

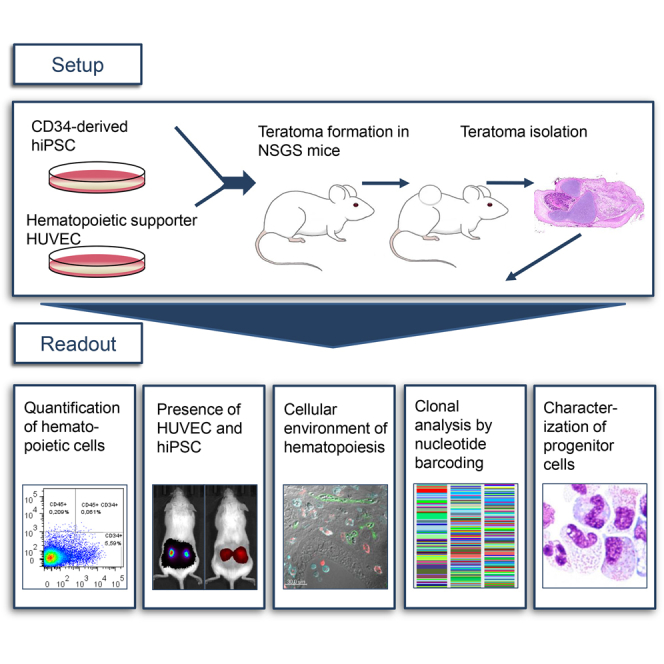
Summary
Hematopoietic stem cells (HSCs) ensure a life-long regeneration of the blood system and are therefore an important source for transplantation and gene therapy. The teratoma environment supports the complex development of functional HSCs from human pluripotent stem cells, which is difficult to recapitulate in culture. This model mimics various aspects of early hematopoiesis, but is restricted by the low spontaneous hematopoiesis rate. In this study, a feasible protocol for robust hematopoiesis has been elaborated. We achieved a significant increase of the teratoma-derived hematopoietic population when teratomas were generated in the NSGS mouse, which provides human cytokines, together with co-injection of human umbilical vein endothelial cells. Since little is known about hematopoiesis in teratomas, we addressed localization and clonality of the hematopoietic lineage. Our results indicate that early human hematopoiesis is closely reflected in teratoma formation, and thus highlight the value of this model.
Keywords: hematopoiesis, teratoma, hiPSC, EHT, genetic barcoding, HUVECs, embryogenesis
Graphical Abstract
Highlights
-
•
Robust human hematopoiesis in teratomas with co-injected HUVECs in NSGS mice
-
•
Hematopoietic progenitors localize inside vascular structures in teratomas
-
•
CD45+ cells are present in mesenchymal tissue in teratomas
-
•
Teratoma formation and subsequent hematopoiesis are polyclonal events
Schambach and colleagues show evidence that hematopoiesis in hiPSC-derived teratomas occurs by endothelial-to-hematopoietic transition in a highly polyclonal manner. This process was supported by the presence of HUVECs in the NSGS mouse, which provides human SCF, IL-3, and GM-CSF. The established hematopoiesis model can potentially be used for disease modeling and improvement of hematopoietic differentiation protocols.
Introduction
Tissue stem cells are the life-long source of organ regeneration. Unlike the origin of solid tissue, development of the hematopoietic stem cells (HSCs) proceeds through distinct stages during embryogenesis (Godin and Cumano, 2002, Mikkola, 2006). The definitive HSCs in vertebrates emerge during a process called endothelial-to-hematopoietic transition (EHT), from hemogenic endothelium (Bertrand et al., 2010, Kissa and Herbomel, 2010). Due to their long-term self-renewal capacities, HSCs are valuable for gene and cell therapies (Morgan et al., 2017). Although many aspects of HSC biology have been elucidated, it remains difficult to generate, maintain, or expand human HSCs. The possibility for de novo generation from human induced pluripotent stem cells (hiPSCs) would especially help to relieve the limitation of donor material.
Most differentiation protocols for modeling hematopoiesis are based on small molecules, cytokine supplementation, and timed expression of transcription factors (TFs), following general lines of developmental steps from PSCs to hematopoietic progenitors (Sugimura et al., 2017, Vo and Daley, 2015). These important models, however, fail to mimic the complexity of the cellular environment necessary to support HSC development. The teratoma formation assay offers a three-dimensional (3D) embryonic microenvironment with corresponding tissue patterning, epigenetic imprinting, and extracellular matrices (Damjanov and Andrews, 2016, Gertow et al., 2004). In this surrounding, a low level of spontaneous hematopoiesis occurs and gives rise to engraftable hematopoietic progenitors (Amabile et al., 2013, Suzuki et al., 2013). Implanted osmotic pumps delivering cytokines, such as thrombopoietin and stem cell factor (SCF), can increase the level of hematopoiesis in teratomas (Suzuki et al., 2013). Other protocols use co-injection of murine bone marrow stromal cells (OP9) together with hiPSCs. Further improvement was achieved by the co-injection of genetically modified OP9 expressing Notch ligand Delta-like 1 (Dll1) or Wnt3a (Amabile et al., 2013, Suzuki et al., 2013). Alternatively, murine iPSCs, equipped with an inducible TF (Gfi1b), led to elevated hematopoietic levels after administration of doxycycline (Tsukada et al., 2017). To overcome technical hurdles such as implantation of osmotic pumps, we developed a feasible and less demanding teratoma assay as a robust model for hematopoiesis.
Currently little is known about the origin or maintenance of human hematopoietic cells in teratomas. H&E staining identified hematopoietic cell phenotype in bone marrow-like structures in teratomas (Amabile et al., 2013). Others showed putative hematopoietic cells to reside in close proximity to liver cells, mesenchymal stroma, or yolk sac structures (Damjanov and Andrews, 2016). In a murine iPSC teratoma system, there is evidence that hematopoiesis occurs by EHT (Tsukada et al., 2017). Using our optimized system for human hematopoiesis, we extensively evaluated the origin and cellular environment of human hematopoietic cells in teratomas via detailed multicolor fluorescent immunohistochemistry (IHC).
To use teratomas to investigate processes of human embryogenesis or hematopoiesis, improved characterization of clonal development is necessary. Blum and Benvenisty (2007) addressed this by mixing three human embryonic stem cell (hESC) lines for teratoma induction. Microsatellite signatures in microdissections of teratoma structures revealed a mixed composition of hESC lines, indicating a polyclonal origin. This important finding illustrates the high impact of environmental cues on differentiation rather than clonal preferences. In our study, we used lentiviral genetic barcode labeling to monitor several thousand clones throughout teratoma development with specific emphasis on hematopoiesis.
In summary, this study provides detailed analysis of hematopoietic teratoma cell localization and clonal development as well as a feasible protocol to generate elevated hematopoiesis levels.
Results
Low Rate of Hematopoiesis in Subcutaneous Teratomas in NSG Mice
To test the level of spontaneous hematopoiesis in teratomas, we injected 1–5 × 106 CD34-derived hiPSCs (CD34hiPSC16) subcutaneously into the flanks of NSG mice. Teratomas were isolated once they reached a diameter of about 1.5 cm, which took 45 ± 8 days (mean ± SD). Quantification of hematopoietic cells was accomplished by flow cytometry (FC). To exclude false-positive results due to antibody cross-reactions with damaged cells, we applied a gating strategy that relied upon viability staining and isotype controls (Figure S1A). Overall, teratomas (n = 3) displayed a low spontaneous rate of hematopoietic differentiation (Figure 1A). IHC staining of teratoma sections for the specific markers confirmed low abundance of CD45+ cells (Figure 1B). Of note, CD34 expression occurred in cell clusters whereas the rare CD45+ hematopoietic cells were found distributed throughout the tissue (Figure 1B).
Figure 1.

Hematopoiesis in hiPSC-Derived Teratomas Is Improved by Steady Cytokine Supply in NSGS Mice and by Co-injection of HUVECs
(A) Exemplary flow cytometry (FC) detecting hematopoietic marker CD45 and progenitor marker CD34 in a teratoma generated in NSG mice.
(B) Immunohistochemistry on teratomas generated in NSG mice.
(C) FC summary of hematopoietic populations in teratomas generated with or without hematopoietic supporter cells. Median, quartiles, and outer values are depicted.
(D) Summary of FC analyses of teratoma hematopoietic populations generated in NSGS mice that express IL-3, GM-CSF, and SCF (median, quartiles, and outer values).
(E) Summary of FC results of teratoma samples generated with co-injection of OP9 or HUVECs expressing DLL1, DLL4, or WNT3A (mean and SD).
(F) Fold change of all CD45+ cells in teratomas generated with hiPSC and different supporter cell types in NSG or NSGS mice. The median CD45+ population of teratomas generated in NSG with hiPSC + OP9 was set to a value of 1. Graph depicts median and range.
Statistics of (C), (D), and (F) were calculated by Kruskal-Wallis and Dunn's multiple comparisons tests (∗p < 0.05).
Co-injection of HUVECs Supports Hematopoiesis in Teratomas
Due to the low rate of CD45+ cells in NSG mice and to better mimic a suitable environment for hematopoiesis in the teratoma, we co-injected either human umbilical vein endothelial cells (HUVECs) or murine stromal cell line OP9 together with hiPSCs. The latter are known to elevate hematopoietic output in teratomas (Amabile et al., 2013, Chen et al., 2015, Suzuki et al., 2013). HUVECs were selected because of their instructive signals for the development of hematopoietic cells from vascular endothelium in vitro (Gori et al., 2015, Lis et al., 2017). We applied 5 × 106 hiPSCs and 1 × 106 of the respective supporter cell type for these co-injection experiments. Co-transplantation of OP9 (n = 4) generated a human CD34−/CD45+ population with a median below 0.5% (Figure 1C). Co-injection of HUVECs (n = 6) resulted in a hematopoietic population with a median of 0.7%. In contrast to former teratoma studies (Amabile et al., 2013, Suzuki et al., 2013), our FC analyses detected no migration of hiPSC-derived CD34+ or CD45+ hematopoietic cells in peripheral blood (PB) or bone marrow (BM) of teratoma-bearing mice.
Elevated Levels of Hematopoietic Cells by Systemic Administration of Human Cytokines
Since co-injection experiments did not efficiently increase hematopoiesis, we explored whether cytokine application can be supportive in this setting. Human interleukin-3 (IL-3), granulocyte macrophage-colony stimulating factor (GM-CSF), and SCF foster hematopoietic progenitor maintenance and expansion (Lemoli and Gulati, 1993). Instead of additive cytokine administration we employed NSGS mice, which systemically express all three cytokines in the NSG mouse background. Teratomas induced with hiPSCs alone (n = 6) in NSGS showed a median of 1.2% hematopoietic cells (CD34−/CD45+) (Figure 1D). These results were reproducible for a human newborn foreskin fibroblast-derived (Nuff-hiPSC) hiPSC clone. The hematopoietic population comprised a median of 0.9% CD34−/CD45+ cells (n = 2), comparable with teratomas generated with hiPSCs (Figure 1D). Encouraged by these results, we tested whether the contribution of hematopoietic cells could be increased by co-injection of OP9 (n = 4) or HUVECs (n = 5). Indeed, we saw synergistic effects of the supporter cells and cytokine background (Figure 1D). HUVEC co-transplanted teratomas contained a significantly larger hematopoietic population (CD34−/CD45+) with a median of 6.1% than teratomas generated in NSGS mice with hiPSCs alone (1.2%). Although the level of hematopoietic cells was elevated, no evidence for migration of hiPSC-derived hematopoietic cells was detected in these experiments by FC (PB and BM).
Notch Ligands and WNT3A Overexpressed by HUVECs Do Not Further Promote Hematopoiesis
Although the exact mode of action through which Notch and Wnt3a signaling affect hematopoiesis remain to be fully elucidated, studies do support the importance of both pathways for hematopoiesis (Bigas and Espinosa, 2012, Lento et al., 2013). Most importantly, Dll1 or Wnt3a expressed by OP9 were previously used to improve hematopoietic output in teratoma assays (Amabile et al., 2013, Suzuki et al., 2013). To compare the influence of these signaling pathways on hematopoiesis in teratomas, we designed lentiviral vectors to constitutively overexpress WNT3A, DLL1, or DLL4 in HUVECs and OP9 (Figures S1B–S1D). DLL4 was included because it is known to be crucial for arterial specification, representing the starting point for definitive hematopoiesis (Duarte et al., 2004, Park et al., 2018). In fact, teratomas generated with co-transplanted OP9 expressing DLL1 (n = 3), DLL4 (n = 4), or WNT3A (n = 4) yielded higher CD45+ populations than with OP9 alone (Figure 1E). On the other hand, teratomas with co-injected HUVEC-WNT3A resulted in comparable CD45+ populations than with HUVECs alone. Interestingly, HUVEC-DLL1 or -DLL4 did not mediate any hematopoietic support (Figure 1E). In summary, teratomas generated in NSGS mice with co-injection of unmodified HUVECs or OP9-DLL1 resulted in an 80-fold and 86-fold (medians) greater CD45+ population compared with teratomas generated in NSG mice with co-injected OP9 (Figure 1F).
HUVEC Populations Decline during Teratoma Formation
As HUVECs appeared to promote the hematopoietic population in teratomas, we investigated how long HUVECs are present during teratoma formation. Therefore, we generated HUVECs that expressed firefly luciferase (HUVEC-Fluc) and modified hiPSCs to express the fluorescent protein Katushka2S to monitor teratoma growth (Luker et al., 2015) (Figures S1E and S1F). Within 7 weeks the bioluminescent signal of the co-injected 1 × 106 HUVEC-Fluc decreases to 2.5% of the signal obtained at the day of injection (Figures 2A and 2B). This suggests that the hemogenic support by HUVECs mainly occurred during the first 2–4 weeks.
Figure 2.

Hematopoiesis in hiPSC-Derived Teratomas Is Improved by Steady Cytokine Supply in NSGS Mice and by Co-injection of HUVECs
(A) Bioluminescent signal of HUVEC-firefly luciferase (Fluc) and red fluorescent signal of hiPSC-Katushka2S (Kat) during teratoma growth in an exemplary NSGS mouse.
(B) Summary of longitudinal study showing bioluminescent signal of HUVEC-Fluc on left y axis and fluorescence signal of hiPSC-Kat on right y axis. Signal intensities were analyzed in regions of interest covering the teratomas at their largest size. Graph displays mean and SD (until day 31 n = 10, day 47 n = 8).
(C) Summary of clonogenic assays for myeloid and erythroid lineages. CD34+/CD45+ cells were isolated from teratomas (n = 6), generated with hiPSC and HUVECs in NSGS mice. CD34+ cord blood cells were used as control (n = 3). Graph shows mean and SD.
(D) Pappenheim stain of isolated colonies types described in (C).
(E) Pappenheim staining of teratoma-derived CD45+ cells. G, granulocyte; M, macrophage; Mo, monocyte; N, normoblast.
We further evaluated lineage capacity of isolated CD34+/CD45+ teratoma cells generated in this setting. Clonogenic assays (n = 6) demonstrated the potential of these cells to produce granulocytes, macrophages, and few immature erythrocytes, which was comparable with freshly isolated CD34+ cord blood cells (n = 3) (Figures 2C and 2D). In Pappenheim staining, cells showed the expected phenotypes of band neutrophils, eosinophils, monocytes, and normoblasts. Sorted CD45+ teratoma populations appeared to consist mainly of monocytes and neutrophils (Figure 2E).
Localization of Hematopoietic Cells inside Vascular Structures
To improve the use of teratoma-linked hematopoiesis as a developmental model, we investigated the points of origin and localization of human hematopoietic cells inside the teratomas. We applied IHC staining on teratoma sections to localize hematopoietic sites in our human setting. All antibodies were validated on appropriate positive and negative controls to ensure specificity for human antigens in this xenograft model. The majority of committed hematopoietic cells (CD34−/CD43+/CD45+) appeared in groups of single cells. Most hematopoietic cells were located close to vascular structures positive for human CD34 (Figures 3A and 3B). Potential events of EHT were implied by co-localization of hematopoietic progenitor cells (CD34+/CD43+/CD45+) surrounded by CD34+ endothelium. In some sections, we identified CD34+ blood vessels with erythrocytes and hCD43+ cells (Figure 3C). To validate the endothelial nature of the CD34+ cells, we also stained sections for the endothelial marker CD31 (Figure 3D). To obtain a more detailed view of the hemogenic vasculature, we recorded z stacks (∼100 μm) of stained teratoma slices and calculated 3D models. This approach visualized hematopoietic cell clusters embedded inside and around CD34+ vessels (Figure 3E). Further characterization of the cellular environment of hematopoietic sites showed vascular structures (CD31+) and hematopoietic cells (CD45+) prominently in uniform tissue. Cells appeared to be spindle-shaped or round with scant cytoplasm, which is characteristic for mesenchymal tissue (Figures 3B and 3C, +BF; Figures S2A–S2C). Furthermore, these areas were positive for the mesenchymal intermediate filament vimentin (Figure S2D).
Figure 3.

Human Hematopoietic Cells Localize inside and in Proximity to Vascular Structures of Teratoma
Fluorescent IHC analysis of teratomas generated with hiPSCs in NSGS mice.
(A and B) CD45+/CD43+ cells embedded in CD34+/CD43−/CD45− vascular tissue (A); CD43+ cells detected inside or in proximity to CD34+ cell clusters (B).
(C) CD34+ vascular structure contained erythrocytes (white arrows) and human hematopoietic cells (CD43+).
(D) Vascularization visualized by staining of CD31 (PECAM-1) and CD34.
(E) z-stack image of a 150-μm stained teratoma slice recorded with a confocal microscope. 3D model was calculated with Imaris software (Bitplane).
Clonality of Teratoma Formation and Hematopoiesis
Currently it remains to be determined whether teratoma development is subject to clonal pressure or how many hiPSCs of the injected culture contribute to teratoma formation and initiate hematopoiesis. In theory, a teratoma can be generated by a single pluripotent cell. Despite the improvement of hematopoiesis rate, the proportion of hematopoietic progenitors in teratomas remained low in the experiments described above. Therefore, we applied RGB (red, green, blue) labeling combined with nucleotide barcoding (Cornils et al., 2014, Selich et al., 2016) to investigate the frequency of iPSCs, which actually undergo hematopoietic differentiation. This method allowed us to trace clones via genetic labeling. Additional expression of mCherry, Venus, or Cerulean (RGB labeling) also facilitated visual assessment of clonal development (Figure 4A). Transduced hiPSCs were expanded for 21 days (14.6-fold expansion). Additionally, transgene expression was monitored by fluorescence microscopy and FC during the expansion phase and in isolated teratomas, indicating no major transgene silencing in vitro or in vivo (Figure 4B). Samples were harvested and analyzed after transduction, after in vitro expansion, and finally after teratoma formation. The teratoma samples (n = 5) were divided into two sections. One was directly used for sequencing and the second was used to isolate the hematopoietic population by magnetic hCD45 beads followed by sequencing (Figure 4C). Deep sequencing data identified a highly polyclonal composition in hiPSC culture, teratomas, and hematopoietic cells (Figure 4D). Read counts of identical sequences were used as a surrogate marker for clonal contribution of individual barcodes. We observed that only 146 out of 28,300 measured barcodes reoccurred in all CD45+ hematopoietic samples, and 139 out of 21,698 barcodes were detected in all teratoma samples. Hence, thousands of injected cells are capable of forming teratomas and hematopoietic cells (Figures 4E and 4F). In summary, our results suggest that expanded hiPSCs contained a homogeneous pool of cell clones potentially able to form teratomas. Furthermore, several thousand initial hiPSCs revealed the capacity for hematopoietic differentiation.
Figure 4.

Genetic Barcoding Determined Teratoma Formation and Subsequent Hematopoiesis as Highly Polyclonal Events
(A) Design of the lentiviral construct used to transduce hiPSC prior to teratoma formation. Fluorescent reporters Venus, Cerulean, or mCherry are expressed under an EFS promoter.
(B) Micrographs of fluorescent reporter expression during expansion of hiPSC prior to teratoma induction and after teratoma isolation (38 days).
(C) Experimental scheme to access clonality of teratomas and teratoma-derived hematopoiesis.
(D) Barcode variety in hiPSC cultures and teratoma samples.
(E) Venn diagram depicting barcodes detected in CD45+ samples isolated from teratoma.
(F) Venn diagram depicting barcodes detected in teratoma samples.
Discussion
The teratoma model provides human embryonic tissue to study early hematopoiesis in a physiological 3D environment. However, the CD45+ population in teratomas generated with hiPSCs in NSG mice is low and, thus, hampers its application as a hematopoiesis model. In this study, we elaborated a feasible approach to enhance teratoma-linked hematopoiesis by using NSGS mice. The supply of cytokines by the mouse model is time-saving, easy to perform, and cost-effective. Further enhancement was achieved upon co-injection with supporter cells. HUVECs can promote EHT through vascular induction or support expansion of hematopoietic progenitors from cord blood by secretion of cytokines and growth factors in vitro (Lis et al., 2017, Yamaguchi et al., 1996, Yildirim et al., 2005). In the teratoma setting, the presence of HUVECs led to a significant increase in the hematopoietic population in teratomas, although the majority of HUVECs was not maintained beyond 30 days of teratoma formation. Similar levels of hematopoietic cells were obtained with OP9-DLL1, OP9-DLL4, or OP9-WNT3A, but not with OP9 alone. Intriguingly, while co-injection of HUVEC-WNT3A did not further elevate hematopoietic yield compared with HUVECs alone, co-injection of HUVEC-DLL1 or -DLL4 suppressed intra-teratoma hematopoiesis. It remains to be determined whether this was caused by changes in the transcriptome of HUVECs potentially provoked by lentiviral transduction or whether the overexpression of the particular transgenes is not compatible with the support of hematopoiesis by HUVECs.
In summary, the elaborated teratoma formation protocol with HUVECs in NSGS mice provides a decent hematopoietic population and represents a starting point from which to investigate human hematopoiesis with a low technical demand.
The explicit polyclonality of teratoma formation and subsequent hematopoiesis makes this system interesting for additional applications. Future studies could use genetic barcode labeling to mark differentially cultured or transduced hiPSC clones to study their differentiation preferences. Microdissections would allow investigation of potential germ-layer preferences exhibited by different hiPSC clones. In particular, upscaling differentiation protocols could be optimized by selecting the most suitable clone for the specific target lineage. Furthermore, effects of TFs from libraries on teratoma-derived hematopoiesis could be examined closely and might promote valuable insights into developmental states even beyond the hematopoietic system.
Another important asset of the teratoma model is the possibility to localize target cells in tissues assembled in natural structures. In this study, human hematopoietic cell localization was carefully examined by specific detection of surface markers. In rare cases, hematopoietic progenitors were located inside vessels. These results indicate that hiPSC-derived hematopoiesis in teratoma reflects the natural EHT process, which has recently been described for murine teratomas (Tsukada et al., 2017). Further examination of the cellular environment revealed that the majority of hematopoietic and vascular endothelial cells were located in mesenchymal tissue, as identified by cell phenotype and vimentin staining. Whether the hematopoietic cells actively migrate from the vessels into the mesenchyme, or the hemogenic endothelium fully differentiates into CD34−/CD45+ cells remains unclear. Based on our results, we hypothesize that human hematopoietic cells originate from EHT. The endothelium, in turn, derives from the mesenchymal-to-epithelial transition. Both processes are closely related to normal embryogenesis and affirm the use of the teratoma model to study embryonic processes in healthy and diseased backgrounds.
Experimental Procedures
Mice
All animal experiments were performed in accordance with Lower Saxony State Office for Consumer Protection and Food Safety in Germany. Further details can be found in Supplemental Experimental Procedures.
Teratoma Induction
For injection into mice, hiPSC were harvested using trypsin/EDTA. Unless stated otherwise, cell numbers were adjusted to 3 × 106 hiPSCs per injection. In co-injection experiments, 1 × 106 supporter cells were mixed with hiPSCs prior to injection. Harvested cells were centrifuged at 300 × g and gently resuspended in hiPSC medium containing 20 μM Y-27632 (kindly provided by Leibniz University, Hannover). Cell suspensions were cooled to 4°C prior to addition of 100 μL of Matrigel basement membrane matrix (Corning) (Prokhorova et al., 2009). The injection solution was kept at 4°C until subcutaneous injection into the flank. We induced two teratomas per mouse. The injection volume was 150–200 μL. Mice were sacrificed as soon as one teratoma reached 1.5 cm in diameter. After isolation, teratoma tissue was randomly divided into two parts. One was used for IHC and the other for FC.
Single-Cell Preparation from Teratoma Tissue
Teratomas were sliced into 3- to 5-mm pieces and incubated in DMEM containing 160 U/mL dispase I (Roche) and 2 U/mL collagenase IV (Thermo Fisher Scientific) for 40–60 min at 37°C. Thereafter, suspensions were processed through 70-μm cell strainers (Fisher Scientific). After centrifugation (300 × g, 10 min, 4°C), cells were resuspended and incubated with 15–20 μg of DNAse I (STEMCELL Technologies) and kept at room temperature for 15 min. Afterward, cells were washed and used for further experiments.
Statistics
Kruskal-Wallis and Dunn's multiple comparison tests were performed with GraphPad Prism 7 software. Asterisks indicate a p value of less than 0.05.
Further methods are detailed in Supplemental Experimental Procedures.
Author Contributions
F.P. designed and performed experiments and wrote the manuscript. A. Selich provided barcode technology and analysis. D.H. and D.K. cloned lentiviral constructs. M.A.M. generated Nuff-hiPSC clone. A. Schambach designed experiments and contributed to lentiviral vectors. S.G. and S.L. supported teratoma experiments. M.R., D.H., V.N., K.S., S.R., M.A.M., A.B., and A. Schambach provided conceptual advice, edited the manuscript, and discussed results.
Acknowledgments
We thank Gina Geide, Margarethe Schleiss, and Karin Serwatzki for assistance in IHC, and Matthias Meyer and Larissa Buch for support in teratoma experiments. We also thank Mania Ackermann for providing CD34iPSC16 and Bettina Weigel for CB isolation. This work was supported by grants from DFG (REBIRTH Cluster of Excellence and SFB738).
Published: October 18, 2018
Footnotes
Supplemental Information includes Supplemental Experimental Procedures and two figures and can be found with this article online at https://doi.org/10.1016/j.stemcr.2018.09.010.
Supplemental Information
References
- Amabile G., Welner R.S., Nombela-Arrieta C., D’Alise A.M., Di Ruscio A., Ebralidze A.K., Kraytsberg Y., Ye M., Kocher O., Neuberg D.S. In vivo generation of transplantable human hematopoietic cells from induced pluripotent stem cells. Blood. 2013;121:1255–1264. doi: 10.1182/blood-2012-06-434407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bertrand J.Y., Chi N.C., Santoso B., Teng S., Stainier D.Y.R., Traver D. Haematopoietic stem cells derive directly from aortic endothelium during development. Nature. 2010;464:108–111. doi: 10.1038/nature08738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bigas A., Espinosa L. Hematopoietic stem cells: to be or Notch to be. Blood. 2012;119:3226–3235. doi: 10.1182/blood-2011-10-355826. [DOI] [PubMed] [Google Scholar]
- Blum B., Benvenisty N. Clonal analysis of human embryonic stem cell differentiation into teratomas. Stem Cells. 2007;25:1924–1930. doi: 10.1634/stemcells.2007-0073. [DOI] [PubMed] [Google Scholar]
- Chen X., Zhao Q., Li C., Geng Y., Huang K., Zhang J., Wang X., Yang J., Wang T., Xia C. OP9-Lhx2 stromal cells facilitate derivation of hematopoietic progenitors both in vitro and in vivo. Stem Cell Res. 2015;15:395–402. doi: 10.1016/j.scr.2015.08.009. [DOI] [PubMed] [Google Scholar]
- Cornils K., Thielecke L., Hüser S., Forgber M., Thomaschewski M., Kleist N., Hussein K., Riecken K., Volz T., Gerdes S. Multiplexing clonality: combining RGB marking and genetic barcoding. Nucleic Acids Res. 2014;42:e56. doi: 10.1093/nar/gku081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Damjanov I., Andrews P.W. Teratomas produced from human pluripotent stem cells xenografted into immunodeficient mice—a histopathology atlas. Int. J. Dev. Biol. 2016;60:337–419. doi: 10.1387/ijdb.160274id. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duarte A., Hirashima M., Benedito R., Trindade A., Diniz P., Bekman E., Costa L., Henrique D., Rossant J. Dosage-sensitive requirement for mouse Dll4 in artery development. Genes Dev. 2004;18:2474–2478. doi: 10.1101/gad.1239004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gertow K., Wolbank S., Rozell B., Sugars R., Andäng M., Parish C.L., Imreh M.P., Wendel M., Ährlund-Richter L. Organized development from human embryonic stem cells after injection into immunodeficient mice. Stem Cells Dev. 2004;13:421–435. doi: 10.1089/scd.2004.13.421. [DOI] [PubMed] [Google Scholar]
- Godin I., Cumano A. The hare and the tortoise: an embryonic haematopoietic race. Nat. Rev. Immunol. 2002;2:593–604. doi: 10.1038/nri857. [DOI] [PubMed] [Google Scholar]
- Gori J.L., Butler J.M., Chan Y., Chandrasekaran D., Poulos M.G., Ginsberg M., Nolan D.J., Elemento O., Wood B.L., Adair J.E. Vascular niche promotes hematopoietic multipotent progenitor formation from pluripotent stem cells. J. Clin. Invest. 2015;125:1–12. doi: 10.1172/JCI79328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kissa K., Herbomel P. Blood stem cells emerge from aortic endothelium by a novel type of cell transition. Nature. 2010;464:112–115. doi: 10.1038/nature08761. [DOI] [PubMed] [Google Scholar]
- Lemoli R.M., Gulati S.C. Effect of stem cell factor (c-kit ligand), granulocyte-macrophage colony stimulating factor and interleukin 3 on hematopoietic progenitors in human long-term bone marrow cultures. Stem Cells. 1993;11:435–444. doi: 10.1002/stem.5530110511. [DOI] [PubMed] [Google Scholar]
- Lento W., Congdon K., Voermans C., Kritzik M., Reya T. Wnt signaling in normal and malignant hematopoiesis. Cold Spring Harb. Perspect. Biol. 2013;5:a008011. doi: 10.1101/cshperspect.a008011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lis R., Karrasch C.C., Poulos M.G., Kunar B., Redmond D., Duran J.G.B., Badwe C.R., Schachterle W., Ginsberg M., Xiang J. Conversion of adult endothelium to immunocompetent haematopoietic stem cells. Nature. 2017;545:439–445. doi: 10.1038/nature22326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luker K.E., Pata P., Shemiakina I.I., Pereverzeva A., Stacer A.C., Shcherbo D.S., Pletnev V.Z., Skolnaja M., Lukyanov K.A., Luker G.D. Comparative study reveals better far-red fluorescent protein for whole body imaging. Sci. Rep. 2015;5:10332. doi: 10.1038/srep10332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mikkola H.K.A. The journey of developing hematopoietic stem cells. Development. 2006;133:3733–3744. doi: 10.1242/dev.02568. [DOI] [PubMed] [Google Scholar]
- Morgan R.A., Gray D., Lomova A., Kohn D.B. Hematopoietic stem cell gene therapy: progress and lessons learned. Cell Stem Cell. 2017;21:574–590. doi: 10.1016/j.stem.2017.10.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park M.A., Kumar A., Jung H.S., Uenishi G., Moskvin O.V., Thomson J.A., Slukvin I.I. Activation of the arterial program drives development of definitive hemogenic endothelium with lymphoid potential article activation of the arterial program drives development of definitive hemogenic endothelium with lymphoid potential. Cell Rep. 2018;23:2467–2481. doi: 10.1016/j.celrep.2018.04.092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prokhorova T.A., Harkness L.M., Frandsen U., Ditzel N., Schrøder H.D., Burns J.S., Kassem M. Teratoma formation by human embryonic stem cells is site dependent and enhanced by the presence of Matrigel. Stem Cells Dev. 2009;18:47–54. doi: 10.1089/scd.2007.0266. [DOI] [PubMed] [Google Scholar]
- Selich A., Daudert J., Hass R., Philipp F., von Kaisenberg C., Paul G., Cornils K., Fehse B., Rittinghausen S., Schamnbach A. Massive clonal selection and transiently contributing clones during expansion of mesenchymal stem cell cultures revealed by Lentiviral RGB-Barcode technology. Stem Cells Transl. Med. 2016;5:591–601. doi: 10.5966/sctm.2015-0176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugimura R., Jha D.K., Han A., Soria-Valles C., da Rocha E.L., Lu Y.-F., Goettel J.A., Serrao E., Rowe R.G., Malleshaiah M. Haematopoietic stem and progenitor cells from human pluripotent stem cells. Nature. 2017;545:432–438. doi: 10.1038/nature22370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki N., Yamazaki S., Yamaguchi T., Okabe M., Masaki H., Takaki S., Otsu M., Nakauchi H. Generation of engraftable hematopoietic stem cells from induced pluripotent stem cells by way of teratoma formation. Mol. Ther. 2013;21:1424–1431. doi: 10.1038/mt.2013.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsukada M., Ota Y., Wilkinson A.C., Becker H.J., Osato M., Nakauchi H., Yamazaki S. In vivo generation of engraftable murine hematopoietic stem cells by Gfi1b, c-Fos, and Gata2 overexpression within teratoma. Stem Cell Reports. 2017;9:1024–1033. doi: 10.1016/j.stemcr.2017.08.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vo L.T., Daley G.Q. De novo generation of HSCs from somatic and pluripotent stem cell sources. Blood. 2015;125:2641–2648. doi: 10.1182/blood-2014-10-570234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi H., Ishii E., Saito S., Tashiro K., Fujita I., Yoshidomi S., Ohtubo M., Akazawa K., Miyazaki S. Umbilical vein endothelial cells are an important source of c-kit and stem cell factor which regulate the proliferation of haemopoietic progenitor cells. Br. J. Haematol. 1996;94:606–611. doi: 10.1046/j.1365-2141.1996.d01-1855.x. [DOI] [PubMed] [Google Scholar]
- Yildirim S., Boehmler A.M., Kanz L., Möhle R. Expansion of cord blood CD34+ hematopoietic progenitor cells in coculture with autologous umbilical vein endothelial cells (HUVEC) is superior to cytokine-supplemented liquid culture. Bone Marrow Transpl. 2005;36:71–79. doi: 10.1038/sj.bmt.1705001. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.