

Abstract
Steroid hormone receptors are important regulators of development and physiology in bilaterian animals, but the role of steroid signaling in cnidarians has been contentious. Cnidarians produce steroids, including A-ring aromatic steroids with a side-chain, but these are probably made through pathways different than the one used by vertebrates to make their A-ring aromatic steroids. Here we present comparative genomic analyses indicating the presence of a previously undescribed nuclear receptor family within medusozoan cnidarians, that we propose to call NR3E. This family predates the diversification of ERR/ER/SR in bilaterians, indicating that the first NR3 evolved in the common ancestor of the placozoan and cnidarian-bilaterian with lineage-specific loss in the anthozoans, even though multiple species in this lineage have been shown to produce aromatic steroids, whose function remain unclear. We discovered serendipitously that a cytoplasmic factor within epidermal cells of transgenic Hydra vulgaris can trigger the nuclear translocation of heterologously expressed human ERα. This led us to hypothesize that aromatic steroids may also be present in the medusozoan cnidarian lineage, which includes Hydra, and may explain the translocation of human ERα. Docking experiments with paraestrol A, a cnidarian A-ring aromatic steroid, into the ligand-binding pocket of Hydra NR3E indicates that, if an aromatic steroid is indeed the true ligand, which remains to be demonstrated, it would bind to the pocket through a partially distinct mechanism from the manner in which estradiol binds to vertebrate ER.
Keywords: A-ring aromatic steroid, aromatization, steroid receptor, cnidarian
1. Introduction
Nuclear receptors are important regulators of life-history transitions in various phyla of bilaterian animals. They are well documented ligand-activated transcription factors in vertebrates, arthropods and nematodes. However, their role is largely unknown in other metazoan groups such as molluscs or annelids [1], In cnidarians, following initial identification through PCR screens [2, 3], some nuclear receptors have been implicated as regulators of nervous system development [4, 5] and one of them was recently shown to be involved in the molecular cascade triggering the polyp-to-jellyfish transition in the scyphozoan medusa Aurelia aurita [6], However, the endogenous ligands, if any, for these cnidarian nuclear receptors are still unclear.
Strictly speaking, cnidarians do not have a bilaterian-like endocrine system, with signaling molecules transported by means of an internally circulating body fluid [7]. However, they do regulate their physiology in response to nutritional and environmental inputs, which also makes them sensitive to environmental pollution, and diffusible signaling molecules are involved in such processes [8, 9].
Historically, numerous efforts have been made to search for vertebrate-like signaling features in cnidarians, especially components of the estrogen signaling pathway, which, in vertebrates, is primarily mediated through the estrogen nuclear receptor belonging to the NR3 family. In particular, aromatization, the final step in the synthesis of estrogens from testosterone (Figure 1A), has been studied in multiple species. Early attempts to detect aromatase activity in cnidarian tissues (scleractinian corals and octocorals), using vertebrate precursors, were unsuccessful [10–12], but subsequently, aromatization activity has been reported in anemones and scleractinian corals [13–15]. Aromatization of exogenous androgen precursors by tissue homogenates from cnidarians or molluscs has been interpreted as evidence for the ability to endogenously synthesize vertebrate-type estrogens [16]. However, analyses of genomic data indicate that cnidarians cannot synthesize estrogens through the same pathways as vertebrates because essential components are lacking in cnidarians. Specifically the CYP19 aromatase is chordate-specific, and the CYP11A1 enzyme responsible for the early cholesterol side-chain cleavage is vertebrate-specific [17]. Moreover, there is evidence that corals can take up vertebrate steroids from the water column, especially around human polluted sites, so estrogen-like compounds detected in cnidarian tissues may have exogenous origins [18, 19]. These observations are also in line with the reinterpretation of the presence of vertebrate-type steroids in molluscs [20, 21]. Arthropod-like ecdysteroids and nematode-like dafachronic acids have also been isolated from corals [22, 23], but have not been viewed as evidence for ecdysozoan-type hormonal signaling in these cnidarians. In case of ecdysteroids, a defensive role and a dietary origin are the main interpretations that have been used to explain the presence and high abundance of those compounds [22]. Similarly, the presence and activity of gonadotropin-like neuropeptides in cnidarians have been used to draw parallels between vertebrate and cnidarian reproductive signaling cascades [24]. However, here again, phylogenetic analyses indicate that the cnidarian peptides are equally similar to arthropod neuropeptides that do not act in reproductive signaling [25].
Figure 1. A model for A-ring aromatic steroid synthesis pathway in the octocoral cnidarian Dendronephthya studeri, based on the molecules described in Yan et al., 2011, and the knowledge of enzymatic pathways known from other organisms.
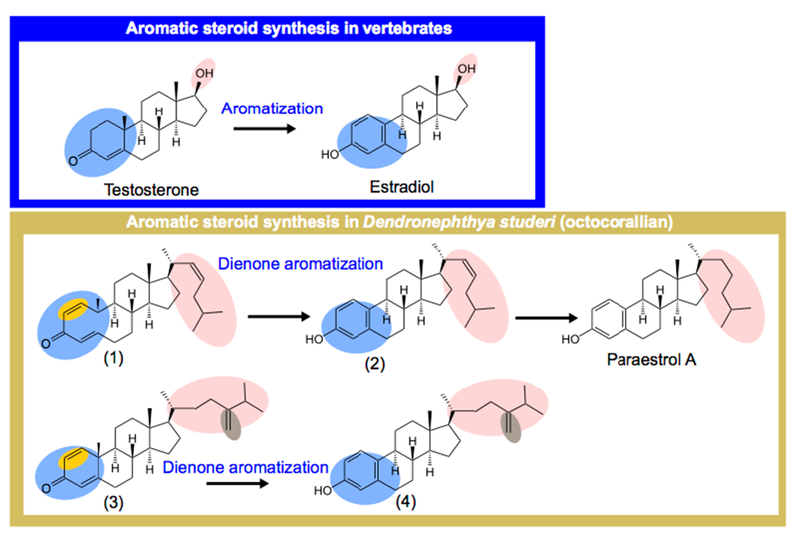
(1): Cholesta-1,4,22-trien-3-one; (2): (22E)-19-Norcholesta-1,3,5(10),22-tetraen-3-ol; (3): 24-Methylenecholesta-1,4,22-trien-3-one; (4): 24-Methylene-19-norcholesta-1,3,5(10),22-tetraen-3-ol. Paraestrol A (19-norcholesta-1,3,5(10)-trien-3-ol) has been proposed as an ancestral steroid [28]. We have named « dienone aromatization » the proposed aromatization reaction to stress the difference from the vertebrate case where there is no delta1–2 double bond (highlighted in yellow) on the A ring.
To date, ten distinct A-ring aromatized steroids have been identified from four different octocoral species belonging to different genera (Alcyonium gracillimum, Capnella sp., Dendronephtya studeri, Scleronephthya pallida). Among these steroids are aromatized pregnanes and aromatized C9-C10 secosteroids, which are compounds that have core structures similar to progesterone and vitamin D, respectively [26]. A third group consists of A-ring aromatized steroids bearing a variety of side chains. These have been found in the Taiwanese octocoral Dendronephthya studeri [27], see also Figure 1. Based on the co-occurrence of A-ring aromatic steroids and corresponding dienones (see compounds (1) and (3) on Figure 1), that bear an additional unsaturation (in yellow on Figure 1) compared to testosterone, we hypothesize that the aromatization reaction in those corals is distinct from that in vertebrates, as proposed for the two other groups [26].
Among the steroids present in Dendronephthya studeri, one compound has an identical structure to what we hypothesize to be the ancestral NR3 ligand in chordates. This compound indeed binds an ancestral steroid receptor with low affinity [28]. Due to the combination of data discussed above, we have previously interpreted paraestrol A (19-norcholesta-1,3,5(10)-trien-3-ol) and other aromatized steroids from corals as defensive compounds [28]. The diversity of steroids in cnidarians led us to reevaluate the distribution and potential signaling functions of NR3 in the phylum Cnidaria. Our data suggest that NR3 orthologs are present in three of the four major cnidarian lineages and that an endogenous ligand may be present based on heterologous expression of vertebrate ERα in Hydra. Together, our data suggest that some ring-A aromatic steroids might function as endogenous signaling molecules in cnidarians, particularly paraestrol A that has the potential to act as a ligand for cnidarian NR3 receptors.
2. Material and Methods
2.1. Sequence analysis
Cnidarian NR3 sequences were identified by tBLASTn and reciprocal BLASTp queries at NCBI GenBank using human ERα and Trichoplax adhaerens NR3 (previously identified [29]). In addition to NCBI data, putative NHRs were identified by tBLASTn searches of the de novo assembled transcriptomes of Aurelia, Morbakka, Chironex, Tripedalia and Copula (Khalturin, unpublished data). We required that identified sequences had to be complete or nearly complete predicted nuclear receptor proteins (i.e., both DNAand ligand binding domains) to facilitate phylogenetic analyses. All sequences were downloaded and, if necessary, translated into the correct reading frame. Collected sequences were aligned using Clustal Omega [30] and alignments were checked by eye and edited with Seaview [31], A phylogenetic tree (Figure 2) was made using PHYML [32], a fast and accurate maximum likelihood heuristic method, using the LG model [33] with a gamma law. Reliability of nodes was assessed by likelihood-ratio test [34],
Figure 2. A new NR3E family at the base of the bilaterian ER/ERR/SR clade.
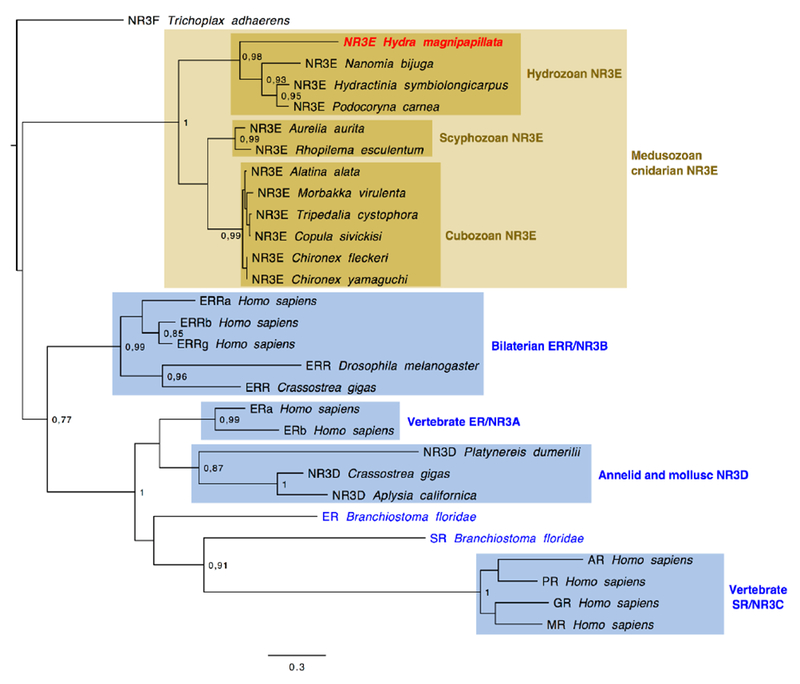
The tree was calculated using maximum-likelihood. Likelihood ratio test branch support values upper than 0.70 are indicated. Bilaterian sequences are shown in blue and cnidarian sequences in brown. The Hydra NR3E is highlighted in red. Mollusc and annelid NR3Ds correspond to proteins that were previously labelled as ERs [46], Similar analyses using the LBD alone gave largely similar results and did not affect the placement of the cnidarian nuclear receptors.
2.2. Heterologous hERα-GFP expression in Hydra
Nucleotide sequence of human estrogen receptor (hERα) was adjusted to fit the codon usage of Hydra vulgaris with the average GC content reduced to 33%. A synthetic gene (shERα) produced by GeneArt (Regensburg, Germany) was inserted into Hydra expression vector (ligAC-6) between the actin promoter and EFGP coding region using the Sbfl and PacI restriction sites (see Figure 3a). The resulting plasmid ligAL-5 was propagated in E. coli and sequenced. DNA for microinjection was prepared using the Plasmid Midi Kit (QIAGEN). Construct DNA was microinjected into fertilized eggs of Hydra vulgaris strain AEP as described previously [35], In about 70% of injected embryos EGFP-positive nuclei were detectable 48 hours after microinjection. Polyps started to hatch 2–3 weeks after microinjection and contained small patches of ectodermal epithelial cells with clear nuclear localisation of the shERα-EFGP protein. Polyps were cultivated under intensive feeding conditions and transgenic patches were enriched by selection of EGFP-positive buds and removal of non-transgenic areas. Within 3 months of selection a polyp culture where −80% of ectodermal cells were transgenic was obtained (see Figure 3).
Figure 3. An endogenous cytoplasmic factor induces hERα-GFP translocation from cytoplasm to nucleus in Hydra epithelial cells.
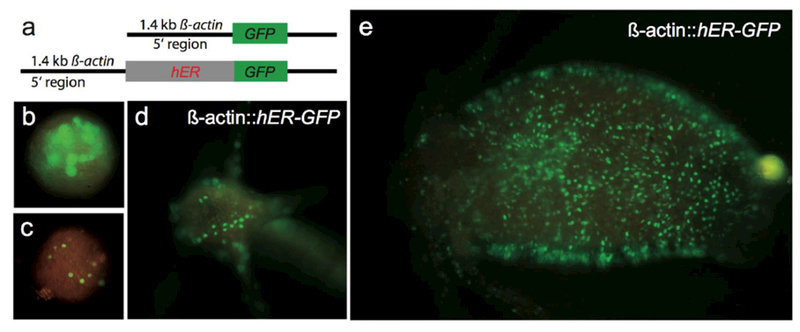
a) Schematic representation of transgenic constructs with GFP only and with human ERα-GFP driven by 1.4 kb of Hydra ß-actin promoter, b) One week old Hydra embryo injected with the control construct. GFP positive blastomeres are visible, c) Hydra embryo injected with actin::ERα-GFP construct. Several GFP positive nuclei are visible and the cytoplasm of blastomeres is not GFP positive, d) Head of transgenic Hydra polyp, e) Body column of a transgenic polyp. Green spots are the nuclei of ectodermal epithelia cells which are filled with hERα-GFP without addition of any ligand.
2.3. Homology modeling of NR3E andparaestrol A docking
Homology modeling was performed using the program Modeller [36] (version 9.18). The selected structural templates for the homology modeling were the crystal structure of ERα bound to 17β-estradiol (PDB id: 1ERE) [37], ERRγ in the constitutively active conformation (PDB id: 1KV6) [38] and apo ERRα in its active conformation (PDB id: 3D24) [39], Missing loops in the ERα structure, namely between residues 331–336 and 462–464, were built prior to homology modeling [40], Sequence alignment of target and template was performed using the align2d procedure in Modeller, taking into account structural information from the template when constructing the alignment, resulting in 26.2, 29.5 and 25.8% of sequence identity for the modeling using ERα, ERRγ and ERRα respectively. A total of 50 models were generated and the structure with the best DOPE score [41], a statistical potential optimized for model assessment in Modeller, was selected for ligand docking. Examination of the best homology models based on ERα, ERRγ and ERRα revealed that the C-alpha traces nicely follow their respective crystal structure templates with an RMSD of 0.172, 0.289 and 0.293Ά respectively after sequence-structure superposition using Pymol software and with 96.48, 95.87 and 96.36% of residues estimated in the Ramachandran favored space using Molprobity server [42], We noticed that homology modelling of NR3E LBD based on the crystal structure of ERRγ, led to a slight local deformation in a helix turn belonging to helix H10. This is to maintain the structure/sequence conservation in the rest of the model, but this does not affect the ligand-binding pocket and thus does not impact the docking results.
Docking was performed with Autodock4 [43] using the Lamarckian genetic algorithm and consisted of 10 runs per search, with a maximum of 2.5 million energy evaluation per run and a population size of 150. Affinity maps for the receptor were computed using Autogrid4 prior to docking, using the default grid map spacing of 0.375Ά. Paraestrol A was superimposed on the 17β-estradiol ligand in the 1ERE structure, and the homology models on the ERα protein in the same structure. Based on this, residues in the models displaying steric clashes were selected as flexible residues in the docking procedure. In order to compare the estimated free energies of binding, a docking run was performed using the 17β-estradiol ligand in the ERα crystal structure with the same docking parameters. Interaction diagram was drawn using LIGPLOT v.4.5.3 [44], All structure figures were prepared using PyMOL (The PyMOL Molecular Graphics System, Version 1.7.4.0 , Schrödinger LLC).
3. Results
3.1. NR3E is a novel nuclear receptor type present in three of the four main cnidarian lineages
We identified new nuclear receptor NR3 sequences from cnidarians that group together as a separate family at the base of the bilaterian NR3s, suggesting they are not particularly related to one of the bilaterian NR3 families (i.e., ER, ERR, or SR). This is also true for the Trichoplax sequence initially described as an ERR [29]. We have previously suggested, based on the uncertainty in the phylogenetic relationships between vertebrate ER and sequences from molluscs and annelids described as ERs to rename the protostome clade as NR3D, to avoid misleading inferences that estrogen would be their natural physiological ligand [45]. The most recent analyses of bilaterian NR3s, based on extensive sampling, are in agreement with this view, putting the molluscan and annelid NR3Ds at the base of a clade grouping the vertebrate NR3A and NR3C [46, 47]. Here, NR3A and NR3D group together in a weakly supported node, so we do not think this grouping contradicts these previous studies. Most importantly, the cnidarian sequences follow the known subdivisions between the three clades in which they are present : cubozoans, scyphozoans and hydrozoans (Figure 3). Therefore we propose to name this clade as NR3E. For the same reason, we propose that the Trichoplax ERR should be renamed NR3F.
3.2. Heterologous Human ERα-GFP construct translocates into the Hydra cytoplasm
In the context of functional studies based on transgenesis in Hydra [35], we made an inducible reporter system for Hydra based on human ERα. This was based on the assumption that the vertebrate estrogen signaling system does not naturally function within Hydra. The approach was to make transgenic animals where the translocation of transcription factors into the nucleus is controlled by addition of estrogen (in the construct hERα would be fused to the gene of interest). Initially, a construct was made containing hERα fused to green fluorescent protein (hERα-GFP), which theoretically should stay in the cytoplasm in the absence of exogenous estrogen. Surprisingly, hERα-GFP was translocated to the nucleus in Hydra even without addition of estrogen (Figure 3). At the same time pure GFP protein never concentrates in the nuclei of transgenic Hydra.
There might be several explanations for the translocation of the hERα-GFP construct. For example, hsp90 and hsp70 of Hydra may not bind hERα-GFP properly and thus ERa would go into the nucleus by default. However, the high level of conservation between human and cnidarian Hsp70/90 speaks against “default” translocation. Another plausible scenario is the presence of a cytoplasmic factor in Hydra which can specifically bind the ligand-binding pocket of the human estrogen receptor and thus trigger nuclear translocation.
3.3. Docking of paraestrol A inside the ligand-binding pocket of ERα-, ERRγ- and ERRα-based homology models of NR3E
Docking experiments were performed to assess the potential binding of paraestrol A to Hydra NR3E. This was done using homology models for Hydra NR3E that were developed using the crystal structures of ERα, ERRγ and ERRα as templates. For comparison, we employed the same parameters (see Materials and Methods) for docking the 17β-estradiol in the ERα crystal structure, estimated the corresponding binding free energy and compared it to that of paraestrol A in the different homology models. In the case of 17β-estradiol in ERα, the conformation obtained for 17β-estradiol was nearly identical to the one observed in the crystal structure (rmsd of 0.7Å) and the calculated binding free energy was estimated to be −10.32 kcal/mol.
Considering the Hydra NR3E ERα-based homology model, superimposition of paraestrol A onto the steroid core of 17β-estradiol in the ERα-based homology model shows that paraestrol A fits inside the ligand-binding pocket without any steric clash with the surrounding residues. As a consequence, the initial conformations of the latter residues were maintained throughout the whole docking procedure. The best docking pose obtained had a binding free energy of −11.28 kcal/mol after 10 runs of docking attempts. These observations together with a comparison of the free energy with that of 17β-estradiol in ERα suggest that paraestrol A would nicely fit inside the ligand-binding pocket of the ERα-based homology model with interactions as described below.
For the ERRγ-based model, ligand superimposition of paraestrol A onto the steroid core of 17β-estradiol indicates that residues W80 and F35 are in steric clash with the ligand. We therefore considered them as flexible in the docking procedure, while the conformation of the other residues inside the binding cavity was kept as modeled initially. The binding free energy of the best docking pose was estimated to be −9.38 kcal/mol. As in the case of the ERα-based homology model, the results suggest that paraestrol A could be well-accommodated inside the ligand-binding pocket of the ERRγ-based homology model of Hydra NR3E LBD with an interaction pattern rather similar to the one observed for the docking in the ERα-based homology model
We further applied the same procedure to the ERRα-based homology model and observed that several residues, especially aromatic ones, were in steric clash with the paraestrol A initial conformation. Despite the fact that the residues in steric clash were allowed to be flexible in the docking procedure, no reasonable docking pose was obtained, suggesting that successful docking of paraestrol A strongly depends on the nature of the homology model used and further supports the validity of the docking predictions obtained in the ERα- and ERRγ-based homology models.
3.4. Interactions ofparaestrol A with residues inside the ligand-binding pocket
In the ERα-based homology model, paraestrol A is globally positioned in a similar manner as is 17β-estradiol in its cognate receptor (Figure 4a). In fact, the aliphatic side chain of paraestrol A is oriented towards the helices H7 and Hll, while the steroid core is positioned between helices H3 and H5 and rather close to the first strand of the β-sheet (Figure 4a). Several hydrophobic residues, including aromatic ones form stabilizing interactions with the ligand. In addition, a remarkable π-π interaction is observed between Trp80 in helix H5 and the steroid core (Supplemental Figure 1); this residue and the corresponding stabilizing interaction are specific to NR3E (Figure 5).
Figure 4. Paraestrol A docking into the ligand binding pocket of an estrogen receptor a homology-based model of Hydra NR3E.
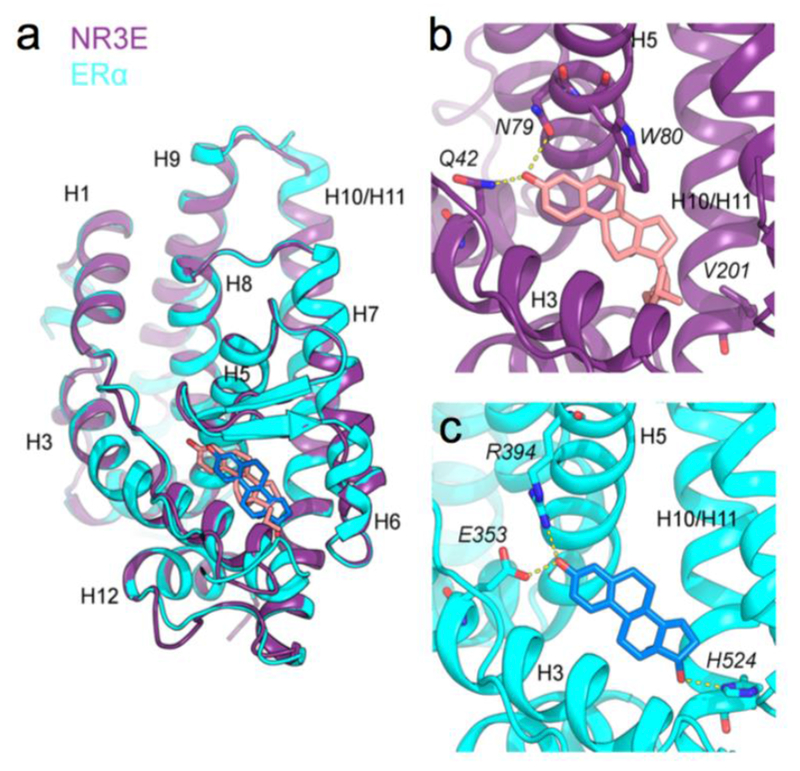
(a) Superposition of the NR3E structural model using ERα structure (in violet) where the paraestrol A is represented in pink, the ERα structure used as template (PDB id: 1ERE) is represented in cyan and the 17β-estradiol in blue, (b) Detailed view of the binding pocket of the NR3E ERα-based model, highlighting interactions (in yellow dashed lines) between the paraestrol and residues Q42 and N79. W80 involved in the cnidarian-specific π-π interaction is also shown, (c) Detailed view of the binding pocket of ERα, highlighting interactions (in yellow dashed lines) between the 17β-estradiol and residues E353, R394 and H524 (corresponding to V201 in NR3E model).
Figure 5. Alignment of H3, H4-H5 and H10-H11 helices in the NR3 family, compared to NR3E from Hydra.
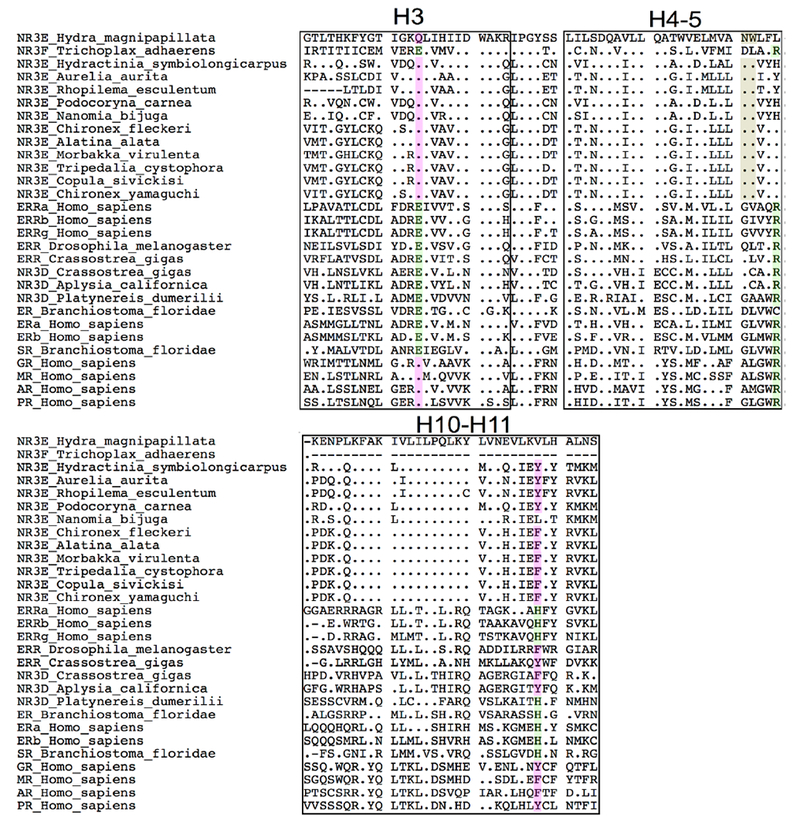
Residues involved in estrogen binding in ER are highlighted in green. Homologous residues that are shared by the cnidarian NR3E and the vertebrate oxosteroid receptors (SRs) are highlighted in pink. The cnidarian-specific N79-W80 anchor is highlighted in brown. An alignment for the entire DNA-binding domain and ligand-binding domain is shown in Supplemental Figure 2. The helices from the ligand-binding domain are mapped according to the structure of human ERα [48], Calculations of identity percentages for both domains are also provided in Supplemental Table1.
We further observe that the 3-OH group of paraestrol A is hydrogen-bonded to Asn79 (H5) with a donor-acceptor distance of 2.9Å, and examination of the surroundings of this interaction shows a potential additional hydrogen bond with Gln42 (H3) (Supplemental Figure 1). Both residues could form a clamp that would strongly position the ligand inside the pocket. To test this hypothesis, we started with the paraestrol A docked conformation in the ERα-based model and performed additional docking runs where both Gln42 and Asn79 were allowed to be flexible. Interestingly, the best docked conformation in this case showed structural adaptation of these two residues, which are then both implicated in hydrogen bond formation with the 3-OH group of paraestrol A (Figure 4b), with estimated binding free energy of −11.89 kcal/mol, and a similar π-π interaction with Trp80 in H5 as in the previous docking result.
The relevance of these observed interactions is emphasized by the remarkable conservation of residues W80, Q42 andN79 in cnidarian NR3 sequences (Figure 5). In fact, W80 is strictly conserved and specific to the cnidarian NR3E, while being replaced by M/L/V/I/C7A in bilaterian steroid receptors and ERRs as well as in Trichoplax NR3F. Furthermore, the anchoring residues Q42 and N79 are strictly conserved in all cnidarian NR3E sequences. Q42 is also present in human oxosteroid receptors, but replaced by E in human ERs and ERRs, as well as in Trichoplax NR3F. N79 is also lineage-specific, being strictly conserved in cnidarian NR3E, and replaced by G/D/A/Q/N in bilaterian steroid receptors and in Trichoplax NR3F. In human ERα bound to 17β-estradiol, the 3-OH group is stabilized by an electrostatic anchor formed by two conserved residues E353 (H3) and R394 (H5) (Figure 4c). These residues are crucial for the positioning and the stabilization of the natural 17β-estradiol ligand inside its cognate ligand-binding pocket. In the case of cnidarian NR3E, the anchor is made of Q42 and N79. In this case, the 3-OH group of paraestrol A acts both as an acceptor and a donor for these residues. An inspection of the multiple sequence alignment together with the superimposition of the homology model with the crystal structure indicate that Q42 in H3 is the cnidarian equivalent of human E353 that forms the first half of the electrostatic anchor. On the other hand, the second half of the anchor differs in the two cases. Residue N79 of Hydra NR3E is located one helix turn upstream of the position occupied by R394 and replaced by G in human ERα (Figure 4b and 4c). Altogether, we observed that the 3-OH functional group of paraestrol A is in an adequate environment to form complementary interactions with the conserved electrostatic Q(H3)/N(H5) anchor and further stabilized by π-π interaction with the cnidarian-specific W80 residue located in H5.
It is noteworthy that in our second set of docking experiments, the two best docked structures display hydrogen-bonding with Q42 and N79, however in the third one, the distance between 3-OH group of paraestrol A and the amide group of Q42 increases to 4.8 Å, leading to a severe displacement of the ligand in the pocket, with the aliphatic side chain replacing the position of the aromatized A-ring. This structure supports the crucial role of the anchor in positioning of the ligand in the pocket, as strongly suggested by the sequence conservation of these residues.
The positioning of paraestrol A inside the ligand-binding pocket of the ERRγ-based homology model is similar as is its general interaction pattern with surrounding residues in the best docked conformation with an estimated binding free energy of −9.38 kcal/mol. Even though the exact position of paraestrol A is slightly more towards the outside of the pocket and slightly shifted between helices H3 and H5, W80 still can form a π-π interaction and a hydrogen bond with Q(H3) (Supplemental Figures 3a and 3b). We then performed a similar docking experiment where this latter structure was used and both residues Q42 and N79 are allowed to be flexible in the search. Similarly to the previous results, adjustment of the side chains of these residues permits in the best model hydrogen bonding with the 3-OH group of the paraestrol, leading to a more favorable binding free energy of −11.50 kcal/mol (Supplemental Figure 3b). The three conserved residues that were shown to be crucial for the interaction of the ligand with the ERα-based homology model hence fulfill the same role in the ERRγ-based homology model.
Altogether, our docking studies of paraestrol Ain NR3E homology models suggest that paraestrol A can be readily accommodated in the ERα-based model and depicts a binding free energy that is of the same order of magnitude as the one calculated for 17β-estradiol in ERα. Paraestrol A can also fit inside the ligand-binding pocket of ERRγ, with comparable energy. In both cases, we have highlighted cnidarian-specific residues that interact with the ligand, among which two polar residues, Q(H3)/N(H5), that define a novel anchor that stabilizes paraestrol A inside the pocket.
4. Discussion
4.1. Novel cnidarian-specific candidate players for a steroid signaling pathway
All evidence to date has shown that the vertebrate-type estrogen-ER signaling pathway is not present in cnidarians [28], However, recent findings, in the literature and the data presented in this study indicate the presence both of aromatic steroids and nuclear receptors within a novel subclade of the NR3 family (NR3E). Previous surveys of nuclear receptor diversity in cnidarian have largely studied anthozoans due to availability of sequence data as well as interest in possible effects of environmental pollution on reef-building corals. Our analysis here used the availability of sequence data from throughout the phylum and revealed NR3 members in the other three cnidarian classes that form the medusozoan clade. The presence of these genes helps to better resolve the history of the NR3 family in animals because NR3 was present in the placozoan Trichoplax, absent from surveyed ctenophores and sponges, and until this study, inferred to absent from cnidarians. Our hypothesis is thus that NR3 evolved in the common ancestor of the placozoan and cnidarian-bilaterian with lineage-specific loss in the anthozoans.
Despite the presence of NR3 genes in cnidarians, it has not yet been demonstrated whether these aromatic steroids or any other compounds can serve as ligands for NR3E receptors. Interestingly, aromatic steroids have only been documented within a few species of anthozoans but NR3E genes are only present within the medusozoa (Hydrozoa, Cubozoa, Scyphozoa). Similarly, NR2B (RXR) homologs have been lost from the Anthozoa and are only present within Medusozoa. The loss of these two receptors from the anthozoan lineage is of considerable interest and may be related to the loss of the medusa (jellyfish) stage. This step implied the loss of metamorphosis between polyp and medusa, and/or the reduction of sensory organs (eyes). Regarding aromatic steroids in anthozoans, data about their quantitative variation under physiological conditions will be necessary to determine if they play a defensive function, or if they may be involved in endogenous intercellular signaling. If they do play any role in endogenous signaling, it could be mediated through binding to different nuclear receptors from other families than NR3, or to membrane receptors [49], The translocation data on the shERα-GFP heterologous construct expressed in Hydra epithelial cells suggests that there may be a cytoplasmic factor that binds to shERα-GFP and triggers its translocation into the nucleus. Because Hydra, an hydrozoan, as other medusozoans, has an endogenous NR3E, which is the closest relative to ER and other vertebrate steroid receptors, we thought it would be important to explore its ability to bind an aromatic steroid which is present in another cnidarian, even from a distantly related species.
4.2. Cnidarian-specific molecular mechanisms for ligand binding in the NR3E pocket
Docking experiments with paraestrol A inside the ligand-binding pocket of Hydra NR3E indicate that this receptor might indeed bind an aromatic steroid. In this case, the binding would involve π-π interactions with a tryptophan residue, which is conserved in all cnidarian sequences, and a glutamine residue which, to date, was mainly correlated to the binding of oxosteroids, like progesterone, androgens and corticoids. Thus, even the most estrogen-like aromatic steroid from cnidarians shows binding properties that are distinct from the mechanism of estradiol binding to the vertebrate estrogen receptor. This strongly suggests that identification of endogenous ligands for the cnidarian NR3E receptors and their concomitant functional characterization will enlarge our knowledge of the diversity ligand-binding interactions in play between steroid hormones and their cognate nuclear receptors. Over the last few years, a great deal of information has accumulated about physiologically relevant alternative ligands for the human estrogen receptors [50], In particular, steroids without an aromatic A-ring were also reported to be able to bind ER. So far, all those described alternative steroid ligands have a hydroxy group on carbon 3, just like the hydroxy group on carbon 3 of estradiol. Interestingly, the distinction between binding of steroids with a 3-hydroxy group, like estrogens, and binding of steroid with a 3-oxo group, like androgens, progesterone and corticoids, has long been correlated to the distinction between the E and Q ligand-binding residues in the H3 part of the ligand-binding pocket, and interpreted as an evidence for more ancient origin of estrogen as a ligand relative to oxosteroids [51]. Docking experiments with NR3E and paraestrol A indicate that, structurally speaking, nothing prevents a 3-hydroxysteroid from binding the pocket through a Q residue, which is classically related to 3-oxosteroid binding. Therefore, the distinction between the two classes of molecules may not be so clear-cut, and even in vertebrates, we could find unexpected new selective estrogen receptor modulators by looking at endogenous 3-oxosteroids. Many of them are indeed present in the human body as precursors in bile acid synthesis [52].
4.3. Long distance chemical communication in the cnidarian body
Because they lack an internal circulating body fluid, cnidarians cannot have a vertebrate-like hormonal system. This does not necessarily preclude steroid-mediated intercellular signaling in those animals. Indeed, even in vertebrates, some steroids do not act through the canonical hormonal pathway. Classically, hormones are defined as internally circulating molecules, which excludes vertebrate bile acids based on this definition [53]. However, this view is changing with the acknowledgment of their functional role not only as facilitators of lipid digestion, but also as signaling molecules, that allow the coupling of the nutritional state with various parameters, from digestive physiology to behavioural traits such as the regulation of appetite [54]. This example shows that there is no objective reason to strictly limit long-distance intercellular communication to the internal milieu. We thus hypothesize that in cnidarians, the digestive surfaces may be a major vehicle for signaling molecules, and that the steroids identified in these animals, if they really have a role in distant intercellular communication, could be transported in this way. The gut lumen is sometimes viewed as the continuation of the external milieu, because both are communicating through the oral openings. Thus, secretions of molecules in the gut are considered as exocrine secretions. The real chemical composition of some parts of the digestive tract is distinct from the external environment, including secretions and selective absorption processes from the digestive surfaces of the animal. The gut goes throughout the body for some cnidarians and is restricted to particular regions in others (e.g. mesenteries) and could be a carrier of signaling molecules between distant body parts or different ramets in colonial species (Figure 6). Anyways, caution is needed before extrapolating anatomical knowledge from bilaterians to cnidarians, because the homology between the germ layers is debated [55]
Figure 6. A possible pathway for steroid enterocrine signaling in cnidarians.
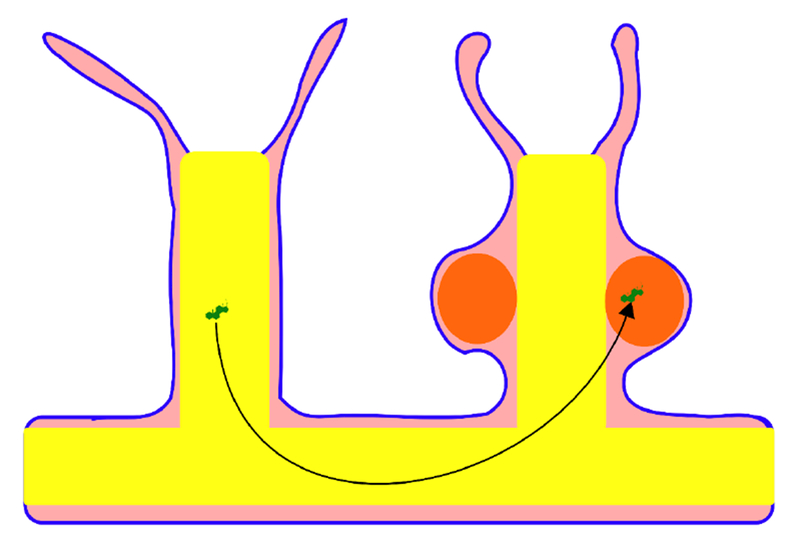
Steroids, in green, could be produced from dietary sterols in the gut (in yellow) and go to the gonad (in orange) along with other nutrients. Gonads may be of ectodermal or endodermal origin, depending on the species. The pink tissue represents the mesoglea, which consists of a gelatinous matrix that contains collagen fibers and usually some cells. The mesoglea forms a hydrostatic skeleton, but does not contain a circulating body fluid or play a known role in circulation.
Regulation of reproduction and metamorphosis have already been proposed as potential roles for cnidarian nuclear hormone receptors. Regulation of energetic metabolism and growth is another prominent function of many nuclear receptors that has not yet been investigated in cnidarians. Indeed, within the NR3 family, ERR plays important and diverse roles in energetic homeostasis in both protostomes and deuterostomes [56, 57]. In cnidarians, regulation of energetic metabolism is of considerable interest, but the mechanisms are still poorly understood. Many species are tightly regulating their body size based on nutrition intake. For example, many anthozoan anemone species can survive several months without food, during which they can exhibit negative growth, consume stored lipids and reduce their metabolic rates [58–61]. Studies with the hydrozoan Hydra suggest an important role of autophagy during starvation. Starving hydras do not die, but simply shrink proportionally [62]. For both anemones and Hydra, growth rapidly resumes following feeding (references cited in two preceding sentences and personal observations of authors). Hence, there must be some regulators and nuclear receptors are among the best candidates.
5. Conclusions
While molecular studies of steroid signaling in cnidarians are still in early stages, the fragmentary data are already sufficient to state that, despite some similarities to bilaterians, cnidarians have lineage-specific receptors and potential ligands. A detailed look at the mechanisms of steroid signaling and receptor function is certain to give new insights on the many different possible ways to achieve interactions between a receptor and a steroid ligand. Obviously, this also holds true for other chemical classes of possible ligands. In comparison to anthozoans, very few chemical analyses have targeted the steroid composition of medusozoan cnidarians [63–65]. There is growing awareness among the global metabolomics community that « time is ripe to focus on model organism metabolomes » [66]. Along these lines, metabolomic approaches have already proven extremely useful in understanding phenotypic plasticity in a mediterranean zoanthid coral [67]. We hope that our results will stimulate interest of natural product chemists in identifying potential endogenous ligands for cnidarian nuclear receptors and particularly in determining whether A ring aromatic sterols are present in Hydra and other medusozoan cnidarians, as well as other molecules that could function as endogenous ligands to cnidarian nuclear receptors. This will not only illuminate the field of cnidarian zoology but, more widely, the field of nuclear receptor pharmacology to uncover new mechanisms of action and novel signaling molecules.
Supplementary Material
Highlights.
Cnidarians do not have vertebrate-type estrogen signaling.
A cytoplasmic factor triggers nuclear translocation of hERα-GFP in Hydra epithelium.
Medusozoan cnidarians have a specific type of steroid-related receptor, called NR3E.
Anthozoan cnidarians have lost the NR3E receptor but can produce aromatic steroids.
NR3E has a cnidarian-specific anchor that can bind to aromatic steroids in a novel way.
Acknowledgments
The authors wish to express their sincere thanks to Jorg Wittlieb (CAU, Kiel) for microinjection of transgenic construct into Hydra AEP embryos. KK is supported by grant from Japan Society for the Promotion of Science (JSPS 17K07420). I.M.L.B and Y.C. acknowledge the support and the use of resources of the French Infrastructure for Integrated Structural Biology FRISBI ANR-10-INBS-05 and of Instruct-ERIC. AMR was supported by NIH Award R15GM114740. AMT was supported by an Internal Research and Development Award from the Woods Hole Oceanographic Institution. The work from the VL laboratory is supported by CNRS, Sorbonne Université, ANSES and EMBRC-France. GVM is supported by the National Research Agency investment expenditure program IDEALG (ANR-10-BTBR-04) and by the Region Bretagne via the grant « SAD2016-METALG (9673) ».
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- [1].Handberg-Thorsager M, Gutierrez-Mazariegos J, Nadendla EK, Bertucci PY, Arold ST, Tomançak P, Pierzchalski K, Jones JW, Albalat R, Kane MA,Bourguet W, Laudet V, Arendt D, Schubert M, The ancestral retinoic acid receptor was a low-affinity sensor triggering neuronal differentiation, Sci Adv, 4 (2018) eaaol261. [Google Scholar]
- [2].Escriva H, Safi R, Hänni C, Langlois MC, Saumitou-Laprade P, Stehelin D, Capron A, Pierce R, Laudet V, Ligand binding was acquired during evolution of nuclear receptors, Proc Natl Acad Sci U S A, 94 (1997) 6803–6808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Grasso LC, Hayward DC, Trueman JW, Hardie KM, Janssens PA, Ball EE, The evolution of nuclear receptors: evidence from the coral Acropora, Mol Phylogenet Evol, 21 (2001) 93–102. [DOI] [PubMed] [Google Scholar]
- [4].Kostrouch Z, Kostrouchova M, Love W, Jannini E, Piatigorsky J, Rall JE, Retinoic acid X receptor in the diploblast, Tripedalia cystophora, Proc Natl Acad Sci U S A, 95 (1998) 13442–13447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Gauchat D, Escriva H, Miljkovic-Licina M, Chera S, Langlois M-C, Begue A, Laudet V, Galliot B, The orphan COUP-TF nuclear receptors are markers for neurogenesis from cnidarians to vertebrates, Dev Biol, 275 (2004) 104–123. [DOI] [PubMed] [Google Scholar]
- [6].Fuchs B, Wang W, Graspeuntner S, Li Y, Insua S, Herbst E-M, Dirksen P, Böhm A-M, Hemmrich G, Sommer F, Domazet-Lošo T, Klostermeier UC, Anton-Erxleben F, Rosenstiel P, Bosch TCG, Khalturin K, Regulation of polyp-to-jellyfish transition in Aurelia aurita, Curr Biol, 24 (2014) 263–273. [DOI] [PubMed] [Google Scholar]
- [7].Tarrant AM, Endocrine-like Signaling in Cnidarians: Current Understanding and Implications for Ecophysiology, Integr Comp Biol, 45 (2005) 201–214. [DOI] [PubMed] [Google Scholar]
- [8].Tarrant AM, Hormonal signaling in cnidarians: do we understand the pathways well enough to know whether they are being disrupted?, Ecotoxicology, 16 (2007) 5–13 [DOI] [PubMed] [Google Scholar]
- [9].Bosch TCG, Adamska M, Augustin R, Domazet-Loso T, Foret S, Fraune S, Funayama N, Grasis J, Hamada M, Hatta M, Hobmayer B, Kawai K, Klimovich A, Manuel M, Shinzato C, Technau U, Yum S, Miller DJ, How do environmental factors influence life cycles and development? An experimental framework for early-diverging metazoans. BioEssays, 36 (2014) 1185–1194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Gassman NJ, Kennedy CJ, Cytochrome P-450 content and xenobiotic metabolizing enzyme activities in the scleractinian coral, Favia fragum (Esper). Bull Mar Sci 50 (1992) 320–330. [Google Scholar]
- [11].Slattery M, Hines GA, Watts SA, Steroid metabolism in Antarctic soft corals. Polar Biol 18 (1997) 76–82. [Google Scholar]
- [12].Tarrant AM, Blomquist CH, Lima PH, Atkinson MJ, Atkinson S, Metabolism of estrogens and androgens by scleractinian corals, Comp Biochem Physiol B Biochem Mol Biol, 136 (2003) 473–485. [DOI] [PubMed] [Google Scholar]
- [13].Twan W-H, Hwang J-S, Chang C-F, Sex steroids in scleractinian coral, Euphyllia ancora: implication in mass spawning, Biol Reprod, 68 (2003) 2255–2260. [DOI] [PubMed] [Google Scholar]
- [14].Rougee LRA, Richmond RH, Collier AC, Molecular reproductive characteristics of the reef coral Pocillopora damicornis, Comp Biochem Physiol A Mol Integr Physiol, 189 (2015) 38–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Goodall CP, Heller I, Schwarz J, Weis VM, Lutterschmidt D, Chappell PE , Characterization of reproductive hormonal pathways in cnidarians, Endocrine Society’s 96th Annual Meeting and Exop, June 21–2104, Chicago: Abstract MON-0416. [Google Scholar]
- [16].Armoza-Zvuloni R, Kramarsky-Winter E, Loya Y, Schlesinger A, Rosenfeld H, Trioecy, a unique breeding strategy in the sea anemone Aiptasia diaphana and its association with sex steroids, Biol Reprod, 90 (2014) 122. [DOI] [PubMed] [Google Scholar]
- [17].Markov GV, Tavares R, Dauphin-Villemant C, Demeneix BA, Baker ME, Laudet V, Independent elaboration of steroid hormone signaling pathways in metazoans. Proc Natl Acad Sci U S A, 106 (2009) 11913–11918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Tarrant AM, Atkinson MJ, Atkinson S, Uptake of estrone from the water column by a coral community, Mar Biol 139 (2001) 321–325. [Google Scholar]
- [19].Armoza-Zvuloni R, Kramarsky-Winter E, Rosenfeld H, Shore LS, Segal R, Sharon D, Loya Y, Reproductive characteristics and steroid levels in the scleractinian coral Oculina patagonica inhabiting contaminated sites along the Israeli Mediterranean coast, Mar Pollut Bull, 64 (2012) 1556–1563. [DOI] [PubMed] [Google Scholar]
- [20].Scott AP, Do mollusks use vertebrate sex steroids as reproductive hormones? Part I: Critical appraisal of the evidence for the presence, biosynthesis and uptake of steroids, Steroids, 77 (2012)1450–1468. [DOI] [PubMed] [Google Scholar]
- [21].Schwarz TI, Katsiadaki I, Maskrey BH, Scott AP, Mussels (Mytilus spp.) display an ability for rapid and high capacity uptake of the vertebrate steroid, estradiol-17beta from water, J Steroid Biochem Mol Biol, 165 (2017) 407–420. [DOI] [PubMed] [Google Scholar]
- [22].Lafont R, Koolman J, Diversity of Ecdysteroids in Animal Species in Ecdysone: Structures and Functions, Smaaghe G (Ed.) Springer, (2009) 47–71. [Google Scholar]
- [23].Saini R,Boland S, Kataeva O,Schmidt AW, Kurzchalia TV, Knölker H-J, Org. Biomol. Chem, 2012, 10, 4159–4163. [DOI] [PubMed] [Google Scholar]
- [24].Twan W-H, Wu H-F, Hwang J-S, Lee Y-H, Chang C-F, Corals have already evolved the vertebrate-type hormone system in the sexual reproduction, Fish Physiol Biochem, 31 (2005) 111–115. [DOI] [PubMed] [Google Scholar]
- [25].Tsai P-S, Gonadotropin-releasing hormone by any other name would smell as sweet, Gen Comp Endocrinol, 2018, in press. [DOI] [PubMed] [Google Scholar]
- [26].Sarma NS, Krishna MS, Pasha SG, Rao TSP, Venkateswarlu Y, Parameswaran PS, Marine metabolites: the sterols of soft coral, Chem Rev, 109 (2009) 2803–2828. [DOI] [PubMed] [Google Scholar]
- [27].Yan X-H, Liu H-L, Huang H,; Li X-B, & Guo Y-W, Steroids with aromatic A-rings from the Hainan soft coral Dendronephthya staderi Ridley, J Nat Prod, 74 (2011) 175–180. [DOI] [PubMed] [Google Scholar]
- [28].Markov GV, Gutierrez-Mazariegos J, Pitrat D, Billas IML, Bonneton F, Moras D, Hasserodt J, Lecointre G, Laudet V, Origin of an ancient hormone/receptor couple revealed by resurrection of an ancestral estrogen, Sci Adv, 3 (2017) el601778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Baker ME, Trichoplax, the simplest known animal, contains an estrogen-related receptor but no estrogen receptor: Implications for estrogen receptor evolution, Biochem Biophys Res Commun, 375 (2008) 623–627. [DOI] [PubMed] [Google Scholar]
- [30].Sievers F, Higgins DG, Clustal Omega, accurate alignment of very large numbers of sequences, Methods in Molecular Biology, 1079 (2014) 105–116. [DOI] [PubMed] [Google Scholar]
- [31].Gouy M, Guindon S, Gascuel O, SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building, Mol Biol Evol, 27 (2010) 221–224. [DOI] [PubMed] [Google Scholar]
- [32].Guindon S, Gascuel O, A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst Biol, 52 (2003) 696–704. [DOI] [PubMed] [Google Scholar]
- [33].Le SQ, Gascuel O, Accounting for solvent accessibility and secondary structure in protein phylogenetics is clearly beneficial, Syst Biol, 59 (2010) 277–287. [DOI] [PubMed] [Google Scholar]
- [34].Anisimova M, Gascuel O, Approximate likelihood-ratio test for branches: A fast, accurate, and powerful alternative, Syst Biol, 55 (2006) 539–552. [DOI] [PubMed] [Google Scholar]
- [35].Wittlieb J, Khalturin K, Lohmann JU, Anton-Erxleben F, Bosch TCG, Transgenic Hydra allow in vivo tracking of individual stem cells during morphogenesis, Proc Natl Acad Sci USA, 103 (2006) 6208–6211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Šali A, Blundell TL, Comparative protein modelling by satisfaction of spatial restraints, J Mol Biol, 234 (1993) 779–815. [DOI] [PubMed] [Google Scholar]
- [37].Brzozowski AM, Pike AC, Dauter Z, Hubbard RE, Bonn T, Engström O, Ohman L, Greene GL, Gustafsson JA, Carlquist M, Molecular basis of agonism and antagonism in the oestrogen receptor, Nature, 389 (1997) 753–758. [DOI] [PubMed] [Google Scholar]
- [38].Greschik H, Wurtz J-M, Sanglier S, Bourguet W, van Dorsselaer A, Moras D, Renaud J-P, Structural and functional evidence for ligand-independent transcriptional activation by the estrogen-related receptor 3, Mol Cell, 9 (2002) 303–313. [DOI] [PubMed] [Google Scholar]
- [39].Greschik H, Althage M, Flaig R, Sato Y, Chavant V, Peluso-Iltis V, Choulier L, Cronet P, Rochel N, Schüle R, Strömstedt PE, Moras D, Communication between the ERRalpha homodimer interface and the PGC-lalpha binding surface via the helix 8-9 loop, J Biol Chem, 283 (2008) 20220–20223. [DOI] [PubMed] [Google Scholar]
- [40].Fiser A, Do RK, Šali A, Modeling of loops in protein structures, Protein Sci, 9 (2000) 1753–1773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Shen MY, Šali A A, Statistical potential for assessment and prediction of protein structures. Protein Sci, 15 (2006) 2507–2524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Chen VB, Arendall WB 3rd, Headd JJ, Keedy DA, Immormino RM, Kapral GJ, Murray L, Richardson JS, Richardson DC, MolProbity: all-atom structure validation for macromolecular crystallography, Acta Crystallogr D Biol Crystallogr, 66 (2010) 12–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Morris GM, Huey R, Lindstrom W, Sanner MF, Belew RK, Goodsell DS, Olson AJ, Autodock4 and AutoDockTools4: automated docking with selective receptor flexiblity. J Comp Chem, 16(2009)2785–2791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Wallace AC, Laskowski RA, Thornton JM. LIGPLOT: a program to generate schematic diagrams of protein-ligand interactions. Protein Eng, 8 (1996) 127–134. [DOI] [PubMed] [Google Scholar]
- [45].Markov GV, Laudet V, Origin and evolution of the ligand-binding ability of nuclear receptors. Mol Cell Endocrinol, 334 (2011) 21–30. [DOI] [PubMed] [Google Scholar]
- [46].Bridgham JT, Keay J, Ortlund EA, Thornton JW, Vestigialization of an allosteric switch: genetic and structural mechanisms for the evolution of constitutive activity in a steroid hormone receptor, PLoS Genet, 10 (2014) el004058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Katsu Y, Cziko PA, Chandsawangbhuwana C, Thornton JW, Sato R, Oka K, Takei Y, Baker ME, Iguchi T, A second estrogen receptor from Japanese lamprey (Lethenteron jciponicum) does not have activities for estrogen binding and transcription, Gen Comp Endocrinol, 236 (2016) 105–114. [DOI] [PubMed] [Google Scholar]
- [48].Tanenbaum DM, Wang Y, Williams SP, Sigler PB, Crystallographic comparison of the estrogen and progesterone receptor’s ligand binding domains. Proc Natl Acad Sci U S A 95 (1998) 5998–6003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Schwartz N, Verma A, Bivens CB, Schwartz Z, Boyan BD, Rapid steroid hormone actions via membrane receptors, BB A Molecular Cell Research, 1863 (2016) 2289–2298. [DOI] [PubMed] [Google Scholar]
- [50].Baker ME, Lathe R, The promiscuous estrogen receptor : evolution of physiological estrogens and response to phytochemicals and endocrine disruptors, J Steroid Biochem Mol Biol (2018), this issue. [DOI] [PubMed] [Google Scholar]
- [51].Thornton JW, Evolution of vertebrate steroid receptors from an ancestral estrogen receptor by ligand exploitation and serial genome expansions, Proc Natl Acad Sci U S A, 98 (2001) 5671–5676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Russell DW, The Enzymes, Regulation, and Genetics of Bile Acid Synthesis, Annu Rev Biochem, 72 (2003) 137–174. [DOI] [PubMed] [Google Scholar]
- [53].Norris DO, Vertebrate endocrinology. Elsevier Academic Press, 2007. [Google Scholar]
- [54].Thomas C, Auwerx J, Schoonjans K, Bile acids and the membrane bile acid receptor TGR5 - connecting nutrition and metabolism, Thyroid, 18 (2008) 167–174. [DOI] [PubMed] [Google Scholar]
- [55].Steinmetz PRH, Aman A, Kraus JEM, Technau U, Gut-like ectodermal tissue in a sea anemone challenges germ layer homology, Nat Ecol Evol, 10 (2017) 1535–1542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Giguere V, Transcriptional control of energy homeostasis by the estrogen-related receptors, Endocr Rev, 29 (2008) 677–696. [DOI] [PubMed] [Google Scholar]
- [57].Tennessen JM, Baker KD, Lam G, Evans J, Thummel CS, The Drosophila estrogen-related receptor directs a metabolic switch that supports developmental growth, Cell Metab, 13 (2011) 139–148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Hand C, Uhlinger KR, Asexual reproduction by transverse fission and some anomalies in the sea anemone Nematostella vectensis, Invertebr Biol, 114 (1995) 9–18. [Google Scholar]
- [59].Chia F-S, Spaulding JG, Development and juvenile growth of the sea anemone Tealia crassicornis, Biol Bull, 142 (1972) 206–218. [DOI] [PubMed] [Google Scholar]
- [60].Sebens KP, The regulation of asexual reproduction and indeterminate body size in the sea anemone Anthopleura. Biol Bull, 158 (1980) 370–82. [Google Scholar]
- [61].Fitt WK, Pardy RL, Effects of starvation, and light and dark on the energy metabolism of symbiotic and aposymbiotic sea anemones, Anthopleura elegantissima, Mar Biol, 61 (1981) 199–202. [Google Scholar]
- [62].Chera S, Buzgariu W, Ghila L, Galliot B. Autophagy in Hydra. A response to starvation and stress in early animal evolution, Biochim Biophys Acta, 1793 (2009) 1432–1443. [DOI] [PubMed] [Google Scholar]
- [63].Cimino G, Rosa SD, Stefano SD, Sodano G, G, Cholest-4-en-4,16β18,22R-tetrol-3-one 16,18-diacetate, a novel polyhydroxylated steroid from the hydroid Eudendrium sp., Tetrahedron Lett, 21 (1980)3303–3304. [Google Scholar]
- [64].Milkova T, Popov S, Marekov N, Andreev S, Sterols from black sea invertebrates—I. Sterols from Scyphozoa and Anthozoa (Coelenterata). Comp Biochem Physiol B, 67 (1980) 633–638. [Google Scholar]
- [65].De Rosa S, Milone A, Popov S, Andreev S, Sterol composition of the Black Sea Hydrozoan, Obelia longissima (Pallas 1766), Comp Biochem Physiol B Biochem Mol Biol, 123 (1999) 229–233. [Google Scholar]
- [66].Edison AS, Hall RD, Junot C, Karp PD, Kurland IJ, Mistrik R, Reed LK, Saito K, Salek RM, Steinbeck C, Sumner LW, Viant MR, M., The Time Is Right to Focus on Model Organism Metabolomes, Metabolites, 6 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [67].Cachet N, Genta-Jouve G, Ivanisevic J, Chevaldonne P, Sinniger F, Culioli G, Perez T, Thomas OP, Metabolomic profiling reveals deep chemical divergence between two morphotypes of the zoanthid Parazoanthus axineiiae, Sci Rep, 5 (2015) 8282. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.