

Abstract
Dorsal root ganglia (DRG) contain cell bodies of sensory neurons. This type of neuron is pseudo-unipolar, with two axons that innervate peripheral tissues, such as skin, muscle and visceral organs, as well as the spinal dorsal horn of the central nervous system. Sensory neurons transmit somatic sensation, including touch, pain, thermal, and proprioceptive sensations. Therefore, DRG primary cultures are widely used to study the cellular mechanisms of nociception, physiological functions of sensory neurons, and neural development. The cultured neurons can be applied in studies involving electrophysiology, signal transduction, neurotransmitter release, or calcium imaging. With DRG primary cultures, scientists may culture dissociated DRG neurons to monitor biochemical changes in single or multiple cells, overcoming many of the limitations associated with in vivo experiments. Compared to commercially available DRG-hybridoma cell lines or immortalized DRG neuronal cell lines, the composition and properties of the primary cells are much more similar to sensory neurons in tissue. However, due to the limited number of cultured DRG primary cells that can be isolated from a single animal, it is difficult to perform high-throughput screens for drug targeting studies. In the current article, procedures for DRG collection and culture are described. In addition, we demonstrate the treatment of cultured DRG cells with an agonist of neuropeptide FF receptor type 2 (NPFFR2) to induce the release of peptide neurotransmitters (calcitonin gene-related peptide (CRGP) and substance P (SP)).
Keywords: Retraction, Issue 140, Dorsal root ganglia, DRG, primary culture, neuronal cultures, CGRP, substance P, neurotransmitter, sensory neuron, pain, pain transmission, nociception
Introduction
The cell bodies of sensory neurons are contained within DRG. These neurons are pseudo-unipolar and innervate both peripheral tissues and the central nervous system. The peripheral nerve endings of sensory neurons are found in muscle, skin, visceral organs, and bone, among other tissues. They transmit peripheral sensation signals to nerve endings in the spinal dorsal horn and the signals are then transmitted to the brain via different ascending pathways of somatic sensation1,2. Somatic sensation enables the body to feel (i.e., touch, pain, and thermal sensations) and perceive movement and spatial orientation (proprioceptive sensations)1,3. There are four subclasses of primary afferent axons, including group I (Aα) fibers that respond to proprioception of skeletal muscles, group II (Aβ) fibers that respond to mechanoreceptors of the skin, and group III (Aδ) and group V (C) fibers that respond to pain and temperature. Only the C fibers are unmyelinated, while the rest are myelinated to different degrees.
Nociceptors are primary sensory neurons, which are activated by noxious stimuli (mechanical, thermal, and chemical stimulation) that carry potential for tissue damage. These neurons are composed of myelinated Aδ fibers and unmyelinated C fibers1,4. The Aδ fibers express the receptors for nerve growth factor (NGF, trkA receptor), CGRP, and SP. The C fibers are classified as either peptidergic and non-peptidergic C fibers. On the other hand, the non-peptidergic C fibers express the receptors for glial-derived neurotrophic factor (GDNF, RET, and GFR receptors), isolectin IB4, and ATP-gated ion channel subtype (P2X3)5,6,7. Nociceptors can be distinguished by the expression of ion channels and activated by neurotrophic factors, cytokines, neuropeptides, ATP, or other chemical compounds8. Upon stimulation, neurotransmitters, including CGRP, SP, and glutamate may be released from sensory neuron terminals in the spinal dorsal horn to transmit nociceptive signals2. DRG are not only composed of neurons, but also contain satellite glial cells. Satellite cells surround the sensory neurons and provide mechanical and metabolic support9,10. Interestingly, there is a growing body of evidence indicating that satellite glial cells in the DRG may be involved in regulating pain sensation11.
Sensory neurons have been reported to be the most frequently used primary neuronal cells12 and have been utilized for electrophysiology, signal transduction, and neurotransmitter release studies. They are also commonly used to explore the cellular mechanisms of neuronal development, inflammatory pain, neuropathic pain, skin sensation (like itch), and axon outgrowth12,13,14,15. DRG primary cultures can be cultured as dissociated neurons to assess biochemical changes in single or multiple cells, allowing scientists to perform studies that cannot be performed in experimental subjects. Recently, DRG were successfully cultured from human organ donors which might greatly benefit translational research16. On the other hand, sensory neurons can also be cultured as DRG explants. The DRG explants preserve the original tissue architecture of the neurons, including Schwann cells and satellite glial cells, and are especially useful to study interactions between neuronal and non-neuronal cells17. DRG primary cultures can be easily prepared within 2.5 h. The cell composition and properties are highly reflective of the source DRG, and as such, specific DRG (lumbar or thoracic DRG) can be collected according to experimental demands. Cultures of embryonic and neonatal DRG neurons require NGF to survive and induce axon outgrowth, but cultures of adult neurons do not require the addition of neurotrophic factors to the media12,17. There are also commercially available DRG-hybridoma cell lines such as ND7/23 and F11, which do not require the use of experimental animals. However, the lack of the transient receptor potential cation channel subfamily V member 1 (TRPV1) expression (an important marker for small sensory nociceptive neurons) and incongruent gene expression profiles limit their applications18. Recently, immortalized DRG neuronal cell lines have been derived from rat (50B11)19 and mouse (MED17.11)20, which are suitable for use in high-throughput screens for drug targeting studies. However, gene expression profiling for these cell lines has yet to be performed. Thus, the validation experiments comparing these immortalized cells to sensory neurons are still ongoing.
NPFFR2 is synthesized in the DRG and translocated to the sensory nerve terminals in the spinal dorsal horn21. In this article, we provide a protocol for culturing lumbar DRG cells and treating them with an agonist of NPFFR2 to induce the release of neurotransmitters, CGRP and SP. The dependence on NPFFR2 is further tested using NPFFR2 small interfering RNA (siRNA), which may be transfected into the cultured DRG cells.
Protocol
All methods described herein that use experimental animals were approved by the Institutional Animal Care and Use Committee (IACUC) of Chang Gung University (CGU 13-014).
1. Collect Lumbar DRG from Experimental Rats
Use 2 to 3 week-old Sprague-Dawley (SD) rats for lumbar DRG collection. NOTE: DRG neurons collected from rats over 4 weeks of age do not grow well under the culture conditions described herein.
Sterilize all surgical instruments in an autoclave.
Anesthetize the rat with a 1:1 mixture of tiletamine and zolazepam (20 mg/kg; intraperitoneal injection (IP)) and wait until the animal shows no foot-withdrawal response in a toe-pinch test. NOTE: Different anesthesia strategies can be used successfully in this protocol.
Sacrifice the rat by decapitation with a commercial guillotine.
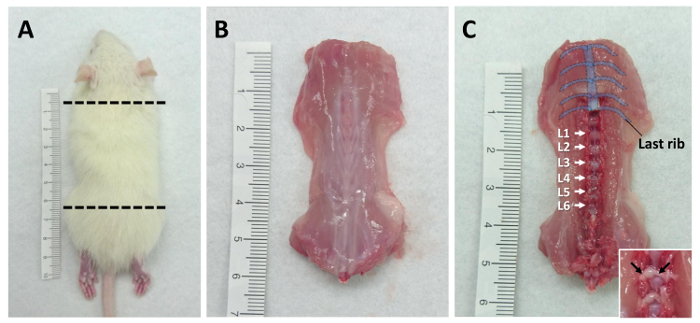
Use the guillotine to isolate the body trunk of the rat between the forelimb and femur. See Figure 1A for a diagram of the region to be collected. NOTE: The caudal cut line should be just rostral to the femur. The lumbar L6 DRG will be excised if the cut site is too high in the spinal column.
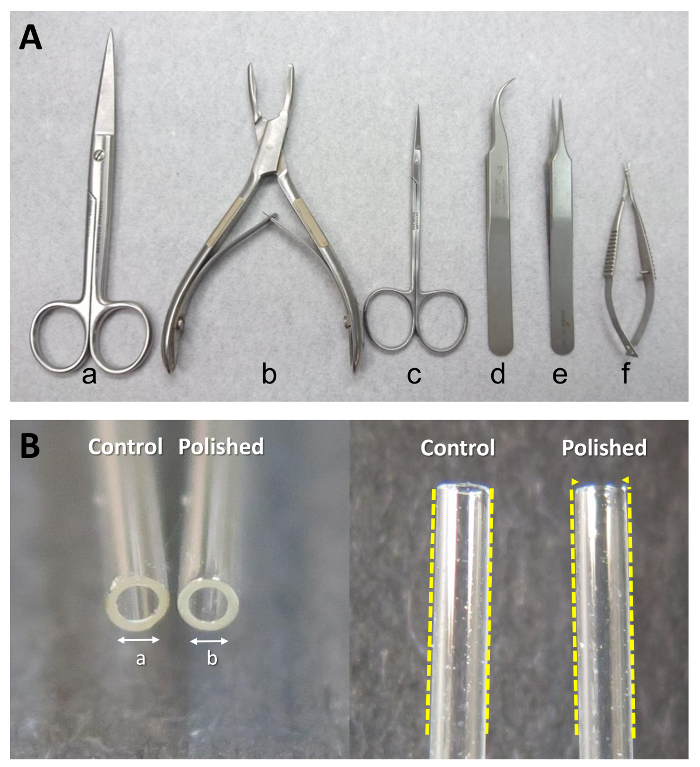
Cut along the sternum and remove all organs/tissues with dissection scissors (Figure 2A-a).
Cut along the side of trunk to collect the dorsal part of the rat and remove the skin. See Figure 1B for a photograph of the dissected dorsal trunk.
Prepare the tissue on ice before collecting DRG. Clean the fur and blood from gloves, and sterilize them with 75% ethanol before proceeding to the next step.
Remove the muscles covering the lumbar spine. First, make two cuts along the sides of the spinal column (left and right) and one lateral cut to mark the rostral extent of the lumbar spine. Then, remove the dorsal muscles of the spine with bone cutting forceps (Figure 2A-b).
Remove the dorsal portion of the vertebrae with bone cutting forceps and expose the spinal cord.
Remove the spinal cord with dissection scissors (Figure 2A-c) and forceps (Figure 2A-d).
Identify the lumbar DRG by counting vertebrae from the last rib (Thoracic Vertebra 13). See Figure 1C for a diagram of the vertebrae positions.
Collect the bilateral lumbar DRG (L1-L6) with micro-scissors (Figure 2A-f) into a 35-mm culture dish with 2 mL ice-cold serum-free medium. Remove the neuronal fibers (as indicated in Figure 1C) from connecting DRG, then transfer it into the culture dish to improve the purity of the cultures. NOTE: The collected DRG can be kept in medium on ice for about 1 h. Meanwhile, multiple rats can be euthanized to create a larger pool of DRG.
2. Primary Culture of Rat Lumber DRG
NOTE: The following steps should be performed in a laminar flow hood.
Prepare culture medium containing 10% fetal bovine serum, 100 mM sodium pyruvate, and 1x penicillin/streptomycin in 1x DMEM-F12.
Coat the cell-culture treated 24-well plate with 200 µg/mL poly-L-lysine for 2 h then wash with sterilized water.
Pre-incubate the culture dish with 1 mL culture medium in a 37 °C CO2 incubator before use for least 30 min.
Transfer the DRG-containing 35 mm dish into a laminar hood, and wash the DRG with serum-free medium 3 times by pipette. NOTE: The outside of the dish should be cleaned with 75% ethanol before transferring into the hood. The 35 mm dish can contain DRG from a number of rats (this will depend on the demands of the experimental design).
Move the DRG (from a single rat or combined from multiple rats) to a new 35 mm culture dish, which contains 2 mL of collagenase type IA (1 mg/mL in serum-free medium) with sterile tweezers (Figure 2A-e). NOTE: The collagenase solution should be sterilized by passing it through a 0.22 µm syringe filter.
Digest the DRG in the collagenase solution in a 37 °C CO2 incubator for 30 min.
Remove the collagenase solution and wash the DRG 3 times in 2 mL Hank's balanced salt solution (HBSS). NOTE: There may be residual fibers or tissues that come off the DRG into the solution. Remove them by pipette with the washing solution.
Add 2 mL pre-warmed 0.05% trypsin-EDTA into the DRG-containing 35 mm dish and digest the DRG in a 37 °C CO2 incubator for 30 min.
Transfer the 2 mL of DRG-containing solution to a 15 mL centrifuge tube by glass pipette. NOTE: The DRG might stick to the glass pipette so this step should be performed with care. DRG loss can be avoided by keeping the DRG-containing solution in the tapered end of a glass pipette (about 0.5 mL) and transferring the solution into the centrifuge tube slowly but without pause.
Centrifuge the solution at 290 x g for 5 min at 4 °C. Remove the supernatant and add another 2-mL serum-free medium to resuspend the DRG.
Repeat step 2.10 2 times but change the serum-free medium to pre-warmed culture medium on the last time.
Manually triturate the DRG approximately 60 times using a flame-polished Pasteur pipette (length 230 mm and tip head inner diameter 1 mm). See Figure 2B for a photograph comparing the orifice of a flame-polished Pasteur pipette to a non-polished pipette. NOTE: The inside diameter of the flame-polished Pasteur pipette is approximately 10% smaller than the control pipette and the inside of the tapered end should be smoother. Be careful not to create bubbles when triturating the cells.
Remove the poly-L-lysine-coated dish from the CO2 incubator. Aspirate the incubated culture medium from the dish, and seed the dissociated cells onto the coated dish.
Seed the DRG cells from one rat (bilateral collection from L1-L6, for 12 total DRG) into four wells of a 24-well plate; there are approximately 5 x 104 cells in one well of a 24-well plate. NOTE: This density is suitable for the detection of the released CGRP or SP and also suitable for immunostaining. For Western blot or RNA extraction, seed the DRG cells from one rat (bilateral L1-L6) into one well of a 6-well plate.
Replace the culture medium on the following day with the addition of 10 µM cytarabine (Ara-C) and 100 ng/mL NGF, and refresh the medium every two days thereafter. NOTE: The thoracic DRG also can also be cultured by this protocol, if they have been collected from the thoracic spine.
3. Transfection of NPFFR2 siRNA in DRG Cells
Perform the transfection of NPFFR2 siRNA and control siRNA according the manufacturer's protocol. NOTE: The protocol will need to be adapted if the chosen transfection reagent is different from the one we used (see the Table of Materials).
On Day 3 after cell plating, change the medium to 0.5 mL pre-warm serum-free medium and incubate the DRG in a 37 °C CO2 incubator for 1 h.
Add 50 nM of siRNA (in 1 µL RNase-free water) into 12.5 µL serum-free medium.
Add 2.5 µL transfection reagent into 10 µL serum-free medium.
Mix the solution from steps 3.3 and 3.4 by pipette, and incubate this mixed transfection solution for 10 min at room temperature.
Add the transfection solution into one DRG-containing 24-well plate and mix the solution with medium by gentle shaking. NOTE: Multiple transfection solutions should be deployed at the same time if multiple wells need to be transfected.
Incubate the DRG in a 37 °C CO2 incubator for 6 h.
Add 0.5 mL/well of culture medium containing 20% fetal bovine serum, 100 mM sodium pyruvate, and 1x penicillin/streptomycin in 1x DMEM-F12, with the addition of 10 µM Ara-C and 100 ng/mL NGF, into the 24-well plate.
Incubate the DRG in a 37 °C CO2 incubator for another 66 h (refresh the medium at 48 h).
4. Release of Neurotransmitters from Primary DRG Cells
On Day 6 after cells were plated (72 h after siRNA transfection), change the culture medium to 200 µL serum-free medium, and incubate the cells in a 37 °C CO2 incubator for 30 min.
Add 1 µL stimulation chemical(s) and gently mix the media by pipetting. Incubate the dish in a 37 °C CO2 incubator for the designated time. NOTE: In this article, the cultured cells were stimulated with the NPFFR2 agonist, dNPA (D.Asn-Pro-(N-Me)Ala-Phe-Leu-Phe-Gln-Pro-Gln-Arg- Phe-NH2, 5 nmol), for 1 h.
Collect the culture medium from the culture dish and centrifuge at 5,000 x g for 5 min at 4 °C to remove any suspended impurities.
Collect the supernatant from the centrifugation and dilute the samples with phosphate-buffered saline (PBS), as needed. Assay the levels of neurotransmitters with enzyme immunoassay (EIA) kits. NOTE: Here, the supernatants were diluted 1:100 before analyzing the level of CGRP. The supernatant was not diluted before analyzing the level of SP.
5. CGRP and SP EIA
Analyze the samples immediately according to the CGRP or SP EIA kit manufacturer's protocol. NOTE: The protocol will vary depending on the kit used.
Rinse the CGRP EIA wells 5 times with wash buffer supplied within the kit.
Add 100 µL samples with 100 µL anti-CGRP acetylcholinesterase (AChE) tracer into the CGRP EIA wells, and add 50 µL samples, 50 µL anti-SP AChE tracer and 50 µL anti-SP antiserum into the SP EIA wells.
Seal the CGRP and SP wells with plastic film which is supplied within the kits.
Incubate the wells overnight at 4 °C.
Wash the wells 5 times with CGRP or SP wash buffer and remove all the residual solution from the wells.
Add 200 µL Ellman's reagent into the CGRP or SP wells which is supplied within the corresponding EIA kits.
Incubate the CGRP wells for 30 min at room temperature, and incubate the SP wells for 90 min at room temperature. Protect the wells from light for both assays.
Read the plates at wavelength 414 nm and calculate the results according to the corresponding EIA instrument. NOTE: Avoid touching the bottom of the wells by hand all the time and clean the water stains from the well bottom by lens cleaning wipes before adding the Ellman's reagent.
Representative Results
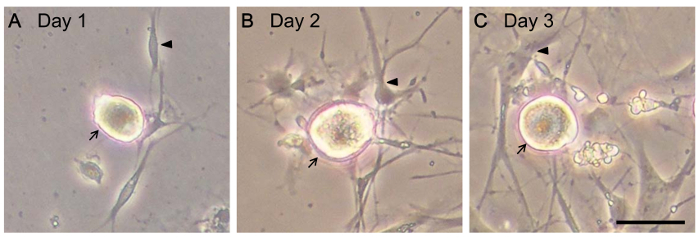
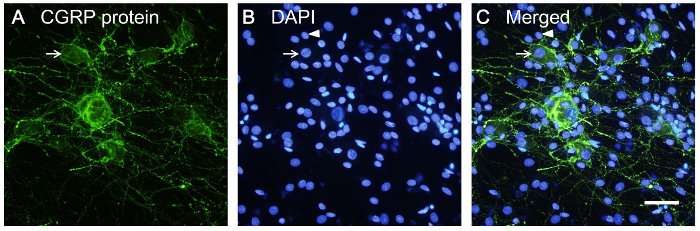
Rat lumbar DRG neurons, cultured in a 24-well plate, were grown in culture medium with additional Ara-C to inhibit glial cell proliferation and NGF to support neuronal growth. The morphology of living DRG cells was observed. As shown in Figure 3, the cell body of a single neuron was attached on the bottom of a dish at Day 1 and selected for observation. Axon growth was monitored from Day 1–3. The glial cells duplicated and extended processes to surround the cell body of the sensory neuron. In another culture, CGRP protein was stained to reveal the shape of neurons. In Figure 4, CGRP protein staining appears in the cytoplasm and axons of sensory neurons. The nuclear morphologies of neurons and glial cells are distinct when stained with DAPI. The neurons have a larger and more rounded nucleus than glial cells. By comparison, the nuclei of glia are more oval in shape (Figure 4B).
The selective NPFFR2 agonist, dNPA, was used to stimulate the release of CGRP and SP. Moreover, the dependence of dNPA-stimulated neurotransmitter release on NPFFR2 was tested by transfecting cells with NPFFR2 siRNA. NPFFR2 siRNA or control siRNA were transfected into the primary DRG cells 72 h prior to agonist treatment. DRG cells were treated with dNPA (5 nmol) for 1 h and the release of CGRP and SP was measured by separate EIA kits. The simulation of DRG with dNPA increased the level of CGRP and SP in the media (Figure 5A, B). However, only the dNPA-induced CGRP release was inhibited by expression of NPFFR2 siRNA in cultured DRG cells. The results shown in Figure 5 were modified from a previous publication and are used here with permission22.
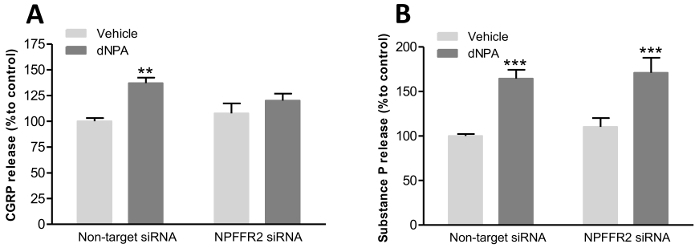
Figure 1: Tissue processing diagrams. Lumbar DRG are collected from 3 week-old rats. (A) The positions where the guillotine should be used to cut the animal are indicated by dotted lines. (B) The dorsal trunk with skin removed and (C) the locations of lumbar DRG (from L1-L6) are shown. The insert represents the DRG and the connecting fibers (which are indicated by the arrows). Please click here to view a larger version of this figure.
Figure 2: Special equipment needed for isolating DRG primary cultures. (A) Surgical instruments used in the collection of DRG. From left to right: (a) dissection scissors (large), (b) bone cutting forceps, (c) dissection scissors (small), (d, e) point tweezers, and (f) micro-scissors. (B) A regular Pasteur pipette and a flame-polished Pasteur pipette. "a" denotes the inside diameter of regular Pasteur pipette, and "b" denotes the inside diameter of the flame-polished Pasteur pipette. b/a ≒ 0.9. Please click here to view a larger version of this figure.
Figure 3: The morphology of living DRG cells. Live DRG cells were monitored by microscopy. Cells are shown (A) one day after seeding, (B) two days after seeding, and (C) three days after seeding. Arrows indicate neuron and arrowheads indicate glia. Scale bar = 20 µm. Please click here to view a larger version of this figure.
Figure 4: Immunostaining of cultured DRG cells. DRG cells were immunostained with anti-CGRP antibody to show neurons, and 4',6-diamidino-2-phenylindole (DAPI) for the nuclei of neurons and glial cells. (A) CGRP protein was expressed in sensory neuron cell bodies and axon fibers. (B) Nuclei of neurons and glia were stained with DAPI. (C) Merged picture from A and B. Arrow indicates a neuron and arrowhead indicates a glial cell. Scale bar = 30 µm. Please click here to view a larger version of this figure.
Figure 5: The release of neurotransmitters from cultured DRG cells. The selective NPFFR2 agonist, dNPA, was used to stimulate the release of CGRP and SP from DRG cultures. The dependence of neurotransmitter release on NPFFR2 was verified by transfecting cells with NPFFR2 siRNA. (A and B) After DRG cells were transfected with NPFFR2 siRNA or non-targeting control siRNA (72 h), dNPA (5 nmol) was applied for 1 h to induce the release of CGRP and SP. Data are expressed as mean ± standard error of the mean (SEM) and were analyzed by two-way analysis of variance (ANOVA) with Bonferroni post hoc tests. **p <0.01, ***p <0.001; compared to corresponding vehicle controls (N = 12 per group). Panels A and B have been modified from Lin et al.22 Please click here to view a larger version of this figure.
Discussion
In the present article, we demonstrate the collection, enzyme-dissociation, and culture of rat lumbar DRG. With the neurotrophic support from NGF, the axons of DRG neurons extended within 3 days after cell seeding. The extended axons were clearly observable after cells were stained for CGRP protein, which is synthesized in the cell soma and transported along the axon fibers. The processes of satellite cells also extended, allowing these dividing glial cells to surround the neurons within days. The primary DRG cells grown by this protocol are suitable for investigations into the cellular mechanisms that regulate sensory neurons. Here, we stimulate the release of neuropeptides, CGRP and SP, from cultured DRG neurons by a selective NPFFR2 agonist, dNPA. NPFFR2 is the cognate receptor for NPFF and has been reported to participate in pain sensation and regulation pathways22,23. The NPFFR2-dependence of CGRP and SP release was further verified by the use of NPFFR2 siRNA.
DRG cultures contain both sensory neurons and satellite glial cells. The satellite glial cells provide metabolic support to neurons and maintain neuronal functions9,10. In the immunostaining pictures, it is easy to identify the neurons and glial cells, since their nuclei are shaped differently (show in Figure 4B). The existence of satellite cells in the culture dish might become problematic if there is an experimental demand to distinguish between the specific function of neurons and glia. For example, it is undeniable that satellite cells are involved in the development and maintenance of pain11,24, and in some studies, the actions of neurons and glial cells would be difficult to distinguish using DRG cultures.
In this protocol, there are a few critical steps that require extra caution. First, since the DRG are collected outside of the laminar hood, extra care should be taken during the tissue collection process to avoid cell contamination. There should be no need to create a sterile space, like in a human surgical room, but instrument sterilization and a clean operating space are essential. Tips to avoid contamination include keeping the sterilized instruments on the sterilization pouch when not in use and avoiding the touching of any unnecessary items. Also, contaminating organisms may be carried on fur that may stick to the rat body trunk or gloves. As such, fur and bloodstains on gloves should be removed by cleaning with 75% ethanol. It is also helpful for the operator to wear a surgical mask to prevent transfer of organisms from the breath or saliva. Furthermore, the 35-mm dish should be kept closed at all the times and only opened when placing dissected DRG inside. It is important to replace the 35 mm dish with a new dish before enzyme digestion. During enzyme digestion, do not extend the incubation time, since over-digestion may damage the neurons. Make sure to pre-warm the trypsin-EDTA to 37 °C in order to achieve appropriate digestion efficiency. The efficiency will be dramatically reduced in lower temperatures, and it will be difficult to achieve a single cell suspension when triturating the DRG by flame-polished Pasteur pipette. Flame polishing the pipette will smooth the orifice and prevent the sharp glass edge from injuring the neurons. However, overheating the pipette with a flame will make the inside diameter too small, and tissue- or cell-containing solution will become difficult to pass through. This reduced diameter may also cause many bubbles to form during the trituration stage, greatly reducing the collectable number of DRG neurons. Finally, DRG cultures should be handled gently at all times, especially when changing medium or performing drug treatment.
The DRG neurons are reported to be the most frequently used primary cultured neuronal cells12. They can be utilized for a variety of different studies, ranging from electrophysiology or cell biology to exploring the physiological or pathological functions of sensory neurons. The major limitation of DRG primary cultures is that they are not well-suited for high-throughput screening. The number of cells that may be collected from the DRG of a single rat are limited, and the neurons are unable to duplicate in culture. Because of the limited cell number, several DRG-hybridoma cell lines or immortalized DRG neuronal cell lines have been developed to replace the primary cultures18,19,20. However, the protein expression profiles of DRG cell lines might not be the same as the original DRG and, thus, each model system needs to be carefully verified. Aside from isolating the cells in the lab, rat embryonic or neonatal DRG neurons have been made commercially available. Therefore, purchase from commercial sources may be a viable alternative to freshly prepared DRG cultures.
DRG primary cultures have been used for many years as a valuable experimental tool that is mostly adopted in pain-related studies. This model system is unlikely to be replaced in the near future. With good quality DRG neurons, scientists can obtain stable and reproducible results that benefit many areas of neuroscience study.
Disclosures
The authors have nothing to disclose.
Acknowledgments
We thank Dr. M. Calkins for English editing. This work was supported by the Chang Gung Memorial Hospital (CMRPD1F0482), Chang Gung University, Healthy Aging Research Center (EMRPD1G0171) and Ministry of Science and Technology (105-2320-B-182-012-MY2).
References
- Bear MF, Connors BW, Paradiso MA. Neuroscience: exploring the brain. 3 edn. Lippincott Williams & Wilkins; 2007. [Google Scholar]
- Hunt SP, Mantyh PW. The molecular dynamics of pain control. Nat Rev Neurosci. 2001;2(2):83–91. doi: 10.1038/35053509. [DOI] [PubMed] [Google Scholar]
- Kandel ER, Schwartz JH, Jessell TM. Principles of neural science. 3 edn. McGraw-Hill, Health Professions Division; 2000. [Google Scholar]
- Julius D, Basbaum AI. Molecular mechanisms of nociception. Nature. 2001;413(6852):203–210. doi: 10.1038/35093019. [DOI] [PubMed] [Google Scholar]
- Sah DW, Ossipo MH, Porreca F. Neurotrophic factors as novel therapeutics for neuropathic pain. Nat Rev Drug Discov. 2003;2(6):460–472. doi: 10.1038/nrd1107. [DOI] [PubMed] [Google Scholar]
- Coutaux A, Adam F, Willer JC, Le Bars D. Hyperalgesia and allodynia: peripheral mechanisms. Joint Bone Spine. 2005;72(5):359–371. doi: 10.1016/j.jbspin.2004.01.010. [DOI] [PubMed] [Google Scholar]
- Basbaum AI, Bautista DM, Scherrer G, Julius D. Cellular and molecular mechanisms of pain. Cell. 2009;139(2):267–284. doi: 10.1016/j.cell.2009.09.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marchand F, Perretti M, McMahon SB. Role of the immune system in chronic pain. Nat Rev Neurosci. 2005;6(7):521–532. doi: 10.1038/nrn1700. [DOI] [PubMed] [Google Scholar]
- Hanani M. Satellite glial cells in sensory ganglia: from form to function. Brain Res Brain Res Rev. 2005;48(3):457–476. doi: 10.1016/j.brainresrev.2004.09.001. [DOI] [PubMed] [Google Scholar]
- Nascimento RS, Santiago MF, Marques SA, Allodi S, Martinez AM. Diversity among satellite glial cells in dorsal root ganglia of the rat. Braz J Med Biol Res. 2008;41(11):1011–1017. doi: 10.1590/s0100-879x2008005000051. [DOI] [PubMed] [Google Scholar]
- Costa FA, Moreira Neto FL. Satellite glial cells in sensory ganglia: its role in pain. Rev Bras Anestesiol. 2015;65(1):73–81. doi: 10.1016/j.bjan.2013.07.013. [DOI] [PubMed] [Google Scholar]
- Malin SA, Davis BM, Molliver DC. Production of dissociated sensory neuron cultures and considerations for their use in studying neuronal function and plasticity. Nat Protoc. 2007;2(1):152–160. doi: 10.1038/nprot.2006.461. [DOI] [PubMed] [Google Scholar]
- Lin YT, Ro LS, Wang HL, Chen JC. Up-regulation of dorsal root ganglia BDNF and trkB receptor in inflammatory pain: an in vivo and in vitro study. J Neuroinflammation. 2011;8:126. doi: 10.1186/1742-2094-8-126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liem L, van Dongen E, Huygen FJ, Staats P, Kramer J. The Dorsal Root Ganglion as a Therapeutic Target for Chronic Pain. Reg Anesth Pain Med. 2016;41(4):511–519. doi: 10.1097/AAP.0000000000000408. [DOI] [PubMed] [Google Scholar]
- Lee JS, Han JS, Lee K, Bang J, Lee H. The peripheral and central mechanisms underlying itch. BMB Rep. 2016;49(9):474–487. doi: 10.5483/BMBRep.2016.49.9.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valtcheva MV, et al. Surgical extraction of human dorsal root ganglia from organ donors and preparation of primary sensory neuron cultures. Nat Protoc. 2016;11(10):1877–1888. doi: 10.1038/nprot.2016.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melli G, Hoke A. Dorsal Root Ganglia Sensory Neuronal Cultures: a tool for drug discovery for peripheral neuropathies. Expert Opin Drug Discov. 2009;4(10):1035–1045. doi: 10.1517/17460440903266829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin K, Baillie GJ, Vetter I. Neuronal cell lines as model dorsal root ganglion neurons: A transcriptomic comparison. Mol Pain. 2016;12 doi: 10.1177/1744806916646111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen W, Mi R, Haughey N, Oz M, Hoke A. Immortalization and characterization of a nociceptive dorsal root ganglion sensory neuronal line. J Peripher Nerv Syst. 2007;12(2):121–130. doi: 10.1111/j.1529-8027.2007.00131.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doran C, Chetrit J, Holley MC, Grundy D, Nassar MA. Mouse DRG Cell Line with Properties of Nociceptors. PLoS One. 2015;10(6):e0128670. doi: 10.1371/journal.pone.0128670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gouarderes C, Roumy M, Advokat C, Jhamandas K, Zajac JM. Dual localization of neuropeptide FF receptors in the rat dorsal horn. Synapse. 2000;35(1):45–52. doi: 10.1002/(SICI)1098-2396(200001)35:1<45::AID-SYN6>3.0.CO;2-0. [DOI] [PubMed] [Google Scholar]
- Lin YT, et al. Activation of NPFFR2 leads to hyperalgesia through the spinal inflammatory mediator CGRP in mice. Exp Neurol. 2017;291:62–73. doi: 10.1016/j.expneurol.2017.02.003. [DOI] [PubMed] [Google Scholar]
- Yang HY, Tao T, Iadarola MJ. Modulatory role of neuropeptide FF system in nociception and opiate analgesia. Neuropeptides. 2008;42(1):1–18. doi: 10.1016/j.npep.2007.06.004. [DOI] [PubMed] [Google Scholar]
- Takeda M, Takahashi M, Matsumoto S. Contribution of the activation of satellite glia in sensory ganglia to pathological pain. Neurosci Biobehav Rev. 2009;33(6):784–792. doi: 10.1016/j.neubiorev.2008.12.005. [DOI] [PubMed] [Google Scholar]