

Abstract
Twice a year in southwestern Nigeria, during a traditional bat festival, community participants enter designated caves to capture bats, which are then consumed for food or traded. We investigated the presence of Bartonella species in Egyptian fruit bats (Rousettus aegyptiacus) and bat flies (Eucampsipoda africana) from these caves and assessed whether Bartonella infections had occurred in persons from the surrounding communities. Our results indicate that these bats and flies harbor Bartonella strains, which multilocus sequence typing indicated probably represent a novel Bartonella species, proposed as Bartonella rousetti. In serum from 8 of 204 persons, we detected antibodies to B. rousetti without cross-reactivity to other Bartonella species. This work suggests that bat-associated Bartonella strains might be capable of infecting humans.
Keywords: Bartonella, bats, bat festival, bat flies, human, zoonoses, vector-borne infections, bacteria, Nigeria
Bats are natural reservoirs for a variety of pathogens (1). However, despite the risk to human health, persons around the world still intentionally handle bats, often without taking appropriate precautions. This lack of precautions is particularly evident in the tropics, where bats are abundant and frequently roost within or in close proximity to humans and domestic animals. In Asia and Africa, larger fruit bats (family Pteropodidae) are used as food, for either cultural reasons or subsistence (2). In some cultures, bat caves serve as spiritual sanctuaries (3).
One particular situation that has attracted the attention of scientists is a bat festival that takes place biannually in the Idanre Hills area of Nigeria. During the festival, which has occurred for many years, men enter designated caves, often without appropriate personal protective equipment, to capture bats. Local customs forbid persons from entering the caves outside of these festivities without permission from the community leadership. The captured bats are then eaten, used in cultural rituals, or sold as bushmeat (3). The predominant bat species within the caves is the Egyptian fruit bat (Rousettus aegyptiacus); colony sizes can reach >1,000 (4). Egyptian fruit bats are known reservoirs of zoonotic pathogens including Lagos bat virus, Marburg virus, and Yersinia pseudotuberculosis (5–8). Given the close human-to-bat contact that occurs during the festival, there is a risk for spillover of batborne pathogens to humans.
The genus Bartonella currently includes >30 species of bacteria (9), many of which have been described only recently. Various arthropod vectors seem to play an essential role in the maintenance and transmission of most known Bartonella species (9,10). In recent years, recognition of multiple Bartonella species as human pathogens responsible for a wide range of clinical manifestations has grown. Numerous novel strains of Bartonella have been discovered in bats of various species around the globe, including the human pathogen Candidatus Bartonella mayotimonensis, which was originally detected in aortic valve tissue of a person with endocarditis (11–13). In addition, a novel Bartonella genotype found in bats from the country of Georgia clustered with genotypes found in human forest workers from Poland (14).
During 2010 and 2013, we researched the health risk to humans participating in the Idanre bat festival. We sampled bats and their ectoparasites from the caves and used them to identify a variety of zoonotic pathogens, including Bartonella. We recruited human participants from the surrounding community and surveyed them (through an orally administered questionnaire and serologic testing) to understand risk factors and the occurrence of pathogen spillover from bats to humans. We examined whether bats and ectoparasites of these bats within the caves used in the Idanre bat festival are infected with Bartonella, characterized any Bartonella species identified in bats or bat flies, and screened human serum samples for evidence of Bartonella infection.
Materials and Methods
Human subjects work was approved by the Centers for Disease Control and Prevention (CDC) Institutional Review Board, the Ahmadu Bello University Human Ethics Board, and the National Health Research Ethics Committee of Nigeria. All animal procedures were conducted in compliance with a field protocol approved by the CDC Animal Institutional Care and Use Committee.
Field Sites, Bat Capture, and Sample Collection
We captured bats by nets in 2 caves in Idanre Hills, Ondo State, southwestern Nigeria, in September 2010 (n = 106) and February 2013 (n = 71). We identified all bats by morphologic characteristics as Egyptian fruit bats (R. aegyptiacus). Captured bats were anesthetized by intramuscular injection of ketamine hydrochloride (0.05–0.1 mg/g bat weight) and exsanguinated via cardiac puncture after surface sterilization with 75% alcohol. Serum and blood clots were separated by centrifugation. Clots were stored at −80°C except while still in the field or being shipped, during which time they were stored on dry ice.
Bat Blood Culture and Characterization of Bartonella Strain
We plated bat blood clots on heart infusion agar containing 10% rabbit blood and incubated in an aerobic atmosphere with 5% carbon dioxide at 35°C for up to 4 weeks. Bacterial colonies morphologically identified as Bartonella were subcultured to obtain pure cultures.
We prepared crude genomic DNA by heating a heavy suspension of pure culture for 10 minutes at 95°C, followed by centrifugation of the lysed cells for 1 minute at 3,000 rpm. The supernatant was then transferred to a clean centrifuge tube to be used as the template DNA. We first verified all isolates obtained from the blood clots as Bartonella spp. by PCR amplification targeting a fragment of the citrate synthase gene (gltA) (15). Positive (B. doshiae) and negative (deionized water) controls were included to ensure that the PCR worked properly.
We purified and sequenced all PCR products of gltA in both directions by using an ABI 3130 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA). We used the Lasergene software package (DNASTAR, Madison, WI, USA) to compare the generated gltA sequences with all available Bartonella species/genotypes. Once the sequences were identified, we selected 1 representative strain (R-191) for further characterization with multilocus sequence typing on the basis of sequence analysis of 8 molecular markers (ftsZ, gltA, nuoG, ribC, rpoB, ssrA, 16S rRNA, and internal transcribed spacers [ITS]) (16). For phylogenetic analyses, we used the neighbor-joining method by the Kimura 2-parameter distance method and bootstrap calculations with 1,000 replicates.
Bat Ectoparasite Collection and Detection of Bartonella DNA
We collected ectoparasites from the skin and pelage of bats and stored them in microcentrifuge tubes with 70% ethanol. Ectoparasite species were identified by using available morphologic keys (17), and identifications were later confirmed by sequencing of the mitochondrial 16S rRNA and cytochrome oxidase I (COI) genes (18,19).
Using a Bullet Blender Gold homogenizer (Next Advance, Averill Park, NY, USA), we homogenized whole ectoparasites in Navy Eppendorf bead tubes (Next Advance) containing 400 μL brain–heart infusion broth (CDC, Atlanta, GA, USA). We extracted DNA from the homogenates by using the KingFisher Flex Purification System and the associated MagMAX Pathogen RNA/DNA Kit (both ThermoFisher, Waltham, MA, USA) according to the manufacturer’s protocols. Detection of Bartonella DNA in ectoparasite samples was performed by nested PCR for gltA (20) because of low concentrations of DNA and by conventional PCR for ITS (21), followed by sequencing and sequence analysis of amplicons.
Preparation of Antigen from the Bartonella Strain Obtained from Bats
We produced a whole-cell antigen by co-cultivating Vero E6 cells with the pure culture (≈106 agar-grown organisms) of the Bartonella strain (R-191) obtained from Egyptian fruit bats. Both were put into T-150 flasks that contained minimum essential medium supplemented with 10% fetal calf serum, 10 mmol HEPES buffer solution, 10 mmol nonessential amino acids, and 2 mmol L-glutamine. The flasks were incubated at 35°C and harvested on postinoculation day 4. At harvest, all but 2 mL of the medium was removed from the flask, sterile glass beads were added, and the flask was gently rocked to remove the Vero E6 cell monolayer. Drops (≈15 μL) of the cell suspension were mounted on each well of 12-well glass slides, which were then air dried, fixed in acetone for 15 minutes, and stored at −70°C until use.
Human Serum Collection and Testing for Antibodies
Persons in communities surrounding the caves who gave consent were enrolled in the study 11–15 days after the first bat festival of 2013 (February 19, 2013); not all of these persons had participated in all bat festival activities. Participants were asked about their contact with bats and their role in the festival, and some provided a blood sample (considered an acute-phase specimen). About 69–78 days later, a follow-up survey was conducted and a second blood sample (considered a convalescent-phase specimen) was collected (the second bat festival of 2013 did not take place between collection of the acute- and convalescent-phase samples). Serum and blood clots were separated by centrifugation; serum was stored at −80°C except while in the field or being shipped, during which time it was stored on dry ice.
To screen human serum, we used an indirect immunofluorescence assay at an initial dilution of 1:32 for IgG against the specific Bartonella antigen from the bat-associated isolate. Antigen-covered wells of the slide were overlaid with dilutions of human serum. Separate slides were included with positive and negative controls. The positive control against the Egyptian fruit bat–associated Bartonella species was produced in laboratory mice via mouse immunization with heat-inactivated bacterium (ProSci Incorporated, Poway, CA, USA). All slides were incubated at 35°C for 30 minutes and then washed in phosphate-buffered saline for 15 minutes. We used anti-human and anti-mouse conjugates (Kirkegaard & Perry Laboratories Inc., Gaithersburg, MD, USA) for human and control serum samples, respectively. Each human serum sample reactive at the initial dilution was further titrated in 2-fold dilutions to endpoint; to check for cross-reactivity, we tested the final positive samples (defined as a titer >1:64) for 3 other Bartonella antigens (B. elizabethae, B. henselae, and B. quintana) previously reported in Africa (22–24).
Results
Bartonella in Egyptian Fruit Bats
We recovered Bartonella isolates from 22 of 177 Egyptian fruit bat blood clots, giving an overall prevalence of 12.4%. The gltA sequences of all Bartonella strains obtained from Egyptian fruit bats were identical or similar (>97% identity) to each other and represented 4 unique variants (GenBank accession nos. HM363764, MH069693–MH069695). A variant is defined when it differs by >1 nt from others. Together with Bartonella strains obtained from Egyptian fruit bats in Kenya (25), these variants constitute a monophyletic genogroup that is distant from all other genotypes previously found in other bat species and any other described Bartonella species.
Multilocus sequence typing of the type strain (gltA; GenBank accession no. HM363764) with 7 additional genetic loci (ftsZ, nuoG, ribC, rpoB, ssrA, 16S rRNA, and ITS) further confirmed the uniqueness of this strain. Sequencing information for each genetic marker demonstrated that the Bartonella strain from the Egyptian fruit bats was distant from all other known Bartonella species and genotypes, including those reported from other bats from Africa. Sequences of all genetic loci obtained during the analyses were deposited in GenBank (accession nos. HM363769, KM387321, HM363779, HM363774, KM382247, HM363784, and KM382255). We compared the fragment sequences of each target with those from other Bartonella species/genotypes. The Egyptian fruit bat–associated Bartonella formed a separate genetic group that was distant from all other Bartonella species with >20% genetic distance and probably represents a novel Bartonella species, according to the definition of La Scola et al. (26). We proposed that this bacterial species be named Bartonella rousetti, to reflect the Egyptian fruit bat (Rousettus aegyptiacus) as the natural host. A phylogenetic tree based on the ITS locus illustrates the relationship of this proposed novel species to other Bartonella species (Figure).
Figure.
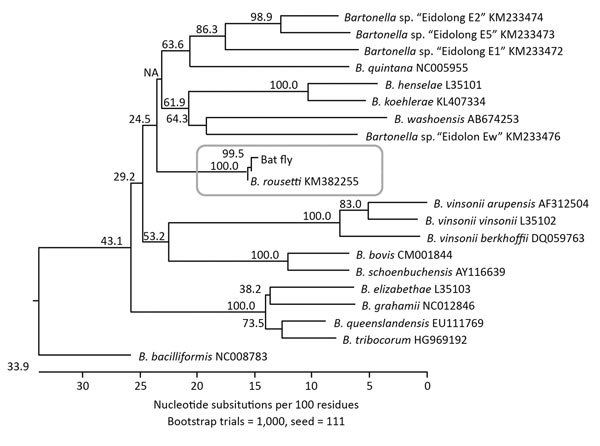
Phylogenetic relationships of Bartonella rousetti (proposed name) obtained from Egyptian fruit bats (Rousettus aegyptiacus) collected in Nigeria, 2010 and 2013, and other Bartonella species and bat-associated Bartonella based on internal transcribed spacer sequences. The neighbor-joining method by the Kimura 2-parameter distance method and bootstrap calculation was conducted with 1,000 replicates for phylogenetic analysis. The internal transcribed spacer sequence obtained from the bat flies was closely clustered with B. rousetti. GenBank accession numbers are provided for the B. rousetti sequence and the comparison sequences.
Identification of Bat Flies and Detection of Bartonella DNA
In 2013, we collected 51 ectoparasites from Egyptian fruit bats. With the exception of 1 unidentified mite, all arthropods were identified as the bat fly Eucampsipoda africana Theodor (Diptera: Nycteribiidae). The morphologic identification of every bat fly was confirmed by 1 or both mitochondrial markers (16S rRNA or COI). Representative 16S rRNA (accession nos. MH138030–MH138037) and COI (accession nos. MH151059–MH151066) sequences have been deposited in GenBank.
Of the 50 DNA extracts from bat flies, 21 (42%) produced >1 ITS or gltA sequence that was confirmed via BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) as Bartonella. Positive samples yielded 19 ITS sequences and 18 gltA sequences; 16 samples yielded sequences for both loci and 5 samples yielded only 1 sequence. All but 1 of the 19 ITS sequences matched closely to the proposed B. rousetti (>95% sequence identity) (Figure); the remaining sequence was identical to B. tamiae (DQ395180). Of the 18 (66.7%) gltA sequences, 12 were close matches for B. rousetti (>98.3% sequence identity); all 12 of these samples also produced ITS sequences matching this strain.
The remaining 6 gltA sequences were identical to Bartonella sequences detected in a louse (Neohaematopinus sciuri) collected from a dead Eastern gray squirrel (Sciurus carolinensis) at a zoo in Greenville, SC, USA (GenBank accession no. EU368000) and an unidentified tick collected from a sheep in Peru (GenBank accession no. AF415209). These sequences were also closely (>99% sequence identity) related to other sequences from fleas (Ctenocephalides felis and C. canis) collected from dogs in Tunisia (GenBank accession nos. KP126468–74), a louse pool (Polyplax spp. and Hoplopleura spp.) collected from rodents in Thailand (GenBank accession no. KT324560), and an unidentified flea collected from a dog in Peru (GenBank accession no. GU583843). Of the specimens with this particular gltA sequence, 2 yielded no ITS sequence, 3 yielded ITS sequences matching B. rousetti, and 1 yielded the single B. tamiae sequence. Representative ITS (GenBank accession nos. MH14262–MH142639) and gltA (GenBank accession nos. MH151067–79) sequences for each novel sequence variant were submitted to GenBank.
Human Exposure to B. rousetti
A total of 305 serum samples from 204 participants were tested for IgG against B. rousetti; 12 samples from different persons showed reactivity at an initial dilution of 1:32. Further 2-fold titration confirmed that 8 were positive, with titers >1:64 (Table). The positive samples were retested for 3 other Bartonella species—B. henselae, B. quintana, and B. elizabethae, all of which have been reported in Africa (22–24); antibodies against these Bartonella species were not detected in any of the samples. Five seropositive participants reported having eaten bats and having either touched bats or been scratched or bitten by them, although not all reported having ever participated in the bat festival. Three seropositive participants reported never having eaten bats, touched bats, or been scratched or bitten by bats; in addition, these 3 participants claimed to have never participated in the bat festival. Of the 8 seropositive participants, only 1 reported having experienced a febrile illness since the bat festival that had taken place earlier in the year.
Table. Epidemiologic data for persons with antibodies to Bartonella rousetti detected in study of human exposure to a novel Bartonella species from contact with fruit bats, Nigeria, 2013*.
| Participant age, y/sex | Titer in acute-phase serum† | Titer in convalescent-phase serum† | Ever ate bat | Ever participated in bat festival | Last time touched, scratched, or bitten by bat | Febrile illness since first bat festival of 2013 |
|---|---|---|---|---|---|---|
| 45/F | <1:32 | 1:64 | Yes | No | 6–12 mo ago | No |
| 37/M | <1:32 | 1:64 | Yes | No | >12 mo ago | No |
| 25/F | <1:32 | 1:512 | Yes | Yes | <1 mo ago | No |
| 30/F | <1:32 | 1:512 | No | No | Never | No |
| 21/M | 1:64 | <1:32 | No | No | Never | No |
| 44/F | 1:64 | <1:32 | Yes | Yes | <1 mo ago | No |
| 70/M | 1:256 | No sample | Yes | No | >12 mo ago | Yes |
| 32/F | 1:256 | No sample | No | No | Never | No |
*Bartonella rousetti is the proposed name for the novel Bartonella species identified in Egyptian fruit bats in Nigeria. †Acute-phase samples collected within 11–15 d after first bat festival of 2013; convalescent-phase samples collected 69–78 d after acute-phase sample collection (the second bat festival of 2013 did not take place between collections of acute- and convalescent-phase samples).
Discussion
We made several observations during this investigation. First, Egyptian fruit bats carry a unique Bartonella strain that probably represents a new species, for which we propose the name Bartonella rousetti. Second, bat flies, the common ectoparasites of bats, carry this same strain of Bartonella. Because this organism was detected by PCR only, the presence of the DNA does not necessarily indicate that the organism is viable. Last, persons from the communities surrounding the bat caves were exposed to this particular Bartonella strain, which might cause human infection.
Since 2010, several reports have described finding diverse Bartonella genotypes in bats of many species (25,27–30). The relationships between Bartonella genotypes and bat species that harbored these bacteria are not always simple. The same Bartonella species may circulate among different bat species, showing no specific relationship between the bats and the Bartonella species (27). Sometimes, multiple Bartonella species are associated with bats of only 1 species. For example, 6 Bartonella species have been identified in straw-colored fruit bats (Eidolon helvum) in Africa (16,29). Our study indicates that Egyptian fruit bats carry a specific Bartonella strain that has not been identified in bats of other species.
Similarly, we found that the most prevalent Bartonella species found in bat flies parasitizing Egyptian fruit bats is B. rousetti. The ectoparasite bat flies E. africana are predominantly associated with Egyptian fruit bats (17,31,32). Although sequences matching other Bartonella species were identified in the bat flies, these genogroups may be primarily associated with arthropods and not mammals. One sequence from a bat fly was identified as B. tamiae. The presence of B. tamiae in bat flies from Algeria has been recorded (33), and the bacterium reportedly has been identified in chigger mites collected from rodents in Thailand (34). It is possible that Bartonella species found only in arthropods and not their associated mammal hosts may represent facultative symbionts that are uniquely adapted to live in the arthropod gut or other body system (35,36). The risks posed to humans by these primarily arthropod-associated Bartonella species are still unclear, although B. tamiae is a reported human pathogen that may cause febrile illness and other clinical signs and symptoms (37).
Detection of antibodies against B. rousetti in serum samples from several study participants indicates their exposure to the bacteria. However, with serologic results, cross-reactivity is a concern. For example, phylogenetically closely related B. henselae and B. quintana (the causative agents of cat-scratch disease and trench fever, respectively) exhibit a high level of serologic cross-reactivity (26,38,39). We tested the positive human serum samples for 3 other Bartonella species (B. henselae, B. quintana, and B. elizabethae) that circulate in Africa, and we did not detect any positive results. Given that immunofluorescence assays have good discriminatory ability for a wide range of antigens (40–42), the results lead us to conclude that the antibodies in these participants were indeed reactive with B. rousetti but not the other Bartonella species tested, although cross-reactivity with other non-Bartonella agents cannot be ruled out.
Our study is not the first attempt to identify antibodies against bat-associated Bartonella in humans. Mannerings et al. (43) conducted a serologic survey of 335 volunteers from Ghana for antibodies against 6 species of Bartonella, including Bartonella strains isolated from straw-colored fruit bats. In that study, only 2 serum samples were positive for B. henselae antibodies at low titers, whereas none was positive against the bat strains.
All known species of Bartonella are transmitted between natural animal hosts by arthropods (29,44). The presence of B. rousetti DNA in E. africana bat flies parasitizing Egyptian fruit bats suggests that these ectoparasites may act as vectors for the transmission of Bartonella infection among bats, but it is unclear how bat flies would play a role in transmitting the bacterium to humans because bat flies do not commonly bite humans (C. McKee, unpub. data). Instead, human exposure may potentially occur via other routes, such as 1) directly by bat bites or scratches, which is similar to how humans acquire infections with B. henselae through cat scratches (10,45); 2) indirectly by contamination of open wounds with blood or other materials (e.g., saliva, urine, feces) of infected bats; or 3) indirectly by contamination of open wounds with bat fly excreta. Several studies have reported detecting Bartonella DNA in bat feces (12,46,47), and Dietrich et al. (47) detected Bartonella DNA in bat saliva and urine, providing support for routes 1 and 2 above, although no attempts have been made to culture viable bacteria from these fluids. However, viable Bartonella bacteria have been cultured from experimentally infected ectoparasites, including fleas and bedbugs (48,49), although such studies have yet to be performed for bat flies. Nevertheless, evidence is accumulating that Bartonella could spread from infected mammalian hosts through multiple routes. Therefore, it may not be necessary for humans to interact directly with live bats to be exposed to bat-associated Bartonella. Persons might be at risk when interacting with bat carcasses, guano, or other contaminated products. Of note, we do not provide definitive evidence of the route of exposure for any of the 8 seropositive participants. Indeed, 3 of these participants reported no interactions at all with bats.
Future studies should continue to evaluate the relative correlations of exposure routes, the pathobiology of batborne B. rousetti in humans, and vector competency of bat flies for transmitting Bartonella. Results should provide guidance to communities for mitigating the risks to humans interacting with animals and their arthropod vectors.
Acknowledgments
Idanre Bat Festival Investigation Team members: Ivan V. Kuzmin, Dianna Blau, James Ellison, Lauren Greenberg, Marissa Person, Ryan Wallace, Panayampalli S. Satheshkumar, Abimbola Aman-Oloniyo, Elizabeth B. Adedire, Mariat O. Soleye, Gloria C. Okara, Sebastian Yennan, Mohammed Abdurrahman, Munir A. Sani, Solomon. W Audu, Maruf Lawal, and Philip P. Mshelbwala.
Acknowledgments
We thank J.D. Kirby, Mary Reynolds, Todd Smith, the Vice Chancellor and Management of Ahmadu Bello University, the Federal Ministry of Health (Abuja, Nigeria), the Owa of Idanre Oba Fredrick Adegunle Aroloye IV, and the chiefs of the Idanre community, Ondo State, Nigeria, for their helpful comments and assistance with logistics.
This study was supported by the Biosecurity Engagement Program of the US Department of State, Bureau of International Security and Nonproliferation, and the Office of Cooperative Threat Reduction’s Global Threat Reduction Programs; One Health funding; and the Global Disease Detection Program of the Center for Global Health at CDC.
Biography
Dr. Bai is a microbiologist in the Division of Vector-Borne Infectious Diseases, National Center for Emerging and Zoonotic Infectious Diseases, CDC, Fort Collins. Her research interests include microbiology, epidemiology, and ecology of zoonotic infectious diseases.
Footnotes
Suggested citation for this article: Bai Y, Osinubi MOV, OsikowiczL, McKee C, Vora NM, Rizzo MR, et al. Human exposure to novel Bartonella species from contact with fruit bats, Nigeria. Emerg Infect Dis. 2018 Dec [date cited]. https://doi.org/10.3201/eid2412.181204
Team members are listed at the end of this article.
References
- 1.Calisher CH, Childs JE, Field HE, Holmes KV, Schountz T. Bats: important reservoir hosts of emerging viruses. Clin Microbiol Rev. 2006;19:531–45. 10.1128/CMR.00017-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Mickleburgh S, Waylen K, Racey P. Bats as bushmeat: a global review. Oryx. 2009;43:217–34. 10.1017/S0030605308000938 [DOI] [Google Scholar]
- 3.Osinubi MO, Recuenco S, Kuzmin I, Haberling DL, Blau DM, Davis LB, et al. Knowledge, attitudes and practices among populations exposed to bats in southern Nigeria. Revista de Educação Continuada em Medicina Veterinária e Zootecnia do CRMV-SP. 2012;10, n. 2/3.
- 4.Kwiecinski GG, Griffiths TA. Rousettus egyptiacus. Mamm Species. 1999;611:1–9. 10.2307/3504411 [DOI] [Google Scholar]
- 5.Kuzmin IV, Niezgoda M, Franka R, Agwanda B, Markotter W, Beagley JC, et al. Lagos bat virus in Kenya. J Clin Microbiol. 2008;46:1451–61. 10.1128/JCM.00016-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Towner JS, Amman BR, Sealy TK, Carroll SA, Comer JA, Kemp A, et al. Isolation of genetically diverse Marburg viruses from Egyptian fruit bats. PLoS Pathog. 2009;5:e1000536. 10.1371/journal.ppat.1000536 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Nakamura S, Settai S, Hayashidani H, Urabe T, Namai S, Une Y. Outbreak of yersiniosis in Egyptian rousette bats (Rousettus aegyptiacus) caused by Yersinia pseudotuberculosis serotype 4b. J Comp Pathol. 2013;148:410–3. 10.1016/j.jcpa.2012.07.007 [DOI] [PubMed] [Google Scholar]
- 8.Amman BR, Jones ME, Sealy TK, Uebelhoer LS, Schuh AJ, Bird BH, et al. Oral shedding of Marburg virus in experimentally infected Egyptian fruit bats (Rousettus aegyptiacus). J Wildl Dis. 2015;51:113–24. 10.7589/2014-08-198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chomel BB, Boulouis HJ, Breitschwerdt EB, Kasten RW, Vayssier-Taussat M, Birtles RJ, et al. Ecological fitness and strategies of adaptation of Bartonella species to their hosts and vectors. Vet Res. 2009;40:29. 10.1051/vetres/2009011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Chomel BB, Kasten RW, Floyd-Hawkins K, Chi B, Yamamoto K, Roberts-Wilson J, et al. Experimental transmission of Bartonella henselae by the cat flea. J Clin Microbiol. 1996;34:1952–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lin EY, Tsigrelis C, Baddour LM, Lepidi H, Rolain JM, Patel R, et al. Candidatus Bartonella mayotimonensis and endocarditis. Emerg Infect Dis. 2010;16:500–3. 10.3201/eid1603.081673 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Veikkolainen V, Vesterinen EJ, Lilley TM, Pulliainen AT. Bats as reservoir hosts of human bacterial pathogen, Bartonella mayotimonensis. Emerg Infect Dis. 2014;20:960–7. 10.3201/eid2006.130956 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lilley TM, Wilson CA, Bernard RF, Willcox EV, Vesterinen EJ, Webber QM, et al. Molecular detection of Candidatus Bartonella mayotimonensis in North American bats. Vector Borne Zoonotic Dis. 2017;17:243–6. 10.1089/vbz.2016.2080 [DOI] [PubMed] [Google Scholar]
- 14.Urushadze L, Bai Y, Osikowicz L, McKee C, Sidamonidze K, Putkaradze D, et al. Prevalence, diversity, and host associations of Bartonella strains in bats from Georgia (Caucasus). PLoS Negl Trop Dis. 2017;11:e0005428. 10.1371/journal.pntd.0005428 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Norman AF, Regnery R, Jameson P, Greene C, Krause DC. Differentiation of Bartonella-like isolates at the species level by PCR-restriction fragment length polymorphism in the citrate synthase gene. J Clin Microbiol. 1995;33:1797–803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bai Y, Hayman DTS, McKee CD, Kosoy MY. Classification of Bartonella strains associated with straw-colored fruit bats (Eidolon helvum) across Africa using a multi-locus sequence typing platform. PLoS Negl Trop Dis. 2015;9:e0003478. 10.1371/journal.pntd.0003478 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Theodor O. The Nycteribiidae of the Ethiopian region and Madagascar. Parasitology. 1957;47:457–543. 10.1017/S0031182000022162 [DOI] [PubMed] [Google Scholar]
- 18.Quetglas J, Balvín O, Lučan RK, Benda P. First records of the bat bug Cacodmus vicinus (Heteroptera: Cimicidae) from Europe and further data on its distribution. Vespertilio. 2012;16:243–8. [Google Scholar]
- 19.Szalanski AL, Austin JW, Scheffrahn RH, Messenger MT. Molecular diagnostics of the Formosan subterranean termite (Isoptera: Rhinotermitidae). Fla Entomol. 2004;87:145–51. 10.1653/0015-4040(2004)087[0145:MDOTFS]2.0.CO;2 [DOI] [Google Scholar]
- 20.Bai Y, Gilbert A, Fox K, Osikowicz L, Kosoy M. Bartonella rochalimae and B. vinsonii subsp. berkhoffii in wild carnivores from Colorado, USA. J Wildl Dis. 2016;52:844–9. 10.7589/2016-01-015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Diniz PP, Maggi RG, Schwartz DS, Cadenas MB, Bradley JM, Hegarty B, et al. Canine bartonellosis: serological and molecular prevalence in Brazil and evidence of co-infection with Bartonella henselae and Bartonella vinsonii subsp. berkhoffii. Vet Res. 2007;38:697–710. 10.1051/vetres:2007023 [DOI] [PubMed] [Google Scholar]
- 22.Kelly PJ, Rooney JJ, Marston EL, Jones DC, Regnery RL. Bartonella henselae isolated from cats in Zimbabwe. Lancet. 1998;351:1706. 10.1016/S0140-6736(05)77744-8 [DOI] [PubMed] [Google Scholar]
- 23.Boutellis A, Veracx A, Angelakis E, Diatta G, Mediannikov O, Trape JF, et al. Bartonella quintana in head lice from Sénégal. Vector Borne Zoonotic Dis. 2012;12:564–7. 10.1089/vbz.2011.0845 [DOI] [PubMed] [Google Scholar]
- 24.Kamani J, Morick D, Mumcuoglu KY, Harrus S. Prevalence and diversity of Bartonella species in commensal rodents and ectoparasites from Nigeria, West Africa. PLoS Negl Trop Dis. 2013;7:e2246. 10.1371/journal.pntd.0002246 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kosoy M, Bai Y, Lynch T, Kuzmin IV, Niezgoda M, Franka R, et al. Bartonella spp. in bats, Kenya. Emerg Infect Dis. 2010;16:1875–81. 10.3201/eid1612.100601 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.La Scola B, Raoult D. Serological cross-reactions between Bartonella quintana, Bartonella henselae, and Coxiella burnetii. J Clin Microbiol. 1996;34:2270–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Bai Y, Kosoy M, Recuenco S, Alvarez D, Moran D, Turmelle A, et al. Bartonella spp. in Bats, Guatemala. Emerg Infect Dis. 2011;17:1269–72. 10.3201/eid1707.101867 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bai Y, Recuenco S, Gilbert AT, Osikowicz LM, Gómez J, Rupprecht C, et al. Prevalence and diversity of Bartonella spp. in bats in Peru. Am J Trop Med Hyg. 2012;87:518–23. 10.4269/ajtmh.2012.12-0097 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kamani J, Baneth G, Mitchell M, Mumcuoglu KY, Gutiérrez R, Harrus S. Bartonella species in bats (Chiroptera) and bat flies (Nycteribiidae) from Nigeria, West Africa. Vector Borne Zoonotic Dis. 2014;14:625–32. 10.1089/vbz.2013.1541 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Brook CE, Bai Y, Dobson AP, Osikowicz LM, Ranaivoson HC, Zhu Q, et al. Bartonella spp. in fruit bats and blood-feeding Ectoparasites in Madagascar. PLoS Negl Trop Dis. 2015;9:e0003532. 10.1371/journal.pntd.0003532 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Nartey NAN. Common parasites of fruit-eating bats in southern Ghana. Accra (Ghana): University of Ghana; 2015. p. 1–144. [Google Scholar]
- 32.Charles AN. Haemoparasites and ectoparasites of fruit bat species in Amurum Forest Reserve and their effects on host physiologic and morphometric parameters. Jos (Nigeria): University of Jos; 2015. p. 1–54. [Google Scholar]
- 33.Leulmi H, Aouadi A, Bitam I, Bessas A, Benakhla A, Raoult D, et al. Detection of Bartonella tamiae, Coxiella burnetii and rickettsiae in arthropods and tissues from wild and domestic animals in northeastern Algeria. Parasit Vectors. 2016;9:27. 10.1186/s13071-016-1316-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kabeya H, Colborn JM, Bai Y, Lerdthusnee K, Richardson JH, Maruyama S, et al. Detection of Bartonella tamiae DNA in ectoparasites from rodents in Thailand and their sequence similarity with bacterial cultures from Thai patients. Vector Borne Zoonotic Dis. 2010;10:429–34. 10.1089/vbz.2009.0124 [DOI] [PubMed] [Google Scholar]
- 35.Neuvonen MM, Tamarit D, Näslund K, Liebig J, Feldhaar H, Moran NA, et al. The genome of Rhizobiales bacteria in predatory ants reveals urease gene functions but no genes for nitrogen fixation. Sci Rep. 2016;6:39197. 10.1038/srep39197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Segers FH, Kešnerová L, Kosoy M, Engel P. Genomic changes associated with the evolutionary transition of an insect gut symbiont into a blood-borne pathogen. ISME J. 2017;11:1232–44. 10.1038/ismej.2016.201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kosoy M, Morway C, Sheff KW, Bai Y, Colborn J, Chalcraft L, et al. Bartonella tamiae sp. nov., a newly recognized pathogen isolated from three human patients from Thailand. J Clin Microbiol. 2008;46:772–5. 10.1128/JCM.02120-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Baneth G, Kordick DL, Hegarty BC, Breitschwerdt EB. Comparative seroreactivity to Bartonella henselae and Bartonella quintana among cats from Israel and North Carolina. Vet Microbiol. 1996;50:95–103. 10.1016/0378-1135(96)00006-5 [DOI] [PubMed] [Google Scholar]
- 39.Vermeulen MJ, Verbakel H, Notermans DW, Reimerink JH, Peeters MF. Evaluation of sensitivity, specificity and cross-reactivity in Bartonella henselae serology. J Med Microbiol. 2010;59:743–5. 10.1099/jmm.0.015248-0 [DOI] [PubMed] [Google Scholar]
- 40.Iralu J, Bai Y, Crook L, Tempest B, Simpson G, Mckenzie T, et al. Rodent-associated Bartonella febrile illness, Southwestern United States. Emerg Infect Dis. 2006;12:1081–6. 10.3201/eid1207.040397 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Laudisoit A, Iverson J, Neerinckx S, Shako JC, Nsabimana JM, Kersh G, et al. Human seroreactivity against Bartonella species in the Democratic Republic of Congo. Asian Pac J Trop Med. 2011;4:320–2. 10.1016/S1995-7645(11)60094-1 [DOI] [PubMed] [Google Scholar]
- 42.Myint KS, Gibbons RV, Iverson J, Shrestha SK, Pavlin JA, Mongkolsirichaikul D, et al. Serological response to Bartonella species in febrile patients from Nepal. Trans R Soc Trop Med Hyg. 2011;105:740–2. 10.1016/j.trstmh.2011.08.002 [DOI] [PubMed] [Google Scholar]
- 43.Mannerings AO, Osikowicz LM, Restif O, Nyarko E, Suu-Ire R, Cunningham AA, et al. Exposure to bat-associated Bartonella spp. among humans and other animals, Ghana. Emerg Infect Dis. 2016;22:922–4. 10.3201/eid2205.151908 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Billeter SA, Hayman DT, Peel AJ, Baker K, Wood JL, Cunningham A, et al. Bartonella species in bat flies (Diptera: Nycteribiidae) from western Africa. Parasitology. 2012;139:324–9. 10.1017/S0031182011002113 [DOI] [PubMed] [Google Scholar]
- 45.Mosbacher M, Elliott SP, Shehab Z, Pinnas JL, Klotz JH, Klotz SA. Cat scratch disease and arthropod vectors: more to it than a scratch? J Am Board Fam Med. 2010;23:685–6. 10.3122/jabfm.2010.05.100025 [DOI] [PubMed] [Google Scholar]
- 46.Banskar S, Bhute SS, Suryavanshi MV, Punekar S, Shouche YS. Microbiome analysis reveals the abundance of bacterial pathogens in Rousettus leschenaultii guano. Sci Rep. 2016;6:36948. 10.1038/srep36948 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Dietrich M, Kearney T, Seamark ECJ, Markotter W. The excreted microbiota of bats: evidence of niche specialisation based on multiple body habitats. FEMS Microbiol Lett. 2017;364:fnw284. 10.1093/femsle/fnw284 [DOI] [PubMed] [Google Scholar]
- 48.Kernif T, Leulmi H, Socolovschi C, Berenger J-M, Lepidi H, Bitam I, et al. Acquisition and excretion of Bartonella quintana by the cat flea, Ctenocephalides felis felis. Mol Ecol. 2014;23:1204–12. 10.1111/mec.12663 [DOI] [PubMed] [Google Scholar]
- 49.Leulmi H, Bitam I, Berenger J-M, Lepidi H, Rolain J-M, Almeras L, et al. Competence of Cimex lectularius bed bugs for the transmission of Bartonella quintana, the agent of trench fever. PLoS Negl Trop Dis. 2015;9:e0003789. 10.1371/journal.pntd.0003789 [DOI] [PMC free article] [PubMed] [Google Scholar]