

Abstract
Taste modalities are conserved in insects and mammals. Sweet gustatory signals evoke attractive behaviors while bitter gustatory information drive aversive behaviors. Salt (NaCl) is an essential nutrient required for various physiological processes, including electrolyte homeostasis, neuronal activity, nutrient absorption, and muscle contraction. Not only mammals, even in Drosophila melanogaster, the detection of NaCl induces two different behaviors: Low concentrations of NaCl act as an attractant, whereas high concentrations act as repellant. The fruit fly is an excellent model system for studying the underlying mechanisms of salt taste due to its relatively simple neuroanatomical organization of the brain and peripheral taste system, the availability of powerful genetic tools and transgenic strains. In this review, we have revisited the literature and the information provided by various laboratories using invertebrate model system Drosophila that has helped us to understand NaCl salt taste so far. We hope that this compiled information from Drosophila will be of general significance and interest for forthcoming studies of the structure, function, and behavioral role of NaCl-sensitive (low and high concentrations) gustatory circuitry for understanding NaCl salt taste in all animals.
Keywords: Taste, Drosophila melanogaster, neural circuits, salt, gustatory neurons, brain
Introduction
Dietary sodium is consumed as a common salt—sodium chloride (NaCl). Sodium, which is present in dietary table salt, is an essential nutrient required for many physiological processes including electrolyte homeostasis, nutrition absorption, maintenance of cell plasma volume, acid-base balance, transmission of nerve impulses, and normal cell physiology. Sodium contributes to the establishment of the membrane potential of most cells and plays a direct role in the action potential required for the transmission of nerve impulses and muscle contraction.
Sodium is a mineral that occurs naturally in foods like flour, mushrooms, celery, beets, and milk and is added in ionized form to table salt (40% sodium and 60% chloride). Packaged and prepared foods like canned soups and frozen eatery items often have added salt during their processing as a measure of preservation. The presence of salt makes food more palatable than the same food with no salt. According to World Health Organization (WHO) details, people in various parts of the world are consuming too much salt in their diets—on an average 9 to 12 g/d which is much more than WHO salt intake guidelines (less than 1500 mg/d, ie, 0.75 teaspoon—3.75 g/d should be consumed). Shown by many groups, extreme intake of salt results in various health issues and causes hypertension, strokes, stomach cancer, osteoporosis, autoimmune diseases, kidney stone, water retention, and bone weakening. Lack of dietary salt intake is also associated with health problems like low blood pressure. Hence, an appropriate amount of salt should be consumed for proper functioning of our body parts and to live longer and healthier.
It is ironic that despite the high incidence of cardiovascular diseases, stroke, elevated blood pressure, and high hypertension-related mortality, we currently do not fully understand the molecular and cellular mechanisms by which low and/or high salt concentrations are perceived or differentially encoded throughout peripheral tissues and in the brain. Furthermore, it is not clear how low or high salt dietary salt intake influences complex feeding behaviors. More research is required in this direction.
This review highlights the studies within Drosophila melanogaster that have begun to shed light on the mechanisms of salt detection, how salt influences feeding behaviors, and the influence of salt on other physiological functions. The findings from these studies hold potential to help us understand similar mechanisms that exist in higher order species and may therefore lead to the identification of targetable pathways in human disease.
Drosophila Taste System
Food palatability, or how food tastes, is the main driving factor for initiating a bout of feeding. Like mammals, insects can detect and discriminate among different gustatory stimuli, such as sugars, bitter substances, and various salt concentrations, which induce an attractive or a repulsive response in behavioral tests. Gustatory signals have been shown to play vital roles in controlling behavior, such as searching for food or finding sexual partners.1 Drosophila is among the most highly studied genetic model systems for investigating feeding behaviors and peripheral and central taste coding. A total of 60 genes in the gustatory receptor (GR) gene family encode 68 receptor proteins.2-4 A number of studies within the past decade have focused on understanding the molecular and cellular mechanism by which different taste modalities (i.e. sweet, bitter, water, salt) are perceived in Drosophila. The response profiles of gustatory neurons have been described, revealing their specificity, tuning breadth, and behavioral roles,5-7 highlighting the complexity of stimulus representation in the taste system as a whole. These findings have invited experiments to probe the extent to which the fly can use this information for both hard-wired and experience-guided behaviors.8 Unexpected interactions between aversive tastants and appetitive neurons have emerged by performing functional analysis of taste neurons and have revealed at least 2 distinct mechanisms by which sweet taste neuron activity is inhibited by bitter tastants.9,10
Expression profiles of GRs in various taste organs such as the labellum, legs, anterior margin of wings, pharynx, genitalia, and in internal organs including intestine in adult Drosophila11,12 have been shown by various laboratories. (Figure 1A). GRs also express in organs of Drosophila larvae such as the terminal organ and pharyngeal sense organs14 (Figure 1B) and are shown to be involved in sensing sweet and bitter compounds.15-17 Drosophila larval olfactory and gustatory chemosensory organs located on the head surface are dorsal organ (DO), terminal organ (TO), and ventral organ (VO), and 3 pharyngeal organs12 (Figure 1B). Olfactory receptor neurons (ORNs) located in the DO project into glomeruli of the antennal lobe (AL), whereas gustatory receptor neurons (GRNs) project via 4 different nerves to the subesophageal zone12 (SEZ) (Figure 1B). During metamorphosis in Drosophila, almost the entire larval peripheral nervous system disappears to be reconstructed into the adult peripheral nervous system.18 The pharyngeal sensory neurons in Drosophila have been demonstrated as the primary gate keepers of the taste system1,19,20 (Figure 1A) are an exception to this rebuilding during metamorphosis.21 It is not clear though what this means for the development and physiology of the organism. More detailed functional analyses are required to understand the involvement of the pharyngeal sensory neurons of the labral sense organ (LSO), the ventral cibarial sensory organ (VCSO), and of the dorsal cibarial sensory organ (DCSO). Identifying chemicals these neurons detect may provide insight to understand why these neurons are maintained throughout development and if the function is conserved during larval and adult stages.
Figure 1.
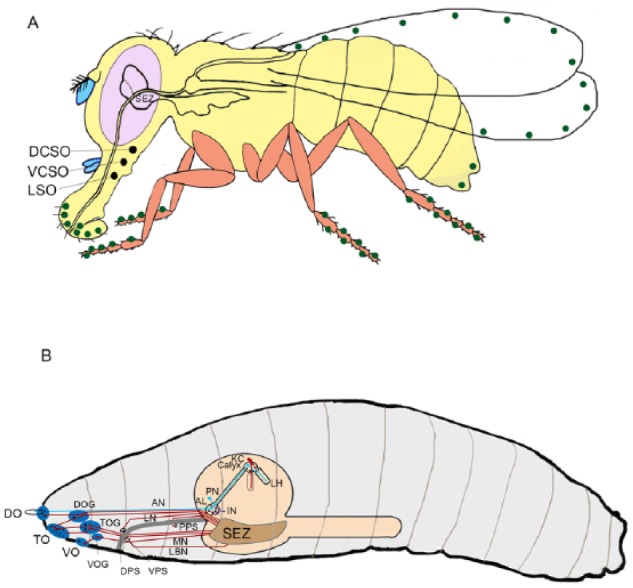
Gustatory system of Drosophila. (A) Cartoon showing the presence of GRs on various taste organs (labellum, legs, anterior margin of wings, pharynx, and genitalia) in adult fly as green spots. The pharyngeal organs are LSO (labral sense organ), VCSO (ventral cibarial sensory organ), and DCSO (dorsal cibarial sensory organ). The peripheral taste neurons from various organs terminate in the brain in specialized areas called SEZ. (B) Chemosensory system of Drosophila larva. The 3 external chemosensory organs, the dorsal organ (DO), terminal organ (TO), ventral organ (VO), and the dorsal, ventral, and posterior pharyngeal sense organs (DPS, VPS, PPS) include mainly gustatory sensilla. The sensory neurons cell bodies are collected in ganglia below each sense organ (DOG, TOG, VOG). Some neurons innervating the TO are also located in the DOG. Olfactory receptor neurons project into individual glomeruli of the larval antennal lobe (AL), which are interconnected by local interneurons (IN). Projection neurons (PN) link the AL with 2 higher olfactory centers, the mushroom body (MB) calyx and the lateral horn (LH). An intrinsic MB Kenyon cell (KC) is shown in red. GRN afferents (brown) extend via 4 different nerves to the SEZ. The pharynx is shown in gray.
Source: Adapted from Gerber and Stocker.13
AN indicates antennal nerve; LN, labral nerve; MN, maxillary nerve; PPS, posterior pharyngeal sensillae; LBN, labial nerve.
In adult Drosophila, 3 sensory organs exist in the pharynx: LSO, VCSO, and DCSO.11,22 The DCSO and VCSO occupy the dorsal and ventral parts of the cibarium, and the LSO is located near the labellum (Figure 1A). The pharyngeal sense organs are situated in an anatomically ideal position to act as additional regulators of feeding that is between the external sensory system to sense the quality of food, and the post-ingestive internal nutrient sensing system which determines whether to continue or stop feeding. Adult GRNs that express sugar receptors promote and maintain feeding,7 while bitter sensing by larval pharyngeal GRNs inhibit ingestion.15 Although emerging evidences suggest that the pharyngeal sense organs indeed regulate feeding, but much is still undetermined.
Salt: An Unavoidable Taste
Among the various taste modalities, NaCl plays an important supportive roles in a multitude of physiological processes, including neuronal function. Both sodium and chloride must be ingested and maintained at proper concentrations throughout the body by homeostatic mechanisms that ensure ideal osmolarity. Therefore, consumption of just the right amount of salt is required; one may expect the behavioral effects of salt being tightly regulated according to concentration. Indeed, the appetitive responses to low salt gradually turn into aversion as the concentration increases,23-25 and these opposing behavioral responses are generated by discrete molecular and cellular processes.
As mentioned elsewhere not only sodium, potassium, calcium, and magnesium are other important nutrients required in our body and act as electrolytes. Chlorine plays a fundamental role in digestion by helping to maintain acid-base balance, and it also aids in the absorption of potassium. Magnesium chloride is consum.29ed as a supplement version of magnesium, a mineral found naturally in the body, and is essential for muscle and nerve function as well as heart and bone health. Magnesium chloride supplements are not necessary unless one is deficient for the mineral. Salts like potassium chloride provides similar properties like NaCl but with several unwanted side effects, of which the most important have relatively offensive side tastes: bitter, acrid, and metallic. The other salt is calcium chloride which provides a small amount of dietary calcium important for maintaining strong bones and plays a role in nerve impulse and muscle function. Sodium bicarbonate (baking soda) is similar to that of table salt for maintaining optimal health and helps in neutralizing stomach acids. When these salts are consumed sodium, chloride, and bicarbonate become electrolytes that carry electrical impulses in our body.
Role of DEG/ENaC Channels in Salt Taste
Like other taste modalities, the gustatory system of animals recognize NaCl and other salts. This chemical sensory modality allows animals to detect and ingest salt, discriminate between different salts and allows them to avoid high salt concentrations that can have deleterious effects on the body. It has been shown in different organisms that moderate and low concentrations of Na+ salt (<100 mM) are appetitive and are mainly sensed by amiloride-sensitive epithelial sodium channel (ENaC).26 ENaC-KO mice lack responses in low salt (amiloride-sensitive) pathway.26
Amiloride-sensitive degenerin/epithelial Na+ channels (DEG/ENaC) form constitutively open and cation selective pores.27 The involvement of the DEG/ENaC channels in sensing salt taste as receptors are supported by the localization of the α, β, and γ ENaC subunits in mammalian taste receptor cells, as well as by the observations that aldosterone increases the apical localization of these subunits.28-32 This is further supported by the observations that amiloride impairs gustatory responses to NaCl.33,34 Extracellular amiloride blocks most DEG/ENaC channels.35 Although amiloride is not a specific inhibitor of DEG/ENaC channels, it can also inhibit other membrane transport processes. Amiloride-insensitive mechanisms have also been proposed to participate in mammalian NaCl taste and include Na/H antiporters36 and cation diffusion through tight junctions.29
Drosophila genome has a large number of predicted DEG/ENaC genes,37 and these channels have been later tested for their role in salt taste. The degenerin epithelial Na+ channel gene family, known as the pickpocket genes in Drosophila, encode subunits of non-voltage gated, amiloride-sensitive cation channels. It has been suggested that DEG/ENaC channels may be formed by homo- or heteromeric arrangements of subunits. Each subunit has 2 transmembrane domains and a large cysteine-rich extracellular loop domain. DEG/ENaC channels are functionally diverse, with roles in fluid and salt absorbance, mechanosensation, and chemosensation. The degenerin/epithelium sodium channels Pickpocket 11(ppk11) and Pickpocket 19 (ppk19) have roles in the detection of sodium and potassium and are also expressed in the tracheal system for liquid clearance.25,38 In larva, ppk11 and ppk19 are found on taste sensing terminal organ and in adults they are present on labellum, legs, and wing margins. Both the larval terminal organ and the adult labellum taste bristles consist of bipolar taste receptor neurons surrounded by supporting cells; this arrangement forms a specialized structure containing a pore at its tip.39 Thus, the localization of PPK subunits in these specialized sensory neurons position them where they could detect changes in salt concentration. Flies with disruption of PPK11 or PPK19 fail to recognize low concentration of sodium or potassium in water.25 Expression of these genes is necessary for the appetitive behavioral responses to low salt, but are dispensable for the aversive responses to high salt.25 The evidence provided by Liu et al25suggests that the PPK genes are involved in salt taste raises the possibility that salt detection mechanisms are conserved from flies to mammals. A deeper understanding of how salt is detected requires the identification of additional DEG/ENaC subunits and associated proteins (beyond PPK11/PPK19).
The Low Sodium Permeable Channel Ir76b
Ionotropic receptors (IRs—comprise a subgroup of the ionotropic glutamate receptor (iGluR) family) have been recognized as critical sensory receptors in insects for detecting environmental stimuli such as chemical compounds,40,41 temperature changes,42 and humidity.43 Recently, it has been established that the salt attractive pathway for low salt relies on a Na+ permeable channel ionotropic receptor IR76b, formerly not known to function in taste.44 This channel bears no relationship to ENaC channels which are necessary for sensing low salt and showing appetitive responses to Na+ at low concentrations in mice.26 In particular, IR76b is expressed in L-bristle GRNs of labellum (Figure 2A and B) and is proposed to serve as a functional Drosophila counterpart of mammalian ENaC. It has been proposed that some ENaC channels may be constitutively active27 leading to depolarization of taste receptor cells following a rise in cation levels at the cell surface. Thus, despite the divergence between fly IRs and mammalian ENaC channels, they may mediate salt taste through similar mechanisms. Zhang et al44 have described competition between taste neurons in the S- and L-type taste sensilla (Figure 2A and B) accounts for the bidirectional behavioral responses to salt. At low salt concentrations, the low-salt GRNs dominate over the high-salt GRNs, thereby causing the animals to prefer low salt. Inversely, At high salt levels, the high-salt GRNs overwhelm the low-salt GRNs, resulting in salt rejection. This competition model presented for low and high salt taste detection may represent a widely used mechanism for salt taste coding in other animals, including mammals.44 In the same study, GRNs in a few s-bristles have been suggested to act as receptors of high NaCl taste in an IR76b-independent manner, while others have shown cells in L- and i-bristles respond to high NaCl concentrations in an earlier study45 (Figure 2). Hence, it has been proposed44 that NaCl perception in Drosophila adults is determined by a bimodal switch system operating in taste neurons that allows detection of low- and high-NaCl concentrations separately. It has been demonstrated though that the ionotropic channel IR76b is selectively involved in the attractive pathway.
Figure 2.

Schematic diagram of the adult Drosophila labellum showing sensillar classes (lateral view). (A) Cartoon showing location of the 31 chemosensilla identified by various groups. (B) Confocal image of the labellum showing various taste sensilla types. Anterior is top and dorsal to the right. Color code for taste sensilla: orange (intermediate-I type sensilla), sky blue (small-s sensilla type), and red (large-L type sensilla).
Little is known about the molecular mechanism of high-salt taste in Drosophila as in most other animals. More recently, another group46 has discovered that suppression of feeding behavior by high sodium-rich food also require Ir76b which is necessary for neuronal Na+ responses in the s-bristle taste cells besides the previously reported L-bristle GRNs in Drosophila labellar gustatory receptor neurons. Together, the results from Zhang et al44 and Lee et al46 suggest that IR76b plays a central role in gustation of both attractive and aversive Na+ concentrations possibly in combination with IRs instructing respective functions46 (Figure 3).
Figure 3.

Two different models of feeding preference decision in response to Na+ concentrations (adapted from Lee et al46. (A) According to Zhang et al,44 Ir76b GRNs in large (L)-bristles signal brain for feeding attraction in response to low salt concentrations. At high salt concentrations, the interneurons of unknown identity, may lead to activation of L-bristle GRNs which may in turn aid in robust excitation of the aversive s-bristles GRNs. In this manner, Ir76b GRNs in L-bristles, are required for both attraction and aversion to increasing concentrations of Na+. The other possibility would be that the L-bristle GRNs code for repulsion, as opposed to the conclusion by Zhang et al.44 The aversion could be also be through GRNs in L and i- sensillae45 or through Ir76b independent mechanisms. In any case, there is a possibility that unidentified Ir76b cells exist for feeding attraction, as in the absence of the L-bristle GRNs the balance of the whole Ir76b cell population move towards attraction (B).
Heterologous expression of IR76b in HEK293 cells has shown intrinsic Na+ sensitivity of the IR76b protein.44 In addition, 2 recent studies demonstrated that IR76b is critical for taste to amino - acids.47,48 Such dual responsiveness to salt and amino acids is reminiscent of extracellular Ca2+-sensing receptor (CaSR)49 and mammalian kainate receptor iGluRs, GRIK1 and GRIK2.50 CaSR, a G protein-coupled receptor (GPCR), is activated by another divalent cation Ca2+ as well as by L-amino acids. The iGluRs homologous to IR76b seem to be functionally conserved in salt sensitivity, as they bind Na+ ions in order to be activated by glutamate. In addition, iGluRs have been reported to be present in taste receptor cells.51,52 Thus, the findings that IR76b critically contribute to high-Na+ taste suggests the interesting possibility that iGluRs may in part mediate the molecular mechanism of high Na+ taste in mammals which has yet to be identified. Thus, these results highlight the complexity of IR76b-dependent Na+ gustation in Drosophila. Various Gal4 transgenes covering different subpopulations of the IR76b cells would facilitate future studies to further dissecting the cellular substrates for IR76b-dependent Na+ feeding behavior.46
The Role of Pharyngeal GR2a
In the vertebrates taste system, taste cells are present not only on the surface of the tongue but also in other organs including the pharynx.53-55 The Zebrafish T1R2/T1R3 sweet taste receptors are expressed in the pharynx, the lip and in gill raker.54 Although expression of T1R2/T1R3 in the pharynx has not been examined in mammals, it is known that the cranial nerve fibers innervating pharyngeal taste cells respond to sugar.56,57 These neurons project to the nucleus of solitary tract (NST) in the hindbrain.56-58 The NST also receives peripheral satiety signals transmitted via vagal nerve fibers, indicating that this brain region is involved in many regulatory processes that regulate food intake including the decision to ingest.58,59 It has been suggested that due to similarities of insect and vertebrate taste processing systems, interneuron populations in the NST with the physiological properties of cholinergic local interneurons (IN1) neurons identified recently20 may exist. Understanding how the pharynx perceives taste would permit rapid evaluation of food intake volume and quality and would provide real-time feedback to the central brain to regulate ingestion in both vertebrates and insects.
Using capillary feeder (CAFÉ) assay and GR expression analysis, recently it has been1 reported that a pair of pharyngeal gustatory receptor neurons is involved in feeding inhibition in response to high concentrations of sodium ions and function as modulators of feeding. Molecular genetic tools, RNAi experiments, and mutant analyses have shown that the neurons of gustatory receptor GR2a are involved in feeding inhibition. This feeding suppression suggests that Drosophila find highly concentrated salty food unappetizing, but is not manifested when flies are starved and hungry suggesting starvation influences feeding preferences. These results provide evidence that factors involved in feeding, function in a context-dependent manner since feeding is influenced by many factors including nutritional status or hunger.1 Although data suggest that GR2a is involved in the aversion to high salt, it is unclear whether GR2a is a salt receptor per se or a modulator of the response.
Serrano in the Larval Taste System
Serrano (Sano) is an apically enriched cytosolic protein required for neuronal activity in response to high-salt concentrations. Sano coexpress with GR66a in 4 gustatory neurons in the terminal organ of third instar larvae.60 Absence of the DEG/ENaC channel PPK19 that function in the same set of neurons eliminates the cellular response to high-salt concentrations. Thus, both PPK19 and Sano are required on the larval gustatory neurons for the detection of high-salt concentrations. Disruption of sano gene expression in gustatory neurons leads to specific loss of high-salt concentration avoidance behavior in larvae. Inactivation of sano-expressing GRNs induces an attraction to high-salt concentrations, suggesting that disruption of only aversive salt pathways determines the opposing behavioral responses to low and high salt.44
Previous studies have shown that Sano interacts with other proteins, such as Grb261 (a protein involved in the signaling pathways of tracheal and wing development) and with Epac, (exchange protein directly activated by cAMP) a member of the Rap1 signal transduction pathway. It is involved in cell adhesion and differentiation, as well as in neuronal activity by regulating calcium levels or neurotransmitter release.62 The genetic and molecular interactions between Sano and Epac is not well understood and remain to be fully demonstrated by future in vitro and in vivo studies. Such studies will provide new insights into the cellular mechanisms taking place downstream of the DEG/ENaC channels in salt-detecting neurons.
Post Mating Circuit
An animal’s nutritional requirements change over their life time to meet the nutritional demands and animals display specific behavioral adaptations to increase their intake of particular nutrients they require to maintain the homeostasis. An animal’s specific nutrient intake can be adapted to needs in the current state by need-dependent and need-independent mechanisms.63 It has been extensively shown that during reproductive period, females’ nutritional requirements change drastically and they tend to invest enormous resources for their progeny. A specific appetite for sodium increases during the reproductive and lactation phases.64-67 For the reproductive success, sodium provides ions required for nutrient balance in newly formed eggs or an increase in total food intake which ultimately contributes to egg production.68,69
A study conducted by Walker et al70 has provided insights into the physiological regulation of salt intake. In particular they have dissected the feed-forward regulation of sensory processing in Drosophila female representing a mechanism through which reproductive state-sensitive circuits modify complex behaviors. Authors in this article have demonstrated that mating induces a salt appetite in Drosophila as seen in many other species during reproduction. They have also dissected the neuronal mechanisms through which animal’s reproductive state drives salt appetite which does not require octopamine as needed in post - mating yeast appetite. Data suggests that during the reproductive period, female flies crave more for sodium and their feeding rises more toward it. The increased attractiveness for salt positively affects reproductive output in flies as in many animals. This appetite for salt is induced independently of salt requirements for egg laying and by a feed-forward change in taste processing, driven by a male-derived signal acting on female post - mating circuitry. The authors explain that male derive sex peptides after copulation transfers into the neuronal sex peptide receptor (SPR) and also inhibit the action of downstream sex peptide abdominal ganglion (SAG) neurons, which have a role in increasing egg laying and inhibiting remating. Sex peptides acts on a small set of sensorimotor system. This study highlights the importance of reproduction as a critical modulator of taste processing and brings new insight into the mechanistic basis of this state-dependent nutritional modulation. Future investigations are required to explore to what extent feed-forward regulation is employed to control specific behavioral strategies used to acquire nutrients depending on differential internal state signals.70
The flexibility of circuit analysis in Drosophila offers unique opportunity to understand the circuit mechanisms through which internal state signals modulate taste processing in the brain, and thus bring about adaptive changes in food preference.71 To attain this, mating may modulate the response of sensory neurons to salt taste, as demonstrated in the olfactory pheromone system of moths.72 Similarly, GRN responses are shown to be modulated by hunger73-75 and the sensitivity of pheromone-sensitive olfactory receptor neurons in mice are modulated across the estrus cycle.76 Conversely, the mating state could lead to a combination of modulation at the receptor neuron level and modification of higher order processing. Understanding how alliesthesia is implemented at the circuit level will provide an opportunity to understand how internal state changes affect sensory processing to mediate adaptive behaviors.70
Learning and Memory
The knowledge of salt handling in larva is still very limited. A study conducted by Niewalda et al77 has explored salt processing in the larval taste system of Drosophila. Their data highlight the behavioral effects of NaCl in choice behavior, feeding behavior, and learning shift from appetitive to aversive as the concentration of salt increases. Regarding feeding behavior, others45 have reported that in adult flies feeding is upregulated by salt at 0.1 M but is downregulated by 0.4 M salt, with the strongest “appetizing” effect between 0.05 and 0.1 M. These findings fit reasonably well with Niewalda et al77 results in larva and suggest some functional conservation of salt processing between larva and adult. Based on the observation that most pharyngeal gustatory sensory neurons of the larva are retained into adulthood, such conserved function had already been proposed.21 Studies conducted by Niewalda et al77 has dissociated parametrically the reflex releasing (choice, feeding) from the reinforcing function of salt in terms of their respective dose-effect characteristics: the reinforcing effect shifts by one order of magnitude toward higher concentrations. Interestingly, a similar shift between these 2 kinds of behavioral effects is also found for sugars,78 suggesting some degree of generality of such parametric dissociation. Thus, for both salt and sugars, the input pathways for gustatory behavior appear to be more sensitive than the ones supporting gustatory reinforcement.
Conclusions and Future Prospective
Understanding higher order low and high salt circuitry in the brain
Drosophila are capable of detecting taste modalities that are associated with food acceptance or rejection behavior79,80 by various taste cells present on the proboscis, legs, and wings of the adult fly.11 The different classes of taste cells include bitter cells marked by the GR, namely, GR66a and sweet cells marked by GR64f.81-86 PPK28 ion channel expressing water taste cells87,88 activated by low osmolarity and are inhibited by high osmolarity. IR76b-expressing taste neurons respond to low salt.44
Different peripheral taste neurons expressing taste receptors from the labellum and pharynx target discrete regions of the SEZ,11,89 the taste center of the fly brain (Figure 4).84 This densely innervated brain structure houses various projection neurons, interneurons, and motor neurons required for taste acceptance and rejection, along with motor circuits that regulate ingestion73,90-97suggesting the presence of local circuits in the SEZ that process taste cues from detection to behavior. In addition, neuromodulators, namely, dopamine, serotonin, neuropeptide F, and short-neuropeptide F modulate food intake by altering the activity of sensory neurons that detect food stimuli, or of homeostatic neurons that regulate hunger.74,75,98,99 Many tastants that inhibit proboscis extension and feeding act not only via activating bitter taste neurons5 but also by inhibiting sweet taste neurons.9,100
Figure 4.
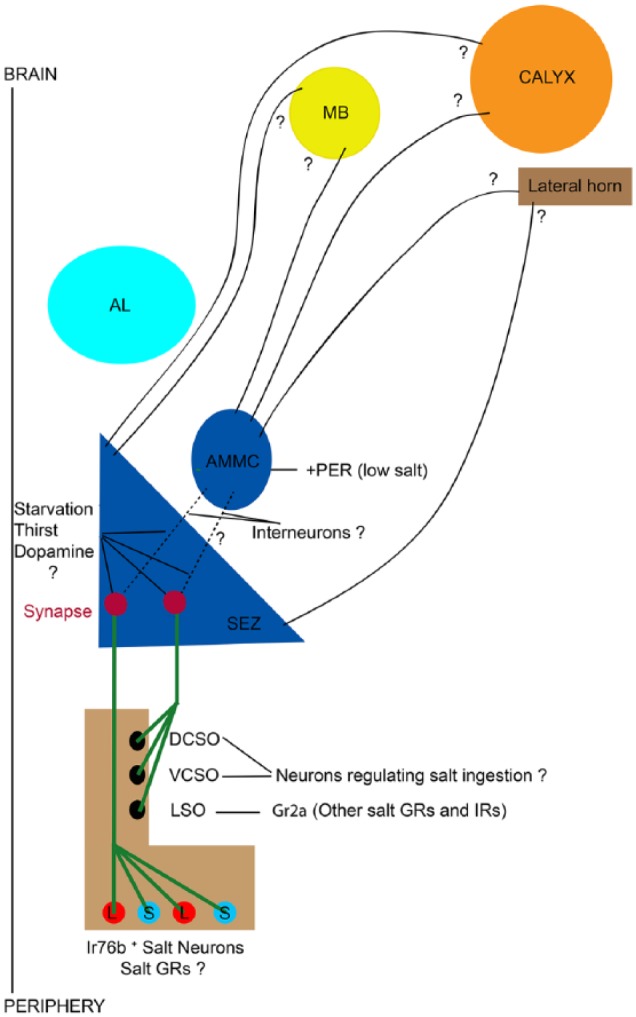
Proposed model. Shown is a schematic illustrating the identified and unidentified components of salt taste circuit in Drosophila. IR76b neurons from the periphery send information about salt taste to SEZ. Direct sensors involving gustatory receptors (GRs) for salt remain to be identified. Pharyngeal taste neurons of LSO (Gr2a neurons are involved in feeding inhibition in response to high concentration of sodium ions), VCSO, and DCSO also send processes to SEZ. The identity of other salt receptors and IRs as well as salt taste neurons of VCSO and DCSO are not known yet. Interneurons in the SEZ (black dotted lines) that receive and process gustatory information about salt are largely lacking. Like sweet taste, if any, the role of dopamine signaling in reducing behavioral threshold to salt upon starvation and modulation of feeding responses is not known. It would be interesting to determine if there would be state-dependent (starvation and thirst) alterations in salt taste circuit activity that could lead to more salt eating or eating of high salt concentrations. One needs to verify the possibilities if the information about the starvation state is amplified during the relay to salt second-order neurons or that these neurons may also be targets of signaling pathways that convey information about the starvation state. Role of AMMC as a secondary center for low salt taste as in case of sweet taste is a future question. It is not known where the information from salt taste neurons input upon stimulation of labellum and tarsi taste neurons with low salt concentrations is integrated, either upstream or at second-order neurons. Since salt taste projections to higher brain centers have not yet been characterized, questions regarding the salt circuitry providing gustatory inputs from SEZ or AMMC or both to motor neurons, MB, calyx and lateral horn to control feeding behavior and associations with appetitive and aversive learning remain unaddressed.
AL indicates antennal lobe; AMMC, antennal mechanosensory and motor center; DCSO, dorsal cibarial sensory organ; LSO, labral sense organ; MB, mushroom body; PER, proboscis extension response; SEZ, subesophageal zone; VCSO, ventral cibarial sensory organ.
Only a limited understanding of central neural circuits to examine the modality processing have been attained so far beyond sensory neurons and MNs that induce behaviorally appropriate motor output. Also, the identification of neural circuits that integrate and process information of taste, hunger, and metabolism to control food ingestion remain poorly understood.58,101 The presence of a large number of interneurons in the SEZ that appear to respond to gustatory input, it is remarkable that only very few of these have been identified at the cellular level. Few gustatory interneuron types that respond to sweet taste input have been identified in the adult SEZ recently.1,20,92 Kain and Dahanukar92 reported the first identified second-order sweet gustatory projection neuron (NP1562) that make synaptic connections with GR5a in the SEZ. This set of neurons is unique as they relay sweet taste information from the SEZ to the antennal mechanosensory and motor center (AMMC) in Drosophila. These neurons show increased sucrose sensitivity upon starvation and dopamine administration.92 Similar to sugars, low salt is innately attractive to most animals. To understand how low salt taste activates appropriate feeding behaviors, higher order neurons in taste circuits must be identified and studied (Figure 4). Proboscis extension can be induced by stimulation of either labellar or tarsal taste hairs in wild-type flies.101 Taste neurons originating from these 2 organs target distinct areas in the SEZ.11,85,86 Where the information from the 2 organs is integrated is not known either upstream or at second-order neurons (Figure 4).
In a recent report,102 large-scale analysis of pan-neural activity in the fly brain suggested that taste modalities in the periphery activate different pathways in the brain. Sweet and bitter tastes are processed by segregated pathways, consistent with labeled line taste processing suggesting strategies that ensure innate responses to essential compounds. Information processing in separate streams is also maintained in the higher brain and is mutually inhibitory. Supporting the studies in the mammalian gustatory system that argues for a modality-specific representation in the gustatory cortex and support labeled line models.103-105 These studies suggest that faithful pathways may be a general strategy to process tastes used throughout evolution. The field is still highly controversial though and evidences supporting both “distributed and labeled line model of taste coding” exist and need future examination. Work by Harris et al102 has provided a population overview of gustatory processing in the fly that will help to determine the functional role of each neuron during different steps of feeding behavior, the anatomy and connectivity of taste-responsive neurons.
With the exception of 2 identified bitter-sensitive projection interneurons types,1,106 information about first-order interneurons that receive and process gustatory information about other tastants categories such as bitter, salt, and water is largely lacking (Figure 4). In a recent study,97 the neural connections for bitter taste processing has been investigated. This study has identified a pair of gustatory local interneurons (bGLNs) involved in bitter taste aversion in flies. bGLN dendrites stay in close proximity to axonal termini of bitter-sensory neurons in the SEZ. It is incredible that the bitter taste modality is conserved and evokes aversive behavior in insects and mammals. The identification of bGLN is a significant step towards understanding how bitter taste modalities are processed by the gustatory circuitry in the SEZ of the brain. Whether these or other yet-unidentified SEZ neurons with roles in gustation or feeding are, indeed, post-synaptic targets of the first-order bitter-sensitive interneurons and whether they receive excitatory or inhibitory input from these cells must await further investigation.97 Whether the same pathways are involved in detecting high salt, and evoke aversion toward high concentrations is the focus for future studies (Figure 4). Unraveling taste circuits, therefore, will be important not only for understanding how sensory inputs is translated to behavioral outputs but also how taste associations are formed in reward and aversive learning.8
Identifying salt pharyngeal neurons
To control behavioral feeding decisions, animals must simultaneously integrate external sensory stimuli with their internal state.107,108 Eat neural metabolic control of eating is regulated both by peripheral sensory detection of food and internal states like hunger and satiety.109-113 Dysregulation in these homeostatic systems can lead to metabolic conditions like obesity and other associated health problems. Ingestion is a poorly understood step in feeding behavior. In all animals, the optimization of food intake requires tight regulation of behaviors responsive to food quality and hunger state. After food ingestion, the nutrient sensing signals processed by the intestine take a relatively long period of time to mediate behavioral responses in the brain to change feeding rates.114,115
Stimulation of sweet taste neurons in the labellum and legs triggers an extension of the proboscis in starved flies, followed by initiation of food intake.101,116 Upon ingestion, the food comes in contact with pharyngeal taste neurons.11 Although function of pharyngeal taste neurons is poorly understood, a subset has been shown to regulate sugar ingestion.7 Only limited studies have investigated the dynamics of fly feeding using proboscis extension as a proxy for food intake.73-75,92 Studies performed on blowflies suggests that food intake is controlled by factors that stimulate ingestion, not the one that act on peripheral taste perception or post-ingestive nutrient-sensing.116 Neurons in the fly taste circuit that regulate different aspects of food intake behavior have been identified recently. Neuropeptide F and dopamine signaling enhance the sensitivity of labellar taste sensory neurons in hungry flies and increase their probability of initiating food intake.73-75 Perturbation of labellar sweet taste perception does not affect ingestion7 suggesting the labellar taste neuron circuitry likely regulates initial food evaluation, but not the later decision to ingest food. Recent work has identified interneurons that regulate the feeding motor program,90 GABAergic neurons that suppress nonselective ingestion,95 and motor neurons that regulate fluid ingestion.93 How these neurons connect taste sensory input to the motor output of ingestion, as well as how they interpret top-down information about hunger state is not known. Yapici et al20 propose that 12 cholinergic local interneurons (IN1) participate within this circuit as a key nodes that governs rapid food intake decisions. These neurons in the taste center of the fly brain regulate sucrose ingestion and receive selective input from sweet taste neurons in the pharynx.7 The identity of neurons like IN1 that will respond to high concentrations of salt and bitter compounds is still unknown (Figure 4). Analysis of pharyngeal GRN projections also suggests distinct connectivity to higher order neuronal circuits.19,20 A recently generated molecular map of pharyngeal taste organs, has opened venues for future investigations to study the roles of pharyngeal taste neurons in food evaluation and in controlling feeding behaviors. Further studies investigating the role of pharyngeal GRNs and pharyngeal taste circuits will provide insight into how internal taste signals are integrated with external taste to control various aspects of feeding behavior (Figure 4).
Salt representation in higher brain centers
The Mushroom body (MB) is a site for experiential learning in Drosophila.117-119 The dendrites of the MB principle cells, called Kenyon cells (KCs), receive sparse and random inputs from olfactory projection neurons (PNs). Evidences that the MB processes taste as CS (conditioned stimulus) and US (unconditioned stimuli) comes from behavioral taste conditioning experiments.120-122 Pairing sucrose stimulation to the leg (CS) with an aversive stimulus (US) causes short-term inhibition of proboscis extension in the proboscis extension response (PER) assay. Such learned behaviors requires the MB, but the neural processing in the MB that underlies taste conditioning is unknown. The salt taste projections to higher brain centers have not been characterized yet, therefore questions regarding the salt circuitry providing gustatory inputs to the MB remain unaddressed (Figure 4). However, a study122 provides direct evidence of multimodal inputs into the MB, with different representations for tastants of different modalities and different representations for different taste organs widening our understanding of the neural coding underlying conditioned learning and providing a basis for examining taste circuitry in the higher brain.
Work performed in rats with salt has provided information about how predictive evaluation can be strongly changed by internal nutrient deficits. Trained rats avoid a metal lever paired with aversive salt concentrations and avidly approach the same lever when sodium is deprived.26,123 Establishing similar paradigms in Drosophila could be equally informative.
State-dependent alterations in the salt taste circuit
Peripheral taste processing and the regulation of hunger states in vertebrates have been intensively studied. It has been shown that activation of sweet cells promotes food acceptance in hungry animals, while activation of bitter cells stimulates food avoidance.124,125 Neurons in the hypothalamic neuroendocrine circuits express proopiomelanocortin (POMC), agouti-related peptide (AgRP), and melanocortin receptor (MC4R) that coordinate ingestion in response to the hunger state of the animal.126-129 The mechanisms controlling taste and food intake in insects are remarkably similar as of vertebrates. Recent evidence in Drosophila suggest an increase in dopamine signaling enhancing the sensitivity of sweet gustatory project neurons (NP1562 neurons) to sucrose.92 Previously, it has been shown that starvation leads to increases in sucrose-evoked electrophysiological130,131 or calcium activity in GR5a+ taste neurons.74 It would be of interest to determine if there are state-dependent alterations in salt taste circuit activity that could lead to more consumption of salt like sugar, or consumption of higher salt concentrations (Figure 4). One needs to verify the possibilities if the information about starvation state is amplified during the relay to salt second-order neurons or if these neurons may also be targets of signaling pathways that convey information about the starvation state. How physiological state like hunger or adaptation to high salt act on these neurons that allows eating of high salt (aversive) concentrations in humans is a subject for future investigations.
Ir76b and other mechanisms for low salt detection
There are at least four kinds of chemoreceptors that function in the GRNs of D melanogaster including GRs, IRs, TRP (transient receptor potential) channels, and PPKs. The molecular identity of mammalian taste receptors (TRs) and insect GRs are very different. Taste receptors are GPCRs, but GRs are ion channels. The difference of GRs can be used to target insects while having minimal effects on humans. The other sensors such as TRPs, IRs, and PPKs are somehow conserved in vertebrates and invertebrates. Studies on the role of ionotropic cation channels in Drosophila taste recognition and regulation of attraction and avoidance behavior in taste are becoming more visible recently. The role of limited IRs in the taste system including IR76b, IR25a, and IR62a have been discovered recently. 132 and the function of other IR proteins in the taste system is still unexplored. It will be interesting to determine whether IRs function in concert with GRs, or whether they independently recognize other classes of tastants. The role of TRP channels, PPK proteins, or direct sensors involving GRs for detecting salt remain to be identified as well.8
The behavioral valence to salt depends on its concentration. Low salt is appetitive, whereas high salt is aversive. “Salt” neurons in L-type labellar sensilla display peak responses to around 100 mM NaCl and evoke appetitive behavior. IR76b-positive salt neurons show an attractive response to low salt and confer salt sensitivity when expressed in sweet neurons.44 Expression of IR76b has been observed in non-salt gustatory neurons, and in several classes of olfactory neurons that are likely salt insensitive.40 Whether, and how IR76b channel activity is gated in these neurons remains to be determined.
Similar to adult flies, the high salt responses are genetically separable from low salt response in larvae. Salt taste in larvae appears to be dependent on ppk genes. Both ppk11 and ppk19 genes are required for behavioral attraction to low salt and salt sensitivity in the terminal organ.25 As in adult flies, behavioral aversion to high salt relies on ppk19 and serrano.60 The ppk genes may not be necessary for salt taste in the adult fly, raising questions about why there exist 2 different molecular mechanisms for low salt.8
Understanding the role of sugar, bitter, and sour gustatory pathways in salt detection
Peripheral gustatory neurons in adult Drosophila84 express different members of the GR gene family and can be activated by salt with low threshold and by sugars (GR5a) and by salt with a high threshold and by bitter substances (GR66a). Additional studies are required to understand if such mechanisms operate in the same set of taste neurons that sense sugars and bitter compounds. Such studies will also shed light on mechanisms where loss of neuronal activity in sweet and bitter neurons can modulate behavioral valence to salt.
The taste of highly concentrated salt is shown to be aversive in animals ranging from nematodes to rodents.77,133,134 Even humans find high salt concentrations to have a bitter taste, therefore the aversive response to high salt concentrations may be more complex than previously thought. Electrophysiological studies performed on Drosophila adult taste sensilla have revealed that low- and high-NaCl concentrations are detected by 2 distinct gustatory neurons.45,135 L1 neurons respond to salt with a low threshold between 0.01 and 0.05 M, whereas the threshold for L2 neurons is about one order of magnitude higher concentration.135 In addition, the dpr locus (for defective proboscis extension response), a member of the Ig superfamily, has also been shown to be required for the aversive response to high-salt concentrations in adult flies.136
Recently, it has been shown134 that high-salt recruits 2 primary aversive taste pathways in mice by activating the sour and bitter taste-sensing cells. Genetic silencing of sour and bitter pathways eliminates behavioral aversion to high concentration of salt, without impairing salt attraction. Mice devoid of salt-aversion pathways exhibit unimpeded, continuous attraction even to exceedingly high concentrations of NaCl. These data suggest that “co-opting” of sour and bitter neural pathways evolved as a means to ensure that high levels of salt reliably trigger robust behavioral rejection, thus preventing its potentially detrimental effects in health and well-being. It would be interesting to dissect if similar pathways are involved in insects. As understanding of how low and high salt concentrations are differentially encoded is still unclear, future studies using specific inhibitors and activators of individual pathway should help address the contributions of the ENaC, T2R, and PKD2L1-expressing taste cells to human salt taste perception. These studies may serve as a catalyst for the development of selective receptor cell modulators to help control (and even satisfy) the strong appetite of the Western world for a high-salt diet, but without the potential ill effects of too much sodium.134
Footnotes
Funding:The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work is supported by Wellcome trust/DBT India Alliance Fellowship (grant number IA/I/15/2/502074) awarded to PK.
Declaration of conflicting interests:The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions: SK made the figures with help from PK; SK, RK and PK wrote the review.
ORCID iDs: Shivam Kaushik  https://orcid.org/0000-0001-8548-4136
https://orcid.org/0000-0001-8548-4136
Rahul Kumar
https://orcid.org/0000-0003-4624-1291
Pinky Kain
https://orcid.org/0000-0002-3579-6032
References
- 1. Kim H, Kirkhart C, Scott K. Long-range projection neurons in the taste circuit of Drosophila. Elife. 2017;6:e23386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Clyne PJ, Warr CG, Carlson JR. Candidate taste receptors in Drosophila. Science. 2000;287:1830-1834. [DOI] [PubMed] [Google Scholar]
- 3. Robertson HM, Warr CG, Carlson JR. Molecular evolution of the insect chemoreceptor gene superfamily in Drosophila melanogaster. Proc Natl Acad Sci U S A. 2003;100:14537-14542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Scott K, Brady R, Jr, Cravchik A, et al. A chemosensory gene family encoding candidate gustatory and olfactory receptors in Drosophila. Cell. 2001;104:661-673. [DOI] [PubMed] [Google Scholar]
- 5. Weiss LA, Dahanukar A, Kwon JY, Banerjee D, Carlson JR. The molecular and cellular basis of bitter taste in Drosophila. Neuron. 2011;69:258-272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Ling F, Dahanukar A, Weiss LA, Kwon JY, Carlson JR. The molecular and cellular basis of taste coding in the legs of Drosophila. J Neurosci. 2014;34:7148-7164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. LeDue EE, Chen YC, Jung AY, Dahanukar A, Gordon MD. Pharyngeal sense organs drive robust sugar consumption in Drosophila. Nat Commun. 2015;6:6667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Freeman EG, Dahanukar A. Molecular neurobiology of Drosophila taste. Curr Opin Neurobiol. 2016;34:140-148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Jeong YT, Shim J, Oh SR, et al. An odorant-binding protein required for suppression of sweet taste by bitter chemicals. Neuron. 2013;79:725-737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. French AS, Sellier MJ, Moutaz AA, et al. Dual mechanism for bitter avoidance in Drosophila. J Neurosci. 2015;35:3990-4004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Stocker RF. The organization of the chemosensory system in Drosophila melanogaster: a review. Cell Tissue Res. 1994;275:3-26. [DOI] [PubMed] [Google Scholar]
- 12. Vosshall LB, Stocker RF. Molecular architecture of smell and taste in Drosophila. Annu Rev Neurosci. 2007;30:505-533. [DOI] [PubMed] [Google Scholar]
- 13. Gerber B, Stocker RF. The Drosophila larva as a model for studying chemosensation and chemosensory learning: a review. Chem Senses. 2007;32:65-89. [DOI] [PubMed] [Google Scholar]
- 14. Kwon JY, Dahanukar A, Weiss LA, Carlson JR. Molecular and cellular organization of the taste system in the Drosophila larva. J Neurosci. 2011;31:15300-15309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Choi J, van Giesen L, Choi MS, Kang K, Sprecher SG, Kwon JY. A pair of pharyngeal gustatory receptor neurons regulates caffeine-dependent ingestion in Drosophila larvae. Front Cell Neurosci. 2016;10:181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Kim H, Choi MS, Kang K, Kwon JY. Behavioral analysis of bitter taste perception in Drosophila larvae. Chem Senses. 2016;41:85-94. [DOI] [PubMed] [Google Scholar]
- 17. Mishra D, Miyamoto T, Rezenom YH, et al. The molecular basis of sugar sensing in Drosophila larvae. Curr Biol. 2013;23:1466-1471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Tissot M, Stocker RF. Metamorphosis in Drosophila and other insects: the fate of neurons throughout the stages. Prog Neurobiol. 2000;62:89-111. [DOI] [PubMed] [Google Scholar]
- 19. Chen YC, Dahanukar A. Molecular and cellular organization of taste neurons in adult Drosophila pharynx. Cell Rep. 2017;21:2978-2991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Yapici N, Cohn R, Schusterreiter C, Ruta V, Vosshall LB. A taste circuit that regulates ingestion by integrating food and hunger signals. Cell. 2016;165:715-729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Gendre N, Luer K, Friche S, et al. Integration of complex larval chemosensory organs into the adult nervous system of Drosophila. Development. 2004;131:83-92. [DOI] [PubMed] [Google Scholar]
- 22. Stocker RF. Taste perception: Drosophila—a model of good taste. Curr Biol. 2004;14:R560-R561. [DOI] [PubMed] [Google Scholar]
- 23. Arora K, Rodrigues V, Joshi S, Shanbhag S, Siddiqi O. A gene affecting the specificity of the chemosensory neurons of Drosophila. Nature. 1987;330:62-63. [DOI] [PubMed] [Google Scholar]
- 24. Miyakawa Y. Bimodal response in a chemotactic behavior of Drosophila larvae to monovalent salts. J Insect Physiol. 1981;27:387-392. [Google Scholar]
- 25. Liu L, Leonard AS, Motto DG, et al. Contribution of Drosophila DEG/ENaC genes to salt taste. Neuron. 2003;39:133-146. [DOI] [PubMed] [Google Scholar]
- 26. Chandrashekar J, Kuhn C, Oka Y, et al. The cells and peripheral representation of sodium taste in mice. Nature. 2010;464:297-301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. McDonald FJ, Price MP, Snyder PM, Welsh MJ. Cloning and expression of the beta and gamma subunits of the human epithelial sodium channel. Am J Physiol. 1995;268:C1157-C1163. [DOI] [PubMed] [Google Scholar]
- 28. Halpern BP. Amiloride and vertebrate gustatory responses to NaCl. Neurosci Biobehav Rev. 1998;23:5-47. [DOI] [PubMed] [Google Scholar]
- 29. Lindemann B. Taste reception. Physiol Rev. 1996;76:719-766. [DOI] [PubMed] [Google Scholar]
- 30. Canessa CM, Schild L, Buell G, et al. Amiloride-sensitive epithelial Na channel is made of three homologous subunits. Nature. 1994;367:463-467. [DOI] [PubMed] [Google Scholar]
- 31. Lin W, Finger TE, Rossier BC, Kinnamon SC. Epithelial Na channel subunits in rat taste cells: localization and regulation by aldosterone. J Comp Neurol. 1999;405:406-420. [DOI] [PubMed] [Google Scholar]
- 32. Kretz O, Barbry P, Bock R, Lindemann B. Differential expression of RNA and protein of the three pore-forming subunits of the amiloride-sensitive epithelial sodium channel in taste buds of the rat. J Histochem Cytochem. 1999;47:51-64. [DOI] [PubMed] [Google Scholar]
- 33. Boughter JD, Jr, Smith DV. Amiloride blocks acid responses in NaCl-best gustatory neurons of the hamster solitary nucleus. J Neurophysiol. 1998;80:1362-1372. [DOI] [PubMed] [Google Scholar]
- 34. Scott TR, Giza BK. Coding channels in the taste system of the rat. Science. 1990;249:1585-1587. [DOI] [PubMed] [Google Scholar]
- 35. Garty H, Palmer LG. Epithelial sodium channels: function, structure, and regulation. Physiol Rev. 1997;77:359-396. [DOI] [PubMed] [Google Scholar]
- 36. Lundy RF, Jr, Pittman DW, Contreras RJ. Role for epithelial Na channels and putative Na/H exchangers in salt taste transduction in rats. Am J Physiol. 1997;273:R1923-R1931. [DOI] [PubMed] [Google Scholar]
- 37. Littleton JT, Ganetzky B. Ion channels and synaptic organization: analysis of the Drosophila genome. Neuron. 2000;26:35-43. [DOI] [PubMed] [Google Scholar]
- 38. Liu L, Johnson WA, Welsh MJ. Drosophila DEG/ENaC pocket genes are expressed in the tracheal system may be involved in liquid clearance. Proc Natl Acad Sci U S A. 2003;100:2128-2133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Singh RN. Neurobiology of the gustatory systems of Drosophila and some terrestrial insects. Microsc Res Tech. 1997;39:547-563. [DOI] [PubMed] [Google Scholar]
- 40. Benton R, Vannice KS, Gomez-Diaz C, Vosshall LB. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell. 2009;136:149-162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Silbering AF, Rytz R, Grosjean Y, et al. Complementary function and integrated wiring of the evolutionarily distinct Drosophila olfactory subsystems. J Neurosci. 2011;31:13357-13375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Ni L, Klein M, Svec KV, et al. The ionotropic receptors IR21a and IR25a mediate cool sensing in Drosophila. Elife. 2016;5:e13254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Knecht ZA, Silbering AF, Ni L, et al. Distinct combinations of variant ionotropic glutamate receptors mediate thermosensation and hygrosensation in Drosophila. Elife. 2016;5:44-60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Zhang YV, Ni J, Montell C. The molecular basis for attractive salt-taste coding in Drosophila. Science. 2013;80:340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Hiroi M, Meunier N, Marion-Poll F, Tanimura T. Two antagonistic gustatory receptor neurons responding to sweet-salty and bitter taste in Drosophila. J Neurobiol. 2004;61:333-342. [DOI] [PubMed] [Google Scholar]
- 46. Lee MJ, Sung HY, Jo H, et al. Ionotropic receptor 76b is required for gustatory aversion to excessive Na+ in Drosophila. Mol Cells. 2017;40:787-795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Croset V, Schleyer M, Arguello JR, Gerber B, Benton R. A molecular and neuronal basis for amino acid sensing in the Drosophila larva. Sci Rep. 2016;6:34871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Ganguly A, Pang L, Duong V-K, et al. A molecular and cellular context- dependent role for Ir76b in detection of amino acid taste. Cell Rep. 2017;18:737-750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Mun HC, Franks AH, Culverston EL, Krapcho K, Nemeth EF, Conigrave AD. The Venus Fly Trap domain of the extracellular Ca2+-sensing receptor is required for L-amino acid sensing. J Biol Chem. 2004;279:51739-51744. [DOI] [PubMed] [Google Scholar]
- 50. Plested AJR, Vijayan R, Biggin PC, Mayer ML. Molecular basis of kainate receptor modulation by sodium. Neuron. 2008;58:720-735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Brand JG, Teeter JH, Kumazawa T, Huque T, Bayley DL. Transduction mechanisms for the taste of amino acids. Physiol Behav. 1991;49:899-904. [DOI] [PubMed] [Google Scholar]
- 52. Chung KM, Lee SB, Heur R, et al. Glutamate-induced cobalt uptake elicited by kainate receptors in rat taste bud cells. Chem Senses. 2005;30:137-143. [DOI] [PubMed] [Google Scholar]
- 53. Henkin RI, Christiansen RL. Taste localization on the tongue, palate, and pharynx of normal man. J Appl Physiol. 1967;22:316-320. [DOI] [PubMed] [Google Scholar]
- 54. Ishimaru Y, Okada S, Naito H, et al. Two families of candidate taste receptors in fishes. Mech Dev. 2005;122:1310-1321. [DOI] [PubMed] [Google Scholar]
- 55. Travers SP, Nicklas K. Taste bud distribution in the rat pharynx and larynx. Anat Rec. 1990;227:373-379. [DOI] [PubMed] [Google Scholar]
- 56. Frank ME. Taste-responsive neurons of the glossopharyngeal nerve of the rat. J Neurophysiol. 1991;65:1452-1463. [DOI] [PubMed] [Google Scholar]
- 57. Hanamori T, Miller IJ, Jr, Smith DV. Gustatory responsiveness of fibers in the hamster glossopharyngeal nerve. J Neurophysiol. 1988;60:478-498. [DOI] [PubMed] [Google Scholar]
- 58. Carleton A, Accolla R, Simon SA. Coding in the mammalian gustatory system. Trends Neurosci. 2010;33:326-334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Morton GJ, Cummings DE, Baskin DG, Barsh GS, Schwartz MW. Central nervous system control of food intake and body weight. Nature. 2006;443:289-295. [DOI] [PubMed] [Google Scholar]
- 60. Alves G, Salle J, Chaudy S, Dupas S, Maniere G. High-NaCl perception in Drosophila melanogaster. J Neurosci. 2014;34:10884-10891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Chung S, Vining MS, Bradley PL, Chan CC, Wharton KA, Jr, Andrew DJ. Serrano (sano) functions with the planar cell polarity genes to control tracheal tube length. PLoS Genet. 2009;5:e1000746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Gloerich M, Bos JL. Epac: defining a new mechanism for cAMP action. Annu Rev Pharmacol Toxicol. 2010;50:355-375. [DOI] [PubMed] [Google Scholar]
- 63. Barton Browne L. Ontogenetic changes in feeding behavior. In: Chapman RF, de Boer G. eds. Regulatory Mechanisms in Insect Feeding. New York, NY: Chapman & Hall; 1995:307-342. [Google Scholar]
- 64. Richter CP, Barelare B. Nutritional requirements of pregnant and lactating rats studied by the self-selection method. Endocrinology. 1938;23:15-24. [Google Scholar]
- 65. Denton DA, Nelson JF. The effects of pregnancy and lactation on the mineral appetites of wild rabbits (Oryctolagus cuniculus (L.)). Endocrinology. 1971;88:31-40. [DOI] [PubMed] [Google Scholar]
- 66. Friend DW, Wolynetz MS. Self-selection of salt by gilts during pregnancy and lactation. Can J Anim Sci. 1981;61:429-438. [Google Scholar]
- 67. McBurnie MI, Blair-West JR, Denton DA, Weisinger RS. Sodium intake and reproduction in BALB/C mice. Physiol Behav. 1999;66:873-879. [DOI] [PubMed] [Google Scholar]
- 68. Whitehead CC, Shannon DWF. The control of egg production using a low-sodium diet. Br Poult Sci. 1974;15:429-434. [Google Scholar]
- 69. Chou R, Hara A, Du D, Shimizu N, Sakuyama H, Uehara Y. Low-salt intake during mating or gestation in rats is associated with low birth and survival rates of babies. J Nutr Metab. 2014;2014:212089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Walker S, Corrales-Carvajal V, Ribeiro C. Postmating circuitry modulates salt taste processing to increase reproductive output in Drosophila. Curr Biol. 2015;25:2621-2630. [DOI] [PubMed] [Google Scholar]
- 71. Itskov PM, Ribeiro C. The dilemmas of the gourmet fly: the molecular and neuronal mechanisms of feeding and nutrient decision making in Drosophila. Front Neurosci. 2013;7:12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Saveer AM, Kromann SH, Birgersson G, et al. Floral to green: mating switches moth olfactory coding and preference. Proc Biol Sci. 2012;279:2314-2322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Marella S, Mann K, Scott K. Dopaminergic modulation of sucrose acceptance behavior in Drosophila. Neuron. 2012;73:941-950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Inagaki HK, Ben-Tabou de-Leon S, Wong AM, et al. Visualizing neuromodulation in vivo: TANGO-mapping of dopamine signaling reveals appetite control of sugar sensing. Cell. 2012;148:583-595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Inagaki HK, Panse KM, Anderson DJ. Independent, reciprocal neuromodulatory control of sweet and bitter taste sensitivity during starvation in Drosophila. Neuron. 2014;84:806-820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Dey S, Chamero P, Pru JK, et al. Cyclic regulation of sensory perception by a female hormone alters behavior. Cell. 2015;161:1334-1344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Niewalda T, Singhal N, Fiala A, Saumweber T, Wegener S, Gerber B. Salt processing in larval Drosophila: choice, feeding, and learning shift from appetitive to aversive in a concentration-dependent way. Chem Senses. 2008;33:685-692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Schipanski A, Yarali A, Niewalda T, Gerber B. Behavioral analyses of sugar processing in choice, feeding, and learning in larval Drosophila. Chem Senses. 2008;33:563-573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Liman ER, Zhang YV, Montell C. Peripheral coding of taste. Neuron. 2014;81:984-1000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Yarmolinsky DA, Zuker CS, Ryba NJ. Common sense about taste: from mammals to insects. Cell. 2009;139:234-244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Dahanukar A, Foster K, van der Goes van Naters WM, Carlson JR. A Gr receptor is required for response to the sugar trehalose in taste neurons of Drosophila. Nat Neurosci. 2001;4:1182-1186. [DOI] [PubMed] [Google Scholar]
- 82. Dahanukar A, Lei Y-T, Kwon JY, Carlson JR. Two Gr genes underlie sugar reception in Drosophila. Neuron. 2007;56:503-516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Jiao Y, Moon SJ, Wang X, Ren Q, Montell C. Gr64f is required in combination with other gustatory receptors for sugar detection in Drosophila. Curr Biol. 2008;18:1797-1801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Marella S, Fischler W, Kong P, Asgarian S, Rueckert E, Scott K. Imaging taste responses in the fly brain reveals a functional map of taste category and behavior. Neuron. 2006;49:285-295. [DOI] [PubMed] [Google Scholar]
- 85. Thorne N, Chromey C, Bray S, Amrein H. Taste perception and coding in Drosophila. Curr Biol. 2004;14:1065-1079. [DOI] [PubMed] [Google Scholar]
- 86. Wang Z, Singhvi A, Kong P, Scott K. Taste representations in the Drosophila brain. Cell. 2004;117:981-991. [DOI] [PubMed] [Google Scholar]
- 87. Cameron P, Hiroi M, Ngai J, Scott K. The molecular basis for water taste in Drosophila. Nature. 2010;465:91-95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Chen Z, Wang Q, Wang Z. The amiloride-sensitive epithelial Na+ channel PPK28 is essential for Drosophila gustatory water reception. J Neurosci. 2010;30:6247-6252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Ito K, Shinomiya K, Ito M, et al. A systematic nomenclature for the insect brain. Neuron. 2014;81:755-765. [DOI] [PubMed] [Google Scholar]
- 90. Flood TF, Iguchi S, Gorczyca M, White B, Ito K, Yoshihara M. A single pair of interneurons commands the Drosophila feeding motor program. Nature. 2013;499:83-87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Gordon MD, Scott K. Motor control in a Drosophila taste circuit. Neuron. 2009;61:373-384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Kain P, Dahanukar A. Secondary taste neurons that convey sweet taste and starvation in the Drosophila brain. Neuron. 2015;85:819-832. [DOI] [PubMed] [Google Scholar]
- 93. Manzo A, Silies M, Gohl DM, Scott K. Motor neurons controlling fluid ingestion in Drosophila. Proc Natl Acad Sci U S A. 2012;109:6307-6312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Miyazaki T, Lin TY, Ito K, Lee CH, Stopfer M. A gustatory second-order neuron that connects sucrose-sensitive primary neurons and a distinct region of the gnathal ganglion in the Drosophila brain. J Neurogenet. 2015;29:144-155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95. Pool AH, Kvello P, Mann K, et al. Four GABAergic interneurons impose feeding restraint in Drosophila. Neuron. 2014;83:164-177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Rajashekhar KP, Singh RN. Organization of motor neurons innervating the proboscis musculature in Drosophila melanogaster meigen (Diptera: Drosophilidae). Int J Insect Morphol Embryol. 1994;23:225-242. [Google Scholar]
- 97. Bohra AA, Kallman BR, Reichert H, VijayRaghavan K. Identification of a single pair of interneurons for bitter taste processing in the Drosophila brain. Curr Biol. 2018;28:19847-19858. [DOI] [PubMed] [Google Scholar]
- 98. Albin SD, Kaun KR, Knapp JM, Chung P, Heberlein U, Simpson JH. A subset of serotonergic neurons evokes hunger in adult Drosophila. Curr Biol. 2015;25:2435-2440. [DOI] [PubMed] [Google Scholar]
- 99. Root CM, Ko KI, Jafari A, Wang JW. Presynaptic facilitation by neuropeptide signaling mediates odor-driven food search. Cell. 2011;145:133-144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100. Charlu S, Wisotsky Z, Medina A, Dahanukak A. Acid sensing by sweet and bitter taste neurons in Drosophila melanogaster. Nat Commun. 2013;4:2042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Dethier VG. The Hungry Fly. Cambridge, MA: Harvard University Press; 1976. [Google Scholar]
- 102. Harris DT, Kallman BR, Mullaney BC, Scott K. Representations of taste modality in the Drosophila brain. Neuron. 2015;86:1449-1460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103. Accolla R, Bathellier B, Petersen CC, Carleton A. Differential spatial representation of taste modalities in the rat gustatory cortex. J Neurosci. 2007;27:1396-1404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Barretto RP, Gillis-Smith S, Chandrashekar J, et al. The neural representation of taste quality at the periphery. Nature. 2015;517:373-376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105. Chen X, Gabitto M, Peng Y, Ryba NJ, Zuker CS. A gustotopic map of taste qualities in the mammalian brain. Science (New York, NY). 2011;333:1262-1266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Hückesfeld S, Peters M, Pankratz MJ. Central relay of bitter taste to the protocerebrum by peptidergic interneurons in the Drosophila brain. Nat Commun. 2016;7:12796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Davis WJ. Behavioural hierarchies. Trends Neurosci. 1979;2:5-7. [Google Scholar]
- 108. Tinbergen N. The Study of Instinct. Oxford, UK: Oxford University Press; 1951. [Google Scholar]
- 109. Burton MJ, Rolls ET, Mora F. Effects of hunger on the responses of neurons in the lateral hypothalamus to the sight and taste of food. Exp Neurol. 1976;51:668-677. [DOI] [PubMed] [Google Scholar]
- 110. Kennedy GC. The role of depot fat in the hypothalamic control of food intake in the rat. Proc R Soc Lond B Biol Sci. 1953;140:578-596. [DOI] [PubMed] [Google Scholar]
- 111. Mayer J. Glucostatic mechanism of regulation of food intake. N Engl J Med. 1953;249:13-16. [DOI] [PubMed] [Google Scholar]
- 112. Raubenheimer D, Simpson SJ. Integrative models of nutrient balancing: application to insects and vertebrates. Nutr Res Rev. 1997;10:151-179. [DOI] [PubMed] [Google Scholar]
- 113. Read N, French S, Cunningham K. The role of the gut in regulating food intake in man. Nutr Rev. 1994;52:1-10. [DOI] [PubMed] [Google Scholar]
- 114. Dus M, Lai JSY, Gunapala KM, et al. Nutrient sensor in the brain directs the action of the brain-gut axis in Drosophila. Neuron. 2015;87:139-151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115. Miyamoto T, Slone J, Song X, Amrein H. A fructose receptor functions as a nutrient sensor in the Drosophila brain. Cell. 2012;151:1113-1125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116. Dethier VG, Evans DR, Rhoades MV. Some factors controlling the ingestion of carbohydrates by the blowfly. Biol Bull. 1956;111:204-222. [Google Scholar]
- 117. Heisenberg M. Mushroom body memoir: from maps to models. Nat Rev Neurosci. 2003;4:266-275. [DOI] [PubMed] [Google Scholar]
- 118. Davis RL. Olfactory memory formation in Drosophila: from molecular to systems neuroscience. Annu Rev Neurosci. 2005;28:275-302. [DOI] [PubMed] [Google Scholar]
- 119. Keene AC, Waddell S. Drosophila olfactory memory: single genes to complex neural circuits. Nat Rev Neurosci. 2007;8:341-354. [DOI] [PubMed] [Google Scholar]
- 120. Masek P, Scott K. Limited taste discrimination in Drosophila. Proc Natl Acad Sci U S A. 2010;107:14833-14838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Keene AC, Masek P. Optogenetic induction of aversive taste memory. Neuroscience. 2012;222:173-180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122. Kirkhart C, Scott K. Gustatory learning and processing in the Drosophila mushroom bodies. J Neurosci. 2015;35:5950-5958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123. Robinson MJF, Berridge KC. Instant transformation of learned repulsion into motivational “wanting.” Curr Biol. 2013;23:282-289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Mueller KL, Hoon MA, Erlenbach I, Chandrashekar J, Zuker CS, Ryba NJP. The receptors and coding logic for bitter taste. Nature. 2005;434:225-229. [DOI] [PubMed] [Google Scholar]
- 125. Zhao GQ, Zhang Y, Hoon MA, et al. The receptors for mammalian sweet and umami taste. Cell. 2003;115:255-266. [DOI] [PubMed] [Google Scholar]
- 126. Aponte Y, Atasoy D, Sternson SM. AGRP neurons are sufficient to orchestrate feeding behavior rapidly and without training. Nat Neurosci. 2011;14:351-355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Atasoy D, Betley JN, Su HH, Sternson SM. Deconstruction of a neural circuit for hunger. Nature. 2012;488:172-177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128. Carter ME, Soden ME, Zweifel LS, Palmiter RD. Genetic identification of a neural circuit that suppresses appetite. Nature. 2013;503:111-114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129. Fan W, Boston BA, Kesterson RA, Hruby VJ, Cone RD. Role of melanocortinergic neurons in feeding and the agouti obesity syndrome. Nature. 1997;385:165-168. [DOI] [PubMed] [Google Scholar]
- 130. Meunier N, Belgacem YH, Martin JR. Regulation of feeding behaviour and locomotor activity by takeout in Drosophila. J Exp Biol. 2007;210:1424-1434. [DOI] [PubMed] [Google Scholar]
- 131. Nishimura A, Ishida Y, Takahashi A, et al. Starvation-induced elevation of taste responsiveness and expression of a sugar taste receptor gene in Drosophila melanogaster. J Neurogenet. 2012;26:206-215. [DOI] [PubMed] [Google Scholar]
- 132. Lee Y, Poudel S, Kim Y, Thakur D, Montell C. Calcium taste avoidance in Drosophila. Neuron. 2018;97:67-74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133. Chatzigeorgiou M, Bang S, Hwang SW, Schafer WR. tmc-1 encodes a sodium-sensitive channel required for salt chemosensation in C. elegans. Nature. 2013;494:95-99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134. Oka Y, Butnaru M, von Buchholtz L, Ryba NJP, Zuker CS. High salt recruits aversive taste pathways. Nature. 2013;494:472-475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135. Ishimoto H, Tanimura T. Molecular neurophysiology of taste in Drosophila. Cell Mol Life Sci. 2004;61:10-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136. Nakamura M, Baldwin D, Hannaford S, Palka J, Montell C. Defective proboscis extension response (DPR), a member of the Ig superfamily required for the gustatory response to salt. J Neurosci. 2002;22:3463-3472. [DOI] [PMC free article] [PubMed] [Google Scholar]