

Abstract
Much evidence for non-human culture comes from vocally learned displays, such as the vocal dialects and song displays of birds and cetaceans. While many oscine birds use song complexity to assess male fitness, the role of complexity in humpback whale (Megaptera novaeangliae) song is uncertain owing to population-wide conformity to one song pattern. Although songs change gradually each year, the eastern Australian population also completely replaces their song every few years in cultural ‘revolutions’. Revolutions involve learning large amounts of novel material introduced from the Western Australian population. We examined two measures of song structure, complexity and entropy, in the eastern Australian population over 13 consecutive years. These measures aimed to identify the role of complexity and information content in the vocal learning processes of humpback whales. Complexity was quantified at two hierarchical levels: the entire sequence of individual sound ‘units’ and the stereotyped arrangements of units which comprise a ‘theme’. Complexity increased as songs evolved over time but decreased when revolutions occurred. No correlation between complexity and entropy estimates suggests that changes to complexity may represent embellishment to the song which could allow males to stand out amidst population-wide conformity. The consistent reduction in complexity during song revolutions suggests a potential limit to the social learning capacity of novel material in humpback whales.
Keywords: animal culture, humpback whale, song complexity, social learning, cultural revolutions
1. Background
Vocally learned traits, such as song displays in bird and mammal species, are an important focus in non-human cultural studies (reviewed by Laland & Janik [1]). These vocal learning displays fall along a broad gradient of complexity in their arrangements [2], from the simple blue whale (Balaenoptera musculus) songs [3] to highly complex birdsongs such as those produced by the zebra finch (Taeniopygia guttate) [4,5]. Where a display fits on this gradient can provide insights into the learning mechanisms involved or the function behind a display. For example, high song complexity correlates with sexual selection in several oscine species [6,7], thus functioning as a metric of male fitness. Increased song complexity or repertoire size in some species may also reflect an increased learning capacity [8,9]. Known as the ‘cognitive capacity hypothesis' [9,10], this suggests that highly complex songs might indicate more developed cognitive abilities [11] and therefore drive female sexual selection for males with more complex songs [9].
A well-studied song display, which is culturally learned, is that of male humpback whales (Megaptera novaeangliae) [12–14]. These songs have a hierarchical structure and are sung in a stereotyped pattern [13,15]. Sounds or ‘units’ of various types are arranged into a ‘phrase’, and phrases are repeated multiple times to create a ‘theme’. Four to seven different themes sung in a consistent order create a ‘song’ [13]. Despite males in a given population conforming to the same song [16], the pattern of the song itself constantly changes [17]. In most humpback whale populations, songs change progressively each year through cultural evolutions [14,16,17]. In the eastern Australian population however, ‘cultural revolutions' also occur every few years where the song is rapidly and completely replaced by a song from another population [12,18]. Songs from eastern Australia then spread eastwards across the South Pacific populations over 1–3 years [14]. This demonstrates cultural transmissions in an animal on spatial and temporal scales that have previously only been documented in humans [14].
The current study quantified several metrics of song complexity in the eastern Australian population over 13 consecutive years in order to clarify the underlying song learning mechanism. Complexity scores were derived from several arrangement features and used to measure changes in song complexity over time [10]. Three sets of complexity scores were calculated to reflect the song's hierarchical structure: (i) song-level (number of units, number of types of units and song duration), (ii) theme-level (number of themes, mean phrase duration and mean theme complexity), and (iii) a score combining song-level and theme-level scores. Complexity was also evaluated in terms of both pattern predictability (using second-order entropy) and individual-level variation. Quantifying these components of complexity in humpback whale song will inform our understanding of the social learning capacity in humpback whales. As one of the few examples of vocal learning in a mammalian species, humpback whales add an important piece to the knowledge of vocal learning use across multiple taxa.
2. Material and methods
(a). Song collection and transcription
Recordings of humpback whale song were made every year from 2002 to 2014 using fixed hydrophone arrays, autonomous recorders and boat-based recordings. Study sites were along the migratory corridor for eastern Australian humpback whales on the coast of southeast Queensland, Australia [19]. Songs were visualized as spectrograms and transcribed by a human classifier (J.A.A.) into a numeric sequence of units using a repertoire ‘dictionary’ as a guide [20]. This dictionary was created based on acoustic features of a representative subset of units from the eastern Australian population [20]. Song cycles from each year were transcribed and analysed [20], with a ‘song cycle’ defined here as a stereotyped set of themes sung in a specific order [13,15,16]. Thirty-six song cycles were transcribed from each year, except for 2006 (12 cycles) and 2007 (four cycles) owing to lack of high-quality recordings. No more than six song cycles were taken from any single individual. Because these recordings were made during migration, it was assumed that singers on separated days had not been previously recorded [21–23]. Song cycles were considered to be from one individual if they occurred in succession with no break in singing.
(b). Verifying themes and transitions
Unit sequences were grouped into phrase variants and assigned to themes. Themes were quantitatively identified and verified using the Levenshtein distance similarity index (LSI) [24,25]. This analysis was weighted (β = 1) to account for acoustic similarity of units [20,26]. To ensure that qualitative theme assignments were robust, LSI was calculated between every possible pair of phrase repetitions in a given year and hierarchically clustered [20]. See Garland et al. [26] for details on the LSI analysis, acoustic dictionary and unit classification.
A single phrase for each theme per year was used as a theme representative in further analysis (known as the ‘set median’). Each set median was calculated by finding the phrase repetition with the highest similarity to all other phrases of a given theme [27]. LSI values were calculated between every pair of theme set medians in adjacent years. These values generated a cost matrix for a weighted LSI analysis of each year's song based on theme sequences. This analysis was used to classify each year's song as either an evolution or a revolution [18]. Songs were classified as an evolution if similarity to the previous year's song was greater than 0%. Revolutions were songs that displayed 0% similarity between itself and the preceding song.
(c). Complexity scores
‘Complexity scores’ were generated for each year's song following Boogert et al. [9] and Templeton et al. [10]. Song-level variables represented the full sequence of units (i.e. the complexity of the entire song irrespective of themes), and theme-level variables represented complexity with respect to the presence and content of themes. Three complexity scores were generated: one using song-level variables, one using theme-level variables and one using all variables from both levels. Three song-level variables were used: number of units per song cycle, number of unit types per song cycle and duration of each complete song cycle. Three theme-level variables were used: number of themes per song cycle, mean phrase duration per song cycle and mean individual theme complexity (calculated similarly to song complexity—number of units for each phrase, number of types of units per phrase and theme duration). All variables were condensed using a principal component analysis and represented by a single composite score [9,10]. For themes present in multiple years (i.e. evolving themes), individual theme complexity scores were compared among years.
(d). Individuality
Variability in the songs of individual whales may present another potential source of song complexity [28]. While a low number of high-quality recordings in 2006 and 2007 prevented the inclusion of the individual variable in complexity scores, it was examined separately using the presence of individually unique phrase variants (i.e. subtle variants of phrases sung only by a single individual) as a metric of ‘individuality’. An additional LSI analysis separated all phrases in each given year into ‘phrase variants’, representing slightly different arrangements of the same theme. Unique phrase variants were sung by a single individual, while shared phrase variants were sung by at least two individuals [25]. Data from 2006 (n = 2 singers) and 2007 (n = 1) were excluded to ensure a representative sample size per year [18,29]. The total phrase repetitions recorded each year (including all recorded repetitions of each phrase variant) were counted. Of these, the proportion that was unique phrase variants was calculated. These proportions were then averaged by the number of individual singers to standardize for sampling effort in each year.
(e). Information entropy estimates
Second-order entropy quantifies how predictably a sequence is arranged [30,31], which addresses an aspect of song arrangement that complexity scores miss. In the context of vocalizations, lower entropy levels typically represent a more redundant message within a display [32]. Entropy estimates were calculated using the following equation:
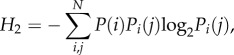 |
where H2 is second-order entropy, P(i) is the probability of occurrence of element i, Pi(j) is the probability of occurrence of unit j given the preceding unit is element i and N is the number of elements (types of units) in the repertoire [30]. Entropy is measured as the number of bits per song cycle. Entropy estimates for each year were calculated separately for: (i) sequences of themes (i.e. the pattern of different themes making up a song, omitting the units that made up those themes), and (ii) sequences of units (i.e. the pattern of units making up the entire song).
3. Results
Complexity scores formed a quasi-sinusoidal pattern over the 13-year study period (figure 1), based on a total of 412 song cycles from 95 singers (range: 1–13 singers per year, mean: 7). Complexity increased as songs evolved, resulting in longer songs containing more sound units, unit types and themes. Following revolutions, complexity decreased so that new songs were shorter and contained fewer units, unit types and themes. All three sets of scores were found to be highly correlated (figure 1), indicating that these trends were present at both the song and theme hierarchical levels.
Figure 1.

Song complexity scores for each year (2002–2014) representing complexity at the (i) song-level, (ii) theme-level, and (iii) total complexity. Revolution and evolution transitions are demarcated.
During periods of evolutionary song change, some themes persisted, but evolved across multiple years [14,17]. To quantify changes in complexity specific to this level of the song, complexity scores were generated for each of these themes year by year. Complexity increased for all evolving themes, except for those from 2011 to 2012 (figure 2), with most themes becoming longer and containing more units and unit types. These results show that during evolutionary song change, complexity increased within each of the themes, not just in the arrangement of the song as a whole.
Figure 2.

Individual theme complexity scores for evolving themes. Each theme's complexity scores over time are within its respective box. Boxes contain each set of evolutionary themes, meaning that the theme progressively evolves from 1 year to the next (as indicated by the years on the x-axis). Each theme was labelled with two numbers indicating the year (02–14) and a letter to differentiate it from the other themes in each respective year. All other themes occurred in a single year.
Of the phrase repetitions recorded in each year (range: 703–1863, mean: 1158), the proportion that was unique variants and thus represented ‘individuality’ ranged from 13% to 59% (mean = 34%). These proportions represent an average per singer in each year (range: 6–13, mean: 8) to standardize the metric for sampling effort. Individuality was correlated with all three complexity scores, most notably during evolutions when both individuality and complexity increased over time. Conversely, individuality did not consistently decrease when complexity decreased. However, the paucity of data from 2006 and 2007 reduced the strength of these results, warranting caution in their interpretation.
Sequential predictability, measured by second-order entropy, determined if more complex songs were less predictable in the arrangement of their theme or unit sequences. Theme sequence entropy (predictability of theme order in a song) had an average of 0.3 bits per song cycle (range 0.04–0.6 bits), while unit sequence entropy for entire song cycles had an average of 1.2 bits per song cycle (range 0.7–1.8 bits). There was no significant relationship between complexity and entropy estimates (Pearson correlation coefficient with Bonferroni correction, p = 0.52 for unit sequences, p = 0.17 for theme sequences) and entropy showed no clear patterns over time (figure 3). Therefore, predictability in a song arrangement was not strongly correlated with the complexity of that arrangement.
Figure 3.

Second-order entropy estimates for theme sequences and unit sequences from 2002 to 2014 in relation to revolution and evolution transitions.
4. Discussion
A long-term analysis of multiple song complexity metrics found that songs increased in complexity as they evolved through small and progressive changes. Parallel trends were also observed in individual themes; complexity usually increased over time with evolutionary changes (excepting 2011 and 2012). During the radical changes of revolution events, songs were always completely replaced with a simpler song. This relationship remained consistent at both the song-level (units, unit types and song duration) and theme-level (number of themes, mean phrase duration and mean theme complexity). Changes at both levels of the song's structure therefore contribute to changes in complexity.
Progressive song changes are thought to come from the novel material each singer adds to his own song [12,14,15]. Singers may embellish their own songs (thus increasing complexity) in order to stand out amidst the population's persistent conformity [28]. The positive correlation between complexity and individuality found here supports this, suggesting that as songs become more complex, they also become more individually unique. Therefore, humpback whale song changes could possibly complement the ‘anti-habituation hypothesis', which proposes that variation in bird song prevents individuals from becoming so accustomed to a song that it no longer serves its function [33]. Individuals may also be attempting to maintain this individuality even when adopting a new, simpler song, accounting for the lack of parallel decreases in complexity and individuality observed. However, the degree of individuality present in a population may also be impacted by other variables, such as the substantial increase in population size documented over this study period [34,35].
The absence of correlation between entropy estimates and complexity indicates that there may be independent variation between the complexity of a song pattern and how predictably that pattern is arranged. Previous unit-level entropy estimates (eastern Australian 2003 song mean = 0.97 bits [36], Hawaiian song mean = 0.99 bits [32]) using first-order Markov models (equivalent to the methods used here) [32,36] fall within our reported range (0.7–1.8 bits, mean = 1.2 bits). While the predictability demonstrated by these levels of entropy is expected in a stereotyped display such as humpback whale song, it was unexpected that predictability would remain relatively consistent over time despite changes to song complexity or arrangement [37]. Such predictability is likely to make songs easier to learn and remember, even with complex or novel arrangements, therefore possibly increasing chances for its spread.
Cultural revolution events in eastern Australia originate from new songs introduced from Western Australia with a 1 or 2 year delay [18]. Within eastern Australia, these revolutionary songs are less complex than their predecessors. The reasons for this are not clear, but several possibilities exist. One is that revolutions contain large amounts of novel material, and singers may be constrained by how much new material they can learn in a finite period [38]. This suggests a potential limit to the social learning rate in humpback whales. A second possibility is that Western Australian songs are simpler than eastern Australian ones and are learned with high fidelity. This seems unlikely as previous work suggests no overt differences in structure or complexity between Western and eastern Australian songs [39]. A third possibility is that simplification during revolutions occurs owing to preference rather than necessity. It may be more advantageous to take up a completely novel (albeit simpler) song than to add complexity to the current song [28]. It is also possible that simplification is a by-product of a limited number of song models. Few Western Australian individuals are likely to be involved in song transmission, regardless of whether Western Australian individuals immigrate into the eastern Australian population, or whether song transmission occurs on shared feeding grounds or migratory routes [40]. This would result in the transfer of only part of the full diversity of Western Australian songs to eastern Australia. This seems unlikely, however, as research suggests equal variation in song both within and between individuals [28,41]. Future research should compare song complexity between analogous songs in east and west Australia to determine which of the proposed explanations is most probable. Additionally, quantifying complexity patterns in other populations without revolution events would determine how cultural revolutions influence song learning and clarify what learning constraints might be present across populations.
5. Conclusion
Our study presents a consistent pattern in which songs become more complex as they evolve but are simplified following cultural revolution events. Large amounts of novel material presented in these revolutions, combined with the fluctuating song complexity identified here, suggest that simplification may result from a possible limit on the ability of humpback whales to learn new material. Additionally, the lack of correlation with entropy, despite changes to song complexity and arrangement, suggests embellishment in the song pattern rather than changes to underlying structure. The complexity of vocal displays provides important insights into the structure, learning mechanisms and evolution of animal communication [31,42]. Although studied to varying degrees in several species [9,37,43–45], limited research exists on structural complexity in humpback whale song [25,32]. The song is a striking example of a culturally transmitted display in animals. Changes in complexity over time can help clarify influential factors in the processes of song learning and cultural transmission, such as possible constraints on social learning capacity. This will lead to a more comprehensive grasp of cultural learning in animals, which can, in turn, help us to better understand its evolution and current role in humans.
Acknowledgements
We thank the staff and volunteers of the Cetacean Ecology and Acoustics Laboratory for data collection, maintenance and storage over the course of the study. Data were collected during the HARC project (2002–2004 and 2008–2009) and the BRAHSS project (2010–2012 and 2014). Thanks to Douglas Cato for his significant role in instigating and participating in all these field programmes. Thanks to Anita Murray for her advice regarding the theme verification technique and to Melinda (Mindi) Rekdahl for her invaluable insights into revolution events and the Western Australian song. We thank Adam Pack and Christopher Templeton for providing comments on a previous version of the manuscript.
Ethics
The following ethics approval numbers pertain to data collected for this study (2002–2004), the University of Queensland Office of Research and Postgraduate Studies (2005–2007), the University of Queensland Research and Research Training Division (2008–2011), the University of Queensland Research Management Office (2012–2014): 2002–2003: ZOO/ENT/250/02/USNR/DSTO; 2003–2004: ZOO/ENT/216/03/UNNR/DSTO; 2004–2005: ZOO/ENT/239/04/USNR/DSTO; 2005–2006: SVS/381/05/DSTO & US ONR; 2006–2007: SVS/870/06/DSTO & US ONR; 2007–2008: SVS/203/07/DSTO & US ONR; 2008–2009: SVS/299/08/ACAMMS; 2010–2012: SVS/230/10/(NF); 2012–2013: SVS/403/12/EPSML; 2013–2014: CURTIN/SVS/283/13. Certificates available upon request
Data accessibility
Data for this manuscript are available on Dryad Digital Repository at: http://dx.doi.org/10.5061/dryad.69161bg [46].
Authors' contributions
R.A.D. provided extensive raw song data, assisted with interpretation of results and edited the final manuscript. E.C.G. contributed to the development of the research concept and methodology, assisted with interpretation of results and edited the final manuscript. M.J.N. provided extensive raw song data, contributed to the development of the research concept and methodology, assisted with interpretation of results and edited the final manuscript. J.A.A. contributed to the development of the research concept, transcribed all song recordings, developed methodology, undertook data analysis and interpretation, and was responsible for the final manuscript.
Competing interests
We have no competing interests.
Funding
J.A.A. was funded by an Australian Government Research Training Programme Scholarship and the Australian American Association University of Queensland Fellowship. E.C.G. was funded by a Royal Society Newton International Fellowship and a Royal Society University Research Fellowship. The HARC project was funded by the US Office of Naval Research, the Australian Defence Science and Technology Organisation and the Australian Marine Mammal Centre. The BRAHSS project was funded by the E&P Sound and Marine Life Joint Industry Programme and the US Bureau of Ocean Energy Management.
References
- 1.Laland KN, Janik VM. 2006. The animal cultures debate. Trends Ecol. Evol. 21, 542–547. ( 10.1016/j.tree.2006.06.005) [DOI] [PubMed] [Google Scholar]
- 2.Janik VM, Slater PJ. 2000. The different roles of social learning in vocal communication. Anim. Behav. 60, 1–11. ( 10.1006/anbe.2000.1410) [DOI] [PubMed] [Google Scholar]
- 3.McDonald MA, Mesnick SL, Hildebrand JA. 2006. Biogeographic characterization of blue whale song worldwide: using song to identify populations. J. Cetacean Res. Manage. 8, 55–65. [Google Scholar]
- 4.Catchpole CK, Slater PJ. 2008. Bird song: biological themes and variations, 2nd edn Cambridge, UK: Cambridge University Press. [Google Scholar]
- 5.Airey DC, DeVoogd TJ. 2000. Greater song complexity is associated with augmented song system anatomy in zebra finches. Neuroreport 11, 1749–1754. ( 10.1097/00001756-200006050-00030) [DOI] [PubMed] [Google Scholar]
- 6.Nowicki S, Peters S, Podos J. 1998. Song learning, early nutrition and sexual selection in songbirds. Am. Zool. 38, 179–190. ( 10.1093/icb/38.1.179) [DOI] [Google Scholar]
- 7.Woodgate JL, Mariette MM, Bennett AT, Griffith SC, Buchanan KL. 2012. Male song structure predicts reproductive success in a wild zebra finch population. Anim. Behav. 83, 773–781. ( 10.1016/j.anbehav.2011.12.027) [DOI] [Google Scholar]
- 8.Nowicki S, Hasselquist D, Bensch S, Peters S. 2000. Nestling growth and song repertoire size in great reed warblers: evidence for song learning as an indicator mechanism in mate choice. Proc. R. Soc. Lond. B 267, 2419–2424. ( 10.1098/rspb.2000.1300) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Boogert NJ, Giraldeau L-A, Lefebvre L. 2008. Song complexity correlates with learning ability in zebra finch males. Anim. Behav. 76, 1735–1741. ( 10.1016/j.anbehav.2008.08.009) [DOI] [Google Scholar]
- 10.Templeton CN, Laland KN, Boogert NJ. 2014. Does song complexity correlate with problem-solving performance in flocks of zebra finches? Anim. Behav. 92, 63–71. ( 10.1016/j.anbehav.2014.03.019) [DOI] [Google Scholar]
- 11.Catchpole C. 1996. Song and female choice: good genes and big brains? Trends Ecol. Evol. 11, 358–360. ( 10.1016/0169-5347(96)30042-6) [DOI] [PubMed] [Google Scholar]
- 12.Noad MJ, Cato DH, Bryden MM, Jenner MN, Jenner KC. 2000. Cultural revolution in whale songs. Nature 408, 537 ( 10.1038/35046199) [DOI] [PubMed] [Google Scholar]
- 13.Payne RS, McVay S. 1971. Songs of humpback whales. Science 173, 585–597. ( 10.1126/science.173.3997.585) [DOI] [PubMed] [Google Scholar]
- 14.Garland EC, Goldizen AW, Rekdahl ML, Constantine R, Garrigue C, Hauser ND, Poole MM, Robbins J, Noad MJ. 2011. Dynamic horizontal cultural transmission of humpback whale song at the ocean basin scale. Curr. Biol. 21, 687–691. ( 10.1016/j.cub.2011.03.019) [DOI] [PubMed] [Google Scholar]
- 15.Cholewiak DM, Sousa-Lima RS, Cerchio S. 2013. Humpback whale song hierarchical structure: historical context and discussion of current classification issues. Mar. Mammal Sci. 29, E312–EE32. ( 10.1111/mms.12005) [DOI] [Google Scholar]
- 16.Payne K, Payne R. 1985. Large scale changes over 19 years in songs of humpback whales in Bermuda. Ethology 68, 89–114. [Google Scholar]
- 17.Payne K, Tyack P, Payne R. 1983. Progressive changes in the songs of humpback whales (Megaptera novaeangliae): a detailed analysis of two seasons in Hawaii. AAAS selected symposium series, Westview Press, Boulder, CO: Commun. Behav. Whales 9–57. [Google Scholar]
- 18.Rekdahl M. 2012. Humpback whale vocal communication: use and stability of social calls and revolutions in the songs of east Australian whales. Doctoral dissertation, School of Veterinary Science, University of Queensland, Gatton, Queensland, Australia.
- 19.Paterson R, Paterson P. 1984. A study of the past and present status of humpback whales in east Australian waters. Biol. Conserv. 29, 321–343. ( 10.1016/0006-3207(84)90003-X) [DOI] [Google Scholar]
- 20.Allen JA, Garland EC, Murray A, Noad MJ, Dunlop R. 2017. Using self-organizing maps to classify humpback whale song units and quantify their similarity. J. Acoust. Soc. Am. 142, 1943–1952. ( 10.1121/1.4982040) [DOI] [PubMed] [Google Scholar]
- 21.Horton TW, Holdaway RN, Zerbini AN, Hauser N, Garrigue C, Andriolo A, Clapham PJ. 2011. Straight as an arrow: humpback whales swim constant course tracks during long-distance migration. Biol. Lett. 7, rsbl20110279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kavanagh AS, Noad MJ, Blomberg SP, Goldizen AW, Kniest E, Cato DH, Dunlop RA. 2017. Factors driving the variability in diving and movement behavior of migrating humpback whales (Megaptera novaeangliae): implications for anthropogenic disturbance studies. Mar. Mammal Sci. 33, 413–439. ( 10.1111/mms.12375) [DOI] [Google Scholar]
- 23.Noad MJ, Cato DH. 2007. Swimming speeds of singing and non-singing humpback whales during migration. Mar. Mammal Sci. 23, 481–495. ( 10.1111/j.1748-7692.2007.02414.x) [DOI] [Google Scholar]
- 24.Garland EC, Lilley MS, Goldizen AW, Rekdahl ML, Garrigue C, Noad MJ. 2012. Improved versions of the Levenshtein distance method for comparing sequence information in animals' vocalisations: tests using humpback whale song. Behaviour 149, 1413–1441. ( 10.1163/1568539X-00003032) [DOI] [Google Scholar]
- 25.Murray A, Dunlop RA, Noad MJ, Goldizen AW. 2018. Stereotypic and complex phrase types provide structural evidence for a multi-message display in humpback whales (Megaptera novaengliae). J. Acoust. Soc. Am. 143, 980–994. [DOI] [PubMed] [Google Scholar]
- 26.Garland EC, Rendell L, Lilley MS, Poole MM, Allen JA, Noad MJ. 2017. The devil is in the detail: quantifying vocal variation in a complex, multileveled, and rapidly evolving display. J. Acoust. Soc. Am. 142, 460–472. ( 10.1121/1.4991320) [DOI] [PubMed] [Google Scholar]
- 27.Kohonen T. 1985. Median strings. Pattern Recognit. Lett. 3, 309–313. ( 10.1016/0167-8655(85)90061-3) [DOI] [Google Scholar]
- 28.Noad MJ. 2002. The use of song by humpback whales (Megaptera novaeangliae) during migration off the east coast of Australia. Doctoral dissertation, University of Sydney, Sydney, Australia. [Google Scholar]
- 29.Garland EC, et al. 2013. Quantifying humpback whale song sequences to understand the dynamics of song exchange at the ocean basin scale. J. Acoust. Soc. Am. 133, 560–569. ( 10.1121/1.4770232) [DOI] [PubMed] [Google Scholar]
- 30.Shannon CE. 1948. A mathematical theory of communication. Bell Syst. Tech. J. 27, 379–423, 623–656. ( 10.1002/j.1538-7305.1948.tb01338.x) [DOI] [Google Scholar]
- 31.Kershenbaum A, et al. 2014. Acoustic sequences in non-human animals: a tutorial review and prospectus. Biol. Rev. 91, 13–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Suzuki R, Buck JR, Tyack PL. 2006. Information entropy of humpback whale songs. J. Acoust. Soc. Am. 119, 1849 ( 10.1121/1.2161827) [DOI] [PubMed] [Google Scholar]
- 33.Krebs JR. 1977. The significance of song repertoires: the Beau Geste hypothesis. Anim. Behav. 25, 475–478. ( 10.1016/0003-3472(77)90022-7) [DOI] [Google Scholar]
- 34.Noad MJ, Dunlop R, Paton D, Cato D. 2011. Absolute and relative abundance estimates of Australian east coast humpback whales (Megaptera novaeangliae). J. Cetacean Res. Manage. Special Issue 3, 243–252. [Google Scholar]
- 35.Noad MJ, Dunlop RA, Mack AK. 2017. Changes in humpback whale singing behavior with abundance: implications for the development of acoustic surveys of cetaceans. J. Acoust. Soc. Am. 142, 1611–1618. ( 10.1121/1.5001502) [DOI] [PubMed] [Google Scholar]
- 36.Miksis-Olds JL, Buck JR, Noad MJ, Cato DH, Dale Stokes M. 2008. Information theory analysis of Australian humpback whale song. J. Acoust. Soc. Am. 124, 2385–2393. ( 10.1121/1.2967863) [DOI] [PubMed] [Google Scholar]
- 37.Briefer E, Osiejuk TS, Rybak F, Aubin T. 2010. Are bird song complexity and song sharing shaped by habitat structure? An information theory and statistical approach. J. Theor. Biol. 262, 151–164. ( 10.1016/j.jtbi.2009.09.020) [DOI] [PubMed] [Google Scholar]
- 38.Petkov CI, Jarvis E. 2012. Birds, primates, and spoken language origins: behavioral phenotypes and neurobiological substrates. Front. Evolution. Neurosci. 4, 12 ( 10.3389/fnevo.2012.00012) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Murray A, Cerchio S, McCauley R, Jenner CS, Razafindrakoto Y, Coughran D, McKay S, Rosenbaum H. 2012. Minimal similarity in songs suggests limited exchange between humpback whales (Megaptera novaeangliae) in the southern Indian Ocean. Mar. Mammal Sci. 28, E41–E57. ( 10.1111/j.1748-7692.2011.00484.x) [DOI] [Google Scholar]
- 40.Garland EC, Gedamke J, Rekdahl ML, Noad MJ, Garrigue C, Gales N. 2013. Humpback whale song on the southern ocean feeding grounds: implications for cultural transmission. PLoS ONE 8, e79422 ( 10.1371/journal.pone.0079422) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Allen J. 2018. Fine-scale song exchange in two neighbouring populations of humpback whales. Doctoral dissertation, School of Veterinary Science, University of Queensland, Gatton, Queensland, Australia. [Google Scholar]
- 42.Patricelli GL, Hebets EA. 2016. New dimensions in animal communication: the case for complexity. Curr. Opin. Behav. Sci. 12, 80–89. ( 10.1016/j.cobeha.2016.09.011) [DOI] [Google Scholar]
- 43.Bohn KM, Schmidt-French B, Schwartz C, Smotherman M, Pollak GD. 2009. Versatility and stereotypy of free-tailed bat songs. PLoS ONE 4, e6746 ( 10.1371/journal.pone.0006746) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kershenbaum A, Ilany A, Blaustein L, Geffen E. 2012. Syntactic structure and geographical dialects in the songs of male rock hyraxes. Proc. R. Soc. B 279, 2974–2981. ( 10.1098/rspb.2012.0322) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Stephan C, Zuberbühler K. 2008. Predation increases acoustic complexity in primate alarm calls. Biol. Lett. 4, 641–644. ( 10.1098/rsbl.2008.0488) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Allen JA, Garland EC, Dunlop RA, Noad MJ. 2018. Data from: Cultural revolutions reduce complexity in the songs of humpback whales Dryad Digital Repository. ( 10.5061/dryad.69161bg) [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Citations
- Allen JA, Garland EC, Dunlop RA, Noad MJ. 2018. Data from: Cultural revolutions reduce complexity in the songs of humpback whales Dryad Digital Repository. ( 10.5061/dryad.69161bg) [DOI] [PMC free article] [PubMed]
Data Availability Statement
Data for this manuscript are available on Dryad Digital Repository at: http://dx.doi.org/10.5061/dryad.69161bg [46].