

Abstract
In recent decades, advances in lighting technology have precipitated exponential increases in night sky brightness worldwide, raising concerns in the scientific community about the impact of artificial light at night (ALAN) on crepuscular and nocturnal biodiversity. Long‐term records show that insect abundance has declined significantly over this time, with worrying implications for terrestrial ecosystems. The majority of investigations into the vulnerability of nocturnal insects to artificial light have focused on the flight‐to‐light behavior exhibited by select insect families. However, ALAN can affect insects in other ways as well. This review proposes five categories of ALAN impact on nocturnal insects, highlighting past research and identifying key knowledge gaps. We conclude with a summary of relevant literature on bioluminescent fireflies, which emphasizes the unique vulnerability of terrestrial light‐based communication systems to artificial illumination. Comprehensive understanding of the ecological impacts of ALAN on diverse nocturnal insect taxa will enable researchers to seek out methods whereby fireflies, moths, and other essential members of the nocturnal ecosystem can coexist with humans on an increasingly urbanized planet.
Keywords: artificial light at night, bioluminescence, fireflies, light pollution, nocturnal insects, visual ecology
1. INTRODUCTION
Localized illumination of nocturnal landscapes by anthropogenic sources of light such as street lamps, path lights, and vehicle headlights, hereafter referred to collectively as artificial light at night (ALAN), is likely to disrupt populations of crepuscular and nocturnal animal species present in affected habitats (Davies & Smyth, 2018; Gaston & Holt, 2018; Navara & Nelson, 2007; Rich & Longcore, 2006). Recent global surveys of night sky brightness have concluded that 23% of land surfaces between 75°N and 60°S, including 88% of Europe and 47% of the United States, experience nightly “light pollution” in the form of an increase in night sky brightness at least 8% above the natural level (Falchi et al., 2016) and that this artificial illumination is gradually invading biodiversity hot spots (Guetté, Godet, Juigner, & Robin, 2018). A meta‐analysis of night sky brightness over time has found regional increases ranging from 0% to 20% per year, averaging 6% (Hölker, Moss, et al., 2010). Such increases are expected to match the pace of urban expansion (Elvidge et al., 2014). In the past two decades, however, some regions have experienced net decreases in night sky brightness, which may be due to environmental and economic incentives to reduce ALAN, as well as to technological innovations such as shielded lights that mitigate light trespass by restricting artificial illumination to desired areas (Bennie, Davies, Duffy, Inger, & Gaston, 2014, but see Kyba et al., 2017).
The ecological effects of ALAN depend on both its intensity and its spectral composition (as well as its flicker rate; see Inger, Bennie, Davies, & Gaston, 2014 for a review). Historically, ALAN sources have mainly comprised low‐ and high‐pressure sodium lamps, which emit characteristic emission spectra concentrated in the yellow‐to‐orange region of the visible spectrum, in addition to “whiter” broad‐spectrum mercury vapor lamps, which also emit a large amount of UV radiation (Figure 1). Most recently, environmental and economic concerns have galvanized movements to replace these traditional, energy‐intensive light sources with energy‐efficient alternatives, primarily LEDs (Davies, Bennie, Inger, & Gaston, 2013; Gaston, Davies, Bennie, & Hopkins, 2012). LEDs can emit monochromatic light of any desired wavelength within or adjacent to the visible spectrum; white light of a given color temperature is produced by coating blue LEDs in a phosphor material that absorbs a percentage of this light and re‐emits it in longer wavelengths (Krames et al., 2007). As a result of this process, most commercial white LED street lamps emit more of their light in the blue region of the visible spectrum than do other ALAN types (Figure 1). Exposure to blue light at night is known to cause insomnia and increased disease risk in humans (American Medical Association, 2016). However, the effects of this widespread spectral shift on other organisms have only recently attracted the attention of researchers (Davies et al., 2017; Donners et al., 2018; Gaston, Visser, & Hölker, 2015; Justice & Justice, 2016; Lewanzik & Voigt, 2017; Longcore et al., 2015, 2018 ; Pawson & Bader, 2014; Plummer, Hale, O'Callaghan, Sadler, & Siriwardena, 2016; Somers‐Yeates, Hodgson, McGregor, Spalding, & Ffrench‐Constant, 2013; Spoelstra et al., 2015; van Grunsven et al., 2014; van Langevelde, Grunsven, Veenendaal, & Fijen, 2017; Wakefield, Broyles, Stone, Jones, & Harris, 2016).
Figure 1.
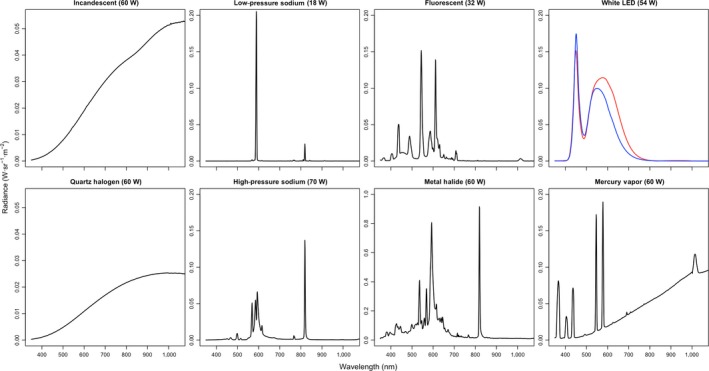
Spectral emission of different ALAN types. ALAN sources, such as incandescent and halogen bulbs, and mercury vapor lamps emit large amounts of energy as infrared radiation (heat); mercury vapor and metal halide lamps also emit a non‐negligible amount of UV radiation. Low‐pressure sodium lamps and LEDs are comparatively efficient ALAN sources, both capable of emitting nearly monochromatic visible light (see text). Neutral (red) and cool (blue) temperature white LEDs have been plotted on the same graph for comparison. Modified from Elvidge, Keith, Tuttle, and Baugh (2010) with permission
Worldwide, around 30% of vertebrates and more than 60% of invertebrates are nocturnal (Hölker, Wolter, Perkin, & Tockner, 2010). These species are undoubtedly most vulnerable to artificial illumination; they also tend to be underrepresented in the scientific literature. To bring greater attention to the ecological consequences of human activity in an often ignored temporal niche, herein we review known individual‐level impacts of ALAN on nocturnal animals, organized into the following five categories: temporal and spatial disorientation, attraction, desensitization, and recognition. Temporal disorientation covers both alterations in circadian clocks and photoperiodism as well as the partitioning of activity between day and night (sensu Gaston, Bennie, Davies, & Hopkins, 2013). To distinguish between discrete effects of ALAN on nocturnal insect taxa, visual perception (sensu Gaston et al., 2013) has been subdivided into the categories of recognition and desensitization; similarly, spatial orientation (sensu Gaston et al., 2013) has been subdivided into spatial disorientation and attraction. As the behavioral responses of individuals determine how ALAN will affect populations, and then entire ecological communities, we believe that this level of assessment is illuminating.
Recent studies have suggested that insect diversity and abundance are both undergoing a rapid decline (Hallmann et al., 2017). Insects are essential components of all terrestrial food webs, and any losses in insect biomass are likely to have widespread ecological ramifications. ALAN affects insects in unique ways related to their body size and visual system. We have therefore chosen to focus our review around nocturnal and crepuscular insects, although our classification may be applicable to other organisms as well. Previous reviews cover the effects of ALAN on plants (Bennie, Davies, Cruse, & Gaston, 2016), butterflies (Seymoure, 2018), and stream and riparian ecosystems (Perkin et al., 2011), as well as the effects of artificial light regimes on pest insects (Johansen, Vänninen, Pinto, Nissinen, & Shipp, 2011) and poultry (Van Nuffel, Bujis, & Delezie, 2015); for general reviews of the impacts of ALAN on all species, see Rich and Longcore (2006) and Gaston et al. (2013).
Fireflies (Coleoptera: Lampyridae), click beetles (Coleoptera: Elateridae), and glowworms (Diptera: Keroplatidae) are among the most charismatic of all nocturnal insects, and their unique light‐based communication system may make them especially vulnerable to artificial illumination. We therefore conclude with a review and synthesis of existing studies concerning the impact of ALAN on bioluminescent insects, followed by recommendations for future research.
2. INSECT VISION
As background for understanding the ecological effects of ALAN, some familiarity with how light is measured, and how nocturnal insects perceive light, is necessary. These topics are briefly introduced below; for a more in‐depth primer, see Cronin et al. (2014) and Honkanen, Immonen, Salmela, Heimonen, and Weckström (2017).
Light behaves both as a wave and as a particle (or photon). Wavelength is measured in nanometers (nm), with wavelengths between 400 and 700 nm corresponding to colors visible to humans. Intensity (particle density) is measured in units of counts per unit area (photons cm−2 s−1), with more photons corresponding to greater intensity. Brightness, a subjective perception of intensity, is influenced by individual spectral sensitivity, which is generally viewed as fixed for a given species. In trichromatic humans, who have blue‐, green‐, and red‐sensitive photoreceptors, luminance sensitivity peaks around 555 nm under photopic (well‐lit) conditions (Figure 2a,b). Photon counts can be weighted by this function to obtain measurements of brightness as perceived by the human eye, given in units of lumens (lm). Light emitted from a point source can be standardized to candelas, or lumens per solid angle (cd = lm/sr), and light incident on a flat surface is converted to lux, or lumens per area (lux = lm/m2). Although lux is widely used by engineers and policymakers, and is easiest to measure, it poorly approximates brightness as it is perceived by non‐human animals (Longcore & Rich, 2004).
Figure 2.
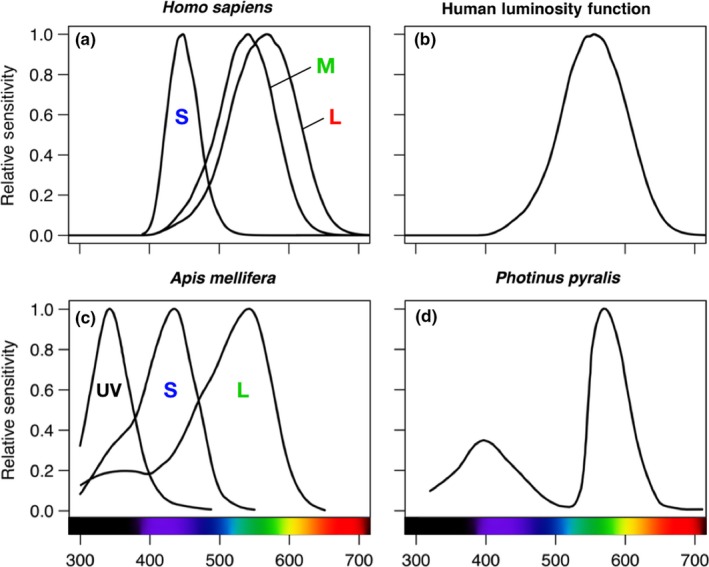
Spectral sensitivity in humans and insects. (a) Spectral sensitivities of blue (S, short‐wavelength), green (M, mid‐wavelength), and red (L, long‐wavelength) sensitive photoreceptors in humans presented as 10° cone fundamentals, calculated from Stiles and Burch (1959) color matching functions (Stockman & Sharpe, 2000; www.cvrl.org/). Humans are insensitive to UV wavelengths (<390 nm), perceive visible light wavelengths (390–700 nm) as color, and perceive infrared wavelengths (>700 nm) as heat. (b) Human luminosity function, used to predict relative “brightness” as perceived by humans. (c) Spectral sensitivity of the photoreceptors of honeybee workers, thought to have retained the trichromatic color vision of ancestral insects (Briscoe & Chittka, 2001; modified from Peitsch et al., 1992). (d) Electroretinography of a male Big Dipper firefly Photinus pyralis suggests that this species may have lost its short‐wavelength sensitive opsin (modified from Lall, Chapman, Ovid Trouth, & Holloway, 1980)
Ancestral insects likely possessed three types of photoreceptors or opsins (Briscoe & Chittka, 2001; Figure 2c): one sensitive to ultraviolet wavelengths (UV, 300–400 nm), another to short wavelengths (blue, 400–480 nm), and a third to long wavelengths (green to amber, 480–600 nm). Subsequent evolution into different ecological niches and light environments then selected for variation in spectral sensitivity (Endler, 1993; Lall, Seliger, Biggley, & Lloyd, 1980; Théry, Pincebourde, & Feer, 2008). In butterflies, dragonflies, and other diurnal insects, more visual opsins improve color discrimination (Briscoe, 2008; Feuda, Marlétaz, Bentley, & Holland, 2016; Futahashi et al., 2015; Lunau, 2014; Sharkey et al., 2017). In contrast, species occupying nocturnal and subterranean (aphotic) habitats often lose one or more opsins, reducing their capacity for color vision (reviewed in Feuda et al., 2016, Tierney et al., 2017; but see Kelber, Balkenius, & Warrant, 2003; Meyer‐Rochow, 2007; Somanathan, Borges, Warrant, & Kelber, 2008; White, Xu, Münch, Bennett, & Grable, 2008).
In addition to reduced spectral sensitivity, nocturnal insects often sacrifice spatial and temporal resolution in order to optimize total visual sensitivity under low‐light conditions (but see Kelber et al., 2011). Moths and some nocturnal beetles have evolved superposition eyes with rhabdoms that collect light from multiple facets, sacrificing resolution to gain up to 1,000× more sensitivity than apposition eyes of similar dimension (Cronin et al., 2014; Horridge, 1969). Some crepuscular bees, ants, and dung beetles have more recently transitioned to a nocturnal niche, and possess apposition eyes with larger lenses and wider rhabdoms that sum photons over time and/or space to increase sensitivity (Baird, Fernandez, Wcislo, & Warrant, 2015; Warrant & Dacke, 2011). Among dung beetles, larger eye size, smoother facets, and absence of screening pigment correlate to nocturnal activity (McIntyre & Caveney, 1998). Nocturnal bees, wasps, and ants have dorsal ocelli—simple eyes used to detect ambient light, movement, and/or orientation—that are significantly larger than those of their diurnal relatives (Narendra & Ribi, 2017; Somanathan, Kelber, Borges, Wallén, & Warrant, 2009; Warrant, Kelber, Wallén, & Wcislo, 2006). The ocelli of nocturnal bees and cockroaches are sensitive, but poor at perceiving fast movements; they also lack UV‐sensitive opsins, perhaps as an adaptation to their UV‐poor light environments (Berry, Wcislo, & Warrant, 2011); similarly, the ocelli of nocturnal ants may have lost polarization sensitivity in response to the loss of polarized skylight signals (Narendra & Ribi, 2017).
3. EFFECTS OF ALAN ON NOCTURNAL INSECTS
When considering the environmental impact of ALAN, it is vital to distinguish between astronomical and ecological light pollution (Longcore & Rich, 2004). The former, often referred to as “skyglow,” is the result of upwelling illumination of the night sky. Skyglow may spread to cover an area far beyond its origin and is not blocked by local terrain (Gaston et al., 2015). In contrast, ecological light pollution refers to the infiltration of point sources of light into habitats on the ground, which can affect local species without necessarily influencing night sky brightness. Although shielded light technology may ameliorate skyglow, this light may still impact local biodiversity. Similarly, important is the position of insects relative to light sources; downwelling light may affect species on the ground more than those in the air, while upwelling light (path lights, tree lights, etc.) is more likely to affect insects in flight.
To provide a comprehensive framework for understanding the ecological impacts of artificial light on nocturnal insects, we have organized the relevant literature into five categories, described below.
4. TEMPORAL DISORIENTATION
ALAN may cause temporal disorientation, desynchronization of organisms from their typical biorhythms. The vast majority of terrestrial species have circadian, circamensual, and/or circannual patterns of activity (foraging, reproduction, migration, etc.) that are synchronized to daily, monthly, and yearly light cycles, respectively. Specialized photoperiodic photoreceptors use external light signals (Zeitgebers or “time‐givers”) to entrain the internal clock to the outside environment (Numata, Miyazaki, & Ikeno, 2015; Numata, Shiga, & Morita, 1997). In nocturnal insects, daily emergence time and duration of feeding and courtship activity are dictated by internal clocks entrained by ambient light or temperature (Saunders, 2009; Tataroglu & Emery, 2014), while monthly or yearly cycles of eclosion, mating, and oviposition can be entrained by moonlight or day length cues (for a review of lunar entrainment, see Kronfeld‐Schor et al., 2013). If ALAN is sufficiently intense and/or sustained in time, and of a specific spectral composition, it can desynchronize the internal clock (reviewed by Saunders, 2012). For example, the pink bollworm Pectinophora gossypiella can be artificially entrained by monochromatic yellow light emitted by low‐pressure sodium (LPS) lamps (Pittendrigh & Minis, 1971), and diurnal Bombus terrestris bumblebees by UV illumination (Chittka , Stelzer, & Stanewsky, 2013). Desynchronization reduces reproductive fitness of Drosophila melanogaster fruit flies (Xu, DiAngelo, Hughes, Hogenesch, & Sehgal, 2011, see also McLay, Nagarajan‐Radha, Green, & Jones, 2018) and has the potential to disrupt vital biological processes in other taxa (Dominoni, Borniger, & Nelson, 2016; Gaston et al., 2013; Saunders, 2012).
Exposure to artificial light in the laboratory inhibits a variety of biological processes in nocturnal insects (e.g., Botha, Jones, & Hopkins, 2017), possibly due to a buildup of melatonin (Honnen, Johnston, & Monaghan, 2016; Jones, Durrant, Michaelides, & Green, 2015). In moths, constant light can inhibit female sex pheromone release (Fatzinger, 1973; Geffen, Groot, et al., 2015; Sower, Shorey, & Gaston, 1970), reduce male attraction (Geffen, Eck, et al., 2015), induce male sterility (Bebas, Cymborowski, & Giebultowicz, 2001; Giebultowicz, 2001; Riemann, Johnson, & Thorson, 1981), and disrupt female oviposition (Yamaoka & Hirao, 1981). Because ALAN rarely completely conceals natural day–night cycles (Longcore & Rich, 2004; but see Honnen et al., 2016), it is unclear how these findings apply to natural populations. If skyglow is sufficiently constant and intense, some animals may respond by adopting around‐the‐clock activity patterns, as polar animals do during seasons with continuous light (Bloch, Barnes, Gerkema, & Helm, 2013). Even low‐level ALAN increases perceived day length (Kyba, Ruhtz, Fischer, & Hölker, 2011a), and could thereby induce seasonal polyphenism in stink bugs (Niva & Takeda, 2003) and aphids (Hardie, 2010; Sanders et al., 2015), or alter the calling songs of katydids (Whitesell & Walker, 1978). Low‐level ALAN is known to accelerate development in a range of insect taxa (Kehoe, Cruse, Sanders, Gaston, & Veen, 2018, van Geffen, Grunsven, Ruijven, Berendse, & Veenendaal, 2014, but see Durrant, Botha, Green, & Jones, 2018), with varying effects on fitness.
ALAN has been shown to conceal monthly and seasonal regimes of lunar sky brightness in urban areas (Davies, Bennie, Inger, Ibarra, & Gaston, 2013; Kyba, Ruhtz, Fischer, & Hölker, 2011b), and this could affect circamensual and circatidal rhythms of certain insect species (but see Satoh, Yoshioka, & Numata, 2008). Some populations of the mayfly Povilla adusta eclose 2 days after the full moon, which provides increased visibility in which to court, copulate, and oviposit (Corbet, Sellick, & Willoughby, 1974). Intertidal midges Clunio marinus (Neumann, 1989) and Pontomyia oceana (Soong, Lee, & Chang, 2011) use moonlight cues to synchronize activities (e.g., eclosion, mating, and oviposition) during intertidal periods. The corn earworm Helicoverpa zea will not mate unless its eyes are dark‐adapted to ambient light below 0.05 lux, similar to that produced by a quarter moon (Agee, 1969). Artificial light also suppresses H. zea oviposition, perhaps because this behavior is synchronized to the lunar cycle (Nemfc, 1971). Masking of lunar cycles by ALAN may disrupt these vital circamensual rhythms.
In addition to disrupting natural rhythms, ALAN can desynchronize ecological interactions of mutualist species. Different species have evolved to use either day length and temperature as Zeitgebers (Bale et al., 2002; Jayatilaka, Narendra, Reid, Cooper, & Zeil, 2011). Because ALAN alters day length but not temperature, it can lead to ecological mismatches. For example, the nightly foraging activity of the nocturnal carpenter bee Xylocopa tranquebarica is triggered by twilight and coincides with the opening of night‐blooming flowers (Somanathan et al., 2008). By delaying the onset of foraging in this bee (a “phase shift”; Pittendrigh, 1993), ALAN may disrupt this vital pollination mutualism, especially if flower opening is entrained by ambient temperature instead of light (van Doorn & van Meeteren, 2003, see also Seymoure, 2018).
ALAN may also prolong the foraging activity of diurnal and/or crepuscular competitors, which generally become more active with increasing illumination (Kempinger, Dittmann, Rieger, & Helfrich‐Forster, 2009; Kronfeld‐Schor et al., 2013). Initiation of foraging activity in moths (Dreisig, 1980; Eaton, Tignor, & Holtzman, 1983; Riley, Reynolds, & Farmery, 1983) and crickets (Campbell, 1976) is triggered when ambient light intensity declines to a species‐specific level. Foraging activity of nocturnal and crepuscular bees is both initiated and inhibited by specific light levels (Dyer, 1985; Kelber et al., 2006). In dung beetle communities, temporal partitioning helps reduce competition. Each species is physiologically adapted to a particular temporal niche: Nocturnal dung beetles have larger eyes and greater body size, which reduces radiant heat loss on cold nights (Caveney, Scholtz, & McIntyre, 1995). Crepuscular insects that phase shift into nocturnal niches may experience cold stress and could be slow to adapt (Urbanski et al., 2012). In this way, the disconnect of temperature and light environment may further disrupt ecological interactions by shifting species into temporal niches in which they experience greater competition and/or lowered fitness.
5. SPATIAL DISORIENTATION
ALAN may also result in spatial disorientation, disrupting an organism's ability to navigate in three‐dimensional space. In nocturnal landscapes, the lack of visual information makes navigation difficult. The most consistently visible landmarks are the moon and stars, followed by the dim pattern of polarized moonlight produced by atmospheric filtering (Dacke, Byrne, Baird, Scholtz, & Warrant, 2011). These orientation cues vary predictably throughout the night and seasonally, and nocturnal insects often use the moon or stars to calculate navigational bearings (e.g., black carpenter ants, Klotz & Reid, 1993; earwigs, Ugolini & Chiussi, 1996; heart‐and‐dart moths, Baker, 1987; and harvester termites, Leuthold, Bruinsma, & Huis, 1976).
Sand hoppers use the moon to orient along beaches and are known to orient to artificial fiber optic moons in the laboratory (Ugolini, Boddi, Mercatelli, & Castellini, 2005). Other animals can navigate by stars alone (reviewed by Foster, Smolka, Nilsson, & Dacke, 2018). On moonless nights, Noctua pronuba yellow underwing moths orient with respect to the north star (Sotthibandhu & Baker, 1979), and Scarabaeus satyrus dung beetles use the Milky Way as a cue to guide themselves away from dung piles in a maximally efficient straight line (Dacke, Baird, Byrne, Scholtz, & Warrant, 2013). On moonlit nights, S. satyrus and its relative Scarabaeus zambesianus (Dacke, Byrne, Scholtz, & Warrant, 2004; Dacke, Nilsson, Scholtz, Byrne, & Warrant, 2003) navigate using polarized moonlight; other nocturnal insects, including wasps (Warrant et al., 2006), bees (Greiner, Cronin, Ribi, Wcislo, & Warrant, 2007; Warrant & Dacke, 2011), and crickets (Herzmann & Labhart, 1989), may do so as well. The bull ant Myrmecia pyriformis uses polarized moonlight in addition to visual landmarks in the surrounding terrain to navigate to and from its nest during nightly foraging trips (Narendra, Reid, & Raderschall, 2013; Reid, Narendra, Hemmi, & Zeil, 2011).
ALAN has potential to interfere with all these forms of nocturnal navigation in two ways: Ecological light pollution introduces new sources of light into the nocturnal landscape, which could be confused for the moon or stars (see Attraction for one example), while atmospheric light pollution reduces the visibility of existing cues. Skyglow dramatically reduces star visibility in urban areas (Falchi et al., 2016), and artificial lighting has been shown to disrupt polarization signals as well (Kyba et al., 2011b).
Some nocturnal insects possess highly sensitive eyes capable of color discrimination under minimal illumination, including the hawkmoth Deilephila elpenor (Kelber et al., 2003) and the nocturnal bee X. tranquebarica (Somanathan et al., 2008). Both X. tranquebarica and the nocturnal sweat bee M. genalis use optic flow and visual landmarks such as trees and flowers to navigate to and from their nests (Baird, Kreiss, Wcislo, Warrant, & Dacke, 2011; Warrant et al., 2004). Several nocturnal ants (Hölldobler & Taylor, 1983; Kaul & Kopteva, 1982; Klotz & Reid, 1993), bees (Warrant et al., 2006), and the shield bug Parastrachia japonensis (Hironaka, Inadomi, Nomakuchi, Filippi, & Hariyama, 2008) have been shown to navigate by canopy patterns, and the site‐specific shape of leaves and branches silhouetted against the bright night sky overhead. Urban skyglow associated with ALAN could increase this contrast, while artificial illumination beneath the canopy is likely to erase it entirely.
Nocturnal insects such as wasps and bees use their enlarged ocelli for navigation (Berry et al., 2011; Goodman, 1965), and direct illumination may cause navigation problems for these and other species. Because these insects adjust their wing angle so that the lighter half of their visual field is always overhead, upward‐directed illumination could cause a maladaptive response. Indeed, light‐reflecting mulching films are used in open crop fields to suppress the arrival of alate aphids, thrips, and whiteflies (Shimoda & Honda, 2013). Disoriented insects might wander into unsuitable habitat, and those without the ability to “beeline” back to their nests at the end of each night may fall prey to overheating, desiccation, or predation the next day.
6. ATTRACTION
Through positive phototaxis, many flying insects are attracted to ALAN (Verheijen, 1960). Some exhibit characteristic spiraling flight patterns, while others approach the light directly. Some orbit the light source, frequently changing their angular velocity and direction to remain within its vicinity (Muirhead‐Thompson, 1991), while others perch on or under the light, apparently stunned. Physiological and behavioral explanations of this phenomenon abound (see Nowinszky, 2004), and their explanatory power varies with species. The light compass theory (Baker & Sadovy, 1978; Sotthibandhu & Baker, 1979) suggests insects that orient themselves by maintaining a constant angle to light rays, historically emitted only by the moon or stars, will spiral into artificial light sources. Other theories involve the illusion of open sky (Goldsmith, 1990) or dark “Mach bands” at light–dark borders (Hsiao, 1973), or disorientation due to “dazzling” (Robinson, 1952, Verheijen, 1960, Hamdorf & Höglund, 1981; see Desensitization below).
Historically, light traps have been used by scientists to survey community composition, monitor beneficial insects (Nabli, Bailey, & Necibi, 1999), and control insect pest populations (e.g., Goretti, Coletti, Veroli, Giulio, & Gaino, 2011; Pawson, Watt, & Brockerhoff, 2009; Wallner & Baranchikov, 1992). The most common insect orders attracted to and captured in light traps are Diptera, Coleoptera, and Lepidoptera (Mikkola, 1972; van Grunsven et al., 2014; Wakefield et al., 2016). Light‐trapping equipment can differ from ALAN in important ways: Experimental light traps usually emit more short wavelengths, are often without glass shields (which filter UV), and are placed near the ground (Degen et al., 2016). However, experiments that vary the intensity and spectral composition of light traps can still offer insight into the potential effects of ALAN on positively phototactic insects. The impact of ALAN on negatively phototactic insects such as cockroaches and earwigs has not yet been well‐explored (Bruce‐White & Shardlow, 2011, but see Farnworth, Innes, Kelly, Littler, & Waas, 2018), despite a clear potential for adverse effects (see Siderhurst, James, & Bjostad, 2006).
Among the common positively phototactic insects, moths (Frank, 1988, 2006 ; MacGregor, Pocock, Fox, & Evans, 2015) and aquatic insects (Perkin, Hölker, & Tockner, 2014; Yoon, Kim, Kim, Jo, & Bae, 2010) are best studied. Comparative surveys have shown that, relative to their calculated visibility, short wavelengths are disproportionately attractive to many insects (Barghini & de Medeiros, 2012; Mikkola, 1972; see also Wakefield et al., 2016 for a discussion of infrared wavelengths). Although most insects can perceive short wavelengths (Briscoe & Chittka, 2001; Kelber & Roth, 2006), certain families of moths are more attracted to them than others (van Langevelde, Ettema, Donners, WallisDeVries, & Groenendijk, 2011; Somers‐Yeates et al., 2013, see also Wölfling, Becker, Uhl, Traub, & Fiedler, 2016). LPS lamps rarely attract moths (Plummer et al., 2016; Robinson, 1952; Rydell, 1992), even though most species can detect the yellow wavelengths they emit (Briscoe & Chittka, 2001; Mikkola, 1972). Some nocturnal insects are disproportionately attracted to polarized light sources as well (Danthanarayana & Dashper, 1986; see Recognition below).
About 30%–40% of insects that approach street lamps die soon thereafter (Eisenbeis, 2006), as a result of collision, overheating, dehydration, or predation (Minnaar, Boyles, Minnaar, Sole, & McKechnie, 2015; Yoon et al., 2010). The presence of foraging bats does not repel moths from ALAN sources (Acharya & Fenton, 1999), and under mercury vapor light, Operophtera brumata and O. fagata moths lacked their normal evasive responses to simulated ultrasonic bat signals (Svensson & Rydell, 1998). Depending on its placement, ALAN may also impede the movement of insects among habitat patches, lure them into bodies of water, or divert them into traffic (Frank, 2006). Insects not killed immediately may become trapped in a “light sink,” unable to forage (Langevelde, Grunsven, et al., 2017), search for mates, or reproduce—especially when different sexes are disproportionately attracted to ALAN, as is the case for many moth species (Altermatt, Baumeyer, & Ebert, 2009; Altermatt & Ebert, 2016; Degen et al., 2016; Frank, 1988; Garris & Snyder, 2010; see also Farnworth et al., 2018). Ecological traps that result in mortality or reproductive failure are predicted to lead to rapid population decline and ultimately extinction (Kokko & Sutherland, 2001; Robertson, Rehage, & Sih, 2013). Long‐term records confirm that positively phototactic macro‐moths (Langevelde, Braamburg‐Annegarn, et al., 2017) in lit habitats (Wilson et al., 2018) have undergone disproportionate declines in abundance over the past 50 years.
Perhaps due to selection, when newly eclosed moths from urban populations are tested under standardized conditions, they are less attracted to ALAN (Altermatt & Ebert, 2016). ALAN in urban settings may also be generally less attractive due to a reduction in background contrast (Frank, 2006), although one study comparing declines in macro‐moth abundance at light‐trap sites with and without artificial night sky brightness did not support this suggestion (Conrad, Warren, Fox, Parsons, & Woiwod, 2006, see also White, 2018).
7. DESENSITIZATION
The highly sensitive visual systems of nocturnal insects may not always function well in illuminated environments. Crepuscular and nocturnal Myrmecia ants are capable of flexible, rapid light adaptation (Narendra, Greiner, Ribi, & Zeil, 2016), and some even forage more effectively under illumination (Narendra et al., 2013). However, the photoreceptors of other insects, such as nocturnal flies, bees, and cockroaches, are saturated at modest light levels (see Honkanen et al., 2017). When exposed to too many photons at once, some insects may be temporarily dazzled or even permanently blinded (Stark, Walker, & Eidel, 1985). In Gryllus bimaculatus crickets, the photoreceptors of nocturnal adults show structural degeneration after exposure to bright UV light, while those of diurnal nymphs are less affected (Meyer‐Rochow, Kashiwagi, & Eguchi, 2002). If not bright enough to cause permanent damage, a discrete ALAN source may still temporarily blind a nocturnal insect to other orientation cues by inducing a rapid process of light adaptation (see Laughlin & Hardie, 1978), perhaps causing it to fly directly into said light (McGeachie, 1988; Robinson, 1952; Verheijen, 1960). In the hawkmoth D. elpenor, an 8‐s exposure to bright blue light can reduce visual sensitivity by two to three orders of magnitude in minutes (Hamdorf & Höglund, 1981); even 0.125 s of white light exposure can effectively blind cockroach ocelli for 15–20 s (Ruck, 1958). Should an affected insect escape into darkness, it may take hours for it to completely recover its original visual sensitivity (Bernhard & Ottoson, 1960). Light adaptation can also have behavioral consequences: When moths encounter yellow or green light above a certain brightness, light adaptation of their eyes suppresses nocturnal behaviors, including flying, foraging, and mating, a response that has been successfully used in pest control (Shimoda & Honda, 2013).
8. RECOGNITION
The effects of environmental illumination on the ability of nocturnal insects to recognize objects in their environment (conspecifics, predators, food plants, etc.) will depend on both the wavelength and intensity of the ALAN source under consideration. Nocturnal insects may be able to detect nearby objects more easily if the source emits (at sufficient intensity) wavelengths to which their visual systems are sensitive. However, some taxa may gain or lose their ability to discriminate colors, depending on the range of wavelengths emitted (Davies, Bennie, Inger, Ibarra, et al., 2013, but see Johnsen et al., 2006). As a result, ALAN has the potential to impede visual signaling and/or undermine camouflage (reviewed by Delhey & Peters, 2017). The body color of dusk‐active beetles is most apparent in a purplish light environment (Endler, 1993; Théry et al., 2008) and may become less visible to conspecifics under broad‐spectrum ALAN illumination. Similarly, the aposematic coloration of Heliconius butterfly wings, which is especially apparent in their typical light environments, may be obscured by ALAN (Seymoure, 2016). The UV emissions of mercury vapor lamps accentuate UV‐reflective markings on flowers and wings, which may benefit bees, moths, and other nocturnal insects sensitive to these signals (Kevan, Chittka, & Dyer, 2001). In contrast, illumination by LPS lamps could obscure these markers (Frank, 2006). Many aquatic insects use polarized light to locate suitable oviposition sites (reviewed by Horváth, Kriska, Malik, & Robertson, 2009, Perkin et al., 2014, Villalobos Jiménez, 2017). Artificial illumination of smooth dark surfaces such as asphalt simulates the polarization of light reflected off bodies of water, causing some aquatic insects to maladaptively oviposit on bridges or cars (Egri et al., 2017; Szaz et al., 2015), or to congregate on windows (Horváth et al., 2009; Kriska, Malik, Szivák, & Horváth, 2008).
Increased visibility is thought to benefit predators over prey (Kronfeld‐Schor et al., 2013; Youthed & Moran, 1969), but evidence of this phenomenon in arthropod systems is mixed (see Grenis, Tjossem, & Murphy, 2015; Skutelsky, 1996; Tigar & Osborne, 1999). By simulating bright moonlit nights every night of the month (Davies, Bennie, Inger, & Gaston, 2013), ALAN may reduce foraging time and increase the starvation risk of nocturnal prey insects (Schmitz, Beckerman, & O'Brien, 1997). Conversely, it may prolong the foraging activity of diurnal and crepuscular insects such as the sweat bee Lasioglossum texanum (Kerfoot, 1967), the desert ant Veromessor pergandei (Hunt, 1977), and the hemipteran Nilaparvata lugens (Riley, Reynolds, & Farrow, 1987), all of which engage in nocturnal foraging during the full moon.
9. COMMUNITY‐LEVEL IMPACTS
By altering the light environment experienced by nocturnal insects, ALAN can disrupt their temporal patterns, interfere with their spatial orientation, act as a fatal attraction, reduce their visual sensitivity, and alter foraging activity and species interactions. When populations of such abundant and ubiquitous organisms are disrupted (Gaston & Bennie, 2014; Kurvers & Hölker, 2015), entire communities will be affected (Davies et al., 2017; Davies, Bennie, Inger, Ibarra, et al., 2013; Sanders & Gaston, 2018). The population‐ and community‐level effects of ALAN have been relatively understudied, despite their great potential for use in predicting the future composition of artificially illuminated habitats; we summarize existing research below.
Insects that are attracted to ALAN sources are readily exploited by predators: Orb‐weaving spiders prefer artificially lit web sites (Enders, 1977), which may net them more prey (Heiling, 1999, but see Yuen & Bonebrake, 2017). Bats (Jung & Kalko, 2010; Minnaar et al., 2015; Rydell, 2006), birds (Robertson, Kriska, Horvath, & Horvath, 2010), and invasive cane toads (González‐Bernal, Greenlees, Brown, & Shine, 2016) congregate around streetlights and lit buildings for a similar reason. Usually, diurnal anole lizards and jumping spiders have been observed hunting for insects at night in artificially illuminated locations (Frank, 2009; Garber, 1978; Wolff, 1982). Moths frozen under illumination may provide stable search images that enable birds to recognize the camouflaged wing patterns of these species in other contexts (Frank, 2006). Decreases in moth abundance have a negative impact on nocturnal pollen transport (Fox, 2013; Frank, 1988; Knop et al., 2017; Macgregor et al., 2015; Macgregor, Evans, Fox, & Pocock, 2017), with cascading effects on populations of plants and insect herbivores.
Artificial illumination in urban areas may reduce the population persistence of nocturnal species by preventing movement between habitat patches (Farnworth et al., 2018; Gaston & Bennie, 2014; Guetté et al., 2018). For example, moths attempting to cross road networks were impeded by a line of closely spaced street lights (Degen et al., 2016). In a riparian ecosystem, emerging aquatic insects were drawn to artificially illuminated patches rather than the surrounding habitat, potentially reducing nutrient exchange and species dispersal (Manfrin et al., 2017). At the same time, illumination of the water may increase predation risk for invertivorous fish, reducing predation on aquatic insects and leading to locally increased insect abundance (Manfrin et al., 2017). Riparian predators respond to increased prey availability by congregating around light sources (Meyer & Sullivan, 2013; Perkin et al., 2011).
Several studies have noted an influx of predatory and scavenging arthropods into lit areas (Davies, Bennie, & Gaston, 2012; Šustek, 1999), though this response appears to be taxon‐specific (Eccard, Scheffler, Franke, & Hoffmann, 2018; Manfrin et al., 2017; Meyer & Sullivan, 2013; van Grunsven, Jähnichen, Grubisic, & Hölker, 2018). Broad‐spectrum LED lights in combination with urban heat reduced pea aphid populations by increasing visibility and lengthening the activity period of their visually oriented coccinellid predators (Miller et al., 2017); however, in similar experiments, bright illumination decreased or did not affect rates of parasitism by parasitoid wasps (Kehoe et al., 2018; Sanders et al., 2015, see Sanders, Kehoe, Cruse, Veen, & Gaston, 2018). The introduction of artificial light and noise causes parasitic frog‐biting midges to be unable to locate and feed from their túngara frog hosts (McMahon, Rohr, & Bernal, 2017). Herbivorous insects may additionally be affected by changes in their food plants (reviewed by Vänninen, Pinto, Nissinen, Johansen, & Shipp, 2010). For example, long‐wavelength ALAN reduced the population sizes of pea aphids by inhibiting flowering in their host plant via the phytochrome pathway (Bennie, Davies, Cruse, Inger, & Gaston, 2015), and light from high‐pressure sodium (HPS) lamps increased plant toughness and decreased the mass of cutworm larvae (Grenis & Murphy, 2018). The resultant absence of midges, pea aphids, and other prey insects in urban areas is likely to have widespread effects on populations of their host plants, pollinated plants, and predators.
10. EFFECTS OF ALAN ON BIOLUMINESCENT INSECTS
Firefly beetles (Coleoptera: Lampyridae) comprise the most widespread and diverse group of bioluminescent species on land. The approximately 2000 lampyrid species, all of which glow aposematically as larvae (Branham & Wenzel, 2003), enjoy a worldwide distribution. In many adult fireflies, either one or both sexes employ bioluminescence as a courtship signal (Lloyd, 2008). Other bioluminescent insect taxa, primarily click beetles (Coleoptera: Elateridae), railroad worms (Coleoptera: Phengodidae), and fungus gnats (Diptera: Keroplatidae), employ bioluminescence as an aposematic signal or predatory lure (Meyer‐Rochow, 2007, Redford, 1982). All of these essential signals may be masked by ALAN, which has been suggested as one of several factors contributing to a worldwide decline in firefly populations (Khoo, 2014; Lewis, 2016; Lloyd, 2006). Recent studies on bioluminescent ostracods (Gerrish, Morin, Rivers, & Patrawala, 2009) and fungus gnats (Merritt & Clarke, 2013; Mills, Popple, Veidt, & Merritt, 2016) have shown inhibitory effects of artificial light on signaling activity, raising similar concerns. In this section, we briefly discuss firefly visual ecology then use our framework to examine how ALAN may affect the courtship and reproductive success of these and other bioluminescent insects.
11. FIREFLY VISION AND BIOLUMINESCENCE
Bioluminescent fireflies employ a diverse range of courtship signaling systems as part of their sexual communication (Lewis, 2009; Takatsu, Minami, Tainaka, & Yoshimura, 2012). In some taxa, sedentary females produce long‐lasting glows that attract flying males, the latter sometimes incapable of producing light. In other taxa, including most North American species, both sexes use flash signals—discrete bursts of light—to communicate with potential mates. North American Photinus fireflies engage in courtship dialogs that involve precisely timed flash signals encoding species identity and sex. Typically, sedentary females respond to advertisement flashes emitted by flying males. In congregating South‐East Asian species, however, clusters of stationary males emit synchronous flashes to attract flying females.
Bioluminescence can be a highly efficient visual signal: against a black background, its contrast is effectively infinite, and the distances across which it can be perceived limited only by habitat structure and the visual sensitivity of the receiver (Cronin et al., 2014). However, fireflies do not always signal against a black background. While nocturnal species initiate courtship flashing long after nightfall, crepuscular species become active shortly after sunset (Lloyd, 1966). Some species flash in shady patches during the daytime (Vencl, Luan, Fu, & Maroja, 2017). The color of firefly bioluminescence varies as follows: Among crepuscular species, flashes are generally yellower compared to the greener flashes emitted by nocturnal fireflies (Seliger, Lall, Lloyd, & Biggley, 1982); this is thought to maximize signal contrast against the green foliage dominating the background at dusk. However, signal color also shows intraspecific variation, perhaps related to differences in habitat type (Hall, Sander, Pallansch, & Stanger‐Hall, 2016).
Due to their temporally restricted courtship activity periods, fireflies are highly sensitive to ambient light cues indicating time of day. When ambient light intensity descends to a certain species‐specific threshold, courtship signaling begins (Buck, 1937a; Table 1). Intracerebral ocelli described from Japanese Luciola cruciata and Luciola lateralis fireflies may assist in entraining this circadian behavior (Hariyama, 2000). Lampyris noctiluca females usually begin emitting courtship signals when ambient light levels fall below 1.3 lux and are never active above 15 lux; for Photuris congener males, these values are 0.25 and 2.5 lux, respectively, reflecting the shorter duration of twilight in their habitat (Dreisig, 1975). Ambient light intensity also affects other aspects of courtship behavior. In crepuscular Photinus species, males fly higher as light levels decline, and when passing under the shade of trees (Lewis & Wang, 1991; Lloyd, 2000).
Table 1.
Effect of ambient light on flash activity of various firefly speciesa
| Species | Flash (lux) | No flash (lux) | Location | References |
|---|---|---|---|---|
| Aspisoma lineatum | 0.85 | Southeast Region, Brazil | Hagen and Viviani (2009) | |
| >0.05 | Hagen et al. (2015) | |||
| Apisoma physonotum | <0.2 | Hagen and Viviani (2009) | ||
| >0.05 | Hagen et al. (2015) | |||
| Apisoma sp2 | <0.2 | Hagen and Viviani (2009) | ||
| Apisoma sp4 | <0.2 | Hagen and Viviani (2009) | ||
| Bicellonychia lividpennis | 4.5 | Hagen and Viviani (2009) | ||
| Bicellonychia ornaticollis | <0.2 | Hagen and Viviani (2009) | ||
| Cratomorphus concolor | <0.2 | Hagen and Viviani (2009) | ||
| Cratomorphus sp4 | <0.2 | Hagen and Viviani (2009) | ||
| Lampyris noctiluca (♀) | 1.3 | 10 | Ebeltoft, Denmark | Dreisig (1975) |
| 0.28 | Dreisig (1975) | |||
| Lampyris noctiluca (larva) | 6.85 | Dreisig (1975) | ||
| Luciola italica | 0.25 | 0.47 ± 0.26 | Turin, Italy | Picchi, Avolio, Azzani, Brombin and Camerini (2013) |
| Photinus interdius (diurnal) | 339.82 ± 88 | >1,000 | Darien Province, Panama | Vencl, Luan, Fu, and Maroja (2017) |
| Photinus pyralis | 210–320 | Laboratory | Buck (1937a) | |
| 301.24 ± 89.07 | Clarke County, Virginia, USA | Firebaugh and Haynes (2016) | ||
| Photinus sp1 | <0.2 | >0.234 | Southeast Region, Brazil | Hagen et al. (2015) |
| >1.5 | Hagen and Viviani (2009) | |||
| Photinus spp. | 1.2 | Piedmont Region, Maryland, USA | Costin and Boulton (2016) | |
| Photinus umbratus | 1 | Highlands County, Florida, USA | Dreisig (1975) | |
| 4 | Dreisig (1975) | |||
| Photuris "A" | 0.38 | Dreisig (1975) | ||
| Photuris congener | 0.25 | 2.5 | Dreisig (1975) | |
| Photuris pennsylvanica | 160,000 | Laboratory | Harvey (1926) | |
| Photuris missouriensis | 3,800 | Laboratory | Case and Trinkle (1968) | |
| Photuris versicolor (♀) | Highlands County, Florida, USA | Dreisig (1975) | ||
| 301.24 ± 89.07 | Clarke County, Virginia, USA | Firebaugh and Haynes (2016) | ||
| Pteroptyx maipo | 0.2–0.3 | Tin Shui Wai, Hong Kong | Yiu (2012) | |
| Pteroptyx valida | 7–14 | Samut Prakan Province, Thailand | Prasertkul (2018) | |
| Pyrogaster moestus | >0.05 | Southeast Region, Brazil | Hagen et al. (2015) | |
| Pyrogaster sp1 | <0.2 | Hagen and Viviani (2009) |
“Flash” column gives ambient light levels shown to be dim enough to induce bioluminescence for each species; “No flash” gives ambient light levels shown to inhibit firefly flash activity.
The superposition compound eyes of fireflies are finely attuned to conspecific signals: They are highly sensitive, but only to a narrow range of wavelengths. To date, only UV‐ and long‐wavelength‐sensitive (UVS and LWS) opsin genes have been isolated from fireflies (Martin, Lord, Branham, & Bybee, 2015; Sander & Hall, 2015), although blue sensitivity has been observed in both North American and Japanese species using electroretinography (ERG) recordings (Eguchi, Nemoto, Meyer‐Rochow, & Ohba, 1984; Lall, Lord, & Ovid Trouth, 1982; Lall, Strother, Cronin, & Seliger, 1988). The peak sensitivity of the LWS photoreceptor generally corresponds to the peak wavelength of signals produced by that species (Sander & Hall, 2015). Signal reception is further tuned using filter pigments: In species that emit yellow bioluminescence, reddish filter pigments narrow visual sensitivity to the yellow region of the spectrum by blocking out green wavelengths (Booth, Stewart, & Osorio, 2004; Cronin, Järvilehto, Weckström, & Lall, 2000). This screening could allow some lampyrids to communicate visually even within light‐polluted habitats. However, current evidence suggests that ALAN does have a demonstrable impact on firefly signaling (Table 2).
Table 2.
Effect size of experimental studies examining impact of ALAN on firefly courtship
| Species | Treatment (ALAN type) | Metric | Intensity (lux) | Effect size (G's Δ) | Study |
|---|---|---|---|---|---|
| Lampyris noctiluca | High‐pressure sodium street lamps | Green LED lure (# of trapped males) | L1: 46–64 | L1: −0.74 | Ineichen and Rüttimann (2012) |
| L2: 0.1–0.4 | L2: −0.97 | ||||
| Lampyris noctiluca | Incandescent flashlight | Green LED lure (# of trapped males) | L1: 0.3 | L1: −0.37 | Bird and Parker (2014) |
| L2: 0.18 | L2: −0.37 | ||||
| L3: 0.09 | L3: −0.23 | ||||
| L4: 0.07 | L4: −0.02 | ||||
| Photinus sp1 | Multi‐metal vapor floodlights | Transect count (# of flashing individuals) | T1: 4.45 | T1: −0.25 | Hagen et al. (2015) |
| T2: 1.5 | T2: −0.22 | ||||
| T3: 0.05 | T3: −0.17 | ||||
| Photuris versicolor | White LED floodlights | Flash count (flashes/min) | 301 at plot center | −0.459 | Firebaugh and Haynes (2016) |
| Photinus pyralis | Flash count (flashes/min) | 301 at plot center | NS | ||
| Flash count, tethered females (15 min total) | 167.21 on average | −0.617 | |||
| Photinus spp. | Mercury vapor bulb | Flash count (flashes/min) | 1.2 at plot edge | −0.653 | Costin and Boulton (2016) |
Because all studies involve comparison of groups of the same size (e.g., firefly populations before and after ALAN exposure), Glass's Δ is an appropriate estimate of effect size, as it uses only the standard deviation of the control group (Kline, 2013). NS indicates that no significant effect on firefly courtship activity was observed.
Studies in urban and rural regions in São Paulo, Brazil, have found that several lampyrid species are limited to areas with ambient light levels below 0.2 lux, approximately equivalent to that of the night sky during a full moon (Hagen & Viviani, 2009; Viviani, Rocha, & Hagen, 2010). However, the crepuscular species Aspisoma lineatum and Bicellonychia lividipennis were found signaling near sodium vapor lamps, in areas with illumination measuring 0.85 and 4.5 lux, respectively; note that B. lividipennis emerges early in the evening when ambient light levels (without ALAN) reach about 4.5 lux. Similarly, surveys of Luciola italica in Turin, Italy, have found populations concentrated in dimly illuminated areas, with a negative correlation between firefly abundance and certain indices of urbanization (Picchi et al., 2013). In descriptive studies such as these, the effects of ALAN are confounded by many other variables associated with urbanization. It is also unclear whether population loss occurs due to movement away from illuminated habitats or to reduced reproductive success within urban populations.
Below, we summarize all available evidence concerning the impact of ALAN on fireflies, organized according to the five categories of impact described earlier. Many of these findings may be applicable to all bioluminescent insects, although data on other taxa are sparse. Should these insects be unable to cope with the following challenges, their survivorship, foraging success, and mating success—and therefore population persistence—will be reduced.
11.1. Temporal disorientation
ALAN has the potential to disrupt both larval and adult activity cycles, but little is known about the magnitude of this impact. Pyractomena borealis larvae kept indoors under continuous light pupated several months earlier than anticipated (Lloyd, 2006), but whether this occurs in natural populations remains unclear. Although adult fireflies rarely live long enough to express circamensual rhythms of activity, L. noctiluca larvae in the field have been shown to hide during the full moon (Gunn & Gunn, 2012), and may reduce their foraging activity if night sky brightness is increased by skyglow.
Nightly onset of courtship activity by adult fireflies is determined by ambient light intensity. P. pyralis adults begin flashing earlier in the evening on cloudy days (Buck, 1937a), although the opposite is predicted for urban areas, where reflection of ALAN makes the sky brighter on cloudy days than on clear ones (Kyba et al., 2011a). In light‐polluted habitats, the flight periods of crepuscular species might be extended into those normally occupied by nocturnal species. If males or females are responsive to heterospecific signals, this could impact their reproductive success by reducing dialog efficiency and/or increasing the frequency of heterospecific matings. Nocturnal Photuris fireflies are a major predator of other firefly species (Eisner, Goetz, Hill, Smedley, & Meinwald, 1997; Eisner, Wiemer, Haynes, & Meinwald, 1978; Lloyd, 1997). If ALAN causes crepuscular species to extend their courtship into the nocturnal niche occupied by Photuris, they may experience higher predation rates. It has been posited that, over evolutionary time, predation by nocturnal Photuris fireflies drove some fireflies into crepuscular and diurnal niches (Deyrup et al., 2017; Gronquist et al., 2006).
11.2. Spatial disorientation
ALAN may disorient fireflies that navigate with respect to the sun or moon, such as the larvae of P. borealis, which choose an aerial pupation site located on the southern side of trees to maximize their exposure to direct sunlight (Gentry, 2003). However, little is known about the balance of positive and negative phototaxis in fireflies. In Photinus fireflies, males produce courtship signals while searching within their habitat, and immediately orient toward a female when they detect her response flash. Because these fireflies do not appear capable of distinguishing between small signals that are nearby and large signals farther away (Cratsley & Lewis, 2003), they could possibly mistake artificial lights with certain emission spectra for receptive conspecifics (see below).
11.3. Attraction
In the European glowworm L. noctiluca, flying non‐bioluminescent males are attracted to larviform females that glow steadily from perches on raised display sites. Numerous experimental studies conducted with this species have employed glowing light lures to elucidate their signaling system (e.g., Bird & Parker, 2014; Mikkola, 1972; Schwalb, 1961). By measuring the attraction of males to different LEDs or chemiluminescent lures, these studies have described male preferences for the color and spatial pattern of female glow signals, and have demonstrated male attraction to light traps that emit yellow‐green light (reviewed by De Cock, 2009).
Although L. noctiluca is geographically widespread in Europe, its populations have been declining for some time (Gardiner, 2011). One potential contributing factor could be that males are more attracted to certain point sources of ALAN than they are to conspecific female glow signals. In North Wales, Bek (2015) reported significant attraction of male L. noctiluca to LPS street lamps, compared to HPS, LED, or unlit lamps: The vast majority of males (556 of 564) were found under LPS lamps, while none were found below LED lamps or those that had been switched off. A notable difference between LPS and LED bulbs is that the latter emit a large percentage of their light in blue wavelengths; binary choice experiments on L. noctiluca suggest that the addition of blue wavelengths significantly decreases male attraction to light lures (Booth et al., 2004).
Though L. noctiluca is not representative of all firefly species, other glowworms may also be attracted to artificial lights. In Uganda, Bowden and Church (1973) found multiple Lamprigera specimens caught in a Robinson trap illuminated by a mercury vapor bulb. In Rwanda, Diaphanes males were collected from a light trap emitting red light (Pacheco, Martin, & Bybee, 2016). Pleotomus and Pleotomodes glowworms are also attracted to light traps, including those exclusively emitting UV wavelengths (Faust, 2017; Lloyd, 2006).
Beyond glowworms, other firefly species could also be attracted to artificial light sources. Male Pteroptyx fireflies in South‐East Asia congregate in particular display trees and flash together en masse, and may be attracted to flashing string lights imitating these mating congregations (Cratsley, Prasertkul, & Thancharoen, 2012; Thancharoen, Srinual, & Laksanawimol, 2017). If these light sources draw individuals away from traditional display sites or differentially attract males and females, they could disrupt Pteroptyx courtship and reduce mating success.
11.4. Desensitization
Due to their highly sensitive visual systems (Horridge, 1969), fireflies may be vulnerable to blinding by bright ALAN sources. Numerous previous studies have used ERG recordings to measure firefly spectral sensitivity, by observing the level of electrical activity in the eye triggered by exposure to small point sources of light (e.g., Lall, 1981; Lall et al., 1982, 1988; Lall, Chapman, et al., 1980; Lall & Lloyd, 1989). These studies show that the compound eyes of Photinus fireflies can take several hours to become fully dark‐adapted (Lall, 1993); intermittent light exposures may decrease resulting gains in visual sensitivity, but the duration and intensity of exposure sufficient to cause a complete reversal to the light‐adapted state, let alone to cause dazzling and/or blinding, have yet to be investigated (A. Lall, pers. comm.). Nonetheless, it appears likely that bright sources of ALAN such as LED street lamps could at minimum slow the dark adaptation process. This may disproportionately diminish long‐wavelength sensitivity in female fireflies (Oba & Kainuma, 2009), reducing their ability to recognize potential mates.
11.5. Recognition
While some background illumination is often present in their signaling milieu, especially for crepuscular firefly species, artificial illumination could reduce courtship success by interfering with the perception of male signals by receptive females, or vice‐versa. In addition, females might be less responsive to male signals that they perceive as dimmer under ALAN. Lloyd (2006) suggested that monochromatic yellow light from LPS lamps might disproportionately impact the signal exchange of yellow‐flashing (crepuscular) species. Because broad‐spectrum illumination from white LED street lamps includes many wavelengths, these ALAN sources may obscure the bioluminescent signals of green‐flashing (nocturnal) firefly species as well.
Several recent field studies have examined how ALAN affects firefly flash activity and courtship behavior (Table 2). Ineichen and Rüttimann (2012) assessed the impact of HPS street lighting on L. noctiluca males in suburban Switzerland. They compared male attraction to LED lures simulating the glows of conspecific females, placed either under (46–64 lux) or between (0.1–0.4 lux) HPS street lamps. Significantly, more males were attracted to lures in darker locations between the HPS street lamps. Perhaps males were unable to detect the lures in the highly illuminated surroundings, or perhaps males prefer signals with greater contrast against the background (Hopkins, Baudry, Candolin, & Kaitala, 2015). In either case, these results suggest HPS street lighting interferes with L. noctiluca courtship communication. Ineichen and Rüttimann (2012) noted that female display sites appeared to be uniformly distributed with respect to street lighting. L. noctiluca females select their display sites during daytime as larvae and rarely relocate in their flightless adult form (Tyler, 2013). Thus, females located underneath ALAN sources are likely to experience lower mating success.
In another observational study on a university campus in Sorocaba, Brazil, Hagen, Santos, Schlindwein, and Viviani (2015) found that courtship flashing by several firefly species was affected by multi‐metal vapor spotlights illuminating an outdoor sport court. On nights when the spotlights were turned on, flash activity by the most common species, Photinus sp1, was significantly reduced. ALAN reduced the number of flashing individuals encountered along transects that were both directly and indirectly illuminated by the spotlights; Photinus sp1 flashed only below measured light intensities of 0.23 lux. While this and other studies clearly demonstrate that ALAN reduces flash activity (Table 1), its impact on firefly mating success remains unknown.
Experimental field studies of ALAN are useful for removing confounding variables associated with urbanization. Working in an undisturbed British chalk grassland, Bird and Parker (2014) introduced a point source of light and then measured attraction of L. noctiluca males to glowing LED lures. Lures were placed 0.5, 1.0, 1.5, and 2 m from an upward‐directed filament bulb flashlight (its emission spectra resembling that of a mercury vapor lamp) with measured intensities of 0.3–0.07 lux. Compared to a non‐illuminated control, levels of ALAN as low as 0.09 lux significantly reduced the number of males attracted to simulated female glows. When ambient illumination was brighter than 0.18 lux, none of the males approached the simulated females.
Although both studies showed that ALAN can interfere with the ability of male L. noctiluca to locate females, Bird and Parker (2014) found effects at much lower ALAN levels compared to Ineichen and Rüttimann (2012). Several explanations are possible. Bird and Parker attributed this difference to the orientation of the ALAN source, suggesting that upward‐directed light is more likely to dazzle males or reduce trap visibility than downward‐directed street lamps. The studies also differed in ALAN type as well as design of the female lures. Furthermore, urban and suburban firefly populations may have adapted to cope with higher levels of ALAN. Male choice is another potential factor: In both studies, males chose the comparatively brightest lures of those presented.
Firebaugh and Haynes (2016) conducted an experimental study of the impact of ALAN on the flash activity of both male and female fireflies at a rural site in Virginia, USA. They established four pairs of 20‐m‐diameter plots, placing a downward‐facing LED floodlight (301 lux at plot center) at the center of each plot. The flash activity of Photuris versicolor, a nocturnal species, declined significantly in the illuminated plots compared to non‐illuminated control plots. However, no difference in flash activity was detected for males of P. pyralis, a crepuscular species. In addition, ALAN did not alter the timing of the nightly onset of P. pyralis flash activity.
In a separate experiment, the authors tethered P. pyralis females to display platforms and observed the impact of LED floodlights (approximately 167 lux) on male approach frequency and flash count, as well as on the frequency of female response flashes. Although ALAN again had no significant effect on the flash activity of P. pyralis males, it significantly reduced the female response rate. This differential effect on male and female flash behavior could be due to the downward directionality of the ALAN source. Flying males may be able to detect a female flashing below them, even against more brightly illuminated vegetation. However, females perch close to the ground and look upward to detect males, typically against a dark sky. Therefore, the females may have been unable to detect male signals viewed against the downward‐facing ALAN. Alternatively, since P. pyralis females are known to prefer brighter simulated flashes (Vencl & Carlson, 1998), they may be less responsive to male signals that, when set against an illuminated background, appear less bright.
In an experimental field study in rural Maryland, USA, Costin and Boulton (2016) introduced ALAN using a mercury vapor bulb (1.2 lux at plot edge). At each of the six sites starting 30 min after sunset, they counted firefly flashes for 30 min one night, then added ALAN, and repeated the count the next night. On nights when ALAN was introduced, the authors found a significant decline in flash rates. Multiple species were present at each site, with the crepuscular Photinus marginellus most frequently observed. In contrast, Firebaugh and Haynes (2016) found no effect of ALAN on flash activity of P. pyralis. The discrepancy between these studies could reflect differing emission spectra of their artificial light sources, and/or differences between firefly species. Although both are crepuscular, P. marginellus emerges slightly later in the evening (Lloyd, 1966) and might be more sensitive to ALAN than P. pyralis. Because control counts were always made on the night preceding ALAN treatment at each site, the results may also reflect temporal changes in firefly abundance.
Yiu (2012) performed a similar experiment on synchronously flashing males of a Hong Kong firefly Pteroptyx maipo, introducing a compact fluorescent lamp for several minute intervals and counting flashes before and during exposure. ALAN illumination (0.21–2.0 lux) significantly decreased the average flash frequency of this species, and these decreases were reversed when the lamp was switched off. However, during a survey of Pteroptyx malaccae and Pteroptyx valida in Thailand, Prasertkul (2018) found robust congregations of both species flashing within close proximity to white fluorescent street lamps (up to 7–14 lux), suggesting that ALAN does not prevent aggregation or courtship flashing in these species.
In contrast to the complex light environments and shifting conditions typical of field experiments, laboratory experiments allow for more precise manipulation of ambient light levels. Early light exposure experiments conducted in the laboratory demonstrated that sufficiently bright light (from 50 to 1,000 lux) completely inhibits bioluminescent signaling in several firefly species (Table 1). More recently, Thancharoen (2007) exposed pairs of the Thai firefly Sclerotia (formerly Luciola) aquatilis to different intensities of fluorescent lighting (0.05, 0.1, 0.2, and 0.3 lux) and observed their mating behavior. Artificial illumination prolonged courtship, mounting, and mating duration in this species. Though all pairs ultimately mated successfully in the laboratory, ALAN could reduce mating success under field conditions by increasing the difficulty of locating a mate.
Using monochromatic LEDs, Owens, Meyer‐Rochow and Yang (2018) investigated the impact of ambient light on the alarm flash behavior of an aquatic Taiwanese firefly, Aquatica ficta. Short‐wavelength light (444–533 nm) caused males to flash more brightly, but less frequently, while long‐wavelength light (597–663 nm) had no significant effect on alarm flash behavior. These results indicate that fireflies can respond to increased background illumination by producing light signals of greater intensity. These results also suggest that, at least for this species, long‐wavelength artificial light (>597 nm, amber to red) is less disruptive than short‐wavelength (444–533 nm, violet to green) ALAN (but see Buck, 1937b, Pacheco et al., 2016).
12. CONCLUSIONS
Widespread nocturnal artificial illumination radically disrupts the habitats of night‐active species. Nocturnal and crepuscular insects are abundant and important components of these ecosystems. Thus, the impact of ALAN on insect fitness and abundance can provide a useful metric of overall ecosystem disturbance. The potential effects of ALAN on insects can be categorized as temporal disorientation, spatial disorientation, attraction, desensitization, and recognition. The severity of impact will depend on the degree of overlap between the spectral sensitivity of the insect in question and spectral emission (and intensity) of the particular ALAN source (Gaston et al., 2015). Recently, many urban areas have begun to phase out monochromatic long‐wavelength LPS lamps in favor of broad‐spectrum white LED lighting. This spectral shift represents an ecological experiment on a global scale, with potentially devastating results.
One recommendation is of paramount importance: future studies concerning the impact of ALAN on nocturnal organisms should employ more objective, less anthropocentric methods for measuring light. Light meter measurements in lux are adjusted for human luminous sensitivity and can be greatly affected by differences in distance from and angle to a light source. Insects often occupy discrete microhabitats within larger light environments. Therefore, researchers should strive to objectively measure how light varies within, as well as among, nocturnal habitats.
Among night‐active insects, bioluminescent fireflies are conspicuous and charismatic flagship species that can attract support for conservation efforts aimed at minimizing excess nocturnal illumination in urban areas. As a result, recent studies conducted in several geographic locations have focused on the impact of ALAN on fireflies. Laboratory and field studies reviewed here demonstrate that ambient light can inhibit the courtship flashing of several firefly species. ALAN reduced courtship signaling by both sexes in a North American firefly and reduced the ability of male European glowworms to locate females. Laboratory studies testing the effects of monochromatic light found that an Asian firefly is capable of increasing its flash intensity in response to ALAN and that long wavelengths (amber to red) were least disruptive.
This review also identifies several key gaps in our knowledge concerning the impacts of ALAN on fireflies, and highlights several important directions for future research. We currently know little about temporal disorientation or desensitization by ALAN, and these effects deserve further study. For example, comparative studies of firefly phenology and nightly emergence times could reveal the degree to which ALAN delays and/or shortens the temporal scope of courtship activity. Furthermore, despite evidence from several firefly species that ALAN interferes with mate location, it remains to be seen whether this disruption has consequences for mating and reproductive success. We also need information on how these behavioral impacts translate into longer‐term effects on population size and persistence.
Fireflies may be able to successfully cope with ALAN in various ways, including increased dispersal or evolutionary adaptation. Can fireflies move away from artificial lights toward darker habitat? We currently know little about dispersal abilities in either the adult or larval stages for any firefly species. While evidence suggests that the colors of firefly bioluminescence are under selection to maximize signal contrast in different natural light environments (Hall et al., 2016; Lall, Seliger, et al., 1980), whether ALAN selects for genetic changes in firefly signals remains unknown. Comparison of urban and rural populations may provide insight into adaptations that allow fireflies to cope with high levels of ALAN.
It will also be important to investigate how different intensities and colors of ALAN affect the courtship signaling of different firefly species. While physiological studies (e.g., ERGs) have revealed variation among species in their spectral sensitivity, the impact of different wavelengths on courtship success has yet to be determined. One particularly important aspect of this issue is understanding how the blue wavelengths emitted by commercial LED street lamps influence courtship signals and mate location across different firefly species. This research can help guide development of new lighting technology that balances the need for public safety, energy efficiency, and conservation, and inform policy recommendations for firefly‐friendly ALAN sources that can be deployed on public, commercial, and private lands in or near firefly habitat.
CONFLICT OF INTEREST
None declared.
AUTHORS CONTRIBUTION
ACSO conceived and drafted the first version of this manuscript and performed all data analyses. Both authors contributed significantly to revisions and have given final approval of the version to be published.
DATA ACCESSIBILITY
Effect size calculations: Dryad https://doi.org/10.5061/dryad.jm8ps6b.
ACKNOWLEDGMENTS
We thank Tufts University and the Zoological Lighting Institute for supporting this work.
Owens ACS, Lewis SM. The impact of artificial light at night on nocturnal insects: A review and synthesis. Ecol Evol. 2018;8:11337–11358. 10.1002/ece3.4557
REFERENCES
- Acharya, L. , & Fenton, M. B. (1999). Bat attacks and moth defensive behaviour around street lights. Canadian Journal of Zoology, 77, 27–33. 10.1139/z98-202 [DOI] [Google Scholar]
- Agee, H. R. (1969). Mating behavior of bollworm moths. Annals of the Entomological Society of America, 62, 1120–1122. [DOI] [PubMed] [Google Scholar]
- Altermatt, F. , Baumeyer, A. , & Ebert, D. (2009). Experimental evidence for male biased flight‐to‐light behavior in two moth species. Entomologia Experimentalis Et Applicata, 130, 259–265. 10.1111/j.1570-7458.2008.00817.x [DOI] [Google Scholar]
- Altermatt, F. , & Ebert, D. (2016). Reduced flight‐to‐light behaviour of moth populations exposed to long‐term urban light pollution. Biology Letters, 12, 20160111 10.1098/rsbl.2016.0111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- American Medical Association . (2016). Human and Environmental Effects of Light Emitting Diode (LED) Community Lighting. House of Delegates.
- Baird, E. , Fernandez, D. C. , Wcislo, W. T. , & Warrant, E. J. (2015). Flight control and landing precision in the nocturnal bee Megalopta is robust to large changes in light intensity. Frontiers in Physiology, 6, 305 10.3389/fphys.2015.00305 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baird, E. , Kreiss, E. , Wcislo, W. , Warrant, E. , & Dacke, M. (2011). Nocturnal insects use optic flow for flight control. Biology Letters, 7, 499–501. 10.1098/rsbl.2010.1205 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baker, R. (1987). Integrated use of moon and magnetic compasses by the heart‐and‐dart moth, Agrotis exclamationis . Animal Behaviour, 35, 94–101. 10.1016/S0003-3472(87)80214-2 [DOI] [Google Scholar]
- Baker, R. , & Sadovy, Y. (1978). The distance and nature of the light‐trap response of moths. Nature, 276, 818–821. 10.1038/276818a0 [DOI] [Google Scholar]
- Bale, J. S. , Masters, G. J. , Hodkinson, I. D. , Bale, J. S. , Masters, G. J. , Hodkinson, I. D. , … Whittaker, J. B. (2002). Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Global Change Biology, 8, 1–16. 10.1046/j.1365-2486.2002.00451.x [DOI] [Google Scholar]
- Barghini, A. , & de Medeiros, B. A. S. (2012). UV radiation as an attractor for insects. LEUKOS, 9, 47–56. [Google Scholar]
- Bebas, P. , Cymborowski, B. , & Giebultowicz, J. M. (2001). Circadian rhythm of sperm release in males of the cotton leafworm, Spodoptera littoralis: In vivo and in vitro studies. Journal of Insect Physiology, 47, 859–866. 10.1016/S0022-1910(01)00058-0 [DOI] [Google Scholar]
- Bek, R. J. (2015). Investigating the impact of artificial night lighting on the common European glow‐worm, Lampyris noctiluca (L.) (Coleoptera: Lampyridae) (ed Altringham J). BSc, University of Leeds.
- Bennie, J. , Davies, T. W. , Cruse, D. , & Gaston, K. J. (2016). Ecological effects of artificial light at night on wild plants. Journal of Ecology, 104, 611–620. 10.1111/1365-2745.12551 [DOI] [Google Scholar]
- Bennie, J. , Davies, T. W. , Cruse, D. , Inger, R. , & Gaston, K. J. (2015). Cascading effects of artificial light at night: resource-mediated control of herbivores in a grassland ecosystem. Philosophical transactions of the Royal Society of London. Series B, Biological sciences, 370, 20140131 10.1098/rstb.2014.0131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennie, J. , Davies, T. W. , Duffy, J. P. , Inger, R. , & Gaston, K. J. (2014). Contrasting trends in light pollution across Europe based on satellite observed night time lights. Scientific Reports, 4, 3789 10.1038/srep03789 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernhard, C. G. , & Ottoson, D. (1960). Comparative studies on dark adaptation in the compound eyes of nocturnal and diurnal Lepidoptera. The Journal of General Physiology, 44(1), 195–203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berry, R. P. , Wcislo, W. T. , & Warrant, E. J. (2011). Ocellar adaptations for dim light vision in a nocturnal bee. Journal of Experimental Biology, 214, 1283–1293. 10.1242/jeb.050427 [DOI] [PubMed] [Google Scholar]
- Bird, S. , & Parker, J. (2014). Low levels of light pollution may block the ability of male glow‐worms (Lampyris noctiluca L.) to locate females. Journal of Insect Conservation, 18, 737–743. 10.1007/s10841-014-9664-2 [DOI] [Google Scholar]
- Bloch, G. , Barnes, B. M. , Gerkema, M. P. , & Helm, B. (2013). Animal activity around the clock with no overt circadian rhythms: Patterns, mechanisms and adaptive value. Proceedings of the Royal Society of London B, 280, 20130019 10.1098/rspb.2013.0019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Booth, D. , Stewart, A. J. A. , & Osorio, D. (2004). Colour vision in the glow‐worm Lampyris noctiluca (L.) (Coleoptera: Lampyridae): Evidence for a green‐blue chromatic mechanism. Journal of Experimental Biology, 207, 2373–2378. 10.1242/jeb.01044 [DOI] [PubMed] [Google Scholar]
- Botha, L. M. , Jones, T. M. , & Hopkins, G. R. (2017). Effects of lifetime exposure to artificial light at night on cricket (Teleogryllus commodus) courtship and mating behaviour. Animal Behaviour, 129, 181–188. 10.1016/j.anbehav.2017.05.020 [DOI] [Google Scholar]
- Bowden, J. , & Church, B. M. (1973). The influence of moonlight on catches of insects in light‐traps in Africa. Part II. The effect of moon phase on light‐trap catches. Bulletin of Entomological Research, 63, 129–142. 10.1017/S0007485300050938 [DOI] [Google Scholar]
- Branham, M. A. , & Wenzel, J. W. (2003). The origin of photic behavior and the evolution of sexual communication in fireflies (Coleoptera: Lampyridae). Cladistics, 19, 1–22. 10.1111/j.1096-0031.2003.tb00404.x [DOI] [PubMed] [Google Scholar]
- Briscoe, A. D. (2008). Reconstructing the ancestral butterfly eye: Focus on the opsins. Journal of Experimental Biology, 211, 1805–1813. 10.1242/jeb.013045 [DOI] [PubMed] [Google Scholar]
- Briscoe, A. D. , & Chittka, L. (2001). The evolution of color vision in insects. Annual Review of Entomology, 46, 471–510. [DOI] [PubMed] [Google Scholar]
- Bruce‐White, C. , & Shardlow, M. (2011). A review of the impact of artificial light on invertebrates. Buglife – The Invertebrate Conservation Trust.
- Buck, J. B. (1937a). Studies on the firefly. I. The effects of light and other agents on flashing in Photinus pyralis, with special reference to periodicity and diurnal rhythm. Physiological Zoology, 10, 45–58. 10.1086/physzool.10.1.30151403 [DOI] [Google Scholar]
- Buck, J. B. (1937b). Studies on the firefly. II. The signal system and color vision in Photinus pyralis . Physiological Zoology, 10, 412–419. 10.1086/physzool.10.4.30151427 [DOI] [Google Scholar]
- Campbell, G. D. (1976). Activity rhythm in the cave cricket, Ceuthophilus conicaudus Hubbell. The American Midland Naturalist, 96(2), 350–366. 10.2307/2424075 [DOI] [Google Scholar]
- Case, J. , & Trinkle, M. S. (1968). Light‐inhibition of flashing in the firefly Photuris missouriensis . Biological Bulletin, 135, 476–485. 10.2307/1539710 [DOI] [Google Scholar]
- Caveney, S. , Scholtz, C. H. , & McIntyre, P. (1995). Patterns of daily flight Activity in onitine dung beetles (Scarabaeinae: Onitini). Oecologia, 103, 444–452. 10.1007/BF00328682 [DOI] [PubMed] [Google Scholar]
- Chittka, L. , Stelzer, R. J. & Stanewsky, R. (2013). Daily changes in ultraviolet light levels can synchronize the circadian clock of bumblebees (Bombus terrestris). Chronobiology International, 30, 434–442. [DOI] [PubMed] [Google Scholar]
- Conrad, K. F. , Warren, M. S. , Fox, R. , Parsons, M. S. , & Woiwod, I. P. (2006). Rapid declines of common, widespread British moths provide evidence of an insect biodiversity crisis. Biological Conservation, 132, 279–291. 10.1016/j.biocon.2006.04.020 [DOI] [Google Scholar]
- Corbet, S. A. , Sellick, R. D. , & Willoughby, N. G. (1974). Notes on the biology of the mayfly Povilla adusta in West Africa. Journal of Zoology, 172, 491–502. 10.1111/j.1469-7998.1974.tb04381.x [DOI] [Google Scholar]
- Costin, K. J. , & Boulton, A. M. (2016). A field experiment on the effect of introduced light pollution on fireflies (Coleoptera: Lampyridae) in the Piedmont Region of Maryland. The Coleopterists’ Bulletin, 70, 84–86. 10.1649/072.070.0110 [DOI] [Google Scholar]
- Cratsley, C. K. , & Lewis, S. M. (2003). Female preference for male courtship flashes in Photinus ignitus fireflies. Behavioral Ecology, 14, 135–140. 10.1093/beheco/14.1.135 [DOI] [Google Scholar]
- Cratsley, C. , Prasertkul, T. , & Thancharoen, A. (2012). Male aggregation behaviour in the fireflies Photinus ignitus and Pteroptyx malaccae. Lampyrid, 2. [Google Scholar]
- Cronin, T. W. , Järvilehto, M. , Weckström, M. , & Lall, A. B. (2000). Tuning of photoreceptor spectral sensitivity in fireflies (Coleoptera: Lampyridae). Journal of Comparative Physiology A, 186, 1–12. 10.1007/s003590050001 [DOI] [PubMed] [Google Scholar]
- Cronin T. W., Johnsen S., Justin Marshall N., & Warrant E. J. (Eds.). (2014). Visual ecology. Princeton: Princeton University Press, 432 pp. [Google Scholar]
- Dacke, M. , Baird, E. , Byrne, M. , Scholtz, C. H. , & Warrant, E. J. (2013). Dung beetles use the Milky Way for orientation. Current Biology, 23, 298–300. 10.1016/j.cub.2012.12.034 [DOI] [PubMed] [Google Scholar]
- Dacke, M. , Byrne, M. J. , Baird, E. , Scholtz, C. H. , & Warrant, E. J. (2011). How dim is dim? Precision of the celestial compass in moonlight and sunlight. Philosophical Transactions of the Royal Society of London. Series B, 366, 697–702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dacke, M. , Byrne, M. J. , Scholtz, C. H. , & Warrant, E. J. (2004). Lunar orientation in a beetle. Proceedings of the Royal Society of London B, 271, 361–365. 10.1098/rspb.2003.2594 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dacke, M. , Nilsson, D.‐E. , Scholtz, C. H. , Byrne, M. , & Warrant, E. J. (2003). Animal behaviour: Insect orientation to polarized moonlight. Nature, 424, 33 10.1038/424033a [DOI] [PubMed] [Google Scholar]
- Danthanarayana, W. , & Dashper, S. (1986). Response of some night‐flying insects to polarized light In Insect flight (pp. 120–127). Berlin, Heidelberg: Springer. [Google Scholar]
- Davies, T. W. , Bennie, J. , Cruse, D. , Blumgart, D. , Inger, R. , & Gaston, K. J. (2017). Multiple night‐time light‐emitting diode lighting strategies impact grassland invertebrate assemblages. Global Change Biology, 23, 2641–2648. 10.1111/gcb.13615 [DOI] [PubMed] [Google Scholar]
- Davies, T. W. , Bennie, J. , & Gaston, K. J. (2012). Street lighting changes the composition of invertebrate communities. Biology Letters, 8, 764–767. 10.1098/rsbl.2012.0216 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies, T. W. , Bennie, J. , Inger, R. , de Ibarra, N. H. , & Gaston, K. J. (2013). Artificial light pollution: Are shifting spectral signatures changing the balance of species interactions? Global Change Biology, 19, 1417–1423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies, T. W. , Bennie, J. , Inger, R. , & Gaston, K. J. (2013). Artificial light alters natural regimes of night‐time sky brightness. Scientific Reports, 3, 1722. [Google Scholar]
- Davies, T. W. , & Smyth, T. (2018). Why artificial light at night should be a focus for global change research in the 21st century. Global Change Biology, 24, 872–882. 10.1111/gcb.13927 [DOI] [PubMed] [Google Scholar]
- De Cock, R. (2009). Biology and behaviour of European lampyrids In Meyer‐Rochow V. B. (Ed.), Bioluminescence in focus—a collection of illuminating essays (pp. 161–200). Kerala, India: Research Signpost. [Google Scholar]
- Degen, T. , Mitesser, O. , Perkin, E. K. , Weiß, N.‐S. , Oehlert, M. , Mattig, E. , & Hölker, F. (2016). Street lighting: Sex‐independent impacts on moth movement. Journal of Animal Ecology, 85, 1352–1360. 10.1111/1365-2656.12540 [DOI] [PubMed] [Google Scholar]
- Delhey, K. , & Peters, A. (2017). Conservation implications of anthropogenic impacts on visual communication and camouflage. Conservation Biology, 31, 30–39. 10.1111/cobi.12834 [DOI] [PubMed] [Google Scholar]
- Deyrup, S. T. , Risteen, R. G. , Tonyai, K. K. , Farrar, M. A. , D'Antonio, B. E. , Ahmed, Z. B. , … Smedley, S. R. (2017). Escape into winter: Does a phenological shift by Ellychnia corrusca (winter firefly) shield it from a specialist predator (Photuris)? Northeastern Naturalist, 24, B147–B166. [Google Scholar]
- Dominoni, D. M. , Borniger, J. C. , & Nelson, R. J. (2016). Light at night, clocks and health: From humans to wild organisms. Biology Letters, 12, 20160015 10.1098/rsbl.2016.0015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donners, M. , van Grunsven, R. H. A. , Groenendijk, D. , van Langevelde, F. , Bikker, J. W. , Longcore, T. , & Veenendaal, E. (2018). Colors of attraction: Modeling insect flight to light behavior. Journal of Experimental Zoology A, 2018, 1–7. 10.1002/jez.2188. [DOI] [PubMed] [Google Scholar]
- Dreisig, H. (1975). Environmental control of the daily onset of luminescent activity in glowworms and fireflies (Coleoptera: Lampyridae). Oecologia, 18, 85–99. 10.1007/BF00348090 [DOI] [PubMed] [Google Scholar]
- Dreisig, H. (1980). The importance of illumination level in the daily onset of flight activity in nocturnal moths. Physiological Entomology, 5, 327–342. 10.1111/j.1365-3032.1980.tb00243.x [DOI] [Google Scholar]
- Durrant, J. , Botha, L. M. , Green, M. P. , & Jones, T. M. (2018). Artificial light at night prolongs juvenile development time in the black field cricket, Teleogryllus commodus . The Journal of Experimental Zoology, 12, 202. [DOI] [PubMed] [Google Scholar]
- Dyer, F. C. (1985). Nocturnal orientation by the Asian honey bee. Apis Dorsata. Animal Behaviour, 33(3), 769–774. 10.1016/S0003-3472(85)80009-9 [DOI] [Google Scholar]
- Eaton, J. L. , Tignor, K. R. , & Holtzman, G. I. (1983). Role of moth ocelli in timing flight initiation at dusk. Physiological Entomology, 8, 71–375. 10.1111/j.1365-3032.1983.tb00370.x [DOI] [Google Scholar]
- Eccard, J. A. , Scheffler, I. , Franke, S. , & Hoffmann, J. (2018). Off‐grid: Solar powered LED illumination impacts epigeal arthropods. Insect Conservation and Diversity, 12, 20160111 10.1111/icad.12303 [DOI] [Google Scholar]
- Egri, Á. , Száz, D. , Farkas, A. , Pereszlényi, Á. , Horváth, G. , & Kriska, G. (2017). Method to improve the survival of night‐swarming mayflies near bridges in areas of distracting light pollution. Royal Society Open Science, 4(11), 171166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eguchi, E. , Nemoto, A. , Meyer‐Rochow, V. B. , & Ohba, N. (1984). A comparative study of spectral sensitivity curves in three diurnal and eight nocturnal species of Japanese fireflies. Journal of Insect Physiology, 30, 607–612. 10.1016/0022-1910(84)90044-1 [DOI] [Google Scholar]
- Eisenbeis, G. (2006). Artificial night lighting and insects: Attraction of insects to streetlamps in a rural setting in Germany In Rich C., & Longcore T. (Eds.), Ecological consequences of artificial night lighting (pp. 281–304). Washington, D.C.: Island Press. [Google Scholar]
- Eisner, T. , Goetz, M. A. , Hill, D. E. , Smedley, S. R. , & Meinwald, J. (1997). Firefly “femmes fatales” acquire defensive steroids (lucibufagins) from their firefly prey. Proceedings of the National Academy of Sciences of the United States of America, 94, 9723–9728. 10.1073/pnas.94.18.9723 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eisner, T. , Wiemer, D. F. , Haynes, L. W. , & Meinwald, J. (1978). Lucibufagins: Defensive steroids from the fireflies Photinus ignitus and P. marginellus (Coleoptera: Lampyridae). Proceedings of the National Academy of Sciences of the United States of America, 75, 905–908. 10.1073/pnas.75.2.905 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elvidge, C. D. , Keith, D. M. , Tuttle, B. T. , & Baugh, K. E. (2010). Spectral identification of lighting type and character. Sensors, 10, 3961–3988. 10.3390/s100403961 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elvidge, C. D. , Sutton, P. C. , Baugh, K. , Ziskin, D. C. , Ghosh, T. , & Anderson, S. (2014). National Trends in Satellite Observed Lighting: 1992–2009 In Weng Q. (Ed.), Global urban monitoring and assessment through earth observation. Boca Raton, Florida: CRC Press. [Google Scholar]
- Enders, F. (1977). Web‐site selection by orb‐web spiders, particularly Argiope aurantia lucas . Animal Behaviour, 25, 694–712. 10.1016/0003-3472(77)90119-1 [DOI] [Google Scholar]
- Endler, J. A. (1993). The color of light in forests and its implications. Ecological Monographs, 63, 2–27. 10.2307/2937121 [DOI] [Google Scholar]
- Falchi, F. , Cinzano, P. , Duriscoe, D. , Kyba, C. C. , Elvidge, C. D. , Baugh, K. , … Furgoni, R. (2016). The new world atlas of artificial night sky brightness. Science Advances, 2, e1600377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farnworth, B. , Innes, J. , Kelly, C. , Littler, R. , & Waas, J. R. (2018). Photons and foraging: Artificial light at night generates avoidance behaviour in male, but not female, New Zealand weta. Environmental Pollution, 236, 82–90. 10.1016/j.envpol.2018.01.039 [DOI] [PubMed] [Google Scholar]
- Fatzinger, C. W. (1973). Circadian rhythmicity of sex pheromone release by Dioryctria abietella (Lepidoptera: Pyralidae (Phycitinae)) and the effect of a diel light cycle on its precopulatory behavior. Annals of the Entomological Society of America, 66, 1147–1153. [Google Scholar]
- Faust, L. F. (2017). Fireflies, glow‐worms, and lightning bugs: Identification and natural history of the fireflies of the Eastern and Central United States and (376 pp.). Canada: University of Georgia Press. [Google Scholar]
- Feuda, R. , Marlétaz, F. , Bentley, M. A. , & Holland, P. W. H. (2016). Conservation, duplication, and divergence of five opsin genes in insect evolution. Genome Biology and Evolution, 8, 579–587. 10.1093/gbe/evw015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Firebaugh, A. , & Haynes, K. J. (2016). Experimental tests of light‐pollution impacts on nocturnal insect courtship and dispersal. Oecologia, 182, 1203–1211. 10.1007/s00442-016-3723-1 [DOI] [PubMed] [Google Scholar]
- Foster, J. J. , Smolka, J. , Nilsson, D.‐E. , & Dacke, M. (2018). How animals follow the stars. Proceedings of the Royal Society of London B, 285, 20172322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox, R. (2013). The decline of moths in Great Britain: A review of possible causes. Insect Conservation and Diversity, 6, 5–19. 10.1111/j.1752-4598.2012.00186.x [DOI] [Google Scholar]
- Frank, K. D. (1988). Impact of outdoor lighting on moths: An assessment. Journal of the Lepidopterists’ Society, 42, 63–93. [Google Scholar]
- Frank, K. D. (2006). Effects of artificial night lighting on moths In Rich C., & Longcore T. (Eds.), Ecological consequences of artificial night lighting (pp. 305–344). Washington, D.C.: Island Press. [Google Scholar]
- Frank, K. D. (2009). Exploitation of artificial light at night by a diurnal jumping spider. Peckhamia, 78, 1–3. [Google Scholar]
- Futahashi, R. , Kawahara‐Miki, R. , Kinoshita, M. , Yoshitake, K. , Yajima, S. , Arikawa, K. , & Fukatsu, T. (2015). Extraordinary diversity of visual opsin genes in dragonflies. Proceedings of the National Academy of Sciences USA, 112, E1247–E1256. 10.1073/pnas.1424670112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garber, S. D. (1978). Opportunistic feeding behavior of Anolis cristatellus (Iguanidae: Reptilia) in Puerto Rico. Transactions of the Kansas Academy of Science, 81, 79 10.2307/3627360 [DOI] [Google Scholar]
- Gardiner, T. A. (2011). Glowing, glowing, gone? The plight of the Glow‐worm in Essex. Colby: British Naturalists’ Association, 80 pp. [Google Scholar]
- Garris, H. W. , & Snyder, J. A. (2010). Sex‐specific attraction of moth species to ultraviolet light traps. Southeastern Naturalist, 9, 427–434. 10.1656/058.009.0302 [DOI] [Google Scholar]
- Gaston, K. J. , & Bennie, J. (2014). Demographic effects of artificial nighttime lighting on animal populations. Environmental Reviews, 22, 323–330. 10.1139/er-2014-0005 [DOI] [Google Scholar]
- Gaston, K. J. , Bennie, J. , Davies, T. W. , & Hopkins, J. (2013). The ecological impacts of nighttime light pollution: A mechanistic appraisal. Biological Reviews of the Cambridge Philosophical Society, 88, 912–927. 10.1111/brv.12036 [DOI] [PubMed] [Google Scholar]
- Gaston, K. J. , Davies, T. W. , Bennie, J. , & Hopkins, J. (2012). Reducing the ecological consequences of night‐time light pollution: Options and developments. Journal of Applied Ecology, 49, 1256–1266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaston, K. J. , & Holt, L. A. (2018). Nature, extent and ecological implications of night‐time light from road vehicles. The Journal of Applied Ecology, 55, 2296–2307. 10.1111/1365-2664.13157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaston, K. J. , Visser, M. E. , & Hölker, F. (2015). The biological impacts of artificial light at night: The research challenge. Philosophical Transactions of the Royal Society of London. Series B, 370, 20140133 10.1098/rstb.2014.0133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gentry, E. (2003). On sexual selection in Florida’s Pyractomena borealis (Coleoptera: Lampyridae). The Florida Entomologist, 86, 114–123. 10.1653/0015-4040(2003)086[0114:OSSIFP]2.0.CO;2 [DOI] [Google Scholar]
- Gerrish, G. A. , Morin, J. G. , Rivers, T. J. , & Patrawala, Z. (2009). Darkness as an ecological resource: The role of light in partitioning the nocturnal niche. Oecologia, 160, 525–536. 10.1007/s00442-009-1327-8 [DOI] [PubMed] [Google Scholar]
- Giebultowicz, J. M. (2001). Peripheral clocks and their role in circadian timing: Insights from insects. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 356, 1791–1799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldsmith, T. H. (1990). Optimization, constraint, and history in the evolution of eyes. Quarterly Review of Biology, 65, 281–322. 10.1086/416840 [DOI] [PubMed] [Google Scholar]
- González‐Bernal, E. , Greenlees, M. J. , Brown, G. P. , & Shine, R. (2016). Toads in the backyard: Why do invasive cane toads (Rhinella marina) prefer buildings to bushland? Population Ecology, 58(2), 293–302. 10.1007/s10144-016-0539-0 [DOI] [Google Scholar]
- Goodman, L. J. (1965). The role of certain optomotor reactions in regulating stability in the rolling plane during flight in the desert locust, Schistocerca gregaria . Journal of Experimental Biology, 42, 385–407. [Google Scholar]
- Goretti, E. , Coletti, A. , Veroli, A. D. , Giulio, A. M. D. , & Gaino, E. (2011). Artificial light device for attracting pestiferous chironomids (Diptera): A case study at Lake Trasimeno (Central Italy). Italian Journal of Zoology, 78, 336–342. 10.1080/11250003.2010.534115 [DOI] [Google Scholar]
- Greiner, B. , Cronin, T. W. , Ribi, W. A. , Wcislo, W. T. , & Warrant, E. J. (2007). Anatomical and physiological evidence for polarisation vision in the nocturnal bee Megalopta genalis . Journal of Comparative Physiology. A, Neuroethology, Sensory, Neural, and Behavioral Physiology, 193, 591–600. 10.1007/s00359-007-0214-1 [DOI] [PubMed] [Google Scholar]
- Grenis, K. , & Murphy, S. M. (2018). Direct and indirect effects of light pollution on the performance of an herbivorous insect. Insect Science, 1–7. 10.1111/1744-7917.12574. [DOI] [PubMed] [Google Scholar]
- Grenis, K. , Tjossem, B. , & Murphy, S. M. (2015). Predation of larval Lepidoptera in habitat fragments varies spatially and temporally but is not affected by light pollution. Journal of Insect Conservation, 19, 559–566. 10.1007/s10841-015-9777-2 [DOI] [Google Scholar]
- Gronquist, M. , Schroeder, F. C. , Ghiradella, H. , Hill, D. , McCoy, E. M. , Meinwald, J. , & Eisner, T. (2006). Shunning the night to elude the hunter: Diurnal fireflies and the “femmes fatales”. Chemoecology, 16, 39–43. 10.1007/s00049-005-0325-5 [DOI] [Google Scholar]
- Guetté, A. , Godet, L. , Juigner, M. , & Robin, M. (2018). Worldwide increase in Artificial Light At Night around protected areas and within biodiversity hotspots. Biological Conservation, 223, 97–103. 10.1016/j.biocon.2018.04.018 [DOI] [Google Scholar]
- Gunn, P. , & Gunn, B. (2012). Lunar effects on the bioluminescent activity of the glow‐worm Lampyris noctiluca and its larvae. Lampyrid, 3, 1–16. [Google Scholar]
- Hagen, O. , & Viviani, V. R. (2009). Investigation of the artificial night lighting influence in firefly occurrence in the urban areas of Campinas and Sorocaba municipalities. Anais do IX Congresso de Ecologia do. Brasil.
- Hagen, O. , Santos, R. M. , Schlindwein, M. N. , & Viviani, V. R. (2015). Artificial night lighting reduces firefly (Coleoptera: Lampyridae) occurrence in Sorocaba, Brazil. Advances in Entomology, 03, 24–32. 10.4236/ae.2015.31004 [DOI] [Google Scholar]
- Hall, D. W. , Sander, S. E. , Pallansch, J. C. , & Stanger‐Hall, K. F. (2016). The evolution of adult light emission color in North American fireflies. Evolution, 70, 2033–2048. 10.1111/evo.13002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hallmann, C. A. , Sorg, M. , Jongejans, E. , Siepel, H. , Hofland, N. , Schwan, H. , … de Kroon, H. (2017). More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS One, 12, e0185809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamdorf, K. , & Höglund, G. (1981). Light induced retinal screening pigment migration independent of visual cell activity. Journal of Comparative Physiology A, 143, 305–309. 10.1007/BF00611166 [DOI] [Google Scholar]
- Hardie, J. (2010). Photoperiodism in insects: Aphid polyphenism In Nelson R. J., Denlinger D. L., & Somers D. E. (Eds.), Photoperiodism: The biological calendar. New York, NY: Oxford University Press. [Google Scholar]
- Hariyama, T. (2000). The brain as a photoreceptor: Intracerebral ocelli in the firefly. Naturwissenschaften, 87, 327–330. 10.1007/s001140050732 [DOI] [PubMed] [Google Scholar]
- Harvey, E. N. (1926). On the inhibition of animal luminescence by Light. Biological Bulletin, 51, 85–88. 10.2307/1536539 [DOI] [Google Scholar]
- Heiling, A. M. (1999). Why do nocturnal orb‐web spiders (Araneidae) search for light? Behavioral Ecology and Sociobiology, 46, 43–49. 10.1007/s002650050590 [DOI] [Google Scholar]
- Herzmann, D. , & Labhart, T. (1989). Spectral sensitivity and absolute threshold of polarization vision in crickets: A behavioral study. Journal of Comparative Physiology, 165, 315–319. 10.1007/BF00619350 [DOI] [PubMed] [Google Scholar]
- Hironaka, M. , Inadomi, K. , Nomakuchi, S. , Filippi, L. , & Hariyama, T. (2008). Canopy compass in nocturnal homing of the subsocial shield bug, Parastrachia japonensis (Heteroptera: Parastrachiidae). Naturwissenschaften, 95, 343–346. 10.1007/s00114-007-0324-1 [DOI] [PubMed] [Google Scholar]
- Hölker, F. , Moss, T. , Griefahn, B. , Kloas, W. , Voigt, C. C. , Henckel, D. , … Tockner, K. (2010). The dark side of light: A transdisciplinary research agenda for light pollution policy. Ecology and Society, 15, 13. [Google Scholar]
- Hölker, F. , Wolter, C. , Perkin, E. K. , & Tockner, K. (2010). Light pollution as a biodiversity threat. Trends in Ecology & Evolution, 25, 681–682. [DOI] [PubMed] [Google Scholar]
- Hölldobler, B. , & Taylor, R. W. (1983). A behavioral study of the primitive ant Nothomyrmecia macrops Clark. Insectes Sociaux, 30, 384–401. 10.1007/BF02223970 [DOI] [Google Scholar]
- Honkanen, A. , Immonen, E. V. , Salmela, I. , Heimonen, K. , & Weckström, M. (2017). Insect photoreceptor adaptations to night vision. Philosophical Transactions of the Royal Society of London B, 372(1717), 20160077 10.1098/rstb.2016.0077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honnen, A.‐C. , Johnston, P. R. , & Monaghan, M. T. (2016). Sex‐specific gene expression in the mosquito Culex pipiens f. molestus in response to artificial light at night. BMC Genomics, 17, 22 10.1186/s12864-015-2336-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hopkins, J. , Baudry, G. , Candolin, U. , & Kaitala, A. (2015). I’m sexy and I glow it: Female ornamentation in a nocturnal capital breeder. Biology Letters, 11, 20150599 10.1098/rsbl.2015.0599 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horridge, G. A. (1969). The eye of the firefly Photuris. Proceedings of the Royal Society London B, 171, 445–463. 10.1098/rspb.1969.0005 [DOI] [Google Scholar]
- Horváth, G. , Kriska, G. , Malik, P. , & Robertson, B. (2009). Polarized light pollution: A new kind of ecological photopollution. Frontiers in Ecology and the Environment, 7, 317–325. 10.1890/080129 [DOI] [Google Scholar]
- Hsiao, H. S. (1973). Flight paths of night‐flying moths to light. Journal of Insect Physiology, 19, 1971–1976. 10.1016/0022-1910(73)90191-1 [DOI] [PubMed] [Google Scholar]
- Hunt, G. L. (1977). Low preferred foraging temperatures and nocturnal foraging in a desert harvester ant. American Naturalist, 111, 589–591. 10.1086/283187 [DOI] [Google Scholar]
- Ineichen, S. , & Rüttimann, B. (2012). Impact of artificial light on the distribution of the common European glow‐worm, Lampyris noctiluca (Coleoptera: Lampyridae). Lampyrid, 2, 31–36. [Google Scholar]
- Inger, R. , Bennie, J. , Davies, T. W. , & Gaston, K. J. (2014). Potential biological and ecological effects of flickering artificial light. PloS One, 9(5), e98631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jayatilaka, P. , Narendra, A. , Reid, S. F. , Cooper, P. , & Zeil, J. (2011). Different effects of temperature on foraging activity schedules in sympatric Myrmecia ants. Journal of Experimental Biology, 214, 2730–2738. 10.1242/jeb.053710 [DOI] [PubMed] [Google Scholar]
- Johansen, N. S. , Vänninen, I. , Pinto, D. M. , Nissinen, A. I. , & Shipp, L. (2011). In the light of new greenhouse technologies: 2. Direct effects of artificial lighting on arthropods and integrated pest management in greenhouse crops. Annals of Applied Biology, 159, 1–27. 10.1111/j.1744-7348.2011.00483.x [DOI] [Google Scholar]
- Johnsen, S. , Kelber, A. , Warrant, E. , Sweeney, A. M. , Widder, E. A. , Lee, R. L. Jr. , & Hernández‐Andrés, J. (2006). Crepuscular and nocturnal illumination and its effects on color perception by the nocturnal hawkmoth Deilephila elpenor. Journal of Experimental Biology, 209, 789–800. 10.1242/jeb.02053 [DOI] [PubMed] [Google Scholar]
- Jones, T. M. , Durrant, J. , Michaelides, E. B. , & Green, M. P. (2015). Melatonin: A possible link between the presence of artificial light at night and reductions in biological fitness. Philosophical Transactions of the Royal Society of London. Series B, 370, 20140122. 10.1098/rstb.2014.0122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jung, K. , & Kalko, E. K. V. (2010). Where forest meets urbanization: Foraging plasticity of aerial insectivorous bats in an anthropogenically altered environment. Journal of Mammalogy, 91(1), 144–153. 10.1644/08-MAMM-A-313R.1. [DOI] [Google Scholar]
- Justice, M. J. , & Justice, T. C. (2016). Attraction of insects to incandescent, compact fluorescent, halogen, and LED lamps in a light trap: Implications for light pollution and urban ecologies. Entomological News, 125(5), 315–326. [Google Scholar]
- Kaul, R. M. , & Kopteva, G. A. (1982). Night orientation of ants Formica rufa (Hymenoptera: Formicidae) upon movement on routes. Zoologicheskij Zhurnal. Moscow, 61, 1351–1358. [Google Scholar]
- Kehoe, R. C. , Cruse, D. , Sanders, D. , Gaston, K. J. , & van Veen, F. J. F. (2018). Shifting daylength regimes associated with range shifts alter aphid‐parasitoid community dynamics. Ecology and Evolution, 147, 381 10.1002/ece3.4401 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelber, A. , Balkenius, A. , & Warrant, E. J. (2003). Colour vision in diurnal and nocturnal hawkmoths. Integrative and Comparative Biology, 43, 571–579. 10.1093/icb/43.4.571 [DOI] [PubMed] [Google Scholar]
- Kelber, A. , Jonsson, F. , Wallén, R. , Warrant, E. , Kornfeldt, T. , & Baird, E. (2011). Hornets can fly at night without obvious adaptations of eyes and ocelli. PloS One, 6, e21892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelber, A. , & Roth, L. S. V. (2006). Nocturnal colour vision–not as rare as we might think. Journal of Experimental Biology, 209, 781–788. 10.1242/jeb.02060 [DOI] [PubMed] [Google Scholar]
- Kelber, A. , Warrant, E. J. , Pfaff, M. , Wallén, R. , Theobald, J. C. , Wcislo, W. T. , & Raguso, R. A. (2006). Light intensity limits foraging activity in nocturnal and crepuscular bees. Behavioral Ecology, 17, 63–72. 10.1093/beheco/arj001 [DOI] [Google Scholar]
- Kempinger, L. , Dittmann, R. , Rieger, D. , & Helfrich‐Forster, C. (2009). The nocturnal activity of fruit flies exposed to artificial moonlight is partly caused by direct light effects on the activity level that bypass the endogenous clock. Chronobiology International, 26, 151–166. 10.1080/07420520902747124 [DOI] [PubMed] [Google Scholar]
- Kerfoot, W. B. (1967). The lunar periodicity of Sphecodogastra texana, a nocturnal bee (Hymenoptera: Halictidae). Animal Behaviour, 15, 479–486. 10.1016/0003-3472(67)90047-4 [DOI] [PubMed] [Google Scholar]
- Kevan, P. G. , Chittka, L. , & Dyer, A. G. (2001). Limits to the salience of ultraviolet: Lessons from colour vision in bees and birds. Journal of Experimental Biology, 204, 2571–2580. [DOI] [PubMed] [Google Scholar]
- Khoo, V. (2014). Conservation of the Selangor River population of Pteroptyx tener in Malaysia: Results of seven years of monitoring. International Firefly Symposium 2014.
- Kline, R. B. (2013). Beyond significance testing: Statistics reform in the behavioral sciences, 2nd ed Washington, DC: American Psychological Association. [Google Scholar]
- Klotz, J. H. , & Reid, B. L. (1993). Nocturnal orientation in the black carpenter ant Camponotus pennsylvanicus (DeGeer) (Hymenoptera: Formicidae). Insectes Sociaux, 40, 95–106. 10.1007/BF01338835 [DOI] [Google Scholar]
- Knop, E. , Zoller, L. , Ryser, R. , Gerpe, C. , Hörler, M. , & Fontaine, C. (2017). Artificial light at night as a new threat to pollination. Nature, 548, 206–209. 10.1038/nature23288 [DOI] [PubMed] [Google Scholar]
- Kokko, H. , & Sutherland, W. J. (2001). Ecological traps in changing environments: Ecological and evolutionary consequences of a behaviourally mediated Allee effect. Evolutionary Ecology Research, 3, 603–610. [Google Scholar]
- Krames, M. R. , Shchekin, O. B. , Mueller‐Mach, R. , Mueller, G. O. , Zhou, L. , Harbers, G. , & Craford, M. G. (2007). Status and future of high‐power light‐emitting diodes for solid‐State lighting. Journal of Display Technology, 3, 160–175. 10.1109/JDT.2007.895339 [DOI] [Google Scholar]
- Kriska, G. , Malik, P. , Szivák, I. , & Horváth, G. (2008). Glass buildings on river banks as “polarized light traps” for mass‐swarming polarotactic caddis flies. Naturwissenschaften, 95, 461–467. 10.1007/s00114-008-0345-4 [DOI] [PubMed] [Google Scholar]
- Kronfeld‐Schor, N. , Dominoni, D. , de la Iglesia, H. , Levy, O. , Herzog, E. D. , Dayan, T. , & Helfrich‐Forster, C. (2013). Chronobiology by moonlight. Proceedings of the Royal Society London B, 280, 20123088 10.1098/rspb.2012.3088 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurvers, R. H. J. M. , & Hölker, F. (2015). Bright nights and social interactions: A neglected issue. Behavioral Ecology, 26, 334–339. 10.1093/beheco/aru223 [DOI] [Google Scholar]
- Kyba, C. , Kuester, T. , Sanchez de Miguel, A. , Baugh, K. , Jechow, A. , Hölker, F. , … Guanter, L. (2017). Artificially lit surface of Earth at night increasing in radiance and extent. Science Advances, 3, e1701528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyba, C. C. M. , Ruhtz, T. , Fischer, J. , & Hölker, F. (2011a). Lunar skylight polarization signal polluted by urban lighting. Journal of Geophysical Research, 116, D24106. [Google Scholar]
- Kyba, C. C. M. , Ruhtz, T. , Fischer, J. , & Hölker, F. (2011b). Cloud coverage acts as an amplifier for ecological light pollution in urban ecosystems. PLoS One, 6, e17307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lall, A. B. (1981). Electroretinogram and the spectral sensitivity of the compound eyes in the firefly Photuris versicolor (Coleoptera‐Lampyridae): A correspondence between green sensitivity and species bioluminescence emission. Journal of Insect Physiology, 27, 461–468. 10.1016/0022-1910(81)90097-4 [DOI] [Google Scholar]
- Lall, A. B. (1993). Nightly increase in visual sensitivity correlated with bioluminescent flashing activity in the firefly Photuris versicolor (Coleoptera: Lampyridae). The Journal of Experimental Zoology, 265, 609–612. 10.1002/jez.1402650518 [DOI] [PubMed] [Google Scholar]
- Lall, A. B. , Chapman, R. M. , Ovid Trouth, C. , & Holloway, J. A. (1980). Spectral mechanisms of the compound eye in the firefly Photinus pyralis (Coleoptera: Lampyridae). Journal of Comparative Physiology, 135, 21–27. [Google Scholar]
- Lall, A. B. , & Lloyd, J. E. (1989). Spectral sensitivity of the compound eyes in two day‐active fireflies (Coleoptera: Lampyridae: Lucidota). Journal of Comparative Physiology. A, Sensory, Neural, and Behavioral Physiology, 166, 257–260. [DOI] [PubMed] [Google Scholar]
- Lall, A. B. , Lord, E. T. , & Ovid Trouth, C. (1982). Vision in the firefly Photuris lucicrescens (Coleoptera: Lampyridae): Spectral sensitivity and selective adaptation in the compound eye. Journal of Comparative Physiology, 147, 195–200. 10.1007/BF00609844 [DOI] [Google Scholar]
- Lall, A. B. , Seliger, H. H. , Biggley, W. H. , & Lloyd, J. E. (1980). Ecology of colors of firefly bioluminescence. Science, 210, 560–562. [DOI] [PubMed] [Google Scholar]
- Lall, A. B. , Strother, G. K. , Cronin, T. W. , & Seliger, H. H. (1988). Modification of spectral sensitivities by screening pigments in the compound eyes of twilight‐active fireflies (Coleoptera: Lampyridae). Journal of Comparative Physiology. A, 162, 23–33. [DOI] [PubMed] [Google Scholar]
- Laughlin, S. B. , & Hardie, R. C. (1978). Common strategies for light adaptation in the peripheral visual systems of fly and dragonfly. Journal of Comparative Physiology, 128, 319–340. [Google Scholar]
- Leuthold, R. H. , Bruinsma, O. , & van Huis, A. (1976). Optical and pheromonal orientation and memory for homing distance in the harvester termite Hodotermes mossambicus (Hagen). Behavioral Ecology and Sociobiology, 1, 127–139. 10.1007/BF00299194 [DOI] [Google Scholar]
- Lewanzik, D. , & Voigt, C. C. (2017). Transition from conventional to light‐emitting diode street lighting changes activity of urban bats. The Journal of Applied Ecology, 54, 264–271. 10.1111/1365-2664.12758 [DOI] [Google Scholar]
- Lewis, S. M. (2009). Bioluminescence and sexual signaling in fireflies In Meyer‐Rochow V. B. (Ed.), Bioluminescence in focus: A collection of illuminating essays. Kerala, India: Research Signpost. [Google Scholar]
- Lewis, S. (2016). Silent SPARKS: The wondrous world of fireflies, Vol. 240 (240 pp.). Princeton: Princeton University Press. [Google Scholar]
- Lewis, S. M. , & Wang, O. T. (1991). Reproductive ecology of two species of Photinus fireflies (Coleoptera: Lampyridae). Psyche, 98, 293–307. 10.1155/1991/76452 [DOI] [Google Scholar]
- Lloyd, J. E. (1997). Firefly mating ecology, sexual selection and evolution In Choe J. C., & Crespi B. J. (Eds.), The evolution of mating systems in insects and arachnids (pp. 184–192). Cambridge: Cambridge University Press. [Google Scholar]
- Lloyd, J. E. (2000). On quantifying mate search in a perfect insect— Seeking true facts and insight (Coleoptera: Lampyridae, Photinus). Florida Entomologist, 83, 211–228. [Google Scholar]
- Lloyd, J. E. (2006). Stray light, fireflies, and fireflyers In Rich C., & Longcore T. (Eds.), Ecological consequences of artificial night lighting (pp. 345–364). Washington, D.C.: Island Press. [Google Scholar]
- Lloyd, J. E. (2008). Fireflies In Capinera J. L. (Ed.), Encyclopedia of Entomology (pp. 1429–1452). New York, NY: Springer. [Google Scholar]
- Lloyd, J. E. (1966). Studies on the flash communication system in Photinus fireflies. Unpublished PhD thesis, University of Michigan.
- Longcore, T. , Aldern, H. L. , Eggers, J. F. , Flores, S. , Franco, L. , Hirshfield‐Yamanishi, E. , … Barroso, A. M. (2015). Tuning the white light spectrum of light emitting diode lamps to reduce attraction of nocturnal arthropods. Philosophical Transactions of the Royal Society of London. Series B, 370, 20140125. 10.1098/rstb.2014.0125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Longcore, T. , & Rich, C. (2004). Ecological light pollution. Frontiers in Ecology and the Environment, 2, 191–198. 10.1890/1540-9295(2004)002[0191:ELP]2.0.CO;2 [DOI] [Google Scholar]
- Longcore, T. , Rodríguez, A. , Witherington, B. , Penniman, J. F. , Herf, L. , & Herf, M. (2018). Rapid assessment of lamp spectrum to quantify ecological effects of light at night. Journal of Experimental Zoology A, 1–11. 10.1002/jez.2184. [DOI] [PubMed] [Google Scholar]
- Lunau, K. (2014). Visual ecology of flies with particular reference to colour vision and colour preferences. Journal of Comparative Physiology A, 200, 497–512. 10.1007/s00359-014-0895-1 [DOI] [PubMed] [Google Scholar]
- Macgregor, C. J. , Evans, D. M. , Fox, R. , & Pocock, M. J. O. (2017). The dark side of street lighting: Impacts on moths and evidence for the disruption of nocturnal pollen transport. Global Change Biology, 23, 697–707. 10.1111/gcb.13371 [DOI] [PubMed] [Google Scholar]
- Macgregor, C. J. , Pocock, M. J. O. , Fox, R. , & Evans, D. M. (2015). Pollination by nocturnal Lepidoptera, and the effects of light pollution: A review. Ecological Entomology, 40, 187–198. 10.1111/een.12174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manfrin, A. , Singer, G. , Larsen, S. , Weiß, N. , van Grunsven, R. H. A. , Weiß, N.‐S. , … Hölker, F. (2017). Artificial light at night affects organism flux across ecosystem boundaries and drives community structure in the recipient ecosystem. Frontiers of Environmental Science & Engineering in China, 5, 61 10.3389/fenvs.2017.00061 [DOI] [Google Scholar]
- Martin, G. J. , Lord, N. P. , Branham, M. A. , & Bybee, S. M. (2015). Review of the firefly visual system (Coleoptera: Lampyridae) and evolution of the opsin genes underlying color vision. Organisms, Diversity & Evolution, 15, 513–526. 10.1007/s13127-015-0212-z [DOI] [Google Scholar]
- McGeachie, W. J. (1988). A remote sensing method for the estimation of light‐trap efficiency. Bulletin of Entomological Research, 78, 379–385. 10.1017/S0007485300013158 [DOI] [Google Scholar]
- McIntyre, P. , & Caveney, S. (1998). Superposition optics and the time of flight in onitine dung beetles. Journal of Comparative Physiology A, 183, 45–60. 10.1007/s003590050233 [DOI] [Google Scholar]
- McLay, L. K. , Nagarajan‐Radha, V. , Green, M. P. , & Jones, T. M. (2018). Dim artificial light at night affects mating, reproductive output, and reactive oxygen species in Drosophila melanogaster . Journal of Experimental Zoology A, 1–10. 10.1002/jez.2164. [DOI] [PubMed] [Google Scholar]
- McMahon, T. A. , Rohr, J. R. , & Bernal, X. E. (2017). Light and noise pollution interact to disrupt interspecific interactions. Ecology, 1–10. 10.1002/ecy.1770 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merritt, D. J. , & Clarke, A. K. (2013). The impact of cave lighting on the bioluminescent display of the Tasmanian glow‐worm Arachnocampa tasmaniensis . Journal of Insect Conservation, 17, 147–153. 10.1007/s10841-012-9493-0 [DOI] [Google Scholar]
- Meyer, L. A. , & Sullivan, S. M. P. (2013). Bright lights, big city: Influences of ecological light pollution on reciprocal stream–riparian invertebrate fluxes. Ecological Applications, 23, 1322–1330. 10.1890/12-2007.1 [DOI] [PubMed] [Google Scholar]
- Meyer‐Rochow, V. B. (2007). Glowworms: A review of Arachnocampa spp. and kin. Luminescence, 22, 251–265. 10.1002/bio.955 [DOI] [PubMed] [Google Scholar]
- Meyer‐Rochow, V. B. , Kashiwagi, T. , & Eguchi, E. (2002). Selective photoreceptor damage in four species of insects induced by experimental exposures to UV‐irradiation. Micron, 33, 23–31. 10.1016/S0968-4328(00)00073-1 [DOI] [PubMed] [Google Scholar]
- Mikkola, K. (1972). Behavioural and electrophysiological responses of night‐flying insects, especially Lepidoptera, to near‐ultraviolet and visible light. Annales Zoologici Fennici, 9, 225–254. [Google Scholar]
- Miller, C. R. , Barton, B. T. , Zhu, L. , Radeloff, V. C. , Oliver, K. M. , Harmon, J. P. , & Ives, A. R. (2017). Combined effects of night warming and light pollution on predator‐prey interactions. Proceedings of the Royal Society London B, 284, 20171195. 10.1098/rspb.2017.1195 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mills, R. , Popple, J.‐A. , Veidt, M. , & Merritt, D. J. (2016). Detection of light and vibration modulates bioluminescence intensity in the glowworm, Arachnocampa flava. Journal of Comparative Physiology A, 202, 313–327. 10.1007/s00359-016-1077-0 [DOI] [PubMed] [Google Scholar]
- Minnaar, C. , Boyles, J. G. , Minnaar, I. A. , Sole, C. L. , & McKechnie, A. E. (2015). Stacking the odds: Light pollution may shift the balance in an ancient predator–prey arms race. Journal of Applied Ecology, 52, 522–531. 10.1111/1365-2664.12381 [DOI] [Google Scholar]
- Muirhead‐Thompson, R. (1991). Trap responses of flying insects: The influence of trap design on capture efficiency. London: Academic Press, 287 pp. ISBN 0-12-509755-7. [Google Scholar]
- Nabli, H. , Bailey, W. C. , & Necibi, S. (1999). Beneficial insect attraction to light traps with different wavelengths. Biological Control, 16, 185–188. 10.1006/bcon.1999.0748 [DOI] [Google Scholar]
- Narendra, A. , Greiner, B. , Ribi, W. A. , & Zeil, J. (2016). Light and dark adaptation mechanisms in the compound eyes of Myrmecia ants that occupy discrete temporal niches. The Journal of Experimental Biology, 219(16), 2435–2442. [DOI] [PubMed] [Google Scholar]
- Narendra, A. , Reid, S. F. , & Raderschall, C. A. (2013). Navigational efficiency of nocturnal Myrmecia ants suffers at low light levels. PloS One, 8, e58801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Narendra, A. , & Ribi, W. A. (2017). Ocellar structure is driven by the mode of locomotion and activity time in Myrmecia ants. The Journal of Experimental Biology, 220(23), 4383–4390. [DOI] [PubMed] [Google Scholar]
- Navara, K. J. , & Nelson, R. J. (2007). The dark side of light at night: Physiological, epidemiological, and ecological consequences. Journal of Pineal Research, 43, 215–224. 10.1111/j.1600-079X.2007.00473.x [DOI] [PubMed] [Google Scholar]
- Nemfc, S. J. (1971). Effects of lunar phases on light‐trap collections and populations of bollworm moths. Journal of Economic Entomology, 64, 860–864. [Google Scholar]
- Neumann, D. (1989). Circadian components of semilunar and lunar timing mechanisms. Journal of Biological Rhythms, 4, 285–294. 10.1177/074873048900400213 [DOI] [PubMed] [Google Scholar]
- Niva, C. C. , & Takeda, M. (2003). Effects of photoperiod, temperature and melatonin on nymphal development, polyphenism and reproduction in Halyomorpha halys (Heteroptera: Pentatomidae). Zoological Science, 20, 963–970. 10.2108/zsj.20.963 [DOI] [PubMed] [Google Scholar]
- Nowinszky, L. (2004). Nocturnal illumination and night flying insects. Applied Ecology and Environmental Research, 2, 17–52. 10.15666/aeer/02017052 [DOI] [Google Scholar]
- Numata, H. , Miyazaki, Y. , & Ikeno, T. (2015). Common features in diverse insect clocks. Zoological Letters, 1, 10 10.1186/s40851-014-0003-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Numata, H. , Shiga, S. , & Morita, A. (1997). Photoperiodic receptors in arthropods. Zoological Science, 14, 187–197. 10.2108/zsj.14.187 [DOI] [Google Scholar]
- Oba, Y. , & Kainuma, T. (2009). Diel changes in the expression of long wavelength‐sensitive and ultraviolet‐sensitive opsin genes in the Japanese firefly, Luciola cruciata . Gene, 436, 66–70. 10.1016/j.gene.2009.02.001 [DOI] [PubMed] [Google Scholar]
- Owens, A. C. S. , Meyer-Rochow, V. B. & Yang, E.-C. (2018). Short- and mid-wavelength artificial light influences the flash signals of Aquatica ficta fireflies (Coleoptera: Lampyridae). PloS One, 13, e0191576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacheco, Y. M. , Martin, G. J. , & Bybee, S. M. (2016). On the phototactic response of Rwandan Diaphanes Motschulsky (Coleoptera: Lampyridae) to a trap with a 630nm red light. Coleopterists’ Bulletin, 70, 559–561. [Google Scholar]
- Pawson, S. M. , & Bader, M.‐K.‐F. (2014). LED lighting increases the ecological impact of light pollution irrespective of color temperature. Ecological Applications, 24, 1561–1568. 10.1890/14-0468.1 [DOI] [PubMed] [Google Scholar]
- Pawson, S. M. , Watt, M. S. , & Brockerhoff, E. G. (2009). Using differential responses to light spectra as a monitoring and control tool for Arhopalus ferus (Coleoptera: Cerambycidae) and other exotic wood‐boring pests. Journal of Economic Entomology, 102, 79–85. [DOI] [PubMed] [Google Scholar]
- Peitsch, D. , Fietz, A. , Hertel, H. , de Souza, J. , Ventura, D. F. , & Menzel, R. (1992). The spectral input systems of hymenopteran insects and their receptor‐based colour vision. Journal of Comparative Physiology A, 170, 23–40. 10.1007/BF00190398 [DOI] [PubMed] [Google Scholar]
- Perkin, E. K. , Hölker, F. , Richardson, J. S. , Sadler, J. P. , Wolter, C. , & Tockner, K. (2011). The influence of artificial light on stream and riparian ecosystems: Questions, challenges, and perspectives. Ecosphere, 2, 1–16. 10.1890/ES11-00241.1 [DOI] [Google Scholar]
- Perkin, E. K. , Hölker, F. , & Tockner, K. (2014). Effects of artificial lighting on adult aquatic and terrestrial insects. Freshwater Biology, 59, 368–377. [Google Scholar]
- Picchi, M. S. , Avolio, L. , Azzani, L. , Brombin, O. , & Camerini, G. (2013). Fireflies and land use in an urban landscape: The case of Luciola italica L. (Coleoptera: Lampyridae) in the city of Turin. Journal of Insect Conservation, 17, 797–805. 10.1007/s10841-013-9562-z [DOI] [Google Scholar]
- Pittendrigh, C. S. (1993). Temporal organization: Reflections of a Darwinian clock‐watcher. Annual Review of Physiology, 55, 16–54. 10.1146/annurev.ph.55.030193.000313 [DOI] [PubMed] [Google Scholar]
- Pittendrigh, C. S. , & Minis, D. H. (1971). The photoperiodic time measurement in Pectinophora gossypiella and its relation to the circadian system in that species In Menaker M. (Ed.), Biochronometry (pp. 212–250). Washington, DC: National Academy of Sciences. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plummer, K. E. , Hale, J. D. , O’Callaghan, M. J. , Sadler, J. P. , & Siriwardena, G. M. (2016). Investigating the impact of street lighting changes on garden moth communities. Journal of Urban Ecology, 2, 1–10. 10.1093/jue/juw004 [DOI] [Google Scholar]
- Prasertkul, T. (2018). Characteristics of Pteroptyx firefly congregations in a human dominated habitat. Journal of Insect Behavior, 31, 436–457. 10.1007/s10905-018-9687-8. [DOI] [Google Scholar]
- Redford, K. H. (1982). Prey attraction as a possible function of bioluminescence in the larvae of Pyrearinus termitilluminans (Coleoptera: Elateridae). Revista Brasileira De Zoologia, 1, 31–34. 10.1590/S0101-81751982000100004 [DOI] [Google Scholar]
- Reid, S. F. , Narendra, A. , Hemmi, J. M. , & Zeil, J. (2011). Polarised skylight and the landmark panorama provide night‐active bull ants with compass information during route following. Journal of Experimental Biology, 214, 363–370. 10.1242/jeb.049338 [DOI] [PubMed] [Google Scholar]
- Rich, C. , & Longcore, T. (2006). Ecological Consequences of Artificial Night Lighting. Washington, DC: Island Press. [Google Scholar]
- Riemann, J. G. , Johnson, M. , & Thorson, B. (1981). Recovery of fertility by Mediterranean flour moths transferred from continuous light to light:dark. Annals of the Entomological Society of America, 74, 274–278. [Google Scholar]
- Riley, J. R. , Reynolds, D. R. , & Farmery, M. J. (1983). Observations of the flight behaviour of the army worm moth, Spodoptera exempta, at an emergence site using radar and infra‐red optical techniques. Ecological Entomology, 8, 395–418. 10.1111/j.1365-2311.1983.tb00519.x [DOI] [Google Scholar]
- Riley, J. R. , Reynolds, D. R. , & Farrow, R. A. (1987). The migration of Nilaparvata lugens (stål) (Delphacidae) and other hemiptera associated with rice during the dry season in the Philippines: A study using radar, visual observations, aerial netting and ground trapping. Bulletin of Entomological Research, 77, 145–169. 10.1017/S0007485300011627 [DOI] [Google Scholar]
- Robertson, B. , Kriska, G. , Horvath, V. , & Horvath, G. (2010). Glass buildings as bird feeders: Urban birds exploit insects trapped by polarized light pollution. Acta Zoologica, 56, 283–293. [Google Scholar]
- Robertson, B. A. , Rehage, J. S. , & Sih, A. (2013). Ecological novelty and the emergence of evolutionary traps. Trends in Ecology & Evolution, 28(9), 552–560. 10.1016/j.tree.2013.04.004 [DOI] [PubMed] [Google Scholar]
- Robinson, H. S. (1952). On the behaviour of night‐flying insects in the neighbourhood of a bright source of light. Proceedings of the Royal Entomological Society of London A, 27, 13–21. 10.1111/j.1365-3032.1952.tb00139.x [DOI] [Google Scholar]
- Ruck, P. (1958). Dark adaptation of the ocellus in Periplaneta Americana: A study of the electrical response to illumination. Journal of Insect Physiology, 2(3), 189–198. 10.1016/0022-1910(58)90004-0 [DOI] [Google Scholar]
- Rydell, J. (1992). Exploitation of insects around streetlamps by bats in Sweden. Functional Ecology, 6, 744–750. 10.2307/2389972 [DOI] [Google Scholar]
- Rydell, J. (2006). Bats and their insect prey at streetlights In Rich C., & Longcore T. (Eds.), Ecological consequences of artificial night lighting. Washington, D.C: Island Press. [Google Scholar]
- Sander, S. E. , & Hall, D. W. (2015). Variation in opsin genes correlates with signalling ecology in North American fireflies. Molecular Ecology, 24, 4679–4696. 10.1111/mec.13346 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders, D. , & Gaston, K. J. (2018). How ecological communities respond to artificial light at night. Journal of Experimental Zoology A, 1–7. 10.1002/jez.2157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders, D. , Kehoe, R. , Cruse, D. , van Veen, F. J. F. , & Gaston, K. J. (2018). Low levels of artificial light at night strengthen top‐down control in insect food web. Current Biology, 28(15), 2474–2478. 10.1016/j.cub.2018.05.078 [DOI] [PubMed] [Google Scholar]
- Sanders, D. , Kehoe, R. , Tiley, K. , Bennie, J. , Cruse, D. , Davies, T. W. , … Gaston, K. J. (2015). Artificial nighttime light changes aphid‐parasitoid population dynamics. Scientific Reports, 5, 15232 10.1038/srep15232 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Satoh, A. , Yoshioka, E. , & Numata, H. (2008). Circatidal activity rhythm in the mangrove cricket Apteronemobius asahinai . Biology Letters, 4, 233–236. 10.1098/rsbl.2008.0036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saunders, D. S. (2009). Circadian rhythms and the evolution of photoperiodic timing in insects. Physiological Entomology, 34, 301–308. 10.1111/j.1365-3032.2009.00699.x [DOI] [Google Scholar]
- Saunders, D. S. (2012). Insect photoperiodism: Seeing the light. Physiological Entomology, 37, 207–218. 10.1111/j.1365-3032.2012.00837.x [DOI] [Google Scholar]
- Schmitz, O. J. , Beckerman, A. P. , & O’Brien, K. M. (1997). Behaviorally mediated trophic cascades: Effects of predation risk on food web interactions. Ecology, 78, 1388–1399. 10.1890/0012-9658(1997)078[1388:BMTCEO]2.0.CO;2 [DOI] [Google Scholar]
- Schwalb, H. H. (1961). Contributions to the biology of the domestic lampyridae Lampyris noctiluca and Phausis splendidula and experimental analysis of their predatory and sexual behavior. Zoologiscbee Jahrbuch, 88, 99–550. [Google Scholar]
- Seliger, H. H. , Lall, A. B. , Lloyd, J. E. , & Biggley, W. H. (1982). The colors of firefly bioluminescence—I. Optimization Model. Photochemistry and Photobiology, 36, 673–680. 10.1111/j.1751-1097.1982.tb09488.x [DOI] [Google Scholar]
- Seymoure, B. M. (2018). Enlightening butterfly conservation efforts: The importance of natural lighting for butterfly behavioral ecology and conservation. Insects, 9(1). 10.3390/insects9010022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seymoure, B. M. (2016). Heliconius in a new light: The effects of light environments on mimetic coloration, behavior, and visual systems. PhD, Arizona State University.
- Sharkey, C. R. , Fujimoto, M. S. , Lord, N. P. , Shin, S. , McKenna, D. D. , Suvorov, A. , … Bybee, S. M. (2017). Overcoming the loss of blue sensitivity through opsin duplication in the largest animal group, beetles. Scientific Reports, 7, 8 10.1038/s41598-017-00061-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimoda, M. , & Honda, K.‐I. (2013). Insect reactions to light and its applications to pest management. Applied Entomology and Zoology, 48, 413–421. 10.1007/s13355-013-0219-x [DOI] [Google Scholar]
- Siderhurst, M. S. , James, D. M. , & Bjostad, L. B. (2006). Ultraviolet light induced autophototoxicity and negative phototaxis in Reticulitermes termites (Isoptera: Rhinotermitidae). Sociobiology, 48, 27–49. [Google Scholar]
- Skutelsky, O. (1996). Predation risk and state‐dependent foraging in scorpions: Effects of moonlight on foraging in the scorpion Buthus occitanus . Animal Behaviour, 52, 49–57. 10.1006/anbe.1996.0151 [DOI] [Google Scholar]
- Somanathan, H. , Borges, R. M. , Warrant, E. J. , & Kelber, A. (2008). Visual ecology of Indian carpenter bees I: Light intensities and flight activity. Journal of Comparative Physiology A, 194, 97–107. 10.1007/s00359-007-0291-1 [DOI] [PubMed] [Google Scholar]
- Somanathan, H. , Kelber, A. , Borges, R. M. , Wallén, R. , & Warrant, E. J. (2009). Visual ecology of Indian carpenter bees II: Adaptations of eyes and ocelli to nocturnal and diurnal lifestyles. Journal of Comparative Physiology A, 195, 571–583. 10.1007/s00359-009-0432-9 [DOI] [PubMed] [Google Scholar]
- Somers‐Yeates, R. , Hodgson, D. , McGregor, P. K. , Spalding, A. , & Ffrench‐Constant, R. H. (2013). Shedding light on moths: Shorter wavelengths attract noctuids more than geometrids. Biology Letters, 9, 20130376 10.1098/rsbl.2013.0376 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soong, K. , Lee, Y. J. , & Chang, I. H. (2011). Short‐lived intertidal midge Pontomyia oceana have semilunar eclosion rhythm entrained by night light. Marine Ecology Progress Series, 433, 121–130. 10.3354/meps09181 [DOI] [Google Scholar]
- Sotthibandhu, S. , & Baker, R. (1979). Celestial orientation by the large yellow underwing moth Noctua pronuba . Animal Behavior, 27, 786–800. [Google Scholar]
- Sower, L. L. , Shorey, H. H. , & Gaston, L. K. (1970). Sex pheromones of noctuid moths. XXI. Light: Dark cycle regulation and light inhibition of sex pheromone release by females of Trichoplusia ni . Annals of the Entomological Society of America, 63, 1090–1092. [DOI] [PubMed] [Google Scholar]
- Spoelstra, K. , van Grunsven, R. H. A. , Donners, M. , Gienapp, P. , Huigens, M. E. , Slaterus, R. , ... Veenendaal, E. (2015). Experimental illumination of natural habitat—an experimental set-up to assess the direct and indirect ecological consequences of artificial light of different spectral composition. Philosophical Transactions of the Royal Society of London. Series B, 370, 20140129 10.1098/rstb.2014.0129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stark, W. S. , Walker, K. D. , & Eidel, J. M. (1985). Ultraviolet and blue light induced damage to the Drosophila retina: Microspectrophotometry and electrophysiology. Current Eye Research, 4, 1059–1075. [DOI] [PubMed] [Google Scholar]
- Stiles, W. S. , & Burch, J. M. (1959). N.P.L. Colour‐matching investigation: Final report (1958). Optica Acta, 6, 1–26. 10.1080/713826267 [DOI] [PubMed] [Google Scholar]
- Stockman, A. , & Sharpe, L. T. (2000). The spectral sensitivities of the middle‐ and long‐wavelength‐sensitive cones derived from measurements in observers of known genotype. Vision Research, 40, 1711–1737. 10.1016/S0042-6989(00)00021-3 [DOI] [PubMed] [Google Scholar]
- Šustek, Z. (1999). Light attraction of carabid beetles and their survival in the city centre. Biologia Bratislava, 43, 539–551. [Google Scholar]
- Svensson, A. M. , & Rydell, J. (1998). Mercury vapour lamps interfere with the bat defence of tympanate moths (Operophtera spp.; Geometridae). Animal Behaviour, 55, 223–226. 10.1006/anbe.1997.0590 [DOI] [PubMed] [Google Scholar]
- Szaz, D. , Horvath, G. , Barta, A. , Robertson, B. A. , Farkas, A. , Egri, A. , … Kriska, G. (2015). Lamp‐lit bridges as dual light‐traps for the night‐swarming mayfly, Ephoron virgo: Interaction of polarized and unpolarized light pollution. PLoS One, 10, e0121194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takatsu, H. , Minami, M. , Tainaka, K.‐I. , & Yoshimura, J. (2012). Spontaneous flash communication of females in an Asian firefly. Journal of Ethology, 30, 355–360. 10.1007/s10164-012-0332-2 [DOI] [Google Scholar]
- Tataroglu, O. , & Emery, P. (2014). Studying circadian rhythms in Drosophila melanogaster . Methods, 68, 140–150. 10.1016/j.ymeth.2014.01.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thancharoen, A. , Srinual, A. , & Laksanawimol, P. (2017). Relationship between synchronous fireflies, Pteroptyx spp. and their firefly trees. International Firefly Symposium 2017.
- Thancharoen, A. (2007). The biology and mating behaviour of an aquatic firefly species, Luciola aquatilis sp. nov. Thancharoen (Coleoptera: Lampyridae) unpublished Ph.D thesis, Mahidol University, Bangkok.
- Théry, M. , Pincebourde, S. , & Feer, F. (2008). Dusk light environment optimizes visual perception of conspecifics in a crepuscular horned beetle. Behavioral Ecology, 19, 627–634. 10.1093/beheco/arn024 [DOI] [Google Scholar]
- Tierney, S. M. , Friedrich, M. , Humphreys, W. F. , Jones, T. M. , Warrant, E. J. , & Wcislo, W. T. (2017). Consequences of evolutionary transitions in changing photic environments. Austral Entomology, 56, 23–46. 10.1111/aen.12264 [DOI] [Google Scholar]
- Tigar, B. J. , & Osborne, P. E. (1999). The influence of the lunar cycle on ground‐dwelling invertebrates in an Arabian desert. Journal of Arid Environments, 43, 171–182. 10.1006/jare.1999.0541 [DOI] [Google Scholar]
- Tyler, J. (2013). What do glow‐worms do on their day off? The diurnal behaviour of adult Lampyris noctiluca (L.) (Coleoptera: Lampyridae). Lampyrid, 3, 23–34. [Google Scholar]
- Ugolini, A. , Boddi, V. , Mercatelli, L. , & Castellini, C. (2005). Moon orientation in adult and young sandhoppers under artificial light. Proceedings of the Royal Society of London B, 272, 2189–2194. 10.1098/rspb.2005.3199 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ugolini, A. , & Chiussi, R. (1996). Astronomical orientation and learning in the earwig Labidura riparia. Behavioural Processes, 36, 151–161. 10.1016/0376-6357(95)00025-9 [DOI] [PubMed] [Google Scholar]
- Urbanski, J. , Mogi, M. , O’Donnell, D. , DeCotiis, M. , Toma, T. , & Armbruster, P. (2012). Rapid adaptive evolution of photoperiodic response during invasion and range expansion across a climatic gradient. American Naturalist, 179, 490–500. 10.1086/664709 [DOI] [PubMed] [Google Scholar]
- van Doorn, W. G. , & Van Meeteren, U. (2003). Flower opening and closure: A review. Journal of Experimental Botany, 54, 1801–1812. 10.1093/jxb/erg213 [DOI] [PubMed] [Google Scholar]
- van Geffen, K. G. , Groot, A. T. , Van Grunsven, R. H. A. , Donners, M. , Berendse, F. , & Veenendaal, E. M. (2015). Artificial night lighting disrupts sex pheromone in a noctuid moth. Ecological Entomology, 40, 401–408. [Google Scholar]
- van Geffen, K. G. , van Eck, E. , de Boer, R. A. , van Grunsven, R. H. A. , Salis, L. , Berendse, F. , & Veenendaal, E. M. (2015). Artificial light at night inhibits mating in a Geometrid moth. Insect Conservation and Diversity, 8, 282–287. [Google Scholar]
- van Geffen, K. G. , van Grunsven, R. H. A. , van Ruijven, J. , Berendse, F. , & Veenendaal, E. M. (2014). Artificial light at night causes diapause inhibition and sex‐specific life history changes in a moth. Ecology and Evolution, 4, 2082–2089. 10.1002/ece3.1090 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Grunsven, R. H. A. , Donners, M. , Boekee, K. , Tichelaar, I. , van Geffen, K. G. , Groenendijk, D. , … Veenendaal, E. M. (2014). Spectral composition of light sources and insect phototaxis, with an evaluation of existing spectral response models. Journal of Insect Conservation, 18, 225–231. 10.1007/s10841-014-9633-9 [DOI] [Google Scholar]
- van Grunsven, R. H. A. , Jähnichen, D. , Grubisic, M. , & Hölker, F. (2018). Slugs (Arionidae) benefit from nocturnal artificial illumination. The Journal of Experimental Zoology, 104, 611. [DOI] [PubMed] [Google Scholar]
- van Langevelde, F. , Braamburg‐Annegarn, M. , Huigens, M. E. , Groendijk, R. , Poitevin, O. , van Deijk, J. R. , … WallisDeVries, M. F. (2017). Declines in moth populations stress the need for conserving dark nights. Global Change Biology, 24, 925–932. 10.1111/gcb.14008. [DOI] [PubMed] [Google Scholar]
- van Langevelde, F. , Ettema, J. A. , Donners, M. , WallisDeVries, M. F. , & Groenendijk, D. (2011). Effect of spectral composition of artificial light on the attraction of moths. Biological Conservation, 144, 2274–2281. 10.1016/j.biocon.2011.06.004 [DOI] [Google Scholar]
- van Langevelde, F. , van Grunsven, R. H. A. , Veenendaal, E. M. , & Fijen, T. P. M. (2017). Artificial night lighting inhibits feeding in moths. Biology Letters, 13, 20160874. 10.1098/rsbl.2016.0874 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Nuffel A., Buijs S. & Delezie E. (2015). Literature review: Effects of different types of artificial light on poultry. Merelbeke: Institute for Agricultural, Fisheries and Food Research (ILVO), 32 pp. [Google Scholar]
- Vänninen, I. , Pinto, D. M. , Nissinen, A. I. , Johansen, N. S. , & Shipp, L. (2010). In the light of new greenhouse technologies: 1. Plant‐mediated effects of artificial lighting on arthropods and tritrophic interactions. Annals of Applied Biology, 157, 393–414. 10.1111/j.1744-7348.2010.00438.x [DOI] [Google Scholar]
- Vencl, F. V. , & Carlson, A. D. (1998). Proximate mechanisms of sexual selection in the firefly Photinus pyralis (Coleoptera: Lampyridae). Journal of Insect Behavior, 11, 191–207. [Google Scholar]
- Vencl, F. V. , Luan, X. , Fu, X. , & Maroja, L. S. (2017). A day‐flashing Photinus firefly (Coleoptera: Lampyridae) from central Panamá: An emergent shift to predator‐free space? Insect Systematics & Evolution, 48, 1–20. 10.1163/1876312X-48022162 [DOI] [Google Scholar]
- Verheijen, F. J. (1960). The mechanisms of the trapping effect of artificial light sources upon animals. Archives Néerlandaises De Zoologie, 13, 1–107. 10.1163/036551660X00017 [DOI] [Google Scholar]
- Villalobos Jiménez, G. J. (2017). The impacts of urbanisation on the ecology and evolution of dragonflies and damselflies (Insecta: Odonata). Unpublished PhD thesis, University of Leeds, United Kingdom.
- Viviani, V. R. , Rocha, M. Y. , & Hagen, O. (2010). Fauna de besouros bioluminescentes (Coleoptera: Elateroidea: Lampyridae; Phengodidae, Elateridae) nos municípios de Campinas, Sorocaba‐Votorantim e Rio Claro‐Limeira (SP, Brasil): Biodiversidade e influência da urbanização. Biota Neotropica, 10, 103–116. 10.1590/S1676-06032010000200013 [DOI] [Google Scholar]
- Wakefield, A. , Broyles, M. , Stone, E. L. , Jones, G. , & Harris, S. (2016). Experimentally comparing the attractiveness of domestic lights to insects: Do LEDs attract fewer insects than conventional light types? Ecology and Evolution, 6(22), 8028–8036. 10.1002/ece3.2527 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wallner, W. E. , & Baranchikov, Y. N. (1992). Response of adult lymantriid moths to illumination devices in the Russian Far East. Forest Entomology, 88, 337–342. [Google Scholar]
- Warrant, E. , & Dacke, M. (2011). Vision and visual navigation in nocturnal insects. Annual Review of Entomology, 56, 239–254. 10.1146/annurev-ento-120709-144852 [DOI] [PubMed] [Google Scholar]
- Warrant, E. J. , Kelber, A. , Gislén, A. , Greiner, B. , Ribi, W. , & Wcislo, W. T. (2004). Nocturnal vision and landmark orientation in a tropical halictid bee. Current Biology, 14, 1309–1318. 10.1016/j.cub.2004.07.057 [DOI] [PubMed] [Google Scholar]
- Warrant, E. J. , Kelber, A. , Wallén, R. , & Wcislo, W. T. (2006). Ocellar optics in nocturnal and diurnal bees and wasps. Arthropod Structure & Development, 35, 293–305. 10.1016/j.asd.2006.08.012 [DOI] [PubMed] [Google Scholar]
- White, P. J. T. (2018). An aerial approach to investigating the relationship between macromoths and artificial nighttime lights across an urban landscape. Journal of Agricultural and Urban Entomology, 34, 1–14. 10.3954/1523-5475-34.1.1 [DOI] [Google Scholar]
- White, R. H. , Xu, H. , Münch, T. A. , Bennett, R. R. , & Grable, E. A. (2003). The retina of Manduca sexta: Rhodopsin expression, the mosaic of green‐, blue‐ and UV‐sensitive photoreceptors, and regional specialization. Journal of Experimental Biology, 206, 3337–3348. 10.1242/jeb.00571 [DOI] [PubMed] [Google Scholar]
- Whitesell, J. J. , & Walker, T. J. (1978). Photoperiodically determined dimorphic calling songs in a katydid. Nature, 274, 887 10.1038/274887a0 [DOI] [Google Scholar]
- Wilson, J. F. , Baker, D. , Cheney, J. , Cook, M. , Ellis, M. , Freestone, R. , … Young, H. (2018). A role for artificial night‐time lighting in long‐term changes in populations of 100 widespread macro‐moths in UK and Ireland: A citizen‐science study. Journal of Insect Conservation, 22, 189 10.1007/s10841-018-0052-1. [DOI] [Google Scholar]
- Wolff, R. J. (1982). Nocturnal activity under artificial lights by the jumping spider Sitticus fasciger . Peckhamia, 2, 32. [Google Scholar]
- Wölfling, M. , Becker, M. C. , Uhl, B. , Traub, A. , & Fiedler, K. (2016). How differences in the settling behaviour of moths (Lepidoptera) may contribute to sampling bias when using automated light traps. European Journal of Entomology, 113, 502–506. 10.14411/eje.2016.066 [DOI] [Google Scholar]
- Xu, K. , DiAngelo, J. R. , Hughes, M. E. , Hogenesch, J. B. , & Sehgal, A. (2011). The circadian clock interacts with metabolic physiology to influence reproductive fitness. Cell Metabolism, 13, 639–654. 10.1016/j.cmet.2011.05.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaoka, K. , & Hirao, T. (1981). Mechanisms of ovipositional behaviour in Bombyx mori: Time‐gating and accumulation of the internal factor. International Journal of Invertebrate Reproduction, 4, 169–180. [Google Scholar]
- Yiu, V. (2012). Effects of artificial light on firefly flashing activity. Insect News, 4, 5–9. [Google Scholar]
- Yoon, T. J. , Kim, D. G. , Kim, S. Y. , Jo, S. I. , & Bae, Y. J. (2010). Light‐attraction flight of the giant water bug, Lethocerus deyrolli (Hemiptera: Belostomatidae), an endangered wetland insect in East Asia. Aquatic Insects, 32, 195–203. [Google Scholar]
- Youthed, G. J. , & Moran, V. C. (1969). The lunar‐day activity rhythm of myrmeleontid larvae. Journal of Insect Physiology, 15, 1259–1271. 10.1016/0022-1910(69)90235-2 [DOI] [Google Scholar]
- Yuen, S. W. , & Bonebrake, T. C. (2017). Artificial night light alters nocturnal prey interception outcomes for morphologically variable spiders. PeerJ, 5, e4070. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Effect size calculations: Dryad https://doi.org/10.5061/dryad.jm8ps6b.