, the Babesia sp. Hue-1 ama-1 sequence generated in the present study clustered with those previously reported in Vietnam and formed a sister clade to B. ovata. To further confirm that cow V was infected with Babesia sp. Mymensingh, 18S rRNA and cox3 sequences were amplified, cloned, and sequenced. The 18S rRNA sequence (GenBank accession number LC385888) from cow V shared 99.6% identity with the Babesia sp. Mymensingh sequence from cow R (GenBank accession number LC385886), whereas the cox3 sequence from cow V (GenBank accession number LC385891) was identical to that from cow R (GenBank accession number LC385889), confirming that cow V was also infected with Babesia sp. Mymensingh.
FIG 4.
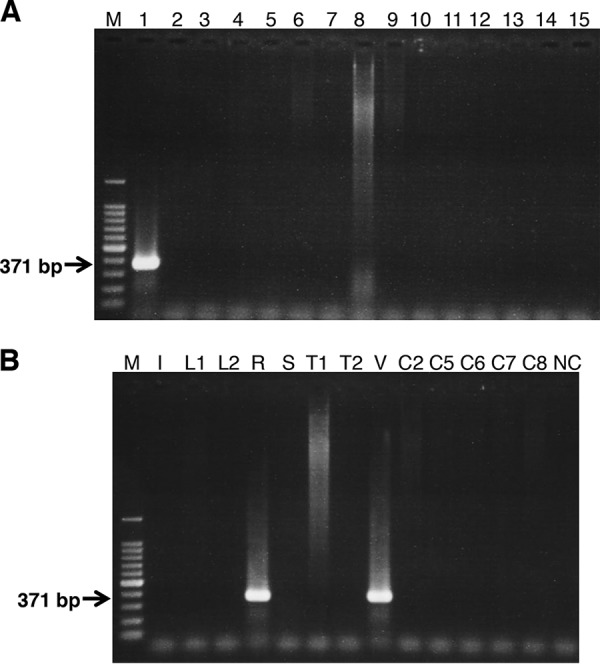
A PCR assay specific to Babesia sp. Mymensingh was developed. (A) Specificity testing. The specificity of the newly developed PCR assay was tested using DNA samples from Babesia sp. Mymensingh, B. bigemina, B. bovis, B. ovata, B. divergens, Babesia sp. Hue-1, Theileria annulata, Theileria parva, Th. orientalis, Trypanosoma evansi, Trypanosoma theileri, Tr. brucei, Anaplasma marginale, Anaplamsa bovis, and uninfected cattle (lanes 1 to 15, respectively). Lane M, 100-bp DNA marker. Note that the amplicon with the expected size was observed only with Babesia sp. Mymensingh. (B) Screening of 13 clinical samples for Babesia sp. Mymensingh. The PCR assay specific to Babesia sp. Mymensingh was used to screen DNA samples from 13 clinical cases. Lanes M and NC contained the 100-bp DNA marker and nontemplate control, respectively. Note that Babesia sp. Mymensingh was also detected in cow V.
FIG 5.

Phylogeny of ama-1. The ama-1 sequences from Babesia sp. Mymensingh from cows R and V (amplified by Babesia sp. Mymensingh-specific PCR) and from Babesia sp. Hue-1 from cow C8 (amplified by B. ovata PCR) together with those retrieved from GenBank were used to construct a maximum likelihood phylogeny. The gene sequences determined in the present study are indicated by boldface type. Note that the Babesia sp. Hue-1 sequences clustered with those previously reported in Vietnam and formed a sister clade to B. ovata, while the Babesia sp. Mymensingh formed a separate clade.
DISCUSSION
Various species of Babesia infect a wide range of host species worldwide, including wild and domestic animals, humans, birds, and reptiles (66). The Babesia species that infect cattle are of great economic importance, as they cause severe clinical diseases leading to significant production losses (1). Among them, B. bovis, B. bigemina, and B. divergens are highly virulent species that cause a severe form of bovine babesiosis. Although bovine babesiosis is very common in Sri Lanka, a detailed examination of clinical cases using molecular techniques has never been carried out in this country. Therefore, in the present study, we used molecular diagnostic tools to investigate clinical babesiosis in Sri Lanka.
Anemia was detected in 10 of the 13 animals with fever and hemoglobinuria investigated in the present study, while 3 animals from the Badulla district were not anemic at the time of sampling, suggesting that these animals were sampled in the early stage of disease development. Three animals with very low RBC indices died even after treatment with an antibabesial drug (diminazene aceturate), suggesting that early veterinary intervention is of paramount importance in bovine babesiosis (7). The neurological signs observed in five animals might have been due to B. bovis infection, which can cause cerebral babesiosis in cattle (1, 5). However, such neurological signs can also occur in anemic animals due to hypoxia-related brain injury caused by low hemoglobin levels (67, 68). Postmortem examination could have clarified whether the actual cause of the neurological signs in cows S and C7 was cerebral babesiosis or hypoxic brain injury. Unfortunately, however, postmortem examinations were not carried out for any of the animals.
The PCR assays and sequencing analyses detected B. bovis and B. bigemina, as well as two other Babesia species (Babesia sp. Mymensingh and Babesia sp. Hue-1) that had not been previously reported in Sri Lanka. In addition to Babesia parasite species, previously described species-specific PCR assays (69–72) determined that the samples contained Theileria annulata, Theileria orientalis, Trypanosoma theileri, and Anaplasma marginale (Tables S1 and S2). However, the involvement of these parasite species in the animals' clinical disease was unclear, as none of these parasite species induce hemoglobinuria.
Based on morphological observation, Babesia sp. Mymensingh can be classified as a large Babesia organism. However, the piroplasms were morphologically different from those of B. bigemina, as the paired pyriforms of the latter usually form an acute angle (73). In addition, the size of the ring forms for Babesia sp. Mymensingh was smaller than that for B. bigemina. Babesia sp. Mymensingh was initially identified based on an 18S rRNA sequence from an apparently asymptomatic cow in Bangladesh (22). In contrast, the parasite in the present study was detected as the sole Babesia species in a cow (R) with typical signs of clinical babesiosis, suggesting that Babesia sp. Mymensingh is a virulent species.
The phylogenetic position of Babesia sp. Mymensingh was further analyzed in a cox3-based phylogeny, in which it formed a sister clade to B. bigemina. The sister clades formed in 18S rRNA and cox3 phylogenies may identify Babesia sp. Mymensingh as a new genotype of B. bigemina. However, the low identity scores shared between the 18S rRNA, cox3, and ama-1 gene sequences from these parasite species, the ama-1 phylogeny in which Babesia sp. Mymensingh formed a separate clade, and the morphological differences confirm that Babesia sp. Mymensingh is a distinct Babesia species. The PCR detection of Babesia sp. Mymensingh in an additional cow suggested that Babesia sp. Mymensingh infection might be common among cattle in Sri Lanka. Babesia sp. Mymensingh may have a wide distribution, as this parasite species was also detected in Bangladesh. Therefore, the PCR assay developed in the present study will be a useful diagnostic tool for specific detection of Babesia sp. Mymensingh in different geographical territories.
Cow R, which was infected with Babesia sp. Mymensingh, was reexamined 1 week after sampling. No hemoglobinuria was present, and the anemia was milder than previously. Furthermore, laboratory examination showed improvements in the HGB concentration (8 g/dl), HCT (23.4%), and RBC count (4.53 × 106/μl) compared with the first sampling, indicating that diminazene aceturate is effective against Babesia sp. Mymensingh. However, Babesia sp. Mymensingh might have implications for the immune control and diagnostics of bovine babesiosis. For example, live attenuated vaccines are used in some countries of endemicity, including Sri Lanka, to immunize cattle against bovine babesiosis caused by B. bovis and B. bigemina (13). If Babesia sp. Mymensingh is not immunologically cross-reactive to these parasite species, especially to B. bigemina, Babesia sp. Mymensingh may cause clinical disease in vaccinated animals. However, if they are immunologically cross-reactive parasites, the serodiagnostic techniques commonly used in epidemiological surveys might generate unreliable data.
The PCR assay targeting B. ovata detected Babesia sp. Hue-1. Phylogenetically, 18S rRNA from Babesia sp. Hue-1 formed a clade that was clearly separate from the other Babesia species, identifying Babesia sp. Hue-1 as a separate Babesia species. The morphology of Babesia sp. Hue-1 was not analyzed, as the blood smear from the infected cattle was unfit for microscopic examination. Even if high-quality blood smears had been available, microscopic detection might have been impossible, as the animal was coinfected with B. bovis and B. bigemina. Babesia sp. Hue-1 was recently reported in Vietnam based on 18S rRNA and ama-1 sequences from healthy cattle (21). Therefore, the clinical significance of Babesia sp. Hue-1 remains unknown.
In conclusion, the present study demonstrated the presence of two Babesia species not previously detected in Sri Lanka and also identified severe clinical bovine babesiosis caused by a Babesia species other than B. bovis, B. bigemina, and B. divergens. Priorities in Babesia research include isolation of the newly detected Babesia species, experiments to investigate their virulence in different cattle breeds and immunological cross-reactivity with B. bovis and B. bigemina, and identification of specific tick vectors.
Supplementary Material
ACKNOWLEDGMENTS
We thank the owners and staff of the cattle farms involved in this study. We also thank the staff members at the Veterinary Research Institute, Peradeniya, Sri Lanka, and Hiroko Yamamoto (National Research Center for Protozoan Diseases, Obihiro University of Agriculture and Veterinary Medicine, Japan) for their excellent technical assistance.
This study was supported by grants from the Japan Society for Promotion of Science (JSPS) Grant-in-Aid for Scientific Research (JSPS KAKENHI numbers 26257417 and 16H05033), Open Partnership Joint Projects of the JSPS Bilateral Joint Research Projects, and AMED/JICA Science and Technology Research Partnership for Sustainable Development (SATREPS) project (grant number 17jm0110006h0005).
We declare no conflicts of interest associated with the present study.
Footnotes
Supplemental material for this article may be found at https://doi.org/10.1128/JCM.00895-18.
REFERENCES
- 1.Bock R, Jackson L, de Vos A, Jorgensen W. 2004. Babesiosis of cattle. Parasitology 129(Suppl):S247–S269. doi: 10.1017/S0031182004005190. [DOI] [PubMed] [Google Scholar]
- 2.Zintl A, Mulcahy G, Skerrett HE, Taylor SM, Gray JS. 2003. Babesia divergens, a bovine blood parasite of veterinary and zoonotic importance. Clin Microbiol Rev 16:622–636. doi: 10.1128/CMR.16.4.622-636.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Homer MJ, Aguilar-Delfin I, Telford SR, Krause PJ, Persing DH. 2000. Babesiosis. Clin Microbiol Rev 13:451–469. doi: 10.1128/CMR.13.3.451-469.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hunfeld KP, Hildebrandt A, Gray JS. 2008. Babesiosis: recent insights into an ancient disease. Int J Parasitol 38:1219–1237. doi: 10.1016/j.ijpara.2008.03.001. [DOI] [PubMed] [Google Scholar]
- 5.Everitt JI, Shadduck JA, Steinkamp C, Clabaugh G. 1986. Experimental Babesia bovis infection in Holstein calves. Vet Pathol 23:556–562. doi: 10.1177/030098588602300503. [DOI] [PubMed] [Google Scholar]
- 6.Mosqueda J, Olvera-Ramirez A, Aguilar-Tipacamu G, Canto GJ. 2012. Current advances in detection and treatment of babesiosis. Curr Med Chem 19:1504–1518. doi: 10.2174/092986712799828355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Vial HJ, Gorenflot A. 2006. Chemotherapy against babesiosis. Vet Parasitol 138:147–160. doi: 10.1016/j.vetpar.2006.01.048. [DOI] [PubMed] [Google Scholar]
- 8.McLeod R, Kristjanson P. 1999. Economic impact of ticks and tick-borne diseases to livestock in Africa, Asia and Australia. Report to the International Livestock Research Institute, Nairobi, Kenya, July 1999. Australian Centre for International Agricultural Research, Canberra, Australia. [Google Scholar]
- 9.World Organisation for Animal Health. 2018. Bovine babesiosis. In Terrestrial animal health code, vol II, 27th ed World Organisation for Animal Health, Paris, France: http://www.rr-africa.oie.int/docspdf/en/Codes/en_csat-vol2.pdf. [Google Scholar]
- 10.Zintl A, Gray JS, Skerrett HE, Mulcahy G. 2005. Possible mechanisms underlying age-related resistance to bovine babesiosis. Parasite Immunol 27:115–120. doi: 10.1111/j.1365-3024.2005.00748.x. [DOI] [PubMed] [Google Scholar]
- 11.Goff WL, Johnson WC, Parish SM, Barrington GM, Tuo W, Valdez RA. 2001. The age-related immunity in cattle to Babesia bovis infection involves the rapid induction of interleukin-12, interferon-gamma and inducible nitric oxide synthase mRNA expression in the spleen. Parasite Immunol 23:463–471. doi: 10.1046/j.1365-3024.2001.00402.x. [DOI] [PubMed] [Google Scholar]
- 12.Bock RE, Kingston TG, de Vos AJ. 1999. Effect of breed of cattle on transmission rate and innate resistance to infection with Babesia bovis and B. bigemina transmitted by Boophilus microplus. Aust Vet J 77:461–464. [DOI] [PubMed] [Google Scholar]
- 13.Shkap V, de Vos AJ, Zweygarth E, Jongejan F. 2007. Attenuated vaccines for tropical theileriosis, babesiosis, and heartwater: the continuing necessity. Trends Parasitol 23:420–426. doi: 10.1016/j.pt.2007.07.003. [DOI] [PubMed] [Google Scholar]
- 14.de Vos AJ, Bock RE. 2000. Vaccination against bovine babesiosis. Ann N Y Acad Sci 916:540–545. doi: 10.1111/j.1749-6632.2000.tb05333.x. [DOI] [PubMed] [Google Scholar]
- 15.Bock RE, de Vos AJ, Kingston TG, Shiels IA, Dalgliesh RJ. 1992. Investigations of breakdowns in protection provided by living Babesia bovis vaccine. Vet Parasitol 43:45–56. doi: 10.1016/0304-4017(92)90047-D. [DOI] [PubMed] [Google Scholar]
- 16.Minami T, Ishihara T. 1980. Babesia ovata sp.n. isolated from cattle in Japan. Nat Inst Anim Health Q (Tokyo) 20:101–113. [PubMed] [Google Scholar]
- 17.Gray JS, de Vos AJ. 1981. Studies on a bovine Babesia transmitted by Hyalomma marginatum rufipes Koch, 1844. Onderstepoort J Vet Res 48:215–223. [PubMed] [Google Scholar]
- 18.Luo JX, Yin H, Guan GQ, Yang DY, Liu AH, Ma ML, Liu ZJ, Dang ZS, Bai Q, Lu WS, Chen PY. 2005. A comparison of small-subunit ribosomal RNA gene sequence of bovine Babesia species transmitted by Haemaphysalis spp. in China. Parasitol Res 95:145–149. doi: 10.1007/s00436-004-1268-4. [DOI] [PubMed] [Google Scholar]
- 19.Ohta M, Tsuji M, Tsuji N, Fujisaki K. 1995. Morphological, serological and antigenic characteristics, and protein profile of newly isolated Japanese bovine Babesia parasite with particular reference to those of B. ovata. J Vet Med Sci 57:671–675. [DOI] [PubMed] [Google Scholar]
- 20.Luo J, Yin H, Liu Z, Yang D, Guan G, Liu A, Ma M, Dang S, Lu B, Sun C, Bai Q, Lu W, Chen P. 2005. Molecular phylogenetic studies on an unnamed bovine Babesia sp. based on small subunit ribosomal RNA gene sequences. Vet Parasitol 133:1–6. doi: 10.1016/j.vetpar.2005.02.014. [DOI] [PubMed] [Google Scholar]
- 21.Weerasooriya G, Sivakumar T, Lan DT, Long PT, Takemae H, Igarashi I, Inoue N, Yokoyama N. 2016. Epidemiology of bovine hemoprotozoa parasites in cattle and water buffalo in Vietnam. J Vet Med Sci 78:1361–1367. doi: 10.1292/jvms.16-0099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Roy BC, Krücken J, Ahmed JS, Majumder S, Baumann MP, Clausen PH, Nijhof AM. 2018. Molecular identification of tick-borne pathogens infecting cattle in Mymensingh district of Bangladesh reveals emerging species of Anaplasma and Babesia. Transbound Emerg Dis 65:e231–e242. doi: 10.1111/tbed.12745. [DOI] [PubMed] [Google Scholar]
- 23.Thomas SE, Mason TE. 1981. Isolation and transmission of an unidentified Babesia sp. infective for cattle. Onderstepoort J Vet Res 48:155–158. [PubMed] [Google Scholar]
- 24.de Waal DT, Potgieter FT, Combrink MP, Mason TE. 1990. The isolation and transmission of an unidentified Babesia sp. to cattle by Hyalomma truncatum Koch 1844. Onderstepoort J Vet Res 57:229–232. [PubMed] [Google Scholar]
- 25.Jorgensen WK, Weilgama DJ, Navaratne M, Dalgliesh RJ. 1992. Prevalence of Babesia bovis and Anaplasma marginale at selected localities in Sri Lanka. Trop Anim Health Prod 24:9–14. doi: 10.1007/BF02357227. [DOI] [PubMed] [Google Scholar]
- 26.Regassa A, Penzhorn BL, Bryson NR. 2003. Attainment of endemic stability to Babesia bigemina in cattle on a South African ranch where non-intensive tick control was applied. Vet Parasitol 116:267–274. doi: 10.1016/j.vetpar.2003.07.011. [DOI] [PubMed] [Google Scholar]
- 27.Abeygunawardena H, Rathnayaka D, Jayathilaka WMAP. 1997. Characteristics of cattle farming systems in Sri Lanka. J Natl Sci Found Sri Lanka 25:25–38. doi: 10.4038/jnsfsr.v25i1.5016. [DOI] [Google Scholar]
- 28.Department of Animal Production and Health, Sri Lanka. 2012. Bovine babesiosis. Vet Epidemiol Bull Sri Lanka 5:1–3. [Google Scholar]
- 29.Weilgama DJ, Jorgensen WK, Dalgliesh RJ, Navaratne M, Weerasinghe C. 1989. Comparison between Sri Lankan and Australian strains of Babesia bovis in the vaccination of imported cattle in Sri Lanka. Trop Anim Health Prod 21:141–145. doi: 10.1007/BF02236195. [DOI] [PubMed] [Google Scholar]
- 30.Department of Animal Production and Health, Sri Lanka. 2015. Bovine babesiosis. Vet Epidemiol Bull Sri Lanka 8:3. [Google Scholar]
- 31.Sivakumar T, Kothalawala H, Abeyratne AS, Vimalakumar SC, Meewawe AS, Hadirampela DT, Puvirajan T, Sukumar S, Kuleswarakumar K, Chandrasiri ADN, Igarashi I, Yokoyama N. 2012. A PCR-based survey of selected Babesia and Theileria parasites in cattle in Sri Lanka. Vet Parasitol 190:263–267. doi: 10.1016/j.vetpar.2012.05.014. [DOI] [PubMed] [Google Scholar]
- 32.Sivakumar T, Yoshinari T, Igarashi I, Kothalawala H, Abeyratne AS, Vimalakumar SC, Meewawe AS, Kuleswarakumar K, Chandrasiri ADN, Yokoyama N. 2013. Genetic diversity within Theileria orientalis parasites detected in Sri Lankan cattle. Ticks Tick Borne Dis 4:235–241. doi: 10.1016/j.ttbdis.2012.11.009. [DOI] [PubMed] [Google Scholar]
- 33.Sivakumar T, Okubo K, Igarashi I, de Silva WK, Kothalawala H, Silva SSP, Vimalakumar SC, Meewewa AS, Yokoyama N. 2013. Genetic diversity of merozoite surface antigens in Babesia bovis detected from Sri Lankan cattle. Infect Genet Evol 19:134–140. doi: 10.1016/j.meegid.2013.07.001. [DOI] [PubMed] [Google Scholar]
- 34.Yokoyama N, Sivakumar T, Fukushi S, Tattiyapong M, Tuvshintulga B, Hayashida K, Kothalawala H, Silva SSP, Igarashi I, Inoue N. 2015. Genetic diversity in Trypanosoma theileri from Sri Lankan cattle and water buffaloes. Vet Parasitol 207:335–341. doi: 10.1016/j.vetpar.2014.12.006. [DOI] [PubMed] [Google Scholar]
- 35.Sivakumar T, Kothalawala H, Weerasooriya G, Silva SSP, Puvanendiran S, Munkhjargal T, Igarashi I, Yokoyama N. 2016. A longitudinal study of Babesia and Theileria infections in cattle in Sri Lanka. Vet Parasitol Reg Stud Rep 6:20–27. [DOI] [PubMed] [Google Scholar]
- 36.Figueroa JV, Chieves LP, Johnson GS, Buening GM. 1993. Multiplex polymerase chain reaction based assay for the detection of Babesia bigemina, Babesia bovis and Anaplasma marginale DNA in bovine blood. Vet Parasitol 50:69–81. doi: 10.1016/0304-4017(93)90008-B. [DOI] [PubMed] [Google Scholar]
- 37.Sivakumar T, Altangerel K, Battsetseg B, Battur B, AbouLaila M, Munkhjargal T, Yoshinari T, Yokoyama N, Igarashi I. 2012. Genetic detection of Babesia bigemina from Mongolian cattle using apical membrane antigen-1 gene based PCR technique. Vet Parasitol 187:17–22. doi: 10.1016/j.vetpar.2012.01.008. [DOI] [PubMed] [Google Scholar]
- 38.Sivakumar T, Tagawa M, Yoshinari T, Ybanez AP, Igarashi I, Ikehara Y, Hata H, Kondo S, Matsumoto K, Inokuma Yokoyama HN. 2012. PCR detection of Babesia ovata from cattle reared in Japan and clinical significance of coinfection with Theileria orientalis. J Clin Microbiol 50:2111–2113. doi: 10.1128/JCM.00220-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Shibata S, Sivakumar T, Igarashi I, Umemiya-Shirafuji R, Inokuma H, Fukumoto S, Yokoyama N. 2018. Epidemiological survey of a cervine Theileria in wild deer, questing ticks, and cattle in Hokkaido, Japan. Tick Tick Borne Dis 9:1235–1240. doi: 10.1016/j.ttbdis.2018.05.006. [DOI] [PubMed] [Google Scholar]
- 40.Katoh K, Misawa K, Kuma K, Miyata T. 2002. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res 30:3059–3066. doi: 10.1093/nar/gkf436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. 2013. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 30:2725–2729. doi: 10.1093/molbev/mst197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Tamura K, Nei M. 1993. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol Biol Evol 10:512–526. [DOI] [PubMed] [Google Scholar]
- 43.Nei M, Kumar S. 2000. Molecular evolution and phylogenetics. Oxford University Press, New York, NY. [Google Scholar]
- 44.Jain K. 1993. Essentials of veterinary hematology. Blackwell Publishing, Hoboken, NJ. [Google Scholar]
- 45.Greay TL, Zahedi A, Krige AS, Owens JM, Rees RL, Ryan UM, Oskam CL, Irwin PJ. 2018. Endemic, exotic and novel apicomplexan parasites detected during a national study of ticks from companion animals in Australia. Parasit Vectors 11:197. doi: 10.1186/s13071-018-2775-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Wu S, Xiong J, Yu Y. 2015. Taxonomic resolutions based on 18S rRNA genes: a case study of subclass Copepoda. PLoS One 10:e0131498. doi: 10.1371/journal.pone.0131498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ishaq SL, Wright A-DG. 2014. Design and validation of four new primers for next generation sequencing to target the 18S rRNA gene of gastrointestinal ciliate protozoa. Appl Environ Microbiol 80:5515–5521. doi: 10.1128/AEM.01644-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Criado-Fornelio A, Buling A, Asenzo G, Benitez D, Florin-Christensen M, Gonzalez-Oliva A, Henriques G, Silva M, Alongi A, Agnone A, Torina A, Madruga CR. 2009. Development of fluorogenic probe-based PCR assays for the detection and quantification of bovine piroplasmids. Vet Parasitol 162:200–206. doi: 10.1016/j.vetpar.2009.03.040. [DOI] [PubMed] [Google Scholar]
- 49.Byaruhanga C, Collins NE, Knobel D, Chaisi ME, Vorster I, Steyn HC, Oosthuizen MC. 2016. Molecular investigation of tick-borne haemoparasite infections among transhumant zebu cattle in Karamoja region, Uganda. Vet Parasitol Reg Stud Rep 3-4:27–35. [DOI] [PubMed] [Google Scholar]
- 50.Altay K, Aydin MF, Dumanli N, Aktas M. 2008. Molecular detection of Theileria and Babesia infections in cattle. Vet Parasitol 158:295–301. doi: 10.1016/j.vetpar.2008.09.025. [DOI] [PubMed] [Google Scholar]
- 51.Rittipornlertrak A, Nambooppha B, Simking P, Punyapornwithaya V, Tiwananthagorn S, Jittapalapong S, Chung YT, Sthitmatee N. 2017. Low levels of genetic diversity associated with evidence of negative selection on the Babesia bovis apical membrane antigen 1 from parasite populations in Thailand. Infect Genet Evol 54:447–454. doi: 10.1016/j.meegid.2017.08.009. [DOI] [PubMed] [Google Scholar]
- 52.Sivakumar T, Igarashi I, Yokoyama N. 2016. Babesia ovata: taxonomy, phylogeny and epidemiology. Vet Parasitol 229:99–106. doi: 10.1016/j.vetpar.2016.10.006. [DOI] [PubMed] [Google Scholar]
- 53.Cho SH, Kim TS, Lee HW, Tsuji M, Ishihara C, Kim JT, Wee SH, Lee CG. 2002. Identification of newly isolated Babesia parasites from cattle in Korea by using the Bo-RBC-SCID mice. Korean J Parasitol 40:33–40. doi: 10.3347/kjp.2002.40.1.33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Schreeg ME, Marr HS, Tarigo JL, Cohn LA, Bird DM, Scholl EH, Levy MG, Wiegmann BM, Birkenheuer AJ. 2016. Mitochondrial genome sequences and structures aid in the resolution of piroplasmida phylogeny. PLoS One 11:e0165702. doi: 10.1371/journal.pone.0165702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Templeton TJ, Asada M, Jiratanh M, Ishikawa SA, Tiawsirisup S, Sivakumar T, Namangala B, Takeda M, Mohkaew K, Ngamjituea S, Inoue N, Sugimoto C, Inagaki Y, Suzuki Y, Yokoyama N, Kaewthamasorn M, Kaneko O. 2016. Ungulate malaria parasites. Sci Rep 6:23230. doi: 10.1038/srep23230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Tian Z, Liu G, Yin H, Luo J, Guan G, Xie J, Luo J, Zheng J, Tian M, Yuan X, Wang F, Chen R, Wang H. 2013. Cytochrome c oxidase subunit III (COX3) gene, an informative marker for phylogenetic analysis and differentiation of Babesia species in China. Infect Genet Evol 18:13–17. doi: 10.1016/j.meegid.2013.04.002. [DOI] [PubMed] [Google Scholar]
- 57.Tian Z, Luo J, Zheng J, Xie J, Shen H, Yin H, Luo J, Tian M, Yuan X, Wang F, Liu G. 2013. Phylogenetic analysis of Babesia species in China based on cytochrome b (COB) gene. Infect Genet Evol 13:36–40. doi: 10.1016/j.meegid.2012.09.001. [DOI] [PubMed] [Google Scholar]
- 58.Jackson AP, Otto TD, Darby A, Ramaprasad A, Xia D, Echaide IE, Farber M, Gahlot S, Gamble J, Gupta D, Gupta Y, Jackson L, Malandrin L, Malas TB, Moussa E, Nair M, Reid AJ, Sanders M, Sharma J, Tracey A, Quail MA, Weir W, Wastling JM, Hall N, Willadsen P, Lingelbach K, Shiels B, Tait A, Berriman M, Allred DR, Pain A. 2014. The evolutionary dynamics of variant antigen genes in Babesia reveal a history of genomic innovation underlying host-parasite interaction. Nucleic Acids Res 42:7113–7131. doi: 10.1093/nar/gku322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Moreau E, Bonsergent C, Al Dybiat I, Gonzalez LM, Lobo CA, Montero E, Malandrin L. 2015. Babesia divergens apical membrane antigen-1 (BdAMA-1): a poorly polymorphic protein that induces a weak and late immune response. Exp Parasitol 155:40–45. doi: 10.1016/j.exppara.2015.04.024. [DOI] [PubMed] [Google Scholar]
- 60.Torina A, Agnone A, Sireci G, Mosqueda JJ, Blanda V, Albanese I, La Farina M, Cerrone A, Cusumano F, Caracappa S. 2010. Characterization of the apical membrane antigen-1 in Italian strains of Babesia bigemina. Transbound Emerg Dis 57:52–56. doi: 10.1111/j.1865-1682.2010.01118.x. [DOI] [PubMed] [Google Scholar]
- 61.Niu Q, Liu Z, Yu P, Yang J, Abdallah MO, Guan G, Liu G, Luo J, Yin H. 2015. Genetic characterization and molecular survey of Babesia bovis, Babesia bigemina and Babesia ovata in cattle, dairy cattle and yaks in China. Parasit Vectors 8:518. doi: 10.1186/s13071-015-1110-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.AbouLaila M, Yokoyama N, Igarashi I. 2009. Data from “Molecular cloning and phylogenitic analysis of Babesia bigemina apical membrane antigen-1 (AMA-1).” GenBank https://www.ncbi.nlm.nih.gov/nuccore/AB481200 (accession no. AB481200).
- 63.Parker ML, Penarete-Vargas DM, Hamilton PT, Guérin A, Dubey JP, Perlman SJ, Spano F, Lebrun M, Boulanger MJ. 2016. Dissecting the interface between apicomplexan parasite and host cell: insights from a divergent AMA-RON2 pair. Proc Natl Acad Sci U S A 113:398–403. doi: 10.1073/pnas.1515898113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Muehlenbein MP, Pacheco MA, Taylor JE, Prall SP, Ambu L, Nathan S, Alsisto S, Ramirez D, Escalante AA. 2015. Accelerated diversification of nonhuman primate malarias in Southeast Asia: adaptive radiation or geographic speciation? Mol Biol Evol 32:422–439. doi: 10.1093/molbev/msu310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lauron EJ, Oakgrove KS, Tell LA, Biskar K, Roy SW, Sehgal RN. 2014. Transcriptome sequencing and analysis of Plasmodium gallinaceum reveals polymorphisms and selection on the apical membrane antigen-1. Malar J 13:382. doi: 10.1186/1475-2875-13-382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Schnittger L, Rodriguez AE, Florin-Christensen M, Morrison DA. 2012. Babesia: a world emerging. Infect Genet Evol 12:1788–1809. doi: 10.1016/j.meegid.2012.07.004. [DOI] [PubMed] [Google Scholar]
- 67.Arciniegas DB. 2012. Hypoxic-ischemic brain injury. International Brain Injury Association, Alexandria, VA. http://www.internationalbrain.org/articles/hypoxicischemic-brain-injury/ Accessed 23 July 2018.
- 68.Hare GM, Tsui AK, McLaren AT, Ragoonanan TE, Yu J, Mazer CD. 2008. Anemia and cerebral outcomes: many questions, fewer answers. Anesth Analg 107:1356–1370. doi: 10.1213/ane.0b013e318184cfe9. [DOI] [PubMed] [Google Scholar]
- 69.Kirvar E, Ilhan T, Katzer F, Hooshmand-Rad P, Zweygarth E, Gerstenberg C, Phipps P, Brown CGD. 2000. Detection of Theileria annulata in cattle and vector ticks by PCR using the Tams1 gene sequences. Parasitology 120:245–254. doi: 10.1017/S0031182099005466. [DOI] [PubMed] [Google Scholar]
- 70.Ota N, Mizuno D, Kuboki N, Igarashi I, Nakamura Y, Yamashina H, Hanzaike T, Fujii K, Onoe S, Hata H, Kondo S, Matsui S, Koga M, Matsumoto K, Inokuma H, Yokoyama N. 2009. Epidemiological survey of Theileria orientalis infection in grazing cattle in the Eastern Part of Hokkaido, Japan. J Vet Med Sci 71:937–944. doi: 10.1292/jvms.71.937. [DOI] [PubMed] [Google Scholar]
- 71.Rodrigues AC, Garcia HA, Ortiz PA, Cortez AP, Martinkovic F, Paiva F, Batista JS, Minervino AH, Campaner M, Pral EM, Alfieri SC, Teixeira MM. 2010. Cysteine proteases of Trypanosoma (Megatrypanum) theileri: cathepsin L-like gene sequences as targets for phylogenetic analysis, genotyping diagnosis. Parasitol Int 59:318–325. doi: 10.1016/j.parint.2010.03.002. [DOI] [PubMed] [Google Scholar]
- 72.Ybañez AP, Sivakumar T, Battsetseg B, Battur B, Altangerel K, Matsumoto K, Yokoyama N, Inokuma H. 2013. Specific molecular detection and characterization of Anaplasma marginale in Mongolian cattle. J Vet Med Sci 75:399–406. doi: 10.1292/jvms.12-0361. [DOI] [PubMed] [Google Scholar]
- 73.Lempereur L, Beck R, Fonseca I, Marques C, Duarte A, Santos M, Zúquete S, Gomes J, Walder G, Domingos A, Antunes S, Baneth G, Silaghi C, Holman P, Zintl A. 2017. Guidelines for the detection of Babesia and Theileria parasites. Vector Borne Zoonotic Dis 17:51–65. doi: 10.1089/vbz.2016.1955. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.