

Abstract
Findings from studies using animal models expressing amyotrophic lateral sclerosis (ALS) mutations in RNA-binding proteins, such as Transactive Response DNA-binding protein-43 (TDP-43), indicate that this protein, which is involved in multiple functions, including transcriptional regulation and pre-mRNA splicing, represents a key candidate in ALS development. This study focuses on characterizing, in a Drosophila genetic model of ALS (TDP-43), the effects of Mucuna pruriens (Mpe) and Withania somnifera (Wse). Electrophysiological and behavioural data in TDP-43 mutant flies revealed anomalous locomotion (i.e. impaired climbing with unexpected hyperactivity) and sleep dysregulation. These features, in agreement with previous findings with a different ALS model, were at least partially, rescued by treatment with Mpe and Wse. In addition, electrophysiological recordings from dorsal longitudinal muscle fibers and behavioral observations of TDP-43 flies exposed to the volatile anaesthetics, diethyl ether or chloroform, showed paradoxical responses, which were normalized upon Mpe or Wse treatment. Hence, given the involvement of some potassium channels in the effects of anaesthetics, our results also hint toward a possible dysregulation of some potassium channels in the ALS-TDP-43 Drosophila model, that might shed new light on future therapeutic strategies pertaining to ALS.
Introduction
Alterations in the expression levels of the highly conserved transactive response DNA binding protein 43 (TDP-43), cause amyotrophic lateral sclerosis (ALS) and fronto-temporal dementia (FTD)1–5. Apart from mutations in the genes encoding superoxide dismutase-1 (SOD1) and fused in sarcoma (FUS), variants of TDP-43 have been reported in almost all the patients afflicted with ALS6,7. TDP-43 is an ubiquitously expressed nuclear protein, harboring two RNA recognition motifs: a nuclear localization sequence with a nuclear export signal and a glycine-rich C-terminus1. Along with regulating transcriptional repression and splicing, TPD-43 also modulates micro RNA biogenesis8–13.
Both, the over-expression of TDP-43, leading to its cytoplasmic accumulation (i.e. cytotoxic gain-of-function), and the nuclear-loss of TDP-43 (i.e. loss of function), have been shown to progress to neurodegeneration (reviewed in9,14). A number of studies performed in mice, zebrafish and Drosophila have shown that both gain- and loss- of TDP-43 function lead to impaired motor behaviour and loss of motoneurons15–19. Interestingly, in Zebra fish and Drosophila, the loss-of-function phenotype associated with motor impairment could be rescued by expressing wild-type TDP-43, suggestive of plasticity in the neural circuit17,18. Moreover, as suggested by Cragnaz et al.20, the age-related drop in TDP-43 expression might likely be a dramatic event which promotes the onset of the human disease.
Remarkably, albeit Riluzole has been approved more than 20 years ago by FDA and considered the recent approval of Radicava (edaravone) to treat patients with ALS, the most part of the currently available treatment still remains somewhat of a conundrum, making the pursuit for new therapeutic strategies essential.
Several traditional medicinal systems, such as Ayurveda, provide a thorough body of literature on medicinal plants, which can be exploited by rational drug design and development of novel pharmaceutical approaches. Mucuna pruriens (L.) DC. (Mpe) and Withania somnifera (L.) Dunal (Wse) have been widely used in Ayurvedic medicine for their potential effects in treating neurodegenerative disorders21. Although isolated components of Wse have already been studied in detail, pointing out withanolides and withanone for their crucial role in neuroprotection22–24, in our previous studies we tested the effectiveness of methanolic extracts, of Mpe and Wse in Drosophila models of Parkinson’s disease25,26, and of ALS (ALS-hSOD127). In the hSOD1 model, treatment with Mpe and Wse remedied the locomotor defects present in this fly phenotype27. Based on the above observations and according to the study by Cragnaz et al.20 and Feiguin et al.28 that correlated TDP-43 levels with motor impairment, we postulated that Mpe and Wse treatments may prove effective in restoring the function of motoneurons in a partial TDP-43-loss-of-function Drosophila model. Our present study exploits advanced genetic tools in Drosophila, to mimic a phenotype associated to reduced levels of TDP-43 expression in motoneurons and examine whether Mpe and Wse hold therapeutic potential in the treatment of ALS.
Results
Mpe or Wse treatment ameliorates the climbing and locomotion behaviour of TDP-43 mutant flies
The GAL4-Upstream Activation Sequence (UAS) system was used to specifically knockdown TAR DNA-binding protein-43 homolog (TBPH) in motoneurons, in order to obtain a partial depletion of the protein. D42 GAL4 driven RNA interference (RNAi) mediated knockdown of TBPH (hence forth called as TDP-43 mutant flies) impaired the climbing and locomotion behaviour in flies (Fig. 1).
Figure 1.
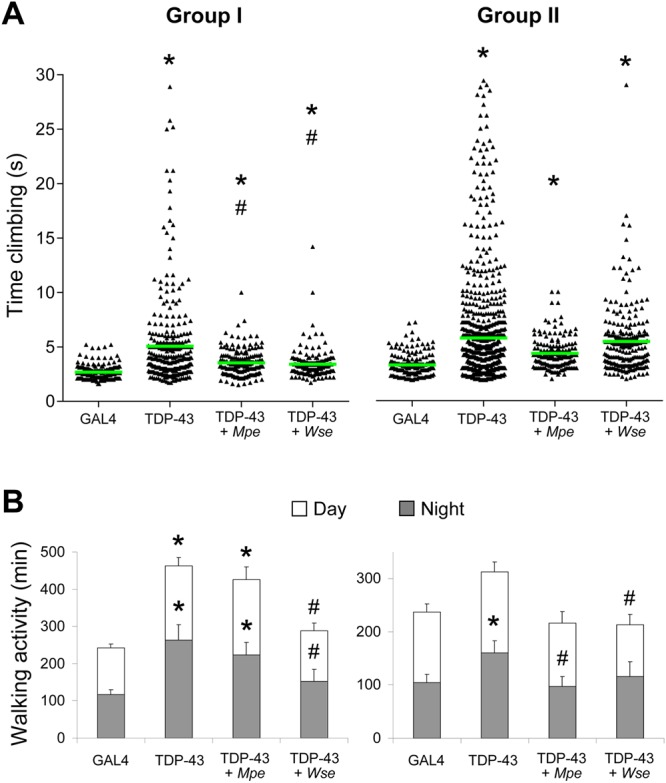
Mpe or Wse treatment ameliorates the climbing and locomotion behaviour of TDP-43 mutant flies. Effects of Mucuna pruriens (Mpe) or Withania somnifera (Wse) extracts on the climbing (A) and walking (B) activity of adult GAL4 flies, untreated mutants (TDP-43) and Mpe- or Wse-treated TDP-43. The treatment was administered during the adult stage and its effect was assayed in two age groups (Group I: 3–6 day old flies, Group II: 10–15 day old flies). In each set of data in (A) the green horizontal line indicates the mean of the corresponding distribution. * and # indicate p < 0.05, one-way ANOVA (non parametric) followed by Dunn’s multiple comparisons test, compared to GAL4 and TDP-43, respectively. The histograms in (B) display the walking activity during the 12-h daytime (white) and 12-h nighttime (black) periods. * and # indicate p < 0.05, one-way ANOVA followed by HSD post-hoc test, compared to GAL4 and TDP-43, respectively.
The behavioural performance of TBPH knockdown was studied in two age groups (I: 3–6 and II: 10–15 day-old male flies). As shown in Fig. 1A, TDP-43 mutant flies showed a significant increase in the climbing time in both age groups tested with respect to the control. We also assessed the effects of Mpe or Wse on the climbing behaviour of TDP-43 mutant flies, treated with Mpe or Wse supplemented in food (0.1% w/w) upon eclosion. Treatment of group I TDP-43 mutant flies with either Mpe or Wse (0.1% w/w) significantly reduced the climbing time (p < 0.05, ANOVA, non parametric), suggesting partial recovery of the motoneuron impairment. In the second group, no significant difference was found between treated and untreated TDP-43 mutant flies. Therefore, the climbing performance of TDP-43 mutant flies declines with age, both in treated and untreated ones, even if in the latter a trend to recovery was found.
After observing a recovery in the climbing behaviour of TDP-43-treated flies, we reasoned that the overall locomotion activity of TDP-43 mutant flies could be restored upon treatment (Fig. 1B). Hence, the locomotion of groups I and II flies was recorded using a drosophila activity monitor (DAM2, trikinetics) and plotted as daytime and nighttime activity. The group I and II TDP-43 mutant flies showed heightened locomotion activity compared to GAL4 control flies. Mpe treated (0.1% w/w) group I TDP-43 flies exhibited a slight reduction in the overall locomotion activity compared to TDP-43 flies. On the other hand, Wse (0.1% w/w) pre-treated group-I TDP-43 flies showed a markedly decreased daytime and nighttime locomotion activity. Interestingly, the overall locomotion activity of Mpe treated group II TDP-43 flies was comparable to that of GAL4 flies, indicating that continuous administration of Mpe may have beneficial effects. In addition, group II TDP-43 flies showed locomotion comparable to GAL4 flies after Wse treatment. Collectively, these findings suggest that Mpe and Wse ameliorate the hyperactive phenotype of TDP-43 flies.
Mpe or Wse restore changes in ePSPs recorded from DLM of TDP-43 mutant flies
We then examined the function of the dorsal longitudinal muscle (DLM) neuromuscular junction in TDP-43 mutant flies. Kinetic properties (i.e. the amplitude and latency) of evoked post-synaptic potentials (ePSPs), recorded from the DLM after Giant Fibre System (GFS) electrical stimulation, were evaluated under basal conditions. The amplitude and latency of ePSPs recorded from the DLM muscle of GAL4 flies had an averaged value of 26.13 ± 2.59 mV and 0.94 ± 0.07 ms, respectively (Fig. 2A–C).
Figure 2.

Effect of TDP-43 loss-of-function mutation and treatment with Mpe or Wse on electrophysiological parameters recorded from DLM. (A) Representative traces obtained from different experimental conditions, in which ePSP latency is calculated as the time (ms) from stimulus application to the peak of post-synaptic potential (PSP) (red scattered lines), and PSP peak is calculated by measuring the maximal amplitude of the response starting from the baseline. Scale bar, 10 mV/1 ms. (B) Ámplitude (mV) and (C) latency (ms) recorded in flies from different experimental groups. Bar graphs represent the mean ± standard error of the mean (SEM) of ePSPs. * indicates p < 0.05 compared to GAL4, one-way ANOVA, followed by Bonferroni post-hoc test. N = 22 for all experimental groups. (D) Representative traces obtained from different experimental groups in which PSPs were evoked in response to 10 stimuli delivered at 100 Hz. Scale bar 10 mV/20 ms. * indicates the presence of the event. (E) Scatter plot graphs showing the changes in ePSPs amplitude following stimulation at increasing frequency (from 10 to 200 Hz) of a train of 10 consecutive stimuli (the effect at 100 Hz is highlighted in yellow). All values are expressed as the mean ± SEM of the % of failure observed in every train. * indicates p < 0.05 vs GAL4. (F) Bar graph representing the averaged % of failure that was plotted in all experimental groups. “Strike” is the % of recordings where failure was not greater that 10%, while “failure” is the number of recordings where the % of failure was greater than 20%. Data corresponds to the % of failures evaluated @ 100 Hz. * indicates p < 0.05 compared to GAL4, unpaired t-test.
Interestingly, ePSPs recorded in TDP-43 mutant flies were characterized by a significant decrease in amplitude (20.32 ± 2.7 mV; p < 0.05 vs GAL4, ANOVA) and an increase in latency (1.4 ± 1.1 ms; p < 0.05 vs GAL4, ANOVA) (Fig. 2A–C). These data suggest that the TDP-43 mutation may hamper the function of GFS-DLM neuronal conduction as well as muscle efficiency. As observed in previous reports with flies carrying different SOD1 mutations27, the treatment of TDP-43 mutants with Mpe or Wse significantly restored the changes in amplitude and latency of ePSPs, with values indistinguishable from those observed in GAL4 flies (p < 0.05; Fig. 2A–C).
Mpe but not Wse ameliorates the ePSP response to high frequency stimulation of GFS in TDP-43 mutant flies
As reported previously26,27,29, recording the “frequency of following”, which consists of applying a train of 10 stimuli to the GFS at different frequencies (from 10 to 200 Hz, with steps of 25 Hz), allows physiological studies on the GFS-DLM muscle fibre. As expected, the number of failures in GAL4 flies was very low (8.7%) at 100 Hz but dramatically increased (28.6%) at 200 Hz (Fig. 2D,E). Remarkably, when TDP-43 flies were exposed to the same protocol, the rate of failures at both 100 and 200 Hz resulted significantly enhanced with respect to GAL4 flies (p < 0.05, ANOVA) (Fig. 2D–F). The treatment of TDP-43 flies with Mpe but not Wse was able to antagonize the increased failures found in TBP-43 flies in response to train stimulations (100 Hz), resulting in values comparable to those observed for the GAL4 flies (Fig. 2D–F).
Mpe or Wse treatment modulates sleep parameters inTDP-43 mutant flies
We next studied the effects of Mpe or Wse treatment (0.1% w/w) on sleep time of TDP-43 mutant flies. Both groups I and II TDP-43 mutant flies showed reduced nighttime sleep duration, compared to GAL4 control flies. In agreement with the locomotion data (Fig. 1B), TDP-43 flies of both age groups treated with Mpe or Wse showed a significantly restored the nighttime sleep duration (Fig. 3A,B).
Figure 3.

Mpe or Wse treatment modulates sleep parameters inTDP-43 mutant flies. Effects of Mucuna pruriens (Mpe) or Withania somnifera (Wse) extracts on total sleep (A,B), the number of sleep episodes (C,D) during the 12-h daytime (white) and 12-h nighttime (gray) periods and on the average of longest sleep episodes (AvLo)/24 h (E,F) of adult GAL4 flies, untreated mutants (TDP-43) and Mpe- and Wse-treated TDP-43 flies. The treatment was administered during the adult stage and its effect assessed in two groups (Group I: 7 days and Group II: 14 days) of the flies’ life-span. The top and bottom of the box and whisker plots show the upper and lower quartiles, respectively. The horizontal line in the middle indicates the median of the corresponding distribution, while the minimum and maximum observed values are indicated by the bars connected to the box. * and # indicate p < 0.05, one-way ANOVA followed by HSD post-hoc test, compared to GAL4 and TDP-43, respectively.
Nighttime sleep is associated with long sleep episodes, which usually result in a reduced number of sleep episodes, a low number of long sleep episodes being indicative of a less fragmented sleep pattern30. TDP-43 mutant flies showed an increased number of sleep episodes compared to GAL4 flies in both age groups (Fig. 3C,D). Mpe treated group I and II TDP-43 mutant flies showed a reduced number of sleep episodes, although this reduction was not statistically significant for the flies of group II. Wse treatment significantly reduced the number of sleep episodes of TDP-43 mutant flies in both groups (Fig. 3C,D). The data are consistent with the notion that the TDP-43 mutation leads to sleep fragmentation, most likely due to hyperactivity31. The decrease in daily sleep (Fig. 3C,D) negatively correlated with the increase in locomotor activity. Moreover, the average of longest sleep episodes in TDP-43 mutants is significantly lower with respect to GAL4 and TDP-43 treated flies, thus suggesting that treatment ameliorates sleep fragmentation, although to a different extent (Fig. 3E,F).
Mpe or Wse treatment prevents the effect of ether or chloroform administration on spontaneous PSPs in TDP-43 mutant flies
By considering that TDP-43 mutants show sleep parameters (fragmented sleep and motor activity) similar to those usually exhibited by Shaker flies (SH)32, in a separate set of these flies, we recorded the DLMs muscle contraction corresponding to a spontaneous wings beat, visualized as spike potentials frequency (Fig. 4), in the absence or after 30 s of diethyl ether (Et) or chloroform (Chl) vapour administration. For the sake of clarity, we specify that the noise level of traces was quite different between animals and strongly related to the experimental protocol by which, at variance with the experimental condition of data shown in Fig. 2, wings are left free to move, in order to also address the spontaneous activity of DLM.
Figure 4.

Mpe or Wse treatment prevents the effect of ether and chloroform administration on spontaneous PSPs in TDP-43 mutant flies. Treatment with Mpe or Wse rescues the effect of TDP-43 loss-of-function mutation to the enhancement of frequency of spike potentials due to Drosophila DLMs spontaneous contractions induced by 30 s of diethyl ether (Et) or chloroform (Chl) exposure. (A–H) Representative traces and plots obtained from GAL4 flies, TPD-43 treated and untreated mutants in which spontaneous PSPs were recorded in the absence, after 30 s and after 4 min wash-out of anaesthetic exposure. Arrows in graphs indicate when vapours of volatile anaesthetics delivery was begun. Scale bars: left panels, 20 mV/2 s, right panels 10 mV/2 s.
Application of both volatile anaesthetics induced a rapid deep anaesthesia. Right after diethyl ether or chloroform application, the spontaneous activity of DLM was completely, but reversibly abolished (only for diethyl ether but not for chloroform) in GAL4 flies (Fig. 4A,E). Interestingly, when TDP-43 mutant flies were tested with the same protocol, the spike potentials frequency was markedly increased after 30 s of diethyl ether or chloroform application. In TDP-43 mutants treated with either Mpe or Wse, the effect of both anaesthetics application on spontaneous DLM contractions was similar to that observed in GAL4 flies (Fig. 4C–H). We tested the same protocol also in SH flies and we observed a dramatic increase of DLM contractions right after diethyl ether application (Supplementary Fig. S1), while chloroform administration failed to induce the same effect.
Discussion
In our earlier studies in a Drosophila model of ALS-SOD1 overexpression, we reported that Mpe and Wse extracts ameliorated SOD1-induced pathologies27. We point out that our ALS TDP-43 mutants used in the present study differ with respect to the above cited genetic Dm model, being the current a model with a knock-down gene, that brings to a reduced level of TDP-43 expression. This leads to impaired climbing and locomotor abilities, although with a great inhomogeneity, that might be likely due to inhomogeneous levels of TDP-43, according to the previous study by Feiguin et al.28. Also, according to Cragnaz et al.20, age-related drop in TDP-43 expression may account for the worsening of climbing and locomotion activity in both untreated and treated mutants. In fact, scattering of data is dramatically increased in the oldest (second) group of flies.
Interestingly, also shorter sleep time and an increased number of nighttime sleep episodes were shown, effects that may be correlated with the fragmented sleep commonly observed in ALS patients33. The overall hyper locomotion of TDP-43 flies was recovered by Wse treatment, while Mpe treatment proved to be effective only in the 10–15 day old flies. In addition, treatment with Mpe and Wse improved the nighttime sleep duration and reduced the number of sleep episodes. The ePSPs recordings from DLM of TDP-43 mutant flies revealed that both Mpe and Wse may restore neuromotor function. In agreement with our results, a recent study on a mouse model of ALS, expressing the humanTDP-43A315T mutation which affects motor function, showed that Wse treatment can reduce Nuclear Factor Kappa Beta (NF-κB) activation and neuroinflammation, and ameliorate motor defects34. An interesting possibility to interpret these results might be that Mpe and Wse optimize TDP-43 expression levels, as suggested also by Cragnaz et al.20 and Feiguin et al.28. Furthermore, TDP-43 mutant flies anaesthetized using both diethyl ether and chloroform (Supplementary Videos S1 and S2) showed a spastic paralysis of the legs coupled to a shaker movement not found in GAL4 flies (Supplementary Video S3); the latter is a phenotype typically seen in voltage-gated potassium channel mutant flies (Shaker and Ether-a’-Go-Go). The voltage-gated potassium channel mutant flies (SH), on the contrary, showed DLM contractions and shaker movements upon diethyl ether but not chloroform treatment (Supplementary Video S4). In mammals, diethyl ether and chloroform are known to activate TREK-1, a two-pore-domain potassium (K2P) channel35. K2P channels maintain hyperpolarized resting membrane potential of various cell types36. Our studies suggest that, similar to the mammalian TREK1 channel, at least one of the Drosophila K2P channels responds to diethyl ether and chloroform vapours and reasonably reduces glutamatergic transmission at the DLM motoneurons of wild-type flies37. In TDP-43 flies, impaired K2P functioning may fit with the disinhibition of presynaptic glutamate release, which eventually leads to increased spike potential frequency after diethyl ether and chloroform application (Fig. 4B). Importantly, the shaking TDP-43 phenotype was remedied in mutant flies treated with Mpe or Wse (see Supplementary Videos S5 and S6). Even if our findings on different effects on volatile anaesthetics suggest an involvement of K2P potassium channels, it would be important to deeply investigate this potential mechanism in TDP-43 mutant flies. Overall, the results reported in the present study suggest a novel and potential therapeutic approach toward the medication or at least the slow-down of the progression of ALS, and provide new evidences on the pathophysiology of this neurodegenerative disease suggesting future experiments to characterize the mechanism of this potential therapeutic action of Mpe and Wse.
All this notwithstanding, we acknowledge that this investigation refers to the effects of two extracts characterized by a complex composition and therefore we cannot make any hypothesis on the possible effects of single constituents that may possibly be responsible of the observed effects. Anyway, we cannot state that the effects of the individual constituents would reproduce the effects of the whole extract as exemplified by the case of Hypericum perforatum whose antidepressant effects cannot be reproduced by the individual active constituents38. In this respect the whole extracts of Mucuna pruriens and Withania somnifera were demonstrated to be effective in counteracting neurodegenerations as shown by us in previous studies25–27. Hence, at the present we can conclude that these whole extracts, besides being therapeutically effective, as shown in the present and other22 studies, are endowed with significant properties such as being safe39–41 as documented by the use in Ayurvedic medicine since many centuries, cheap and easy to be administered.” Obviously, it is necessary to assess the proper concentration and duration of treatment as these phytotherapic extracts exert their effects as a drug following a hormesis-like dose-response curve27.
Methods
Flies
Flies were reared on a standard cornmeal-yeast-agar medium in controlled environmental conditions (24–25 °C; 60% relative humidity; light/dark = 12/12). D42 GAL4 flies and RNAi line against TBPH were obtained from Bloomington Stock Center (TBPH-RNAi: 39014, D42GAL4: 42737). Flies were isogenized by backcrossing for more than five generations. Only male flies were used for the experiments.
Pharmacological treatment
Flies were reared on a standard medium supplemented with Mucuna pruriens (Mpe) or Withania somnifera (Wse) extracts (kindly provided by Natural Remedies Pvt. Ltd., Bangalore, India). In order to follow good practice in publishing studies on herbal medicine42, we provided full details of the batches used (Mpe Batch No. RD/3208; Wse Batch No. PC/FWS1701003; Supplementary Figs S2 and S3).
In agreement with our previous studies25–27, Transactive Response DNA-binding protein-43 (TDP-43) mutants were supplied with Mpe or Wse at 0.1% w/w concentrations as adults (L-/A+), that, according to the life span curves, showed a lack of toxic effects (Supplementary Fig. S4). It is to be pointed out that we tried a higher concentration (1% w/w) and did not see any toxic effect. Even if it was not the aim of this study, at the concentration reported in the Supplementary Fig. S4, we observed a significant amelioration of the life span after Mpe and Wse treatment.
Climbing assay
The climbing assay (negative geotaxis assay) was performed as previously reported25–27,43,44 with some modifications. Briefly, based upon age and in order to verify any age-dependent impairment, flies were divided into two groups (I: 3–6; II: 10–15 days old) of untreated-GAL4, Mpe-treated, and Wse-treated TDP-43 mutants. Cohorts of at least 60 flies from each experimental group were subjected to the assay. Flies were placed individually in a vertically-positioned plastic tube (length 10 cm; diameter 1.5 cm) and tapped to the bottom. Climbing time (seconds) was recorded upon crossing a line drawn at 6 cm from the bottom. Data were expressed as average ± standard error of the mean (SEM) from three to five experiment replications (n flies from 180 to 400). Statistically significant differences (p < 0.05) were analyzed between GAL4 vs. TDP-43 and between untreated-TDP-43 vs. treated ones by means of the one-way ANOVA (non parametric), followed by Dunn’s multiple comparisons test.
Locomotor and sleep assay
The effects of Mpe and Wse were assayed at different age steps (at 7, 14 days treatment, hereafter referred to as group I and group II, respectively) in male TDP-43 e GAL4. Twenty - four (12 light:12 dark, thereafter indicated to as daytime and nighttime, respectively) hours of motor activity were recorded by Drosophila Activity Monitor System (DAMS – Trikinetics, Waltham, MA) from enclosure to 1 and 2 weeks of treatment with Mpe and Wse at 0.1% w/w. After one week and two weeks of treatment both mutants and GAL4 flies were isolated and each fly was housed in glass tubes (5 mm diameter by 65 mm length). Flies were given 24 hr to habituate to the experimental conditions prior to data collection. 10–15 flies were gathered and placed in each monitor for each genotype for each experiment. Motor activity in the tube was recorded and determined as each fly moves back and forth in the tube interrupting the infrared light beam that bisects each tube. Every time the fly crosses the tube is counted as a movement counted over 1-min periods. Drosophila Activity Monitor System was housed inside an environmental chamber where temperature and humidity were kept constant. Data for sleep analysis were acquired and analyzed with a dedicated software (pySolo)45. Sleep was defined as any period of uninterrupted behavioural immobility (0 counts/min) lasting >5 min according to Bushey et al.32. We considered, for each strain and age group, the total sleep, the number of episodes and the average of longest (AvLo) sleep episodes.
Statistically significant differences (p < 0.05) were analyzed between GAL4 vs. TDP-43 (*) and between untreated-TDP-43 vs. treated ones (#) by means of one-way ANOVA followed by HSD post-hoc test.
Electrophysiological recordings
Flies from the different experimental groups were anesthetized by using CO2 and tightly anchored to a wax support with ventral side down, before starting the experiment, as previously reported26, and visualized under a stereomicroscope. Giant fiber system (GFS) was activated by a two tungsten stimulating electrodes, connected to a stimulator (Master 8, A.M.P.I, Jerusalem, IL, USA) triggered by a stimulus isolation unit (DS2A, Digitimer Ltd., Hertfordshire UK), were placed into both eyes of the fly. The intensity of each stimulus was not greater than 10 V and was increased gradually until the postsynaptic potential response was observed. Moreover, no correlation between stimulation intensity needed to evoke the response and type of animal tested was observed. As potential reference electrode, a ground tungsten wire was placed into the fly abdomen. A borosilicate recording electrode, shaped by a horizontal puller (P97, Flaming/Brown, Sutter Instruments, Novato, CA, USA) with a resistance of 4–5 MΩ when filled with 3 M KCl, was placed into the right or left backside of the fly along the 45a and 45b fibres of the dorsal longitudinal muscle fibres (DLMs). Evoked post-synaptic potentials (ePSPs) were recorded with an Axopatch 2-B amplifier (Axon Instruments, Foster City, CA), filtered at 0.5 kHz and digitized at 1 kHz. ePSPs were recorded in bridge mode, measured using peak and event detection software pCLAMP 8.2 (Axon Instruments, Foster City, CA), and analyzed off-line by pCLAMP 8.2 fit software (Axon Instruments, Foster City, CA). Experiments were blind to the treatment. Electrophysiological experiments were performed by applying a protocol consisting in a single GFS stimulation, delivered every 20 s, followed by the ePSP recording. The “frequency of following” was determined by delivering trains of 10 stimuli at increasing frequencies (from 10 to 200 Hz) indicating the failures, as the percentage of lacking responses at each train. As previously published26,27,29, we also evaluated the amplitude (peak of the PSP expressed in mV) as well as the latency (interval between the stimulating artifact and the time at ePSP peak, expressed in ms). In a separate set of flies, the recording electrode was placed in DLMs and events derived from spontaneous muscle contraction, and thus wing beats, were recorded in the absence or after 30 s of ether or chloroform vapour administration to the fly in order to induce a deep anaesthesia. After one minute of spontaneous recording, anaesthetic vapours were applied through a silicon tube placed close to the fly and recording has been continued for at least 5 min. This protocol was applied to all different experimental groups. Data are expressed as mean + SEM and analyzed by one or two-way ANOVAs followed by Bonferroni’s post-hoc tests.
Electronic supplementary material
Acknowledgements
We thank Prof. Dr. Michael Freissmuth for helpful advices and discussions. We also thank Dr. Ignazio Collu (University of Cagliari) for taking care of flies.
Author Contributions
R.M., M.D.S., G.T. and A.L. conceived and designed the experiments. R.M., M.D.S., G.T., S.P. and P.M. performed the experiments. R.M., M.D.S., G.T., P.S. and A.L. analyzed the data. E.S., S.K., E.A. and A.L. contributed reagents/materials/analysis tools. R.M, M.D.S., G.T., A.K., S.S. and A.L. contributed to the interpretation of the data and wrote the manuscript. All the Authors reviewed and approved the manuscript.
Competing Interests
The authors declare no competing interests.
Footnotes
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Riccardo Maccioni, Maria Dolores Setzu and Giuseppe Talani contributed equally.
Electronic supplementary material
Supplementary information accompanies this paper at 10.1038/s41598-018-34452-1.
References
- 1.Wang HY, Wang IF, Bose J, Shen CK. Structural diversity and functional implications of the eukaryotic TDP gene family. Genomics. 2004;83:130–139. doi: 10.1016/S0888-7543(03)00214-3. [DOI] [PubMed] [Google Scholar]
- 2.Ayala YM, et al. Human, Drosophila, and C. elegans TDP43: nucleic acid binding properties and splicing regulatory function. J. Mol. Biol. 2005;348:575–588. doi: 10.1016/j.jmb.2005.02.038. [DOI] [PubMed] [Google Scholar]
- 3.Arai T, et al. TDP-43 is a component of ubiquitin-positive tau-negative inclusions in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Biochem. Biophys. Res. Commun. 2006;351:602–611. doi: 10.1016/j.bbrc.2006.10.093. [DOI] [PubMed] [Google Scholar]
- 4.Neumann M, et al. Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science. 2006;314:130–133. doi: 10.1126/science.1134108. [DOI] [PubMed] [Google Scholar]
- 5.Sreedharan J, et al. TDP-43 mutations in familial and sporadic amyotrophic lateral sclerosis. Science. 2008;319:1668–1672. doi: 10.1126/science.1154584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Mackenzie IR, et al. Pathological TDP-43 distinguishes sporadic amyotrophic lateral sclerosis from amyotrophic lateral sclerosis with SOD1 mutations. Ann. Neurol. 2007;61:427–434. doi: 10.1002/ana.21147. [DOI] [PubMed] [Google Scholar]
- 7.Kwiatkowski TJ, et al. Mutations in the FUS/TLS gene on chromosome 16 cause familial amyotrophic lateral sclerosis. Science. 2009;323:1205–1208. doi: 10.1126/science.1166066. [DOI] [PubMed] [Google Scholar]
- 8.Gregory RI, et al. The microprocessor complex mediates the genesis of microRNAs. Nature. 2004;432:235–240. doi: 10.1038/nature03120. [DOI] [PubMed] [Google Scholar]
- 9.Buratti E, Baralle FE. The molecular links between TDP-43 dysfunction and neurodegeneration. Adv. Genet. 2009;66:1–34. doi: 10.1016/S0065-2660(09)66001-6. [DOI] [PubMed] [Google Scholar]
- 10.Bose JK, Huang CC, Shen CK. Regulation of autophagy by neuropathological protein TDP-43. J. Biol. Chem. 2011;286:44441–44448. doi: 10.1074/jbc.M111.237115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Tollervey JR, et al. Characterizing the RNA targets and position-dependent splicing regulation by TDP-43. Nat. Neurosci. 2011;14:452–458. doi: 10.1038/nn.2778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ayala YM, et al. TDP-43 regulates its mRNA levels through a negative feedback loop. EMBO J. 2011;30:277–288. doi: 10.1038/emboj.2010.310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Polymenidou M, et al. Long pre-mRNA depletion and RNA missplicing contribute to neuronal vulnerability from loss of TDP-43. Nat. Neurosci. 2011;14:459–68. doi: 10.1038/nn.2779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wang IF, Wu LS, Chang HY, Shen CK. TDP-43, the signature protein of FTLD-U, is a neuronal activity-responsive factor. J. Neurochem. 2008;105:797–806. doi: 10.1111/j.1471-4159.2007.05190.x. [DOI] [PubMed] [Google Scholar]
- 15.Wegorzewska I, Bell S, Cairns NJ, Miller TM, Baloh RH. TDP-43 mutant transgenic mice develop features of ALS and frontotemporal lobar degeneration. Proc. Natl. Acad. Sci. USA. 2009;106:18809–18814. doi: 10.1073/pnas.0908767106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Shan X, Chiang PM, Price DL, Wong PC. Altered distributions of Gemini of coiled bodies and mitochondria in motor neurons of TDP-43 transgenic mice. Proc. Natl. Acad. Sci. USA. 2010;107:16325–16330. doi: 10.1073/pnas.1003459107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Diaper DC, et al. Loss and gain of Drosophila TDP-43 impair synaptic efficacy and motor control leading to age-related neurodegeneration by loss-of-function phenotypes. Hum. Mol. Genet. 2014;22:1539–1557. doi: 10.1093/hmg/ddt005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kabashi E, et al. Investigating the contribution of VAPB/ALS8 loss of function in amyotrophic lateral sclerosis. Hum. Mol. Genet. 2013;22:2350–2360. doi: 10.1093/hmg/ddt080. [DOI] [PubMed] [Google Scholar]
- 19.Vanden Broeck L, et al. TDP-43 loss-of-function causes neuronal loss due to defective steroid receptor-mediated gene program switching in Drosophila. Cell. Rep. 2013;3:160–172. doi: 10.1016/j.celrep.2012.12.014. [DOI] [PubMed] [Google Scholar]
- 20.Cragnaz L, et al. An age-related reduction of brain TBPH/TDP-43 levels precedes the onset of locomotion defects in a Drosophila ALS model. Neuroscience. 2015;311:415–421. doi: 10.1016/j.neuroscience.2015.10.037. [DOI] [PubMed] [Google Scholar]
- 21.Katzenschlager R, et al. Mucuna pruriens in Parkinson’s disease: a double blind clinical and pharmacological study. J. Neurol. Neurosurg. Psychiatry. 2004;75:1672–1677. doi: 10.1136/jnnp.2003.028761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Dar NJ, Hamid A, Ahmad M. Pharmacologic overview of Withania somnifera, the Indian Ginseng. Cell. Mol. Life Sci. 2015;72:4445–4460. doi: 10.1007/s00018-015-2012-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Dar NJ, et al. Withanone, an active constituent from Withania somnifera, affords protection against NMDA-induced excitotoxicity in neuron-like cells. Mol. Neurobiol. 2017;54:5061–5073. doi: 10.1007/s12035-016-0044-7. [DOI] [PubMed] [Google Scholar]
- 24.Dar NJ, Satti NK, Dutt P, Hamid A, Ahmad M. Attenuation of glutamate-induced excitotoxicity by Withanolide-A in neuron-like cells: role for PI3K/Akt/MAPK signaling pathway. Mol. Neurobiol. 2018;55:2725–2739. doi: 10.1007/s12035-017-0515-5. [DOI] [PubMed] [Google Scholar]
- 25.Poddighe S, et al. Mucuna Pruriens (Velvet bean) rescues motor, olfactory, mitochondrial and synaptic impairment in PINK1B9 Drosophila melanogaster genetic model of Parkinson’s disease. PLoS ONE. 2014;9:e110802. doi: 10.1371/journal.pone.0110802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.De Rose F, et al. Functional and morphological correlates in the Drosophila LRRK2 loss-of-function model of Parkinson’s disease: drug effects of Withania somnifera (Dunal) administration. PLoS ONE. 2016;11:e0146140. doi: 10.1371/journal.pone.0146140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.De Rose F, et al. Differential effects of phytotherapic preparations in the hSOD1 Drosophila melanogaster model of ALS. Sci. Rep. 2017;7:41059. doi: 10.1038/srep41059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Feiguin F, et al. Depletion of TDP-43 affects Drosophila motoneurons terminal synapsis and locomotive behavior. FEBS Lett. 2009;583:1586–1592. doi: 10.1016/j.febslet.2009.04.019. [DOI] [PubMed] [Google Scholar]
- 29.Augustin H, Allen MJ, Partridge L. Electrophysiological recordings from the giant fiber pathway of D. melanogaster. J. Vis. Exp. 2011;14:2412. doi: 10.3791/2412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Huber R, et al. Sleep homeostasis in Drosophila melanogaster. Sleep. 2004;27:628–639. doi: 10.1093/sleep/27.4.628. [DOI] [PubMed] [Google Scholar]
- 31.Estes PS, et al. Motor neurons and glia exhibit specific individualized responses to TDP-43 expression in a Drosophila model of amyotrophic lateral sclerosis. Dis Model Mech. 2013;6:721–733. doi: 10.1242/dmm.010710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bushey D, Huber R, Tononi G, Cirelli C. Drosophila hyperkinetic mutants have reduced sleep and impaired memory. J. Neurosci. 2007;27:5384–5393. doi: 10.1523/JNEUROSCI.0108-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lo Coco D, Mattaliano P, Spataro R, Mattaliano A, La Bella V. Sleepwake disturbances in patients with amyotrophic lateral sclerosis. J. Neurol. Neurosurg. Psychiatry. 2011;82:839–842. doi: 10.1136/jnnp.2010.228007. [DOI] [PubMed] [Google Scholar]
- 34.Dutta K, Patel P, Rahimian R, Phaneuf D, Julien JP. Withania somnifera reverses transactive response DNA binding protein 43 proteinopathy in a mouse model of amyotrophic lateral sclerosis/frontotemporal lobar degeneration. Neurotherapeutics. 2017;14:447–462. doi: 10.1007/s13311-016-0499-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Patel AJ, et al. Inhalational anesthetics activate two-pore-domain background K+ channels. Nat. Neurosci. 1999;2:422–426. doi: 10.1038/8084. [DOI] [PubMed] [Google Scholar]
- 36.Lesage F, Lazdunski M. Molecular and functional properties of two-pore-domain potassium channels. Am. J. Physiol. Renal. Physiol. 2000;279:793–801. doi: 10.1152/ajprenal.2000.279.5.F793. [DOI] [PubMed] [Google Scholar]
- 37.Franks NP, Honoré E. The TREK K2P channels and their role in general anaesthesia and neuroprotection. Trends Pharmacol. Sci. 2004;25:601–608. doi: 10.1016/j.tips.2004.09.003. [DOI] [PubMed] [Google Scholar]
- 38.Kasture S, et al. Withania somnifera prevents morphine withdrawal-induced decrease in spine density in nucleus accumbens shell of rats: a confocal laser scanning microscopy study. Neurotox. Res. 2009;16:343–355. doi: 10.1007/s12640-009-9069-2. [DOI] [PubMed] [Google Scholar]
- 39.Mishra LC, Singh BB, Dagenais S. Scientific basis for the therapeutic use of Withania somnifera (ashwagandha): A review. Alternative Medicine Review. 2000;5:334–346. [PubMed] [Google Scholar]
- 40.Prabu PC, Panchapakesan S, Raj CD. Acute and sub-acute oral toxicity assessment of the hydroalcoholic extract of Withania somnifera roots in Wistar rats. Phytother Res. 2013;27:1169–78. doi: 10.1002/ptr.4854. [DOI] [PubMed] [Google Scholar]
- 41.Patel SB, Rao NJ, Hingorani LL. Safety assessment of Withania somnifera extract standardized for Withaferin A: Acute and sub-acute toxicity study. J Ayurveda Integr Med. 2016;7:30–37. doi: 10.1016/j.jaim.2015.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Chan K, et al. Good practice in reviewing and publishing studies on herbal medicine, with special emphasis on traditional Chinese medicine and Chinese materia medica. J. Ethnopharmacol. 2012;140:469–475. doi: 10.1016/j.jep.2012.01.038. [DOI] [PubMed] [Google Scholar]
- 43.Liu Z, et al. A Drosophila model for LRRK2-linked parkinsonism. Proc. Natl. Acad. Sci. USA. 2008;105:2693–2698. doi: 10.1073/pnas.0708452105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Poddighe S, et al. Impaired sense of smell in a Drosophila Parkinson’s model. PLoS One. 2013;8:e73156. doi: 10.1371/journal.pone.0073156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Gilestro GF, Cirelli C. PySolo: A complete suite for sleep analysis in Drosophila. Bioinformatics. 2009;25:1466–1467. doi: 10.1093/bioinformatics/btp237. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.