

Abstract
In recent years vitamin B6 has become a focus of research describing the compound’s critical function in cellular metabolism and stress response. For many years the sole function of vitamin B6 was considered to be that of an enzymatic cofactor. However, recently it became clear that it is also a potent antioxidant that effectively quenches reactive oxygen species and is thus of high importance for cellular well-being. In view of the recent findings, the current review takes a look back and summarizes the discovery of vitamin B6 and the elucidation of its structure and biosynthetic pathways. It provides a detailed overview on vitamin B6 both as a cofactor and a protective compound. Besides these general characteristics of the vitamin, the review also outlines the current literature on vitamin B6 derivatives and elaborates on recent findings that provide new insights into transport and catabolism of the compound and on its impact on human health.
Keywords: Pyridoxal phosphate, Vitamin B6, Oxidative stress, Derivatives, Transporter.
Introduction
The Discovery of Vitamin B6
The formula of vitamin B6 (henceforth referred to as vitB6) was first published by Ohdake in 1932. He worked on the isolation from rice-polishings of what he called “Oryzanin” (Vitamin B1) and found vitB6 as a by-product [1,2]. Ohdake described the formula, but he was not aware that he had found a vitamin and did not recognize its physiological importance.
At this time several scientists worked on the characterization of the vitamin B family members [3,4,5,6,7,8,9,10]. These scientists were searching for the so called “rat pellagra prevention factor” that could cure acrodyna, a pellagra-like skin disorder in rats. They discovered that by adding a special yeast eluate to the diet, acrodyna could be cured. Paul György, a Hungarian born scientist, first described vitB6 as the active “rat pellagra prevention factor” in the yeast eluate [3]. Several years later in 1938 five separate groups of researchers, including György, isolated the crystalline vitB6 from yeast [4,5,6,7,8]. After determination of the structure of vitB6 in 1939, György named the vitamin pyridoxine due to its structural homology to pyridine [9]. In the same year, Stanton A. Harris and Karl Folkers, accomplished the synthesis of vitB6 [10].
In further studies it was shown that vitB6 could exist in other chemical forms that differ from pyridoxine by a variable group present at the 4′ position [11]. Pyridoxine (PN) carries a hydroxyl, pyridoxal (PL) an aldehyde and pyridoxamine (PM) an amino group (Figure 1). While all three species can be phosphorylated, it is pyridoxal 5’-phosphate (PLP) that is the biologically most active form and used as cofactor for many important enzymatic reactions.
Figure 1.

Schematic drawing of de novo and salvage pathways.
The Function of VitB6
The discovery and the first publications on vitB6 ascribed a growth function to the vitamers [3,11]. However, further studies clarified that this relatively rough description understated the diversity of crucial functions and importance for living organism that vitB6 has. VitB6, in the form of PLP, plays a primary role acting as a cofactor for a large number of essential enzymes. These PLP-dependent enzymes catalyze more than 140 distinct enzymatic reactions and belong to five (oxidoreductases EC 1, transferases EC 2, hydrolases EC 3, lyases EC 4, isomerases EC 5) of the six enzyme classes defined by the Enzyme Nomenclature Committee of the International Union of Biochemistry and Molecular Biology (http://www.chem.qmul.ac.uk/iubmb/enzyme). This underlines the wide variety of chemical reactions that PLP-dependent enzymes promote in the organisms and shows again the importance of vitB6. The following section will give an overview of the metabolic reactions in which PLP-dependent enzymes are significantly involved.
Many of the PLP-dependent enzymes catalyze important steps in the amino acid metabolism, like co-catalyzing transamination, racemization, decarboxylation, and α,β-elimination reactions [12,13]. For example, transaminases mediate the conversion of α-ketoacids to amino acids and amino acid racemases produce D-amino acids from L-amino acids [14].
Another site of action for the PLP-dependent enzymes is fatty acid metabolism. The enzyme δ-6-desaturase (EC 1.14.19.3) catalyzes the synthesis of vital polyunsaturated fatty acids by the desaturation of linolic acid and γ-linolenic acid, respectively [15,16].
Besides these roles, PLP also represents an important cofactor for the degradation of storage carbohydrates, such as glycogen. The PLP-dependent glycogen phosphorylase (EC 2.4.1.1) mediates the glycogen breakdown by the release of glucose from glycogen [17].
Furthermore, two PLP-dependent enzymes are involved in hemoglobin formation and chlorophyll biosynthesis. In these reactions the rate-limiting step is the primary biosynthesis of δ-aminolevulinic acid. In mammals and birds δ-aminolevulinic acid is synthesized by the action of δ-aminolevulinic acid synthase (EC 2.3.1.37) and in plants and algae by the action of glutamate-1-semialdehyde 2,1-aminomutase (EC 5.4.3.8) [18].
Additionally, in plants the biosynthesis of the phytohormone ethylene is controlled by the synthesis of the precursor 1-aminocyclopropane-1-carboxylic acid from S-adenosylmethionine by PLP-dependent 1-aminocyclopropane-1-carboxylate synthases (EC 4.4.1.14) [19].
Apart from its function as a cofactor for PLP-dependent enzymes, vitB6 is also thought to act directly as a protective agent against reactive oxygen species, such as singlet oxygen which will be discussed in a following section [20,21].
While fungi, plants, archae, and most eubacteria are able to synthesize vitB6 (see next Section), most animals, including humans, lack this ability and rely on the external supply of vitB6.
The Known Pathways of VitB6 Anabolism
Deoxyxylose 5’-phosphate-Dependent and –Independent De Novo Biosynthesis of VitB6
Two existing pathways are known for de novo vitB6 biosynthesis. First, the deoxyxylose 5’-phosphate (DXP)-dependent pathway, which is present in eubacteria, such as Escherichia coli and second the DXP-independent pathway which is described for some bacteria, archaea and eukarya.
The DXP-dependent pathway has been intensively studied in the gram-negative bacterium E. coli [22]. It was shown that in E. coli, vitB6 is synthesized by the action of PdxJ (EC 2.6.99.2) and PdxA (EC 1.1.1.262) (Figure 1) [23,24,25]. These two vitB6 synthase proteins use 4-phospohydroxy-L-threonine (4HPT) and DXP, which are precursors in isoprenoid and thiamine biosynthesis, respectively, as substrates to form PNP [26,27,28]. PdxA catalyzes the oxidation of 4HPT to 3-amino-1-hydroxyacetone 1-phosphate (AHAP), and PdxJ forms PNP with the intermediates AHAP and DXP [29]. PNP is then oxidized to PLP, the biocatalytically active form of vitB6, by PdxH via the salvage pathway (Figure 1) [30]. The vitB6 precursors 4HPT and DXP originate on the one hand from the oxidation plus transamination of D-erythrose-4-phosphate, and on the other hand, by the synthesis from pyruvate and D-glyceraldehyde-3-phosphate by DXP synthase (EC 2.2.1.7) [31].
Analysis of the crystal structure of the participating enzymes showed that PdxA and PdxJ act separately. PdxA dimers create an interface onto which 4HPT binds [29]. In contrast PdxJ forms octamers as tetramers of PdxJ dimers [32]. In every dimer interface a pocket is located where the intermediates DXP and AHAP are converted to PNP.
The second known de novo vitB6 biosynthesis pathway is the DXP-independent pathway, which is found in bacteria, archaea, and eukarya [13,33,34]. The occurrence of this pathway is demonstrated in plants, fungi, Plasmodium falciparum, Thermotoga maritima as well as Bacillus subtilis and involves two proteins, PDX1 and PDX2 (for pyridoxine biosynthesis protein; orthologs for B. subtilis YaaD and YaaE, Geobacillus stearothermophilus PDXS and PDXT, Saccharomyces cerevisiae SNZ and SNO) [33,34,35,36,37]. These two synthase proteins directly synthesized PLP from ribose 5’-phosphate or ribulose 5’-phosphate, in combination with either glyceraldehyde 3’-phosphate or dihydroxyacetone phosphate and glutamine (Figure 1). Here PDX2 acts as a glutaminase, which deaminates glutamine to glutamate in order to supply nitrogen for the PLP heterocycle, and then PDX1 arranges the final ring closure [33,38].
Crystallization studies in the organisms B. subtilis, G. stearothermophilus and P. falciparum demonstrated that the PDX enzymes form a synthase complex with a cogwheel-like structure [37,39,40]. The core of the PLP synthase complex consists of 12 PDX1 enzymes that interact in two hexameric layers joining face-to-face to form a dodecamer onto which 12 PDX2 monomers attach [37,39,40].
The Salvage Pathway
In addition to the direct synthesis of new PNP or PLP, the vitamers are interconvertible via the so called salvage pathway. These conversions are accomplished by the action of either kinases or oxidases [22,41,42]. The salvage pathway has been best analyzed in E. coli, where it has been shown that two different kinases (EC 2.7.1.35) can phosphorylate PN, PL and PM to their respective 5’-phosphates (Figure 1). The two kinases differ in their substrate specifities, with PdxY acting on PL, whereas PdxK can utilize all three non-phosphorylated vitamers as substrates. Most eukaryotes contain a single kinase, and crystal structure of the kinase from several different organisms has revealed a dimer [43,44,45,46]. Each of the two monomers contains an active site that utilizes bound ATP and metal ions, which are required for activity [47].
Contrary to the kinases, only one oxidase, PdxH (EC 1.4.3.5), is shown in E. coli to oxidize the phosphorylated forms of PNP and PMP to PLP (Figure 1) [28,48].
In yeast a pyridoxine phosphate oxidase, PDX3 (EC 1.4.3.5), was identified and mutants in this gene had increased oxidative stress sensitivities [49]. Interestingly, complementation with a recently identified oxidase in Arabidopsis, AtPPOX, rescued this yeast mutant which underscores the highly conserved nature of the pathway [50]. The crystal structure of pyridoxine oxidase has shown that this protein also functions as a dimer, with the cofactor flavin mononucleotide, FMN, bound in the active site of each monomer [51,52,53].
The Other End: VitB6 Catabolism
Besides the biosynthesis of vitB6, catabolism of the vitamin is also an important aspect for cellular homeostasis of the compound. A critical step is represented by the dephosphorylation of PLP/PMP/PNP because this step represents a major control for the pool of available active vitB6 cofactor. There are reports of unspecific dephosphorylation of PLP/PMP/PNP by alkaline phosphatase (EC 3.1.3.1) and acid phosphatase (EC 3.1.3.2) [54,55,56,57,58]. However, additional phosphatases have been annotated for various organisms that specifically target phosphorylated vitB6 as a substrate [59,60]. Of these currently the one best characterized is human pyridoxal phosphatase (PLPP) (EC 3.1.3) (Figure 2). The enzyme is a 64 kDa dimer with a requirement for Mg2+. It is expressed in various tissues but predominantly in brain, liver and testis [60]. PLPP has its highest affinity for PLP, followed by PNP and then PMP. Inorganic phosphate has a strong inhibitory effect, but the enzyme can also be weakly inhibited by PL [60,61]. Although basic biochemical data are well established for human PLPP control of its activity is still open.
Figure 2.

Bacterial catabolic pathways of vitB6. Roman numbers in parenthesis indicate enzymes: (I) pyridoxine 4-oxidase; (II) pyridoxal phosphatase; (III) pyridoxal 4-dehydrogenase; (IV) 4-pyridoxollactonase; (V) pyridoxol oxidase. Arabic numbers indicate compound: (1) PN; (2) PLP; (3) PL; (4) 4-pyridoxolactone; (5) 4-pyridoxic acid; (6) succinic semialdehyde (modified from [62]).
In the gram-negative bacteria Pseudomonas sp. and Mesorhizobium loti a detailed pathway has been described for degradation of vitB6 and its phosphorylated derivatives (see Figure 2 for an overview) [62,63]. Here PL can either be converted in a two-step process from 4-pyridoxolactone to 4-pyridoxic acid (4-PA) via the subsequent activities of pyridoxal-4-dehydrogenase (EC 1.1.1.107) and 4-pyridoxolactonase (EC 3.1.1.27) or directly to 4-PA by pyridoxal oxidase (EC 1.2.3.8); 4-PA in turn is then catabolized in five additional steps to succinic semialdehyde [62,64]. It is noteworthy that 4-PA is also found in rats and human which makes it likely that similar catabolic pathways exist in these organisms [65,66]. In addition, succinic semialdehyde is a common compound that can accumulate in humans if, for example, degradation of γ-amino-butyric acid is impaired by succinic semialdehyde dehydrogenase (EC 1.2.1.24) deficiency [67]. However, it is basically unknown how the vitamin is degraded in eukaryotes after pyridoxine phosphatase activity.
Allocation of VitB6 in Prokaryotes and Eukaryotes
A highly important but poorly understood field is the translocation of vitB6 in prokaryotes and eukaryotes. Some vitB6 auxotrophic prokaryotes and single cell eukaryotes rely on the import of the vitamin, while multicellular eukaryotes that cannot synthesize vitB6 need to translocate the vitamin to their different organs.
Pioneering studies that demonstrated the existence of such vitB6 transporters came from Salmonella typhimurium and S. cerevisiae [68,69]. In higher eukaryotes studies on rat renal proximal tubular cells demonstrated uptake of pyridoxine and N-(4'-pyridoxyl) amines [70]. Here, the work by Zhang and McCormick showed that the tested rat cells have an active Na+-dependent uptake system, which is able to discriminate between the different offered substrates. Interestingly, both pyridoxine and its amine derivatives were substrates of a pyridoxal kinase as they were phosphorylated upon entry into the cytosol [70]. Intestinal absorption in humans comes from two sources, dietary ingestion in the small intestine and uptake of bacteria produced vitB6 in the large intestine [71,72]. Experiments using human intestinal epithelial Caco-2 cells and mammalian colonocytes demonstrated the involvement of carrier-mediated systems for both with pH, temperature and pyridoxine levels affecting the rate of transport. An intercellular protein kinase A (PKA)-mediated pathway appears to regulate transport in the Caco-2 cells, whereas transport in colonocytes appears to be regulated by a Ca2+/CaM- mediated pathway [71,72]. Work in human placental tissue has also indicated the presence of a passive transport system that transfers pyridoxine in and out of the cells but not against a concentration gradient [73]. Both active uptake and export of vitB6 have been demonstrated in yeast [69,74,75]. The fission yeast, Schizosaccharomyces pombe, showed a preferential efflux of PN that was dependent on the internal concentration of PN, and the rate was increased with the addition of Na+ [74]. It was suggested that the membrane potential could be affecting the PN efflux gate or carrier. Additionally, work in S. pombe has linked the PL reductase encoded by prl1+ gene to the excretion of PN following its reduction from PL as part of the maintenance of vitB6 levels in the cell [76]. Overall these findings demonstrated the presence of various mechanisms for transport of the vitB6. However the precise identity of the transport proteins that facilitate such movements has remained open. Described for S. cerevisiae in 2003, Tpn1p was the first example of a eukaryotic vitB6 transporter [77]. The protein belongs to the family of purine-cytosine permeases and functions as a plasma membrane bound proton symporter for vitB6 uptake. It has a high affinity for PN with a Km value of 0.55 μM but also transports PM and PL with lower affinities. A second unrelated transporter, Bsu1p was identified in S. pombe, a yeast strain that does not contain a Tpn1p homlog [78]. While Bsu1p has lower affinity for PN than Tpn1p, it also operates as a proton symporter with similar optimal pH and substrate profiles. Likewise, expression of both transporters increased when PN concentrations decreased [77,78]. Very recently, a novel class of vitamin transporters were identified in prokaryotes [79]. They are composed of different modules that have substrate specific components and an energy-coupling module and were named accordingly as energy-coupling factor (ECF) transporters. The energy-coupling module allows docking of different substrate specific components to facilitate selected transport of metabolites across membranes. Interestingly, the authors also identified an ECF transporter with a high affinity for pyridoxine [79]. The findings of Tpn1p, Bsu1p and the ECF transporters demonstrate that different transport pathways have evolved and it will be exciting to learn what kind of transport proteins are active in other organisms. In addition it will be of importance to understand intracellular transport of vitB6 since many PLP-dependent enzymes are present in mitochondria and chloroplast. However, currently it is open how this is achieved in the cell since corresponding transport proteins remain to be identified.
The Role of VitB6 in Stress Response
Recent work has provided evidence of an expanded role of vitB6 in cells. As stated above, this compound serves a role in enzymatic catalysis. However, vitB6 may play a crucial role in protecting cells from oxidative stress because the vitamin has been shown to exhibit antioxidant activity that even exceeds that of vitamins C and E [80,81,82,83]. Upon mutation of genes involved in the salvage and de novo pathways of vitB6 synthesis, a range of phenotypes are seen in salt and reactive oxygen species (ROS) sensitivity (Table 1). ROS sensitivity in context with vitB6 was originally characterized in the phytopathogen Cercospora nicotianae. Here mutant strains were identified that were particularly vulnerable to their own toxin cercosporin, a photosensitizer that produces singlet oxygen and superoxide upon irradiation [35,81]. Cloning of the mutant genes in C. nicotianae revealed that the mutated fungi were affected in a PDX1 ortholog [35,80,81]. These findings were of importance since so far vitB6 had not been mentioned in the context of singlet oxygen resistance [84]. Subsequent studies in other organisms also showed that vitB6 is crucial for oxidative stress tolerance and other abiotic stressors. For example, loss of Arabidopsis thaliana PDX1.3 causes hypersensitivity towards treatment with Rose Bengal, a ROS inducing chemical [85]. Moreover Arabidopsis pdx1.3 mutants are also hypersensitive towards salt and UV-B treatments [85,86]. It is also interesting to note that mutants in the salvage pathway show aberrant stress sensitivities, demonstrating that vitB6 metabolism in general is critical for abiotic stress tolerance. For example, the Arabidopsis sos4 mutant that is affected in pyridoxine kinase PDXK is highly sensitive to salt stress [87]. However, unlike mutants affected in PDX1 genes that have been demonstrated to contain reduced levels of vitB6, sos4 has overall increased levels of the vitamin [86,88,89,90]. Though the precise reason for increased vitB6 levels in sos4 remains to be answered, the example given demonstrates that vitB6 levels do not strictly correlate with stress tolerance. Hence, one must question whether aberrant vitB6 levels are the primary reason for the observed abiotic stress sensitivities.
Table 1.
Examples of VitB6 De Novo and Salvage Pathway Mutants in Context with Stress.
| Organism | Mutant | Pathway affected | Phenotype | Citation |
|---|---|---|---|---|
| E. coli | ppox/pdxH | Salvage | Reduced growth, aberrant shape | [27] |
| C. nicotianae | sor1/pdx1 | De novo | Increased ROS sensitivity, loss of vitB6 production, increased salt sensitivity, reduced growth | [81] |
| S. cerevisiae | snz1/pdx1 | De novo | Reduced growth in minimal media | [94] |
| S. cerevisiae | sno1/pdx2 | De novo | Reduced growth in minimal media | [94] |
| S. cerevisiae | pdx3 | Salvage | Increased ROS sensitivity | [50] |
| A. thaliana | sos4-1 | Salvage | Increased salt sensitivity | [87, 90] |
| A. thaliana | pdx 1.1, pdx 1.3 | De novo | Increased salt sensitivity | [21, 86] |
| A. thaliana | pdx3/PPOX | Salvage | Reduced aerial & root growth, increased salt sensitivity | [50, 90] |
It is noteworthy that PDX1 and PDXK gene expression is also regulated in response to abiotic stress. For example, B. subtilis PDX1 has been demonstrated to be upregulated in the presence of the singlet oxygen producer, methylene blue [91]. In addition, PYRO A (a PDX1 homologue described in Aspergillus nidulans) and Arabidopsis PDX1.3 are upregulated after exposure to UV radiation [92,93]. Also SNZ1, a S. cerevisiae PDX1 homologue, has been shown to be present in higher amounts during the stationary growth phase in which cultures are more prone to oxidative stress [94,95]. Findings in plants revealed that expression of Arabidopsis PDX1 genes is regulated by drought, chilling, UV-B treatment, and ozone [96]. Finally, SOS4 up-regulation has also been demonstrated as a response to cold stress and abscisic acid (ABA) treatment [87].
Overall there appears to be a broad and beneficial effect of vitB6 on abiotic stress tolerance in the cell, and stressors have been found to result in an increased amount of expression of genes involved in vitB6 biosynthesis. Notably and as mentioned above there are some exceptions to the observation that increased vitB6 availability is beneficial: Herrero and Daub observed negligible changes in vitB6 content in tobacco in response to salt stress, and Gonzalez and coworkers noted significantly higher vitB6 levels in sos4 relative to wild type plants despite the mutant’s increased salt sensitivity [90]. Considering these findings, future work may aim to link additional signals to a response of altered vitB6 production in cells. Additionally, characterization of the ROS quenching capacity and regulation of vitB6 biosynthesis may help to solve the close relationships seen between the vitamin and the described sources of stress.
The Diversity of VitB6 Derivatives
As described in the preceding paragraphs, vitB6 is a well-investigated compound critical for many cellular processes as either a central cofactor or as a potent antioxidant. However, it is noteworthy that a variety of different PN, PM, and PL derivatives have been described, for which the precise function is not understood (Table 2). These derivatives potentially have novel functions, and may be crucial to fully appreciate the biological relevance of vitB6.
Table 2.
Examples of VitB6 Vitamers and Their Derivatives
| Derivative | Structure | Function | Organism found | citation |
|---|---|---|---|---|
| Vitamin B6 |  |
Antioxidant | ubiquitous | [13] |
| Vitamin B6-phosphate | 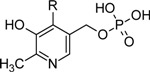 |
Cofactor | ubiquitous | [13] |
| 4’-O-Methyl-pyridoxine (ginkgotoxin) | 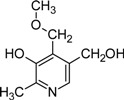 |
Unknown; potential inhibitor of PN/PM/PL kinase | Ginkgo biloba, Albizia tanganyicensis | [23, 97, 99] |
| 5’-O-Acetyl-4’-O-methylpyridoxine | 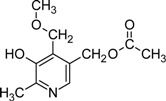 |
Unknown | Albizia tanganyicensis | [100] |
| Julibrine I | 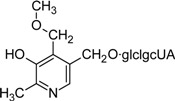 |
Unknown | Albizia julibrissin | [102] |
| Julibrine II | 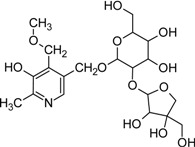 |
Unknown; has been demonstrated to induce arrhythmia | Albizia julibrissin | [102] |
| 5'-0-(β-D-Glucopyranosyl) pyridoxine | 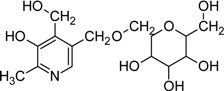 |
Unknown | Various plant products | [111, 112] |
| N-Hexanoyl-pyridoxamine (HAPM) | 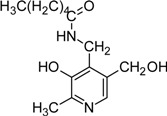 |
Inhibitor of advanced glycation and lipoxidation reactions | PM treated diabetic and obese rats | [9, 10] |
| N-Nonanedioyl-pyridoxamine monoamide (NDAPM) | 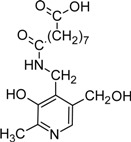 |
Inhibitor of advanced glycation and lipoxidation reactions | PM treated diabetic and obese rats | [9, 10] |
| N-Pentanedioyl-pyridoxamine monoamide (PDAPM) | 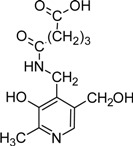 |
Inhibitor of advanced glycation and lipoxidation reactions | PM treated diabetic and obese rats | [104, 105] |
| N-Formyl-pyridoxamine (FAPM) | 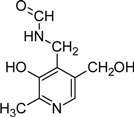 |
Inhibitor of advanced glycation and lipoxidation reactions | PM treated diabetic and obese rats | [104, 105] |
The best known of these derivatives is probably 4’-O-methylpyridoxine or ginkgotoxin from the tree Ginkgo biloba [12,97]. The compound has been found in different tissues with the highest concentrations being present in seeds [98]. Although it has been shown that the additional 4'-O-methyl group most likely derives from methionine, and that both phosphorylated and non-phosphorylated forms of pyridoxine are methylated, the biosynthetic pathway leading to 4’-O-methylpyridoxine is still unresolved [98,99]. Ingestion of the toxin can lead to Gin-nan-sitotoxism, epileptic convulsions, and other neuronal disorders [100]. As seeds from Ginkgo trees are a food source in China and Japan, and extracts from leaves are used in pharmaceutical products, they represent a potential health risk. The PLP-dependent enzyme glutamate decarboxylase (GAD), which is critical for synthesis of the neurotransmitter GABA was discussed as a potential target of 4’-O-methylpyridoxine. However, there is no clear evidence that ginkgotoxin significantly reduces GAD activity when present in physiologically relevant concentrations [101]. In contrast, recent work rather suggests that the toxin is competing with PN/PM/PL for human pyridoxine kinase [97]. This in turn might reduce the pool of available PLP and PMP in the brain and negatively affect GAD activity and GABA biosynthesis [97].
Ginkgotoxin was also found in the African tree Albizia tanganyicensis [98,99,100] demonstrating that the biosynthetic pathway leading to the formation of 4’-O-methylpyridoxine is not unique to Ginkgo. Albizia tanganyicensis and its close relative Albizia julibrissin also synthesize other more complex vitB6 derivatives (see Table 2) [100,102]. Unfortunately, neither for ginkgotoxin nor for the other Albizia derivatives could we find a biological function explaining why these compounds are synthesized. A likely possibility is that they serve as protecting compounds against pathogens due to their toxicity. This poses the attractive question as to what kind of mechanisms these plants employ to protect their own metabolism against toxic vitB6 derivatives. For example, do they utilize specific compartments or organelles for storage of their toxic compounds?
Another aspect that vitB6 derivatives have been brought in context with is the formation of advanced glycation and lipoxygenation end-products (AGE and ALE, respectively). AGE and ALE formation can occur in cells when reduced sugars (e.g. glucose, fructose) or polyunsaturated fatty acids are abundant. In such a situation they can cross-react preferentially with lysine residues of proteins [103,104]. Accumulation of AGE and ALE is also caused by oxidative stress or overload of pathways active in detoxification [103]. Such end-products are often detrimental for protein function and, especially in older tissues, might lead to severe damage. Hence, patients with diabetes or atherosclerosis that have increased contents of blood sugar or blood lipids, respectively, suffer from accumulation of AGEs and ALEs. Here, pyridoxamine is discussed to serve as a protecting compound by bonding with fatty acids (see Table 2) and thereby effectively competing with proteins for ALE formation [104,105]. The vitamin is also discussed to serve as a protecting compound for AGE formation which is of special interest to patients suffering from diabetes [106,107,108].
A significant proportion of vitB6 (ranging from 5-80% of the total vitB6 content) in many fruits and vegetables is glycosylated [109,110]. Glycosylated vitB6 appears to be abundant in plants and has been detected in soybean, rice, and Ginkgo [111,112,113]. Furthermore, in fungi β-fructosyl and β-galactosyl compounds of pyridoxine have been found [114,115]. It is likely that these derivatives of vitB6 are not substrates of a pyridoxine kinase and thus are not accessible for metabolic utilization. Consequently, specific β-glucosidases have been described in plants and human capable of removing the sugar moiety, making the vitamin again accessible for salvage pathway enzymes [111,116,117,118]. Although no precise explanation for the high amount of glycosylated vitB6 is provided in literature, a possibility can be seen in context with AGE accumulation. Here, vitB6 might serve as a protecting compound to prevent reaction of sugar with lysine residues of proteins. Alternatively, glycosylated forms of vitB6 might serve as storage compounds of the vitamin and even carbohydrates that can be mobilized upon demand. Overall the existence of such a diverse variety of vitB6 derivatives indicates that the vitamin is involved in or employed for many other currently unknown processes.
VitB6 Supplements and Human Health: Is it too much or not enough of a good thing?
The importance of vitB6 in human health is reflected in how actively it is studied in relationship to a wide range of disease prevention and control. The current Recommended Dietary Allowance (RDS) from the American National Institute of Health (NIH) for vitB6 is 2 milligrams per day with an upward tolerance of 100 mg per day for adults. High doses can lead to peripheral sensory neuropathy and nerve degeneration [119,120]. These problems are generally reversible when supplementation is stopped. Additionally some studies have suggested that increased levels of the B6 vitamers and some derivatives can generate toxic photoproducts as a result of UV irradiation [121,122,123].
Most problems are stemming from deficiencies of the vitamin and there are several areas of interest actively being pursued. Numerous clinical trials have been conducted to observe the broader impact of vitB6 on human health problems such as cancer prevention and recovery and the amelioration of neurological diseases. Also under investigation are the benefits of increased vitB6 through its role as a cofactor in upstream processes that lead to problems involving heart disease, osteoporosis and diabetes. Several of these topics are addressed in the following section.
Studies of vitB6 supplements in cancer prevention have yielded mixed results ranging from no significant benefits seen in breast cancer to indications of protection against colorectal cancer [124]. A recent study on hairless mice given PN supplements was performed to see if increased vitB6 protected against UV-B induced skin tumors [121]. It was reasoned that vitB6 would help quench reactive oxygen species (ROS), which are associated with the development of cancer. Interestingly though, while higher serum levels of PLP correlated with a higher dosage of dietary PN, neither the amount of PLP nor oxidative stress markers in skin differed in relationship to the dosages. Additionally an increase in tumor induction was seen in the mice given the higher doses of PN, correlating with previous work [122,123]. Interestingly decreased levels of vitB6 accompanied by increased levels of oxidative stress were detected in red blood cells of non-small cell lung cancer patients, also highlighting the vitamin’s potential benefits as an antioxidant [125].
Phosphorylated vitB6 is needed as a cofactor for neurotransmitter synthesis. However, studies on elderly people have shown that a walking program has more benefits on cognitive improvement and increased folate may help prevent Alzheimer’s diseases rather than vitB6 supplements [126,127,128,129]. While an association of low PLP and high symptoms of depression have been reported, supplementation with vitB6 has not been shown to conclusively improve depression in older men [130,131]. In schizophrenic patients with tardive dyskinsia, plasma levels of PLP were significantly lower and treatment with vitB6 supplements reduced the symptoms of this disease along with another schizophrenic associated disease, akathisia [132,133,134]. VitB6 is also studied as a potentially important candidate to improve behavioural disorders of autistic children, although the precise impact of the vitamin remains to be shown. [135,136].
VitB6 is involved in maintenance of normal homocysteine levels, and lower levels of homocysteine are associated with lower rates of coronary heart disease and stroke [137]. However studies are conflicting as to whether giving supplements to lower the homocysteine levels improves protection against these diseases [137,138,139]. High homocysteine levels have also been linked to osteoperosis and bone fragility fractures. In vitro tissue experiments demonstrated that either decreasing vitB6 or increasing homocysteine levels stimulated osteoclast activity, which leads to bone resorption [140].
Blood vitB6 levels are significantly decreased in diabetics. As vitB6 is a cofactor in tryptophan catabolism, disruption of this pathway leads to increased levels of kynurenine metabolites which inhibit insulin secretion and lower glucose tolerance [141]. Studies on different B6 vitamers have shown that supplements can help with problems related to glucose tolerance [142]. Complications from other diabetes-associated diseases are also common, and interestingly high doses of vitB6 normalized endothelial dysfunction, a precursor to vascular disease, in children with type 1 diabetes [143].
Outlook and Perspectives
Though in the last years various aspects in vitB6 biosynthesis and the impact and benefits of the vitamin for general metabolism have been explained, many questions remain unanswered. Five of these are covered in the following section to provide an outlook on future opportunities related to this important research field.
Do regulatory mechanisms exist that control PLP de novo biosynthesis? Although the two biosynthetic pathways – DXP-dependent and –independent – have been resolved, regulatory mechanisms on the transcriptional and posttranscriptional level to control PLP biosynthesis remain to be found. Although transcriptional regulation of PDX1 genes has been described for various organisms after stress treatments, it remains open whether this in turn affects vitB6 levels [13,144]. Considering the central and indispensable role of PLP in metabolism one would expect that such regulatory switches exist. These need to be connected with general metabolism, first because vitB6 exhibits such a central role as a cofactor and second because the de novo biosynthetic machinery competes for the precursors required for PLP synthesis with other pathways.
How are the salvage pathway and PLP phosphatase activities regulated? As for the de novo pathways, it has not been explained how salvage pathway enzymes and PLP phosphatases are regulated. This is surprising as pyridoxine kinase and PNP/PMP oxidase are crucial players in controlling vitB6 homeostasis and the availability of the active cofactor which might even occur in concert with PLP phosphatases. However, only a few studies reveal insights on factors like ions, ATP, or end-products that directly affect activities of these enzymes [61,145,146,147,148,149], while the interplay between the different proteins and the de novo pathway has not been addressed. Again, one would suggest that some higher degree of active control is present that regulates the proteins of the salvage pathway and PLP phosphatases upon demand.
What are the mechanisms of vitamin B6 translocation? For many organisms it is open how vitB6 is translocated within the different organs and tissues. Although the non-phosphorylated forms can to some extent go passively through membranes, this diffusion is most likely insufficient for long distance allocation or rapid uptake of the vitamin when needed [77]. Currently only a few examples are given for vitB6 transporters in yeast and prokaryotes, making this an important question in other organisms like animals or plants [77,79].
How does vitamin B6 metabolism positively affect stress tolerance? For many organisms it has been shown that mutants affected in either the salvage or the de novo pathway are hypersensitive towards abiotic stress conditions. However, this hypersensitivity does not always correlate with the vitB6 content in the cell [86,90]. Hence it is currently unclear and necessary to ask whether the amount of vitB6 is the critical factor to protect against abiotic stress, whether it is the vitB6 homeostasis that is important, or whether it is the proteins that participate in the different pathways that have additional functions connected to stress alleviation.
Are there more de novo pathways present in yet unexplored organisms and how are vitB6 derivatives formed? Currently it appears to be that there are just the two described de novo pathways, and no evidence for additional biosynthetic pathways is at hand. Though one cannot exclude the possibility for a third pathway, it is more likely that only the two described DXP-dependent and DXP-independent pathways exist. In contrast the variety of existing vitB6 derivatives indicates an extensive metabolic ability of organisms to modify vitB6. It will be interesting in the future to have more knowledge generated on the enzymes that modify vitB6, about the biological purposes of these compounds, and how these organisms protect themselves against potentially toxic derivatives. Understanding these points may provide better approaches to utilizing the pharmaceutical potentials of vitB6 and its derivatives for human health.
Acknowledgments
We would like to thank our Russian collaborator Dr. Cleatus for critical reading. We also would like to thank the DFG (DFG grant HE3224/7-1 to HH) for supporting this work.
Footnotes
Sample Availability: Not available.
References and Notes
- 1.Ohdake S. Isolation of "Oryzanin" (Antineuritic Vitamin) from Rice-polishings. Bull. Agri. Chem. Soc. Japan. 1932;8:11–46. doi: 10.1271/bbb1924.8.11. [DOI] [Google Scholar]
- 2.Wiardy P.W. Crystalline Vitamin B6 (Adermin) Nature. 1938;142:1158–1158. doi: 10.1038/1421158b0. [DOI] [Google Scholar]
- 3.György P. Vitamin B2 and the Pellagra-like Dermatitis in Rats. Nature. 1934;133:498–499. doi: 10.1038/133498a0. [DOI] [Google Scholar]
- 4.György P. Crystalline vitamin B6. J. Am. Chem. Soc. 1938;60:983–984. doi: 10.1021/ja01271a505. [DOI] [Google Scholar]
- 5.Ichiba A., Michi K. Cristalline vitamin B6. Sci. Pap. Inst. Phys. Chem. Res. 1938;34:623–626. (Tokyo) [Google Scholar]
- 6.Keresztesy J.C., Stevens J.R. Vitamin B-6. Proc. Soc. Exp. Biol. Med. 1938;38:64–65. doi: 10.3181/00379727-38-9740P. [DOI] [Google Scholar]
- 7.Kuhn R., Wendt G. Über das antidermatitische Vitamin der Hefe. Ber. Chem. Ges. 1938;71:780–782. [Google Scholar]
- 8.Lepkovsky Crystalline factor I. Science. 1938;87:169–170. doi: 10.1126/science.87.2251.169. [DOI] [PubMed] [Google Scholar]
- 9.György P., Eckardt R.E. Vitamin B6 and Skin Lesions in Rats. Nature. 1939;144:512–512. [Google Scholar]
- 10.Harris S.A., Folkers K. Synthetic vitamin B6. Science. 1939;89:347. doi: 10.1126/science.89.2311.347. [DOI] [PubMed] [Google Scholar]
- 11.Snell E.E., Guirard B.M., Williams R.J. Occurrence in natural products of a physiologically active metabolite of pyridoxine. J. Biol. Chem. 1942;143:519–530. [Google Scholar]
- 12.Drewke C., Leistner E. Biosynthesis of vitamin B6 and structurally related derivatives. Vitam. Horm. 2001;61:121–155. doi: 10.1016/s0083-6729(01)61004-5. [DOI] [PubMed] [Google Scholar]
- 13.Mittenhuber G. Phylogenetic analyses and comparative genomics of vitamin B6 (pyridoxine) and pyridoxal phosphate biosynthesis pathways. J. Mol. Microbiol.Biotechnol. 2001;3:1–20. [PubMed] [Google Scholar]
- 14.Grogan D.W. Temperature-sensitive murein synthesis in an Escherichia coli pdx mutant and the role of alanine racemase. Arc. Microbiol. 1988;150:363–367. doi: 10.1007/BF00408308. [DOI] [PubMed] [Google Scholar]
- 15.Nakamura M.T., Nara T.Y. Structure, function, and dietary regulation of delta-6, delta-5, and delta-9 desaturases. Annu. Rev. Nutr. 2004;24:345–376. doi: 10.1146/annurev.nutr.24.121803.063211. [DOI] [PubMed] [Google Scholar]
- 16.Horrobin D.F. Fatty acid metabolism in health and disease: the role of delta-6-desaturase. Am. J. Clin. Nutr. 1993;57:732S-736S; discussion 736S-737S. doi: 10.1093/ajcn/57.5.732S. [DOI] [PubMed] [Google Scholar]
- 17.Helmreich E.J. How pyridoxal 5'-phosphate could function in glycogen phosphorylase catalysis. Biofactors. 1992;3:159–172. [PubMed] [Google Scholar]
- 18.Cheltsov A.V., Guida W.C., Ferreira G.C. Circular permutation of 5-aminolevulinate synthase: effect on folding, conformational stability, and structure. J. Biol. Chem. 2003;278:27945–27955. doi: 10.1074/jbc.M207011200. [DOI] [PubMed] [Google Scholar]
- 19.Rottmann W.H., Peter G.F., Oeller P.W., Keller J.A., Shen N.F., Nagy B.P., Taylor L.P., Campbell A.D., Theologis A. 1-aminocyclopropane-1-carboxylate synthase in tomato is encoded by a multigene family whose transcription is induced during fruit and floral senescence. J. Mol. Biol. 1991;222:937–961. doi: 10.1016/0022-2836(91)90587-V. [DOI] [PubMed] [Google Scholar]
- 20.Bilski P., Li M.Y., Ehrenshaft M., Daub M.E., Chignell C.F. Vitamin B6 (Pyridoxine) and Its Derivatives Are Efficient Singlet Oxygen Quenchers and Potential Fungal Antioxidants. Photochem. Photobiol. 2000;71:129–134. doi: 10.1562/0031-8655(2000)071<0129:SIPVBP>2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 21.Chen H., Xiong L. Pyridoxine is required for post-embryonic root development and tolerance to osmotic and oxidative stresses. Plant J. 2005;44:396–408. doi: 10.1111/j.1365-313X.2005.02538.x. [DOI] [PubMed] [Google Scholar]
- 22.Hill R.E., Himmeldirk K., Kennedy I.A., Pauloski R.M., Sayer B.G., Wolf E., Spenser I.D. The biogenetic anatomy of vitamin B6. A 13C NMR investigation of the biosynthesis of pyridoxol in Escherichia coli. J. Biol. Chem. 1996;271:30426–30435. doi: 10.1074/jbc.271.48.30426. [DOI] [PubMed] [Google Scholar]
- 23.Arenz A., Klein M., Fiehe K., Gross J., Drewke C., Hemscheidt T., Leistner E. Occurrence of neurotoxic 4'-O-methylpyridoxine in Ginkgo biloba leaves, Ginkgo medications and Japanese Ginkgo food. Planta Med. 1996;62:548–551. doi: 10.1055/s-2006-957967. [DOI] [PubMed] [Google Scholar]
- 24.Drewke C., Notheis C., Hansen U., Leistner E., Hemscheidt T., Hill R.E., Spenser I.D. Growth response to 4-hydroxy-L-threonine of Escherichia coli mutants blocked in vitamin B6 biosynthesis. FEBS Lett. 1993;318:125–128. doi: 10.1016/0014-5793(93)80005-F. [DOI] [PubMed] [Google Scholar]
- 25.Laber B., Maurer W., Scharf S., Stepusin K., Schmidt F.S. Vitamin B6 biosynthesis: formation of pyridoxine 5'-phosphate from 4-(phosphohydroxy)-L-threonine and 1-deoxy-D-xylulose-5-phosphate by PdxA and PdxJ protein. FEBS Lett. 1999;449:45–48. doi: 10.1016/S0014-5793(99)00393-2. [DOI] [PubMed] [Google Scholar]
- 26.Cane D.E., Hsiung Y., Cornish J.A., Robinson J.K., Spenser I.D. Biosynthesis of Vitamin B6: The Oxidation of 4-(Phosphohydroxy)-l-threonine by PdxA. J. Am. Chem. Soc. 1998;120:1936–1937. doi: 10.1021/ja9742085. [DOI] [Google Scholar]
- 27.Lam H.M., Winkler M.E. Characterization of the complex pdxH-tyrS operon of Escherichia coli K-12 and pleiotropic phenotypes caused by pdxH insertion mutations. J. Bacteriol. 1992;174:6033–6045. doi: 10.1128/jb.174.19.6033-6045.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zhao G., Winkler M.E. 4-Phospho-hydroxy-L-threonine is an obligatory intermediate in pyridoxal 5'-phosphate coenzyme biosynthesis in Escherichia coli K-12. FEMS Microbiol. Lett. 1996;135:275–280. doi: 10.1111/j.1574-6968.1996.tb08001.x. [DOI] [PubMed] [Google Scholar]
- 29.Sivaraman J., Li Y., Banks J., Cane D.E., Matte A., Cygler M. Crystal structure of Escherichia coli PdxA, an enzyme involved in the pyridoxal phosphate biosynthesis pathway. J. Biol. Chem. 2003;278:43682–43690. doi: 10.1074/jbc.M306344200. [DOI] [PubMed] [Google Scholar]
- 30.Notheis C., Drewke C., Leistner E. Purification and characterization of the pyridoxol-5'-phosphate: oxygen oxidoreductase (deaminating) from Escherichia coli. Biochim. Biophys. Acta. 1995;1247:265–271. doi: 10.1016/0167-4838(94)00235-9. [DOI] [PubMed] [Google Scholar]
- 31.Sprenger G.A., Schorken U., Wiegert T., Grolle S., de Graaf A.A., Taylor S.V., Begley T.P., Bringer-Meyer S., Sahm H. Identification of a thiamin-dependent synthase in Escherichia coli required for the formation of the 1-deoxy-D-xylulose 5-phosphate precursor to isoprenoids, thiamin, and pyridoxol. Proc. Natl. Acad. Sci. USA. 1997;94:12857–12862. doi: 10.1073/pnas.94.24.12857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Franco M.G., Laber B., Huber R., Clausen T. Structural basis for the function of pyridoxine 5'-phosphate synthase. Structure. 2001;9:245–253. doi: 10.1016/S0969-2126(01)00584-6. [DOI] [PubMed] [Google Scholar]
- 33.Burns K.E., Xiang Y., Kinsland C.L., McLafferty F.W., Begley T.P. Reconstitution and biochemical characterization of a new pyridoxal-5'-phosphate biosynthetic pathway. J. Am. Chem. Soc. 2005;127:3682–3683. doi: 10.1021/ja042792t. [DOI] [PubMed] [Google Scholar]
- 34.Tambasco-Studart M., Titiz O., Raschle T., Forster G., Amrhein N., Fitzpatrick T.B. Vitamin B6 biosynthesis in higher plants. Proc. Natl. Acad. Sci. USA. 2005;102:13687–13692. doi: 10.1073/pnas.0506228102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ehrenshaft M., Bilski P., Li M.Y., Chignell C.F., Daub M.E. A highly conserved sequence is a novel gene involved in de novo vitamin B6 biosynthesis. Proc. Natl. Acad. Sci. USA. 1999;96:9374–9378. doi: 10.1073/pnas.96.16.9374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Gengenbacher M., Fitzpatrick T.B., Raschle T., Flicker K., Sinning I., Muller S., Macheroux P., Tews I., Kappes B. Vitamin B6 biosynthesis by the malaria parasite Plasmodium falciparum: biochemical and structural insights. J. Biol. Chem. 2006;281:3633–3641. doi: 10.1074/jbc.M508696200. [DOI] [PubMed] [Google Scholar]
- 37.Zhu J., Burgner J.W., Harms E., Belitsky B.R., Smith J.L. A new arrangement of (beta/alpha)8 barrels in the synthase subunit of PLP synthase. J. Biol. Chem. 2005;280:27914–27923. doi: 10.1074/jbc.M503642200. [DOI] [PubMed] [Google Scholar]
- 38.Tambasco-Studart M., Tews I., Amrhein N., Fitzpatrick T.B. Functional analysis of PDX2 from Arabidopsis, a glutaminase involved in vitamin B6 biosynthesis. Plant Physiol. 2007;144:915–925. doi: 10.1104/pp.107.096784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Zein F., Zhang Y., Kang Y.N., Burns K., Begley T.P., Ealick S.E. Structural insights into the mechanism of the PLP synthase holoenzyme from Thermotoga maritima. Biochemistry. 2006;45:14609–14620. doi: 10.1021/bi061464y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Flicker K., Neuwirth M., Strohmeier M., Kappes B., Tews I., Macheroux P. Structural and thermodynamic insights into the assembly of the heteromeric pyridoxal phosphate synthase from Plasmodium falciparum. J. Mol. Biol. 2007;374:732–748. doi: 10.1016/j.jmb.2007.09.038. [DOI] [PubMed] [Google Scholar]
- 41.Yang Y., Tsui H.C., Man T.K., Winkler M.E. Identification and function of the pdxY gene, which encodes a novel pyridoxal kinase involved in the salvage pathway of pyridoxal 5'-phosphate biosynthesis in Escherichia coli K-12. J. Bacteriol. 1998;180:1814–1821. doi: 10.1128/jb.180.7.1814-1821.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yang Y., Zhao G., Winkler M.E. Identification of the pdxK gene that encodes pyridoxine (vitamin B6) kinase in Escherichia coli K-12. FEMS Microbiol. Lett. 1996;141:89–95. doi: 10.1111/j.1574-6968.1996.tb08368.x. [DOI] [PubMed] [Google Scholar]
- 43.Cao P., Gong Y., Tang L., Leung Y.C., Jiang T. Crystal structure of human pyridoxal kinase. J. Struct. Biol. 2006;154:327–332. doi: 10.1016/j.jsb.2006.02.008. [DOI] [PubMed] [Google Scholar]
- 44.Li M.H., Kwok F., Chang W.R., Lau C.K., Zhang J.P., Lo S.C., Jiang T., Liang D.C. Crystal structure of brain pyridoxal kinase, a novel member of the ribokinase superfamily. J. Biol. Chem. 2002;277:46385–46390. doi: 10.1074/jbc.M208600200. [DOI] [PubMed] [Google Scholar]
- 45.Safo M.K., Musayev F.N., di Salvo M.L., Hunt S., Claude J.B., Schirch V. Crystal structure of pyridoxal kinase from the Escherichia coli pdxK gene: implications for the classification of pyridoxal kinases. J. Bacteriol. 2006;188:4542–4552. doi: 10.1128/JB.00122-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Safo M.K., Musayev F.N., Hunt S., di Salvo M.L., Scarsdale N., Schirch V. Crystal structure of the PdxY Protein from Escherichia coli. J. Bacteriol. 2004;186:8074–8082. doi: 10.1128/JB.186.23.8074-8082.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Musayev F.N., di Salvo M.L., Ko T.P., Gandhi A.K., Goswami A., Schirch V., Safo M.K. Crystal Structure of human pyridoxal kinase: structural basis of M(+) and M(2+) activation. Protein Sci. 2007;16:2184–2194. doi: 10.1110/ps.073022107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zhao G., Winkler M.E. Kinetic limitation and cellular amount of pyridoxine (pyridoxamine) 5'-phosphate oxidase of Escherichia coli K-12. J. Bacteriol. 1995;177:883–891. doi: 10.1128/jb.177.4.883-891.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Loubbardi A., Marcireau C., Karst F., Guilloton M. Sterol uptake induced by an impairment of pyridoxal phosphate synthesis in Saccharomyces cerevisiae: cloning and sequencing of the PDX3 gene encoding pyridoxine (pyridoxamine) phosphate oxidase. J. Bacteriol. 1995;177:1817–1823. doi: 10.1128/jb.177.7.1817-1823.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sang Y., Barbosa J.M., Wu H., Locy R.D., Singh N.K. Identification of a pyridoxine (pyridoxamine) 5'-phosphate oxidase from Arabidopsis thaliana. FEBS Lett. 2007;581:344–348. doi: 10.1016/j.febslet.2006.12.028. [DOI] [PubMed] [Google Scholar]
- 51.Pedelacq J.D., Rho B.S., Kim C.Y., Waldo G.S., Lekin T.P., Segelke B.W., Rupp B., Hung L.W., Kim S.I., Terwilliger T.C. Crystal structure of a putative pyridoxine 5'-phosphate oxidase (Rv2607) from Mycobacterium tuberculosis. Proteins. 2006;62:563–569. doi: 10.1002/prot.20824. [DOI] [PubMed] [Google Scholar]
- 52.Biswal B.K., Au K., Cherney M.M., Garen C., James M.N. The molecular structure of Rv2074, a probable pyridoxine 5'-phosphate oxidase from Mycobacterium tuberculosis, at 1.6 angstroms resolution. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2006;62:735–742. doi: 10.1107/S1744309106025012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.di Salvo M.L., Safo M.K., Musayev F.N., Bossa F., Schirch V. Structure and mechanism of Escherichia coli pyridoxine 5'-phosphate oxidase. Biochim. Biophys. Acta. 2003;1647:76–82. doi: 10.1016/s1570-9639(03)00060-8. [DOI] [PubMed] [Google Scholar]
- 54.Fonda M.L., Trauss C., Guempel U.M. The binding of pyridoxal 5'-phosphate to human serum albumin. Arch. Biochem. Biophys. 1991;288:79–86. doi: 10.1016/0003-9861(91)90167-H. [DOI] [PubMed] [Google Scholar]
- 55.Li T.K., Lumeng L., Veitch R.L. Regulation of pyridoxal 5'-phosphate metabolism in liver. Biochem. Biophys. Res. Commun. 1974;61:677–684. doi: 10.1016/0006-291X(74)91010-9. [DOI] [PubMed] [Google Scholar]
- 56.Lumeng L., Brashear R.E., Li T.K. Pyridoxal 5'-phosphate in plasma: source, protein-binding, and cellular transport. J. Lab. Clin. Med. 1974;84:334–343. [PubMed] [Google Scholar]
- 57.Harris H. The human alkaline phosphatases: what we know and what we don't know. Clin. Chim. Acta. 1990;186:133–150. doi: 10.1016/0009-8981(90)90031-M. [DOI] [PubMed] [Google Scholar]
- 58.Bull H., Murray P.G., Thomas D., Fraser A.M., Nelson P.N. Acid phosphatases. Mol. Pathol. 2002;55:65–72. doi: 10.1136/mp.55.2.65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Lee Y.P., Kim D.W., Lee M.J., Jeong M.S., Kim S.Y., Lee S.H., Jang S.H., Park J., Kang T.C., Won M.H., Cho S.W., Kwon O.S., Eum W.S., Choi S.Y. Human brain pyridoxal-5'-phosphate phosphatase (PLPP):protein transduction of PEP-1-PLPP into PC12 cells. BMB Rep. 2008;41:408–413. doi: 10.5483/BMBRep.2008.41.5.408. [DOI] [PubMed] [Google Scholar]
- 60.Jang Y.M., Kim D.W., Kang T.C., Won M.H., Baek N.I., Moon B.J., Choi S.Y., Kwon O.S. Human pyridoxal phosphatase. Molecular cloning, functional expression, and tissue distribution. J. Biol. Chem. 2003;278:50040–50046. doi: 10.1074/jbc.M309619200. [DOI] [PubMed] [Google Scholar]
- 61.Fonda M.L. Purification and characterization of vitamin B6-phosphate phosphatase from human erythrocytes. J. Biol. Chem. 1992;267:15978–15983. [PubMed] [Google Scholar]
- 62.Mukherjee T., Kinsland C., Begley T.P. PLP catabolism: identification of the 4-pyridoxic acid dehydrogenase gene in Mesorhizobium loti MAFF303099. Bioorg. Chem. 2007;35:458–464. doi: 10.1016/j.bioorg.2007.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Yagi T., Kishore G.M., Snell E.E. The bacterial oxidation of vitamin B6. 4-Pyridoxic acid dehydrogenase: a membrane-bound enzyme from Pseudomonas MA-1. J. Biol. Chem. 1983;258:9419–9425. [PubMed] [Google Scholar]
- 64.Burg R.W., Rodwell V.W., Snell E.E. Bacterial oxidation of vitamin B6. II. Metabolites of pyridoxamine. J. Biol. Chem. 1960;235:1164–1169. [PubMed] [Google Scholar]
- 65.Fukuwatari T., Wada H., Shibata K. Age-related alterations of B-group vitamin contents in urine, blood and liver from rats. J. Nutr. Sci. Vitaminol. (Tokyo) 2008;54:357–362. doi: 10.3177/jnsv.54.357. [DOI] [PubMed] [Google Scholar]
- 66.Rybak M.E., Pfeiffer C.M. Clinical analysis of vitamin B(6): determination of pyridoxal 5'-phosphate and 4-pyridoxic acid in human serum by reversed-phase high-performance liquid chromatography with chlorite postcolumn derivatization. Anal. Biochem. 2004;333:336–344. doi: 10.1016/j.ab.2004.06.036. [DOI] [PubMed] [Google Scholar]
- 67.Pearl P.L., Novotny E.J., Acosta M.T., Jakobs C., Gibson K.M. Succinic semialdehyde dehydrogenase deficiency in children and adults. Ann. Neurol. 2003;54(Suppl. 6):S73–S80. doi: 10.1002/ana.10629. [DOI] [PubMed] [Google Scholar]
- 68.Mulligan J.H., Snell E.E. Transport and metabolism of vitamin B6 in Salmonella typhimurium LT2. J. Biol. Chem. 1976;251:1052–1056. [PubMed] [Google Scholar]
- 69.Shane B., Snell E.E. Transport and metabolism of vitamin B6 in the yeast Saccharomyces carlsbergensis 4228. J. Biol. Chem. 1976;251:1042–1051. [PubMed] [Google Scholar]
- 70.Zhang Z.M., McCormick D.B. Uptake of N-(4'-pyridoxyl) amines and release of amines by renal cells: a model for transporter-enhanced delivery of bioactive compounds. Proc. Natl. Acad. Sci. USA. 1991;88:10407–10410. doi: 10.1073/pnas.88.23.10407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Said Z.M., Subramanian V.S., Vaziri N.D., ad Said H.M. Pyridoxine uptake by colonocytes: a specific and regulated carrier-mediated process. Am. J. Physiol. Cell. Physiol. 2008;294:C1192–C1197. doi: 10.1152/ajpcell.00015.2008. [DOI] [PubMed] [Google Scholar]
- 72.Said H.M., Ortiz A., Ma T.Y. A carrier-mediated mechanism for pyridoxine uptake by human intestinal epithelial Caco-2 cells: regulation by a PKA-mediated pathway. Am. J. Physiol. Cell. Physiol. 2003;285:C1219–C1225. doi: 10.1152/ajpcell.00204.2003. [DOI] [PubMed] [Google Scholar]
- 73.Schenker S., Johnson R.F., Mahuren J.D., Henderson G.I., Coburn S.P. Human placental vitamin B6 (pyridoxal) transport: normal characteristics and effects of ethanol. Am. J. Physiol. 1992;262:R966–R974. doi: 10.1152/ajpregu.1992.262.6.R966. [DOI] [PubMed] [Google Scholar]
- 74.Hirose K., Chumnantana R., Nakashima T., Ashiuchi M., Yagi T. Efflux system for pyridoxine in Schizosaccharomyces pombe. Biosci. Biotechnol. Biochem. 2000;64:2675–2679. doi: 10.1271/bbb.64.2675. [DOI] [PubMed] [Google Scholar]
- 75.Yagi T., Tanouchi A., Hiraoka Y. Growth phase-dependent active transport of pyridoxine in a fission yeast, Schizosaccharomyces pombe. FEMS Microbiol. Lett. 1998;161:145–150. doi: 10.1111/j.1574-6968.1998.tb12941.x. [DOI] [PubMed] [Google Scholar]
- 76.Morita T., Takegawa K., Yagi T. Disruption of the plr1+ gene encoding pyridoxal reductase of Schizosaccharomyces pombe. J. Biochem. 2004;135:225–230. doi: 10.1093/jb/mvh026. [DOI] [PubMed] [Google Scholar]
- 77.Stolz J., Vielreicher M. Tpn1p, the plasma membrane vitamin B6 transporter of Saccharomyces cerevisiae. J. Biol. Chem. 2003;278:18990–18996. doi: 10.1074/jbc.M300949200. [DOI] [PubMed] [Google Scholar]
- 78.Stolz J., Wohrmann H.J., Vogl C. Amiloride uptake and toxicity in fission yeast are caused by the pyridoxine transporter encoded by bsu1+ (car1+) Eukaryot. Cell. 2005;4:319–326. doi: 10.1128/EC.4.2.319-326.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Rodionov D.A., Hebbeln P., Eudes A., Ter Beek J., Rodionova I.A., Erkens G.B., Slotboom D.J., Gelfand M.S., Osterman A.L., Hanson A.D., Eitinger T. A Novel Class of Modular Transporters for Vitamins in Prokaryotes. J. Bacteriol. 2009;191:42–51. doi: 10.1128/JB.01208-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Ehrenshaft M., Jenns A.E., Chung K.R., Daub M.E. SOR1, a gene required for photosensitizer and singlet oxygen resistance in Cercospora fungi, is highly conserved in divergent organisms. Mol. Cell. 1998;1:603–609. doi: 10.1016/S1097-2765(00)80060-X. [DOI] [PubMed] [Google Scholar]
- 81.Ehrenshaft M., Chung K.R., Jenns A.E., Daub M.E. Functional characterization of SOR1, a gene required for resistance to photosensitizing toxins in the fungus Cercospora nicotianae. Curr. Genet. 1999;34:478–485. doi: 10.1007/s002940050423. [DOI] [PubMed] [Google Scholar]
- 82.Bilski P., Li M.Y., Ehrenshaft M., Daub M.E., Chignell C.F. Vitamin B6 (pyridoxine) and its derivatives are efficient singlet oxygen quenchers and potential fungal antioxidants. Photochem. Photobiol. 2000;71:129–134. doi: 10.1562/0031-8655(2000)071<0129:SIPVBP>2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 83.Bilski P., Daub M.E., Chignell C.F. Direct detection of singlet oxygen via its phosphorescence from cellular and fungal cultures. Meth. Enzymol. 2002;352:41–52. doi: 10.1016/s0076-6879(02)52005-x. [DOI] [PubMed] [Google Scholar]
- 84.Graham C.M., Ehrenshaft M., Hausner G., Reid D.M. A highly conserved gene for vitamin B biosynthesis may have consequences for stress and hormone responses in plants. Physiol. Plant. 2004;121:8–14. doi: 10.1111/j.0031-9317.2004.00283.x. [DOI] [PubMed] [Google Scholar]
- 85.Chen H., Xiong L. Pyridoxine is required for post-embryonic root development and tolerance to osmotic and oxidative stresses. Plant J. 2005;44:396–408. doi: 10.1111/j.1365-313X.2005.02538.x. [DOI] [PubMed] [Google Scholar]
- 86.Titiz O., Tambasco-Studart M., Warzych E., Apel K., Amrhein N., Laloi C., Fitzpatrick T.B. PDX1 is essential for vitamin B6 biosynthesis, development and stress tolerance in Arabidopsis. Plant J. 2006;48:933–946. doi: 10.1111/j.1365-313X.2006.02928.x. [DOI] [PubMed] [Google Scholar]
- 87.Shi H., Xiong L., Stevenson B., Lu T., Zhu J.K. The Arabidopsis salt overly sensitive 4 mutants uncover a critical role for vitamin B6 in plant salt tolerance. Plant Cell. 2002;14:575–588. doi: 10.1105/tpc.010417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Wagner S., Bernhardt A., Leuendorf J.E., Drewke C., Lytovchenko A., Mujahed N., Gurgui C., Frommer W.B., Leistner E., Fernie A.R., Hellmann H. Analysis of the Arabidopsis rsr4-1/pdx1-3 mutant reveals the critical function of the PDX1 protein family in metabolism, development, and vitamin B6 biosynthesis. Plant Cell. 2006;18:1722–1735. doi: 10.1105/tpc.105.036269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Leuendorf J.E., Genau A., Szewczyk A., Mooney S., Drewke C., Leistner E., Hellmann H. The Pdx1 family is structurally and functionally conserved between Arabidopsis thaliana and Ginkgo biloba. Febs J. 2008;275:960–969. doi: 10.1111/j.1742-4658.2008.06275.x. [DOI] [PubMed] [Google Scholar]
- 90.Gonzalez E., Danehower D., Daub M.E. Vitamer levels, stress response, enzyme activity, and gene regulation of Arabidopsis lines mutant in the pyridoxine/pyridoxamine 5'-phosphate oxidase (PDX3) and the pyridoxal kinase (SOS4) genes involved in the vitamin B6 salvage pathway. Plant Physiol. 2007;145:985–996. doi: 10.1104/pp.107.105189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Antelmann H., Bernhardt J., Schmid R., Mach H., Volker U., Hecker M. First steps from a two-dimensional protein index towards a response-regulation map for Bacillus subtilis. Electrophoresis. 1997;18:1451–1463. doi: 10.1002/elps.1150180820. [DOI] [PubMed] [Google Scholar]
- 92.Savenstrand H., Brosche M., Strid A. Ultraviolet-B signalling: Arabidopsis brassinosteroid mutants are defective in UV-B regulated defence gene expression. Plant Physiol. Biochem. 2004;42:687–694. doi: 10.1016/j.plaphy.2004.06.011. [DOI] [PubMed] [Google Scholar]
- 93.Osmani A.H., May G.S., Osmani S.A. The extremely conserved pyroA gene of Aspergillus nidulans is required for pyridoxine synthesis and is required indirectly for resistance to photosensitizers. J. Biol. Chem. 1999;274:23565–23569. doi: 10.1074/jbc.274.33.23565. [DOI] [PubMed] [Google Scholar]
- 94.Padilla P.A., Fuge E.K., Crawford M.E., Errett A., Werner-Washburne M. The highly conserved, coregulated SNO and SNZ gene families in Saccharomyces cerevisiae respond to nutrient limitation. J. Bacteriol. 1998;180:5718–5726. doi: 10.1128/jb.180.21.5718-5726.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Braun E.L., Fuge E.K., Padilla P.A., Werner-Washburne M. A stationary-phase gene in Saccharomyces cerevisiae is a member of a novel, highly conserved gene family. J. Bacteriol. 1996;178:6865–6872. doi: 10.1128/jb.178.23.6865-6872.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Denslow S.A., Rueschhoff E.E., Daub M.E. Regulation of the Arabidopsis thaliana vitamin B6 biosynthesis genes by abiotic stress. Plant Physiol. Biochem. 2007;45:152–161. doi: 10.1016/j.plaphy.2007.01.007. [DOI] [PubMed] [Google Scholar]
- 97.Kästner U., Hallmen C., Wiese M., Leistner E., Drewke C. The human pyridoxal kinase, a plausible target for ginkgotoxin from Ginkgo biloba. Febs J. 2007;274:1036–1045. doi: 10.1111/j.1742-4658.2007.05654.x. [DOI] [PubMed] [Google Scholar]
- 98.Arenz A., Klein M., Fiehe K., Gross J., Drewke C., Hemscheidt T., Leistner E. Occurrence of Neurotoxic 4'-O-Methylpyridoxine in Ginkgo biloba Leaves, Ginkgo Medications and Japanese Ginkgo Food. Planta Med. 1996;62:548–551. doi: 10.1055/s-2006-957967. [DOI] [PubMed] [Google Scholar]
- 99.Fiehe K., Arenz A., Drewke C., Hemscheidt T., Williamson R.T., Leistner E. Biosynthesis of 4'-O-methylpyridoxine (Ginkgotoxin) from primary precursors. J. Nat. Prod. 2000;63:185–189. doi: 10.1021/np990414+. [DOI] [PubMed] [Google Scholar]
- 100.Wada K., Ishigaki S., Ueda K., Sakata M., Haga M. An antivitamin B6, 4'-methoxypyridoxine, from the seed of Ginkgo biloba L. Chem. Pharm. Bull. (Tokyo) 1985;33:3555–3557. doi: 10.1248/cpb.33.3555. [DOI] [PubMed] [Google Scholar]
- 101.Buss K., Drewke C., Lohmann S., Piwonska A., Leistner E. Properties and interaction of heterologously expressed glutamate decarboxylase isoenzymes GAD(65kDa) and GAD(67kDa) from human brain with ginkgotoxin and its 5'-phosphate. J. Med. Chem. 2001;44:3166–3174. doi: 10.1021/jm010868f. [DOI] [PubMed] [Google Scholar]
- 102.Higuchi O., Nakagawa K., Tsuzuki T., Suzuki T., Oikawa S., Miyazawa T. Aminophospholipid glycation and its inhibitor screening system: a new role of pyridoxal 5'-phosphate as the inhibitor. J. Lipid Res. 2006;47:964–974. doi: 10.1194/jlr.M500348-JLR200. [DOI] [PubMed] [Google Scholar]
- 103.Negre-Salvayre A., Coatrieux C., Ingueneau C., Salvayre R. Advanced lipid peroxidation end products in oxidative damage to proteins. Potential role in diseases and therapeutic prospects for the inhibitors. Br. J. Pharmacol. 2008;153:6–20. doi: 10.1038/sj.bjp.0707395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Metz T.O., Alderson N.L., Thorpe S.R., Baynes J.W. Pyridoxamine, an inhibitor of advanced glycation and lipoxidation reactions: a novel therapy for treatment of diabetic complications. Arch. Biochem. Biophys. 2003;419:41–49. doi: 10.1016/j.abb.2003.08.021. [DOI] [PubMed] [Google Scholar]
- 105.Metz T.O., Alderson N.L., Chachich M.E., Thorpe S.R., Baynes J.W. Pyridoxamine traps intermediates in lipid peroxidation reactions in vivo: evidence on the role of lipids in chemical modification of protein and development of diabetic complications. J. Biol. Chem. 2003;278:42012–42019. doi: 10.1074/jbc.M304292200. [DOI] [PubMed] [Google Scholar]
- 106.Voziyan P.A., Metz T.O., Baynes J.W., Hudson B.G. A post-Amadori inhibitor pyridoxamine also inhibits chemical modification of proteins by scavenging carbonyl intermediates of carbohydrate and lipid degradation. J. Biol. Chem. 2002;277:3397–3403. doi: 10.1074/jbc.M109935200. [DOI] [PubMed] [Google Scholar]
- 107.Booth A.A., Khalifah R.G., Todd P., Hudson B.G. In vitro kinetic studies of formation of antigenic advanced glycation end products (AGEs). Novel inhibition of post-Amadori glycation pathways. J. Biol. Chem. 1997;272:5430–5437. doi: 10.1074/jbc.272.9.5430. [DOI] [PubMed] [Google Scholar]
- 108.Booth A.A., Khalifah R.G., Hudson B.G. Thiamine pyrophosphate and pyridoxamine inhibit the formation of antigenic advanced glycation end-products: comparison with aminoguanidine. Biochem. Biophys. Res. Commun. 1996;220:113–119. doi: 10.1006/bbrc.1996.0366. [DOI] [PubMed] [Google Scholar]
- 109.Mackey A.D., McMahon R.J., Townsend J.H., Gregory J.F., 3rd Uptake, hydrolysis, and metabolism of pyridoxine-5'-beta-D-glucoside in Caco-2 cells. J. Nutr. 2004;134:842–846. doi: 10.1093/jn/134.4.842. [DOI] [PubMed] [Google Scholar]
- 110.Gregory J.F., Ink S.L. Identification and quantification of pyridoxine-β-glucoside as a major form of vitamin B-6 in plant-derived foods. J. Agric. Food Chem. 1987;35:76–82. doi: 10.1021/jf00073a018. [DOI] [Google Scholar]
- 111.Opassiri R., Hua Y., Wara-Aswapati O., Akiyama T., Svasti J., Esen A., Ketudat Cairns J.R. Beta-glucosidase, exo-beta-glucanase and pyridoxine transglucosylase activities of rice BGlu1. Biochem. J. 2004;379:125–131. doi: 10.1042/BJ20031485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Suzuki Y., Uchida K. Formation of 5'-0-(β-D-Glucopyranosyl) pyridoxine in soybean seedlings and suspension cells cultured in the presence of pyridoxine. Bull. Res. Inst. Bioresour. Okayama Univ. 1998;5:107–120. [Google Scholar]
- 113.Scott P.M., Lau B.P., Lawrence G.A., Lewis D.A. Analysis of Ginkgo biloba for the presence of ginkgotoxin and ginkgotoxin 5'-glucoside. J. AOAC Int. 2000;83:1313–1320. [PubMed] [Google Scholar]
- 114.Suzuki Y., Uchida K. Formation of beta-galactosides of pyridoxine using Sporobolomyces singularis. Methods Enzymol. 1997;280:71–77. doi: 10.1016/s0076-6879(97)80103-6. [DOI] [PubMed] [Google Scholar]
- 115.Suzuki Y., Uchida K. Formation of beta-fructosyl compounds of pyridoxine in growing culture of Aspergillus niger. Biosci. Biotechnol. Biochem. 1993;57:875–880. doi: 10.1271/bbb.57.875. [DOI] [PubMed] [Google Scholar]
- 116.Opassiri R., Pomthong B., Onkoksoong T., Akiyama T., Esen A., Ketudat Cairns J.R. Analysis of rice glycosyl hydrolase family 1 and expression of Os4bglu12 beta-glucosidase. BMC Plant Biol. 2006;6:33. doi: 10.1186/1471-2229-6-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Nakano H., Gregory J.F., 3rd. Pyridoxine and pyridoxine-5'-beta-D-glucoside exert different effects on tissue B-6 vitamers but similar effects on beta-glucosidase activity in rats. J. Nutr. 1995;125:2751–2762. doi: 10.1093/jn/125.11.2751. [DOI] [PubMed] [Google Scholar]
- 118.Trumbo P.R., Banks M.A., Gregory J.F., 3rd. Hydrolysis of pyridoxine-5'-beta-D-glucoside by a broad-specificity beta-glucosidase from mammalian tissues. Proc. Soc. Exp. Biol. Med. 1990;195:240–246. doi: 10.3181/00379727-195-43142. [DOI] [PubMed] [Google Scholar]
- 119.Perry T., Holloway H.W., Weerasuriya A., Mouton P.R., Duffy K., Mattison J.A., Greig N.H. Evidence of GLP-1-mediated neuroprotection in an animal model of pyridoxine-induced peripheral sensory neuropathy. Exp. Neurol. 2007;203:293–301. doi: 10.1016/j.expneurol.2006.09.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Albin R.L., Albers J.W., Greenberg H.S., Townsend J.B., Lynn R.B., Burke J.M., Jr., Alessi A.G. Acute sensory neuropathy-neuronopathy from pyridoxine overdose. Neurology. 1987;37:1729–1732. doi: 10.1212/WNL.37.11.1729. [DOI] [PubMed] [Google Scholar]
- 121.Lu T., Xu Y., Monttinen E.S., Kato N. Supplementing vitamin B6 to a low vitamin B6 diet exaggerates UVB-Induced skin tumorigenesis in DMBA-treated hairless mice. J. Nutr. Sci. Vitaminol. (Tokyo) 2008;54:262–265. doi: 10.3177/jnsv.54.262. [DOI] [PubMed] [Google Scholar]
- 122.Wondrak G.T., Roberts M.J., Jacobson M.K., Jacobson E.L. 3-hydroxypyridine chromophores are endogenous sensitizers of photooxidative stress in human skin cells. J. Biol. Chem. 2004;279:30009–30020. doi: 10.1074/jbc.M404379200. [DOI] [PubMed] [Google Scholar]
- 123.Maeda T., Taguchi H., Minami H., Sato K., Shiga T., Kosaka H., Yoshikawa K. Vitamin B6 phototoxicity induced by UVA radiation. Arch. Dermatol. Res. 2000;292:562–567. doi: 10.1007/s004030000174. [DOI] [PubMed] [Google Scholar]
- 124.Theodoratou E., Farrington S.M., Tenesa A., McNeill G., Cetnarskyj R., Barnetson R.A., Porteous M.E., Dunlop M.G., Campbell H. Dietary vitamin B6 intake and the risk of colorectal cancer. Cancer Epidemiol. Biomarkers Prev. 2008;17:171–182. doi: 10.1158/1055-9965.EPI-07-0621. [DOI] [PubMed] [Google Scholar]
- 125.Tsao S.M., Yin M.C., Liu W.H. Oxidant stress and B vitamins status in patients with non-small cell lung cancer. Nutr. Cancer. 2007;59:8–13. doi: 10.1080/01635580701365043. [DOI] [PubMed] [Google Scholar]
- 126.van Uffelen J.G., Chinapaw M.J., van Mechelen W., Hopman-Rock M. Walking or vitamin B for cognition in older adults with mild cognitive impairment? A randomised controlled trial. Br. J. Sports Med. 2008;42:344–351. doi: 10.1136/bjsm.2007.044735. [DOI] [PubMed] [Google Scholar]
- 127.van Uffelen J.G., Chin A.P.M.J., Hopman-Rock M., van Mechelen W. The effect of walking and vitamin B supplementation on quality of life in community-dwelling adults with mild cognitive impairment: a randomized, controlled trial. Qual.Life Res. 2007;16:1137–1146. doi: 10.1007/s11136-007-9219-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Luchsinger J.A., Tang M.X., Miller J., Green R., Mayeux R. Relation of higher folate intake to lower risk of Alzheimer disease in the elderly. Arch. Neurol. 2007;64:86–92. doi: 10.1001/archneur.64.1.86. [DOI] [PubMed] [Google Scholar]
- 129.Balk E.M., Raman G., Tatsioni A., Chung M., Lau J., Rosenberg I.H. Vitamin B6, B12, and folic acid supplementation and cognitive function: a systematic review of randomized trials. Arch. Intern. Med. 2007;167:21–30. doi: 10.1001/archinte.167.1.21. [DOI] [PubMed] [Google Scholar]
- 130.Ford A.H., Flicker L., Thomas J., Norman P., Jamrozik K., Almeida O.P. Vitamins B12, B6, and folic acid for onset of depressive symptoms in older men: results from a 2-year placebo-controlled randomized trial. J. Clin. Psychiatry. 2008;69:1203–1209. doi: 10.4088/JCP.v69n0801. [DOI] [PubMed] [Google Scholar]
- 131.Merete C., Falcon L.M., Tucker K.L. Vitamin B6 is associated with depressive symptomatology in Massachusetts elders. J. Am. Coll. Nutr. 2008;27:421–427. doi: 10.1080/07315724.2008.10719720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Miodownik C., Meoded A., Libov I., Bersudsky Y., Sela B.A., Lerner V. Pyridoxal plasma level in schizophrenic and schizoaffective patients with and without tardive dyskinesia. Clin. Neuropharmacol. 2008;31:197–203. doi: 10.1097/WNF.0b013e3181506623. [DOI] [PubMed] [Google Scholar]
- 133.Lerner V., Miodownik C., Kaptsan A., Bersudsky Y., Libov I., Sela B.A., Witztum E. Vitamin B6 treatment for tardive dyskinesia: a randomized, double-blind, placebo-controlled, crossover study. J. Clin. Psychiatry. 2007;68:1648–1654. doi: 10.4088/JCP.v68n1103. [DOI] [PubMed] [Google Scholar]
- 134.Miodownik C., Lerner V., Statsenko N., Dwolatzky T., Nemets B., Berzak E., Bergman J. Vitamin B6 versus mianserin and placebo in acute neuroleptic-induced akathisia: a randomized, double-blind, controlled study. Clin. Neuropharmacol. 2006;29:68–72. doi: 10.1097/00002826-200603000-00002. [DOI] [PubMed] [Google Scholar]
- 135.Rimland B., Edelson S.M. Parent ratings of behavior effects of biomedical interventions. Autism Research Institute; San Diego: 2005. p. 34. [Google Scholar]
- 136.Audhya T. Laboratory indices of vitamin and mineral deficiency in autism; Presented at Defeat Autism Now!; San Diego, California, USA. 2002. [Google Scholar]
- 137.Lonn E., Yusuf S., Arnold M.J., Sheridan P., Pogue J., Micks M., McQueen M.J., Probstfield J., Fodor G., Held C., Genest J., Jr. Homocysteine lowering with folic acid and B vitamins in vascular disease. N. Engl. J. Med. 2006;354:1567–1577. doi: 10.1056/NEJMoa060900. [DOI] [PubMed] [Google Scholar]
- 138.Ishihara J., Iso H., Inoue M., Iwasaki M., Okada K., Kita Y., Kokubo Y., Okayama A., Tsugane S. Intake of folate, vitamin B6 and vitamin B12 and the risk of CHD: the Japan Public Health Center-Based Prospective Study Cohort I. J. Am. Coll. Nutr. 2008;27:127–136. doi: 10.1080/07315724.2008.10719684. [DOI] [PubMed] [Google Scholar]
- 139.Albert C.M., Cook N.R., Gaziano J.M., Zaharris E., MacFadyen J., Danielson E., Buring J.E., Manson J.E. Effect of folic acid and B vitamins on risk of cardiovascular events and total mortality among women at high risk for cardiovascular disease: a randomized trial. JAMA. 2008;299:2027–2036. doi: 10.1001/jama.299.17.2027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Herrmann M., Schmidt J., Umanskaya N., Colaianni G., Al Marrawi F., Widmann T., Zallone A., Wildemann B., Herrmann W. Stimulation of osteoclast activity by low B-vitamin concentrations. Bone. 2007;41:584–591. doi: 10.1016/j.bone.2007.06.005. [DOI] [PubMed] [Google Scholar]
- 141.Toyota T., Kai Y., Kakizaki M., Ohtsuka H., Shibata Y., Goto Y. The endocrine pancreas in pyridoxine deficient rats. Tohoku J. Exp. Med. 1981;134:331–336. doi: 10.1620/tjem.134.331. [DOI] [PubMed] [Google Scholar]
- 142.Jain S.K. Vitamin B6 (pyridoxamine) supplementation and complications of diabetes. Metabolism. 2007;56:168–171. doi: 10.1016/j.metabol.2006.09.002. [DOI] [PubMed] [Google Scholar]
- 143.MacKenzie K.E., Wiltshire E.J., Gent R., Hirte C., Piotto L., Couper J.J. Folate and vitamin B6 rapidly normalize endothelial dysfunction in children with type 1 diabetes mellitus. Pediatrics. 2006;118:242–253. doi: 10.1542/peds.2005-2143. [DOI] [PubMed] [Google Scholar]
- 144.Denslow S.A., Rueschhoff E.E., Daub M.E. Regulation of the Arabidopsis thaliana vitamin B(6) biosynthesis genes by abiotic stress. Plant Physiol. Biochem. 2007;2:152–161. doi: 10.1016/j.plaphy.2007.01.007. [DOI] [PubMed] [Google Scholar]
- 145.Musayev F.N., Di Salvo M.L., Ko T.P., Schirch V., Safo M.K. Structure and properties of recombinant human pyridoxine 5'-phosphate oxidase. Protein Sci. 2003;12:1455–1463. doi: 10.1110/ps.0356203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Zhao G., Winkler M.E. Kinetic limitation and cellular amount of pyridoxine (pyridoxamine) 5'-phosphate oxidase of Escherichia coli K-12. J. Bacteriol. 1995;177:883–891. doi: 10.1128/jb.177.4.883-891.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Laine-Cessac P., Cailleux A., Allain P. Mechanisms of the inhibition of human erythrocyte pyridoxal kinase by drugs. Biochem. Pharmacol. 1997;54:863–870. doi: 10.1016/s0006-2952(97)00252-9. [DOI] [PubMed] [Google Scholar]
- 148.Hanna M.C., Turner A.J., Kirkness E.F. Human pyridoxal kinase. cDNA cloning, expression, and modulation by ligands of the benzodiazepine receptor. J. Biol. Chem. 1997;272:10756–10760. doi: 10.1074/jbc.272.16.10756. [DOI] [PubMed] [Google Scholar]
- 149.Li M.H., Kwok F., An X.M., Chang W.R., Lau C.K., Zhang J.P., Liu S.Q., Leung Y.C., Jiang T., Liang D.C. Crystallization and preliminary crystallographic studies of pyridoxal kinase from sheep brain. Acta Crystallogr. D. Biol. Crystallogr. 2002;58:1479–1481. doi: 10.1107/S0907444902011034. [DOI] [PubMed] [Google Scholar]