

Abstract
The sirtuin family of nicotinamide adenine dinucleotide (NAD+)-dependent deacylases (SIRT1–7) are thought to be responsible, in large part, for the cardiometabolic benefits of lean diets and exercise and when upregulated can delay key aspects of aging. SIRT1, for example, protects against a decline in vascular endothelial function, metabolic syndrome, ischemia-reperfusion (IR) injury, obesity and cardiomyopathy, and SIRT3 is protective against dyslipidemia and IR injury. With increasing age, however, NAD+ levels and sirtuin activity steadily decrease and the decline is further exacerbated by obesity and sedentary lifestyles. Activation of sirtuins or NAD+ repletion induces angiogenesis, insulin sensitivity and other health benefits in a wide range of age-related cardiovascular and metabolic disease models. Human clinical trials testing agents that activate SIRT1 or boost NAD+ levels are in progress and show promise in their ability to improve the health of cardiovascular and metabolic disease patients.
Keywords: STACs, NAD+ booster, atherosclerosis, ischemia-reperfusion injury, obesity, metabolic syndrome, cardiomyopathy, dyslipidemia, insulin resistance, aging
1. Introduction
The lifespan extending abilities of the sirtuins were first discovered in yeast in the 1990s1,2 when the silent information regulator (SIR2) gene was shown to increase the replicative lifespan of yeast when upregulated2. Sir2 is a yeast gene silencing protein, which silences transcription at the HM mating-type loci in young yeast but re-localizes to the ribosomal DNA as cells age to prevent DNA damage that contributes to yeast aging3. Upregulating the expression of Sir2 reduces DNA damage and increases the lifespan of yeast2. Subsequently, Sir2 was shown to have histone deacetylase activity which requires nicotinamide adenine dinucleotide (NAD+)4,5 and sirtuins in mammals (SIRT1–7) were identified. Mammalian sirtuins were also shown to have important beneficial roles in aging, longevity and stress responses6–8. Increasing the activity of the sirtuins is associated with the delay of age-related diseases9–13 and, in some cases, increased longevity14–17.
Mammalian sirtuins are NAD+-dependent deacylases with a huge range of roles in transcription regulation, energy metabolism modulation, cell survival, DNA repair, inflammation and circadian rhythm regulation8,18. SIRT1, which is the most extensively studied sirtuin, is found in the nucleus and cytosol and along with histone deacetylation also modulates transcription factors such as p5319, nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB)20, FOXOs21 and peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC1α)22 and DNA repair proteins such as Poly ADP-ribose polymerase 1 (PARP1)23. SIRT2 is a cytosolic sirtuin, whilst SIRT3, 4, and 5 are located in the mitochondria and have roles in oxidative stress and lipid metabolism24–26. SIRT6 and 7 are nuclear sirtuins with roles in gene expression and DNA repair27,28. There is growing evidence that sirtuins, along with deacetylation, also have roles in catalysing other reactions such as demalonylation, desuccinylation and mono-ADP-ribosylation29,30, collectively termed deacylase reactions.
The activity of SIRT1 can be increased with pharmaceuticals known as sirtuin activating compounds (STACs). One of the first STACs, resveratrol, was discovered in a screen for molecules that increased the activity of human SIRT1 and extended the lifespan of yeast31. Since then there have been several generations of STACs, with increasing potency and specificity, including SRT1720 and SRT21046,9,32. STACs act as allosteric activators of SIRT1, binding to the STAC-binding domain in the N terminus and, via a ‘bend-at-the-elbow’ model, increasing the binding affinity of a substrate for SIRT133,34. Whether STACs activate sirtuins other than SIRT1, and the mechanism of this activation, is still not completely understood35. In addition to SIRT1, resveratrol has been reported to activate SIRT336 and SIRT537 as well as non-sirtuin targets38,39. Honokiol is believed to be primarily a SIRT3 activator40,41, although it may also activate SIRT142,43. Sirtuin transgenic mice should be used in combination with STACs to confirm the roles of individual sirtuins in mechanistic studies.
For almost a century NAD+ has been known as a fundamental housekeeping molecule that catalyzes electron transfer in metabolic reduction-oxidation (redox) reactions. In the past 10 years NAD+ has also emerged as a key modulator of cell signalling and survival pathways7,44. The discovery that NAD+ is an essential substrate of sirtuin deacetylation created renewed interest in NAD+ as a potential modulator of longevity and health. The sirtuin deactylation reaction involves the removal of an acetyl group from target substrates via the conversion of NAD+ to nicotinamide (NAM) and O-acetyl-ADP-ribose4,30 (Figure 1).
Figure 1. Synthesis, salvage and metabolism of NAD+.
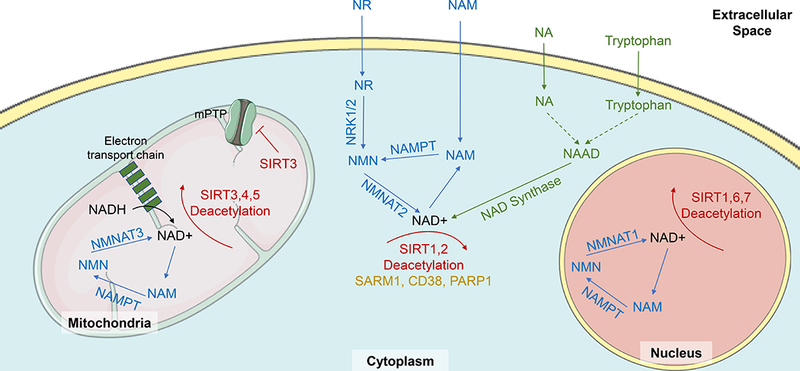
NAD+ supplies in the body come from either de novo synthesis, or via salvage pathways. NAD+ is synthesized de novo from dietary tryptophan via a series of enzymatic reactions ultimately resulting in the production of nicotinic acid adenine dinucleotide (NAAD), which is converted to NAD+ by NAD-synthase. However, in the heart the vast majority of NAD+ is synthesized via salvage routes from nicotinamide (NAM), nicotinamide riboside (NR) and nicotinic acid (NA). NAM and NR are converted to nicotinamide mononucleotide (NMN) by nicotinamide phosphoribosyltransferase (NAMPT) and nicotinamide riboside kinases (NRKs) respectively. NMN is then converted to NAD+ by nicotinamide mononucleotide adenylyltransferease (NMNAT)2 in the cytoplasm, NMNAT3 in the mitochondria, and NMNAT1 in the nucleus. NA is converted, via a series of reactions into NAAD and finally to NAD+ by NAD-synthase. NAD+ is a necessary substrate of sirtuin deacetylation reactions, by which it is converted into NAM. NAD+ is broken down by sterile α and TIR motif–containing protein 1 (SARM1), cluster of differentiation 38 (CD38), Poly ADP-ribose polymerase 1 (PARP1) and SIRTS1–7. SIRTs 3, 4 and 5 are located in the mitochondria, SIRT 2 is located in the cytoplasm, SIRT6 and 7 are found in the nucleus and SIRT1 is found in both the cytoplasm and nucleus. Sirtuins and NAD+ have a variety of roles in cardiac and vascular cells that can effect cardiovascular function. For example, SIRT3 acts to block opening of the mitochondrial permeability transition pore (mPTP), to reduce cell swelling and hypertrophy. Figure adapted from Rajman et al 2018 and Bonkowksi & Sinclair 2016, using adapted images from http://smart.servier.com.
The first evidence that NAD+ boosting was beneficial to health came from yeast studies. Overexpression of yeast PNC1, which encodes a nicotinamidase that catalyzes the first step in the NAD+ salvage pathway, increased stress resistance and lifespan of yeast cells by mimicking caloric restriction45. Increasing age is associated with significantly reduced levels of NAD+46, which is coupled with an increase in NADH47,48, suggesting that redox imbalance may contribute to decreased NAD+ with aging. Increasing NAD+ levels, especially in old age, is associated with a range of beneficial health effects in a variety of models7,49.
NAD+ supplies in the cells are replenished by either de novo synthesis from dietary tryptophan or via salvage pathways from precursors including NAM, nicotinamide riboside (NR) and nicotinic acid (NA)50 (Figure 1). In the heart more than 99% of NAD+ is synthesized via the salvage pathway51. Levels of NAD+ can be raised by providing precursors to NAD+ such as NR, NAM or NMN or by increasing the levels of enzymes involved in the NAD+ synthesis pathways such as nicotinamide phosphoribosyltransferase (NAMPT)52. Another potential approach is to inhibit enzymes that consume NAD+ including cluster of differentiation 38 (CD38)53–55, sterile α and TIR motif–containing protein 1 (SARM1)56,57 and PARP158.
As described below, the sirtuins and NAD+ have major roles in maintaining homeostasis of metabolic processes and cardiovascular function, which are disturbed in aging and in cardiovascular and metabolic diseases. The manipulation of sirtuins and NAD+ could lead to radical new medicines to prevent and treat these and other age-related diseases.
2. Sirtuins and NAD+ in age-related cardiovascular and metabolic diseases
Loss of sirtuin activity and NAD+ levels with age are implicated in the etiology of a wide variety of cardiovascular and metabolic diseases, including atherosclerosis, endothelial dysfunction, acute cardiac syndromes, cardiomyopathy, hypertrophy and heart failure, arrhythmias, hypertension, metabolic syndrome, obesity and fatty liver, diabetes and dyslipidemia. The majority of these studies have been completed in animal models but human studies are increasing. Preclinical studies were completed in either genetically manipulated mice (summarised in Table 1) or using STACs and NAD+ boosters to modulate sirtuin activity (summarised in Table 2).
Table 1.
Effect of over or under expression of SIRT1–7 on cardiac and metabolic function in mice
| Key cardiac and metabolic effects | ||
|---|---|---|
| Gain of Function | Loss of Function | |
| SIRT1 | ↑/↓ atherosclerosis ↓ endothelial inflammation ↓ IR injury susceptibility ↑/↓ hypertrophy and cardiomyopathy ↓ hepatic steatosis ↓ obesity ↑/↔ glucose homeostasis ↑/↓ serum lipid levels ↓ hypertension |
↑/↓ atherosclerosis ↑ endothelial dysfunction, ↓ angiogenesis ↑ IR injury susceptibility ↑/↓ hypertrophy ↑/↓ hepatic steatosis ↑ obesity ↑/↓ glucose homeostasis ↑/↓ serum lipid levels ↑ arrhythmias |
| SIRT2 | ↓ hypertrophy | ↑ hypertrophy and fibrosis |
| SIRT3 | ↓ hypertrophy and fibrosis | ↑ hypertrophy and fibrosis ↑ IR injury susceptibility ↑ obesity ↑/↔ insulin resistance ↑ serum lipid levels |
| SIRT4 | ↑ hypertrophy and cardiac dysfunction | ↓ hypertrophy and fibrosis ↓ obesity ↑ insulin resistance |
| SIRT5 | - | ↑ IR injury susceptibility ↑ hypertrophy and fibrosis |
| SIRT6 | ↓ hypertrophy ↓ obesity ↑ glucose homeostasis, ↑ insulin sensitivity ↓ serum lipid levels |
↑ hypertrophy ↑ obesity ↓ glucose homeostasis ↑ serum lipid levels |
| SIRT7 | - | ↑ hypertrophy and fibrosis ↓ systolic function ↓ hepatic steatosis ↓ glucose intolerance |
Table 2.
Preclinical animal studies of STACs and NAD+ boosters in cardiovascular and metabolic diseases
| Drug | Disease | Model | Effects | Reference |
|---|---|---|---|---|
| NR | Metabolic Syndrome | HFD in mice | ↓ fat mass ↑ insulin sensitivity ↓ LDL | 59 |
| NMN | Endothelial dysfunction | Aging mice | ↑ capillary density ↑ endothelial function | 60 |
| Aging mice | ↑ capillary density ↑ endurance | 61 | ||
| Hypertrophy | Cardiomyopathy mouse | ↑ cardiac function | 62 | |
| TAC in mice | ↓ hypertrophy | 63 | ||
| Cardiomyopathy | Ndufs4 KO mice | ↓ sensitivity to pressure overload | 64 | |
| IR injury | Ischemia in mice | ↓ ischemic damage | 65 | |
| NR | Cardiomyopathy | Aged MDX mice | ↓ inflammation and fibrosis | 66 |
| Resveratrol | Hypertrophy | Rats + doxorubicin | ↓ fibrosis ↓ diastolic dysfunction | 67 |
| TAC in rats | ↑ cardiac function | 68 | ||
| Dahl salt-sensitive rats | ↑ cardiac function ↓ hypertrophy | 69 | ||
| Spontaneously HT rats | ↑ cardiac function ↓ hypertrophy | 70 | ||
| TAC in mice | ↑ cardiac function ↓ hypertrophy | 36 | ||
| Cardiomyopathy | TO-2 hamster | ↓ fibrosis ↑ cardiac function | 71 | |
| Type 1 diabetes model | ↑ cardiac function | 72 | ||
| Autoimmune myocarditis | ↑ cardiac function ↓ hypertrophy ↓ fibrosis | 73 | ||
| Endothelial dysfunction | Rats + cigarette smoke | ↓ ROS ↑ vascular relaxation | 74 | |
| Rhesus monkeys + HFD | ↓ arterial stiffening ↓ inflammation | 75 | ||
| Rat MI model | ↑ angiogenesis ↑ cardiac function | 76 | ||
| I/R injury | IR in mice | ↓ ischemic damage | 77 | |
| IR in rats | ↓ ischemic damage | 78 | ||
| Insulin/glucose regulation | Rhesus monkeys + HFD | ↑ insulin sensitivity | 79 | |
| Mice + HFD | ↑ lifespan ↑ insulin sensitivity | 80 | ||
| Mice + HFD | ↓ obesity ↑ insulin sensitivity | 81 | ||
| SRT2104 | Insulin/glucose regulation | Mice | ↑ lifespan ↑ insulin sensitivity | 17 |
| SRT1720 | Metabolic syndrome | Mice | ↑ lifespan ↓ metabolic syndrome ↓ cholesterol | 16 |
| Genetically obese mice | ↑ lifespan ↓ serum glucose | 32 | ||
| Mice + HFD | ↓ obesity ↑ endurance ↓ insulin resistance | 82 | ||
| Mice + HFD | ↑ lifespan ↓ insulin resistance ↓ hepatic steatosis | 83 | ||
| Endothelial dysfunction | Aging mice | ↑ vascular relaxation ↓ superoxide ↓ inflammation | 84 | |
| SRT3025 | Astherosclerosis | ApoE KO + HFD | ↓ serum LDL and cholesterol ↓ astherosclerosis | 85 |
| Curcumin | I/R injury | IR in rats + curcumin | ↑ cardiac function ↓ ischemic damage | 86 |
| Honokiol | Hypertrophy | TAC in mice | ↓ hypertrophy ↓ fibrosis | 40 |
| Mice + Doxorubicin | ↓ oxidative stress | 41 | ||
| 4-aminobenzamide | Endothelial dysfunction | Diabetes rat model | ↓ endothelial dysfunction ↓ inflammation | 87 |
| Luteolinidin | I/R injury | Isolated rat hearts | ↓ endothelial dysfunction ↓ contractile dysfunction | 88 |
| Apigenin | Metabolic syndrome | Mice + HFD | ↓ glucose dysregulation | 89 |
KO, knockout; MI, myocardial infarction; HT, hypertensive
2.1. Atherosclerosis
Atherosclerosis is the major risk factor for myocardial infarction, coronary artery disease and stroke in the older population. It is the number one cause of death in the developed world and the incidence of atherosclerosis increases with age90. Atherosclerosis results from increased low density lipoprotein (LDL) cholesterol in the plasma, dysfunctional endothelial cells, endothelial oxidative stress, an inflammatory response and ultimately plaque formation91,92 (Figure 2). Risk factors for atherosclerosis, along with age, include hypercholesterolemia, metabolic syndrome, hypertension, obesity and type 2 diabetes90. Sirtuins and NAD+ play a role in the modulation of many of these risk factors and there is increasing evidence that sirtuins may also modulate atherosclerosis itself (Figure 2). SIRT1 levels are decreased in atherosclerotic regions of patients93,94. Monocytes isolated from patients with coronary artery disease also have lower SIRT1 levels than control patients95 suggesting a systemic decrease in SIRT1 in this disease.
Figure 2. The role of Sirtuins and NAD+ in the aging vasculature.
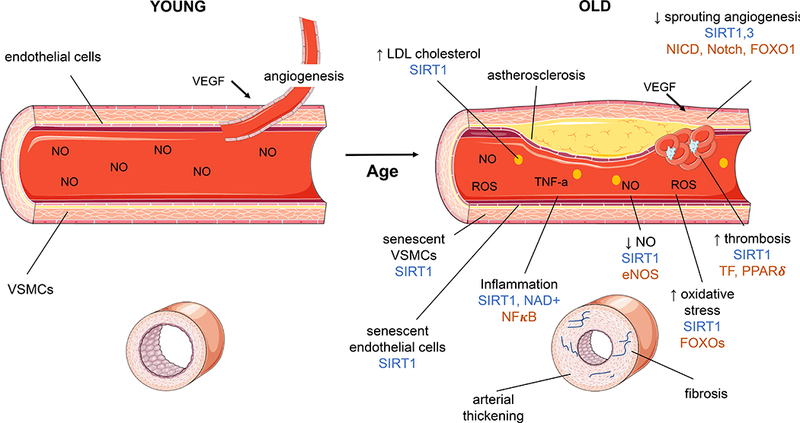
(A) A young blood vessel shows healthy vascular smooth muscle cells (VSMCs), lined with health endothelial cells. Stimulation with nitric oxide (NO) results in vessel contraction. Angiogenesis, the growth of new blood vessels, is induced when vascular endothelial growth factor (VEGF) acts on SIRT1 to relieve notch intracellular domain (NICD) inhibition of Notch. (B) Aging results in changes to the structure and function of the vasculature. With increasing age there are increased atherosclerotic plaques, increased low density lipoprotein (LDL) cholesterol in the blood stream, decreased sprouting angiogenesis, increased thrombosis, increased oxidative stress, decreased nitric oxide, increased inflammation, senescence of endothelial and VSMCs, arterial thickening and fibrosis. These changes contribute to reduced blood flow, and reduced reactivity of the blood vessels in aging. Sirtuins and NAD+ can improve or reverse some aspects of vascular aging (marked in blue on the figure) through modulation of targets (marked in brown). In particular SIRT1 decreases LDL cholesterol levels, reduces vascular cell senescence, increases NO through endothelial nitric oxide synthase (eNOS), reduces inflammation through nuclear factor kappa-light-chain-enhancer of activated B cells (NFκB) modulation, decreases oxidative stress by modulating FOXOs and thrombosis via tissue factor (TF) and peroxisome proliferator-activated receptor delta (PPARδ), and increases angiogenesis through modulation of FOXO1 and NICD-Notch. SIRT3 also has a protective role in angiogenesis and NAD+ reduces vascular inflammation. Figure made using adapted images from http://smart.servier.com.
Pre-clinical studies have shown a protective role for SIRT1 in atherosclerosis, at least in vascular endothelial and smooth muscle cells. Apolipoprotein E (ApoE) knockout mice have increased serum cholesterol levels and can be used as a model of atherosclerosis, especially when fed a high fat diet (HFD). ApoE knockout mice have reduced SIRT1 expression96, particularly in atherosclerotic plaques94. Endothelial cell-specific overexpression of SIRT1 in ApoE knockout mice protects against atherosclerotic plaques97. Additionally, ApoE knockout mice crossed with smooth muscle cell-specific SIRT1 knockout mice, showed increased atherosclerosis94. Treatment of ApoE knockout mice with the STAC SRT3025 reduced atherosclerosis in these mice by increasing LDL uptake85. On the other hand, however, whole-body SIRT1 over-expressors that were placed on an atherogenic diet to induce plaque formation had worse lipid profiles and more severe atherosclerosis than wild-type mice on the same diet98. The proposed mechanism was increased lipid accumulation and secretion in the liver as a result of Creb modulation which resulted in overall pro-atherogenic effects, despite potential benefits of SIRT1 overexpression in the vasculature98. More research is needed to understand the complex role of SIRT1 in lipid metabolism in the liver, especially in the context of high-fat or high-cholesterol diets. This is discussed further in section 2.8 and 2.9.
Other factors associated with atherosclerosis are also modulated by SIRT1. Age-related de-differentiation of vascular smooth muscle cells is implicated in the disease and the induction of SIRT1 with resveratrol can promote differentiation of these cells, and may contribute to protection against atherosclerosis99. Thrombosis contributes to an increased risk of ruptured plaques and clots in atherosclerosis. In mice, inhibition of SIRT1 with sirtinol results in increased tissue factor expression and thrombi promotion indicating SIRT1 may protect against thrombosis100,101. Interestingly treatment with resveratrol to increase sirtuin activity reversed the pro-thrombotic status of cyclooxygenase (COX)-2 knockout mice100.
Overall, the majority of studies show a protective role of SIRT1 in modulating atherosclerosis and many of the factors that contribute to its development. Increasing SIRT1 activity should be further investigated as a promising treatment option for atherosclerosis and its risk factors. More research is needed, however, to confirm the role of SIRT1 in lipid modulation in the liver in the context of different diets, and the effect of this on atherosclerosis.
There are limited studies exploring the effect of the other sirtuins and NAD+ in atherosclerosis. Knockout of SIRT3 in mice lacking the LDL receptor had no effect102, while SIRT6 gene variants have been associated with the development of plaques in patients103. Inhibition of NAMPT with the drug FK866 in ApoE knockout mice resulted in reduced CXCL1 chemokine levels and thus reduced neutrophil infiltration in atherosclerotic plaques104, indicating a potential role of NAD+ in increasing inflammation in atherosclerosis.
2.2. Endothelial dysfunction and artery stiffness
A hallmark of vascular aging is increased dysfunction of the vascular endothelium, characterised by a reduced nitric oxide (NO)-mediated vasodilatory response to stimuli. Additionally, with aging there are changes to the vascular smooth muscle cells which result in increased arterial stiffness and reduced elasticity105,106. These changes reduce the ability of the vasculature to relax and dilate in old age and are associated with an increased risk of a range of cardiovascular diseases including hypertension, heart failure and coronary artery disease105. The role of SIRT1 has been explored in the context of endothelial dysfunction and appears to contribute to normal endothelial activity and protect against dysfunction in aging and other models of damage (Figure 2).
Inhibition of SIRT1 is associated with poor vascular function and increased artery stiffness. Mice with knockout of both ApoE and SIRT1 show increased endothelial superoxide production and increased NF-κB compared to the ApoE knockout alone, indicating a role of SIRT1 in reducing endothelial dysfunction, at least in the context of atherosclerosis107. SIRT1 inhibition with sirtinol in young wild-type mice resulted in artery stiffness to a similar level to that seen in aging wild-type mice108. Specific inhibition of SIRT1 in the arterial endothelium of mice resulted in a reduction in NO and an inhibition of endothelium-dependent vasodilation109. Importantly, the same study showed in vitro that SIRT1 increased endothelial NO by directly deacetylating and thus stimulating the activity of endothelial nitric oxide synthase (eNOS)109.
Several studies have explored the protective effects of increasing SIRT1 activity on the vascular endothelium. A cigarette-smoke model of vascular aging in rats resulted in impaired artery relaxation, increased reactive oxidative species (ROS) and increased inflammatory markers. Activation of SIRT1 either genetically or with resveratrol prevented these effects on the vasculature74. SIRT1-overexpressing mice fed a HFD had better endothelium-dependent vaso-relaxation, and increased eNOS expression, compared to wild-type mice fed a HFD97. SIRT1 activation with SRT1720 in old mice restored endothelial function by increasing COX-2 mediated dilation, and also reduced superoxide production and decreased inflammation84. SIRT1 also acts to prevent senescence of endothelial cells, which may contribute to vascular endothelial dysfunction in aging110,111. Importantly for clinical translation, in rhesus monkeys increasing SIRT1 activity with resveratrol treatment prevented arterial stiffening and inflammation75 and a human study found that in older patients with stiffer vessels there was reduced SIRT1 expression in artery endothelial cells108.
The role of the other sirtuins in endothelial dysfunction has not been well characterised. SIRT6 appears to have a role in protecting against endothelial cell senescence112,113 which may contribute to protection against age-related endothelial dysfunction. NAD+ may also have a protective role in the aging vasculature, as NMN treatment of old mice improved endothelial relaxation in arteries and reduced artery stiffness60. Treatment with a PARP1 inhibitor in a diabetes-induced vascular dysfunction rat model was shown to prevent endothelial dysfunction and inflammation, although NAD+ levels were not explicitly investigated87. Targeting detrimental changes to the endothelium in aging with SIRT1 activators or NAD+ boosters shows promise as an area of focus for treating age-related vascular dysfunction and arterial stiffening.
2.3. Angiogenesis
Angiogenesis, which involves endothelial cell proliferation and migration in response to vascular endothelial growth factor (VEGF), is also decreased in aging114. Angiogenesis may have a protective role in cardiovascular disease by increasing blood flow and myocardial function, especially in ischemic heart disease115,116.
SIRT1 and SIRT3 have both been shown to have roles in inducing angiogenesis. SIRT1 acts as a negative regulator of Notch and FOXO1, key regulators of blood vessel growth117. In vitro studies show that SIRT1 knockout results in reduced angiogenesis of endothelial cells118. In mice and zebrafish a reduction in SIRT1 reduces sprouting blood vessel formation118. A more recent study showed that in aged endothelial cell-specific SIRT1 knockout mice there is reduced capillary density which was exacerbated by transverse aortic constriction (TAC), an intervention to model hypertrophy and heart failure in mice119. In vitro studies showed that SIRT1 knockout resulted in reduced angiogenic response to VEGF, possibly due to reduced expression of the VEGF receptors FLT1 and FLK2119. Resveratrol pre-treatment to increase SIRT1 activity in a rat myocardial infarction model upregulated modulators of angiogenesis and increased capillary density which ultimately resulted in improved cardiac function76. Bone marrow cell (BMC) treatment increased angiogenesis and improved cardiac function in a mouse model of myocardial infarction120. Interestingly, it appears that SIRT3 is required for this protection as treatment with BMCs isolated from SIRT3 knockout mice did not result in angiogenesis or the associated cardiac benefits120.
Additionally, recent findings showed that NMN treatment in aging mice resulted in a reversal of the age-related decline in capillary density and blood flow in muscles which resulted in increased mobility and endurance61. Importantly, the authors demonstrated that the mechanism of reduced blood flow with aging is related to reduced NAD+-dependent SIRT1 activity and that increasing the NAD+-SIRT1 axis with NMN resulted in SIRT1-dependent inhibition of Notch1 intracellular domain (NICD) and promoted VEGF stimulated sprouting angiogenesis61 (Figure 3). Hydrogen sulphide (H2S) was also shown to improve angiogenesis via NAD+- and SIRT1-dependent mechanisms61, although the exact mechanism of the H2S-SIRT1 interaction remains to be elucidated121. Furthermore, Das and colleagues found that NMN treatment also increased sprouting of aortic rings from aging wild-type, but not SIRT1 knockout mice, indicating that NMN may also be beneficial in inducing SIRT1-dependent angiogenesis in tissues other than skeletal muscle61.
Figure 3. The role of SIRT1 and NAD+ in age-related endothelial dysfunction.
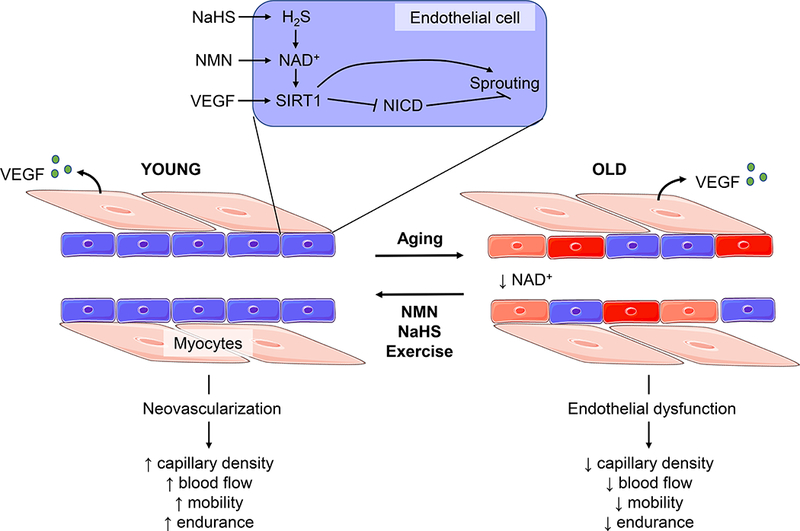
In young healthy capillaries angiogenesis occurs after stimulation of endothelial cells with VEGF released from myocytes. Characteristic changes of the vasculature with old age include dysfunctional endothelial cells, and reduced VEGF-induced angiogenesis. These age-related changes result in reduced capillary density, reduced blood flow and ultimately reduced mobility and endurance. A mechanism of this reduced angiogenesis in old age is reduced endothelial SIRT1 and NAD+, which ostensibly reduces the sensitivity of endothelial cells to VEGF. Increasing the hydrogen sulfide (H2S)-NAD+-SIRT1 axis in endothelial cells, either with NaHS to boost H2S, NMN to boost NAD+ or with exercise, causes SIRT1-dependent inhibition of Notch Intracellular Domain (NICD) which results in increased sprouting angiogenesis. These interventions can reverse endothelial dysfunction and increase angiogenesis in old age, to return capillary density and endurance to the levels of that seen in young mice. Figure adapted from Das et al 2018, using adapted images from http://smart.servier.com.
These promising results indicate that NAD+ boosters and sirtuin activators can be used to increase angiogenesis in cardiovascular diseases and age-related blood flow dysfunction.
2.4. Ischemia-Reperfusion Injury
Many studies over the last decade have investigated the role of sirtuins and NAD+ in the context of ischemia-reperfusion (IR) injury. IR injury occurs clinically in the context of myocardial infarction or during cardiovascular surgery when blood supply to the heart, or part of the heart, is stopped. The period of ischemia can result in tissue injury and cell death as a result of rapidly reduced ATP levels. This damage is exacerbated by rapid reperfusion, which results in the induction of oxidative stress and apoptosis. IR injury increases with age122 and periods of ischemia or hypoxia in cardiomyocytes are associated with a reduction in endogenous NAMPT65,123, as well as downregulation of SIRT1124, SIRT3125 and SIRT4126.
Increasing SIRT1 has a protective role in IR injury (Figure 4). Cardiac-specific SIRT1 knockout mice display increased cardiac damage after exposure to IR, whilst mice overexpressing SIRT1 in the heart are protected from IR damage124. Overexpression of SIRT1 resulted in upregulation of anti-oxidant pathways mediated by FOXO1 and manganese superoxide dismutase (MnSOD) and downregulation of pro-apoptotic pathways mediated by caspase 3 and Bax124. A similar role of SIRT1 was observed in rats exposed to IR injury86, and resveratrol pre-treatment protected both mice and rats from IR injury77,78. Other in vitro studies have shown the same anti-apoptotic role of SIRT1 by mediating FOXO1 expression, in the context of hypoxia127.
Figure 4. The role of Sirtuins and NAD+ in age-related metabolic and heart diseases.
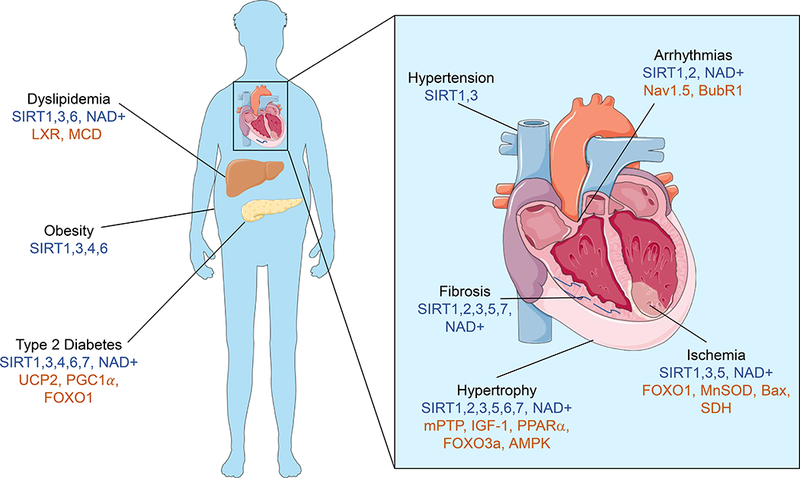
In old age there is increased risk of cardiovascular and metabolic diseases including fatty liver, dyslipidemia, obesity, type 2 diabetes, hypertension, arrhythmias, fibrosis, hypertrophy and ischemia/reperfusion injury. Sirtuins and NAD+ have roles in protecting against or preventing the development of these diseases (shown in blue on figure), through modulation of a variety of proteins (shown in brown on figure). SIRT1, 3, 6 and NAD+ have roles in modulating dyslipidemia, with the liver X receptor (LXR) and malonyl-CoA decarboxylase (MCD) identified as targets, and SIRT1, 3, 4 and 6 have roles in preventing obesity. SIRT1, 3, 4, 6, 7 and NAD+ may have protective roles in Type 2 Diabetes and identified targets include mitochondrial uncoupling protein 2 (UCP2), PGC1α and FOXO1. SIRT1 and 3 have protective roles in hypertension, and SIRT1, 2, 3, 5, 7 and NAD+ have been associated with reduced cardiac fibrosis. SIRT1, 2, 3, 4, 6, 7 and NAD+ protect against cardiac hypertrophy with identified targets including the mitochondrial permeability transition pore (mPTP), insulin-like growth factor (IGF)-1, PPARα, eukaryotic initiation Factor 2 (elF2a), FOXO3a and AMP-activated protein kinase (AMPK). SIRT1, 3, 5 and NAD+ protect against ischemia/reperfusion injury via FOXO1, manganese superoxide dismutase (MnSOD), Bcl-2-associated X protein (Bax) and succinate dehydrogenase (SDH). SIRT1, 2 and NAD+ improve arrhythmias via sodium channel Nav1.5 and BubR1. Figure made using adapted images from http://smart.servier.com.
The mitochondrial sirtuins SIRT3, 4 and 5 also have protective roles in IR injury. In vitro studies of SIRT3 overexpression in cardiomyocytes exposed to hypoxia showed protection against cell damage via a downregulation of apoptosis involving Ku70 and Bax128. The same role is seen in vivo, where SIRT3 knockout mice are more susceptible to IR injury and have increased damage and reduced mitochondrial function post injury when compared to wild-type mice122. Overexpression of SIRT4 in cardiomyoblast cells increased their viability post hypoxia by decreasing caspase activity and apoptosis, while knock-down of SIRT4 in the same cells results in reduced viability and an induction of apoptosis126. SIRT5 knockout mice have increased infarct size following IR injury compared to wild-type mice and an inhibitor of succinate dehydrogenase protected against this injury, implying a role of SIRT5 in modulating protein succinylation in the heart in the context of IR injury129. In vitro studies also support a role of SIRT5 in protecting against IR injury, at least in part by reducing apoptosis130.
Little is known about the role of the nuclear sirtuins SIRT6 and SIRT7, and the other cytoplasmic sirtuin SIRT2, in IR injury. One study showed that cardiomyocytes isolated from mice overexpressing SIRT6 had less damage when exposed to hypoxia than those isolated from wild-type mice, via a downregulation of apoptosis and oxidative stress through modulation of B-cell lymphoma 2 (Bcl-2), NF-κB, phosphorylated-Akt and phosphorylated-5’ adenosine monophosphate-activated protein kinase (AMPK)131. An in vitro study suggested that SIRT2 may play an contributing role in IR injury with downregulation of SIRT2 protecting against hypoxia in H9c2 cells132. This supports other evidence suggesting that SIRT2 acts as a negative regulator of biological stress133.
Cardiac-specific overexpression of NAMPT in mice results in an expected increase in cardiac NAD+ levels and protection against IR injury123. Additionally ischemic preconditioning, which is known to reduce damage to subsequent IR injury, appears to induce protection, at least in part, through NAMPT upregulation65. NAMPT knockout mice show no protection against IR injury with preconditioning65. Increasing NAD+ levels with the precursor NMN also protects against IR injury65. PARP inhibitors also protect against IR injury in a variety of animal models134, possibly through increased NAD+ levels. The CD38 inhibitor luteolinidin also prevented left ventricular contractile and endothelial dysfunction after IR injury in isolated rat hearts88.
Overall sirtuins and NAD+ appear to have a strongly protective role in IR injury. There is the potential for the development of STACs or NAD boosting therapeutics for use during cardiac surgery, or to attenuate damage post-myocardial infarction in older patients.
2.5. Hypertrophy, fibrosis and heart failure
Aging is associated with distinct structural changes in the heart including increased fibrosis, left ventricular hypertrophy and dilation135–137. Hypertrophy and fibrosis in the aging heart cause systolic and diastolic dysfunction and can ultimately result in heart failure135–137. Age-related hypertension can contribute to the development of hypertrophy and heart failure in humans. Mice do not naturally develop age-related hypertension so this is modelled in rodents with interventions including TAC which causes pressure overload, angiotensin II exposure, doxorubicin exposure, transgenic cardiomyopathy models, diabetic cardiomyopathy and other stress-induced cardiomyopathy models.
SIRT1 has apparently opposing roles in age-related structural changes of the heart. Several mouse studies have observed the development of cardiac hypertrophy in mice with SIRT1 overexpression138, with one study showing the degree of impairment was dependent on the degree of overexpression139. High levels of SIRT1 overexpression (20 fold) resulted in dilated cardiomyopathy, whilst moderate overexpression (6.8 fold) caused diastolic dysfunction, and low overexpression (3.2 fold) resulted in normal function at baseline, but the development of cardiac dysfunction with pressure overload139. Knockout of either SIRT1 or peroxisome proliferator-activated receptor α (PPARα) reduced hypertrophy in a TAC mouse study, whilst the overexpression of both proteins exacerbated it140, ostensibly due to upregulation of estrogen-related receptors and resulting mitochondrial dysfunction140.
Conversely, SIRT1 can also have a protective function (Figure 4). Another study exploring levels of cardiac-specific SIRT1 overexpression on age-related remodeling in 18 month old mice, found that low to moderate overexpression (2.5–7.5 fold) protected against age-related hypertrophy, fibrosis and cardiac dysfunction, whilst only a high level of overexpression (12.5 fold) resulted in cardiomyopathy and hypertrophy141. The authors suggest that the detrimental effect of a high level of SIRT1 overexpression is due to induction of mitochondrial dysfunction141, which may also contribute to hypertrophic damage in the context of TAC pressure overload and increased SIRT activity139. Cardiomyocytes with SIRT1 activation either genetically or with resveratrol, were protected against phenylephrine-induced hypertrophy and related inflammation. Interestingly, in direct opposition to that seen above, the inhibition of PPARα stopped this protective effect142. The same study also showed that SIRT1 over expression in mice protected against isoproterenol-induced hypertrophy142. Mouse studies using either stress-induced or diabetes models of cardiomyopathy observed that SIRT1 knockout increased hypertrophy and decreased cardiac function and treatment with either the sirtuin activator STAC-3 or resveratrol, was protective against this hypertrophy72,143. Consistent with this, resveratrol treatment in rat or hamster models of heart failure and cardiomyopathy also prevented hypertrophy and a decline in cardiac function, though the exact target of resveratrol is difficult to prove without the use of SIRT1 knockouts67–71,73.
The conflicting roles of SIRT1 in these studies imply a potentially different effect of SIRT1 on cardiac remodeling in different contexts and models. Investigation of the role of sirtuins in more clinically relevant models of cardiac aging, or studies in aging humans themselves, will be essential. Importantly, the one study discussed above that did use naturally aging mice found a protective effect of low to moderate SIRT1 overexpression on hypertrophy141. More studies are required to fully understand the complex effects of SIRT1 in cardiomyopathy and hypertrophy in animal models and humans.
SIRT3 has a clear protective role in cardiac hypertrophy. Doxorubicin-induced cardiomyopathy, TAC-induced hypertrophy, and a Friedreich’s ataxia model of cardiomyopathy all resulted in reduced SIRT3 levels, which were associated with hyper-acetylation of mitochondrial proteins62,144,145. SIRT3 knockout mice develop hypertrophy and fibrosis as they age as a result of increased mitochondrial permeability transition pore (mPTP) opening (Figure 1) and transforming growth factor (TGF)-β1 signaling146,147. SIRT3 knockout mice are also more sensitive to the hypertrophic effects of TAC, angiotensin II145,146,148, doxorubicin144,149, diabetes150 and isoprotenerol151 than wild-type mice. SIRT3 overexpression results in resistance to hypertrophy-inducing stimuli, as a result of increased FOXO3a, MnSOD, oxoguanine glycosylase (OGG)1, parkin and catalase activation144,148,150. HFD is also associated with hypertrophy and cardiac dysfunction, which is accompanied by a decrease in cardiac SIRT3. Moreover, SIRT3 KO mice with HFD had greater cardiac dysfunction than HFD-fed wild-type mice152. The known protective effects of resveratrol on hypertrophy and fibrosis may be as a result of both SIRT1 and SIRT3 activation, as the protective effects of resveratrol were not observed in SIRT3 knockout mice in one study36. Honokiol is a poly-phenolic compound that is believed to activate SIRT3. Honokiol treatment in mice exposed to pressure overload and drug-induced heart failure prevented hypertrophy and fibrosis40 as well as the associated mitochondrial damage and cell death41.
The role of the other sirtuins in cardiac hypertrophy and fibrosis has not been as extensively studied. SIRT2 knockout mice have increased hypertrophy and fibrosis and decreased systolic function compared with wild-type mice153. Moreover, cardiac-specific SIRT2 overexpression protected against angiotensin II-induced hypertrophy by activating liver kinase B1 (LKB1) which ultimately resulted in maintenance of AMPK levels in hypertrophy153. SIRT4 appears to contribute to remodeling with SIRT4 knockout mice having reduced fibrosis and hypertrophy in response to angiotensin II, and cardiac-specific SIRT4 overexpression increasing hypertrophy and reducing cardiac function154. SIRT5 prevents age-related hypertrophy with aging SIRT5 knockout mice displaying more fibrosis, hypertrophy and systolic dysfunction than age-matched wild-types155. SIRT6 knockout mice have increased hypertrophy, whilst over expression of SIRT6 was associated with protection against hypertrophic stimuli in mice156. Overexpression of nicotinamide mononucleotide adenylyltransferease (NMNAT)2 in rat cardiomyocytes to increase NAD+, protected against hypertrophy in a SIRT6- but not SIRT1-dependent manner, further suggesting a protective role of SIRT6157. Finally, SIRT7 knockout in mice results in hypertrophy, fibrosis, inflammation, systolic dysfunction and increased apoptosis158,159, indicating a protective role of SIRT7 against cardiomyopathy and hypertrophy as well.
Several studies suggest that increasing NAD+ may protect against hypertrophy. Increasing NAD+ in mice by either dosing with NMN or overexpressing NAMPT prevented hypertrophy induced by TAC and improved cardiac function63, by increasing energy production in the mitochondria and reducing mPTP opening63. The addition of NAD+ prevented isoproterenol-induced hypertrophy in mice and interestingly this was mediated by SIRT3, but not SIRT1, through activation of LKB1-AMPK signaling151. NMN treatment protected against cardiac dysfunction in a Friedreich’s ataxia cardiomyopathy mouse model in a SIRT3-dependent manner62 and increased cardiac function in Ndufs4 knockout mice exposed to pressure overload64. NR treatment also reduced inflammation and fibrosis in aged MDX mice, a muscular dystrophy model that develops cardiomyopathy66. Finally, PARP inhibitors protected against hypertrophy and improved cardiac function in several rodent models134, possibly in part by increasing NAD+ levels.
The role of sirtuins and NAD+ in heart failure, rather than remodelling, has not been as extensively studied. Levels of SIRT1 increase in failing hamster71, rat68, mouse141 and dog hearts160 but whether this increase is cardio-protective or contributes to the development of heart failure is not clear. The association between heart failure and the other sirtuins is not well understood although one study found reduced SIRT6 expression in failing human hearts156. Failing hearts also have extensive mitochondrial hyper-acetylation which may contribute to dysfunction in heart failure161 and NAD+ levels are reduced in failing mouse and human hearts63,161.
Overall, moderately increasing SIRT1 activity, increasing SIRT3 activity or boosting NAD+ levels appear to be promising approaches for protection against age-related hypertrophy and fibrosis. More research in clinically relevant models of heart aging is needed, however, to understand the potentially detrimental effects of high overexpression of SIRT1.
2.6. Arrhythmias
The risk of atrial fibrillation and other types of arrhythmias increases significantly with age136. The role of sirtuins and NAD+ in arrhythmias has been minimally investigated. A recent study showed that SIRT1 deacetylases the voltage gated Na+ channel (Nav1.5), which is necessary for normal cardiac electrophysiology, to increase its activity. Cardiac-specific SIRT1 knockout mice had decreased Nav1.5 expression, which resulted in abnormal cardiac conduction162. One study in BubR1 knockout mice, which are short-lived in part because of defective cardiac electrophysiology, showed a potential role of NAD+ and SIRT2 in treating arrhythmias. They found that either overexpressing SIRT2 or increasing NAD+ levels with NMN increased BubR1 and partly reversed the repolarisation defects observed163. Given the association between reduced NAD+ and sirtuin activity in the heart with age and the increased risk of arrhythmias, this is an area that warrants further research.
2.7. Hypertension
Hypertension is a risk factor for other cardiovascular diseases and the risk of hypertension increases with age, especially in women164,165. Few studies have explored the effects of the sirtuins on hypertension. Angiotensin II exposure, which is a commonly used model of hypertrophy in mice, also causes hypertension although most studies of angiotensin II-induced hypertrophy discussed above did not investigate blood pressure changes. One study of SIRT1 overexpression in the context of angiotensin II found that SIRT1 protected against the systolic blood pressure increase seen in wild-type mice166. In addition, a study in rats with spontaneous pulmonary artery hypertension found that they had reduced SIRT3 expression and that SIRT3 overexpression prevented the development of hypertension167. Polymorphisms in SIRT3 were also associated with pulmonary artery hypertension in patients167. More research in this area will identify whether boosting sirtuins or NAD+ may protect against hypertension in aging.
2.8. Diabetes, glucose/insulin dysregulation and metabolic syndrome
The metabolic syndrome is a set of risk factors for cardiovascular diseases, including obesity, diabetes, increased LDL levels, increased triglyceride (TG) levels and hypertension168,169. Type 2 diabetes is characterised by high blood glucose levels, low levels of insulin, and insulin resistance. The risk of diabetes is increased by obesity and a lack of exercise, and diabetes is associated with an increased risk of cardiovascular disease, stroke and kidney failure170. Increasing age is a significant risk factor for insulin resistance and impaired glucose tolerance171–173. The role of sirtuins in glucose and insulin control, obesity and the development and prevention of diabetes has been well explored, at least for SIRT1, 3, 4 and 6. A variety of animal models are used to study diabetes and insulin resistance, including aging mice, transgenic mouse models of diabetes such as db/db and the induction of obesity and diabetes with a HFD. Human studies are limited but a specific SIRT1 mutation was associated with type 1 diabetes in one patient study174 and low SIRT1 has been associated with insulin resistance in the offspring of those with type 2 diabetes175. Two human studies found associations between SIRT3 polymorphisms and the metabolic syndrome, suggesting a role of SIRT3 in this syndrome in humans167,176
SIRT1 transgenic overexpressors fed a diet high in fat and cholesterol have better glucose homeostasis than wild-type mice fed the same diet, but with no difference in body weight12,98,177. SIRT1-overexpressing mice fed a standard diet also show increased glucose tolerance, which is accompanied by reduced cholesterol, reduced fat mass and lower body weight178. SIRT1 overexpression in the db/db diabetic mouse model also improved glucose tolerance and reduced hepatic glucose production177. Pancreatic β-cell-specific overexpression of SIRT1 was associated with increased insulin secretion and improved glucose tolerance, even as the mice aged179. Both mice and rhesus monkeys with insulin resistance induced by a HFD had increased insulin sensitivity when SIRT1 activity was increased with resveratrol or SRT1720 treatment79–83. Mice fed a standard diet and dosed with the SIRT1 activators SRT2104 or SRT1720 also had increased insulin sensitivity16,17. The role of reduced SIRT1 function in mice is less clear. One study found that SIRT1 knockout mice displayed reduced basal and glucose stimulated insulin levels as a result of upregulation of uncoupling protein (UCP)2 expression180. Another study, however, showed that on a standard diet liver-specific SIRT1 knockout mice had no change in insulin or glucose modulation and that on a HFD they maintained insulin sensitivity, implying a role of SIRT1 in contributing to insulin resistance in this model181.
Whilst overexpression of SIRT1 in high caloric exposure generally acts to decrease glucose and increase insulin response, increased SIRT1 in fasting appears to increase glucose production. Acute SIRT1 knockout in the liver of fasted mice actually resulted in increased insulin sensitivity, decreased serum glucose and decreased glucose production, and overexpression reversed these effects182. In vitro studies show that SIRT1 negatively regulates glycolysis through phosphoglycerate mutase 1 (PGAM1) and that in fasted cells there is increased SIRT1 activity and thus reduced glucose breakdown183. Fasting in mice also resulted in increased SIRT1 modulation of the glycolytic and gluconeogenesis pathways via FOXO1 and PGC-1α to potentially increase glucose levels22. Thus, there is clearly a complex and potentially tissue-specific relationship between nutritional state, SIRT1 and insulin and glucose regulation that requires further research.
Unlike SIRT1, SIRT3 knockout mice fed a high cholesterol diet gained more weight but had the same glucose tolerance as wild-type mice102. One study of HFD-fed SIRT3 knockout mice found increased fatty acid oxidation, implying that SIRT3 may act to protect against insulin resistance in high calorie environments by decreasing fatty acid oxidation184. Interestingly, SIRT3 appears to have the opposite effect in low calorie environments as SIRT3 knockout mice have decreased levels of fatty acid oxidation in fasting24. SIRT4 knockout mice have upregulation of fatty acid oxidation and metabolism25,185,186 which results in protection against diet-induced obesity187. SIRT4 knockout mice also have increased insulin secretion in response to amino acids25 which results in insulin resistance and glucose intolerance188. Interestingly, SIRT4 knockout mice also have increased SIRT1 activity so there are likely several sirtuin pathways involved in the regulation of glucose and insulin homeostasis185.
Whole-body SIRT6 knockout mice die prematurely from hypoglycaemia. The mechanism appears to be increased AKT activation and reduced insulin-like growth factor (IGF)-1 levels which results in upregulation of glucose transporters and increased glucose uptake in skeletal muscle and brown adipose tissue189–191. SIRT6 knockout mice also have increased glucose production as normally SIRT6 downregulates hepatic glucose production via PGC-1α192, as well as increased glycolysis, possibly through increased activity of hypoxia-inducible factor (HIF)-1α191,193. Interestingly, overexpression of SIRT6 improved glucose homeostasis in aging mice15 and increased glucose tolerance and insulin sensitivity in the context of a HFD, along with a reduction in visceral fat11. The authors suggest that the HFD caused defective insulin secretion as a result of increased fat and inflammatory cells in the pancreas and that SIRT6 overexpression improved insulin sensitivity by reducing this inflammatory pancreatic infiltration11. Neural-specific SIRT6 knockout mice develop obesity overtime implying that SIRT6 may act as a central regulator of growth and obesity194. As such, SIRT6 may be a promising target for therapeutics to modulate obesity, increase insulin sensitivity and reduce glucose levels in patients. SIRT7 may also have a role in regulating glucose levels as SIRT7 KO mice were resistant to glucose intolerance195. Supplementation of mice on a HFD with NR increased insulin sensitivity and reduced fat mass compared to HFD alone. Interestingly NR also increased the activity of SIRT2 and SIRT359. Similarly, supplementation with NMN also prevented the insulin resistance and glucose intolerance associated with a HFD in mice and improved glucose tolerance in aging mice196. Finally, treatment with the CD38 inhibitor apigenin in mice with diet-induced obesity, resulted in increased NAD+ levels as well as protection against glucose dysregulation89.
Overall sirtuins appear to have a complex but generally protective effect on obesity but whether this effect is related to or independent from effects on insulin and glucose regulation is still not known. Boosting NAD+ levels or increasing SIRT1 and/or SIRT6 activity in the context of the metabolic syndrome, type 2 diabetes and/or insulin resistance could be an important therapeutic approach for preventing and treating these diseases in aging.
2.9. Dyslipidemia
Dyslipidemia is characterised by abnormal levels of lipids including TGs and cholesterol in the blood. The most common of these abnormalities in the elderly is hypercholesterolemia which is an increased level of LDL cholesterol197,198. The role of sirtuins and NAD+ in dyslipidemia has not been well characterized. SIRT1 has a role in cholesterol homeostasis as a positive regulator of the cholesterol sensing proteins, liver X receptors (LXRs)199. SIRT1 knockout results in reduced LXR expression and reduced HDL and TG lipid levels, implying a protective role of SIRT1 in increasing the levels of beneficial HDL cholesterol199. The role of SIRT1 in lipid homeostasis also appears to depend on the nutritional state of the mice. Acute SIRT1 knockout in the liver of fasted mice results in decreased serum cholesterol levels and increased liver cholesterol182. The opposite role for SIRT1 was seen on standard or HFD, however, where SIRT1 overexpression resulted in decreased serum cholesterol levels178,200.
SIRT3 knockout mice with HFD exposure develop hyperlipidemia implying a role of SIRT3 in maintaining lipid homeostasis24. SIRT4 also has a role in lipid metabolism by regulating mast cell degranulating (MCD) peptide activity187. SIRT6 plays a role in negatively regulating glycolysis and TG synthesis193 and may also have a role in protecting against increased LDL cholesterol levels201. SIRT6 knockout mice have increased LDL whilst SIRT6 over-expressing mice fed a HFD have lower LDL and TG levels than wild-type mice11,201. NR in HFD-fed mice was associated with reduced LDL cholesterol levels compared to HFD alone59. Niacin, which is a collective term for NAM and NA, has for many years been used as a treatment for hypercholesterolemia202 but the mechanism of this protection is still not known203. NAD+-dependent activation of sirtuins is a possible mechanism204 but the effect may also be driven by NAD+-independent activation of the G-coupled receptor GPR109A203.
NAD+ and sirtuin boosters may have beneficial roles in treating dyslipidemia, especially in the context of western diets, but more research is required to fully understand the mechanisms behind this protection and the role of each of the sirtuins in lipid homeostasis.
3. Clinical trials of molecules to modulate sirtuins and NAD+ in cardiovascular and metabolic diseases
The use of both STACs and NAD+ boosters in cardiovascular and metabolic diseases has shown beneficial effects in a variety of animal models (Table 2). Given the preclinical evidence for benefits of sirtuins and NAD+ in preventing and treating cardiovascular and metabolic diseases, many researchers recognise the potential of these pathways as medicines. As a result, there are a growing number of human trials of sirtuin and NAD+-boosting drugs, some of which are showing protective effects in cardiovascular and metabolic diseases (Table 3).
Table 3.
Clinical trials of STACs and NAD+ boosters in cardiovascular and metabolic diseases
| Drug | Effect | Study Population | Reference |
|---|---|---|---|
| Resveratrol | No change in walking performance | Patients with peripheral artery disease | 213 |
| ↑ Vascular function No change in glucose metabolism |
Glucose intolerant adults | 214 | |
| ↓ systolic blood pressure ↑ Insulin sensitivity |
Healthy obese men | 207 | |
| No effect | Healthy non-obese men | 208 | |
| ↓ Body weight ↓ Fat mass ↓ Total insulin secretion |
Metabolic syndrome patients | 209 | |
| ↓ Insulin resistance ↓ Fasting glucose |
Type 2 diabetes patients | 210 | |
| ↑ Endothelial function ↑ Diastolic function |
Patients with coronary artery disease | 211 | |
| ↓ Serum LDL ↓ Insulin resistance |
NAFLD patients | 224 | |
| ↓ Hepatic steatosis ↓ Inflammation |
NAFLD patients | 212 | |
| SRT2104 | ↓ Body weight Changes to glucose control |
Type 2 diabetes patients | 217 |
| ↓ Serum LDL ↓ Serum TGs |
Healthy older volunteers | 215 | |
| ↓ Serum LDL ↓ Serum TGs |
Healthy smokers | 216 | |
| Niacin | ↓ Dyslipidemia | Varied | 202 |
| NRPT | Safely tolerated | Healthy volunteers | 221 |
| NR | Safely tolerated | Healthy volunteers | 218,219 |
| ↓ blood pressure ↓ arterial stiffening |
Healthy volunteers | 220 | |
| Underway, aim to improve astherosclerosis and heart failure | Varied | #NCT02812238 #NCT03423342 (Clinicaltrials.gov) |
|
| NMN | Underway, aim to improve cardiometabolic outcomes | Women 55+ with dyslipidemia | #NCT03151239 (Clinicaltrials.gov) |
| INO-1001 | Safely tolerated | Patients with myocardial infarction | 223 |
3.1. Sirtuin Activating Compounds
There have been over 20 clinical trials of resveratrol, including at least six investigating cardiovascular outcomes, and eight with primary metabolic outcomes including diabetes205. Clinical trials of resveratrol have been recently and thoroughly reviewed6,205,206 with most trials showing beneficial cardiovascular and metabolic effects of resveratrol6,205,206. For example, administration of resveratrol to healthy obese men for 30 days improved insulin sensitivity and decreased systolic blood pressure207, whilst no effect was seen in healthy non-obese men208. Resveratrol treatment in patients with metabolic syndrome resulted in reduced body weight, reduced fat mass and decreased total insulin secretion209 and patients with type 2 diabetes treated with resveratrol for 45 days had lower fasting glucose and decreased insulin resistance than controls210. Resveratrol has also shown improvements in patients with coronary artery disease211 and non-alcoholic fatty liver disease36,212. Not all studies however, have seen improvements with resveratrol treatment205,206,213. Resveratrol treatment in glucose intolerant patients for six weeks resulted in improved vascular function with no effect on glucose tolerance or insulin sensitivity214 and resveratrol treatment in patients with peripheral artery diseases did not improve the primary outcome of walking performance213. Potential differences in clinical trial design such as drug formulation, dose and timing, may explain these varied results6.
Clinical trials of the STAC SRT2104 have shown beneficial serum lipid lowering effects in patients. A trial in healthy older volunteers found that SRT2014 decreased serum total cholesterol, LDL and TG levels with no change in glucose tolerance215. SRT2104 also reduced LDL and TG levels in healthy smokers but had no effect on the vasculature216. A small cross-over trial of SRT2104 in patients with type 2 diabetes found that drug treatment was associated with weight loss but there was little effect on endothelial function or serum lipid profiles and there was a potentially adverse effect on glucose control217. The authors suggest that this negative effect may be a result of acute rather than chronic exposure to SRT2104 and that long term exposure may mimic what is seen in preclinical studies217.
3.1. NAD+ Boosters
There have been a few clinical trials of NAD+ boosters in treating cardiovascular or metabolic diseases with more currently underway6,44. Niacin has been used for many years for the treatment of hypercholesteremia and acts to decrease very low density lipoprotein (VLDL) triglycerides by suppressing lipolysis202, although this effect may be NAD+-independent203. The first study of NR pharmacokinetics in healthy volunteers demonstrated safety and efficacy of single doses of NR in boosting NAD+ levels218. A second pharmacokinetic study investigated the effect of 8 days of increased dosing of NR in healthy volunteers, and also found that NR was well-tolerated and increased circulating NAD+ levels219. Recently, these safety and pharmacokinetic results were confirmed in a 6 week clinical trial of NR in healthy older adults and secondary analyses suggest it also reduced blood pressure and arterial stiffness220. A combination of NR and the polyphenol compound pterostilbene was also found to be tolerated without side effects in a recent Phase I trial221. There are many more trials investigating NR in cardiovascular outcomes including heart failure and atherosclerosis currently underway6,44. The first human trials of NMN are also ongoing222. Safety and pharmacokinetics of the PARP inhibitor INO-1001 were investigated in a small study of patients with myocardial infarction, and it was found to be effective and well tolerated223.
The overall promising results of STACs and NAD+ boosters in cardiovascular and metabolic diseases will result in further clinical trials of these compounds over the next decade and beyond.
Conclusions
This review has shown the clear role of sirtuins and NAD+ in a range of cardiovascular and metabolic diseases and identified exciting opportunities to boost NAD+ and sirtuin activity to treat or prevent these diseases. In particular there is extensive preclinical, and recently clinical, evidence for increasing SIRT1 activity to target type 2 diabetes and dyslipidemia. Additionally, there is promising preclinical evidence for increasing SIRT6 activity in insulin resistance, increasing SIRT3 activity to protect against age-related cardiac hypertrophy and boosting NAD+ levels to promote angiogenesis and increase blood flow, that may translate into clinical studies in the near future.
There are still many questions that remain unanswered, however, and more research to be done. There is very little know about sex differences in the roles of NAD+ and sirtuins in cardiovascular and metabolic diseases and most preclinical studies have only been completed in males. Given the important sex differences in the cardiovascular system with aging225,226 and sex differences in response to longevity interventions227 this area warrants further research. Additionally, more research is needed on unravelling the role of each of the sirtuins in these age-related diseases, especially SIRT2, 4, 5 and 7 where little is currently known. The potentially divergent roles of different sirtuins in these diseases are not well understood, especially the possible detrimental roles of SIRT2 in IR injury and SIRT4 in hypertrophy. More research could improve understanding of how STACs may act differently on different sirtuins, Moreover, future studies should provide a better understanding of the effect of nutritional status on the role of sirtuins in glucose and insulin regulation and in dyslipidemia, to optimise the use of sirtuin modulating therapies in these conditions. In addition, there is still much to learn about the basic physiology and pharmacology of the NAD+ boosting drugs, such as their cellular transport, metabolism and dosing before they can be widely used clinically. Although not discussed in this review there is also the potential that diets which improve cardiovascular or metabolic outcomes228,229, may modulate NAD+ levels and increase sirtuin activity. For example it has been suggested that increased NAD+ may be a significant mechanism contributing to the benefits of a high ketone diet230.
In conclusion, there is clear preclinical evidence for NAD+ boosting and sirtuin activating therapies in treating and preventing age-related cardiovascular and metabolic diseases, which is resulting in growing translation of these promising interventions into clinical trials.
Acknowledgements, Funding and Disclosures:
D.A. Sinclair is a consultant and inventor on patents licensed to Metro International Biotech, Cohbar, Caudalie, Jumpstart Fertility, Life Biosciences and Liberty Biosecurity and D.A.S. is a consultant to EdenRoc Sciences, Senolytic Therapeutics, Spotlight Biosciences, and Continuum Biosciences. This work was supported by the Glenn Foundation for Medical Research and grants from the NIH (RO1 AG028730 and RO1 DK100263). AEK is supported by an NHMRC CJ Martin biomedical fellowship (GNT1122542).
Non-standard Abbreviations and Acronyms
- AMPK
adenosine monophosphate-activated protein kinase
- ApoE
apolipoprotein E
- Bcl2
B-cell lymphoma 2
- BMC
bone marrow cell
- COX
cyclooxygenase
- eIF
eukaryotic initiation factor
- eNOS
endothelial nitric oxide synthase
- H2S
hydrogen sulfide
- HFD
high fat diet
- HIF-1α
hypoxia-inducible factor 1 alpha
- IGF-1
insulin-like growth factor
- IR
ischemia-reperfusion
- LDL
low density lipoprotein
- LKB1
liver kinase B1
- LXR
liver X receptors
- MCD
mast cell degranulating
- MnSOD
manganese superoxide dismutase
- mPTP
mitochondrial permeability transition pore
- NA
nicotinic acid
- NAAD
nicotinic acid adenine dinucleotide
- NAD+
nicotinamide adenine dinucleotide
- NAM
nicotinamide
- NaMN
nicotinic acid mononucleotide
- NAMPT
nicotinamide phosphoribosyltransferase
- NAPRT
nicotinic acid phosphoribosyltransferase
- Nav1.5
voltage gated Na+ channel 1.5
- NF-κB
nuclear factor kappa-light-chain-enhancer of activated B cells
- NICD
Notch1 intracellular domain
- NMN
nicotinamide mononucleotide
- NMNATs
nicotinamide mononucleotide adenylyltransfereases
- NMRK
nicotinamide riboside kinase
- NO
nitric oxide
- NR
nicotinamide riboside
- NRF1
nuclear respiratory factor 1
- OGG1
oxoguanine glycosylase 1
- PARP1
Poly ADP-ribose polymerase 1
- PGAM1
phosphoglycerate mutase 1
- PGC1α
peroxisome proliferator-activated receptor gamma coactivator 1-alpha
- PPARα
peroxisome proliferator-activated receptor α
- Redox
reduction-oxidation
- ROS
reactive oxygen species
- SIR2
silent information regulator
- SREPB
sterol-regulatory-element-binding protein
- STACs
sirtuin activating compounds
- TAC
transverse aortic constriction
- TG
triglyceride
- TGF
transforming growth factor
- UCP2
uncoupling protein 2
- VEGF
vascular endothelial growth factor
- VLDL
very low density lipoprotein
References:
- 1.Sinclair DA, Guarente L. Extrachromosomal rDNA circles - a cause of aging in yeast. Cell. 1997;91:1033–1042. [DOI] [PubMed] [Google Scholar]
- 2.Kaeberlein M, McVey M, Guarente L. The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes Dev. 1999;13:2570–2580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kennedy B, Gotta M, Sinclair D, Mills K, McNab D, Murthy M, Pak S, Laroche T, Gasser S, Guarente L. Redistribution of silencing proteins from telomeres to the nucleolus is associated with extension of life span in S. cerevisiae. Cell. 1997;89:381–391. [DOI] [PubMed] [Google Scholar]
- 4.Imai S, Armstrong CM, Kaeberlein M, Guarente L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature. 2000;403:795–800. [DOI] [PubMed] [Google Scholar]
- 5.Landry J, Sutton A, Tafrov S, Heller R, Stebbins J, Pillus L, Sternglanz R. The silencing protein SIR2 and its homologs are NAD-dependent protein deacetylases. Proc Natl Acad Sci USA. 2000;97:5807–5811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bonkowski MS, Sinclair DA. Slowing ageing by design: the rise of NAD+ and sirtuin-activating compounds. Nat Rev Mol Cell Biol. 2016;17:679–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Imai S ichiro, Guarente L. NAD+ and sirtuins in aging and disease. Trends Cell Biol. 2014;24:464–471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Haigis MC, Sinclair DA. Mammalian sirtuins: biological insights and disease relevance. Annu Rev Pathol. 2010;5:253–295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hubbard BP, Sinclair DA. Small molecule SIRT1 activators for the treatment of aging and age-related diseases. Trends Pharmacol Sci. 2014;35:146–154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kugel S,CS Fitamant J, Ross K, Saha S, Jain E, Gladden A, Arora K, Kato Y, Rivera M, Ramaswamy S, Sadreyev R, Goren A, Deshpande V, Bardeesy N, Mostoslavsky R. SIRT6 suppresses pancreatic cancer through control of Lin28b. Cell. 2016;165:1401–1415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kanfi Y, Peshti V, Gil R, Naiman S, Nahum L, Levin E, Kronfeld-Schor N, Cohen HY. SIRT6 protects against pathological damage caused by diet-induced obesity. Aging Cell. 2010;9:162–173. [DOI] [PubMed] [Google Scholar]
- 12.Pfluger PT, Herranz D, Velasco-Miguel S, Serrano M, Tschop MH, Tschöp MH. Sirt1 protects against high-fat diet-induced metabolic damage. Proc Natl Acad Sci USA. 2008;105:9793–9798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Banks AS, Kon N, Knight C, Matsumoto M, Gutierrez-Juarez R, Rossetti L, Gu W, Accili D. SirT1 gain-of-function increases energy efficiency and prevents diabetes in mice. Cell Metab. 2011;8:333–341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Satoh A, Brace CS, Rensing N, Clifton P, Wozniak DF, Erik D, Yamada KA, Imai S. Sirt1 extends life span and delays aging in mice through the regulation of Nk2 homeobox 1 in the DMH and LH. Cell Metab. 2013;18:416–430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kanfi Y, Naiman S, Amir G, Peshti V, Zinman G, Nahum L, Bar-Joseph Z, Cohen HY. The sirtuin SIRT6 regulates lifespan in male mice. Nature. 2012;483:218–221. [DOI] [PubMed] [Google Scholar]
- 16.Mitchell SJ, Martin-Montalvo A, Mercken EM, Palacios HH, Ward TM, Abulwerdi G, Minor RK, Vlasuk GP, Ellis JL, Sinclair D a, Dawson J, Allison DB, Zhang Y, Becker KG, Bernier M, de Cabo R. The SIRT1 activator SRT1720 extends lifespan and improves health of mice fed a standard diet. Cell Rep. 2014;6:836–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mercken E, Mitchell S, Martin-Montalvo A, Minor R, Almeida M, Gomes A, Scheibye-Knudson M, Palacios H, Licata J, Zhang Y, Becker K, Khraiwesh H, Gonzalez-Reyes J, Villalba J, Baur J, Elliott P, Westphal C, Vlasuk G, Ellis J, Sinclair D, Bernier M, de Cabo R. SRT2104 extends survival of male mice on a standard diet and preserves bone and muscle mass. Aging Cell. 2014;13:787–796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Chang HC, Guarente L. SIRT1 and other sirtuins in metabolism. Trends Endocrinol Metab. 2014;25:138–145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Vaziri H, Dessain S, Ng Eaton E, Imai S, Frye R, Pandita T, Guarente L, Weinberg R. SIRT1 functions as an NAD-dependent p53 deacetylase. Cell. 2001;107:149–159. [DOI] [PubMed] [Google Scholar]
- 20.Yeung F, Hoberg J, Ramsey C, Keller M, Jones D, Frye R, Mayo M. Modulation of NF-[kappa]B-dependent transcription and cell survival by the SIRT1 deacetylase. EMBO J. 2004;23:2369–2380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mouchiroud L, Houtkooper RH, Moullan N, Katsyuba E, Ryu D, Cantó C, Mottis A, Jo Y, Viswanathan M, Schoonjans K, Guarente L, Auwerx J. The NAD+/sirtuin pathway modulates longevity through activation of mitochondrial UPR and FOXO signaling. Cell. 2013;154:430–441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rodgers JT, Lerin C, Haas W, Gygi SP, Spiegelman BM, Puigserver P. Nutrient control of glucose homeostasis through a complex of PGC-1a and SIRT1. Nature. 2005;434:113–118. [DOI] [PubMed] [Google Scholar]
- 23.Rajamohan SB, Pillai VB, Gupta M, Sundaresan NR, Birukov KG, Samant S, Hottiger MO, Gupta MP. SIRT1 Promotes Cell Survival under Stress by Deacetylation-Dependent Deactivation of Poly(ADP-Ribose) Polymerase 1. Mol Cell Biol. 2009;29:4116–4129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hirschey MD, Shimazu T, Goetzman E, Jing E, Lombard DB, Grueter C a, Harris C, Biddinger S, Ilkayeva OR, Stevens RD, Li Y, Saha AK, Ruderman NB, James R, Newgard CB, RVF Jr, Alt FW, Kahn CR, Verdin E. SIRT3 regulates fatty acid oxidation via reversible enzyme deacetylation. Nature. 2010;464:121–125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Haigis MC, Mostoslavsky R, Haigis K, Fahie K, Christodoulou D, Murphy A, Valenzuela D, Yancopoulos G, Karow M, Blander G, Wolberger C, Prolla T, Weindruch R, Alt F, Guarente L. SIRT4 inhibits glutamate dehydrogenase and opposes the effects of calorie restriction in pancreatic beta cells. Cell. 2006;126:941–954. [DOI] [PubMed] [Google Scholar]
- 26.Rardin M, He W, Nishida Y, Newman J, Carrico C, Danielson SR, Guo A, Gut P, Sahu AK, Li B, Fitch M, Riiff T, Zhu L, Zhou J, Mulhern D, Robert D, Ilkayeva OR, Newgard CB, Jacobson MP, Goetzman ES, Gibson BW, Verdin E. SIRT5 regulates the mitochondrial lysine succinylome and metabolic networks. Cell Metab. 2013;18:920–933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Li L, Shi L, Yang S, Yan R, Zhang D, Yang J, He L, Li W, Yi X, Sun L, Liang J, Cheng Z, Shi L, Shang Y, Yu W. SIRT7 is a histone desuccinylase that functionally links to chromatin compaction and genome stability. Nat Commun. 2016;7:1–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.McCord RA, Michishita E, Hong T, Berber E, Boxer LD, Kusumoto R, Guan S, Shi X, Gozani O, Burlingame AL, Bohr VA, Chua KF. SIRT6 stabilizes DNA-dependent protein kinase at chromatin for DNA double-strand break repair. Aging (Albany NY). 2009;1:109–121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Du J, Zhou Y, Su X, Yu JJ, Khan S, Jiang H, Kim JH, Woo J, Kim JH, Choi BH, He B, Chen W, Zhang S, Cerione RA, Auwerx J, Hao Q, Lin H. Sirt5 is a NAD-dependent protein lysine demalonylase and desuccinylase. Science (80- ). 2011;334:806–809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Tanner KG, Landry J, Sternglanz R, Denu JM. Silent information regulator 2 family of NAD- dependent histone/protein deacetylases generates a unique product, 1-O-acetyl-ADP-ribose. Proc Natl Acad Sci USA. 2000;97:14178–14182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Howitz KT, Bitterman K, Cohen H, Lamming D, Lavu S, Wood J, Zipkin R, Chung P, Kisielewski A, Zhang L, Scherer B, Sinclair D. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature. 2003;425:191–196. [DOI] [PubMed] [Google Scholar]
- 32.Milne JC, Lambert PD, Schenk S, Carney DP, Smith JJ, Gagne DJ, Jin L, Boss O, Perni RB, Vu CB, Bemis JE, Disch JS, Ng PY, Nunes JJ, Lynch A V, Yang H, Galonek H, Israelian K, Choy W, Iffland A, Lavu S, Sinclair DA, Olefsky JM, Jirousek MR, Elliott PJ, Westphal CH. Small molecule activators of SIRT1 as therapeutics for the treatment of type 2 diabetes. Nature. 2007;450:712–716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Hubbard BP, Gomes AP, Dai H, Li J, Case AW, Considine T, Riera T V, Lee JE, E SY, Lamming DW, Pentelute BL, Schuman ER, Stevens L a, Ling AJY, Armour SM, Michan S, Zhao H, Jiang Y, Sweitzer SM, Blum C a, Disch JS, Ng PY, Howitz KT, Rolo AP, Hamuro Y, Moss J, Perni RB, Ellis JL, Vlasuk GP, Sinclair D a. Evidence for a Common Mechanism of SIRT1 Regulation by Allosteric Activators. Science (80- ). 2013;339:1216–1219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Dai H, Case A, Riera T, Considine T, Lee J, Hamuro Y, Zhao H, Jiang Y, Sweitzer S, Pietrak B, Schwartz B, Blum C, Disch J, Caldwell R, Szczepankiewicz B, Oalmann C, Yee Ng P, White B, Casaubon R, Narayan R, Koppetsch K, Bourbonais F, Wu B, Wang J, Qian D, Jiang F, Mao C, Wang M, Hu E, Wu J, Perni R, Vlasuk G, Ellis J. Crystallographic structure of a small molecule SIRT1 activator-enzyme complex. Nat Commun. 2015;6:7645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sinclair D, Guarente L. Small-Molecule Allosteric Activators of Sirtuins. Annu Rev Pharmacol Toxicol. 2014;54:363–380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chen T, Li J, Liu J, Li N, Wang S, Liu H, Zeng M, Zhang Y, Bu P. Activation of SIRT3 by resveratrol ameliorates cardiac fibrosis and improves cardiac function via the TGF-β/Smad3 pathway. Am J Physiol - Hear Circ Physiol. 2015;308:H424–H434. [DOI] [PubMed] [Google Scholar]
- 37.Gertz M, Nguyen GTT, Fischer F, Suenkel B, Schlicker C, Fränzel B, Tomaschewski J, Aladini F, Becker C, Wolters D, Steegborn C. A Molecular Mechanism for Direct Sirtuin Activation by Resveratrol. PLoS One. 2012;7:1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sajish M, Schimmel P. A human tRNA synthetase is a potent PARP1-activating effector target for resveratrol. Nature. 2015;519:370–373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Cantó C, Gerhart-Hines Z, Feige JN, Lagouge M, Noriega L, Milne JC, Elliott PJ, Puigserver P, Auwerx J. AMPK regulates energy expenditure by modulating NAD + metabolism and SIRT1 activity. Nature. 2009;458:1056–1060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Pillai VB, Samant S, Sundaresan NR, Raghuraman H, Kim G, Bonner MY, Arbiser JL, Walker DI, Jones DP, Gius D, Gupta MP. Honokiol blocks and reverses cardiac hypertrophy in mice by activating mitochondrial SIRT3. Nat Commun. 2015;6:6656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Pillai VB, Kanwal A, Fang YH, Sharp WW, Samant S, Arbiser J, Gupta MP, Pillai VB, Kanwal A, Fang YH, Sharp WW, Samant S, Arbiser J, Gupta MP. Honokiol, an activator of Sirtuin-3 (SIRT3) preserves mitochondria and protects the heart from doxorubicin-induced cardiomyopathy in mice. Oncotarget. 2017;8:34082–34098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Avtanski DB, Nagalingam A, Bonner MY, Arbiser JL, Saxena NK, Sharma D. Honokiol activates LKB1-miR-34a axis and antagonizes the oncogenic actions of leptin in breast cancer. Oncotarget. 2015;6:29947–29962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Seo MS, Kim JH, Kim HJ, Chang KC, Park SW. Honokiol activates the LKB1-AMPK signaling pathway and attenuates the lipid accumulation in hepatocytes. Toxicol Appl Pharmacol. 2015;284:113–124. [DOI] [PubMed] [Google Scholar]
- 44.Hershberger K, Martin A, Hirschey M. Role of NAD+ and mitochondrial sirtuins in cardiac and renal diseases. Nat Rev Nephrol. 2017;13:213–225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Anderson RM, Bitterman KJ, Wood JG, Medvedik O, Sinclair DA. Nicotinamide and PNC1 govern lifespan extension by calorie restriction in Saccharomyces cerevisiae. Nature. 2003;423:181–185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Camacho-Pereira J, Tarragó MG, Chini CCS, Nin V, Escande C, Warner GM, Puranik AS, Schoon RA, Reid JM, Galina A, Chini EN. CD38 dictates age-related NAD decline and mitochondrial dysfunction through an SIRT3-dependent mechanism. Cell Metab. 2016;23:1127–1139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Zhu X-H, Lu M, Lee B-Y, Ugurbil K, Chen W. In vivo NAD assay reveals the intracellular NAD contents and redox state in healthy human brain and their age dependences. Proc Natl Acad Sci. 2015;112:2876–2881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Braidy N Age related changes in NAD+ metabolism oxidative stress and Sirt1 activity in Wistar rats. PLoS One. 2011;6:e19194. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 49.Rajman L, Chwalek K, Sinclair D. Therapeutic potential of NAD-boosting molecules: the in vivo evidence. Cell Metab. 2018;27:529–547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Revollo JR, Grimm AA, Imai S. The regulation of nicotinamide adenine dinucleotide biosynthesis by Nampt/PBEF/visfatin in mammals. Curr Opin Gastroenterol. 2007;23:164–170. [DOI] [PubMed] [Google Scholar]
- 51.Mori V, Amici A, Mazzola F, Di Stefano M, Conforti L, Magni G, Ruggieri S, Raffaelli N, Orsomando G. Metabolic profiling of alternative NAD biosynthetic routes in mouse tissues. PLoS One. 2014;9:1–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Yang H, Yang T, Baur J a, Perez E, Matsui T, Carmona JJ, Lamming DW, Souza-Pinto NC, Bohr V a, Rosenzweig A, de Cabo R, Sauve A a, Sinclair D a. Nutrient-sensitive mitochondrial NAD+ levels dictate cell survival. Cell. 2007;130:1095–1107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Escande C Flavonoid apigenin is an inhibitor of the NAD+ase CD38: implications for cellular NAD+ metabolism, protein acetylation, and treatment of metabolic syndrome. Diabetes. 2013;62:1084–1093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Haffner CD, Ha CD, Becherer JD, Boros EE, Cadilla R, Cowan D, Deaton DN, Guo Y, Harrington W, Henke BR, Jeune MR, Kaldor I, Milliken N, Petrov KG, Preugschat F, Schulte C, Shearer BG, Shearer T, Smalley TL, Stewart EL, Stuart JD, Ulrich JC, Drive M, Box PO, Carolina N. Discovery, synthesis, and biological evaluation of thiazoloquin(az)olin(on)es as potent CD38 inhibitors. J Med Chem. 2015;58:3548–3571. [DOI] [PubMed] [Google Scholar]
- 55.Chini E CD38 as a Regulator of Cellular NAD: A Novel Potential Pharmacological Target for Metabolic Conditions. Curr Pharm Des. 2009;15:57–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Essuman K, Summers DW, Sasaki Y, Mao X, Diantonio A, Milbrandt J, Essuman K, Summers DW, Sasaki Y, Mao X, Diantonio A, Milbrandt J. The SARM1 Toll / Interleukin-1 Receptor Domain Possesses Intrinsic NAD + Cleavage Activity that Promotes Pathological Axonal Degeneration. Neuron. 2017;93:1334–1343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Gerdts J, Brace EJ, Sasaki Y, Diantonio A, Milbrandt J, Louis S, Louis S, Louis S. SARM1 activation triggers axon degeneration locally via NAD+ destruction. Science (80- ). 2015;348:453–457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Brown JS, Kaye SB, Yap TA. PARP inhibitors: The race is on. Br J Cancer. 2016;114:713–715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Canto C, Houtkooper RH, Pirinen E, Youn DY, Oosterveer MH, Cen Y, Fernandez-Marcos PJ, Yamamoto H, Andreux PA, Cettour-Rose P, Gademann K, Rinsch C, Schoonjans K, Sauve AA, Auwerx J. The NAD+ precursor nicotinamide riboside enhances oxidative metabolism and protects against high-fat diet-induced obesity. Cell Metab. 2012;15:838–847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.de Picciotto NE, Gano LB, Johnson LC, Martens CR, Sindler AL, Mills KF, Imai S ichiro, Seals DR. Nicotinamide mononucleotide supplementation reverses vascular dysfunction and oxidative stress with aging in mice. Aging Cell. 2016;15:522–530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Das A, Huang GX, Bonkowski MS, Longchamp A, Li C, Schultz MB, Kim L-J, Osborne B, Joshi S, Lu Y, Treviño-Villarreal JH, Kang M-J, Hung T, Lee B, Williams EO, Igarashi M, Mitchell JR, Wu LE, Turner N, Arany Z, Guarente L, Sinclair DA. Impairment of an Endothelial NAD+-H2S Signaling Network Is a Reversible Cause of Vascular Aging. Cell. 2018;173:74–89.e20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Martin AS, Abraham DM, Hershberger KA, Bhatt DP, Mao L, Cui H, Liu J, Liu X, Muehlbauer MJ, Grimsrud PA, Locasale JW, Payne RM, Hirschey MD. Nicotinamide mononucleotide requires SIRT3 to improve cardiac function and bioenergetics in a Friedreich ‘ s ataxia cardiomyopathy model. JCI Insight. 2017;2:e93885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Lee CF, Chavez JD, Garcia-menendez L, Chiao YA, Edgar JS, Goodlett DR, Bruce JE, Roe ND, Chiao YA, Edgar JS, Ah Y, Goodlett DR, Bruce JE, Tian R. Normalization of NAD+ Redox Balance as a Therapy for Heart Failure. Circulation. 2016;134:883–894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Karamanlidis G, Lee CF, Garcia-Menendez L, Kolwicz SC, Suthammarak W, Gong G, Sedensky MM, Morgan PG, Wang W, Tian R, Kolwicz SC Jr, Suthammarak W, Gong G, Sedensky MM, Morgan PG, Wang W, Tian R. Mitochondrial complex i deficiency increases protein acetylation and accelerates heart failure. Cell Metab. 2013;18:239–250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Yamamoto T, Byun J, Zhai P, Ikeda Y, Oka S, Sadoshima J. Nicotinamide mononucleotide, an intermediate of NAD+ synthesis, protects the heart from ischemia and reperfusion. PLoS One. 2014;9:e98972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Ryu D, Zhang H, Ropelle ER, Sorrentino V, Davi A, Mázala G, Mouchiroud L, Marshall PL, Campbell MD, Ali AS, Knowels GM, Bellemin S, Iyer SR, Wang X, Gariani K, Sauve AA, Cantó C, Conley KE, Walter L, Richard M, Chin ER, Jasmin BJ, Marcinek DJ, Menzies KJ, Mázala DAG, Mouchiroud L, Marshall PL, Campbell MD, Ali AS, Knowels GM, Bellemin S, Iyer SR, Wang X, Gariani K, Sauve AA, Cantó C, Conley KE, Walter L, Lovering RM, Chin ER, Jasmin BJ, Marcinek DJ, Menzies KJ, Auwerx J. NAD+ repletion improves muscle function in muscular dystrophy and counters global PARylation. Sci Transl Med. 2016;8:361ra139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Cappetta D, Esposito G, Piegari E, Russo R, Ciuffreda LP, Rivellino A, Berrino L, Rossi F, De Angelis A, Urbanek K. SIRT1 activation attenuates diastolic dysfunction by reducing cardiac fibrosis in a model of anthracycline cardiomyopathy. Int J Cardiol. 2016;205:99–110. [DOI] [PubMed] [Google Scholar]
- 68.Gu XS, Wang ZB, Ye Z, Lei JP, Li L, Su DF, Zheng X. Resveratrol, an activator of SIRT1, upregulates AMPK and improves cardiac function in heart failure. Genet Mol Res. 2014;13:323–35. [DOI] [PubMed] [Google Scholar]
- 69.Rimbaud S, Ruiz M, Piquereau J, Mateo P, Fortin D, Veksler V, Garnier A, Ventura-Clapier R. Resveratrol improves survival, hemodynamics and energetics in a rat model of hypertension leading to heart failure. PLoS One. 2011;6:1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Thandapilly SJ, Wojciechowski P, Behbahani J, Louis XL, Yu L, Juric D, Kopilas MA, Anderson HD, Netticadan T. Resveratrol prevents the development of pathological cardiac hypertrophy and contractile dysfunction in the SHR without lowering blood pressure. Am J Hypertens. 2010;23:192–196. [DOI] [PubMed] [Google Scholar]
- 71.Tanno M, Kuno A, Yano T, Miura T, Hisahara S, Ishikawa S, Shimamoto K, Horio Y. Induction of manganese superoxide dismutase by nuclear translocation and activation of SIRT1 promotes cell survival in chronic heart failure. J Biol Chem. 2010;285:8375–8382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Sulaiman M, Matta MJ, Sunderesan NR, Gupta MP, Periasamy M, Gupta M. Resveratrol, an activator of SIRT1, upregulates sarcoplasmic calcium ATPase and improves cardiac function in diabetic cardiomyopathy. AJP Hear Circ Physiol. 2010;298:H833–H843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Yoshida Y, Shioi T, Izumi T. Resveratrol ameliorates experimental autoimmune myocarditis. Circ J. 2007;71:397–404. [DOI] [PubMed] [Google Scholar]
- 74.Csiszar A, Labinskyy N, Podlutsky A, Kaminski PM, Wolin MS, Zhang C, Mukhopadhyay P, Pacher P, Hu F, De R, Ballabh P, Ungvari Z. Vasoprotective effects of resveratrol and SIRT1: attenuation of cigarette smoke-induced oxidative stress and proinflammatory phenotypic alterations. Am J Physiol Hear Circ Physiol. 2008;294:H2721–H2735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Mattison JA, Wang M, Bernier M, Zhang J, Park S-S, Maudsley S, An S, Santhanam L, Martin B, Faulkner S, Morrell C, Baur J, Peshkin L, Sosnowska D, Csiszar A, Herbert R, Tilmont E, Ungvari Z, Pearson K, Lakatta E, de Cabo R. Resveratrol prevents high fat/sucrose diet-induced central arterial wall inflammation and stiffening in nonhuman primates. Cell Metab. 2014;20:183–190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Fukuda S, Kaga S, Zhan L, Bagchi D, Das DK, Bertelli A, Maulik N. Resveratrol ameliorates myocardial damage by inducing vascular endothelial growth factor-angiogenesis and tyrosine kinase receptor Flk-1. Cell Biochem Biophys. 2006;44:43–49. [DOI] [PubMed] [Google Scholar]
- 77.Shalwala M, Zhu SG, Das A, Salloum FN, Xi L, Kukreja RC. Sirtuin 1 (SIRT1) activation mediates sildenafil induced delayed cardioprotection against ischemia-reperfusion injury in mice. PLoS One. 2014;9:3–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Hung LM, Su MJ, Chen JK. Resveratrol protects myocardial ischemia-reperfusion injury through both NO-dependent and NO-independent mechanisms. Free Radic Biol Med. 2004;36:774–781. [DOI] [PubMed] [Google Scholar]
- 79.Jimenez-Gomez Y Resveratrol improves adipose insulin signaling and reduces the inflammatory response in adipose tissue of rhesus monkeys on high-fat, high-sugar diet. Cell Metab. 2013;18:533–545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Baur JA, Pearson KJ, Price NL, Jamieson H a, Lerin C, Kalra A, Prabhu VV, Allard JS, Lopez-Lluch G, Lewis K, Pistell PJ, Poosala S, Becker KG, Boss O, Gwinn D, Wang M, Ramaswamy S, Fishbein KW, Spencer RG, Lakatta EG, Le Couteur D, Shaw RJ, Navas P, Puigserver P, Ingram DK, de Cabo R, Sinclair D a. Resveratrol improves health and survival of mice on a high-calorie diet. Nature. 2006;444:337–342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Lagouge M, Argmann C, Gerhart-Hines Z, Meziane H, Lerin C, Daussin F, Messadeq N, Milne J, Lambert P, Elliott P, Geny B, Laakso M, Puigserver P, Auwerx J. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1[alpha]. Cell. 2006;127:1109–1122. [DOI] [PubMed] [Google Scholar]
- 82.Feige JN, Lagouge M, Canto C, Strehle A, Houten SM, Milne JC, Lambert PD, Mataki C, Elliott PJ, Auwerx J. Specific SIRT1 activation mimics low energy levels and protects against diet-induced metabolic disorders by enhancing fat oxidation. Cell Metab. 2008;8:347–358. [DOI] [PubMed] [Google Scholar]
- 83.Minor RK, Baur JA, Gomes AP, Ward TM, Csiszar A, Mercken EM, Abdelmohsen K, Shin YK, Canto C, Scheibye-Knudsen M, Krawczyk M, Irusta PM, Martin-Montalvo A, Hubbard BP, Zhang YQ, Lehrmann E, White AA, Price NL, Swindell WR, Pearson KJ, Becker KG, Bohr VA, Gorospe M, Egan JM, Talan MI, Auwerx J, Westphal CH, Ellis JL, Ungvari Z, Vlasuk GP, Elliott PJ, Sinclair DA, de Cabo R. SRT1720 improves survival and healthspan of obese mice. Sci Rep. 2011;70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Gano LB, Donato AJ, Pasha HM, Hearon CM, Sindler AL, Seals DR. The SIRT1 activator SRT1720 reverses vascular endothelial dysfunction, excessive superoxide production, and inflammation with aging in mice. AJP Hear Circ Physiol. 2014;307:H1754–H1763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Miranda M, van Tits L, Lohmann C, Arsiwala T, Winnik S, Tailleux A, Stein S, Gomes A, Suri V, Ellis J, Lutz T, Hottiger M, Sinclair D, Auwerx J, Schoonjans K, Staels B, Luscher T, Matter C. The Sirt1 activator SRT3025 provides atheroprotection in Apoe−/− mice by reducing hepatic Pcsk9 secretion and enhancing Ldlr expression. Eur Hear J. 2015;36:51–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Yang Y, Duan W, Lin Y, Yi W, Liang Z, Yan J, Wang N, Deng C, Zhang S, Li Y, Chen W, Yu S, Yi D, Jin Z. SIRT1 activation by curcumin pretreatment attenuates mitochondrial oxidative damage induced by myocardial ischemia reperfusion injury. Free Radic Biol Med. 2013;65:667–679. [DOI] [PubMed] [Google Scholar]
- 87.Zakaria EM, El-Bassossy HM, El-Maraghy NN, Ahmed AF, Ali AA. PARP-1 inhibition alleviates diabetic cardiac complications in experimental animals. Eur J Pharmacol. 2016;791:444–454. [DOI] [PubMed] [Google Scholar]
- 88.Boslett J, Hemann C, Zhao Y, Lee H, Zweier J. Luteolinidin Protects the Postischemic Heart through CD38 Inhibition with Preservation of NAD(P)(H). J Pharmacol Exp Ther. 2017;361:99–108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Escande C, Nin V, Price NL, Capellini V, Gomes AP, Barbosa MT, O’Neil L, White TA, Sinclair DA, Chini EN. Flavonoid apigenin is an inhibitor of the NAD+ase CD38: Implications for cellular NAD+ metabolism, protein acetylation, and treatment of metabolic syndrome. Diabetes. 2013;62:1084–1093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Wang JC, Bennett M. Aging and atherosclerosis: Mechanisms, functional consequences, and potential therapeutics for cellular senescence. Circ Res. 2012;111:245–259. [DOI] [PubMed] [Google Scholar]
- 91.Winnik S, Auwerx J, Sinclair DA, Matter CM. Protective effects of sirtuins in cardiovascular diseases: From bench to bedside. Eur Heart J. 2015;36:3404–3412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Hansson GK. Inflammation, atherosclerosis, and coronary artery disease. N Engl J Med. 2005;352:1685–95. [DOI] [PubMed] [Google Scholar]
- 93.Thompson AM, Wagner R, Rzucidlo EM. Age-related loss of SirT1 expression results in dysregulated human vascular smooth muscle cell function. AJP Hear Circ Physiol. 2014;307:H533–H541. [DOI] [PubMed] [Google Scholar]
- 94.Gorenne I, Kumar S, Gray K, Figg N, Yu H, Mercer J, Bennett M. Vascular smooth muscle cell sirtuin 1 protects against dna damage and inhibits atherosclerosis. Circulation. 2013;127:386–396. [DOI] [PubMed] [Google Scholar]
- 95.Breitenstein A, Wyss CA, Spescha RD, Franzeck FC, Hof D, Riwanto M, Hasun M, Akhmedov A, von Eckardstein A, Maier W, Landmesser U, Lüscher TF, Camici GG. Peripheral Blood Monocyte Sirt1 Expression Is Reduced in Patients with Coronary Artery Disease. PLoS One. 2013;8:8–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Rodella LF, Favero G, Rossini C, Foglio E, Bonomini F, Reiter RJ, Rezzani R. Aging and vascular dysfunction: Beneficial melatonin effects. Age (Omaha). 2013;35:103–115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Zhang QJ, Wang Z, Chen HZ, Zhou S, Zheng W, Liu G, Wei YS, Cai H, Liu DP, Liang CC. Endothelium-specific overexpression of class III deacetylase SIRT1 decreases atherosclerosis in apolipoprotein E-deficient mice. Cardiovasc Res. 2008;80:191–199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Qiang L, Lin HV., Kim-Muller JY, Welch CL, Gu W, Accili D. Proatherogenic abnormalities of lipid metabolism in SirT1 transgenic mice are mediated through Creb deacetylation. Cell Metab. 2011;14:758–767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Thompson AM, Martin KA, Rzucidlo EM. Resveratrol induces vascular smooth muscle cell differentiation through stimulation of SirT1 and AMPK. PLoS One. 2014;9:1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Barbieri SS, Amadio P, Gianellini S, Tarantino E, Zacchi E, Veglia F, Howe LR, Weksler BB, Mussoni L, Tremoli E. Cyclooxygenase-2-derived prostacyclin regulates arterial thrombus formation by suppressing tissue factor in a sirtuin-1-dependent-manner. Circulation. 2012;126:1373–1384. [DOI] [PubMed] [Google Scholar]
- 101.Breitenstein A, Stein S, Holy EW, Camici GG, Lohmann C, Akhmedov A, Spescha R, Elliott PJ, Westphal CH, Matter CM, Lscher TF, Tanner FC. Sirt1 inhibition promotes in vivo arterial thrombosis and tissue factor expression in stimulated cells. Cardiovasc Res. 2011;89:464–472. [DOI] [PubMed] [Google Scholar]
- 102.Winnik S, Gaul DS, Preitner F, Lohmann C, Weber J, Miranda MX, Liu Y, Van Tits LJ, Mateos JM, Brokopp CE, Auwerx J, Thorens B, Lüscher TF, Matter CM. Deletion of Sirt3 does not affect atherosclerosis but accelerates weight gain and impairs rapid metabolic adaptation in LDL receptor knockout mice: Implications for cardiovascular risk factor development. Basic Res Cardiol. 2014;109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Dong C, Della-Morte D, Wang L, Cabral D, Beecham A, McClendon MS, Luca CC, Blanton SH, Sacco RL, Rundek T. Association of the sirtuin and mitochondrial uncoupling protein genes with carotid plaque. PLoS One. 2011;6:e27157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Nencioni A, da Silva R, Fraga-Silva R, Steffens S, Fabre M, Bauer I, Caffa I, Magnone M, Sociali G, Quercioli A, Pelli G, Lenglet S, Galan K, Burger F, Vázquez Calvo S, Bertolotto M, Bruzzone S, Ballestrero A, Patrone F, Dallegri F, Santos R, Stergiopulos N, Mach F, Vuilleumier N, Montecucco F. Nicotinamide phosphoribosyltransferase inhibition reduces intraplaque CXCL1 production and associated neutrophil infiltration in atherosclerotic mice. Thromb Haemost. 2014;111:189–380. [DOI] [PubMed] [Google Scholar]
- 105.Fleg JL, Strait J. Age-associated changes in cardiovascular structure and function: a fertile milieu for future disease. Hear Fail Rev. 2012;17:545–554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Gavish B, Izzo JL. Arterial Stiffness: Going a Step beyond. Am J Hypertens. 2016;29:1223–1233. [DOI] [PubMed] [Google Scholar]
- 107.Stein S, Schäfer N, Breitenstein A, Besler C, Winnik S, Lohmann C, Heinrich K, Brokopp CE, Handschin C, Landmesser U, Tanner FC, Lüscher TF, Matter CM. SIRT1 reduces endothelial activation without affecting vascular function in ApoE−/−mice. Aging (Albany NY). 2010;2:344–352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Donato AJ, Magerko KA, Lawson BR, Durrant JR, Lesniewski LA, Seals DR. SIRT-1 and vascular endothelial dysfunction with ageing in mice and humans. J Physiol. 2011;589:4545–4554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Mattagajasingh I, Kim C-S, Naqvi A, Yamamori T, Hoffman TA, Jung S-B, DeRicco J, Kasuno K, Irani K. SIRT1 promotes endothelium-dependent vascular relaxation by activating endothelial nitric oxide synthase. Proc Natl Acad Sci. 2007;104:14855–14860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Ota H, Eto M, Ogawa S, Iijima K, Akishita M, Ouchi Y. SIRT1/eNOS axis as a potential target against vascular senescence, dysfunction and atherosclerosis. J Atheroscler Thromb. 2010;17:431–435. [DOI] [PubMed] [Google Scholar]
- 111.Zu Y, Liu L, Lee MYK, Xu C, Liang Y, Man RY, Vanhoutte PM, Wang Y. SIRT1 promotes proliferation and prevents senescence through targeting LKB1 in primary porcine aortic endothelial cells. Circ Res. 2010;106:1384–1393. [DOI] [PubMed] [Google Scholar]
- 112.Liu R, Liu H, Ha Y, Tilton RG, Zhang W. Oxidative Stress Induces Endothelial Cell Senescence via Downregulation of Sirt6. Biomed Res Int. 2014;2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Cardus A, Uryga AK, Walters G, Erusalimsky JD. SIRT6 protects human endothelial cells from DNA damage, telomere dysfunction, and senescence. Cardiovasc Res. 2013;97:571–579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Lahteenvuo J, Rosenzweig A. The Role of Angiogenesis in Cardiovascular Aging. Circ Res. 2012;110:1252–1264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Ware JA, Simons M. Angiogenesis in ischemic heart disease. Nat Med. 1997;3:158–164. [DOI] [PubMed] [Google Scholar]
- 116.Tabibiazar R, Rockson SG. Angiogenesis and the ischaemic heart. Eur Heart J. 2001;22:903–918. [DOI] [PubMed] [Google Scholar]
- 117.Guarani V, Deflorian G, Franco CA, Bentley K, Toussaint L, Dequiedt F, Mostoslavsky R, Schmidt HH, Zimmermann B, Brandes RP, Mione M, Christoph H, Braun T, Zeiher AM, Gerhardt H, Dimmeler S, Potente M. Acetylation-dependent regulation of endothelial Notch signalling by the SIRT1 deacetylase. Nature. 2011;473:234–238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Potente M, Ghaeni L, Baldessari D, Mostoslavsky R, Rossig L, Dequiedt F, Haendeler J, Mione M, Dejana E, Alt FW, Zeiher AM, Dimmeler S. SIRT1 controls endothelial angiogenic functions during vascular growth. Genes Dev. 2007;21:2644–2658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Maizel J, Xavier S, Chen J, Lin CHS, Vasko R, Goligorsky MS. Sirtuin 1 ablation in endothelial cells is associated with impaired angiogenesis and diastolic dysfunction. AJP Hear Circ Physiol. 2014;307:H1691–H1704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Zeng H, Li L, Chen JX. Loss of Sirt3 limits bone marrow cell-mediated angiogenesis and cardiac repair in post-myocardial infarction. PLoS One. 2014;9:e107011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Longchamp A, Mirabella T, Arduini A, MacArthur MR, Das A, Treviño-Villarreal JH, Hine C, Ben-Sahra I, Knudsen NH, Brace LE, Reynolds J, Mejia P, Tao M, Sharma G, Wang R, Corpataux J-M, Haefliger J-A, Ahn KH, Lee C-H, Manning BD, Sinclair DA, Chen CS, Ozaki CK, Mitchell JR. Amino Acid Restriction Triggers Angiogenesis via GCN2/ATF4 Regulation of VEGF and H 2 S Production. Cell. 2018;173:117–129.e14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Porter GA, Urciuoli WR, Brookes PS, Nadtochiy SM. SIRT3 deficiency exacerbates ischemia-reperfusion injury: implication for aged hearts. AJP Hear Circ Physiol. 2014;306:H1602–H1609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Hsu C-PP, Oka S, Shao D, Hariharan N, Sadoshima J. Nicotinamide phosphoribosyltransferase regulates cell survival through nad+ synthesis in cardiac myocytes. Circ Res. 2009;105:481–491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Hsu C-P, Zhai P, Yamamoto T, Maejima Y, Matsushima S, Hariharan N, Shao D, Takagi H, Oka S, Sadoshima J. Sirt1 Protects the Heart from Ischemia Reperfusion. Circulation. 2010;122:2170–2182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Klishadi MS, Zarei F, Hejazian SH, Moradi A, Hemati M, Safari F. Losartan protects the heart against ischemia reperfusion injury: Sirtuin3 involvement. J Pharm Pharm Sci. 2015;18:112–123. [DOI] [PubMed] [Google Scholar]
- 126.Liu B, Che W, Xue J, Zheng C, Tang K, Zhang J, Wen J, Xu Y. SIRT4 prevents hypoxia-induced apoptosis in H9c2 cardiomyoblast cells. Cell Physiol Biochem. 2013;32:655–662. [DOI] [PubMed] [Google Scholar]
- 127.Chen C-J, Yu W, Fu Y-C, Wang X, Li J-L, Wang W. Resveratrol protects cardiomyocytes from hypoxia-induced apoptosis through the SIRT1-FoxO1 pathway. Biochem Biophys Res Commun. 2009;378:389–93. [DOI] [PubMed] [Google Scholar]
- 128.Sundaresan NR, Samant SA, Pillai VB, Rajamohan SB, Gupta MP. SIRT3 Is a Stress-Responsive Deacetylase in Cardiomyocytes That Protects Cells from Stress-Mediated Cell Death by Deacetylation of Ku70. Mol Cell Biol. 2008;28:6384–6401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Boylston JA, Sun J, Chen Y, Gucek M, Sack MN, Murphy E. Characterization of the cardiac succinylome and its role in ischemia-reperfusion injury. J Mol Cell Cardiol. 2015;88:73–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Liu B, Che W, Zheng C, Liu W, Wen J, Fu H, Tang K, Zhang J, Xu Y. SIRT5: A safeguard against oxidative stress-induced apoptosis in cardiomyocytes. Cell Physiol Biochem. 2013;32:1050–1059. [DOI] [PubMed] [Google Scholar]
- 131.Maksin-Matveev A, Kanfi Y, Hochhauser E, Isak A, Cohen HY, Shainberg A. Sirtuin 6 protects the heart from hypoxic damage. Exp Cell Res. 2015;330:81–90. [DOI] [PubMed] [Google Scholar]
- 132.Lynn EG, Mcleod CJ, Gordon JP, Bao J, Sack MN. SIRT2 is a negative regulator of anoxia-reoxygenation tolerance via regulation of 14–3-3 zeta and BAD in H9c2 cells. FEBS Lett. 2008;582:2857–2862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Wang J, Ruotsalainen S, Moilanen L, Lepisto P, Laakso M, Kuusisto J. The metabolic syndrome predicts cardiovascular mortality: A 13-year follow-up study in elderly non-diabetic Finns. Eur Heart J. 2007;28:857–864. [DOI] [PubMed] [Google Scholar]
- 134.Pacher P, Szabó C. Role of poly(ADP-ribose) polymerase 1 (PARP-1) in cardiovascular diseases: The therapeutic potential of PARP inhibitors. Cardiovasc Drug Rev. 2007;25:235–260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Steenman M, Lande G. Cardiac aging and heart disease in humans. Biophys Rev. 2017;9:131–137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Strait J, Lakatta E. Aging-associated cardiovascular changes and their relationship to heart failure. Heart Fail Clin. 2012;8:143–164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Olivetti G, Melissari M, Capasso J, Anversa P. Cardiomyopathy of the Aging Human Heart. Circ Res. 1991;68:1560–1569. [DOI] [PubMed] [Google Scholar]
- 138.Sundaresan N, Pillai VB, Gupta MP. Emerging roles of SIRT1 deacetylase in regulating cardiomyocyte survival and hypertrophy. J Mol Cell Cardiol. 2011;51:614–618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Kawashima T, Inuzuka Y, Okuda J, Kato T, Niizuma S, Tamaki Y, Iwanaga Y, Kawamoto A, Narazaki M, Matsuda T, Adachi S, Takemura G, Kita T, Kimura T, Shioi T. Constitutive SIRT1 overexpression impairs mitochondria and reduces cardiac function in mice. J Mol Cell Cardiol. 2011;51:1026–1036. [DOI] [PubMed] [Google Scholar]
- 140.Oka S, Alcendor R, Zhai P, Park JY, Shao D, Cho J, Yamomoto T, Tian B, Sadoshima J. PPARα-Sirt1 complex mediates cardiac hypertrophy and failure through suppression of the ERR transcriptional pathway. Cell Metab. 2011;14:598–611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Alcendor RR, Gao S, Zhai P, Zablocki D, Holle E, Yu X, Tian B, Wagner T, Vatner SF, Sadoshima J. Sirt1 regulates aging and resistance to oxidative stress in the heart. Circ Res. 2007;100:1512–1521. [DOI] [PubMed] [Google Scholar]
- 142.Planavila A, Iglesias R, Giralt M, Villarroya F. Sirt1 acts in association with PPARα to protect the heart from hypertrophy, metabolic dysregulation, and inflammation. Cardiovasc Res. 2011;90:276–284. [DOI] [PubMed] [Google Scholar]
- 143.Prola A, Da Silva JP, Guilbert A, Lecru L, Piquereau J, Ribeiro M, Mateo P, Gressette M, Fortin D, Boursier C, Gallerne C, Caillard A, Samuel JL, François H, Sinclair DA, Eid P, Ventura-Clapier R, Garnier A, Lemaire C. SIRT1 protects the heart from ER stress-induced cell death through eIF2α deacetylation. Cell Death Differ. 2017;24:343–356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Pillai VB, Bindu S, Sharp WW, Fang YH, Kim GH, Gupta M, Samant S, Gupta MP. Sirt3 protects mitochondrial DNA damage and blocks the development of doxorubicin-induced cardiomyopathy in Mice. Am J Physiol - Hear Circ Physiol. 2016;310:H962–H972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Chen T, Liu J, Li N, Wang S, Liu H, Li J, Zhang Y, Bu P. Mouse SIRT3 attenuates hypertrophy-related lipid accumulation in the heart through the deacetylation of LCAD. PLoS One. 2015;10:1–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Hafner A V, Dai J, Gomes AP, Xiao CY, Palmeira KCM, Rosenzweig A, Sinclair DA. Regulation of the mPTP by SIRT3-mediated deacetylation of CypD at lysine 166 suppresses age-related cardiac hypertrophy. Aging (Albany NY). 2010;2:914–923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Sundaresan N, Bindu S, Pillai V, Saman S, Pan Y, Huang J, Gupta M, Nagalingam R, Wolfgether D, Verdin E, Cgupta M. SIRT3 blocks aging-associated tissue fibrosis in mice by deacetylating and activating glycogen synthase kinase 3 β. Mol Cell Biol. 2016;36:678–692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Sundaresan NR, Gupta M, Kim G, Rajamohan SB, Isbatan A, Gupta MP. Sirt3 blocks the cardiac hypertrophic response by augmenting Foxo3a-dependent antioxidant defense mechanisms in mice. J Clin Invest. 2009;119:2758–2771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Samant SA, Zhang HJ, Hong Z, Pillai VB, Sundaresan NR, Wolfgeher D, Archer SL, Chan DC, Gupta MP. SIRT3 Deacetylates and Activates OPA1 To Regulate Mitochondrial Dynamics during Stress. Mol Cell Biol. 2014;34:807–819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Yu W, Gao B, Li N, Wang J, Qiu C, Zhang G, Liu M, Zhang R, Li C, Ji G, Zhang Y. Sirt3 deficiency exacerbates diabetic cardiac dysfunction: Role of Foxo3A-Parkin-mediated mitophagy. Biochim Biophys Acta - Mol Basis Dis. 2017;1863:1973–1983. [DOI] [PubMed] [Google Scholar]
- 151.Pillai VB, Sundaresan NR, Kim G, Gupta M, Rajamohan SB, Pillai JB, Samant S, Ravindra P V., Isbatan A, Gupta MP. Exogenous NAD blocks cardiac hypertrophic response via activation of the SIRT3-LKB1-AMP-activated kinase pathway. J Biol Chem. 2010;285:3133–3144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Zeng H, Vaka VR, He X, Booz GW, Chen JX. High-fat diet induces cardiac remodelling and dysfunction: Assessment of the role played by SIRT3 loss. J Cell Mol Med. 2015;19:1847–1856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Tang X, Chen XF, Wang NY, Wang XM, Liang ST, Zheng W, Lu YB, Zhao X, Hao DL, Zhang ZQ, Zou MH, Liu DP, Chen HZ. SIRT2 Acts as a Cardioprotective Deacetylase in Pathological Cardiac Hypertrophy. Circulation. 2017;136:2051–2067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Luo YX, Tang X, An XZ, Xie XM, Chen XF, Zhao X, Hao DL, Chen HZ, Liu DP. SIRT4 accelerates Ang II-induced pathological cardiac hypertrophy by inhibiting manganese superoxide dismutase activity. Eur Heart J. 2017;38:1389–1398. [DOI] [PubMed] [Google Scholar]
- 155.Sadhukhan S, Liu X, Ryu D, Nelson OD, Stupinski JA, Li Z, Chen W, Zhang S, Weiss RS, Locasale JW, Auwerx J, Lin H. Metabolomics-assisted proteomics identifies succinylation and SIRT5 as important regulators of cardiac function. Proc Natl Acad Sci. 2016;113:4320–4325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Sundaresan N, Vasudevan P, Zhong L, Kim G, Samant S, Parekh V, Pillai V, Ravindra P, Gupta M, Jeevanandam V, Cunningham J, Deng C, Lombard D, Mostoslavsky R, Gupta M. The sirtuin SIRT6 blocks IGF-Akt signaling and development of cardiac hypertrophy by targeting c-Jun. Nat Med. 2012;18:1643–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Cai Y, Yu SS, Chen SR, Pi RB, Gao S, Li H, Ye JT, Liu PQ. Nmnat2 protects cardiomyocytes from hypertrophy via activation of SIRT6. FEBS Lett. 2012;586:866–874. [DOI] [PubMed] [Google Scholar]
- 158.Ryu D, Jo YS, Lo Sasso G, Stein S, Zhang H, Perino A, Lee JU, Zeviani M, Romand R, Hottiger MO, Schoonjans K, Auwerx J. A SIRT7-dependent acetylation switch of GABPβ1 controls mitochondrial function. Cell Metab. 2014;20:856–869. [DOI] [PubMed] [Google Scholar]
- 159.Vakhrusheva O, Smolka C, Gajawada P, Kostin S, Boettger T, Kubin T, Braun T, Bober E. Sirt7 increases stress resistance of cardiomyocytes and prevents apoptosis and inflammatory cardiomyopathy in mice. Circ Res. 2008;102:703–710. [DOI] [PubMed] [Google Scholar]
- 160.Alcendor RR, Kirshenbaum LA, Imai SI, Vatner SF, Sadoshima J. Silent information regulator 2α, a longevity factor and class III histone deacetylase, is an essential endogenous apoptosis inhibitor in cardiac myocytes. Circ Res. 2004;95:971–980. [DOI] [PubMed] [Google Scholar]
- 161.Horton JL, Martin OJ, Lai L, Riley NM, Richards AL, Vega RB, Leone TC, Pagliarini DJ, Muoio DM, Bedi KC, Margulies KB, Coon JJ, Kelly DP. Mitochondrial protein hyperacetylation in the failing heart. JCI Insight. 2016;1:1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Vikram A, Lewarchik C, Yoon J, Naqvi A, Kumar S, Morgan G, Jacobs J, Li Q, Kim Y, Kassan M, Liu J, Gabani M, Kumar A, Mehdi H, Zhu X, Guan X, Kutschke W, Zhang X, Boudreau R, Dai S, Matasic D, Jung S, Margulies K, Kumar V, Bachschmid M, London B, Irani K. Sirtuin 1 regulates cardiac electrical activity by deacetylating the cardiac sodium channel. Nat med. 2017;23:361–367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.North BJ, Rosenberg MA, Jeganathan KB, Hafner AV., Michan S, Dai J, Baker DJ, Cen Y, Wu LE, Sauve AA, van Deursen JM, Rosenzweig A, Sinclair DA. SIRT2 induces the checkpoint kinase BubR1 to increase lifespan. EMBO J. 2014;33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.AlGhatrif M, Lakatta E. The Conundrum of Arterial Stiffness, Elevated blood pressure, and Aging. Curr Hypertens Rep. 2015;17:96–111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Buford TW. Hypertension and Aging. Ageing Res Rev. 2016;26:96–111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Gao P, Xu TT, Lu J, Li L, Xu J, Hao DL, Chen HZ, Liu DP. Overexpression of SIRT1 in vascular smooth muscle cells attenuates angiotensin II-induced vascular remodeling and hypertension in mice. J Mol Med. 2014;92:347–357. [DOI] [PubMed] [Google Scholar]
- 167.Paulin R, Dromparis P, Sutendra G, Gurtu V, Zervopoulos S, Bowers L, Haromy A, Webster L, Provencher S, Bonnet S, Michelakis ED. Sirtuin 3 deficiency is associated with inhibited mitochondrial function and pulmonary arterial hypertension in rodents and humans. Cell Metab. 2014;20:827–839. [DOI] [PubMed] [Google Scholar]
- 168.Bonomini F, Rodella LF, Rezzani R. Metabolic Syndrome, Aging and Involvement of Oxidative Stress. Aging Dis. 2015;6:109–120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Grundy SM, Brewer HB, Cleeman JI, Smith SC, Lenfant C. Definition of Metabolic Syndrome: Report of the National Heart, Lung, and Blood Institute/American Heart Association Conference on Scientific Issues Related to Definition. Arterioscler Thromb Vasc Biol. 2004;24:e13–e18. [DOI] [PubMed] [Google Scholar]
- 170.Halter JB, Musi N, McFarland Horne F, Crandall JP, Goldberg A, Harkless L, Hazzard WR, Huang ES, Kirkman MS, Plutzky J, Schmader KE, Zieman S, High KP. Diabetes and Cardiovascular Disease in Older Adults: Current Status and Future Directions. Diabetes. 2014;63:2578–2589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Fink RI, Kolterman OG, Griffin J, Olefsky JM. Mechanisms of insulin resistance in aging. J Clin Invest. 1983;71:1523–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Ryan A Insulin resistance with aging: effects of diet and exercise. Sport Med. 2000;30:327–346. [DOI] [PubMed] [Google Scholar]
- 173.DeFronzo R GLucose Intolerance and Aging. Diabetes Care. 1981;4:493–502. [DOI] [PubMed] [Google Scholar]
- 174.Biason-lauber A, Böni-schnetzler M, Hubbard BP, Bouzakri K, Brunner A, Cavelti-weder C, Keller C, Meyer-böni M, Daniel T, Brorsson C, Timper K, Leibowitz G, Patrignani A, Boily G, Zulewski H, Geier A, Cermak JM, Ellis JL, Westphal C, Knobel U, Eloranta JJ, Rogler G, Schlapbach R, Regairaz C, Carballido JM. Identification of a SIRT1 Mutation in a Family with Type 1 Diabetes. Cell Metab. 2013;17:448–455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Rutanen J, Yaluri N, Modi S, Pihlajamäki J, Vänttinen M, Itkonen P, Kainulainen S, Yamamoto H, Lagouge M, Sinclair DA, Elliott P, Westphal C, Auwerx J, Laakso M. SIRT1 mRNA expression may be associated with energy expenditure and insulin sensitivity. Diabetes. 2010;59:829–835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Hirschey MD, Shimazu T, Jing E, Grueter CA, Amy M, Aouizerat B, Stan A, Goetzman E, Lam MM, Schwer B, Stevens RD, Muehlbauer MJ, Kakar S, Nathan M, Kuusisto J, Laakso M, Alt FW, Christopher B. SIRT3 Deficiency and Mitochondrial Protein Hyperacetylation Accelerate the Development of the Metabolic Syndrome. Mol Cell. 2013;44:177–190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Banks AS. SirT1 gain of function increases energy efficiency and prevents diabetes in mice. Cell Metab. 2008;8:333–341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Bordone L, Cohen D, Robinson A, Motta MC, Van Veen E, Czopik A, Steele AD, Crowe H, Marmor S, Luo J, Gu W, Guarente L. SIRT1 transgenic mice show phenotypes resembling calorie restriction. Aging Cell. 2007;6:759–767. [DOI] [PubMed] [Google Scholar]
- 179.Moynihan KA, Grimm AA, Plueger MM, Bernal-Mizrachi E, Ford E, Cras-Méneur C, Permutt MA, Imai SI. Increased dosage of mammalian Sir2 in pancreatic β cells enhances glucose-stimulated insulin secretion in mice. Cell Metab. 2005;2:105–117. [DOI] [PubMed] [Google Scholar]
- 180.Bordone L, Motta MC, Picard F, Robinson A, Jhala US, Apfeld J, McDonagh T, Lemieux M, McBurney M, Szilvasi A, Easlon EJ, Lin SJ, Guarente L. Sirt1 regulates insulin secretion by repressing UCP2 in pancreatic β cells. PLoS Biol. 2006;4:210–220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Chen D, Bruno J, Easlon E, Lin S, Cheng H, Alt FW, Guarente L. Tissue-specific regulation of SIRT1 by calorie restriction service Tissue-specific regulation of SIRT1 by calorie restriction. Genes Dev. 2008;22:1753–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Rodgers JT, Puigserver P. Fasting-dependent glucose and lipid metabolic response through hepatic sirtuin 1. Proc Natl Acad Sci U S A. 2007;104:12861–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Hallows WC, Yu W, Denu JM. Regulation of glycolytic enzyme phosphoglycerate mutase-1 by Sirt1 protein-mediated deacetylation. J Biol Chem. 2012;287:3850–3858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Alrob OA, Sankaralingam S, Ma C, Wagg CS, Fillmore N, Jaswal JS, Sack MN, Lehner R, Gupta MP, Michelakis ED, Padwal RS, Johnstone DE, Sharma AM, Lopaschuk GD. Obesity-induced lysine acetylation increases cardiac fatty acid oxidation and impairs insulin signalling. Cardiovasc Res. 2014;103:485–497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Nasrin N, Wu X, Fortier E, Feng Y, Baré OC, Chen S, Ren X, Wu Z, Streeper RS, Bordone L. SIRT4 regulates fatty acid oxidation and mitochondrial gene expression in liver and muscle cells. J Biol Chem. 2010;285:31995–32002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Laurent G, de Boer VCJ, Finley LWS, Sweeney M, Lu H, Schug TT, Cen Y, Jeong SM, Li X, Sauve AA, Haigis MC. SIRT4 Represses Peroxisome Proliferator-Activated Receptor α Activity To Suppress Hepatic Fat Oxidation. Mol Cell Biol. 2013;33:4552–4561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Laurent G, German NJ, Saha AK, de Boer VCJ, Davies M, Koves TR, Dephoure N, Fischer F, Boanca G, Vaitheesvaran B, Lovitch SB, Sharpe AH, Kurland IJ, Steegborn C, Gygi SP, Muoio DM, Ruderman NB, Haigis MC. SIRT4 coordinates the balance between lipid synthesis and catabolism by repressing malonyl CoA decarboxylase. Mol Cell. 2013;50:686–698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Anderson KA, Huynh FK, Fisher-Wellman K, Stuart JD, Peterson BS, Douros JD, Wagner GR, Thompson JW, Madsen AS, Green MF, Sivley RM, Ilkayeva OR, Stevens RD, Backos DS, Capra JA, Olsen CA, Campbell JE, Muoio DM, Grimsrud PA, Hirschey MD. SIRT4 Is a Lysine Deacylase that Controls Leucine Metabolism and Insulin Secretion. Cell Metab. 2017;25:838–855.e15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Xiao C, Kim HS, Lahusen T, Wang RH, Xu X, Gavrilova O, Jou W, Gius D, Deng CX. SIRT6 deficiency results in severe hypoglycemia by enhancing both basal and insulin-stimulated glucose uptake in mice. J Biol Chem. 2010;285:36776–36784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Mostoslavsky R, Chua KF, Lombard DB, Pang WW, Fischer MR, Gellon L, Liu P, Mostoslavsky G, Franco S, Murphy MM, Mills KD, Patel P, Hsu JT, Hong AL, Ford E, Cheng HL, Kennedy C, Nunez N, Bronson R, Frendewey D, Auerbach W, Valenzuela D, Karow M, Hottiger MO, Hursting S, Barrett JC, Guarente L, Mulligan R, Demple B, Yancopoulos GD, Alt FW. Genomic instability and aging-like phenotype in the absence of mammalian SIRT6. Cell. 2006;124:315–329. [DOI] [PubMed] [Google Scholar]
- 191.Zhong L, Urso AD, Toiber D, Sebastian C, Henry RE, Vadysirisack DD, Guimaraes A, Marinelli B, Jakob D, Nir T, Clish CB, Vaitheesvaran B, Iliopoulos O, Kurland I, Dor Y, Weissleder R, Shirihai OS, Ellisen LW, Espinosa M, Mostoslavsky R. The Histone Deacetylase SIRT6 Regulates Glucose Homeostasis via Hif1α. Cell. 2010;140:280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Dominy JE, Jr YL, Jedrychowski MP, Chim H, Michael J, Camporez JP, Ruan H, Feldman J, Pierce K, Denu JM, Clish CB, Yang X, Shulman GI, Gygi SP, Puigserver P. The Deacetylase Sirt6 Activates the Acetyltransferase GCN5 and Suppresses Hepatic Gluconeogenesis. Mol Cell. 2012;48:900–913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Kim H-S, Xiao C, Wang R-H, Lahusen T, Xu X, Vassilopoulos A, Vazquez-Ortiz G, Jeong W-I, Park O, Ki SH, Gao B, Deng C-X. Hepatic specific disruption of SIRT6 in mice results in fatty liver formation due to enhanced glycolysis and triglyceride synthesis. Cell Metab. 2010;12:224–236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Schwer B, Schumacher B, Lombard DB, Xiao C, Kurtev M V, Gao J. Growth and Causes Obesity. PNAS. 2010;107:21790–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Yoshizawa T, Karim MF, Sato Y, Senokuchi T, Miyata K, Fukuda T, Go C, Tasaki M, Uchimura K, Kadomatsu T, Tian Z, Smolka C, Sawa T, Takeya M, Tomizawa K, Ando Y, Araki E, Akaike T, Braun T, Oike Y, Bober E, Yamagata K. SIRT7 controls hepatic lipid metabolism by regulating the ubiquitin-proteasome pathway. Cell Metab. 2014;19:712–721. [DOI] [PubMed] [Google Scholar]
- 196.Yoshino J, Mills KF, Yoon MJ, Imai S. Nicotinamide mononucleotide, a key NAD+ intermediate, treats the pathophysiology of diet- and age-induced diabetes in mice. Cell Metab. 2011;14:528–536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Liu HH, Li JJ. Aging and dyslipidemia: A review of potential mechanisms. Ageing Res Rev. 2015;19:43–52. [DOI] [PubMed] [Google Scholar]
- 198.Félix-Redondo FJ, Grau M, Fernández-Bergés D. Cholesterol and cardiovascular disease in the elderly. Facts and gaps. Aging Dis. 2013;4:154–69. [PMC free article] [PubMed] [Google Scholar]
- 199.Li X, Zhang S, Blander G, Tse JG, Krieger M, Guarente L. SIRT1 Deacetylates and Positively Regulates the Nuclear Receptor LXR. Mol Cell. 2007;28:91–106. [DOI] [PubMed] [Google Scholar]
- 200.Walker AK, Yang F, Jiang K, Ji J, Watts J, Purushotham A, Boss O, Hirsch M, Ribich S, Smith J, Israelian K, Westphal C, Rodgers J, Shioda T, Elson S, Mulligan P, Najafi-Shoushtari H, Black J, Thakur J, Kadyk L, Whetstine J, Mostoslavsky R, Puigserver P, Li X, Dyson N, Hart A, Naar A. Conserved role of SIRT1 orthologs in fasting-dependent inhibition of the lipid/cholesterol regulator SREBP. Genes Dev. 2010;24:1403–1417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Tao R, Xiong X, DePinho RA, Deng CX, Dong XC. FoxO3 transcription factor and Sirt6 deacetylase regulate low density lipoprotein (LDL)-cholesterol homeostasis via control of the proprotein convertase subtilisin/kexin type 9 (Pcsk9) gene expression. J Biol Chem. 2013;288:29252–29259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Karpe F, Frayn KN. The nicotinic acid receptor--a new mechanism for an old drug. Lancet. 2004;363:1892–1894. [DOI] [PubMed] [Google Scholar]
- 203.Canto C, Menzies K, Auwerx J. NAD+ metabolism and the control of energy homeostasis - a balancing act between mitochondria and the nucleus. Cell Metab. 2015;22:31–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Cantó C, Auwerx J. Targeting sirtuin 1 to improve metabolism: all you need is NAD(+)? Pharmacol Rev. 2012;64:166–87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Berman AY, Motechin RA, Wiesenfeld MY, Holz MK. The therapeutic potential of resveratrol: a review of clinical trials. npj Precis Oncol. 2017;1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Öztürk E, Arslan AKK, Yerer MB, Bishayee A. Resveratrol and diabetes: A critical review of clinical studies. Biomed Pharmacother. 2017;95:230–234. [DOI] [PubMed] [Google Scholar]
- 207.Timmers S, Konings E, Bilet L, Houtkooper RH, de Weijer T Van, Goosens GH, Hoeks J, van der Krieken S, Ryu D, Kersten S, Moonen-Kornips E, Hesselink MK, Kunz I, Schrauwen-Hinderling VB, Blaak E, Auwerx J, Schrauwen P. Calorie restriction-like effects of 30 days of resveratrol supplementation on energy metabolism and metabolic profile in obese humans. Cell Metab. 2011;14:612–622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Poulsen MM. High-dose resveratrol supplementation in obese men: an investigator-initiated, randomized, placebo-controlled clinical trial of substrate metabolism, insulin sensitivity, and body composition. Diabetes. 2013;62:1186–1195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Méndez-del Villar M, González-Ortiz M, Martínez-Abundis E, Pérez-Rubio KG, Lizárraga-Valdez R. Effect of Resveratrol Administration on Metabolic Syndrome, Insulin Sensitivity, and Insulin Secretion. Metab Syndr Relat Disord. 2014;12:497–501. [DOI] [PubMed] [Google Scholar]
- 210.Movahed A, Nabipour I, Louis XL, Thandapilly SJ, Yu L, Kalantarhormozi M, Rekabpour SJ, Netticadan T. Antihyperglycemic effects of short term resveratrol supplementation in type 2 diabetic patients. Evidence-Based Complement Altern Med. 2013;2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Magyar K Cardioprotection by resveratrol: a human clinical trial in patients with stable coronary artery disease. Clin Hemorheol Microcirc. 2012;50:179–187. [DOI] [PubMed] [Google Scholar]
- 212.Faghihzadeh F, Adibi P, Rafiei R, Hekmatdoost A. Resveratrol supplementation improves inflammatory biomarkers in patients with nonalcoholic fatty liver disease. Nutr Res. 2014;34:837–843. [DOI] [PubMed] [Google Scholar]
- 213.McDermott MM, Leeuwenburgh C, Guralnik JM, Tian L, Sufit R, Zhao L, Criqui MH, Kibbe MR, Stein JH, Lloyd-Jones D, Anton SD, Polonsky TS, Gao Y, de Cabo R, Ferrucci L. Effect of Resveratrol on Walking Performance in Older People With Peripheral Artery Disease. JAMA Cardiol. 2017;2:902–907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Pollack RM, Barzilai N, Anghel V, Kulkarni AS, Golden A, O’Broin P, Sinclair DA, Bonkowski MS, Coleville AJ, Powell D, Kim S, Moaddel R, Stein D, Zhang K, Hawkins M, Crandall JP. Resveratrol Improves Vascular Function and Mitochondrial Number but Not Glucose Metabolism in Older Adults. J Gerontol A Biol Sci Med Sci. 2017;72:1703–1709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Libri V A pilot randomized, placebo controlled, double blind phase I trial of the novel SIRT1 activator SRT2104 in elderly volunteers. PLoS One. 2012;7:e51395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Venkatasubramanian S, Noh RM, Daga S, Langrish JP, Joshi N V., Mills NL, Hoffmann E, Jacobson EW, Vlasuk GP, Waterhouse BR, Lang NN, Newby DE. Cardiovascular effects of a novel SIRT1 activator, SRT2104, in otherwise healthy cigarette smokers. J Am Hear Assoc. 2013;2:e000042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Noh RM, Venkatasubramanian S, Daga S, Langrish J, Mills NL, Lang NN, Hoffmann E, Waterhouse B, Newby DE, Frier BM. Cardiometabolic effects of a novel SIRT1 activator, SRT2104, in people with type 2 diabetes mellitus. Open Hear. 2017;4:e000647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Trammell SAJ, Schmidt MS, Weidemann BJ, Redpath P, Jaksch F, Dellinger RW, Li Z, Abel ED, Migaud ME, Brenner C. Nicotinamide riboside is uniquely and orally bioavailable in mice and humans. Nat Commun. 2016;7:12948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Airhart SE, Shireman LM, Risler LJ, Anderson GD, Gowda GAN, Raftery D, Tian R, Shen DD, O’Brien KD. An open-label, non-randomized study of the pharmacokinetics of the nutritional supplement nicotinamide riboside (NR) and its effects on blood NAD+ levels in healthy volunteers. PLoS One. 2017;12:1–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Martens CR, Denman BA, Mazzo MR, Armstrong ML, Reisdorph N, Mcqueen MB, Chonchol M, Seals DR. Chronic nicotinamide riboside supplementation is well-tolerated and elevates NAD+ in healthy middle-aged and older adults. Nat Commun. 2018;9:1286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Dellinger RW, Santos SR, Morris M, Evans M, Alminana D, Guarente L, Marcotulli E. Repeat dose NRPT (nicotinamide riboside and pterostilbene) increases NAD+ levels in humans safely and sustainably: a randomized, double-blind, placebo-controlled study. npj Aging Mech Dis. 2017;3:17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Tsubota K The first human clinical study for NMN has started in Japan. npj Aging Mech Dis. 2016;2:16021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Morrow DA, Brickman CM, Murphy SA, Baran K, Krakover R, Dauerman H, Kumar S, Slomowitz N, Grip L, McCabe CH, Salzman AL. A randomized, placebo-controlled trial to evaluate the tolerability, safety, pharmacokinetics, and pharmacodynamics of a potent inhibitor of poly(ADP-ribose) polymerase (INO-1001) in patients with ST-elevation myocardial infarction undergoing primary percutaneous coronary intervention: Results of the TIMI 37 trial. J Thromb Thrombolysis. 2009;27:359–364. [DOI] [PubMed] [Google Scholar]
- 224.Chen S, Zhao X, Ran L, Wan J, Wang X, Qin Y, Shu F, Gao Y, Yuan L, Zhang Q, Mi M. Resveratrol improves insulin resistance, glucose and lipid metabolism in patients with non-alcoholic fatty liver disease: A randomized controlled trial. Dig Liver Dis. 2015;47:226–232. [DOI] [PubMed] [Google Scholar]
- 225.Keller K, Howlett SE. Sex Differences in the Biology and Pathology of the Ageing Heart. Can J Cardiol. 2016; [DOI] [PubMed] [Google Scholar]
- 226.Merz AA, Cheng S. Sex differences in cardiovascular ageing. Heart. 2016;102:825–831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Austad SN, Bartke A. Sex Differences in Longevity and in Responses to Anti-Aging Interventions: A Mini-Review. Gerontology. 2016;62:40–46. [DOI] [PubMed] [Google Scholar]
- 228.Parikh P, McDaniel MC, Ashen MD, Miller JI, Sorrentino M, Chan V, Blumenthal RS, Sperling LS. Diets and cardiovascular disease: An evidence-based assessment. J Am Coll Cardiol. 2005;45:1379–1387. [DOI] [PubMed] [Google Scholar]
- 229.Dietary Mozaffarian D. and Policy Priorities for Cardiovascular Disease, Diabetes, and Obesity. Circulation. 2016;133:187–225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Elamin M, Ruskin DN, Masino SA, Sacchetti P. Ketone-Based Metabolic Therapy: Is Increased NAD+ a Primary Mechanism? Front Mol Neurosci. 2017;10:1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]