

Abstract
Studies of climate effects on ecology often account for non-stationarity in individual physical and biological variables, but rarely allow for non-stationary relationships among variables. Here, we show that non-stationary relationships among physical and biological variables are central to understanding climate effects on salmon (Onchorynchus spp.) in the Gulf of Alaska during 1965–2012. The relative importance of two leading patterns in North Pacific climate, the Pacific Decadal Oscillation (PDO) and North Pacific Gyre Oscillation (NPGO), changed around 1988/1989 as reflected by changing correlations with leading axes of sea surface temperature variability. Simultaneously, relationships between the PDO and Gulf of Alaska environmental variables weakened, and long-standing temperature–salmon and PDO–salmon covariance declined to zero. We propose a mechanistic explanation for changing climate–salmon relationships in terms of non-stationary atmosphere–ocean interactions coinciding with changing PDO–NPGO relative importance. We also show that regression models assuming stationary climate–salmon relationships are inappropriate over the multidecadal time scale we consider. Relaxing assumptions of stationary relationships markedly improved modelling of climate effects on salmon catches and productivity. Attempts to understand the implications of changing climate patterns in other ecosystems might also be aided by the application of models that allow associations among environmental and biological variables to change over time.
Keywords: climate indices, non-stationary relationships, novel climate, North Pacific Gyre Oscillation, Pacific Decadal Oscillation, Pacific salmon
1. Introduction
Ecologists are well aware that physical and biological state variables often exhibit non-stationary mean and variance. However, the possibility of non-stationary relationships among variables is often ignored, and these relationships are typically modelled as stochastic processes with a fixed probability density [1]. There is a reason to expect that understanding of climate effects on ecology might be vulnerable to violations of this assumption of stationary relationships among physical variables. The sets of climate variables acting on ecosystems are typically highly collinear, which makes it difficult or impossible to attribute biological responses to the effect of individual variables [2]. But statistical models lack predictive skill when relationships among covariates change, and models that are parametrized in the presence of high collinearity tend to be particularly unreliable if collinear relationships subsequently weaken [2]. Because of these concerns, non-stationary relationships among different climate variables (‘novel’ climates) are an important issue at the longer time scales considered by paleoecology and by studies of potential future climate states [3]. As observational time series become longer and as anthropogenic forcing increases global rates of environmental change, non-stationary associations among climate variables will become increasingly likely and therefore should be taken into account in analyses [1].
Large-scale climate indices that serve as proxies for conditions across many collinear physical variables have shown great utility for understanding climate effects on both terrestrial and marine ecosystems [4]. The use of these climate indices also typically implies the assumption of stationary relationships among underlying physical variables. Indices are commonly defined by the leading eigenvectors of anomaly fields in primary variables such as sea level pressure (SLP) or sea surface temperature (SST). These definitions are then used to summarize subsequent climate observations, but the underlying definition remains based on a fixed covariance matrix [4,5]. However, climate science suggests at least two reasons that climate variability may violate implicit assumptions of stationary relationships in ecological studies using these indices. First, the amplitude and importance of different climate patterns may vary over time [6–8], and these changes may not be captured by climate indices that assume stationary relationships. Second, climate indices based on eigenvectors are by definition statistical patterns, which may arise from multiple physical processes. If the generating processes operate on independent time scales, the set of physical variables mapping onto the climate index will change over time [9]. Given expectations for changing associations among physical variables associated with climate indices and the potential that such changes have to create ‘ecological surprises' [3], improved awareness of non-stationary relationships among physical variables is an important step for improving our understanding of climate effects on ecology.
Here, we evaluate the implications of non-stationary relationships among physical variables for climate regulation of salmon populations (Onchorynchus spp.) in the Gulf of Alaska. The Pacific Decadal Oscillation (PDO) climate pattern was first recognized as it related to covariance between coastal temperature anomalies and catches of salmon [5], which are numerically and trophically dominant members of North Pacific ecosystems [10] and critical economic and cultural resources for many human communities. For much of the twentieth century, the positive coastal temperature anomalies that characterize a positive PDO state were associated with increased salmon catches in Alaska [5]. However, since the late 1980s, another pattern, the North Pacific Gyre Oscillation (NPGO), has apparently explained an increasing fraction of North Pacific climate variance compared to earlier decades [11–14]. During this time, weakening PDO relationships with some salmon populations and strengthening NPGO relationships with others have been observed, but the mechanisms underlying these changes are unknown [15,16]. A change in relative importance of the PDO and NPGO might imply change between distinct patterns of multivariate environmental variability in North Pacific ecosystems, as these indices capture variability in a broad set of physical variables, including SST, sea surface height (SSH), gyre-scale advection, coastal upwelling and downwelling, surface wind stress and surface salinity [5,17]. The implications of possible changes in the physical variables associated with these climate patterns or changes in their relative importance in the climate have received little attention, and ecologists continue to use the PDO and NPGO indices with the implicit assumption of stationary relationships with underlying physical variables (e.g. [18–20]). We use climate data and salmon catch and productivity data from the Gulf of Alaska to test the hypothesis that PDO-associated climate effects on salmon have weakened as the NPGO pattern has played a more important role in North Pacific climate. We also present a plausible mechanistic explanation for changing climate–salmon relationships in terms of non-stationary relationships among physical variables acting on the Gulf of Alaska ecosystem. Our specific goals are: (i) to evaluate the evidence for changing relative importance of the PDO and NPGO; (ii) to test for non-stationary relationships among physical variables associated with changes in the relative importance of the PDO and NPGO, both in links between large-scale climate indices and local (Gulf of Alaska) environment and in multivariate variability among local environmental variables; and (iii) to test for non-stationary relationships between physical variables and salmon catches and productivity.
2. Material and methods
(a). Data
Biological data included fisheries-dependent and fisheries-independent time series for the four salmon species that are widely targeted in Gulf of Alaska fisheries: pink (O. gorbuscha), sockeye (O. nerka), chum (O. keta) and coho (O. kisutch). Commercial catch data for these species from 1965 (when fisheries were largely developed) through ocean entry year 2012 (to avoid effects of recent heatwave anomalies [21]) were obtained from the Alaska Department of Fish and Game. Catch data were log-transformed and lagged one year for pink and coho salmon, two years for sockeye salmon and three years for chum salmon, to correspond with average ages for ocean entry in the Gulf of Alaska [22]. Salmon productivity was estimated with fisheries-independent spawner-recruit time series obtained from previous studies [23,24] (n = 20 pink salmon runs, 9 sockeye salmon runs, 10 chum salmon runs; electronic supplementary material, table S1). These data relate spawner abundance in each year to the number of recruits produced from that brood year (i.e. offspring surviving to adulthood). Suitable spawner-recruit time series were not available for coho salmon.
Large-scale climate data included the published PDO and NPGO indices [5,17], and SST and SLP fields for the North Pacific (electronic supplementary material, table S2). Relationships between salmon productivity and coastal temperature anomalies associated with the PDO are not thought to reflect direct temperature effects, as observed temperature ranges have been small relative to salmon physiological tolerances, but instead are thought to arise because SST is correlated with environmental variables directly affecting salmon early marine survival [25]. Our climate data included six proxies for processes proposed in the literature as mechanisms that might explain correlations between SST and early marine survival in the Gulf of Alaska [25–30]: (i) the mean of three coastal downwelling indices as proxies for summer upwelling/downwelling and onshore Ekman transport; (ii) winter values of a SLP gradient that is a proxy for coastal precipitation; (iii) an independent estimate of coastal freshwater discharge; (iv) 20 m salinity observations at the GAK1 site; (v) the first principal component (PC1) of Gulf of Alaska wind stress anomalies (electronic supplementary material, figure S1); and (vi) the Papa advection index, a measure of winter gyre-scale advection at Ocean Station Papa in the Gulf of Alaska (details in electronic supplementary material, table S2).
(b). Climate analysis
The PDO and NPGO are defined as the first and second eigenvectors, respectively, of anomalies in SST or SSH for the North Pacific basin [5,17]. These definitions are based on fixed reference periods (1900–1993 for the PDO and 1950–2004 for the NPGO). The first eigenvector (nominally the PDO) explains the most variance in the data, while the second eigenvector (nominally the NPGO) explains the second-most variance. However, since the late 1980s, the NPGO pattern may have played a more important role in North Pacific climate in terms of explaining more variance in climate properties and showing increased association with the first eigenvector of SST anomalies (SSTa) [12,31]. We therefore define the relative importance of the two climate patterns in terms of the strength of their correlations with the first or second eigenvectors of North Pacific SSTa fields as calculated over a specific period of time. We tested for changes in the relative importance of the PDO and NPGO by comparing their correlations with PC1–2 for SSTa fields during 1950–1988 and 1989–2012 (see electronic supplementary material for details). The 1988/1989 division is based on the proposed timing of increasing relative importance of the NPGO [12]. All correlation tests used the modified Chelton method to account for autocorrelation [32]. We also calculated correlations between the PDO and NPGO indices and PC1-2 of North Pacific SSTa variability over 21-yr rolling windows to evaluate the timing of the change in relative importance of the PDO and NPGO.
To understand non-stationary relationships among Gulf of Alaska environmental variables, we focused on the Aleutian Low, an area of low values in the monthly SLP climatology of the North Pacific that is a leading atmospheric driver of the PDO pattern [9]. The six local environmental variables thought to affect salmon survival, listed above, are all related to cyclonic wind stress arising from the Aleutian Low [29,33]. We began evaluating possible changes in the relationship between the Aleutian Low and the Gulf of Alaska ecosystem by regressing fields of North Pacific SLPa and wind stress onto the PDO index in two data blocks (1950–1988 and 1989–2012). We tested for changes in the relationship between the Aleutian Low and environmental processes in the Gulf of Alaska by first testing for changing temporal variance in the SLPa fields associated with the Aleutian Low during periods of differing relative importance of the PDO and NPGO (1950–1988 and 1989–2012). We then tested for changing correlations between mean SLPa values in the Aleutian Low area and individual environmental variables in the Gulf of Alaska (details in electronic supplementary material). Finally, we tested for changing patterns of multivariate variability among Gulf of Alaska environmental variables by comparing the proportion of variance explained by, and loadings on, PC1 for the six environmental time series calculated separately for 1950–1988 and 1989–2012.
(c). Climate–salmon analysis
We first tested the hypothesis of non-stationary climate–salmon relationships by comparing linear regression models of stationary and non-stationary relationships between winter averages of climate variables (SST, PDO and NPGO) and PC1 of commercial salmon catches. Catch PC1 explained 85% of total variance and loaded the four species nearly equally (loadings = 0.49–0.52). Explanatory variables were smoothed with a 3-year running mean to capture conditions the year before, year of and year after ocean entry. Stationary models invoked a single regression coefficient describing climate–salmon relationships across 1965–2012, while non-stationary models allowed coefficients to differ between eras (1965–1988 versus 1989–2012) through climate × era interaction effects as well as main climate and era effects. Model parsimony and fits to data were evaluated with the Akaike information criterion adjusted for small sample size (AICc) [34]. We used first-order autocorrelation in model residuals to evaluate the assumption of independent residuals.
We confirmed that the change in the climate–catch relationship coincided with the proposed late 1980s change in the relative importance of the PDO and NPGO using a model selection approach. Specifically, regression models with SST × era interaction effects as well as SST and era main effects were fitted to the PC1 of catch, with separate models invoking every possible year for a change in the relationship from 1974 to 2002. The support for different possible change points was evaluated with AICc scores. We evaluated the statistical significance of the best-supported timing of non-stationary climate–catch relationships using generalized least-squares regression formulated to allow autocorrelated residuals [35].
Catch data are subject to confounding factors that might produce spurious conclusions. An example is Gulf of Alaska hatchery programmes, which expanded rapidly in the late 1980s and have the stated goal of buffering salmon fisheries against environmental variability. We tested for a hatchery effect by comparing the evidence for non-stationary SST–catch relationships in wild and hatchery-subsidized fisheries. These designations were based on contributions of less than or equal to 9% and greater than or equal to 17%, respectively, of hatchery fish in commercial catches within different management areas (electronic supplementary material, table S3). Non-stationary responses to SST were tested with mixed-effects models [36] fit separately to wild and hatchery-subsidized fisheries. These models included fixed intercepts and SST, era and SST × era fixed effects, in addition to random effects allowing individual species-area combinations to deviate from the fixed intercept (overall productivity) and the fixed SST effect. All mixed-effects models were formulated with autocorrelated residuals [35] (details in electronic supplementary materials).
As a second test for non-stationary climate–salmon relationships, we used mixed-effects models to test for changing SST effects on productivity (recruits per spawner, accounting for density dependence) in wild runs of pink, sockeye and chum salmon. We first fit models with the full set of fixed effects and selected the random effects structure based on AICc scores from model results generated with restricted maximum likelihood. The hypothesis of non-stationary SST effects was then evaluated with likelihood ratio tests (based on maximum-likelihood scores) comparing models fit with and without the SST×era interaction term [36], with era referring to ocean entry years 1960–1988 versus 1989–2012. Selected models were assessed for violations of model assumptions, especially normal distributions of random effect residuals. Because density-dependent effects were not normally distributed among runs, these effects were modelled as run-specific fixed effects rather than as random effects [25]. This analysis used local SST values as covariates, reflecting the scale of previously observed SST effects on spawner-recruit dynamics [25,37] (details in electronic supplementary material). Pink salmon spawner-recruit data were analysed separately for genetically isolated populations spawning in the same river in alternate (even and odd) years.
Finally, an extensive literature has attempted to identify the causal mechanisms that produce PDO–salmon and SST–salmon correlations. This literature almost universally assumes that the underlying mechanistic relationships are fixed in time [26–28,30,37]. To evaluate this assumption, we tested for non-stationary relationships between the six individual Gulf of Alaska environmental time series and spawner-recruit dynamics before and after 1988/1989. Although it typically would not be appropriate to interpret individual effects for collinear covariates [2], in this case we fit separate Ricker models for the effect of each environmental covariate to evaluate the evidence for continuity of effects across 1988/1989. The inflated type I error arising from multiple hypothesis tests was addressed using False Discovery Rate control adjustments to α [38]. All analysis was performed in the computer language R [39].
3. Results
(a). Changing relative importance of the Pacific Decadal Oscillation and North Pacific Gyre Oscillation
After 1988/1989, the classification of the PDO and NPGO as the first and second eigenvectors of North Pacific climate variability, respectively, broke down. During 1989–2012, the NPGO became significantly correlated with SSTa PC1 (|r| = 0.57, p = 8.4 × 10−4) and the PDO became significantly correlated with PC2 (|r| = 0.37, p = 6.9 × 10−4), supporting the previous observations that there were changes in the relative importance of the two patterns (complete results in electronic supplementary material, table S4). Absolute values of correlation coefficients were used because the sign of PC scores is arbitrary and can change between iterations of the EOF. More overall variance was explained by PC1 during the period of significant correlation with both the PDO and NPGO (1989–2012, 29.9%) than during 1950–1988 when this axis was only correlated with the PDO (23.6%). Variance explained by PC2 was similar between eras (13.1% in 1950–1988, 13.7% in 1989–2012). The rolling window analysis was consistent with the hypothesis of a sharp increase in relative importance of the NPGO pattern around 1988/1989 (figure 1a,b). Changing spatial loadings for PC1,2 are plotted in electronic supplementary material, figure S2.
Figure 1.

Non-stationary patterns in North Pacific climate. (a,b) Changing relative importance of the PDO and NPGO patterns. (a) Absolute value of correlation between PDO/NPGO indices and PC1 of North Pacific SSTa calculated on 21-yr rolling windows. (b) Correlation with PC2. Dashed vertical lines indicate 1988/1989. (c–f), Non-stationary atmospheric associations with the PDO. (c) Regression of SLPa fields onto PDO index, 1950–1988. (d) The same regression, 1989–2012. (e) Regression of wind stress fields onto PDO index,1950–1989. (f) Wind stress-PDO regression for 1989–2012. The polygon in (c–f) indicates the Gulf of Alaska.
(b). Non-stationary relationships among physical variables
Temporal differences in the SLPa–PDO regression suggest a change in the atmospheric variability mapping onto the PDO that is synchronous with change in the relative importance of the PDO and NPGO. For 1950–1988 data, the regression highlights the Aleutian Low (figure 1c). After 1988/1989, the area of SLPa most closely linked with the PDO index shifted southeast (figure 1d). Regressions of North Pacific wind stress onto the PDO index also changed after 1988/1989 (figure 1e,f). North-south wind stress–PDO relationships were significantly weaker (p ≤ 0.05) over most of the Gulf of Alaska after 1988/1989 (electronic supplementary material, figure S3). This result is the first indication of non-stationarity in relationships among the physical variables related to Gulf of Alaska ecological variability: identical values of the PDO index reflect significantly different levels of wind stress during the periods of differing PDO–NPGO importance.
The late 1980s change in relationships between the PDO and large-scale atmospheric patterns coincided with declining temporal variability in the Aleutian Low. The standard deviation (SD) in SLPa values in the Aleutian Low area showed a strong drop after 1988/1989 (figure 2; see electronic supplementary material, figure S4 for the area used to calculate this index). Declining variance was coincident with a decline in the correlations between the Aleutian Low SLPa index and the six Gulf of Alaska environmental variables (figure 3a). Correlations between the SLPa index and every local environmental time series except downwelling strength were significant prior to 1988/1989 (|r| ≥ 0.65, p ≤ 8.0 × 10−4), demonstrating the role of Aleutian Low variability as an important driver of these ecologically important processes prior to 1988/1989. These relationships all weakened after 1988/1989, and only the SLPa–salinity correlation remained significant (r = 0.64, p = 0.002, figure 3a). The coherence of local environmental processes changed at the same time. Covariance among the six environmental time series was very strong prior to 1988/1989, with PC1 explaining fully 89% of total variance; after 1988/1989, this proportion fell to 40% (figure 3b). Changing loadings for PC1 further underscore the non-stationary nature of local environmental variability in the two periods (figure 3c). Processes related to freshwater dynamics (the SLP gradient, estimated freshwater discharge and observed 20-m salinity) retain similar loadings on PC1 in the two periods. The other variables either showed weaker loadings in the later period (advection, wind stress) or opposite signs for loadings in the two periods (downwelling; figure 3c). Thus, PC1 of local environmental variability during the two periods represents different sets of physical forcings on salmon survival. Additionally, winter values of the PDO and mean Gulf of Alaska SST served as accurate indices of PC1 of the six processes thought to affect salmon during 1950–1988 (r ≤ −0.75, p ≤ 0.002), but these relationships were non-significant during 1989–2012 (r ≥ −0.32, p ≥ 0.14; figure 3d). We interpret these results with a conceptual model of the Gulf of Alaska system operating during conditions of either high or low Aleutian Low variance. With high variance, positive (negative) PDO conditions co-occur with strong negative (positive) SLP anomalies associated with increased (decreased) cyclonic circulation driving highly collinear variability among individual environmental variables. With low variance, reduced contrast between positive and negative SLP anomalies results in reduced collinear variability among environmental variables and reduced association between the PDO/SST and environmental conditions.
Figure 2.
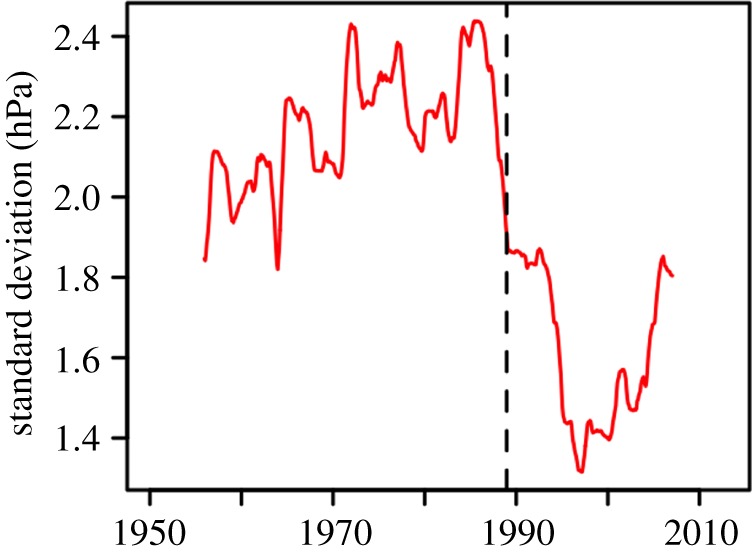
Changing variance in the Aleutian Low SLPa index in periods of differing PDO–NPGO relative importance. Standard deviations of smoothed monthly SLPa averaged over the Aleutian Low area (47°–57° N, 153°–169° W) are calculated over 11 yr rolling windows. The vertical dashed line indicates 1988/1989. (Online version in colour.)
Figure 3.

Non-stationary relationships among physical variables in the Gulf of Alaska during periods of high (1950–1988) and low (1989–2012) Aleutian Low variance. (a) Correlation between individual environmental variables and SLPa in the Aleutian Low area. (b) Variance explained by principal components of environmental variables during each period. PC1 explains more than twice as much variance before 1988/1989 (89%) than after (40%). (c) Loadings for PC1 for the two periods. (d) Correlations between PDO/Gulf of Alaska SST and PC1 of environmental variables by time period. Asterisks in (a,d) indicate significant correlations (p ≤ 0.05).
(c). Non-stationary climate–salmon relationships
We next tested the hypothesis that climate–catch relationships are non-stationary across these two periods. The non-stationary regression models universally out-performed stationary models in terms of parsimony and predictive skill (figure 4a). Furthermore, stationary models universally suffered from violations of the assumption of independent residuals as seen in elevated residual autocorrelation (figure 4b). A model invoking a fixed SST–catch relationship was the best stationary model in terms of AICc score and residual autocorrelation, but even this model suffered from highly significant autocorrelation in residuals (Durbin–Watson test, DW = 0.51, p = 1.77 × 10−7). This model systematically overestimated catches prior to 1988/1989 and underestimated catches after 1988/1989 (figure 4c). These results suggest that models assuming stationary dynamics, whereby climate–salmon relationships can be evaluated without reference to the relative importance of the PDO and NPGO, are misspecified for application to this system.
Figure 4.

Comparing non-stationary and stationary models of climate effects on salmon catches. (a) Model selection results (ΔAICc scores) for non-stationary and stationary linear regression models explaining variability in catch PC1. Stationary models fit a single regression coefficient to the 1965–2012 time series; non-stationary models fit different slopes before and after 1988/1989. (b) First-order autocorrelation in model residuals. Labels on x-axes indicate covariates in each candidate model. (c) Time series of residuals from best stationary model (explaining catch variability as a fixed response to SST) showing the systematic errors in the two periods compared with residuals from non-stationary SST-catch model. The vertical dashed line indicates 1988/1989.
The best-supported change point in the SST–catch relationship was 1988/1989, though we found nearly equal support for changing covariance over a broad period of the late 1980s (figure 5a). SST–catch relationships differed significantly before and after 1988/1989 (SST × era effect: t48 = 6.03, p = 2.6 × 10−7, figure 5b). Nearly identical results were obtained with the PDO (not shown). Likelihood ratio tests showed similar changes across 1988/1989 in SST-catch effects (i.e. a significant SST × era effect) for both wild (L1 = 21.66, p = 3.3 × 10−6) and hatchery-subsidized fisheries (L1 = 13.96, p = 1.9 × 10−4), leading us to conclude that hatchery effects do not explain non-stationary relationships with temperature. Spawner–recruit time series for wild runs provided further support for non-stationary climate–salmon relationships. Likelihood ratio tests showed that SST–productivity relationships changed across 1988/1989 for pink (L1 = 28.46, p = 9.6 × 10−8) and sockeye salmon (L1 = 12.97, p = 3.2 × 10−4). These species showed the same pattern observed for SST and catch: a change from strong positive covariance prior to 1988/1989 to an effect that could not be distinguished from zero afterwards (figure 5c). The SST × era effect was not significant for chum salmon (L1 = 1.70, p = 0.19). The overall probability of these results for the three species, using the chi-squared distribution for evaluating multiple p-values [40], is 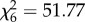 , p = 2.1 × 10−9. Model comparison for spawner–recruit variability in pink salmon, the species providing the most statistical power, showed strongest support for the late 1980s as the timing of change in SST effects (figure 5d).
, p = 2.1 × 10−9. Model comparison for spawner–recruit variability in pink salmon, the species providing the most statistical power, showed strongest support for the late 1980s as the timing of change in SST effects (figure 5d).
Figure 5.

Non-stationary SST–salmon relationships in the Gulf of Alaska. (a) Model selection results comparing support for different timing of changing SST–catch PC1 relationships; lowest values indicate strongest support for a particular year as the timing of change. (b) Catch PC1 versus SST before/after the best threshold year (1988/1989): linear regressions with 95% CI. (c) SST effects on productivity (recruits per spawner) for wild runs before/after 1988/1989: estimated coefficients and 95% CI. (d) Model selection results comparing support for different timing of changing SST effects on pink salmon recruits per spawner. Vertical dashed lines in (a,d) indicate 1988/1989.
Finally, we found little evidence for stationary mechanistic effects on spawner–recruit relationships across the 1988/1989 event. Of the 18 covariate × species comparisons tested, only one (Papa advection index for pink salmon) showed a significant relationship with productivity in both eras (figure 6).
Figure 6.

Non-stationarity in proposed mechanisms controlling salmon productivity: hypothesis testing results (p-values) from mixed-effects Ricker models fit to data from before (a) and after (b) 1988/1989. Dashed horizontal lines indicate value of α (0.014) required to maintain the expected experiment-wide type I error rate at 0.05; values at or below this line are considered evidence for rejecting the null hypothesis of no effect.
4. Discussion
We found support for the hypothesis that the relationships governing ocean climate effects on Gulf of Alaska salmon catches and productivity are non-stationary. Relationships between the PDO and Gulf of Alaska environmental variables, patterns of multivariate environmental variability within the Gulf of Alaska, and relationships between physical variables and salmon catches and productivity all depend on the relative importance of the PDO and NPGO in North Pacific climate and cannot be assumed to be constant over time. It is widely recognized that climate change means that physical and biological time series are increasingly likely to show non-stationary mean and variance [41], but ecologists retain the tendency to model relationships between climatic and biological variables as stationary properties [1]. Our results demonstrate the limitation of this approach for salmon in the Gulf of Alaska ecosystem and suggest that understanding of climate effects on this well-studied system may be rapidly advanced through consideration of non-stationary relationships over time. Similar patterns of changing climate–productivity relationships for several Gulf of Alaska forage fish, groundfish and crustacean populations suggest that non-stationary dynamics governing salmon productivity are likely part of a community-wide pattern [42]. We conclude that the pre- and post-1988/1989 periods are distinct climate states in the Gulf of Alaska system that cannot be directly compared.
These distinct climate states are primarily different in terms of the strength of collinear relationships among physical variables. When the PDO was exclusively associated with PC1 of North Pacific SST variability (1988 and before), strong variance in the Aleutian Low apparently drove coherent variability in Gulf of Alaska ocean physical processes. During this time, the Aleutian Low SLPa index faithfully represents regional wind stress forcing on the Gulf of Alaska. But when the NPGO became more important after 1988 and Aleutian Low variance declined, the strength of association among individual Gulf of Alaska environmental variables also declined. Relationships between physical and biological variables that arise and disappear over time, such as SST–salmon and PDO–salmon relationships in the present study, are a long-standing problem in marine ecology [43]. The propensity of the climate system to display previously unobserved patterns [6,7,44] suggests that the perspective we advance here, of allowing for changes in climatic processes and relationships when analysing historical data, might be broadly useful for explaining ephemeral patterns in climate–biology covariance. The increased importance of the NPGO pattern since the late 1980s may have been part of a basin-wide change in the linked atmosphere–ocean system involving a switch between the East Pacific and Central Pacific El Niño Southern Oscillation patterns, resulting in decreased Aleutian Low variability and increased variability in the North Pacific Oscillation atmospheric pattern that is believed to drive the NPGO [11,45]. The basin-wide spatial scale of this change between contrasting atmosphere–ocean climate patterns suggests the possibility of novel ecological dynamics in other North Pacific ecosystems. Previous suggestions of changing climate–biology relationships in the Bering Sea [15] and California Current [46–48] support this possibility. The PDO and Aleutian Low strongly influence the Gulf of Alaska [5], while the NPGO has a weaker influence on this area [17]. Other North Pacific ecosystems with other degrees of relative association with the PDO and NPGO patterns might therefore show different patterns of non-stationary relationships across this event. Additionally, the 2014–2016 North Pacific marine heatwave, which was not considered in our analysis, appears to reflect increased coupling between the PDO and NPGO patterns [49,50]. These observations suggest the possibility of continuing evolution in the relative importance of leading climate patterns in the North Pacific beyond the pre- and post-1988/1989 states we discuss here. The PDO and NPGO indices are simple statistical representations of complex climate variability. While the utility of these indices is undeniable, ecologists should resist the fallacy of viewing them as more than statistical summaries. Thus while it is acceptable to refer to ‘PDO effects’ when describing statistical relationships with the PDO index, the PDO and NPGO are not themselves physical modes of climate variability [9]. The ecological importance of these indices depends on their relationship to the primary climate variables that actually regulate the population or community of interest [4] and stationarity in these relationships is by no means guaranteed [9]. Attempts to understand the ecological implications of emerging climate patterns will therefore likely be aided by the application of models that allow associations among environmental and biological variables to change over time.
As is always the case with observational studies, we cannot rule out uncontrolled or unmeasured confounding covariates (e.g. changes in population or community dynamics or fishing practices) that might contribute to non-stationary climate–salmon relationships. However, the multiple instances of non-stationary relationships that we document among physical variables, as well as simultaneous non-stationary relationships between climate and non-salmon taxa [42], strongly suggest a role of novel climate associations as a critical driver of non-stationary relationships between climate variables and Gulf of Alaska salmon. Novel climates are an extremely active area of research in global change ecology, but the focus remains on the emergence of these states in the future [3]. However, anthropogenic climate change is emerging from the envelope of natural variability in many ecosystems globally, including the Gulf of Alaska [21]. Anthropogenic forcing produces different rates of change across different environmental variables, suggesting the increasing likelihood of novel relationships. Therefore, recognition of novel ecosystem dynamics within the observational record, including the demise of previously robust climate–biology relationships, is likely to become increasingly common.
Supplementary Material
Acknowledgements
We thank Brigitte Dorner, David Hill, Michael Malick, Randall Peterman and Stephani Zador for sharing data; Nick Bond, Rich Brenner, Bethany Johnson, Emanuele Di Lorenzo, Michael Malick and Franz Mueter for helpful insights and feedback; and Randall Peterman and an anonymous reviewer for helpful comments on this paper.
Data accessibility
Data and code for replicating all reported results are publicly available in the non-stationary climate–salmon repository (https://github.com/mikelitzow/non-stationary_climate-salmon) and on Dryad Digital Repository at http://dx.doi.org/10.5061/dryad.3mg69k5 [51].
Authors' contributions
M.A.L., L.C. and R.R.R. designed the study. M.A.L., L.C. and P.P. designed the biological analysis, and M.A.L., J.J.W., R.R.R. and M.O. designed the climate analysis. M.A.L. conducted analysis and all authors contributed to the interpretation of results. M.A.L. wrote the first draft of the paper and all authors contributed substantially to revisions.
Competing interests
We declare we have no competing interests.
Funding
This article was supported by US National Science Foundation (grant nos OCE-1756081, OCE-1558648, and OCE-1558787).
References
- 1.Wolkovich EM, Cook BI, McLauchlan KK, Davies TJ. 2014. Temporal ecology in the Anthropocene. Ecol. Lett. 17, 1365–1379. ( 10.1111/ele.12353) [DOI] [PubMed] [Google Scholar]
- 2.Dormann CF, et al. 2013. Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography (Cop.) 36, 27–46. ( 10.1111/j.1600-0587.2012.07348.x) [DOI] [Google Scholar]
- 3.Williams JW, Jackson ST. 2007. Novel climates, no-analog communities, and ecological surprises. Front. Ecol. Environ. 5, 475–482. ( 10.1890/070037) [DOI] [Google Scholar]
- 4.Stenseth NC, Ottersen G, Hurrell JW, Mysterud A, Lima M, Chan KS, Yoccoz NG, Adlandsvik B. 2003. Studying climate effects on ecology through the use of climate indices: the North Atlantic Oscillation, El Nino Southern Oscillation and beyond. Proc. R. Soc. Lond. B 270, 2087–2096. ( 10.1098/rspb.2003.2415) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Mantua NJ, Hare SR, Zhang Y, Wallace JM, Francis RC. 1997. A Pacific interdecadal climate oscillation with impacts on salmon production. Bull. Am. Meteorol. Soc. 78, 1069–1079. ( 10.1175/1520-0477(1997)078%3C1069:apicow%3E2.0.co;2) [DOI] [Google Scholar]
- 6.MacDonald GM, Case RA. 2005. Variations in the Pacific Decadal Oscillation over the past millennium. Geophys. Res. Lett. 32 ( 10.1029/2005GL022478) [DOI] [Google Scholar]
- 7.Wittenberg AT. 2009. Are historical records sufficient to constrain ENSO simulations? Geophys. Res. Lett. 36, L12702 ( 10.1029/2009GL038710) [DOI] [Google Scholar]
- 8.Capotondi A, et al. 2015. Understanding ENSO diversity. Bull. Am. Meteorol. Soc. 96, 921–938. ( 10.1175/bams-d-13-00117.1) [DOI] [Google Scholar]
- 9.Newman M, et al. 2016. The Pacific Decadal Oscillation, revisited. J. Clim. 29, 4399–4427. ( 10.1175/JCLI-D-15-0508.1) [DOI] [Google Scholar]
- 10.Springer AM, van Vliet GB. 2014. Climate change, pink salmon, and the nexus between bottom-up and top-down forcing in the subarctic Pacific Ocean and Bering Sea. Proc. Natl Acad. Sci. USA 111, E1880–E1888. ( 10.1073/pnas.1319089111) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Di Lorenzo E, Cobb KM, Furtado JC, Schneider N, Anderson BT, Bracco A, Alexander MA, Vimont DJ. 2010. Central Pacific El Niño and decadal climate change in the North Pacific Ocean. Nat. Geosci. 3, 762–765. ( 10.1038/ngeo984) [DOI] [Google Scholar]
- 12.Yeh SW, Kang YJ, Noh Y, Miller AJ. 2011. The North Pacific climate transitions of the winters of 1976/77 and 1988/89. J. Clim. 24, 1170–1183. ( 10.1175/2010jcli3325.1) [DOI] [Google Scholar]
- 13.Bond NA, Overland JE, Spillane M, Stabeno P. 2003. Recent shifts in the state of the North Pacific. Geophys. Res. Lett. 30, 2183 ( 10.1029/2003gl018597) [DOI] [Google Scholar]
- 14.Sydeman WJ, Santora JA, Thompson SA, Marinovic B, Di Lorenzo E. 2013. Increasing variance in North Pacific climate relates to unprecedented ecosystem variability off California. Glob. Chang. Biol. 19, 1662–1675. ( 10.1111/gcb.12165) [DOI] [PubMed] [Google Scholar]
- 15.Mantua NJ. 2009. Patterns of change in climate and Pacific salmon production. In American fisheries society symposium (eds Krueger CC, Zimmerman CE), pp. 1143–1157. Bethesda, MD: American Fisheries Society. [Google Scholar]
- 16.Kilduff DP, Di Lorenzo E, Botsford LW, Teo SLH. 2015. Changing central Pacific El Niños reduce stability of North American salmon survival rates. Proc. Natl Acad. Sci. USA 112, 10 962–10 966. ( 10.1073/pnas.1503190112) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Di Lorenzo E, et al. 2008. North Pacific Gyre Oscillation links ocean climate and ecosystem change. Geophys. Res. Lett. 32, L08607. [Google Scholar]
- 18.Lindegren M, Checkley DM Jr, Koslow JA, Goericke R, Ohman MD. 2018. Climate-mediated changes in marine ecosystem regulation during El Niño. Glob. Chang. Biol. 24, 796–809. ( 10.1111/gcb.13993) [DOI] [PubMed] [Google Scholar]
- 19.Debertin AJ, Irvine JR, Holt CA, Oka G, Trudel M. 2017. Marine growth patterns of southern British Columbia chum salmon explained by interactions between density-dependent competition and changing climate. Can. J. Fish. Aquat. Sci. 74, 1077–1087. ( 10.1139/cjfas-2016-0265) [DOI] [Google Scholar]
- 20.Jeffrey KM, Cote IM, Irvine JR, Reynolds JD. 2017. Changes in body size of Canadian Pacific salmon over six decades. Can. J. Fish. Aquat. Sci. 74, 191–201. ( 10.1139/cjfas-2015-0600) [DOI] [Google Scholar]
- 21.Walsh JE, et al. 2018. The high latitude heat wave of 2016 and its impacts on Alaska. Bull. Am. Meteorol. Soc. 99, S39–S43. ( 10.1175/BAMS-D-17-0105.1) [DOI] [Google Scholar]
- 22.Hare SR, Mantua NJ. 2000. Empirical evidence for North Pacific regime shifts in 1977 and 1989. Prog. Oceanogr. 47, 103–145. ( 10.1016/s0079-6611(00)00033-1) [DOI] [Google Scholar]
- 23.Malick MJ, Cox SP. 2016. Regional-scale declines in productivity of pink and chum salmon stocks in western North America. PLoS ONE 11, e0146009 ( 10.1371/journal.pone.0146009) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Peterman RM, Dorner B. 2012. A widespread decrease in productivity of sockeye salmon (Oncorhynchus nerka) populations in western North America. Can. J. Fish. Aquat. Sci. 69, 1255–1260. ( 10.1139/f2012-063) [DOI] [Google Scholar]
- 25.Mueter FJ, Peterman RM, Pyper BJ. 2002. Opposite effects of ocean temperature on survival rates of 120 stocks of Pacific salmon (Oncorhynchus spp.) in northern and southern areas. Can. J. Fish. Aquat. Sci. 59, 456–463. ( 10.1139/f02-020) [DOI] [Google Scholar]
- 26.Malick MJ, Cox SP, Mueter FJ, Peterman RM. 2015. Linking phytoplankton phenology to salmon productivity along a north–south gradient in the Northeast Pacific Ocean. Can. J. Fish. Aquat. Sci. 72, 697–708. ( 10.1139/cjfas-2014-0298) [DOI] [Google Scholar]
- 27.Malick MJ, Cox SP, Mueter FJ, Dorner B, Peterman RM. 2016. Effects of the North Pacific Current on the productivity of 163 Pacific salmon stocks. Fish. Oceanogr. 26, 268–281. ( 10.1111/fog.12190) [DOI] [Google Scholar]
- 28.Henson SA. 2007. Water column stability and spring bloom dynamics in the Gulf of Alaska. J. Mar. Res. 65, 715–736. ( 10.1357/002224007784219002) [DOI] [Google Scholar]
- 29.Weingartner TJ, Danielson SL, Royer TC. 2005. Freshwater variability and predictability in the Alaska Coastal Current. Deep. Res. Part Ii-Topical Stud. Oceanogr. 52, 169–191. ( 10.1016/j.dsr2.2004.09.030) [DOI] [Google Scholar]
- 30.Gargett AE. 1997. The optimal stability ‘window’: a mechanism underlying decadal fluctuations in North Pacific salmon stocks? Fish. Oceanogr. 6, 109–117. ( 10.1046/j.1365-2419.1997.00033.x) [DOI] [Google Scholar]
- 31.Yeh SW, Wang X, Wang C, Dewitte B. 2015. On the relationship between the North Pacific Climate variability and the Central Pacific El Niño. J. Clim. 28, 663–677. ( 10.1175/JCLI-D-14-00137.1) [DOI] [Google Scholar]
- 32.Pyper BJ, Peterman RM. 1998. Comparison of methods to account for autocorrelation in correlation analyses of fish data. Can. J. Fish. Aquat. Sci. 55, 2127–2140. ( 10.1139/cjfas-55-9-2127) [DOI] [Google Scholar]
- 33.Stabeno PJ, Bond NA, Hermann AJ, Kachel NB, Mordy CW, Overland JE. 2004. Meteorology and oceanography of the Northern Gulf of Alaska. Cont. Shelf Res. 24, 859–897. ( 10.1016/j.csr.2004.02.007) [DOI] [Google Scholar]
- 34.Burnham KP, Anderson DR. 2002. Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn New York, NY: Springer-Verlag. [Google Scholar]
- 35.Pinheiro JC, Bates DM. 2000. Mixed-effects models in S and S-PLUS. New York, NY: Springer. [Google Scholar]
- 36.Zuur AF, Ieno N, Walker N, Saveliev AA, Smith GM. 2009. Mixed effects models and extensions in ecology with R. Berlin, Germany: Springer. [Google Scholar]
- 37.Pyper BJ, Mueter FJ, Peterman RM. 2005. Across-species comparisons of spatial scales of environmental effects on survival rates of Northeast Pacific salmon. Trans. Am. Fish. Soc. 134, 86–104. ( 10.1577/t04-034.1) [DOI] [Google Scholar]
- 38.Verhoeven KJF, Simonsen KL, McIntyre LM. 2005. Implementing false discovery rate control: increasing your power. Oikos 108, 643–647. ( 10.1111/j.0030-1299.2005.13727.x) [DOI] [Google Scholar]
- 39.R Core Team. 2018. R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; See http://www.r-project.org/. [Google Scholar]
- 40.Sokal RR, Rohlf FJ. 1995. Biometry: the principles and practice of statistics in biological research, 3rd edn New York, NY: Freeman. [Google Scholar]
- 41.Szuwalski CS, Hollowed AB. 2016. Climate change and non-stationary population processes in fisheries management. ICES J. Mar. Sci. 73, 1297–1305. ( 10.1093/icesjms/fsv229) [DOI] [Google Scholar]
- 42.Puerta P, Ciannelli L, Rykaczewski R, Opiekun M, Litzow MA.Do Gulf of Alaska fish and crustacean populations show synchronous non-stationary responses to climate? Prog. Oceanogr. In press.
- 43.Myers RA. 1998. When do environment-recruitment correlations work? Rev. Fish Biol. Fish. 8, 285–305. ( 10.1023/A:1008828730759) [DOI] [Google Scholar]
- 44.Jackson ST, Betancourt JL, Booth RK, Gray ST. 2009. Ecology and the ratchet of events: climate variability, niche dimensions, and species distributions. Proc. Natl Acad. Sci. USA 106, 19 685–19 692. ( 10.1073/pnas.0901644106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yeh SW, Kug J-S, Dewitte B, Kwon M-H, Kirtman BP, Jin F-F. 2009. El Niño in a changing climate. Nature 461, 511–514. ( 10.1038/nature08316) [DOI] [PubMed] [Google Scholar]
- 46.Miller AK, Sydeman WJ. 2004. Rockfish response to low-frequency ocean climate change as revealed by the diet of a marine bird over multiple time scales. Mar. Ecol. Prog. Ser. 281, 207–216. ( 10.3354/meps281207) [DOI] [Google Scholar]
- 47.Schmidt AE, Botsford LW, Eadie JM, Bradley RW, Di Lorenzo E, Jahncke J. 2014. Non-stationary seabird responses reveal shifting ENSO dynamics in the northeast Pacific. Mar. Ecol. Prog. Ser. 499, 249–258. ( 10.3354/meps10629) [DOI] [Google Scholar]
- 48.King JR, Agostini VN, Harvey CJ, McFarlane GA, Foreman MGG, Overland JE, Di Lorenzo E, Bond NA, Aydin KY. 2011. Climate forcing and the California Current ecosystem. ICES J. Mar. Sci. 68, 1199–1216. ( 10.1093/icesjms/fsr009) [DOI] [Google Scholar]
- 49.Joh Y, Di Lorenzo E. 2017. Increasing coupling between NPGO and PDO leads to prolonged marine heatwaves in the Northeast Pacific. Geophys. Res. Lett. 44, 11 663–11 671. ( 10.1002/2017gl075930) [DOI] [Google Scholar]
- 50.Di Lorenzo E, Mantua N. 2016. Multi-year persistence of the 2014/15 North Pacific marine heatwave. Nat. Clim. Chang. 6, 1042–1047. ( 10.1038/nclimate3082) [DOI] [Google Scholar]
- 51.Litzow M, Ciannelli L, Puerta P, Wettstein J, Rykaczewski R, Opiekun M. 2018. Data from: Non-stationary climate–salmon relationships in the Gulf of Alaska Dryad Digital Repository. ( 10.5061/dryad.3mg69k5) [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Citations
- Litzow M, Ciannelli L, Puerta P, Wettstein J, Rykaczewski R, Opiekun M. 2018. Data from: Non-stationary climate–salmon relationships in the Gulf of Alaska Dryad Digital Repository. ( 10.5061/dryad.3mg69k5) [DOI] [PMC free article] [PubMed]
Supplementary Materials
Data Availability Statement
Data and code for replicating all reported results are publicly available in the non-stationary climate–salmon repository (https://github.com/mikelitzow/non-stationary_climate-salmon) and on Dryad Digital Repository at http://dx.doi.org/10.5061/dryad.3mg69k5 [51].