

Abstract
Mitochondria are highly dynamic and regulated organelles that historically have been defined based on their crucial role in cell metabolism. However, they are implicated in a variety of other important functions, making mitochondrial dysfunction an important axis in several pathological contexts. Despite that conventional biochemical and molecular biology approaches have provided significant insight into mitochondrial functionality, innovative techniques that provide a global view of the mitochondrion are still necessary. Proteomics fulfils this need by enabling accurate, systems-wide quantitative analysis of protein abundance. More importantly, redox proteomics approaches offer unique opportunities to tackle oxidative stress, a phenomenon that is intimately linked to aging, cardiovascular disease, and cancer. In addition, cutting-edge proteomics approaches reveal how proteins exert their functions in complex interaction networks where even subtle alterations stemming from early pathological states can be monitored. Here, we describe the proteomics approaches that will help to deepen the role of mitochondria in health and disease by assessing not only changes to mitochondrial protein composition but also alterations to their redox state and how protein interaction networks regulate mitochondrial function and dynamics. This review is aimed at showing the reader how the application of proteomics approaches during the last 20 years has revealed crucial mitochondrial roles in the context of aging, neurodegenerative disorders, metabolic disease, and cancer.
1. Introduction
Mitochondria represent the metabolic dynamism of the cell, and they are present in the cytoplasm of all eukaryotic cells relying on aerobic metabolism [1]. These intracellular organelles are involved in a number of vital processes including the generation of adenosine triphosphate (ATP) by oxidative phosphorylation (OXPHOS), the determination of the redox state through the generation and detoxification of reactive nitrogen and oxygen species (RNOS), the regulation of calcium homeostasis, and cell death [2, 3]. Mitochondrial activity comprises various processes, including the activity of the mitochondrial electron transport chain (ETC) complexes, substrate oxidation through the tricarboxylic acid (TCA) cycle and ATP synthesis, β-oxidation of fatty acids, ketogenesis, and biosynthesis of pyrimidines, heme groups, and urea [4, 5]. Additionally, mitochondria sense oxygen and nutrient levels, providing the energy required for other cellular processes such as muscle contraction or heat production, and maintain ionic gradients and membrane potentials in excitable cells, allowing the secretion of hormones and neurotransmitters [6].
Due to the huge number of cellular processes in which this organelle is implicated, even in the closed context of cellular respiration, the term mitochondrial function can refer to a variety of features. In fact, the definition of mitochondrial capacity has been broadened during the last decade to include additional features such as mitochondrial dynamics, which includes fusion and fission events, turnover (biogenesis and mitophagy), and plasticity [7]. The consideration of these mitochondrial capabilities has advanced our understanding of how the cell may respond to metabolic disturbances such as oxidative stress, inflammation, and gluco- and lipotoxicity [8], among others. Moreover, mitochondrial dynamics is influenced by energy expenditure and nutrient supply, and changes in mitochondrial architecture have emerged as a new mechanism of adaptation to metabolic demand [9]. Therefore, the mitochondrion is both origin and target of several metabolic signals which orchestrate cellular function and homeostasis [4].
Mitochondrial function can be tackled through in vivo, ex vivo, and in vitro methods, which have been extensively reviewed [7, 10–13]. In general, measuring aspects of mitochondrial function in vivo is preferable to in vitro assessments, although this approach might be less specific and even become unattainable depending on the type of sample under study. Despite that these in vivo, ex vivo, and in vitro approaches provide invaluable data on specific mitochondrial functions (e.g., submaximal and maximal ADP-stimulated OXPHOS capacity, TCA cycle flux, rates of fatty acid uptake, and RNOS production), they have limitations. Thus, the invasiveness of the method, the indirect measurement of the parameter, the impossibility to discriminate between mitochondrial and cytosolic shared functions, the expensiveness of the hardware/methodology, and/or the lack of data on mitochondrial density usually hamper the application of these methodologies in the study of mitochondrial function [7]. In addition, while the aforementioned approaches address mitochondrial functions individually, the central role of mitochondria in cellular metabolism and maintenance demands the application of additional innovative techniques that have the capacity to provide a global view of mitochondria in a single experiment. Although in an indirect manner, proteomics offers this functional snapshot thanks to the robustness, velocity, sensitiveness, and precision that mass spectrometry (MS) has acquired during the last decades. MS is recognized as the method of choice for both protein identification and quantification, and it has been the driving force for proteomics research development [14, 15].
Here, we describe innovative approaches to the quantitation of the mitochondrial proteome, with an emphasis on the oxidative modifications involved. In addition, we include cutting-edge experiments aimed at unveiling the mitochondrial interactome, since to deepen the role of mitochondria in health and disease it is essential to monitor not only the alterations to mitochondrial protein composition, but also how these proteins organize in interaction networks that regulate mitochondrial function and dynamics. The scope of the present review is to show the reader how the application of proteomics methodologies during the last 20 years has discovered key mitochondrial roles in the context of aging, neurodegenerative disorders, metabolic disease, and cancer.
2. The Mitochondrial Proteome
As a reminiscence of its prokaryotic past, the mitochondrion hosts its own genome. In mammalian cells, the mitochondrial DNA (mtDNA) only encodes 13 proteins belonging to the mitochondrial respiratory chain, 2 ribosomal RNAs, and 22 transfer RNAs, determining its intramitochondrial translation code, which is markedly different from the extramitochondrial one and is tightly regulated to maintain mitochondrial function [16]. The remaining mitochondrial proteins, i.e., the components of the TCA cycle, β-oxidation and protein transport, the other respiratory chain subunits, and the apoptotic factors, are all nuclear-encoded, which makes the definition of the mitochondrial proteome extremely challenging.
Since most mitochondrial proteins have to be imported into the organelle, there are several mechanisms whereby they can be transported [17, 18]. The import of cytosolically synthesized mitochondrial proteins usually requires cytosolic chaperones, such as heat shock proteins 70 and 90. These chaperones guide pre-proteins to mitochondrial outer membrane receptors (primarily TOM70 and TOM20) of the TOM (translocase of the outer membrane) complex [19]. The successive import into the diverse suborganelle compartments (that is, outer membrane (OM), intermembrane space (IMS), inner membrane (IM), or mitochondrial matrix) depends on several targeting signals recognized by various mitochondrial protein import complexes (reviewed in [17]). Of note, the characterization of key components and pathways involved in protein targeting has been extensively described over the last three decades, including oxidative folding machinery in the IMS, which contributes to the redox-dependent control of proteostasis [20]. Moreover, protein sorting could be differentially regulated among different organisms and physiological or pathological conditions [21], currently constituting an area of intensive research [20, 22, 23].
2.1. Key Aspects of the Mitochondrial Proteome
To date, the wealth of data that have largely contributed to define the mitochondrial proteome comes from proteomics studies performed in a number of eukaryotic organisms. In the human context, the majority of the studies have been carried out in connection with disease research [24]. Hence, in the last 20 years, the study of the mitochondrial proteome has elucidated several properties about this extraordinary organelle, some of which are outlined below.
2.1.1. Evolutionary Origin and Mitochondrial Proteome Composition
Evidence suggests that current mitochondrion derives from an α-proteobacterial endosymbiont which transferred its DNA to the host nucleus [25]. It is estimated that only 15–20% of the current human mitochondrial proteome derives from the original endosymbiont DNA [26], although mammalian mitochondrial proteins are more conserved (nearly 75%) compared to the rest of cellular components (around 48% of evolutionary conservation) [27, 28]. In fact, phylogenetic profiling has been very useful in elucidating the function of several mitochondrial proteins [28, 29].
At present, the number of mammalian mitochondrial proteins ranges from 1100 to 1900 depending on the database and resource used. In the last decade, despite of the broad dynamic range (up to six orders of magnitude) that mitochondrial proteins exhibit, which hinders the detection of less abundant proteins, MS analyses have significantly contributed to the description of the mitochondrial proteome [30, 31]. These results have underscored remarkable quantitative differences across different tissues.
2.1.2. Tissue Heterogeneity
Although the mitochondrial function is essential for almost all cell types (with the exception of red blood cells, which lack mitochondria), there are tissue-specific differences. Large-scale studies have provided valuable information about tissue or cell specificity. Thus, heart mitochondria are not only over 5-fold more abundant than adipose tissue mitochondria but also qualitatively different. In their seminal proteomics comparison across tissues, Mootha et al. found that mitochondria from different tissues shared approximately 75% of their components [32], and these data have been repeatedly confirmed with the subsequent extensive analyses of the organelle. As later revealed by Pagliarini and colleagues, mitochondria from developmentally related organs tend to share more proteins [28]. Out of the ∼1100 mitochondrial proteins they initially described, nearly half of them appear to be core components virtually found in all tissues; however, some of these pathways showed striking patterns of tissue diversity. According to their results, the most representative mitochondrial proteome could be found in skeletal muscle followed by heart tissue, since both proteomes include the least proportion of less abundant mitochondrial proteins [28, 33]. Interestingly, respiratory complexes I (CI), II (CII), III (CIII), and V (CV) are found in high abundance in all the tissues surveyed in contrast to complex IV (CIV), which seems to have a reasonable number of subunits expressed in a tissue-dependent manner. In fact, protein expression strongly differs in complex IV and large ribosomal subunits of mitochondria between heart and adipose tissue, most probably responding to the differences in the energetic demand between these tissues [28, 33].
Apart from the differences in ETC complexes, some mitochondrial proteins and isoenzymes are preferentially, or even uniquely, expressed in certain cell types. For example, uncoupling protein-1 (UCP1) is a distinctive marker of mitochondria from brown adipocytes [34]. This protein supports the nonshivering thermogenesis by the uncoupling of mitochondrial respiration which characterizes brown adipose tissue.
Remarkably, the characterization of tissue-specific mitochondrial proteomes has enabled the description of the biosynthetic capacities of different organs and the construction of flux balance analysis models [35–37]. These innovative studies represent the use of the mitochondrial proteome as a framework for systems physiology analyses [38–40].
2.1.3. Dual Localization of Mitochondrial Proteins
Since different compartments can exhibit similar functions, it is not surprising that proteins can be dually localized in the cell. Generally, dual localization refers to those protein products from a single gene locus that are present in multiple subcellular locations [41]. In the case of mitochondria, it is noteworthy that they actively interact with other subcellular compartments like the endoplasmic reticulum, the nucleus, or the peroxisomes, which favours the dual localization of proteins. Thus, it has been estimated that around 15–30% of mitochondrial proteins are dual-localized [30, 42]. This dual localization can be achieved through several mechanisms, including alternative splicing of the same gene product or alternative start sites from the same gene locus, which produce distinct proteins. Additionally, some proteins may intermittently interact in the surface of mitochondria (e.g., hexokinase binds to the channel protein VDAC1 in order to release intramitochondrial ATP for glucose phosphorylation [43]), while others may translocate from the mitochondria to the cytosol in certain conditions (a typical example is the release of cytochrome c under proapoptotic conditions [44]) or vice versa (e.g., BID protein transfers from the cytosol into the mitochondrion after cell death stimulus [45]). In addition, certain proteins can localize to two different compartments under similar physiological conditions (e.g., the enzyme complex pyruvate dehydrogenase, which participates in the TCA cycle in the mitochondria, whereas in the nucleus it produces histone acetylation as part of the epigenetic regulation [46]).
In this complex scenario, the reports on the mitochondrial localization of proteins not usually associated with mitochondria or the localization of well-known mitochondrial proteins outside the organelle must be carefully considered. The validation of these results could be quite challenging and should involve at least two independent methods. In this regard, quantitative proteomics analysis of the total proteome and the corresponding mitochondrial subproteome could reveal even subtle quantitative differences originated by dual localization of mitochondrial proteins [24]. Additionally, other resources such as The Human Protein Atlas (http://www.proteinatlas.org) could provide orthogonal evidence to proteomic results based on subcellular localization data obtained through antibody-based techniques [47].
2.1.4. The Mitochondrial Subproteome
As expected from cellular complexity, not only organellar localization but also the precise compartmentalization within the proper organelle is important for protein function, adding an additional level of complexity to the study of the mitochondrial proteome. For instance, the IMS is key to ATP production by storing the electromotive force generated by the ETC, which in turn localizes to the OM (and proteins localized to membranes have very different properties from those found in the mitochondrial matrix or the cytosol). Nevertheless, up to date only a few proteomic studies have analysed the human mitochondrial subproteome. In this regard, Rhee and colleagues have greatly contributed to the description of the mitochondrial matrix [38] and IMS proteomes [48] by applying MS-compatible tagging techniques as APEX [49]. Among other advantages, APEX strategy selectively tags proteins that localize to any specific region in living cells, circumventing the requirement for organellar purification when applied to membrane-bound compartments. This in vivo system is based on ascorbate peroxidase (APX) activity, which produces biotin-phenol radicals that can conjugate to the side chain of amino acids in proteins only in its immediate proximity. Thus, once biotin-labelled proteins are obtained, they can be pulled down and subsequently analysed by MS [49]. Thanks to this approach, Rhee and coworkers described 31 [38] and 9 proteins [48] that had not been previously ascribed to the mitochondrion.
2.2. Mitochondrial Databases
Due to the critical roles that mitochondria play in both cell life and cell death, lots of efforts have been dedicated to obtaining a complete map of mitochondrial proteins from different tissues and organisms. To date, the application of MS methods has been the most successful strategy to define the mitochondrial proteome. Pioneering studies combining two-dimensional electrophoresis (2-DE) with MS methods date back to 1998 and 2001, with the analysis of mitochondrial extracts from human placenta [50] and a human neuroblastoma cell line [51], respectively. In 1999, MITOP database was released [52]. It was the first database for both nuclear- and mitochondrial-encoded proteins, comprising genetic and functional information on five species (including mouse and human), with annotated data derived from a variety of online resources and literature. Three years later, Taylor et al. published the first comprehensive human mitochondrial proteome, extending previous information about mitochondrial proteins identified by MS [53]. This study constituted the first step for the achievement of the MitoProteome Project (http://www.mitoproteome.org/) [54]. In its initial release, the Human Mitochondrial Protein Database contained 847 human mitochondrial protein sequences derived from public sequence databases and MS analyses [54]. This set was substantially revised in 2015, and, in the current version (as of February 2016), it covers information for 1705 genes and 3625 proteins. Additional reference databases have been released that list mitochondrial proteins identified via multiple approaches, including MS analysis, literature curations, and bioinformatics evaluations [55]. Some examples are MitoP2 [56] or MitoMiner [57]. The latter contains green fluorescence protein (GFP) localization images, is updated regularly, and provides useful lists of functionally related genes in up to 12 different organisms [58]. Currently, MitoMiner describes 1837 human proteins, providing experimental or bioinformatic prediction for mitochondrial localization. In 2008, Pagliarini et al. reported the largest compendium of mammalian mitochondrial proteins to date [28], the MitoCarta database (http://www.broadinstitute.org/pubs/MitoCarta), which encompassed 1098 genes coding for mouse mitochondrial proteins unveiled by experimental MS-based identification, bioinformatics analyses, and literature curation. This inventory was updated in 2016 after the use of improved transcript models, MS search algorithms, new database versions, and homology detection methods [27]. The current version (MitoCarta 2.0) consists of 1158 human and 1158 mouse genes encoding mitochondrial proteins whose localization and distribution across 14 tissues are provided [27].
Other interesting sources for mitochondrial annotation currently available are the UniProt database (http://www.uniprot.org) [59] and the Gene Ontology (GO) functional repository (http://www.geneontology.org) [60]. Recently, large-scale studies like the Human Proteome Map (HPM) initiative (http://humanproteomemap.org/) [61], the Human Protein Atlas (http://www.proteinatlas.org) [47], and COPaKB (the Cardiac Organellar Protein Atlas Knowledgebase) (http://www.heartproteome.org/) [62] have also provided highly specific new data on the subcellular localization of mitochondrial proteins.
3. Addressing Mitochondrial Dysfunction through High-Throughput Proteomics Techniques
It is agreed that the first attempt to address the mitochondrial proteome was carried out by Rabilloud and colleagues 20 years ago [50]. Since then, the interest on mitochondrial proteomics has gradually increased, as reflected by a simple query on the PubMed database (https://www.ncbi.nlm.nih.gov/pubmed) (Figure 1). The publication of landmark proteomics studies describing the mitochondria of human [53, 63], rat [64, 65], mouse [28, 66], and yeast [67, 68] species, as well as those related to different tissues [32] and cells [69], have critically contributed to the definition of the mitochondrial proteome to date.
Figure 1.
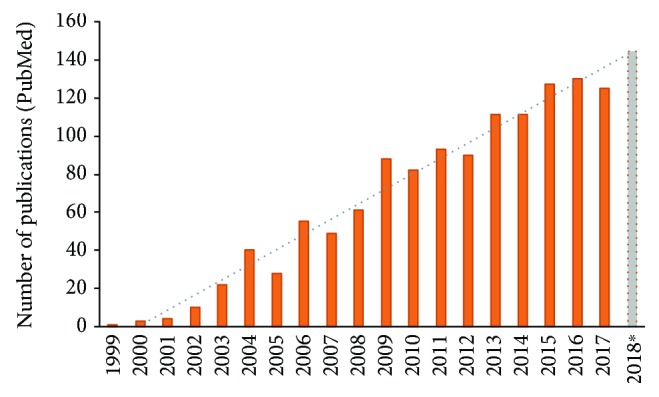
Timeline for “mitoproteomics” publications. Bars show the number of publications per year corresponding to the keywords “mitochondria” and “proteomics” in PubMed database (https://www.ncbi.nlm.nih.gov/pubmed). Note that the first proteomic study of human mitochondria [50] is not indexed. Results for 2018 are not included since they should be constantly updated during the writing process. According to the linear progression (dashed line), in 2018 we will probably reach a total of 144 publications related to mitochondrial proteomics (∗grey bar).
3.1. Isolation of Mitochondria
The starting point in the study of the mitochondrial proteome is mitochondrion isolation. Mitochondrion-enriched fractions have been used as starting material until differential centrifugation [70] became the method of choice. Three key steps can be distinguished in mitochondrial enrichment: (i) permeabilization and disruption of plasma membrane and release of mitochondria without compromising their integrity, (ii) differential centrifugation to remove cellular debris and larger organelles, and (iii) high-speed centrifugation to pellet the mitochondria [71]. The first step is usually performed through mechanical methods (e.g., mortar and pestle) under well-controlled buffer compositions. Some of the disadvantages of mechanical methods are excessive heat generation and stress on mitochondria, as well as increased intersample variability when executed manually [71]. In contrast, differential centrifugation should not compromise the quality or the yield of the preparation. Furthermore, additional gradient centrifugation protocols can be applied after differential centrifugation to obtain mitochondria with higher purity [72].
There are currently a few well-established protocols for isolating mitochondria [71, 73–75], although certain sample types might require specific procedures, as is the case of brain [76], adrenal glands [77], and adipose tissue [78]. Of note, the alternative use of magnetic devices for mitochondrial enrichment has gained popularity in recent years [78, 79].
The most common limitation of isolation strategies is the presence of carried-over nonmitochondrial proteins in the mitochondrial extracts, a phenomenon that is expected by virtue of the physical connection between the mitochondrion and other subcellular organelles [80]. However, high coverage of the total cellular proteome can be achieved thanks to recent improvements in protein identification methods based on liquid chromatography coupled with mass spectrometry (LC-MS) [24]. When combined with bioinformatics sorting of mitochondrial proteins, these data on protein identification reduce the necessity of purified mitochondria, which points out the combination of high-throughput MS analyses and database knowledge as a successful strategy, as previously described [78].
3.2. Sample Preparation for Mitochondrial Proteomics
As the identification of mitochondrial proteins is technically challenging due to their number and dynamic range, sample fractionation is desirable to reduce the complexity before MS analysis. While the first mitochondrial proteomics studies resorted to protein separation by 2-DE and peptide mass fingerprinting MS analysis to identify only a few dozen proteins [50, 51], later developments in LC-MS have facilitated the large-scale identification of mitochondrial proteins. In 2003, Taylor et al. identified 615 mitochondrial proteins from human heart by LC-MS (instead of 2-DE) analysis of 11 sucrose fractions [53], which represented a breakthrough in the identification of mitochondrial proteins. Later that year, gel filtration chromatography was used to separate mitochondrial proteins from brain, heart, kidney, and liver of C57BL6/J mice [32]. From each tissue, the authors obtained 15–20 fractions that were subjected to LC-MS analysis, which allowed the identification of 133 proteins not previously associated with this organelle. However, Mootha et al. did not overlook that their analysis was biased toward abundant molecules and that improvements in sample preparation, protein separation, and sensitiveness of MS were necessary. Many researchers have resorted to protein separation by sodium dodecyl sulfate (SDS) polyacrylamide gel electrophoresis (PAGE) followed by gel slicing and analysis of each band by high-performance LC-MS [28, 64, 66, 81]. Other less frequent fractionation methods have been applied prior to LC-MS, like the aforementioned sucrose gradients [53] and gel filtration chromatography [32].
3.3. Identification of Mitochondrial Proteins
Most proteomics experiments involve the enzymatic digestion of a complex protein extract prior to peptide separation, ionization, and analysis by LC-MS. For the last 15 years, LC-MS in data-dependent acquisition (DDA) mode has been the standard for proteomics identification [82, 83]. DDA is a milestone for high-throughput proteomics due to its efficiency at identifying peptides and proteins. In this method, peptide peaks (precursor ions) that overcome a threshold intensity in a survey MS scan are selected for fragmentation in the MS2 mode. This fragmentation generates an experimental fragment ion pattern characteristic of the peptide amino acid sequence that can be matched against theoretical patterns derived from protein databases for peptide identification [84]. In spite of the power of the DDA method, random peptide sampling for fragmentation entails a bias towards those with the strongest signal, which hampers the reproducible quantitation of especially low-abundance peptides.
Conversely, targeted acquisition methods, most notably selected reaction monitoring (SRM) [85], focus the mass spectrometer capabilities on the fragmentation of a limited number of peptides representing proteins of interest with very high sensitivity and quantitative accuracy. However, owing to the limited fraction of the proteome amenable to SRM analysis, this method is not suitable for most proteomics experiments, where detailed protein composition is required. So far, only a few studies have resorted to targeted methods for the study of human mitochondrial protein profiles [86, 87]. Other authors have applied targeted methods to the study of such posttranslational modifications (PTMs) as acetylation [88, 89] and phosphorylation [90, 91] in the mitochondrial proteome.
In recent years, data-independent acquisition (DIA), which combines the advantages of DDA and SRM, has raised increasing interest. The DIA method performs the fragmentation of all the peptides within a defined mass-to-charge (m/z) window; the window is then progressively shifted until the full m/z range is covered. This enables accurate peptide quantitation without being limited to profiling a predefined set of peptides of interest, but at the cost of losing the link between precursor and fragment ions thereof. An implementation of the DIA method, SWATH, relies on spectral libraries that collect m/z values, relative intensities, retention times, etc., to trace the so-obtained compound fragmentation maps for specific peptides. Since these libraries must be carefully built up based on additional DDA experiments [92], only a few studies have relied on SWATH to address mitochondrial disturbances in disease models [93–95]. A recent development based on DIA, DiS (data-independent scanning), resorts to narrower isolation windows to facilitate peptide identification based on the conventional database search methods developed for DDA [96]. DiS has been successfully used to generate a highly detailed structural map of OXPHOS mitochondrial supercomplexes in several models, including the characterization of protein species and PTMs thereof that regulate complex and supercomplex assembly [96]. DIA methods have enormous potential for mitoproteomics, where highly curated databases (e.g., MitoCarta 2.0) will help improve mapping and quantification of mitochondrial proteins.
3.4. Quantification of Mitochondrial Proteins
Depending on the separation strategy selected, protein quantification can be performed through gel-based or MS-based (gel-free) approaches. In gel-based approaches, the quantification of differentially abundant proteins is conducted by comparison of gel images, and MS methods are set aside for protein identification. Paramount among gel-based strategies is difference gel electrophoresis (DIGE), an improvement on 2-DE in which the proteins from two samples are labelled with different fluorescent dyes and separated in the same gel, allowing a more accurate relative quantification as compared to 2-DE [97].
In contrast, gel-free strategies rely on mass spectrometric data for both protein identification and protein quantitation. At this point, it should be noted that gel-free approaches have higher yield and well-established advantages in quantification over gel-based techniques. The quantification method in MS-based approaches includes label-free and stable isotope labelling (SIL) methods. Most often, label-free quantification relies on the counting of MS2 spectra from the peptides of a given protein [98]. The label-free approach has been used in numerous large-scale studies, as it provides a simple way to carry out relative comparisons across different samples with improved dynamic range of quantification [99]. Conversely, in SIL strategies, the relative quantification is based on the addition of chemically equivalent differential mass tags, which allow the comparison of up to ten samples in a single experiment. Despite that some limitations have been pointed out for SIL-based quantification, namely, limited linear dynamic range, complex sample preparation, and high cost of the reagents [100], this approach has great advantages in quantification accuracy and reproducibility. Some examples of SIL techniques successfully employed in comparative mitoproteomics are (i) metabolic labelling with stable isotope labelling by amino acids in cell culture (SILAC) [101, 102], (ii) enzymatic labelling with 18O during proteolytic digestion [103, 104], and (iii) chemical labelling employing isotope-coded affinity tags (ICAT) [105, 106], isobaric tags for relative and absolute quantitation (iTRAQ) [78, 107–109], or tandem mass tags (TMT) [110, 111].
3.5. Mitoproteomics in Human Disease
Early mitoproteomics studies relied mainly on gel-based techniques for protein separation, such as 2-DE, in combination with a variety of MS methods. After 2-DE separation, the protein spots of interest must be sliced off from the gel and digested for later off-line MS analysis. In these pioneering studies, the underrepresentation of extreme pI, low abundant, or very hydrophobic proteins, which has been pointed out as an important drawback inherent to 2-DE, has hindered the identification of a higher number of mitochondrial proteins [50, 65, 112].
During the first decade after mitoproteomics implementation, alternative separation methods based on SDS-PAGE coupled with LC-MS were developed that allowed significant advances in the investigation of the mitochondrial proteome (reviewed in [100]). In the last decade, there has been a dramatic increase in the number of publications related to mitoproteomics (e.g., until 2008, the total of publications according to PubMed indexing was 273, a number that was overcome in just the last two years) (Figure 1).
Most mitoproteomics studies have been performed on animal models, as these offer more possibilities for mitochondrial isolation and purification. Results on these models give insight into basic functions of proteins and reveal the mechanisms of adaptation under loss of function of the respective proteins (e.g., knockout models), altered nutritional status (e.g., obesity), pharmacological treatment, or physiopathological stressors, shedding light on mitochondrial responses. In the case of human specimens, there are significant limitations regarding sample availability, some of which are related to obvious difficulties in collecting the material, whereas others derive from medical and ethical issues. Despite this bottleneck, during the last years the increased robustness of MS approaches and the application of multiplexing methods (such as iTRAQ or TMT), which substantially reduce the amount of sample needed, have counteracted the limitations inherent to working with human samples, bringing mitoproteomics closer to translational research [78].
Furthermore, the combination of mitoproteomics with other high-resolution techniques has provided new insights into mitochondrial disease. For instance, Walheim and coworkers have recently described the combination of quantitative mitochondrial respirometry (through Seahorse technology) and proteomics (LC-MS-based total protein approach) to understand how protein changes relate to mitochondrial energy capacity during diet-induced obesity [113]. This approach, termed “respiromics,” has shown not only that CI concentration controls mitochondrial lipid oxidation in the liver, but also that obesity affects the functional capacity of this complex by regulating the nuclear- but not mitochondrial-encoded subunit concentrations.
In the following sections, the reader will find a selection of recent illustrative mitoproteomics studies in different pathological contexts, emphasizing the global perspective obtained on mitochondrial research thanks to these cutting-edge techniques.
3.5.1. Aging
Mitochondrial dysfunction is considered one of the hallmarks of aging [114], where several aspects of mitochondrial physiology are involved, such as mitochondrial biogenesis and turnover, calcium dynamics, energy sensing, RNOS production, and apoptosis [115]. In addition, aging is recognized as an independent risk factor for different pathologies such as cardiovascular disease (CVD), type 2 diabetes mellitus (T2DM), neurodegenerative disorders, or cancer [116, 117]. Several mitoproteomics studies have addressed the effect of aging in the context of these pathologies: see [78, 118] for metabolic disease, [119–121] for cardiac disturbances, and [122, 123] for neurodegenerative disorders. Downregulation of complex subunits and mitochondrial respiratory capacity is a common feature of normal aging mitochondria, although particular changes have been highlighted depending on the model, the age differences, and the particular tissue under study. For instance, in C57BL/6 mice, CI subunits have been described both to decrease [124] and to increase [123] with aging, and Stauch et al. have reported different patterns of metabolic and ETC modifications according to the age of mice in both synapses [125] and whole-brain mitochondria [126]. In the context of obesity, a pioneering study on human adipocyte mitochondria showed a general decrease in metabolic enzymes and ETC subunits in older compared to younger individuals, despite that all individuals were middle-aged (from 32 to 52 years) [78].
Of note, mitochondrial dysfunction has been addressed by proteomics in mouse models of premature aging (also known as Hutchinson-Gilford progeria syndrome (HGPS)) [127]. Although the genetic components of this disease are well-known (i.e., a dominant mutation in the LMNA gene leading to the expression of progerin), the mechanisms underlying cellular damage, senescence, and accelerated aging in HGPS are only partly understood. Resorting to SILAC and LC-MS analysis, the authors observed a pronounced downregulation of several components of the mitochondrial ATPase complex together with an upregulation of glycolytic enzymes. Interestingly, the analysis of different tissues revealed that mitochondrial dysfunction was time- and dose-dependent in the HPGS mouse models [127]. Altogether, these results underline the tight regulation undergone by mitochondria with aging, pointing out the importance of the surrounding context for this organelle.
3.5.2. Neurodegenerative Diseases
As previously introduced, mitoproteomics studies in the context of the nervous system have been closely related to aging research. This fact most probably responds to the observation of a decline in brain energy metabolism with aging [128]. This metabolic change is considered part of the physiological aging contributing to age-related neurodegeneration (e.g., Alzheimer's disease (AD) or Parkinson's disease (PD)), where different evidences support a key role of mitochondria in its progression [123, 129, 130]. Several proteomics approaches, including redox proteomics (see Redox Mitoproteomics Section in this review), have pointed out specific mitochondrial proteins as key regulators, providing new insights into their role in aberrant mitochondrial function and structure in AD and PD, among others (for an excellent review, see [131]). Interestingly, alterations to TCA enzymes and OXPHOS subunits are an early feature of mitochondrial dysfunction that can be observed prior to the deposition of significant amyloid plaques and neurofibrillary tangles in AD [132]. Causal links for mitochondrial abnormalities and oxidative damage in synaptic degeneration and amyloid-β deposition have been proposed [133, 134]. Moreover, substantial differences between synaptic and nonsynaptic mitochondria have been previously reported [135, 136]. In the mitoproteomics field, Graham et al. have recently evidenced distinct molecular fingerprints between mitochondrial subpopulations that are likely to influence the synaptic morphology in vivo, highlighting mitochondrial CI as an upstream regulator in neurodegenerative pathologies [137]. Thus, mitoproteomics research is regarded as a promising tool for the investigation of preclinical and clinical biomarkers of neurodegeneration and its potential treatments [131].
3.5.3. CVD
Heart tissue is rich in mitochondria and highly dependent on the catabolism of branched-chain amino acids and fatty acids within this organelle. Mitochondrial function has been closely related to cardiomyopathies [138] and ischemic heart disease [139]. Although the precise underlying mechanisms are not fully understood, it is clear that mitochondrial metabolism is heavily involved in both cardiac injury [140] and cardiac protection [141].
Heart ischemia causes tissue injury through an acute phase followed by secondary damage. Notably, ischemic preconditioning (brief episodes of ischemia preceding prolonged ischemia) provides robust protection against cell death resulting from ischemia and reperfusion (I/R) injury in the heart and is currently explored as a new target for cardiac treatment [142]. In fact, a combination of mitoproteomics with respiratory assays and electron microscopy (EM) described a “mitochondrial preconditioning” independent of cytosol that confers protection against I/R-induced respiratory failure and oxidative damage [143].
The cardiac mitochondrial proteome has been widely explored in models of I/R [144–147]. Most of these studies have also relied on redox approaches (see Redox Mitoproteomics Section in this review), since mitochondrial RNOS generation has been described as a double-edged sword contributing to myocardial infarction and stroke as well as ischemic preconditioning [148].
3.5.4. Metabolic Disease: Obesity and Diabetes
In essence, obesity consists in an excessive fat accumulation due to an imbalance between energy expenditure and uptake. Sedentary lifestyle and overnutrition have greatly contributed to the present epidemic of obesity, which is also aggravated by the aging of population [149]. Mitochondrial dysfunction has long been associated to the development of insulin resistance (IR) and T2DM, since changes in physical activity and overnutrition have a direct impact upon mitochondrial function [4]. During the last decades, most mitoproteomics studies addressing IR and T2DM pathologies were based on skeletal muscle samples, therefore overlooking the relevance of mitochondria in adipose tissue and the systemic implications of its impairment (for an excellent review of mitoproteomics in the context of metabolic disease, see [118]). Most of these studies relied on animal or cellular models, which imposed additional limitations to the extent of their conclusions [150]. The key role for white adipose tissue in regulating energy expenditure and IR was clearly recognized during the first decade of the 2000s [5, 151], but by that time the only comparative study between white and brown adipocyte mitochondria had been carried out in mice [102]. Therefore, the study of human adipocyte mitochondria was of outmost importance.
In 2015, Lindinger et al. published the first mitoproteomics study on this area [152]. Despite the limitations of their analysis (e.g., only 62 putative mitochondrial proteins were identified), the authors validated four proteins (citrate synthase, mitofilin, HADHA, and LETM1) that were inversely associated with body mass index (BMI) in white adipocyte mitochondria from 76 patients. Their results confirmed previous studies regarding whole adipose tissue biopsies [153, 154]. A few years later, Peral and colleagues published two complementary works that constitute the most comprehensive depiction of the human adipocyte mitochondrial proteome to date. In their first study, they described a global reduction of mitochondrial proteins (~150 proteins) in whole visceral adipose tissue biopsies from T2DM individuals by means of a high-throughput multiplexed proteomics approach based on iTRAQ technology [155]. In their second study, the authors deepened the proteome changes in adipocyte mitochondria with an improved redox approach [78] (see Redox Mitoproteomics Section for further details). Their results revealed impaired assembly of mitochondrial complexes together with defective protein import as a new potential cause of mitochondrial dysfunction in T2DM (Figure 2). In fact, the authors underlined the alteration of mitochondrial import pathways at different levels, like the TOM complex or the mitochondrial intermembrane space assembly (MIA) pathway [78], extending their previous results regarding the alteration of heat shock protein 70 in diabetic patients [154]. Additionally, the multiplexing capacity of their proteomics approach allowed the authors to address also the differences associated to aging in the same experiment, pointing out CIV as a common target for mitochondrial remodelling in both the aging and T2DM processes [78].
Figure 2.

High-throughput proteomics for the study of adipose tissue mitochondria. Gomez-Serrano et al. carried out a deep study of mitochondrial alterations in human fat. In a first study based on whole adipose tissue samples, they described proteome-wide downregulation of the identified mitochondrial proteins in both aging and T2DM processes thanks to the application of systems biology analyses (upper panel). Alterations in relation to aging were mostly circumscribed to the electron transport chain (ETC), whereas disturbances in diabetic patients included also other metabolic pathways such as tricarboxylic acid (TCA) cycle, branched-chain amino acid (BCAA), metabolism, or β-oxidation, most likely resembling a decrease in mitochondrial mass with severe consequences on mitochondrial fitness [155]. In a second study, the authors proceeded to the isolation of adipocyte mitochondria in order to further analyse this organelle (lower panel). Resorting to a redox approach, they described thiol oxidative modifications as well as protein abundance changes [78]. The authors found that thiol protein oxidation was inversely correlated to protein levels in adipocyte mitochondria and that this relationship was more dramatic in T2DM compared to the aging process. Additionally, OXPHOS mitochondrial- vs. nuclear-encoded protein modules were altered in T2DM (note that complex modules are represented in different colours to indicate upregulation (yellow) and downregulation (purple), respectively). Thus, their results underscored defects in respiratory capacity and protein import in aging and T2DM. Of note, CIV emerged as a common target of oxidative damage connecting aging and T2DM development. Graphical elements from this figure were taken and adapted from Servier Medical Art Powerpoint image bank (http://smart.servier.com/). Servier Medical Art by Servier is licensed under a Creative Commons Attribution 3.0 Unported License.
3.5.5. Liver Disease
Also, in close relation to aging or metabolic pathologies, we could find excellent works performed in the context of liver disease. Nonalcoholic fatty liver disease (NAFLD) is a chronic liver condition strongly associated with obesity, IR, and T2DM [156]. It can gradually progress to nonalcoholic steatohepatitis (NASH) in 12–40% of cases, cirrhosis (15% of NASH patients), and hepatic cancer (10%) [157], and oxidative damage on mitochondria has been tightly related to this fatal progression [158]. Thus, NAFLD and NASH attract a great interest among the scientific proteomics community (reviewed elsewhere [159]). These studies have revealed the importance of mitochondrial proteins in liver disease together with enzymes involved in methionine metabolism [160, 161], whose alteration may lead to an increase in both protein and lipid oxidative damage influencing the development of NAFLD and NASH. Furthermore, the beneficial action of metformin treatment in NAFLD has been also revealed in a mitoproteomic study of a mouse model [162].
Interestingly, ethanol-induced stress on liver has been also addressed through these approaches [163, 164], revealing an important modification of the mitochondrial proteome under these xenobiotic stimuli.
3.5.6. Cancer
Since the mitochondrion is the powerhouse of the cell, this organelle has long been related to carcinogenesis, cancer progression, and metastasis, although its specific role is still disputed [165]. It has been suggested that mitochondrial pathways such as RNOS signalling or Ca2+ homeostasis could play a critical role in the alteration of energy metabolism in cancer cells [166, 167]. Besides, the Warburg effect [168], a shift from mitochondrial respiration towards glycolysis in cancer cell metabolism, constitutes a canonical observation for almost all cancer types whose mechanism remains obscure. Of note, the wealth of data provided by cancer mitoproteomics has provided new insights into the Warburg effect (outstandingly reviewed in [169]), although proteomic studies in cancer have been mostly used for tumor subtyping [170].
The heterogeneity of tumor tissue and cell composition hampers the analysis of the corresponding mitochondria, hindering the interpretation of the results. Notwithstanding the above, altered structure and protein levels have been reported for mitochondria from cancer cells, and mitochondria have also been found affected by antioncogenic treatments (reviewed in [170]). Interestingly, a recent study of Casal's group has revealed the mechanisms leading to OXPHOS downregulation in colorectal cancer [171]. By using SILAC technology, subcellular fractionation, LC-MS analysis, and bioinformatics, the authors described spatial proteome alterations at the subcellular level that help elucidate the molecular mechanisms underlying colorectal cancer metastasis [171]. Apart from mitochondrial dysfunction, their data suggest that proteins belonging to the TCA cycle and OXPHOS might have secondary localizations such as the nucleus and the cytoskeleton, confirming previous results in MCF7 breast cancer cells [172].
3.5.7. Other Pathologies
Because of the great interest in mitochondrial proteomics shown in the last decade, many examples of models or diseases addressed through these approaches can be found. Thus, proteomics studies have also revealed the importance of mitochondrial proteins in hypertension [173], osteoarthritis [174], and male infertility [175, 176], among others.
4. A Further Step: Redox Mitoproteomics
Since mitochondria contain major producers of RNOS, including components of the respiratory chain and other redox enzymes [177], together with powerful antioxidative defence systems [178], measuring redox damage in this organelle is central to deciphering disease pathogenesis. Thus, significantly increased mitochondrial RNOS levels have been associated with CVD [179–181], cancer [182, 183], neurodegenerative disease [184, 185], and aging [55].
Among many other chemically feasible oxidative PTMs that may occur in proteins, redox reactions involving Cys thiol (-S-H) groups are frequent, mostly reversible, enzymatically controlled, and selective in vivo [186, 187] and are considered the main way whereby oxidants integrate into cellular signal transduction pathways [188]. Common reversible Cys oxoforms include disulphide bonds (-S-S-), S-glutathionylation (-S-SG-), S-nitrosation (-S-NO), and S-sulfenylation (-S-OH), whereas sulfinic (S-O2H) and sulfonic acid (S-O3H) constitute irreversible Cys oxoforms (except for eukaryotic 2-Cys peroxiredoxins, where sulfinic acid may be reduced by sulfiredoxin [189]). Despite that the mechanisms that regulate these processes in the mitochondrion remain obscure, the respiratory chain should play an essential role, as it harbours the main producers of mitochondrial RNOS, CI [190], CII [191], and CIII [192]. Still, the development of redox mitoproteomics, i.e., the large-scale characterization of the specific mitochondrial Cys residues that sense the oxidative milieu, has been obstructed by the lability and low abundance of Cys oxoforms, which for decades could only be tackled by site-directed mutagenesis [193]. Proteomics approaches to protein Cys thiol oxidation provide a variety of methods for the enrichment, detection, and quantitation of such modifications, which can be classified into gel-based and gel-free methods.
4.1. Gel-Based Redox Mitoproteomics
Reversible S-nitrosation (-S-NO) affects a number of mitochondrial proteins and is thought to be a major way in which nitric oxide (NO) metabolism modulates mitochondrial physiology [194]. Galkin et al. used fluorescein-N-ethylmaleimide (NEM) labelling and Blue Native (BN) separation to identify by MS the specific residue of mitochondrially encoded ND3 subunit that becomes accessible to chemical modification only in the deactive form of mitochondrial CI from bovine heart mitochondria [195]. This enzyme had been previously suggested to undergo modification by S-nitrosation under pathological conditions during hypoxia or when the NO : O2 ratio increases [196]. Under these conditions, RNOS and NO generation by the respiratory chain complexes and NO synthases, respectively, could cause the accumulation of the endogenous nitrosating agent peroxynitrite and result in permanent deactivation of the enzyme [196]. Shortly afterwards, S-nitrosation of the Cys switch of CI active/deactive transition was associated with marked cardioprotective effects in mice [197]. In 2007, Hurd et al. adapted DIGE [97] for the detection of proteins differentially oxidized by mitochondrial RNOS in mitochondria isolated from rat heart [198]. Based on the oxidation of a small subset of mitochondrial thiol proteins in the absence of bulk thiol changes, the authors suggested a possible link between mitochondrial RNOS production and the modulation of mitochondrial fatty acid and carbohydrate metabolism. Chouchani et al. further developed the DIGE method for the selective analysis of S-nitrosated proteins in mitochondria from rat heart and liver, where S-nitrosated proteins could be discriminated from proteins oxidized due to NO metabolism [199]. The authors identified 13 S-nitrosated mitochondrial proteins plus four that were oxidized, probably due to evanescent S-nitrosation relaxing to a reversible thiol modification. Interestingly, S-nitrosation was found to selectively and reversibly inhibit the activity of enzymes central to mitochondrial metabolism, namely, aconitase, mitochondrial aldehyde dehydrogenase, and α-ketoglutarate dehydrogenase. The contribution of NO-mediated S-nitrosation to the nerve injury-evoked pathology was shown in mouse spinal cord by Scheving et al., who used DIGE to reveal abundance changes of S-nitrosated proteins after sciatic nerve injury upon pretreatment with a NO synthase inhibitor [200]. A DIGE-based study also enabled quantitation of S-nitrosation in mitochondrial CI in normoxic and ischemic intact mouse hearts [201], where S-nitrosation had been associated with marked cardioprotective effects [197].
It has been suggested that changes to the sulfenome, which encompasses Cys sulfenic (-S-OH) groups derived from the initial oxidation of Cys thiol (-S-H) groups, play a central role in redox signalling. Thus, identifying the sulfenome under oxidative stress has been pointed out as a way to detect potential redox sensors. To date, the sulfenome has only been addressed in Arabidopsis thaliana cell suspensions exposed to H2O2 stress [202–204]. Using the dimedone derivative 4-(pent-4-yn-1-yl)cyclohexane-1,3-dione as a probe, Akter et al. identified by MS more than 200 sulfenylated proteins after the oxidative treatment of Arabidopsis thaliana cells, 14 of which located to the mitochondrion and were involved in the TCA cycle and the ETC [203].
4.2. MS-Based Redox Mitoproteomics
Early MS-based redox proteomics approaches resorted to the so-called biotin switch assay, whereby labile S-nitrosated Cys are converted to stable biotinylated Cys and the corresponding proteins purified by affinity chromatography and detected by immunoblotting or MS [205–208]. This assay allowed Hao et al. to pinpoint by LC-MS 68 S-nitrosation sites from 56 rat cerebellar proteins after treatment with S-nitrosoglutathione [205]. Some of these S-nitrosated Cys were involved in important mitochondrial processes like the formation of the permeability transition pore, the inactivation of sarcomeric creatine kinase, and mitochondrial relocation and neuroprotective activity of protein/nucleic acid deglycase DJ-1 [205]. The biotin switch assay has also been adapted for the analysis of additional Cys oxoforms (e.g., sulfenic acid [209–211]). Moreover, MS-based identification of S-glutathionylated proteins isolated from mouse liver helped to reveal a potential mechanism of resistance to acetaminophen-induced hepatotoxicity [212]. A combination of chemical enrichment methodologies with MS allowed Gould et al. to map 2596 sites of Cys modification including S-nitrosylation, S-glutathionylation, S-acylation, and S-sulfenylation, in wild-type mouse livers under normal physiological conditions [213]. Structural analysis localized these modifications in unique, evolutionary conserved protein segments outside commonly annotated functional regions.
Quantitative LC-MS approaches to Cys oxidation often start by stabilizing the redox status of the biological sample, usually by blocking free thiols with highly thiol-reactive reagents (e.g., iodoacetamide (IAM)). Thereafter, reversibly oxidized Cys residues are treated with a strong reducing agent like dithiothreitol (DTT) and the so-obtained free thiols alkylated with a second reagent to enable specific detection of Cys oxoforms. Differential Cys tagging with SIL reagents produces either chemically indiscernible light and heavy or isobaric peptides, which coelute in the chromatographic separation and exhibit identical ionization properties. Quantitation is then achieved by measuring the intensity of either neighbouring heavy and light precursor mass signals in the MS spectrum or reporter ions in the MS2 spectrum [214–218]. Weerapana et al. used click chemistry conjugation and affinity enrichment to characterize Cys functionality in mouse and human proteomes based on the levels of Cys oxoforms, some of which pertained to mitochondrial proteins [219]. In 2008, Leichert et al. proposed the OxICAT method [220], based on the 2-plex ICAT (isotope-coded affinity tag) technology, to address the thiol redoxome (Figure 3, left). A comparative study of redoxome analysis based on DIGE [198] and OxICAT identified 63 proteins in mouse heart that were prone to thiol oxidation upon H2O2 treatment, many of which were mitochondrial TCA cycle members [221]. Results showed superior sensitivity of the OxICAT approach, which has been progressively used for investigating the thiol redoxome in a variety of nonmitochondrial biological samples, including Saccharomyces cerevisiae [222], Schizosaccharomyces pombe [223, 224], Escherichia coli [225], Caenorhabditis elegans [226], Phaeodactylum tricornutum [227], Drosophila melanogaster [228], and Mus musculus heart [180]. Of note, Go et al. investigated the effect of acute cadmium exposure on the redox proteome and metabolome of mitochondria isolated from mouse liver [229]. OxICAT unveiled significantly increased oxidation relative to controls in 1247 Cys residues from 547 proteins that were mapped to mitochondrial pathways like OXPHOS and pyruvate metabolism. A recent work by Topf et al. used OxICAT to quantify oxidation in more than 4300 Cys residues pertaining to ~2200 proteins from S. cerevisiae upon exogenous and intracellular mitochondria-derived oxidative stress [230]. Site-specifically mapping these redox-active Cys to the cellular translation machinery helped the authors identify redox switches for global translation modulation by mitochondrially produced RNOS. SIL labelling of peptides derived from S-glutathionylated mouse liver proteins based on 6-plex TMT has evidenced that this Cys modification has a fundamental role in energy metabolism [231]. Chouchani et al. also resorted to TMT-based, 6-plex peptide tagging to label reversibly oxidized thiols in mouse brown adipose tissue, where mitochondrial RNOS induction was identified as a mechanism that supports UCP1-dependent thermogenesis and whole-body energy expenditure [232]. Recently, alterations to S-glutathionylated mitochondrial protein levels were linked to fatiguing contractions in mice [233]. For that, the authors resorted to selective reduction with a glutaredoxin enzyme cocktail and resin-assisted enrichment followed by 10-plex TMT labelling of total thiol and S-glutathionylated peptides to unveil about 2200 S-glutathionylation sites.
Figure 3.

The OxICAT and GELSILOX approaches to redox proteomics. Step I: a hypothetical protein, which exists in two different samples A and B in both the reduced (green) and oxidized (disulfide-linked, yellow) form, is incubated with an alkylating agent (12C-IAM for OxICAT, left, and, e.g., IAM, for GELSILOX, right) to block free sulfhydryl groups. Then, a reducing agent is used to cleave the disulfide bonds and the nascent thiol groups blocked using a second alkylating agent (13C-IAM for OxICAT and, e.g., NEM, for GELSILOX). Step II: the proteins are digested with trypsin and the so-obtained peptides either affinity-purified (OxICAT) or tagged with 18O labelling prior to mixing (GELSILOX). Step III: two independent quantitative MS assays reveal the extent of thiol modification in samples A and B (OxICAT), whereas one single MS assay yields not only the relative amount of the reduced and oxidized forms but also the relative quantitation of the corresponding protein between the two samples based on the MS signals from noncysteine peptides.
These quantitative, MS-based methods have enabled not only the identification of redox-regulated thiol proteins, most of them of mitochondrial origin, but also to pinpoint the precise reactive Cys and to quantify their oxidation level. Nevertheless, their dependence on trapping and enrichment of Cys-containing peptides prevents parallel assessment of alterations in protein abundance. Given that redox signalling or oxidative stress could trigger changes in protein abundance owing to protein degradation or protein translocation, their assessment is essential to providing an integrated view of the underlying processes.
4.3. The GELSILOX Approach
To circumvent the above-described deficiencies, in 2012, Martínez-Acedo et al. proposed GELSILOX (gel-based stable isotope labelling of oxidized Cys, Figure 3, right) [234]. By combining differential alkylation of reduced and oxidized Cys with SIL of both Cys-containing and non-Cys peptides, GELSILOX provides simultaneous quantification of reversible Cys oxoforms and protein abundance. For that, the method relies on a statistical framework newly developed for SIL-based quantitative proteomics, the weighted spectrum, peptide, and protein (WSPP) model [235]. GELSILOX was initially applied to characterize the main redox targets of H2O2 in human endothelial cells as well as to study alterations to the thiol redoxome in mitochondria isolated from rat heart cardiomyocytes after the I/R insult [234]. The method was later applied by Ruiz-Meana et al. to show that decreased subsarcolemmal mitochondrial respiration in heart rat after I/R is paralleled by increased Cys oxidation, with ischemic preconditioning preventing both phenomena [143]. In their investigation of the role of altered mitochondrial function in aging, Fernandez-Sanz et al. resorted to GELSILOX to demonstrate increased oxidation of different subunits of mitochondrial ATP synthase in hearts from old mice upon I/R [120]. Moreover, GELSILOX allowed Guaras et al. to hypothesize that specific Cys oxidation is linked to the differential stability and degradation of mitochondrial CI during reoxygenation. For that, the authors assessed the oxidation state of more than 1000 Cys residues from 784 mitochondrial proteins of mouse fibroblasts [96]. Of note, a combination of redox approaches allowed Gomez-Serrano et al. to point out defects in protein translocation to the mitochondrion as responsible for mitochondrial dysfunction in T2DM and aging. For that, the authors relied on a GELSILOX-based method to quantify 244 Cys oxidation sites within a set of 116 bona fide mitochondrial proteins that evidenced for the first time a link between increased thiol oxidation and decreased protein abundance under both conditions (Figure 2).
Despite tremendous advances of recent years, several challenges remain to be addressed by redox mitoproteomics. First, some of the aforementioned methodologies have been demonstrated appropriate for in vitro studies, and therefore their suitability to tackle the mitochondrial redoxosome in vivo is in question. For this reason, and also because redox proteomics studies based on human mitochondria are still scarce, the experimental evidence for physiological readouts of mitochondrial Cys thiol oxidation is yet limited. In this regard, care must be taken if reagents with widespread use like DTT and dimedone are incorporated to in vivo assays, as the former can stimulate RNOS production by mitochondrial enzymes [236] and the latter can react rapidly with sulfenyl amides [237]. Another hurdle to overcome when investigating the mitochondrial redoxome is the isolation of the organelle, which has traditionally been carried out by ultracentrifugation [238] and has the drawback of carrying over organelles with similar size profiles like lysosomes. In this regard, the effective enrichment of mitochondria is often a lengthy procedure, which is detrimental to the preservation of oxidative PTMs. Furthermore, as stated above, prevalent methods like OxICAT fail to provide parallel assessment of protein abundance changes in the same assay that unveils the alterations of Cys oxoforms, which hampers data interpretation.
Finally, notwithstanding the importance of oxidative modifications, many other PTMs take place in the mitochondrion that are responsive to the stressful and changing environmental conditions in which this organelle exerts its functions, e.g., phosphorylation, acetylation, ubiquitination, and succinylation. The functional significance of these PTMs and their interplay in the mitochondrion are still under investigation.
5. The Mitochondrial Interactome
Another step forward on defining the mitochondrial proteome was to understand the roles and consequences of protein-protein interactions (PPIs), as proteins do not usually function in isolation but interact with other molecules. Understanding PPIs is fundamental for the development of systems biology and novel therapeutics. A number of public databases provide comprehensive lists of physical PPIs in human as well as in other species [239]. The STRING (Search Tool for the Retrieval of Interacting Genes/Proteins) database (http://string-db.org/) contains both known and predicted protein-protein interactions [240]. IntAct (http://www.ebi.ac.uk/intact) and MINT databases provide molecular interaction data and curation tools for annotation [241]. Both IntAct and MINT are active contributors to the IMEx consortium (http://www.imexconsortium.org), which provides a nonredundant set of physical molecular interaction data from a broad taxonomic range of organisms. The Database of Interacting Proteins (DIP, http://dip.doe-mbi.ucla.edu/dip/) is an integration of experimentally determined PPIs on different organisms and contains an interesting visualization tool which provides a graphical illustration of these interactions [242]. In The Human Protein Reference Database (HPRD) the information of PPI is manually curated from the published literature and it also integrates the information of domain architecture, PTMs, interaction networks, and disease association for each protein in the human proteome [243].
The information about the physical interactions contained in these and other nonmentioned databases was derived from several methodologies applied to dissect PPIs. Methods based on immunoprecipitation or two-hybrid system in combination with Western blotting have been widely used to identify interaction partners of specific proteins. However, these approaches are limited to a reduced number of proteins.
The recognition that proteomes are highly interconnected networks, or interactomes, has changed biology's view of cause and effect, simultaneously highlighting the importance of systems biology and necessitating its use in deciphering complex biological phenomena for the development of novel therapeutics [64, 244–246]. This concept of connectivity modulating systems explains how functionally dissimilar systems, such as tissues, can have largely similar compositions [61, 247, 248].
In this regard, the combination of affinity purification and MS (AP-MS) has emerged as a powerful approach to delineate biological processes. The method consists in tagging a bait protein with an affinity tag such as His-tag, flag-tag, or tandem affinity purification (TAP) tag for expression in vivo. The group of partners interacting with the bait protein can be purified from the cell lysate by affinity or immunoaffinity techniques and analysed by MS for identification of the interacting components [249]. This technique has been applied to study interactions in a high-throughput manner [246, 250, 251], but also subsets of specific networks have been analysed, such as the interaction partners of COX4 for the study of CIV assembly [252], the characterization of ATPase protein components [253], and the interaction partners of the inner mitochondrial membrane protein LETM1 [254]. In the neurodegeneration research field this technique has unveiled parkin-associated proteins that are implicated in the mechanism of mitophagy of defective mitochondria [255], and a map of mitochondrial proteins involved in neurodegenerative diseases was obtained in 2017 [256]. Other recent works have described the bcl-2 interactome in human lung adenocarcinoma cells [257], as well as the interaction network of a set of 50 mitochondrial proteins that lacked significant functional annotation [258] to gain insight into their role in health and disease.
However, the largest set of AP-MS experiments to date is Bio-Plex [259, 260], consisting of more than 5891 individual experiments with the human ORFeome (the collection of open reading frames (ORFs)). The data can be explored through the web application Bio-Plex (http://bioplex.hms.harvard.edu/index.php) [261]. This platform allows the exploration of individual networks, highly interconnected network communities, subthreshold interactions, and specific experimental information like peptide spectral match counts for individual AP-MS experiments [260].
In spite of the power of AP-MS for studying interactions, it still suffers from some limitations. The global study of PPIs in a given system requires tagging every ORF of interest to provide a measurable readout or to enable purification and identification of the protein complex. In addition, protein tagging can be time-consuming and can disrupt interactions or alter the localization of the protein complex [262]. Moreover, these approaches generally trigger perturbations to the system, such as genetic modification of the native cells or disruption of the original cellular context, which increases the rate of both false-positive and false-negative results [263]. The selection of few cell types under artificial expression conditions also poses limitations to the interactome characterization [246, 259]. For these reasons, current networks remain of limited use in interpreting biological phenomena, as the observed interactions may not reflect the interactome landscapes within other systems or in disease. Alternative approaches are necessary for mapping native, endogenous interactomes under multiple conditions [264], as well as to survey the global organization of protein complexes within mitochondria. For the last years, protein complexes have gathered attention, as proteins arrange to form macromolecular assemblies that facilitate their function and regulation. Although macromolecular complexes are essential players in numerous biological processes, a limited number of methods suitable for defining the composition and functional dynamics of such assemblies have been described. Thus, the comprehensive mapping of these complex networks of stable and transient associations remains a key goal.
To study macromolecular complexes, it is necessary to apply mild, nondestructive techniques during sample processing to obtain the native complexes. Density gradient centrifugation, size exclusion chromatography (SEC), and native electrophoresis have been applied to define large multiprotein complexes, e.g., the respiratory chain supercomplexes in mitochondria [265, 266] and pyruvate dehydrogenase and other complexes from chloroplasts [267]. Density gradient centrifugation requires large protein amounts and suffers from low resolution. SEC provides a robust workflow for the separation of cytoplasmic complexes; however, it is not compatible with membrane complexes as they are extremely sensitive to the separation conditions [264, 268, 269], and size exclusion columns lead to sample dilution and cannot be used for complexes larger than 5 MDa [270]. Notwithstanding these drawbacks, interesting results have been achieved (e.g., the combination of quantitative proteomics and SEC to map 291 coeluting complexes, where the use of triplex labelling enabled monitoring of interactome rearrangements [262]). Due to these reasons, BN electrophoresis has been the preferred method for fractionating mitochondrial membrane protein complexes. Below is a description of the methods available for the study of the mitochondrial interactome.
5.1. BN-PAGE
BN-PAGE was developed by Schägger and von Jagow [271] and was firstly used for the isolation and characterization of the respiratory complexes from bovine mitochondria [272, 273]. BN electrophoresis is a special case of native electrophoresis for high-resolution separation of enzymatically active protein complexes from tissue homogenates and cell fractions, between 10 and 10,000 kDa [274]. Higher resolution can be attained by decreasing the pore size in the polyacrylamide gradient gel. The principle of BN-PAGE is simple, yet powerful: the protein complexes interact with Coomassie dye through hydrophobic interactions and, since the dye is negatively charged, become negatively charged as well. With this negative charge, the protein complexes migrate, at neutral pH, towards the anode, the same way that SDS forces proteins to run towards the anode in SDS-PAGE [249]. The difference between SDS-based SDS-PAGE and Coomassie-based BN-PAGE is that SDS is a strong denaturing agent, while Coomassie dye does not affect the integrity of the native protein complexes. Therefore, due to the external charge induced by Coomassie dye, BN-PAGE separates native protein complexes according to their molecular mass in the 100–1500 (or even higher) kDa mass range. BN-PAGE provides a vast amount of information: complex size, subunit composition and stoichiometry, assembly of protein complexes into supercomplexes, and relative abundance and stability of subcomplexes within protein complexes. In addition, the relative abundance of these protein complexes can also be determined [249].
BN electrophoresis in combination with MS analysis [275] emerged as one of the methods of choice for the analysis of the “complexome” (protein complex proteome) when in 2009 Wessels and coworkers reported the first “complexome profiling” [265]. The authors analysed a mitochondrial enriched fraction from a human HEK cell line and identified 48 of the 71 canonical subunits of the respiratory chain complexes (CI–CIV). Manually cut slices from BN gels were subjected to label-free, semiquantitative LC-MS, and protein abundance profiles were determined across the gel. Protein correlation profiling (PCP) was performed to identify potentially interacting proteins. Later that year, another work confirmed the potential of this technique studying how the composition and formation of respiratory chain complexes change under anaerobic conditions in mitochondrial protein extracts from S. cerevisiae [276].
5.2. PCP-MS-Based Approaches
PCP is based on the generation of individual protein elution or migration profiles, where comparison with other profiles by computational clustering and other approaches allows the identification of putative interacting proteins on the basis of similarities in their elution profiles [277, 278]. Since its discovery, lots of works performed on mitochondria have been published [264, 270, 279–282], and the methodology has been gradually improved: first, regarding the quality of native gel separation and gel slicing [281], and second, based on the evolution of mass spectrometers, which are faster and more sensitive [283].
Combined PCP-MS approaches have been used to characterize mitochondrial complexomes, leading to the discovery of TMEM126B as an assembly factor of respiratory chain CI [270], the analysis of human mitochondrial ribosomal complexes [282], the identification of novel assemblies of voltage-dependent anion channels/porins and TOM proteins [281], the analysis of changes occurring in cytoplasmic and mitochondrial interactomes in response to apoptosis initiation by caspase activity [264], the description of the assembly mechanism for CI [280], and the superassembly of CIII and CIV [279]. Although the aforementioned works performed the correlation profiles based on protein migration in a BN gel, elution profiles derived from other techniques, such as biochemical fractionation or SEC, are also feasible [262, 284, 285].
A potential advantage of the PCP approach is that hundreds to thousands of protein complexes can be simultaneously and rapidly analysed. A disadvantage of the PCP approach is that currently only soluble complexes with interactions that are not markedly weakened by the buffers used can be analysed [278].
5.3. Alternative Techniques: An Insight into Cross-Linking
Finally, another outstanding MS approach that can provide new evidences on the systems-wide organization of proteins in intact mitochondria is cross-linking mass spectrometry (XL-MS). In XL-MS, native protein contacts are captured using a cross-linker, which is typically a small organic molecule composed of a spacer arm and two functional groups that are reactive toward specific residue side chains. After proteolytic digestion of the cross-linked sample, residue-to-residue cross-links can be localized by MS-based peptide sequencing. A cross-link can only occur if the cross-linker can bridge the distance between the residues. Detected cross-links, therefore, reveal maximum residue-to-residue distance constraints within and in between proteins, providing insights into protein conformations, protein complex architectures, and protein interaction networks [286].
Although the identification of cross-linked peptides is not trivial even for purified protein complexes, which may be available in large quantity [263], recent advances in cross-linking methods have enabled high-throughput identification of protein interactions in complex mixtures and living cells [287], allowing, e.g., the determination of the molecular structure of the mammalian mitochondrial ribosome [288]. XL-MS provides data about the location of the interaction site in the biomolecule under study, an information that cannot be derived from the other methods for studying PPIs. Many works have contributed to improving (i) methods to enrich cross-links [289–291], (ii) cross-linking chemistries [263, 292–297], (iii) detection and identification strategies [292, 296, 298–305], and (iv) tools to perform structural analysis based on cross-linking sites [306–310]. The most widely used cross-linkers are active esters, such as disuccinimidyl suberate and bis(sulfosuccinimidyl) suberate, which induce nucleophilic attacks on primary amines and thus rely on the coupling of Lys residues [311].
Recent works have performed high-throughput investigation of protein structures and interactions at the mitochondrial proteome-wide level using XL-MS approaches [286, 287]. By combining cross-link identifications from 11 mitochondrial preparations from 4 different tissues, Schweppe et al. reported 1920 unique residue-to-residue connections and found intercomplex cross-linked peptides, supporting the existence of the respirosome in an excellent agreement with cryo-EM models [287]. Very recently, Liu et al. analysed intact mouse heart mitochondria and identified 3322 unique residue-to-residue contacts, the largest survey of mitochondrial protein interactions reported so far. In addition, the authors revealed the mitochondrial localization of four proteins not yet included in the MitoCarta database [286].
The major limitation of XL-MS from the structural point of view is that it cannot directly determine the relative stoichiometry of subunits in a complex, although this information can be derived from complementary quantitative MS methods. In addition, the field remains heterogeneous, with various experimental protocols and software applications, and this variety may seem intimidating to newcomers. Nevertheless, XL-MS studies of partially purified complexes will certainly provide new insights about protein interaction networks and their changes on perturbation, e.g., as a result of mutations connected to diseases. If this technique becomes optimized for complex samples, it will solidify the relevance of cross-linking-based methods not only in structural biology but also in systems biology, thus advancing the convergence of structural and cell biology [311].
6. Outlook
These are exciting times for exploring the human mitochondrial proteome. Since this highly active organelle contains major producers of RNOS, encompassing redox enzymes and OXPHOS proteins, measuring the redox damage with innovative redox proteomics-based approaches will be crucial to puzzle out human pathology in the near future. Mitochondria and energy metabolism are key components in life and death regulation. It is not surprising that dysfunctional mitochondria lie beneath the physiological process of aging and the unhealthy aging. Despite that research efforts have resulted in significant advances in the knowledge of the mitochondrial proteome, the identification of low-abundance proteins and the detection of very subtle expression changes under human physiological and pathological conditions is still challenging. In the coming years, improved MS instrumentation together with the combination of advanced labelling and fractionation methods will likely render a deeper view of the mitochondrial proteome that will shed light on the human diseases connected to mitochondria. So far, it is noteworthy that the application of systems biology analyses to big data sets accomplished through proteomics has greatly contributed to the depiction of the mitochondrial network. This achievement is particularly interesting, as it enables to assess whether and how protein abundance changes occur in a coordinated manner, a biological event that is well-known [312]. In the near future, interactomics will provide an upper level of information about complex regulation both inside and outside the mitochondrion, revealing the role of intricate protein networks in the whole cell metabolism. Thus, proteomics implementation for the study of the mitochondrial interactome constitutes a step forward that must be taken in the near future, especially in the context of human disease, where such kind of studies is scarce.
Furthermore, to understand the regulation of these proteins and networks, the high-throughput analysis of PTMs in health and disease is also of outmost importance. The unprecedented capacity of current MS-based approaches provides a vast amount of biologically relevant information, producing a molecular picture of the complex interplay between proteins and regulators. Of note, many PTMs have been described as molecular switches directly involved in protein activation and/or inhibition. Oxidative modifications constitute an exciting area of research due to the implications of RNOS in cell signalling and oxidative damage. Given that redox mitoproteomics has just started to be explored, the application of these approaches in different pathological models will probably elucidate novel mechanisms of redox regulation in human disease. Other PTM analyses have been performed in the mitochondrion, notably phosphorylation, acetylation, succinylation, and ubiquitination (reviewed in [313]); nevertheless, high-throughput studies have been hindered due to technical issues (e.g., usually only one, at most two, type of modification can be addressed in a single experiment). Therefore, the development of advanced algorithms for PTM discovery and their application in mitoproteomics will probably constitute a new milestone in mitochondrial research.
Acknowledgments
The proteomic data supporting this systematic review are from previously reported studies and datasets, which have been cited throughout the text. This work was supported by grant SAF2012-33014 from the Ministry of Economy, Industry and Competitiveness (MEIC), Spain, and partially financed with FEDER funds (to B.P.). CNIC is supported by the Ministerio de Ciencia, Innovación y Universidades, and the Pro CNIC Foundation and is a Severo Ochoa Center of Excellence (SEV-2015-0505).
Abbreviations
- 2-DE:
Two-dimensional gel electrophoresis
- AD:
Alzheimer's disease
- AP-MS:
Affinity purification and tandem mass spectrometry
- ATP:
Adenosine triphosphate
- BMI:
Body mass index
- BN:
Blue native
- CI:
Complex I, NADH dehydrogenase
- CII:
Complex II, succinate dehydrogenase
- CIII:
Complex III, cytochrome c-oxidoreductase
- CIV:
Complex IV, cytochrome c oxidase
- CV:
Complex V or ATP synthase
- CVD:
Cardiovascular disease
- DIA:
Data-independent acquisition
- DIGE:
Difference gel electrophoresis
- DiS:
Data-independent scanning
- DDA:
Data-dependent acquisition
- DTT:
Dithiothreitol
- EM:
Electron microscopy
- ETC:
Electron transport chain
- GELSILOX:
Gel-based stable isotope labelling of oxidized Cys
- GFP:
Green fluorescence protein
- GO:
Gene ontology
- HPLC:
High-performance liquid chromatography
- I/R:
Ischemia/reperfusion
- IAA:
Iodoacetic acid
- IAM:
Iodoacetamide
- ICAT:
Isotope-coded affinity tag
- IM:
Inner membrane
- IMS:
Intermembrane space
- IR:
Insulin resistance
- iTRAQ:
Isobaric tags for relative and absolute quantitation
- LC:
Liquid chromatography
- LC-MS:
Liquid chromatography coupled with mass spectrometry
- MIA:
Mitochondrial intermembrane space assembly
- MS2:
Fragmentation spectrum
- MS:
Mass spectrometry
- mtDNA:
Mitochondrial DNA
- m/z:
Mass-to-charge
- NEM:
N-Ethylmaleimide
- NO:
Nitric oxide
- OM:
Outer membrane
- ORF:
Open reading frame
- OXPHOS:
Oxidative phosphorylation
- PAGE:
Polyacrylamide gel electrophoresis
- PD:
Parkinson disease
- PCP:
Protein correlation profiling
- PPIs:
Protein-protein interactions
- PTM:
Posttranslational modification
- RNOS:
Reactive nitrogen and oxygen species
- SDS:
Sodium dodecyl sulfate
- SEC:
Size-exclusion chromatography
- SIL:
Stable isotope labelling
- SILAC:
Stable isotope labelling by amino acids in cell culture
- SRM:
Selected reaction monitoring
- T2DM:
Type 2 diabetes mellitus
- TAP:
Tandem affinity purification
- TCA:
Tricarboxylic acid
- TCEP:
Tris(2-carboxyethyl)phosphine
- TMT:
Tandem mass tags
- TOM:
Translocase of the outer membrane
- UCP1:
Uncoupling protein-1
- XL-MS:
Cross-linking mass spectrometry.
Contributor Information
María Gómez-Serrano, Email: maria.gomez@cnic.es.
Belén Peral, Email: bperal@iib.uam.es.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- 1.Alberts B., Johnson A., Lewis J., Walter P., Raff M., Roberts K. Molecular Biology of the Cell. 4th. Routledge: International Student Edition; 2002. [Google Scholar]
- 2.Cheng Z., Ristow M. Mitochondria and metabolic homeostasis. Antioxidants & Redox Signaling. 2013;19(3):240–242. doi: 10.1089/ars.2013.5255. [DOI] [PubMed] [Google Scholar]
- 3.Starkov A. A. The role of mitochondria in reactive oxygen species metabolism and signaling. Annals of the New York Academy of Sciences. 2008;1147(1):37–52. doi: 10.1196/annals.1427.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Gao A. W., Canto C., Houtkooper R. H. Mitochondrial response to nutrient availability and its role in metabolic disease. EMBO Molecular Medicine. 2014;6(5):580–589. doi: 10.1002/emmm.201303782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kusminski C. M., Scherer P. E. Mitochondrial dysfunction in white adipose tissue. Trends in Endocrinology and Metabolism. 2012;23(9):435–443. doi: 10.1016/j.tem.2012.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Duchen M. R. Mitochondria in health and disease: perspectives on a new mitochondrial biology. Molecular Aspects of Medicine. 2004;25(4):365–451. doi: 10.1016/j.mam.2004.03.001. [DOI] [PubMed] [Google Scholar]
- 7.Koliaki C., Roden M. Alterations of mitochondrial function and insulin sensitivity in human obesity and diabetes mellitus. Annual Review of Nutrition. 2016;36(1):337–367. doi: 10.1146/annurev-nutr-071715-050656. [DOI] [PubMed] [Google Scholar]
- 8.Galloway C. A., Yoon Y. Mitochondrial morphology in metabolic diseases. Antioxidants & Redox Signaling. 2013;19(4):415–430. doi: 10.1089/ars.2012.4779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Liesa M., Shirihai O. S. Mitochondrial dynamics in the regulation of nutrient utilization and energy expenditure. Cell Metabolism. 2013;17(4):491–506. doi: 10.1016/j.cmet.2013.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kalyanaraman B., Darley-Usmar V., Davies K. J. A., et al. Measuring reactive oxygen and nitrogen species with fluorescent probes: challenges and limitations. Free Radical Biology & Medicine. 2012;52(1):1–6. doi: 10.1016/j.freeradbiomed.2011.09.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kemp G. J., Brindle K. M. What do magnetic resonance–based measurements of Pi→ATP flux tell us about skeletal muscle metabolism? Diabetes. 2012;61(8):1927–1934. doi: 10.2337/db11-1725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Perry C. G. R., Kane D. A., Lanza I. R., Neufer P. D. Methods for assessing mitochondrial function in diabetes. Diabetes. 2013;62(4):1041–1053. doi: 10.2337/db12-1219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Szendroedi J., Phielix E., Roden M. The role of mitochondria in insulin resistance and type 2 diabetes mellitus. Nature Reviews Endocrinology. 2011;8(2):92–103. doi: 10.1038/nrendo.2011.138. [DOI] [PubMed] [Google Scholar]
- 14.Gregersen N., Hansen J., Palmfeldt J. Mitochondrial proteomics—a tool for the study of metabolic disorders. Journal of Inherited Metabolic Disease. 2012;35(4):715–726. doi: 10.1007/s10545-012-9480-3. [DOI] [PubMed] [Google Scholar]
- 15.Nikolov M., Schmidt C., Urlaub H. Quantitative mass spectrometry-based proteomics: an overview. Methods in Molecular Biology. 2012;893:85–100. doi: 10.1007/978-1-61779-885-6_7. [DOI] [PubMed] [Google Scholar]
- 16.Gustafsson C. M., Falkenberg M., Larsson N. G. Maintenance and expression of mammalian mitochondrial DNA. Annual Review of Biochemistry. 2016;85(1):133–160. doi: 10.1146/annurev-biochem-060815-014402. [DOI] [PubMed] [Google Scholar]
- 17.Chacinska A., Koehler C. M., Milenkovic D., Lithgow T., Pfanner N. Importing mitochondrial proteins: machineries and mechanisms. Cell. 2009;138(4):628–644. doi: 10.1016/j.cell.2009.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Stojanovski D., Bragoszewski P., Chacinska A. The MIA pathway: a tight bond between protein transport and oxidative folding in mitochondria. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. 2012;1823(7):1142–1150. doi: 10.1016/j.bbamcr.2012.04.014. [DOI] [PubMed] [Google Scholar]
- 19.Neupert W., Herrmann J. M. Translocation of proteins into mitochondria. Annual Review of Biochemistry. 2007;76(1):723–749. doi: 10.1146/annurev.biochem.76.052705.163409. [DOI] [PubMed] [Google Scholar]
- 20.MacPherson L., Tokatlidis K. Protein trafficking in the mitochondrial intermembrane space: mechanisms and links to human disease. Biochemical Journal. 2017;474(15):2533–2545. doi: 10.1042/BCJ20160627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Sokol A. M., Sztolsztener M. E., Wasilewski M., Heinz E., Chacinska A. Mitochondrial protein translocases for survival and wellbeing. FEBS Letters. 2014;588(15):2484–2495. doi: 10.1016/j.febslet.2014.05.028. [DOI] [PubMed] [Google Scholar]
- 22.Harbauer A. B., Zahedi R. P., Sickmann A., Pfanner N., Meisinger C. The protein import machinery of mitochondria-a regulatory hub in metabolism, stress, and disease. Cell Metabolism. 2014;19(3):357–372. doi: 10.1016/j.cmet.2014.01.010. [DOI] [PubMed] [Google Scholar]
- 23.Opalinska M., Meisinger C. Metabolic control via the mitochondrial protein import machinery. Current Opinion in Cell Biology. 2015;33:42–48. doi: 10.1016/j.ceb.2014.11.001. [DOI] [PubMed] [Google Scholar]
- 24.Palmfeldt J., Bross P. Proteomics of human mitochondria. Mitochondrion. 2017;33:2–14. doi: 10.1016/j.mito.2016.07.006. [DOI] [PubMed] [Google Scholar]
- 25.Andersson S. G. E., Zomorodipour A., Andersson J. O., et al. The genome sequence of Rickettsia prowazekii and the origin of mitochondria. Nature. 1998;396(6707):133–140. doi: 10.1038/24094. [DOI] [PubMed] [Google Scholar]
- 26.Huynen M. A., de Hollander M., Szklarczyk R. Mitochondrial proteome evolution and genetic disease. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease. 2009;1792(12):1122–1129. doi: 10.1016/j.bbadis.2009.03.005. [DOI] [PubMed] [Google Scholar]
- 27.Calvo S. E., Clauser K. R., Mootha V. K. MitoCarta2.0: an updated inventory of mammalian mitochondrial proteins. Nucleic Acids Research. 2016;44(D1):D1251–D1257. doi: 10.1093/nar/gkv1003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Pagliarini D. J., Calvo S. E., Chang B., et al. A mitochondrial protein compendium elucidates complex I disease biology. Cell. 2008;134(1):112–123. doi: 10.1016/j.cell.2008.06.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Gabaldon T., Rainey D., Huynen M. A. Tracing the evolution of a large protein complex in the eukaryotes, NADH:ubiquinone oxidoreductase (complex I) Journal of Molecular Biology. 2005;348(4):857–870. doi: 10.1016/j.jmb.2005.02.067. [DOI] [PubMed] [Google Scholar]
- 30.Foster L. J., de Hoog C. L., Zhang Y., et al. A mammalian organelle map by protein correlation profiling. Cell. 2006;125(1):187–199. doi: 10.1016/j.cell.2006.03.022. [DOI] [PubMed] [Google Scholar]
- 31.Kislinger T., Cox B., Kannan A., et al. Global survey of organ and organelle protein expression in mouse: combined proteomic and transcriptomic profiling. Cell. 2006;125(1):173–186. doi: 10.1016/j.cell.2006.01.044. [DOI] [PubMed] [Google Scholar]
- 32.Mootha V. K., Bunkenborg J., Olsen J. V., et al. Integrated analysis of protein composition, tissue diversity, and gene regulation in mouse mitochondria. Cell. 2003;115(5):629–640. doi: 10.1016/S0092-8674(03)00926-7. [DOI] [PubMed] [Google Scholar]
- 33.Calvo S. E., Mootha V. K. The mitochondrial proteome and human disease. Annual Review of Genomics and Human Genetics. 2010;11(1):25–44. doi: 10.1146/annurev-genom-082509-141720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Cannon B., Nedergaard J. Brown adipose tissue: function and physiological significance. Physiological Reviews. 2004;84(1):277–359. doi: 10.1152/physrev.00015.2003. [DOI] [PubMed] [Google Scholar]
- 35.Johnson D. T., Harris R. A., Blair P. V., Balaban R. S. Functional consequences of mitochondrial proteome heterogeneity. American Journal of Physiology-Cell Physiology. 2007;292(2):C698–C707. doi: 10.1152/ajpcell.00109.2006. [DOI] [PubMed] [Google Scholar]
- 36.Johnson D. T., Harris R. A., French S., et al. Tissue heterogeneity of the mammalian mitochondrial proteome. American Journal of Physiology-Cell Physiology. 2007;292(2):C689–C697. doi: 10.1152/ajpcell.00108.2006. [DOI] [PubMed] [Google Scholar]
- 37.Vo T. D., Greenberg H. J., Palsson B. O. Reconstruction and functional characterization of the human mitochondrial metabolic network based on proteomic and biochemical data. Journal of Biological Chemistry. 2004;279(38):39532–39540. doi: 10.1074/jbc.M403782200. [DOI] [PubMed] [Google Scholar]
- 38.Rhee H. W., Zou P., Udeshi N. D., et al. Proteomic mapping of mitochondria in living cells via spatially restricted enzymatic tagging. Science. 2013;339(6125):1328–1331. doi: 10.1126/science.1230593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Thiele I., Price N. D., Vo T. D., Palsson B. O. Candidate metabolic network states in human mitochondria. Impact of diabetes, ischemia, and diet. Journal of Biological Chemistry. 2005;280(12):11683–11695. doi: 10.1074/jbc.M409072200. [DOI] [PubMed] [Google Scholar]
- 40.Williams E. G., Wu Y., Jha P., et al. Systems proteomics of liver mitochondria function. Science. 2016;352(6291, article aad0189) doi: 10.1126/science.aad0189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Yogev O., Pines O. Dual targeting of mitochondrial proteins: mechanism, regulation and function. Biochimica et Biophysica Acta (BBA) - Biomembranes. 2011;1808(3):1012–1020. doi: 10.1016/j.bbamem.2010.07.004. [DOI] [PubMed] [Google Scholar]
- 42.Ben-Menachem R., Tal M., Shadur T., Pines O. A third of the yeast mitochondrial proteome is dual localized: a question of evolution. Proteomics. 2011;11(23):4468–4476. doi: 10.1002/pmic.201100199. [DOI] [PubMed] [Google Scholar]
- 43.Roberts D. J., Miyamoto S. Hexokinase II integrates energy metabolism and cellular protection: Akting on mitochondria and TORCing to autophagy. Cell Death & Differentiation. 2015;22(2):248–257. doi: 10.1038/cdd.2014.173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Liu X., Kim C. N., Yang J., Jemmerson R., Wang X. Induction of apoptotic program in cell-free extracts: requirement for dATP and cytochrome c. Cell. 1996;86(1):147–157. doi: 10.1016/S0092-8674(00)80085-9. [DOI] [PubMed] [Google Scholar]
- 45.Luo X., Budihardjo I., Zou H., Slaughter C., Wang X. Bid, a Bcl2 interacting protein, mediates cytochrome c release from mitochondria in response to activation of cell surface death receptors. Cell. 1998;94(4):481–490. doi: 10.1016/S0092-8674(00)81589-5. [DOI] [PubMed] [Google Scholar]
- 46.Sutendra G., Kinnaird A., Dromparis P., et al. A nuclear pyruvate dehydrogenase complex is important for the generation of acetyl-CoA and histone acetylation. Cell. 2014;158(1):84–97. doi: 10.1016/j.cell.2014.04.046. [DOI] [PubMed] [Google Scholar]
- 47.Uhlen M., Fagerberg L., Hallstrom B. M., et al. Proteomics. Tissue-based map of the human proteome. Science. 2015;347(6220, article 1260419) doi: 10.1126/science.1260419. [DOI] [PubMed] [Google Scholar]
- 48.Hung V., Zou P., Rhee H. W., et al. Proteomic mapping of the human mitochondrial intermembrane space in live cells via ratiometric APEX tagging. Molecular Cell. 2014;55(2):332–341. doi: 10.1016/j.molcel.2014.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Martell J. D., Deerinck T. J., Sancak Y., et al. Engineered ascorbate peroxidase as a genetically encoded reporter for electron microscopy. Nature Biotechnology. 2012;30(11):1143–1148. doi: 10.1038/nbt.2375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Rabilloud T., Kieffer S., Procaccio V., et al. Two-dimensional electrophoresis of human placental mitochondria and protein identification by mass spectrometry: toward a human mitochondrial proteome. Electrophoresis. 1998;19(6):1006–1014. doi: 10.1002/elps.1150190616. [DOI] [PubMed] [Google Scholar]
- 51.Scheffler N. K., Miller S. W., Carroll A. K., et al. Two-dimensional electrophoresis and mass spectrometric identification of mitochondrial proteins from an SH-SY5Y neuroblastoma cell line. Mitochondrion. 2001;1(2):161–179. doi: 10.1016/S1567-7249(01)00007-1. [DOI] [PubMed] [Google Scholar]
- 52.Scharfe C., Zaccaria P., Hoertnagel K., et al. MITOP: database for mitochondria-related proteins, genes and diseases. Nucleic Acids Research. 1999;27(1):153–155. doi: 10.1093/nar/27.1.153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Taylor S. W., Fahy E., Zhang B., et al. Characterization of the human heart mitochondrial proteome. Nature Biotechnology. 2003;21(3):281–286. doi: 10.1038/nbt793. [DOI] [PubMed] [Google Scholar]
- 54.Cotter D., Guda P., Fahy E., Subramaniam S. MitoProteome: mitochondrial protein sequence database and annotation system. Nucleic Acids Research. 2004;32(90001):463D–4467. doi: 10.1093/nar/gkh048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Cui H., Kong Y., Zhang H. Oxidative stress, mitochondrial dysfunction, and aging. Journal of Signal Transduction. 2012;2012:13. doi: 10.1155/2012/646354.646354 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Elstner M., Andreoli C., Ahting U., et al. MitoP2: an integrative tool for the analysis of the mitochondrial proteome. Molecular Biotechnology. 2008;40(3):306–315. doi: 10.1007/s12033-008-9100-5. [DOI] [PubMed] [Google Scholar]
- 57.Smith A. C., Robinson A. J. MitoMiner, an integrated database for the storage and analysis of mitochondrial proteomics data. Molecular & Cellular Proteomics. 2009;8(6):1324–1337. doi: 10.1074/mcp.M800373-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Smith A. C., Robinson A. J. MitoMiner v3.1, an update on the mitochondrial proteomics database. Nucleic Acids Research. 2016;44(D1):D1258–D1261. doi: 10.1093/nar/gkv1001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.The UniProt Consortium. UniProt: a hub for protein information. Nucleic Acids Research. 2015;43:D204–D212. doi: 10.1093/nar/gku989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Ashburner M., Ball C. A., Blake J. A., et al. Gene ontology: tool for the unification of biology. Nature Genetics. 2000;25(1):25–29. doi: 10.1038/75556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kim M.-S., Pinto S. M., Getnet D., et al. A draft map of the human proteome. Nature. 2014;509(7502):575–581. doi: 10.1038/nature13302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Zong N. C., Li H., Li H., et al. Integration of cardiac proteome biology and medicine by a specialized knowledgebase. Circulation Research. 2013;113(9):1043–1053. doi: 10.1161/CIRCRESAHA.113.301151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Lefort N., Yi Z., Bowen B., et al. Proteome profile of functional mitochondria from human skeletal muscle using one-dimensional gel electrophoresis and HPLC-ESI-MS/MS. Journal of Proteomics. 2009;72(6):1046–1060. doi: 10.1016/j.jprot.2009.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Forner F., Foster L. J., Campanaro S., Valle G., Mann M. Quantitative proteomic comparison of rat mitochondria from muscle, heart, and liver. Molecular & Cellular Proteomics. 2006;5(4):608–619. doi: 10.1074/mcp.M500298-MCP200. [DOI] [PubMed] [Google Scholar]
- 65.Fountoulakis M., Berndt P., Langen H., Suter L. The rat liver mitochondrial proteins. Electrophoresis. 2002;23(2):311–328. doi: 10.1002/1522-2683(200202)23:2<311::AID-ELPS311>3.0.CO;2-0. [DOI] [PubMed] [Google Scholar]
- 66.Zhang J., Li X., Mueller M., et al. Systematic characterization of the murine mitochondrial proteome using functionally validated cardiac mitochondria. Proteomics. 2008;8(8):1564–1575. doi: 10.1002/pmic.200700851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Prokisch H., Scharfe C., Camp D. G., et al. Integrative analysis of the mitochondrial proteome in yeast. PLoS Biology. 2004;2(6, article e160) doi: 10.1371/journal.pbio.0020160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Sickmann A., Reinders J., Wagner Y., et al. The proteome of Saccharomyces cerevisiae mitochondria. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(23):13207–13212. doi: 10.1073/pnas.2135385100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Geiger T., Wehner A., Schaab C., Cox J., Mann M. Comparative proteomic analysis of eleven common cell lines reveals ubiquitous but varying expression of most proteins. Molecular & Cellular Proteomics. 2012;11(3, article M111.014050) doi: 10.1074/mcp.M111.014050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Hogeboom G. H., Schneider W. C., Pallade G. E. Cytochemical studies of mammalian tissues; isolation of intact mitochondria from rat liver; some biochemical properties of mitochondria and submicroscopic particulate material. Journal of Biological Chemistry. 1948;172(2):619–635. [PubMed] [Google Scholar]
- 71.Wettmarshausen J., Perocchi F. Isolation of functional mitochondria from cultured cells and mouse tissues. Methods in Molecular Biology. 2017;1567:15–32. doi: 10.1007/978-1-4939-6824-4_2. [DOI] [PubMed] [Google Scholar]
- 72.Graham J. M. Purification of a crude mitochondrial fraction by density-gradient centrifugation. Current Protocols in Cell Biology. 2001;4:3.4.1–3.4.22. doi: 10.1002/0471143030.cb0304s04. [DOI] [PubMed] [Google Scholar]
- 73.Frezza C., Cipolat S., Scorrano L. Organelle isolation: functional mitochondria from mouse liver, muscle and cultured fibroblasts. Nature Protocols. 2007;2(2):287–295. doi: 10.1038/nprot.2006.478. [DOI] [PubMed] [Google Scholar]
- 74.Pallotti F., Lenaz G. Isolation and subfractionation of mitochondria from animal cells and tissue culture lines. Methods in Cell Biology. 2007;80:3–44. doi: 10.1016/S0091-679X(06)80001-4. [DOI] [PubMed] [Google Scholar]
- 75.Schulz S., Lichtmannegger J., Schmitt S., et al. A protocol for the parallel isolation of intact mitochondria from rat liver, kidney, heart, and brain. Methods in Molecular Biology. 2015;1295:75–86. doi: 10.1007/978-1-4939-2550-6_7. [DOI] [PubMed] [Google Scholar]
- 76.Chinopoulos C., Zhang S. F., Thomas B., Ten V., Starkov A. A. Isolation and functional assessment of mitochondria from small amounts of mouse brain tissue. Methods in Molecular Biology. 2011;793:311–324. doi: 10.1007/978-1-61779-328-8_20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Solinas P., Fujioka H., Tandler B., Hoppel C. L. Isolation of rat adrenocortical mitochondria. Biochemical and Biophysical Research Communications. 2012;427(1):96–99. doi: 10.1016/j.bbrc.2012.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Gomez-Serrano M., Camafeita E., Lopez J. A., et al. Differential proteomic and oxidative profiles unveil dysfunctional protein import to adipocyte mitochondria in obesity-associated aging and diabetes. Redox Biology. 2017;11:415–428. doi: 10.1016/j.redox.2016.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Tang B., Zhao L., Liang R., Zhang Y., Wang L. Magnetic nanoparticles: an improved method for mitochondrial isolation. Molecular Medicine Reports. 2012;5(5):1271–1276. doi: 10.3892/mmr.2012.806. [DOI] [PubMed] [Google Scholar]
- 80.Giorgi C., De Stefani D., Bononi A., Rizzuto R., Pinton P. Structural and functional link between the mitochondrial network and the endoplasmic reticulum. The International Journal of Biochemistry & Cell Biology. 2009;41(10):1817–1827. doi: 10.1016/j.biocel.2009.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Adachi J., Kumar C., Zhang Y., Mann M. In-depth analysis of the adipocyte proteome by mass spectrometry and bioinformatics. Molecular & Cellular Proteomics. 2007;6(7):1257–1273. doi: 10.1074/mcp.M600476-MCP200. [DOI] [PubMed] [Google Scholar]
- 82.Hu A., Noble W. S., Wolf-Yadlin A. Technical advances in proteomics: new developments in data-independent acquisition. F1000Res. 2016;5 doi: 10.12688/f1000research.7042.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Walther T. C., Mann M. Mass spectrometry–based proteomics in cell biology. Journal of Cell Biology. 2010;190(4):491–500. doi: 10.1083/jcb.201004052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Washburn M. P., Wolters D., Yates J. R. Large-scale analysis of the yeast proteome by multidimensional protein identification technology. Nature Biotechnology. 2001;19(3):242–247. doi: 10.1038/85686. [DOI] [PubMed] [Google Scholar]
- 85.Lange V., Picotti P., Domon B., Aebersold R. Selected reaction monitoring for quantitative proteomics: a tutorial. Molecular Systems Biology. 2008;4:p. 222. doi: 10.1038/msb.2008.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Fernández-Guerra P., Birkler R. I. D., Merinero B., et al. Selected reaction monitoring as an effective method for reliable quantification of disease-associated proteins in maple syrup urine disease. Molecular Genetics & Genomic Medicine. 2014;2(5):383–392. doi: 10.1002/mgg3.88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Rosello-Lleti E., Tarazon E., Barderas M. G., et al. Heart mitochondrial proteome study elucidates changes in cardiac energy metabolism and antioxidant PRDX3 in human dilated cardiomyopathy. PLoS One. 2014;9(11, article e112971) doi: 10.1371/journal.pone.0112971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Hosp F., Lassowskat I., Santoro V., et al. Lysine acetylation in mitochondria: from inventory to function. Mitochondrion. 2017;33:58–71. doi: 10.1016/j.mito.2016.07.012. [DOI] [PubMed] [Google Scholar]
- 89.Rardin M. J., Newman J. C., Held J. M., et al. Label-free quantitative proteomics of the lysine acetylome in mitochondria identifies substrates of SIRT3 in metabolic pathways. Proceedings of the National Academy of Sciences of the United States of America. 2013;110(16):6601–6606. doi: 10.1073/pnas.1302961110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Kruse R., Hojlund K. Mitochondrial phosphoproteomics of mammalian tissues. Mitochondrion. 2017;33:45–57. doi: 10.1016/j.mito.2016.08.004. [DOI] [PubMed] [Google Scholar]
- 91.Lam M. P. Y., Scruggs S. B., Kim T. Y., et al. An MRM-based workflow for quantifying cardiac mitochondrial protein phosphorylation in murine and human tissue. Journal of Proteomics. 2012;75(15):4602–4609. doi: 10.1016/j.jprot.2012.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Schubert O. T., Gillet L. C., Collins B. C., et al. Building high-quality assay libraries for targeted analysis of SWATH MS data. Nature Protocols. 2015;10(3):426–441. doi: 10.1038/nprot.2015.015. [DOI] [PubMed] [Google Scholar]
- 93.Grois L., Hupf J., Reinders J., et al. Combined inhibition of the renin-angiotensin system and neprilysin positively influences complex mitochondrial adaptations in progressive experimental heart failure. PLoS One. 2017;12(1, article e0169743) doi: 10.1371/journal.pone.0169743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Villeneuve L. M., Purnell P. R., Boska M. D., Fox H. S. Early expression of Parkinson’s disease-related mitochondrial abnormalities in PINK1 knockout rats. Molecular Neurobiology. 2016;53(1):171–186. doi: 10.1007/s12035-014-8927-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Villeneuve L. M., Stauch K. L., Fox H. S. Proteomic analysis of the mitochondria from embryonic and postnatal rat brains reveals response to developmental changes in energy demands. Journal of Proteomics. 2014;109:228–239. doi: 10.1016/j.jprot.2014.07.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Guaras A., Perales-Clemente E., Calvo E., et al. The CoQH2/CoQ ratio serves as a sensor of respiratory chain efficiency. Cell Reports. 2016;15(1):197–209. doi: 10.1016/j.celrep.2016.03.009. [DOI] [PubMed] [Google Scholar]
- 97.Unlu M., Morgan M. E., Minden J. S. Difference gel electrophoresis: a single gel method for detecting changes in protein extracts. Electrophoresis. 1997;18(11):2071–2077. doi: 10.1002/elps.1150181133. [DOI] [PubMed] [Google Scholar]
- 98.Bantscheff M., Lemeer S., Savitski M. M., Kuster B. Quantitative mass spectrometry in proteomics: critical review update from 2007 to the present. Analytical and Bioanalytical Chemistry. 2012;404(4):939–965. doi: 10.1007/s00216-012-6203-4. [DOI] [PubMed] [Google Scholar]
- 99.Calderon-Celis F., Encinar J. R., Sanz-Medel A. Standardization approaches in absolute quantitative proteomics with mass spectrometry. Mass Spectrometry Reviews. 2018;37(6):715–737. doi: 10.1002/mas.21542. [DOI] [PubMed] [Google Scholar]
- 100.Chen X., Li J., Hou J., Xie Z., Yang F. Mammalian mitochondrial proteomics: insights into mitochondrial functions and mitochondria-related diseases. Expert Review of Proteomics. 2010;7(3):333–345. doi: 10.1586/epr.10.22. [DOI] [PubMed] [Google Scholar]
- 101.Bogenhagen D. F., Ostermeyer-Fay A. G., Haley J. D., Garcia-Diaz M. Kinetics and mechanism of mammalian mitochondrial ribosome assembly. Cell Reports. 2018;22(7):1935–1944. doi: 10.1016/j.celrep.2018.01.066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Forner F., Kumar C., Luber C. A., Fromme T., Klingenspor M., Mann M. Proteome differences between brown and white fat mitochondria reveal specialized metabolic functions. Cell Metabolism. 2009;10(4):324–335. doi: 10.1016/j.cmet.2009.08.014. [DOI] [PubMed] [Google Scholar]
- 103.Bienengraeber M., Pellitteri-Hahn M., Hirata N., Baye T. M., Bosnjak Z. J., Olivier M. Quantitative characterization of changes in the cardiac mitochondrial proteome during anesthetic preconditioning and ischemia. Physiological Genomics. 2013;45(5):163–170. doi: 10.1152/physiolgenomics.00117.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Smith J. R., Matus I. R., Beard D. A., Greene A. S. Differential expression of cardiac mitochondrial proteins. Proteomics. 2008;8(3):446–462. doi: 10.1002/pmic.200701009. [DOI] [PubMed] [Google Scholar]
- 105.Liu Y., He J., Ji S., et al. Comparative studies of early liver dysfunction in senescence-accelerated mouse using mitochondrial proteomics approaches. Molecular & Cellular Proteomics. 2008;7(9):1737–1747. doi: 10.1074/mcp.M800109-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Lovell M. A., Xiong S., Markesbery W. R., Lynn B. C. Quantitative proteomic analysis of mitochondria from primary neuron cultures treated with amyloid beta peptide. Neurochemical Research. 2005;30(1):113–122. doi: 10.1007/s11064-004-9692-5. [DOI] [PubMed] [Google Scholar]
- 107.Newton B. W., Cologna S. M., Moya C., Russell D. H., Russell W. K., Jayaraman A. Proteomic analysis of 3T3-L1 adipocyte mitochondria during differentiation and enlargement. Journal of Proteome Research. 2011;10(10):4692–4702. doi: 10.1021/pr200491h. [DOI] [PubMed] [Google Scholar]
- 108.Shi Z., Long W., Zhao C., Guo X., Shen R., Ding H. Comparative proteomics analysis suggests that placental mitochondria are involved in the development of pre-eclampsia. PLoS One. 2013;8(5, article e64351) doi: 10.1371/journal.pone.0064351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Xu Z., Jin X., Cai W., et al. Proteomics analysis reveals abnormal electron transport and excessive oxidative stress cause mitochondrial dysfunction in placental tissues of early-onset preeclampsia. Proteomics Clinical applications. 2018;12(5, article e1700165) doi: 10.1002/prca.201700165. [DOI] [PubMed] [Google Scholar]
- 110.Cheng Y., Hou T., Ping J., Chen G., Chen J. Quantitative succinylome analysis in the liver of non-alcoholic fatty liver disease rat model. Proteome Science. 2016;14(1):p. 3. doi: 10.1186/s12953-016-0092-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Gao W., Xu J., Wang F., et al. Mitochondrial proteomics approach reveals voltage-dependent anion channel 1 (VDAC1) as a potential biomarker of gastric cancer. Cellular Physiology and Biochemistry. 2015;37(6):2339–2354. doi: 10.1159/000438588. [DOI] [PubMed] [Google Scholar]
- 112.Fountoulakis M., Schlaeger E. J. The mitochondrial proteins of the neuroblastoma cell line IMR-32. Electrophoresis. 2003;24(12):260–275. doi: 10.1002/elps.200390022. [DOI] [PubMed] [Google Scholar]
- 113.Walheim E., Wisniewski J. R., Jastroch M. Respiromics - an integrative analysis linking mitochondrial bioenergetics to molecular signatures. Molecular Metabolism. 2018;9:4–14. doi: 10.1016/j.molmet.2018.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Lopez-Otin C., Blasco M. A., Partridge L., Serrano M., Kroemer G. The hallmarks of aging. Cell. 2013;153(6):1194–1217. doi: 10.1016/j.cell.2013.05.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Gonzalez-Freire M., de Cabo R., Bernier M., et al. Reconsidering the role of mitochondria in aging. The Journals of Gerontology Series A, Biological Sciences and Medical Sciences. 2015;70(11):1334–1342. doi: 10.1093/gerona/glv070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Bao Q., Pan J., Qi H., et al. Aging and age-related diseases – from endocrine therapy to target therapy. Molecular and Cellular Endocrinology. 2014;394(1-2):115–118. doi: 10.1016/j.mce.2014.07.005. [DOI] [PubMed] [Google Scholar]
- 117.Niccoli T., Partridge L. Ageing as a risk factor for disease. Current Biology. 2012;22(17):R741–R752. doi: 10.1016/j.cub.2012.07.024. [DOI] [PubMed] [Google Scholar]
- 118.Peinado J. R., Diaz-Ruiz A., Fruhbeck G., Malagon M. M. Mitochondria in metabolic disease: getting clues from proteomic studies. Proteomics. 2014;14(4-5):452–466. doi: 10.1002/pmic.201300376. [DOI] [PubMed] [Google Scholar]
- 119.Chakravarti B., Oseguera M., Dalal N., et al. Proteomic profiling of aging in the mouse heart: altered expression of mitochondrial proteins. Archives of Biochemistry and Biophysics. 2008;474(1):22–31. doi: 10.1016/j.abb.2008.02.001. [DOI] [PubMed] [Google Scholar]
- 120.Fernandez-Sanz C., Ruiz-Meana M., Castellano J., et al. Altered FoF1 ATP synthase and susceptibility to mitochondrial permeability transition pore during ischaemia and reperfusion in aging cardiomyocytes. Thrombosis and Haemostasis. 2015;113(03):441–451. doi: 10.1160/TH14-10-0901. [DOI] [PubMed] [Google Scholar]
- 121.Fernandez-Sanz C., Ruiz-Meana M., Miro-Casas E., et al. Defective sarcoplasmic reticulum-mitochondria calcium exchange in aged mouse myocardium. Cell Death & Disease. 2014;5(12, article e1573) doi: 10.1038/cddis.2014.526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Ingram T., Chakrabarti L. Proteomic profiling of mitochondria: what does it tell us about the ageing brain? Aging. 2016;8(12):3161–3179. doi: 10.18632/aging.101131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Pollard A., Shephard F., Freed J., Liddell S., Chakrabarti L. Mitochondrial proteomic profiling reveals increased carbonic anhydrase II in aging and neurodegeneration. Aging. 2016;8(10):2425–2436. doi: 10.18632/aging.101064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Mao L., Zabel C., Wacker M. A., et al. Estimation of the mtDNA mutation rate in aging mice by proteome analysis and mathematical modeling. Experimental Gerontology. 2006;41(1):11–24. doi: 10.1016/j.exger.2005.09.012. [DOI] [PubMed] [Google Scholar]
- 125.Stauch K. L., Purnell P. R., Fox H. S. Aging synaptic mitochondria exhibit dynamic proteomic changes while maintaining bioenergetic function. Aging. 2014;6(4):320–334. doi: 10.18632/aging.100657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Stauch K. L., Purnell P. R., Villeneuve L. M., Fox H. S. Proteomic analysis and functional characterization of mouse brain mitochondria during aging reveal alterations in energy metabolism. Proteomics. 2015;15(9):1574–1586. doi: 10.1002/pmic.201400277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Rivera-Torres J., Acin-Perez R., Cabezas-Sanchez P., et al. Identification of mitochondrial dysfunction in Hutchinson–Gilford progeria syndrome through use of stable isotope labeling with amino acids in cell culture. Journal of Proteomics. 2013;91:466–477. doi: 10.1016/j.jprot.2013.08.008. [DOI] [PubMed] [Google Scholar]
- 128.Boveris A., Navarro A. Brain mitochondrial dysfunction in aging. IUBMB Life. 2008;60(5):308–314. doi: 10.1002/iub.46. [DOI] [PubMed] [Google Scholar]
- 129.Moon H. E., Paek S. H. Mitochondrial dysfunction in Parkinson’s disease. Experimental Neurobiology. 2015;24(2):103–116. doi: 10.5607/en.2015.24.2.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Onyango I. G., Dennis J., Khan S. M. Mitochondrial dysfunction in Alzheimer’s disease and the rationale for bioenergetics based therapies. Aging and Disease. 2016;7(2):201–214. doi: 10.14336/AD.2015.1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Butterfield D. A., Palmieri E. M., Castegna A. Clinical implications from proteomic studies in neurodegenerative diseases: lessons from mitochondrial proteins. Expert Review of Proteomics. 2016;13(3):259–274. doi: 10.1586/14789450.2016.1149470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Chou J. L., Shenoy D. V., Thomas N., et al. Early dysregulation of the mitochondrial proteome in a mouse model of Alzheimer’s disease. Journal of Proteomics. 2011;74(4):466–479. doi: 10.1016/j.jprot.2010.12.012. [DOI] [PubMed] [Google Scholar]
- 133.Nunomura A., Perry G., Aliev G., et al. Oxidative damage is the earliest event in Alzheimer disease. Journal of Neuropathology and Experimental Neurology. 2001;60(8):759–767. doi: 10.1093/jnen/60.8.759. [DOI] [PubMed] [Google Scholar]
- 134.Swerdlow R. H., Burns J. M., Khan S. M. The Alzheimer’s disease mitochondrial cascade hypothesis. Journal of Alzheimer's Disease. 2010;20(Supplement 2):S265–S279. doi: 10.3233/JAD-2010-100339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Bertoni-Freddari C., Fattoretti P., Casoli T., Spagna C., Meier-Ruge W., Ulrich J. Morphological plasticity of synaptic mitochondria during aging. Brain Research. 1993;628(1-2):193–200. doi: 10.1016/0006-8993(93)90955-M. [DOI] [PubMed] [Google Scholar]
- 136.Brown M. R., Sullivan P. G., Geddes J. W. Synaptic mitochondria are more susceptible to Ca2+ overload than nonsynaptic mitochondria. Journal of Biological Chemistry. 2006;281(17):11658–11668. doi: 10.1074/jbc.M510303200. [DOI] [PubMed] [Google Scholar]
- 137.Graham L. C., Eaton S. L., Brunton P. J., et al. Proteomic profiling of neuronal mitochondria reveals modulators of synaptic architecture. Molecular Neurodegeneration. 2017;12(1):p. 77. doi: 10.1186/s13024-017-0221-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Brunel-Guitton C., Levtova A., Sasarman F. Mitochondrial diseases and cardiomyopathies. The Canadian Journal of Cardiology. 2015;31(11):1360–1376. doi: 10.1016/j.cjca.2015.08.017. [DOI] [PubMed] [Google Scholar]
- 139.Hirschey M. D., DeBerardinis R., Diehl A. M. E., et al. Dysregulated metabolism contributes to oncogenesis. Seminars in Cancer Biology. 2015;35:S129–S150. doi: 10.1016/j.semcancer.2015.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Wojtovich A. P., Nadtochiy S. M., Brookes P. S., Nehrke K. Ischemic preconditioning: the role of mitochondria and aging. Experimental Gerontology. 2012;47(1):1–7. doi: 10.1016/j.exger.2011.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Ertracht O., Malka A., Atar S., Binah O. The mitochondria as a target for cardioprotection in acute myocardial ischemia. Pharmacology & Therapeutics. 2014;142(1):33–40. doi: 10.1016/j.pharmthera.2013.11.003. [DOI] [PubMed] [Google Scholar]
- 142.Perrelli M. G., Pagliaro P., Penna C. Ischemia/reperfusion injury and cardioprotective mechanisms: role of mitochondria and reactive oxygen species. World Journal of Cardiology. 2011;3(6):186–200. doi: 10.4330/wjc.v3.i6.186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Ruiz-Meana M., Nunez E., Miro-Casas E., et al. Ischemic preconditioning protects cardiomyocyte mitochondria through mechanisms independent of cytosol. Journal of Molecular and Cellular Cardiology. 2014;68:79–88. doi: 10.1016/j.yjmcc.2014.01.001. [DOI] [PubMed] [Google Scholar]
- 144.Boylston J. A., Sun J., Chen Y., Gucek M., Sack M. N., Murphy E. Characterization of the cardiac succinylome and its role in ischemia-reperfusion injury. Journal of Molecular and Cellular Cardiology. 2015;88:73–81. doi: 10.1016/j.yjmcc.2015.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Chen Q., Younus M., Thompson J., Hu Y., Hollander J. M., Lesnefsky E. J. Intermediary metabolism and fatty acid oxidation: novel targets of electron transport chain-driven injury during ischemia and reperfusion. American Journal of Physiology-Heart and Circulatory Physiology. 2018;314(4):H787–H795. doi: 10.1152/ajpheart.00531.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Chouchani E. T., James A. M., Methner C., et al. Identification and quantification of protein S-nitrosation by nitrite in the mouse heart during ischemia. Journal of Biological Chemistry. 2017;292(35):14486–14495. doi: 10.1074/jbc.M117.798744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Lichardusova L., Tatarkova Z., Calkovska A., et al. Proteomic analysis of mitochondrial proteins in the guinea pig heart following long-term normobaric hyperoxia. Molecular and Cellular Biochemistry. 2017;434(1-2):61–73. doi: 10.1007/s11010-017-3037-6. [DOI] [PubMed] [Google Scholar]
- 148.Kalogeris T., Bao Y., Korthuis R. J. Mitochondrial reactive oxygen species: a double edged sword in ischemia/reperfusion vs preconditioning. Redox Biology. 2014;2:702–714. doi: 10.1016/j.redox.2014.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.James W. P. T. The epidemiology of obesity: the size of the problem. Journal of Internal Medicine. 2008;263(4):336–352. doi: 10.1111/j.1365-2796.2008.01922.x. [DOI] [PubMed] [Google Scholar]
- 150.Chen X., Wei S., Yang F. Mitochondria in the pathogenesis of diabetes: a proteomic view. Protein & Cell. 2012;3(9):648–660. doi: 10.1007/s13238-012-2043-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Sun K., Kusminski C. M., Scherer P. E. Adipose tissue remodeling and obesity. The Journal of Clinical Investigation. 2011;121(6):2094–2101. doi: 10.1172/JCI45887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Lindinger P. W., Christe M., Eberle A. N., et al. Important mitochondrial proteins in human omental adipose tissue show reduced expression in obesity. Journal of Proteomics. 2015;124:79–87. doi: 10.1016/j.jprot.2015.03.037. [DOI] [PubMed] [Google Scholar]
- 153.Kim S. J., Chae S., Kim H., et al. A protein profile of visceral adipose tissues linked to early pathogenesis of type 2 diabetes mellitus. Molecular & Cellular Proteomics. 2014;13(3):811–822. doi: 10.1074/mcp.M113.035501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Perez-Perez R., Garcia-Santos E., Ortega-Delgado F. J., et al. Attenuated metabolism is a hallmark of obesity as revealed by comparative proteomic analysis of human omental adipose tissue. Journal of Proteomics. 2012;75(3):783–795. doi: 10.1016/j.jprot.2011.09.016. [DOI] [PubMed] [Google Scholar]
- 155.Gomez-Serrano M., Camafeita E., Garcia-Santos E., et al. Proteome-wide alterations on adipose tissue from obese patients as age-, diabetes- and gender-specific hallmarks. Scientific Reports. 2016;6(1, article 25756) doi: 10.1038/srep25756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Perry R. J., Samuel V. T., Petersen K. F., Shulman G. I. The role of hepatic lipids in hepatic insulin resistance and type 2 diabetes. Nature. 2014;510(7503):84–91. doi: 10.1038/nature13478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Golabi P., Locklear C. T., Austin P., et al. Effectiveness of exercise in hepatic fat mobilization in non-alcoholic fatty liver disease: systematic review. World Journal of Gastroenterology. 2016;22(27):6318–6327. doi: 10.3748/wjg.v22.i27.6318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Paradies G., Paradies V., Ruggiero F. M., Petrosillo G. Oxidative stress, cardiolipin and mitochondrial dysfunction in nonalcoholic fatty liver disease. World Journal of Gastroenterology. 2014;20(39):14205–14218. doi: 10.3748/wjg.v20.i39.14205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Nuno-Lambarri N., Barbero-Becerra V. J., Uribe M., Chavez-Tapia N. C. Mitochondrial molecular pathophysiology of nonalcoholic fatty liver disease: a proteomics approach. International Journal of Molecular Sciences. 2016;17(3):p. 281. doi: 10.3390/ijms17030281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Thomas A., Klein M. S., Stevens A. P., et al. Changes in the hepatic mitochondrial and membrane proteome in mice fed a non-alcoholic steatohepatitis inducing diet. Journal of Proteomics. 2013;80:107–122. doi: 10.1016/j.jprot.2012.12.027. [DOI] [PubMed] [Google Scholar]
- 161.Valle A., Catalan V., Rodriguez A., et al. Identification of liver proteins altered by type 2 diabetes mellitus in obese subjects. Liver International. 2012;32(6):951–961. doi: 10.1111/j.1478-3231.2012.02765.x. [DOI] [PubMed] [Google Scholar]
- 162.Stachowicz A., Suski M., Olszanecki R., Madej J., Okon K., Korbut R. Proteomic analysis of liver mitochondria of apolipoprotein E knockout mice treated with metformin. Journal of Proteomics. 2012;77:167–175. doi: 10.1016/j.jprot.2012.08.015. [DOI] [PubMed] [Google Scholar]
- 163.Shi L., Wang Y., Tu S., et al. The responses of mitochondrial proteome in rat liver to the consumption of moderate ethanol: the possible roles of aldo-keto reductases. Journal of Proteome Research. 2008;7(8):3137–3145. doi: 10.1021/pr700853j. [DOI] [PubMed] [Google Scholar]
- 164.Venkatraman A., Landar A., Davis A. J., et al. Modification of the mitochondrial proteome in response to the stress of ethanol-dependent hepatotoxicity. Journal of Biological Chemistry. 2004;279(21):22092–22101. doi: 10.1074/jbc.M402245200. [DOI] [PubMed] [Google Scholar]
- 165.Potter M., Newport E., Morten K. J. The Warburg effect: 80 years on. Biochemical Society Transactions. 2016;44(5):1499–1505. doi: 10.1042/BST20160094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Kang S. W., Lee S., Lee E. K. ROS and energy metabolism in cancer cells: alliance for fast growth. Archives of Pharmacal Research. 2015;38(3):338–345. doi: 10.1007/s12272-015-0550-6. [DOI] [PubMed] [Google Scholar]
- 167.Uzhachenko R., Shanker A., Yarbrough W. G., Ivanova A. V. Mitochondria, calcium, and tumor suppressor Fus1: at the crossroad of cancer, inflammation, and autoimmunity. Oncotarget. 2015;6(25):20754–20772. doi: 10.18632/oncotarget.4537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Warburg O. On the origin of cancer cells. Science. 1956;123(3191):309–314. doi: 10.1126/science.123.3191.309. [DOI] [PubMed] [Google Scholar]
- 169.Zhou W., Liotta L. A., Petricoin E. F. The Warburg effect and mass spectrometry-based proteomic analysis. Cancer Genomics Proteomics. 2017;14(4):211–218. doi: 10.21873/cgp.20032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Jiang Y., Wang X. Comparative mitochondrial proteomics: perspective in human diseases. Journal of Hematology & Oncology. 2012;5(1):p. 11. doi: 10.1186/1756-8722-5-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Mendes M., Pelaez-Garcia A., Lopez-Lucendo M., et al. Mapping the spatial proteome of metastatic cells in colorectal cancer. Proteomics. 2017;17(19) doi: 10.1002/pmic.201700094. [DOI] [PubMed] [Google Scholar]
- 172.Qattan A. T., Radulovic M., Crawford M., Godovac-Zimmermann J. Spatial distribution of cellular function: the partitioning of proteins between mitochondria and the nucleus in MCF7 breast cancer cells. Journal of Proteome Research. 2012;11(12):6080–6101. doi: 10.1021/pr300736v. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Lopez-Campistrous A., Hao L., Xiang W., et al. Mitochondrial dysfunction in the hypertensive rat brain: respiratory complexes exhibit assembly defects in hypertension. Hypertension. 2008;51(2):412–419. doi: 10.1161/HYPERTENSIONAHA.107.102285. [DOI] [PubMed] [Google Scholar]
- 174.Ruiz-Romero C., Calamia V., Mateos J., et al. Mitochondrial dysregulation of osteoarthritic human articular chondrocytes analyzed by proteomics: a decrease in mitochondrial superoxide dismutase points to a redox imbalance. Molecular & Cellular Proteomics. 2009;8(1):172–189. doi: 10.1074/mcp.M800292-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.An T., Wang Y. F., Liu J. X., et al. Comparative analysis of proteomes between diabetic and normal human sperm: insights into the effects of diabetes on male reproduction based on the regulation of mitochondria-related proteins. Molecular Reproduction and Development. 2018;85(1):7–16. doi: 10.1002/mrd.22930. [DOI] [PubMed] [Google Scholar]
- 176.Samanta L., Agarwal A., Swain N., et al. Proteomic signatures of sperm mitochondria in varicocele: clinical use as biomarkers of varicocele associated infertility. The Journal of Urology. 2018;200(2):414–422. doi: 10.1016/j.juro.2018.03.009. [DOI] [PubMed] [Google Scholar]
- 177.Murphy M. P. How mitochondria produce reactive oxygen species. Biochemical Journal. 2009;417(1):1–13. doi: 10.1042/BJ20081386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Murphy M. P. Mitochondrial thiols in antioxidant protection and redox signaling: distinct roles for glutathionylation and other thiol modifications. Antioxidants & Redox Signaling. 2012;16(6):476–495. doi: 10.1089/ars.2011.4289. [DOI] [PubMed] [Google Scholar]
- 179.Halestrap A. P., Clarke S. J., Khaliulin I. The role of mitochondria in protection of the heart by preconditioning. Biochimica et Biophysica Acta (BBA) - Bioenergetics. 2007;1767(8):1007–1031. doi: 10.1016/j.bbabio.2007.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Kumar V., Kleffmann T., Hampton M. B., Cannell M. B., Winterbourn C. C. Redox proteomics of thiol proteins in mouse heart during ischemia/reperfusion using ICAT reagents and mass spectrometry. Free Radical Biology & Medicine. 2013;58:109–117. doi: 10.1016/j.freeradbiomed.2013.01.021. [DOI] [PubMed] [Google Scholar]
- 181.Yellon D. M., Hausenloy D. J. Myocardial reperfusion injury. The New England Journal of Medicine. 2007;357(11):1121–1135. doi: 10.1056/NEJMra071667. [DOI] [PubMed] [Google Scholar]
- 182.Wallace D. C. Mitochondria and cancer. Nature Reviews Cancer. 2012;12(10):685–698. doi: 10.1038/nrc3365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Weinberg F., Chandel N. S. Reactive oxygen species-dependent signaling regulates cancer. Cellular and Molecular Life Sciences. 2009;66(23):3663–3673. doi: 10.1007/s00018-009-0099-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Butterfield D. A., Gnjec A., Poon H. F., et al. Redox proteomics identification of oxidatively modified brain proteins in inherited Alzheimer’s disease: an initial assessment. Journal of Alzheimer's Disease. 2006;10(4):391–397. doi: 10.3233/JAD-2006-10407. [DOI] [PubMed] [Google Scholar]
- 185.Lin M. T., Beal M. F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature. 2006;443(7113):787–795. doi: 10.1038/nature05292. [DOI] [PubMed] [Google Scholar]
- 186.Held J. M., Gibson B. W. Regulatory control or oxidative damage? Proteomic approaches to interrogate the role of cysteine oxidation status in biological processes. Molecular & Cellular Proteomics. 2012;11(4, article R111.013037) doi: 10.1074/mcp.R111.013037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Lindahl M., Mata-Cabana A., Kieselbach T. The disulfide proteome and other reactive cysteine proteomes: analysis and functional significance. Antioxidants & Redox Signaling. 2011;14(12):2581–2642. doi: 10.1089/ars.2010.3551. [DOI] [PubMed] [Google Scholar]
- 188.Winterbourn C. C., Hampton M. B. Thiol chemistry and specificity in redox signaling. Free Radical Biology & Medicine. 2008;45(5):549–561. doi: 10.1016/j.freeradbiomed.2008.05.004. [DOI] [PubMed] [Google Scholar]
- 189.Rhee S. G., Jeong W., Chang T. S., Woo H. A. Sulfiredoxin, the cysteine sulfinic acid reductase specific to 2-Cys peroxiredoxin: its discovery, mechanism of action, and biological significance. Kidney International. 2007;72:S3–S8. doi: 10.1038/sj.ki.5002380. [DOI] [PubMed] [Google Scholar]
- 190.Muller F. L., Liu Y., Abdul-Ghani M. A., et al. High rates of superoxide production in skeletal-muscle mitochondria respiring on both complex I- and complex II-linked substrates. Biochemical Journal. 2008;409(2):491–499. doi: 10.1042/BJ20071162. [DOI] [PubMed] [Google Scholar]
- 191.Drose S. Differential effects of complex II on mitochondrial ROS production and their relation to cardioprotective pre- and postconditioning. Biochimica et Biophysica Acta (BBA) - Bioenergetics. 2013;1827(5):578–587. doi: 10.1016/j.bbabio.2013.01.004. [DOI] [PubMed] [Google Scholar]
- 192.Lanciano P., Khalfaoui-Hassani B., Selamoglu N., Ghelli A., Rugolo M., Daldal F. Molecular mechanisms of superoxide production by complex III: a bacterial versus human mitochondrial comparative case study. Biochimica et Biophysica Acta (BBA) - Bioenergetics. 2013;1827(11-12):1332–1339. doi: 10.1016/j.bbabio.2013.03.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Janssen-Heininger Y. M. W., Mossman B. T., Heintz N. H., et al. Redox-based regulation of signal transduction: principles, pitfalls, and promises. Free Radical Biology & Medicine. 2008;45(1):1–17. doi: 10.1016/j.freeradbiomed.2008.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Moncada S., Erusalimsky J. D. Does nitric oxide modulate mitochondrial energy generation and apoptosis? Nature Reviews Molecular Cell Biology. 2002;3(3):214–220. doi: 10.1038/nrm762. [DOI] [PubMed] [Google Scholar]
- 195.Galkin A., Meyer B., Wittig I., et al. Identification of the mitochondrial ND3 subunit as a structural component involved in the active/deactive enzyme transition of respiratory complex I. Journal of Biological Chemistry. 2008;283(30):20907–20913. doi: 10.1074/jbc.M803190200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Galkin A., Moncada S. S-nitrosation of mitochondrial complex I depends on its structural conformation. Journal of Biological Chemistry. 2007;282(52):37448–37453. doi: 10.1074/jbc.M707543200. [DOI] [PubMed] [Google Scholar]
- 197.Prime T. A., Blaikie F. H., Evans C., et al. A mitochondria-targeted S-nitrosothiol modulates respiration, nitrosates thiols, and protects against ischemia-reperfusion injury. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(26):10764–10769. doi: 10.1073/pnas.0903250106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Hurd T. R., Prime T. A., Harbour M. E., Lilley K. S., Murphy M. P. Detection of reactive oxygen species-sensitive thiol proteins by redox difference gel electrophoresis: implications for mitochondrial redox signaling. The Journal of Biological Chemistry. 2007;282(30):22040–22051. doi: 10.1074/jbc.M703591200. [DOI] [PubMed] [Google Scholar]
- 199.Chouchani E. T., Hurd T. R., Nadtochiy S. M., et al. Identification of S-nitrosated mitochondrial proteins by S-nitrosothiol difference in gel electrophoresis (SNO-DIGE): implications for the regulation of mitochondrial function by reversible S-nitrosation. Biochemical Journal. 2010;430(1):49–59. doi: 10.1042/BJ20100633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Scheving R., Wittig I., Heide H., et al. Protein S-nitrosylation and denitrosylation in the mouse spinal cord upon injury of the sciatic nerve. Journal of Proteomics. 2012;75(13):3987–4004. doi: 10.1016/j.jprot.2012.05.006. [DOI] [PubMed] [Google Scholar]
- 201.Chouchani E. T., Methner C., Nadtochiy S. M., et al. Cardioprotection by S-nitrosation of a cysteine switch on mitochondrial complex I. Nature Medicine. 2013;19(6):753–759. doi: 10.1038/nm.3212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Akter S., Carpentier S., Van Breusegem F., Messens J. Identification of dimedone-trapped sulfenylated proteins in plants under stress. Biochemistry and Biophysics Reports. 2017;9:106–113. doi: 10.1016/j.bbrep.2016.11.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Akter S., Huang J., Bodra N., et al. DYn-2 based identification of Arabidopsis Sulfenomes. Molecular & Cellular Proteomics. 2015;14(5):1183–1200. doi: 10.1074/mcp.M114.046896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Waszczak C., Akter S., Eeckhout D., et al. Sulfenome mining in Arabidopsis thaliana. Proceedings of the National Academy of Sciences of the United States of America. 2014;111(31):11545–11550. doi: 10.1073/pnas.1411607111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Hao G., Derakhshan B., Shi L., Campagne F., Gross S. S. SNOSID, a proteomic method for identification of cysteine S-nitrosylation sites in complex protein mixtures. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(4):1012–1017. doi: 10.1073/pnas.0508412103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Jaffrey S. R., Erdjument-Bromage H., Ferris C. D., Tempst P., Snyder S. H. Protein S-nitrosylation: a physiological signal for neuronal nitric oxide. Nature Cell Biology. 2001;3(2):193–197. doi: 10.1038/35055104. [DOI] [PubMed] [Google Scholar]
- 207.Jaffrey S. R., Snyder S. H. The biotin switch method for the detection of S-nitrosylated proteins. Science's STKE. 2001;2001, article pl1 doi: 10.1126/stke.2001.86.pl1. [DOI] [PubMed] [Google Scholar]
- 208.Murray C. I., Uhrigshardt H., O'Meally R. N., Cole R. N., Van Eyk J. E. Identification and quantification of S-nitrosylation by cysteine reactive tandem mass tag switch assay. Molecular & Cellular Proteomics. 2012;11(2, article M111 013441) doi: 10.1074/mcp.M111.013441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Charles R. L., Schröder E., May G., et al. Protein sulfenation as a redox sensor: proteomics studies using a novel biotinylated dimedone analogue. Molecular & Cellular Proteomics. 2007;6(9):1473–1484. doi: 10.1074/mcp.M700065-MCP200. [DOI] [PubMed] [Google Scholar]
- 210.Gupta V., Carroll K. S. Sulfenic acid chemistry, detection and cellular lifetime. Biochimica et Biophysica Acta (BBA) - General Subjects. 2014;1840(2):847–875. doi: 10.1016/j.bbagen.2013.05.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Poole L. B., Klomsiri C., Knaggs S. A., et al. Fluorescent and affinity-based tools to detect cysteine sulfenic acid formation in proteins. Bioconjugate Chemistry. 2007;18(6):2004–2017. doi: 10.1021/bc700257a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.McGarry D. J., Chakravarty P., Wolf C. R., Henderson C. J. Altered protein S-glutathionylation identifies a potential mechanism of resistance to acetaminophen-induced hepatotoxicity. The Journal of Pharmacology and Experimental Therapeutics. 2015;355(2):137–144. doi: 10.1124/jpet.115.227389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Gould N. S., Evans P., Martinez-Acedo P., et al. Site-specific proteomic mapping identifies selectively modified regulatory cysteine residues in functionally distinct protein networks. Chemistry & Biology. 2015;22(7):965–975. doi: 10.1016/j.chembiol.2015.06.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Gygi S. P., Rist B., Gerber S. A., Turecek F., Gelb M. H., Aebersold R. Quantitative analysis of complex protein mixtures using isotope-coded affinity tags. Nature Biotechnology. 1999;17(10):994–999. doi: 10.1038/13690. [DOI] [PubMed] [Google Scholar]
- 215.Ong S. E., Blagoev B., Kratchmarova I., et al. Stable isotope labeling by amino acids in cell culture, SILAC, as a simple and accurate approach to expression proteomics. Molecular & Cellular Proteomics. 2002;1(5):376–386. doi: 10.1074/mcp.M200025-MCP200. [DOI] [PubMed] [Google Scholar]
- 216.Ross P. L., Huang Y. N., Marchese J. N., et al. Multiplexed protein quantitation in Saccharomyces cerevisiae using amine-reactive isobaric tagging reagents. Molecular & Cellular Proteomics. 2004;3(12):1154–1169. doi: 10.1074/mcp.M400129-MCP200. [DOI] [PubMed] [Google Scholar]
- 217.Stewart I. I., Thomson T., Figeys D. 18O labeling: a tool for proteomics. Rapid Communications in Mass Spectrometry. 2001;15(24):2456–2465. doi: 10.1002/rcm.525. [DOI] [PubMed] [Google Scholar]
- 218.Thompson A., Schafer J., Kuhn K., et al. Tandem mass tags: a novel quantification strategy for comparative analysis of complex protein mixtures by MS/MS. Analytical Chemistry. 2003;75(8):1895–1904. doi: 10.1021/ac0262560. [DOI] [PubMed] [Google Scholar]
- 219.Weerapana E., Wang C., Simon G. M., et al. Quantitative reactivity profiling predicts functional cysteines in proteomes. Nature. 2010;468(7325):790–795. doi: 10.1038/nature09472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Leichert L. I., Gehrke F., Gudiseva H. V., et al. Quantifying changes in the thiol redox proteome upon oxidative stress in vivo. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(24):8197–8202. doi: 10.1073/pnas.0707723105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Fu C., Hu J., Liu T., Ago T., Sadoshima J., Li H. Quantitative analysis of redox-sensitive proteome with DIGE and ICAT. Journal of Proteome Research. 2008;7(9):3789–3802. doi: 10.1021/pr800233r. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Brandes N., Reichmann D., Tienson H., Leichert L. I., Jakob U. Using quantitative redox proteomics to dissect the yeast redoxome. The Journal of Biological Chemistry. 2011;286(48):41893–41903. doi: 10.1074/jbc.M111.296236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Garcia-Santamarina S., Boronat S., Domenech A., Ayte J., Molina H., Hidalgo E. Monitoring in vivo reversible cysteine oxidation in proteins using ICAT and mass spectrometry. Nature Protocols. 2014;9(5):1131–1145. doi: 10.1038/nprot.2014.065. [DOI] [PubMed] [Google Scholar]
- 224.Garcia-Santamarina S., Boronat S., Espadas G., Ayte J., Molina H., Hidalgo E. The oxidized thiol proteome in fission yeast—optimization of an ICAT-based method to identify H2O2-oxidized proteins. Journal of Proteomics. 2011;74(11):2476–2486. doi: 10.1016/j.jprot.2011.05.030. [DOI] [PubMed] [Google Scholar]
- 225.Lindemann C., Leichert L. I. Quantitative redox proteomics: the NOxICAT method. Methods in Molecular Biology. 2012;893:387–403. doi: 10.1007/978-1-61779-885-6_24. [DOI] [PubMed] [Google Scholar]
- 226.Knoefler D., Thamsen M., Koniczek M., Niemuth N. J., Diederich A. K., Jakob U. Quantitative in vivo redox sensors uncover oxidative stress as an early event in life. Molecular Cell. 2012;47(5):767–776. doi: 10.1016/j.molcel.2012.06.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Rosenwasser S., Graff van Creveld S., Schatz D., et al. Mapping the diatom redox-sensitive proteome provides insight into response to nitrogen stress in the marine environment. Proceedings of the National Academy of Sciences of the United States of America. 2014;111(7):2740–2745. doi: 10.1073/pnas.1319773111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Menger K. E., James A. M., Cocheme H. M., et al. Fasting, but not aging, dramatically alters the redox status of cysteine residues on proteins in Drosophila melanogaster. Cell Reports. 2015;13(6):p. 1285. doi: 10.1016/j.celrep.2015.10.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Go Y. M., Roede J. R., Orr M., Liang Y., Jones D. P. Integrated redox proteomics and metabolomics of mitochondria to identify mechanisms of Cd toxicity. Toxicological Sciences. 2014;139(1):59–73. doi: 10.1093/toxsci/kfu018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Topf U., Suppanz I., Samluk L., et al. Quantitative proteomics identifies redox switches for global translation modulation by mitochondrially produced reactive oxygen species. Nature Communications. 2018;9(1):p. 324. doi: 10.1038/s41467-017-02694-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.McGarry D. J., Chen W., Chakravarty P., Lamont D. L., Wolf C. R., Henderson C. J. Proteome-wide identification and quantification of S-glutathionylation targets in mouse liver. Biochemical Journal. 2015;469(1):25–32. doi: 10.1042/BJ20141256. [DOI] [PubMed] [Google Scholar]
- 232.Chouchani E. T., Kazak L., Jedrychowski M. P., et al. Mitochondrial ROS regulate thermogenic energy expenditure and sulfenylation of UCP1. Nature. 2016;532(7597):112–116. doi: 10.1038/nature17399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Kramer P. A., Duan J., Gaffrey M. J., et al. Fatiguing contractions increase protein S-glutathionylation occupancy in mouse skeletal muscle. Redox Biology. 2018;17:367–376. doi: 10.1016/j.redox.2018.05.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Martínez-Acedo P., Núñez E., Gómez F. J. S., et al. A novel strategy for global analysis of the dynamic thiol redox proteome. Molecular & Cellular Proteomics. 2012;11(9):800–813. doi: 10.1074/mcp.M111.016469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Navarro P., Trevisan-Herraz M., Bonzon-Kulichenko E., et al. General statistical framework for quantitative proteomics by stable isotope labeling. Journal of Proteome Research. 2014;13(3):1234–1247. doi: 10.1021/pr4006958. [DOI] [PubMed] [Google Scholar]
- 236.Mailloux R. J., Gardiner D., O’Brien M. 2-Oxoglutarate dehydrogenase is a more significant source of O2·−/H2O2 than pyruvate dehydrogenase in cardiac and liver tissue. Free Radical Biology & Medicine. 2016;97:501–512. doi: 10.1016/j.freeradbiomed.2016.06.014. [DOI] [PubMed] [Google Scholar]
- 237.Forman H. J., Davies M. J., Kramer A. C., et al. Protein cysteine oxidation in redox signaling: caveats on sulfenic acid detection and quantification. Archives of Biochemistry and Biophysics. 2017;617:26–37. doi: 10.1016/j.abb.2016.09.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Welter C., Meese E., Blin N. Rapid step-gradient purification of mitochondrial DNA. Molecular Biology Reports. 1988;13(2):117–120. doi: 10.1007/BF00539059. [DOI] [PubMed] [Google Scholar]
- 239.Caldera M., Buphamalai P., Müller F., Menche J. Interactome-based approaches to human disease. Current Opinion in Systems Biology. 2017;3:88–94. doi: 10.1016/j.coisb.2017.04.015. [DOI] [Google Scholar]
- 240.Szklarczyk D., Morris J. H., Cook H., et al. The STRING database in 2017: quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Research. 2017;45(D1):D362–D368. doi: 10.1093/nar/gkw937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Orchard S., Ammari M., Aranda B., et al. The MIntAct project—IntAct as a common curation platform for 11 molecular interaction databases. Nucleic Acids Research. 2014;42(D1):D358–D363. doi: 10.1093/nar/gkt1115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Salwinski L., Miller C. S., Smith A. J., Pettit F. K., Bowie J. U., Eisenberg D. The database of interacting proteins: 2004 update. Nucleic Acids Research. 2004;32(90001):449D–4451. doi: 10.1093/nar/gkh086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Miryala S. K., Anbarasu A., Ramaiah S. Discerning molecular interactions: a comprehensive review on biomolecular interaction databases and network analysis tools. Gene. 2018;642:84–94. doi: 10.1016/j.gene.2017.11.028. [DOI] [PubMed] [Google Scholar]
- 244.Barabási A.-L., Gulbahce N., Loscalzo J. Network medicine: a network-based approach to human disease. Nature Reviews Genetics. 2011;12(1):56–68. doi: 10.1038/nrg2918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Barabási A.-L., Oltvai Z. N. Network biology: understanding the cell’s functional organization. Nature Reviews Genetics. 2004;5(2):101–113. doi: 10.1038/nrg1272. [DOI] [PubMed] [Google Scholar]
- 246.Ewing R. M., Chu P., Elisma F., et al. Large-scale mapping of human protein–protein interactions by mass spectrometry. Molecular Systems Biology. 2007;3:p. 89. doi: 10.1038/msb4100134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Geiger T., Velic A., Macek B., et al. Initial quantitative proteomic map of 28 mouse tissues using the SILAC mouse. Molecular & Cellular Proteomics. 2013;12(6):1709–1722. doi: 10.1074/mcp.M112.024919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Wilhelm M., Schlegl J., Hahne H., et al. Mass-spectrometry-based draft of the human proteome. Nature. 2014;509(7502):582–587. doi: 10.1038/nature13319. [DOI] [PubMed] [Google Scholar]
- 249.Ngounou Wetie A. G., Sokolowska I., Woods A. G., Roy U., Loo J. A., Darie C. C. Investigation of stable and transient protein-protein interactions: past, present and future. Proteomics. 2013;13(3-4):538–557. doi: 10.1002/pmic.201200328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.Collins M. O., Choudhary J. S. Mapping multiprotein complexes by affinity purification and mass spectrometry. Current Opinion in Biotechnology. 2008;19(4):324–330. doi: 10.1016/j.copbio.2008.06.002. [DOI] [PubMed] [Google Scholar]
- 251.Selbach M., Mann M. Protein interaction screening by quantitative immunoprecipitation combined with knockdown (QUICK) Nature Methods. 2006;3(12):981–983. doi: 10.1038/nmeth972. [DOI] [PubMed] [Google Scholar]
- 252.Böttinger L., Guiard B., Oeljeklaus S., et al. A complex of Cox4 and mitochondrial Hsp70 plays an important role in the assembly of the cytochrome c oxidase. Molecular Biology of the Cell. 2013;24(17):2609–2619. doi: 10.1091/mbc.e13-02-0106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Runswick M. J., Bason J. V., Montgomery M. G., Robinson G. C., Fearnley I. M., Walker J. E. The affinity purification and characterization of ATP synthase complexes from mitochondria. Open Biology. 2013;3(2) doi: 10.1098/rsob.120160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Austin S., Bennett K. L., Nowikovsky K. Interactome of LETM1 using miniaturised affinity purification mass spectrometry. Biochimica et Biophysica Acta (BBA) - Bioenergetics. 2014;1837, article e107 doi: 10.1016/j.bbabio.2014.05.256. [DOI] [Google Scholar]
- 255.Sun Y., Vashisht A. A., Tchieu J., Wohlschlegel J. A., Dreier L. Voltage-dependent anion channels (VDACs) recruit parkin to defective mitochondria to promote mitochondrial autophagy. Journal of Biological Chemistry. 2012;287(48):40652–40660. doi: 10.1074/jbc.M112.419721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256.Malty R. H., Aoki H., Kumar A., et al. A map of human mitochondrial protein interactions linked to neurodegeneration reveals new mechanisms of redox homeostasis and NF-κB signaling. Cell Systems. 2017;5(6):564–577.e12. doi: 10.1016/j.cels.2017.10.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257.Trisciuoglio D., Desideri M., Farini V., et al. Affinity purification-mass spectrometry analysis of bcl-2 interactome identified SLIRP as a novel interacting protein. Cell Death & Disease. 2016;7(2, article e2090) doi: 10.1038/cddis.2015.357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Floyd B. J., Wilkerson E. M., Veling M. T., et al. Mitochondrial protein interaction mapping identifies regulators of respiratory chain function. Molecular Cell. 2016;63(4):621–632. doi: 10.1016/j.molcel.2016.06.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 259.Huttlin E. L., Bruckner R. J., Paulo J. A., et al. Architecture of the human interactome defines protein communities and disease networks. Nature. 2017;545(7655):505–509. doi: 10.1038/nature22366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260.Huttlin E. L., Ting L., Bruckner R. J., et al. The BioPlex network: a systematic exploration of the human interactome. Cell. 2015;162(2):425–440. doi: 10.1016/j.cell.2015.06.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 261.Schweppe D. K., Huttlin E. L., Harper J. W., Gygi S. P. BioPlex display: an interactive suite for large-scale AP–MS protein–protein interaction data. Journal of Proteome Research. 2018;17(1):722–726. doi: 10.1021/acs.jproteome.7b00572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Kristensen A. R., Gsponer J., Foster L. J. A high-throughput approach for measuring temporal changes in the interactome. Nature Methods. 2012;9(9):907–909. doi: 10.1038/nmeth.2131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Tang X., Bruce J. E. A new cross-linking strategy: protein interaction reporter (PIR) technology for protein–protein interaction studies. Molecular BioSystems. 2010;6(6):939–947. doi: 10.1039/b920876c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 264.Scott N. E., Rogers L. D., Prudova A., et al. Interactome disassembly during apoptosis occurs independent of caspase cleavage. Molecular Systems Biology. 2017;13(1):p. 906. doi: 10.15252/msb.20167067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265.Wessels H. J. C. T., Vogel R. O., van den Heuvel L., et al. LC-MS/MS as an alternative for SDS-PAGE in blue native analysis of protein complexes. Proteomics. 2009;9(17):4221–4228. doi: 10.1002/pmic.200900157. [DOI] [PubMed] [Google Scholar]
- 266.Wittig I., Carrozzo R., Santorelli F. M., Schägger H. Supercomplexes and subcomplexes of mitochondrial oxidative phosphorylation. Biochimica et Biophysica Acta (BBA) - Bioenergetics. 2006;1757(9-10):1066–1072. doi: 10.1016/j.bbabio.2006.05.006. [DOI] [PubMed] [Google Scholar]
- 267.Olinares P. D. B., Ponnala L., van Wijk K. J. Megadalton complexes in the chloroplast stroma of Arabidopsis thaliana characterized by size exclusion chromatography, mass spectrometry, and hierarchical clustering. Molecular & Cellular Proteomics. 2010;9(7):1594–1615. doi: 10.1074/mcp.M000038-MCP201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 268.Babu M., Vlasblom J., Pu S., et al. Interaction landscape of membrane-protein complexes in Saccharomyces cerevisiae. Nature. 2012;489(7417):585–589. doi: 10.1038/nature11354. [DOI] [PubMed] [Google Scholar]
- 269.Drew D., Newstead S., Sonoda Y., Kim H., von Heijne G., Iwata S. GFP-based optimization scheme for the overexpression and purification of eukaryotic membrane proteins in Saccharomyces cerevisiae. Nature Protocols. 2008;3(5):784–798. doi: 10.1038/nprot.2008.44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 270.Heide H., Bleier L., Steger M., et al. Complexome profiling identifies TMEM126B as a component of the mitochondrial complex I assembly complex. Cell Metabolism. 2012;16(4):538–549. doi: 10.1016/j.cmet.2012.08.009. [DOI] [PubMed] [Google Scholar]
- 271.Schägger H., von Jagow G. Blue native electrophoresis for isolation of membrane protein complexes in enzymatically active form. Analytical Biochemistry. 1991;199(2):223–231. doi: 10.1016/0003-2697(91)90094-A. [DOI] [PubMed] [Google Scholar]
- 272.Schägger H. [12] Native electrophoresis for isolation of mitochondrial oxidative phosphorylation protein complexes. Methods in Enzymology. 1995;260:190–202. doi: 10.1016/0076-6879(95)60137-6. [DOI] [PubMed] [Google Scholar]
- 273.Schagger H., Cramer W. A., Vonjagow G. Analysis of molecular masses and oligomeric states of protein complexes by blue native electrophoresis and isolation of membrane protein complexes by two-dimensional native electrophoresis. Analytical Biochemistry. 1994;217(2):220–230. doi: 10.1006/abio.1994.1112. [DOI] [PubMed] [Google Scholar]
- 274.Reisinger V., Eichacker L. A. Analysis of membrane protein complexes by blue native PAGE. Proteomics. 2006;6(S2):6–15. doi: 10.1002/pmic.200600553. [DOI] [PubMed] [Google Scholar]
- 275.Wittig I., Braun H.-P., Schagger H. Blue native PAGE. Nature Protocols. 2006;1(1):418–428. doi: 10.1038/nprot.2006.62. [DOI] [PubMed] [Google Scholar]
- 276.Helbig A. O., de Groot M. J. L., van Gestel R. A., et al. A three-way proteomics strategy allows differential analysis of yeast mitochondrial membrane protein complexes under anaerobic and aerobic conditions. Proteomics. 2009;9(20):4787–4798. doi: 10.1002/pmic.200800951. [DOI] [PubMed] [Google Scholar]
- 277.Andersen J. S., Wilkinson C. J., Mayor T., Mortensen P., Nigg E. A., Mann M. Proteomic characterization of the human centrosome by protein correlation profiling. Nature. 2003;426(6966):570–574. doi: 10.1038/nature02166. [DOI] [PubMed] [Google Scholar]
- 278.Larance M., Lamond A. I. Multidimensional proteomics for cell biology. Nature Reviews Molecular Cell Biology. 2015;16(5):269–280. doi: 10.1038/nrm3970. [DOI] [PubMed] [Google Scholar]
- 279.Cogliati S., Calvo E., Loureiro M., et al. Mechanism of super-assembly of respiratory complexes III and IV. Nature. 2016;539(7630):579–582. doi: 10.1038/nature20157. [DOI] [PubMed] [Google Scholar]
- 280.Guerrero-Castillo S., Baertling F., Kownatzki D., et al. The assembly pathway of mitochondrial respiratory chain complex I. Cell Metabolism. 2017;25(1):128–139. doi: 10.1016/j.cmet.2016.09.002. [DOI] [PubMed] [Google Scholar]
- 281.Müller C. S., Bildl W., Haupt A., et al. Cryo-slicing blue native-mass spectrometry (csBN-MS), a novel technology for high resolution complexome profiling. Molecular & Cellular Proteomics. 2016;15(2):669–681. doi: 10.1074/mcp.M115.054080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282.Wessels H. J. C. T., Vogel R. O., Lightowlers R. N., et al. Analysis of 953 human proteins from a mitochondrial HEK293 fraction by complexome profiling. PLoS One. 2013;8(7, article e68340) doi: 10.1371/journal.pone.0068340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 283.Iwamoto N., Shimada T. Recent advances in mass spectrometry-based approaches for proteomics and biologics: great contribution for developing therapeutic antibodies. Pharmacology & Therapeutics. 2018;185:147–154. doi: 10.1016/j.pharmthera.2017.12.007. [DOI] [PubMed] [Google Scholar]
- 284.Havugimana P. C., Hart G. T., Nepusz T., et al. A census of human soluble protein complexes. Cell. 2012;150(5):1068–1081. doi: 10.1016/j.cell.2012.08.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 285.Kirkwood K. J., Ahmad Y., Larance M., Lamond A. I. Characterization of native protein complexes and protein isoform variation using size-fractionation-based quantitative proteomics. Molecular & Cellular Proteomics. 2013;12(12):3851–3873. doi: 10.1074/mcp.M113.032367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 286.Liu F., Lössl P., Rabbitts B. M., Balaban R. S., Heck A. J. R. The interactome of intact mitochondria by cross-linking mass spectrometry provides evidence for coexisting respiratory supercomplexes. Molecular & Cellular Proteomics. 2018;17(2):216–232. doi: 10.1074/mcp.RA117.000470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 287.Schweppe D. K., Chavez J. D., Lee C. F., et al. Mitochondrial protein interactome elucidated by chemical cross-linking mass spectrometry. Proceedings of the National Academy of Sciences of the United States of America. 2017;114(7):1732–1737. doi: 10.1073/pnas.1617220114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 288.Greber B. J., Boehringer D., Leitner A., et al. Architecture of the large subunit of the mammalian mitochondrial ribosome. Nature. 2014;505(7484):515–519. doi: 10.1038/nature12890. [DOI] [PubMed] [Google Scholar]
- 289.Fritzsche R., Ihling C. H., Götze M., Sinz A. Optimizing the enrichment of cross-linked products for mass spectrometric protein analysis. Rapid Communications in Mass Spectrometry. 2012;26(6):653–658. doi: 10.1002/rcm.6150. [DOI] [PubMed] [Google Scholar]
- 290.Leitner A., Reischl R., Walzthoeni T., et al. Expanding the chemical cross-linking toolbox by the use of multiple proteases and enrichment by size exclusion chromatography. Molecular & Cellular Proteomics. 2012;11(3, article M111.014126) doi: 10.1074/mcp.M111.014126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291.Trester-Zedlitz M., Kamada K., Burley S. K., Fenyö D., Chait B. T., Muir T. W. A modular cross-linking approach for exploring protein interactions. Journal of the American Chemical Society. 2003;125(9):2416–2425. doi: 10.1021/ja026917a. [DOI] [PubMed] [Google Scholar]
- 292.Kao A., Chiu C. L., Vellucci D., et al. Development of a novel cross-linking strategy for fast and accurate identification of cross-linked peptides of protein complexes. Molecular & Cellular Proteomics. 2011;10(1, article M110.002212) doi: 10.1074/mcp.M110.002212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 293.Kölbel K., Ihling C. H., Sinz A. Analysis of peptide secondary structures by photoactivatable amino acid analogues. Angewandte Chemie International Edition. 2012;51(50):12602–12605. doi: 10.1002/anie.201205308. [DOI] [PubMed] [Google Scholar]
- 294.Leitner A., Joachimiak L. A., Unverdorben P., et al. Chemical cross-linking/mass spectrometry targeting acidic residues in proteins and protein complexes. Proceedings of the National Academy of Sciences. 2014;111(26):9455–9460. doi: 10.1073/pnas.1320298111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 295.Müller M. Q., Dreiocker F., Ihling C. H., Schäfer M., Sinz A. Fragmentation behavior of a thiourea-based reagent for protein structure analysis by collision-induced dissociative chemical cross-linking. Journal of Mass Spectrometry. 2010;45(8):880–891. doi: 10.1002/jms.1775. [DOI] [PubMed] [Google Scholar]
- 296.Rinner O., Seebacher J., Walzthoeni T., et al. Identification of cross-linked peptides from large sequence databases. Nature Methods. 2008;5(4):315–318. doi: 10.1038/nmeth.1192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 297.Yu C., Kandur W., Kao A., Rychnovsky S., Huang L. Developing new isotope-coded mass spectrometry-cleavable cross-linkers for elucidating protein structures. Analytical Chemistry. 2014;86(4):2099–2106. doi: 10.1021/ac403636b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 298.Chu F., Baker P. R., Burlingame A. L., Chalkley R. J. Finding chimeras: a bioinformatics strategy for identification of cross-linked peptides. Molecular & Cellular Proteomics. 2010;9(1):25–31. doi: 10.1074/mcp.M800555-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 299.Götze M., Pettelkau J., Schaks S., et al. StavroX—a software for analyzing crosslinked products in protein interaction studies. Journal of the American Society for Mass Spectrometry. 2012;23(1):76–87. doi: 10.1007/s13361-011-0261-2. [DOI] [PubMed] [Google Scholar]
- 300.Hoopmann M. R., Weisbrod C. R., Bruce J. E. Improved strategies for rapid identification of chemically cross-linked peptides using protein interaction reporter technology. Journal of Proteome Research. 2010;9(12):6323–6333. doi: 10.1021/pr100572u. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301.Hoopmann M. R., Zelter A., Johnson R. S., et al. Kojak: efficient analysis of chemically cross-linked protein complexes. Journal of Proteome Research. 2015;14(5):2190–2198. doi: 10.1021/pr501321h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 302.Liu F., Rijkers D. T. S., Post H., Heck A. J. R. Proteome-wide profiling of protein assemblies by cross-linking mass spectrometry. Nature Methods. 2015;12(12):1179–1184. doi: 10.1038/nmeth.3603. [DOI] [PubMed] [Google Scholar]
- 303.Weisbrod C. R., Chavez J. D., Eng J. K., Yang L., Zheng C., Bruce J. E. In vivo protein interaction network identified with a novel real-time cross-linked peptide identification strategy. Journal of Proteome Research. 2013;12(4):1569–1579. doi: 10.1021/pr3011638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 304.Xu H., Hsu P.-H., Zhang L., Tsai M.-D., Freitas M. A. Database search algorithm for identification of intact cross-links in proteins and peptides using tandem mass spectrometry. Journal of Proteome Research. 2010;9(7):3384–3393. doi: 10.1021/pr100369y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 305.Yang B., Wu Y. J., Zhu M., et al. Identification of cross-linked peptides from complex samples. Nature Methods. 2012;9(9):904–906. doi: 10.1038/nmeth.2099. [DOI] [PubMed] [Google Scholar]
- 306.Combe C. W., Fischer L., Rappsilber J. xiNET: cross-link network maps with residue resolution. Molecular & Cellular Proteomics. 2015;14(4):1137–1147. doi: 10.1074/mcp.O114.042259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 307.Grimm M., Zimniak T., Kahraman A., Herzog F. xVis: a web server for the schematic visualization and interpretation of crosslink-derived spatial restraints. Nucleic Acids Research. 2015;43(W1):W362–W369. doi: 10.1093/nar/gkv463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 308.Kosinski J., von Appen A., Ori A., Karius K., Müller C. W., Beck M. Xlink analyzer: software for analysis and visualization of cross-linking data in the context of three-dimensional structures. Journal of Structural Biology. 2015;189(3):177–183. doi: 10.1016/j.jsb.2015.01.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 309.Schweppe D. K., Chavez J. D., Bruce J. E. XLmap: an R package to visualize and score protein structure models based on sites of protein cross-linking. Bioinformatics. 2016;32(2):306–308. doi: 10.1093/bioinformatics/btv519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 310.Zheng C., Weisbrod C. R., Chavez J. D., et al. XLink-DB: database and software tools for storing and visualizing protein interaction topology data. Journal of Proteome Research. 2013;12(4):1989–1995. doi: 10.1021/pr301162j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 311.Leitner A., Faini M., Stengel F., Aebersold R. Crosslinking and mass spectrometry: an integrated technology to understand the structure and function of molecular machines. Trends in Biochemical Sciences. 2016;41(1):20–32. doi: 10.1016/j.tibs.2015.10.008. [DOI] [PubMed] [Google Scholar]
- 312.Sprinzak E., Cokus S. J., Yeates T. O., Eisenberg D., Pellegrini M. Detecting coordinated regulation of multi-protein complexes using logic analysis of gene expression. BMC Systems Biology. 2009;3(1):p. 115. doi: 10.1186/1752-0509-3-115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 313.Hofer A., Wenz T. Post-translational modification of mitochondria as a novel mode of regulation. Experimental Gerontology. 2014;56:202–220. doi: 10.1016/j.exger.2014.03.006. [DOI] [PubMed] [Google Scholar]