

Abstract
The “Landscape Of Gut Microbiome - Pan-India Exploration”, or LogMPIE study, is the first large-scale, nationwide record of the Indian gut microbiome. The primary objective of the study was to identify and map the Indian gut microbiome baseline. This observational study was conducted across 14 geographical locations in India. Enrolled subjects were uniformly distributed across geographies (north, east, west and south) and body mass index (obese and non-obese). Furthermore, factors influencing the microbiome, such as age and physical activity, were also considered in the study design. The LogMPIE study recorded data from 1004 eligible subjects and reported 993 unique microorganisms across the Indian microbiome diaspora. The data not only map the Indian gut microbiome baseline but also function as a useful resource to study, analyse and identify signatures characterizing the physiological dispositions of the subjects. Furthermore, they provide insight into the unique features describing the Indian microbiome. The data are open and may be accessed from the European Nucleotide Archive (ENA) portal of the European Bioinformatics Institute (https://www.ebi.ac.uk/ena/data/view/PRJEB25642).
Subject terms: Metagenomics, Microbiome, DNA sequencing, Gastrointestinal system
Background & Summary
The gut microbiome and its host share a primarily symbiotic, commensal relationship that is occasionally pathogenic1. An increasing body of evidence now substantiates that the gut microbiome plays a critical role in digestion, nutrition, and immune system maturation2–7. A non-exhaustive list of physiological disorders associated with gut microbiome dysbiosis includes Crohn’s disease8,9, type II diabetes10,11, colorectal cancer12,13 and metabolic disorders14,15. With advancements in the gut microbiome field, modulation of host physiology and biochemistry by the microbiome is being investigated in greater detail16.
To better understand host physiology modulation by the gut microbiome, it is imperative to know the microbiome composition. Acknowledging this requirement, multiple consortia were set up to map the gut microbiome across different geographies. Pioneering efforts were initiated through the ‘The Human Microbiome Project’ (HMP)17,18 and Metagenome of Human Intestinal Tract (MetaHIT)19,20 studies. Following suit, multiple nationwide and cohort-specific studies were conducted to understand the impact of the gut microbiome. 21,22. These studies led to an exponential rise in available gut microbiome datasets across multiple cohorts.
Among the multiple factors contributing to the compositional diversity of the gut microbiome, two key influencers are geography and diet23–28. Comparative assessments across multiple populations, such as those between Europeans and Americans19, Koreans and other Asians29 and within Africans, confirmed that geography influences gut microbiome diversity30,31. Genome-scale metabolic simulations indicated that diet composition influences the growth rates of microorganisms within the gut32. The differential growth rate of these microorganisms leads to diversity within the gut microbiome33,34. Age as a factor was also reported to influence the gut microbiome composition35,36. An increase in gut microbiome diversity was most pronounced in infants37 and continued until adulthood36,38. Interestingly, the elderly population showed a loss in gut microbiome diversity39. Reduction in the gut microbiome diversity in the elderly population could be a result of dietary restrictions, constrained lifestyle, and medications39.
Over the past few years, multiple studies investigated sub-sections of the Indian gut microbiome40–43. Each of these gut microbiome studies did consider a cohort from different geographies and physiological dispositions. Unfortunately, owing to the lack of protocol consistency and processing pipeline variations, a meta-analysis based on the gut microbiome composite data was limited44,45. A comprehensive gut microbiome study describing the impact of geography, age, sex, BMI and physical activity across the Indian population has yet to be reported. A study along these lines would provide insight into the association of various factors, such as cultural affiliations, geography and changing lifestyle, with the gut microbiome composition.
The ‘Landscape Of Gut Microbiome - Pan-India Exploration’, or the LogMPIE, is to the best of our knowledge the first large-scale, observational, multi-centric, cross-geographic and diverse age group study focusing on the Indian population. This study reports data from 1004 participating subjects. The participants represented a uniform distribution of obese and non-obese subjects. Additionally, the study recorded sex and lifestyle patterns based on physical activity (sedentary and non-sedentary) of the participating subjects.
Adherence to a common standardized operating protocol and centralized sequencing facility reduced possibilities of sample processing and pipeline-related variations. Furthermore, cross-checking and validation at multiple points during sample and data processing assured strict quality control.
Based on a preliminary assessment, operational taxonomic unit (OTU) tables for the individual subjects were shared along with the FASTQ data. The sequence processing pipeline used thresholds for reporting higher specificity against a coverage trade-off (please refer to the Methods section for details). FASTQ data from the study are available from the European Nucleotide Archive portal of the European Bioinformatics Institute (Data Citation 1). Sharing of the FASTQ data enables users to re-compute OTU distributions using a preferred pipeline and customized parameters.
The LogMPIE study reported features specific to the Indian population. In comparison to the Western gut microbiome composition, the Indian gut microbiome reported a higher relative abundance of Prevotella copri (~0.39). Increased abundance of Prevotella copri is attributed to the high content of resistant starch within the standard Indian diet46. Faecalibacterium prausnitzii was the other copious microorganism reported with a relative abundance of ~0.13 (Table 1).
Table 1. Relative abundance and frequency of observation in the study sample of the top 10 microorganisms within the study cohort.
| Organisms (order; family) | Relative Abundance | Frequency of Observation in the Study Sample |
|---|---|---|
| Prevotella copri (o = Bacteroidales; f = Prevotellaceae) | 0.391 | 0.966 |
| Faecalibacterium prausnitzii (o = Clostridiales; f = Ruminococcaceae) | 0.131 | 0.966 |
| Bacteroides plebeius (o = Bacteroidales; f = Bacteroidaceae) | 0.041 | 0.964 |
| Haemophilus parainfluenzae (o = Pasteurellales; f = Pasteurellaceae) | 0.033 | 0.861 |
| Roseburia faecis (o = Clostridiales; f = Lachnospiraceae) | 0.025 | 0.962 |
| Megasphaera elsdenii (o = Veillonellales; f = Veillonellaceae) | 0.024 | 0.933 |
| Lactobacillus rogosae (o = Lactobacillales; f = Lactobacillaceae) | 0.022 | 0.964 |
| Prevotella stercoreacopri (o = Bacteroidales; f = Prevotellaceae) | 0.021 | 0.964 |
| Parasutterella excrementihominis (o = Burkholderiales; f = Sutterellaceae) | 0.020 | 0.924 |
| Ruminococcus gnavus (o = Clostridiales; f = Lachnospiraceae) | 0.017 | 0.958 |
The comparative assessment indicated that 390 out of 993 microorganisms were present in all the geographical zones (Fig. 1). A detailed assessment of the gut microbiome composition and its variation owing to influencing factors is beyond the scope of the current article. However, the LogMPIE data allow an investigation of the gut microbiome composition in response to multiple influencing factors.
Figure 1. Distinct species reported across geographical locations.
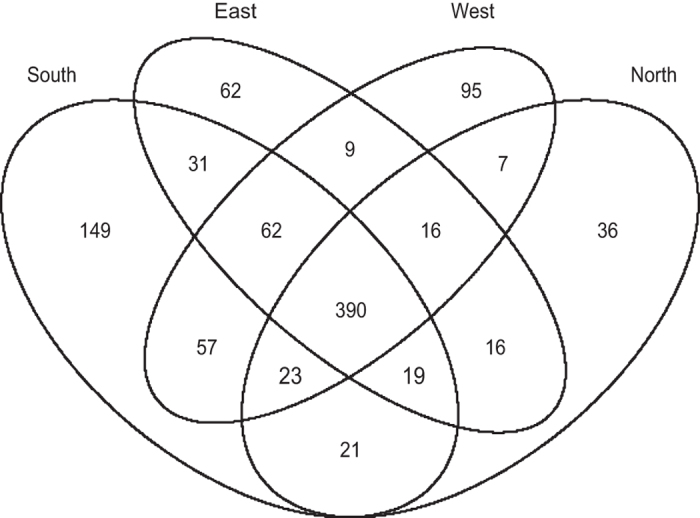
North (number of subjects 243), South (number of subjects 250), East (number of subjects 250) and West (number of subjects 261).
Methods
The LogMPIE study was conducted across 14 geographical locations in India (Table 2). During the study, all pertinent requirements recommended by the Indian Council of Medical Research (ICMR) for Biomedical Research on Human Subjects and by the International Conference on Harmonization-Good Clinical Practice (ICH-GCP) were consulted and adhered to. The study was registered with the Clinical Trial Registry-India (CTRI Number: CTRI/2016/03/007616). Following acceptance of the study protocol by independent ethics committees/institutional review boards (IEC/IRB), the LogMPIE study was initiated. Prior to initiation of the study, willing volunteers from the 14 geographical locations were educated on the study objectives and the sampling procedure, and consent documents were obtained.
Table 2. Sample counts from the multiple study centres.
| Site | Geographical Location | Number of Samples |
|---|---|---|
| Bhopal | North | 65 |
| Ludhiana | North | 65 |
| Lucknow | North | 67 |
| New Delhi | North | 46 |
| Guwahati | East | 83 |
| Kolkata | East | 84 |
| Patna | East | 83 |
| Ahmedabad | West | 65 |
| Ajmer | West | 70 |
| Mumbai | West | 59 |
| Nagpur | West | 67 |
| Chennai | South | 89 |
| Cochin | South | 96 |
| Mangalore | South | 65 |
Study Design
Since LogMPIE was an observational, multi-centric and non-interventional study, the sample size of the study was not statistically derived. Enrolment of subjects for the study was based on the inclusion and exclusion criteria, as listed in Table 3. A schema of the study workflow is included in Fig. 2.
Table 3. Inclusion and exclusion criteria of the study.
| Inclusion Criteria | Exclusion Criteria |
|---|---|
| • Age: 18-65• Certified healthy on physical examination and free from diabetes, acquired immunodeficiency syndrome (AIDS), chronic diarrhoea, inflammatory bowel disease, irritable bowel syndrome, or other gastrointestinal disorders, gastrointestinal surgery [with exception of appendectomy, polypectomy, or herniorrhaphy]. | • Prescription, OTC medications or supplements (e.g., acid anti-secretory drugs, probiotics) known to alter the gut function or microbiome during the 4 weeks prior to study enrolment |
Figure 2. Schematic workflow elucidating sample collection, sample processing and data processing adopted during the LogMPIE study.

A total of 1022 subjects were enrolled and screened. The data from 1004 eligible subjects were obtained, processed and reported. Subject distributions were guided based on geography and body mass index (with obese and non-obese being the two groups) uniformity. Details of subject distribution under individual categories are reported in Table 4. Subject classification under the physical activity category (sedentary and non-sedentary) was based on features adapted from the MOPO study47. Furthermore, the grouping of subjects under the BMI category was based on the recommendations from the National Heart, Lung and Blood Institute48.
Table 4. Distribution of subjects across different study categories.
| Categories | Distribution |
|---|---|
| Age | Range (Years): 18–65 |
| Sex | Male = 591; Female = 431 |
| BMI | Underweight = 31 Normal = 263 Overweight = 277 Obese = 451 |
| Geographical Location | North = 247 South = 262 East = 250 West = 263 |
| Physical Activity | Sedentary = 470 Non-sedentary = 552 |
Sample Collection
Faecal samples were collected in a pre-labelled sterile OMNIgene®•GUT stool collection kit (OMR-200, DNA Genotek, Ottawa, Canada) by individual subjects49. The stool collection kit is an all-in-one system (for details please refer to Supplementary Information, S1.1). It is designed to stabilize and maintain DNA integrity, thus enabling gut microbiome profiling at an ambient temperature.
Prior to stool collection, adequate training and instructions regarding the collection process were provided to individual subjects. Samples collected in the OMNIgene®•GUT stool collection kit were stored at ~20 °C. This temperature adhered to the kit manufacturer’s instructions. The temperature was maintained at ~20°C until sample processing at the central sequencing facility. The samples were processed for genomic DNA isolation within 2 days of collection.
DNA Isolation
The bacterial genomic DNA was isolated and purified from the collected faecal samples using a QiaAmp DNA Stool Mini Kit (Qiagen, Hilden, Germany). ASL buffer was used to lyse the stool samples. Normally, bacterial cells lyse at 70°C in the ASL buffer. For the current protocol, a higher lysis temperature (95°C) was used to account for cells that were difficult to lyse (such as Gram-positive bacteria). Post lysis, DNA-damaging agents and PCR inhibitors were removed using InhibitEX Matrix Tablets (Qiagen, Hilden, Germany). A standardized protocol was used to isolate DNA using QIAamp Mini Spin Columns (Supplementary Information, S1.2). The genomic DNA was eluted in 100 μl of elution buffer. The quality of the isolated genomic DNA was confirmed by agarose gel electrophoresis. DNA concentration was estimated with a Qubit 2.0 instrument and with a Qubit dsDNA HS Assay Kit (Thermo Fisher Scientific, Carlsbad, CA, USA). A detailed standardized protocol adopted for DNA quantification is included within the supplementary section (Supplementary Information, S1.3).
16S Primers and Amplicon Library Generation
The hypervariable regions of the 16S rRNA gene were PCR amplified using extracted DNA as the template. For details regarding the protocol, please refer to the supplementary section (Supplementary Information, S1.4). Primer pair Probio_Uni and Probio_Rev were used to amplify the V3 region50. To target the V4 region of the 16S rRNA gene, a primer pair of 520F and 802R was used51. Primer details are listed in Table 5.
Table 5. Details of the primers used to amplify the 16S rRNA gene.
| Primer Name | Adapter Sequence | Key | Barcode | Barcode Adapter | Primer Sequence (5′-3′) |
|---|---|---|---|---|---|
| Probio_Uni50 | CCATCTCATCCCTGCGTGTCTCCGAC | TCAG | TTACAACCTC | GAT | CCTACGGGRSGCAGCAG |
| Probio_Rev50 | CCTCTCTATGGGCAGTCGGTGAT | ATTACCGCGGCTGCT | |||
| 520F51 | CCATCTCATCCCTGCGTGTCTCCGAC | TCAG | TTACAACCTC | GAT | AYTGGGYDTAAAGNG |
| 802R51 | CCTCTCTATGGGCAGTCGGTGAT | TACNVGGGTATCTAATCC |
Amplitaq Gold 360 MM (Thermo Fisher Scientific, Foster City, CA, USA) was used for 16S rRNA gene amplification, and the PCR conditions were set as follows: initial denaturation at 94°C for 5 min, denaturation at 94°C for 30 sec, annealing at 55°C for 30 sec and extension at 72°C for 90 sec. The final extension was performed at 72°C for 10 min. PCR amplification was performed using a 9700 Thermocycler (Thermo Fisher Scientific, Grand Island, NY, USA). The PCR amplicon was purified using AMPure XP reagent (Beckman Coulter, Brea, CA, USA). The concentration of the amplicon was determined with a Qubit dsDNA HS Assay Kit. The respective size distribution of the amplicon was verified with an Agilent 2100 Bioanalyzer using a high-sensitivity DNA kit (Agilent, Santa Clara, CA, USA). The amplicon library was diluted to 100 pM, and an equimolar pool was prepared for clonal amplification. Protocols used for amplicon library formation are included in the supplementary section (Supplementary Information, S1.3).
Template Preparation and Sequencing
Template preparation for libraries using ion spheres was performed using OneTouch 2 protocols and corresponding reagents (Thermo Fisher Scientific, Carlsbad, CA, USA). The Ion OneTouch 2 system (Thermo Fisher Scientific, Carlsbad, CA, USA) enables automated delivery of Ion Sphere Particle (ISP) templates. Further details regarding the Ion One Touch 2 system are included within the supplementary section (Supplementary Information, S1.5). The ISP templates were loaded either on an Ion 520 or Ion 530 chip kit, and a standardized protocol was followed. Sequencing was performed with the Ion 520 or Ion 530 kit-OT2 (Thermo Fisher Scientific, Carlsbad, CA, USA) using the 200 bp chemistry with 500 flow (125 cycles) for the V3 region and 400 bp chemistry with 800 flow (200 cycles) for the V4 region run format. The sequencing was performed on an Ion S5 System. Using the default pre-processing parameters, reads pertaining to adaptor sequences were filtered out, and the sequence data were stored in the FASTQ format. Individual sequence data for the subjects are available at the European Nucleotide Archive (Data Citation 1).
Bioinformatics Analysis
The raw sequencing data were processed through a customized 16S analysis pipeline to report the taxonomical distribution of species along with their relative abundance in an OTU table (Relative Abundance Table, Data Citation 2). The customized processing was performed using a QIIME workflow on the Ion Reporter Server51. It should be noted that the content of the OTU table depends on the pipeline and the parameters used for processing the data. For customized assessments, users are encouraged to regenerate the OTU table from the shared FASTQ data, employing their preferred pipeline and parameters.
Pre-processing Sequence Data
The sequence data were pre-processed, and the step primers were trimmed. The minimum read length threshold was set to 100 bp. Reads recording lengths shorter than the threshold were dropped from further processing. Read sequences were clustered together and checked for copy number with a minimum threshold of 10 reads. Low copy numbers (threshold <10) were filtered out and dropped from further analysis.
Organism Screening and Assessment
The reads were aligned against two comprehensive 16S databases, the Thermo Fisher Scientific in-house MicroSEQ® 16S rRNA reference database (V2013.1) and the curated Greengenes database (V13.5)52. Reads were aligned against the databases using Megablast (from the BLAST package). The expectation value (E-value) for the searches was set to 0.01, and the max target hits value was set to 10053. To assign taxonomy, the minimum alignment percentage of a read to a subject sequence (homologue) in the database was set to a threshold of 90. A read was assigned to a genus only when the identity score of the sequence alignment (between the read and subject sequence from the database) was at 97% or higher. For species assignment, the minimum percentage identity of the alignment was set to 99%.
Reads assigned to multiple entities (genus or species) at a taxonomy level were further assessed for refinement. In a scenario where the top homologue of a read (the most similar sequence based on the Megablast search) reports a sequence identity of greater than 0.2% in comparison to the next homologue, the read was assigned the taxonomic label of the top homologue. On failure, the read was assigned the taxonomic label of the homologues within 0.2% of the top homologue. For reads with a conflicting assignment, a “slash ID” was issued. The slash ID recorded the multiple taxonomy assessments.
The taxonomy distribution counts or abundance was derived from the clustered reads. The abundance value from the pipeline was further transformed into the relative abundance of the individual species. The shared OTU table (Relative Abundance Table, Data Citation 2) was derived using a QIIME workflow on the Ion Reporter Server and in-house optimized parameters.
Data Analysis Pipeline
Customized assessment of the LogMPIE data was performed using Ion ReporterTM software (please refer to the previous sub-sections for the parameters used in the analysis). To use Ion ReporterTM software, individual users are required to register with the portal (https://ionreporter.thermofisher.com/ir/). For standalone processing of the data, ‘Microbiome Processing Pipeline’, a Python-based tool, is being shared at GitHub (https://github.com/anirbanbhaduri/LogMPIE). Based on user-defined parameters, the pipeline processes input FASTQ data and reports out OTU tables. The tool enables a user to download the microbiome processing tools (QIIME54 and Mothur55). For taxonomic referencing, the tool may use Greengenes56, Silva57, and RDP58 databases. Owing to licensing implications, processing tools and databases need to be obtained separately by the user.
Data Records
The LogMPIE study repository shares 3 data types (explained below). The data are organized to enable multiple forms of assessments. Data type 1 is available at the European Nucleotide Archive (ENA) portal of European Bioinformatics Institute, while the other 2 data types are shared through the Supplementary Information.
Data type 1
FASTQ data obtained from sequencing the V3 and V4 regions of the 16S rRNA gene of microorganisms hosted within individual subjects are shared as a part of the LogMPIE study repository. The data comprises 1004 FASTQ sequence files (Data Citation 1). They are found under the primary accession code, PRJEB25642, and secondary accession code, ERP07577, on the ENA portal. These FASTQ files were processed through a QIIME workflow on the Ion Reporter Server51. It should be noted that FASTQ files enable users to customize their assessments based on selected parameters and pipelines.
Data type 2
The OTU table reporting the relative abundance of microorganism across individuals is available as Supplementary Information (Relative Abundance Table, Data Citation 2). The table reports the relative abundance of all microorganisms at the species level. The strain information is currently not included but may be obtained through a customized FASTQ data processing pipeline by interested users.
Data type 3
Data type 3 reports the study metadata (LogMPIE Study Metadata, Data Citation 2). This comprises the codes of participating subjects along with information regarding their age, sex, physical activity, BMI and geographical locations. Attributes within the metadata would facilitate retrospective studies.
Technical Validation
Several layers of quality assurance and quality control (QC) systems were implemented and maintained. To ensure that the study was conducted and the data were generated, documented and reported in compliance with the ICH-GCP and ICMR guidelines for Biomedical Research on Humans, standard operating protocols were developed. Furthermore, each individual working on the study was trained on the protocols.
Sample Management
Iterative data checks were performed to ensure accuracy in data entry. Manual inspection of the data integrity within the database was performed by the QC team before the database was locked.
DNA Sample Quality Control
All the isolated DNA samples were quantified, checked for quality and further used for the amplicon library preparation. 16S rRNA gene amplicon generation success was assessed by reviewing the amplicon size. The primer pair Probio_Uni and Probio_Rev (V3 Region) led to a PCR product of 194 bp, and the primer pair 520 F and 802 R (V4 region) produced a PCR product of 263 bp according to the Escherichia coli K-12 16S rRNA gene sequence. The absence of contaminants and the respective size distribution of the amplicon were verified using an Agilent 2100 Bioanalyzer and the Qubit dsDNA HS Assay Kit.
Sequence Quality Assessment and Bioinformatics Pipeline
Reads were further assessed based on the quality score. Histograms of the Phred scores for all the reads of the 1004 FASTQ samples are shown in Fig. 3. Geographical location wise plots are shared as Supplementary Information (Supplementary Information, Figure S1). The FASTQ data may be refined based on the Phred score. In the current taxonomic assignment, a Phred score threshold of 20 was considered to reduce both noise and false positives in the data. Using a minimum read length of 100 bp and having a threshold of 10 reads per cluster, reliable read data were obtained. These data were processed and used to generate the OTU table. Parameters used to process the data are sensitive and would influence the produced OTU table.
Figure 3. Plot of the Phred score against the ‘Average read counts with specific Phred score per subject’, across the 1004 subjects.
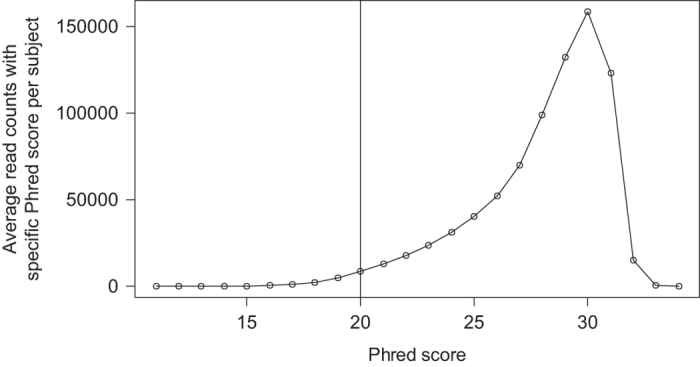
Phred score threshold for the taxonomic assignment was set to 20.
Usage Notes
The primary objective of the LogMPIE study was to report the Indian gut microbiome composition baseline. Additionally, the study recorded geographical location, sex, age, physical activity and body mass index for each participating subject. The repository, to the best of our knowledge, is the most comprehensive gut microbiome dataset representing the Indian population. The authors acknowledge that this study, though comprehensive, may not be exhaustive.
The LogMPIE study acts as a powerful microbiome dataset to enable multiple applications. In addition, the dataset helps to identify and quantify features or descriptors associated with physiological dispositions of the host.
Additional information
How to cite this article: Dubey, A. K. et al. LogMPIE, pan-India profiling of the human gut microbiome using 16S rRNA sequencing. Sci. Data. 5:180232 doi: 10.1038/sdata.2018.232 (2018).
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Material
Acknowledgments
The authors would like to acknowledge Faraz Ul Hasan, Madhavi Kaushal, Jyoti Sharma, Bharat Bhushan and Nitish Jha for their assistance in facilitating sample collection. Furthermore, we acknowledge the contribution of Atima Agarwal for her inputs in DNA sequencing.
Footnotes
The authors of this study are employees of commercial companies. However, this does not alter their adherence to the Scientific Data journal policies on sharing data and materials. The specific role of the individual authors is articulated in the ‘Author contributions’ section.
Data Citations
- 2018. European Nucleotide Archive. PRJEB25642
- Dubey A. K., et al. . 2018. figshare. https://doi.org/10.6084/m9.figshare.c.4147079
References
- Liang D., Leung R. K.-K., Guan W. & Au W. W. Involvement of gut microbiome in human health and disease: brief overview, knowledge gaps and research opportunities. Gut Pathog 10, 3 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Statovci D., Aguilera M., MacSharry J. & Melgar S. The Impact of Western Diet and Nutrients on the Microbiota and Immune Response at Mucosal Interfaces. Front. Immunol 8, 838 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi N., Li N., Duan X. & Niu H. Interaction between the gut microbiome and mucosal immune system. Mil. Med. Res 4, 14 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson E. L., Heaver S. L., Walters W. A. & Ley R. E. Microbiome and metabolic disease: revisiting the bacterial phylum Bacteroidetes. J. Mol. Med. (Berl) 95, 1–8 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michail S. et al. Altered gut microbial energy and metabolism in children with non-alcoholic fatty liver disease. FEMS Microbiol. Ecol 91, 1–9 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Relman D. A. The human microbiome: ecosystem resilience and health. Nutr. Rev. 70(Suppl 1): S2–9 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hehemann J.-H. et al. Transfer of carbohydrate-active enzymes from marine bacteria to Japanese gut microbiota. Nature 464, 908–12 (2010). [DOI] [PubMed] [Google Scholar]
- Prosberg M., Bendtsen F., Vind I., Petersen A. M. & Gluud L. L. The association between the gut microbiota and the inflammatory bowel disease activity: a systematic review and meta-analysis. Scand. J. Gastroenterol. 51, 1407–1415 (2016). [DOI] [PubMed] [Google Scholar]
- Hedin C. R., van der Gast C. J., Stagg A. J., Lindsay J. O. & Whelan K. The gut microbiota of siblings offers insights into microbial pathogenesis of inflammatory bowel disease. Gut Microbes 8, 359–365 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang D. D. & Hu F. B. Precision nutrition for prevention and management of type 2 diabetes. lancet. Diabetes Endocrinol 6, 416–426 (2018). [DOI] [PubMed] [Google Scholar]
- Brunkwall L. & Orho-Melander M. The gut microbiome as a target for prevention and treatment of hyperglycaemia in type 2 diabetes: from current human evidence to future possibilities. Diabetologia 60, 943–951 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu J. et al. Metagenomic analysis of faecal microbiome as a tool towards targeted non-invasive biomarkers for colorectal cancer. Gut 66, 70–78 (2017). [DOI] [PubMed] [Google Scholar]
- Liang Q. et al. Fecal Bacteria Act as Novel Biomarkers for Noninvasive Diagnosis of Colorectal Cancer. Clin. Cancer Res. 23, 2061–2070 (2017). [DOI] [PubMed] [Google Scholar]
- Meijnikman A. S., Gerdes V. E., Nieuwdorp M. & Herrema H. Evaluating Causality of Gut Microbiota in Obesity and Diabetes in Humans. Endocr. Rev 39, 133–153 (2018). [DOI] [PubMed] [Google Scholar]
- Kasselman L. J., Vernice N. A., DeLeon J. & Reiss A. B. The gut microbiome and elevated cardiovascular risk in obesity and autoimmunity. Atherosclerosis 271, 203–213 (2018). [DOI] [PubMed] [Google Scholar]
- Schmidt T. S. B., Raes J. & Bork P. The Human Gut Microbiome: From Association to Modulation. Cell 172, 1198–1215 (2018). [DOI] [PubMed] [Google Scholar]
- Human Microbiome Jumpstart Reference Strains Consortium et al. A catalog of reference genomes from the human microbiome. Science 328, 994–9 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turnbaugh P. J. et al. The human microbiome project. Nature 449, 804–10 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin J. et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 464, 59–65 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J. et al. An integrated catalog of reference genes in the human gut microbiome. Nat. Biotechnol 32, 834–41 (2014). [DOI] [PubMed] [Google Scholar]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 486, 207–14 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Del Savio L., Prainsack B. & Buyx A. Motivations of participants in the citizen science of microbiomics: data from the British Gut Project. Genet. Med. 19, 959–961 (2017). [DOI] [PubMed] [Google Scholar]
- Gupta V. K., Paul S. & Dutta C. Geography, Ethnicity or Subsistence-Specific Variations in Human Microbiome Composition and Diversity. Front. Microbiol 8, 1162 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yadav D., Ghosh T. S. & Mande S. S. Global investigation of composition and interaction networks in gut microbiomes of individuals belonging to diverse geographies and age-groups Gut Pathog 8, 17 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hullar M. A. J. & Fu B. C. Diet, the gut microbiome, and epigenetics. Cancer J. 20, 170–175 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arumugam M. et al. Enterotypes of the human gut microbiome. Nature 473, 174–180 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu G. D. et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 334, 105–108 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- David L. A. et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 505, 559–63 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nam Y.-D., Jung M.-J., Roh S. W., Kim M.-S. & Bae J.-W. Comparative analysis of Korean human gut microbiota by barcoded pyrosequencing. PLoS One 6, e22109 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schnorr S. L. et al. Gut microbiome of the Hadza hunter-gatherers. Nat. Commun. 5 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Filippo C. et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 107, 14691–6 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shoaie S. et al. Understanding the interactions between bacteria in the human gut through metabolic modeling. Sci. Rep 3, 2532 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Queipo-Ortuño M. I. et al. Influence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers. Am. J. Clin. Nutr 95, 1323–1334 (2012). [DOI] [PubMed] [Google Scholar]
- Mutlu E. A. et al. Colonic microbiome is altered in alcoholism. Am. J. Physiol. Liver Physiol 302, G966–G978 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lynch S. V & Pedersen O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 375, 2369–2379 (2016). [DOI] [PubMed] [Google Scholar]
- Kundu P., Blacher E., Elinav E. & Pettersson S. Our Gut Microbiome: The Evolving Inner Self. Cell 171, 1481–1493 (2017). [DOI] [PubMed] [Google Scholar]
- Korpela K. et al. Selective maternal seeding and environment shape the human gut microbiome. Genome Res. 28, 561–568 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Odamaki T. et al. Age-related changes in gut microbiota composition from newborn to centenarian: a cross-sectional study. BMC Microbiol. 16, 90 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Toole P. W. & Jeffery I. B. Gut microbiota and aging. Science 350, 1214–5 (2015). [DOI] [PubMed] [Google Scholar]
- Bhute S. S. et al. Gut Microbial Diversity Assessment of Indian Type-2-Diabetics Reveals Alterations in Eubacteria, Archaea, and Eukaryotes. Front. Microbiol 8, 214 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumbhare S. V et al. A cross-sectional comparative study of gut bacterial community of Indian and Finnish children. Sci. Rep 7, 10555 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Das A. et al. Gastric microbiome of Indian patients with Helicobacter pylori infection, and their interaction networks. Sci. Rep 7, 15438 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chauhan N. S. et al. Western Indian Rural Gut Microbial Diversity in ExtremePrakritiEndo-Phenotypes Reveals Signature Microbes. Front. Microbiol 9, 118 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinha R. et al. Assessment of variation in microbial community amplicon sequencing by the Microbiome Quality Control (MBQC) project consortium. Nat. Biotechnol. 35, 1077–1086 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siegwald L. et al. Assessment of Common and Emerging Bioinformatics Pipelines for Targeted Metagenomics. PLoS One 12, e0169563 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakayama J. et al. Diversity in gut bacterial community of school-age children in Asia. Sci. Rep 5, 8397 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pyky R. et al. Profiles of sedentary and non-sedentary young men - a population-based MOPO study. BMC Public Health 15, 1164 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Managing Overweight and Obesity in Adults: Systematic Evidence Review from the Obesity Expert Panel. https://www.nhlbi.nih.gov/health-topics/managing-overweight-obesity-in-adults (2013).
- Abrahamson M., Hooker E., Ajami N. J., Petrosino J. F. & Orwoll E. S. Successful collection of stool samples for microbiome analyses from a large community-based population of elderly men. Contemp. Clin. Trials Commun 7, 158–162 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milani C. et al. Assessing the fecal microbiota: an optimized ion torrent 16S rRNA gene-based analysis protocol. PLoS One 8, e68739 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Claesson M. J. et al. Comparative analysis of pyrosequencing and a phylogenetic microarray for exploring microbial community structures in the human distal intestine. PLoS One 4, e6669 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDonald D. et al. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 6, 610–8 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgulis A. et al. Database indexing for production MegaBLAST searches. Bioinformatics 24, 1757–64 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caporaso J. G. et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–6 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schloss P. D. et al. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75, 7537–41 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Al-Hebshi N. N., Nasher A. T., Idris A. M. & Chen T. Robust species taxonomy assignment algorithm for 16S rRNA NGS reads: application to oral carcinoma samples. J. Oral Microbiol 7, 28934 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfeiffer S. et al. Improved group-specific primers based on the full SILVA 16S rRNA gene reference database. Environ. Microbiol. 16, 2389–407 (2014). [DOI] [PubMed] [Google Scholar]
- Bacci G. et al. Evaluation of the Performances of Ribosomal Database Project (RDP) Classifier for Taxonomic Assignment of 16S rRNA Metabarcoding Sequences Generated from Illumina-Solexa NGS. J. Genomics 3, 36–9 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Citations
- 2018. European Nucleotide Archive. PRJEB25642
- Dubey A. K., et al. . 2018. figshare. https://doi.org/10.6084/m9.figshare.c.4147079