

Abstract
In spite of significant progress in anti-HIV-1 therapy, current antiviral chemo-therapy still suffers from deleterious side effects and emerging drug resistance. Therefore, the development of novel antiviral drugs remains a crucial issue for the fight against AIDS. HIV-1 integrase is a key enzyme in the replication cycle of the retrovirus since it catalyzes the integration of the reverse transcribed viral DNA into the chromosomal DNA. Efforts to develop anti-integrase drugs started during the early nineties, culminating with the recent approval of Raltegravir. The discovery and the development of the styrylquinoline inhibitor class was an important step in the overall process. In this review we have described the key synthetic issues and the structure-activity relationship of this family of integrase inhibitors. Crystallographic and docking studies that shed light on their mechanism of action are also examined.
Keywords: AIDS, HIV-1, integrase, inhibitor, styrylquinoline
Abbreviations
- HIV
Human immunodeficiency virus
- IN
Integrase
- INSTI
integrase strand transfer inhibitor
- INBI
integrase binding inhibitor
- SQL
styrylquinoline
- LTR
long terminal repeat
- DMAP
4-Dimethylaminopyridine
- DCC
Dicyclohexylcarbodiimide
- DMF
Dimethylformamide
- TFA
Trifluoroacetic acid
- DFT
Density functional theory
- RSV
Rous sarcoma virus
- CoMFA
Comparative Molecular Field Analysis
- QSAR
Quantitative structure-activity relationship
- AZT
Azidothymidine.
1. Introduction
Despite ongoing prevention efforts the number of HIV-infected individuals is still increasing mainly in the developing world. For a long time, there have been only two HIV targets for blocking viral replication, reverse transcriptase and protease. Following an initial breakthrough with the design of the fusion inhibitor, Enfuvirtide (Fuzeon®) [1], newer generations of antiretroviral drugs have emerged to complement existing treatment regimens [2]. Small molecules capable of blocking the CCR5 chemokine receptor raised considerable hope [3], nevertheless the most promising novel class of antiretrovirals is probably the integrase inhibitors [4].
The HIV-1 integrase (IN) is a key enzyme in the replication cycle of the retrovirus since it catalyzes the integration of the reverse transcribed viral DNA into the chromosomal DNA. Integration of the HIV-1 DNA ensures stable maintenance of the viral genome and perpetuation of the virus in the host organism, therefore making integrase an attractive target for antiviral chemotherapy. The integration process requires two distinct catalytic steps. During the first one, named 3’-processing, the integrase specifically removes two nucleotides from each 3'-end of the linear viral DNA. The second step consists in a strand transfer following the translocation of the viral DNA into the nucleus of the infected cells. During this step, IN transfers both extremities of the viral DNA into the target DNA by a one-step transesterification reaction, resulting in full-site integration (Figure 1) [5].
Figure 1.

Schematic representation of the two integrase catalytic reactions (3'-processing and strand transfer).
Since the early 90’s, many compounds that inhibit one of the two steps were identified. However, only strand transfer inhibitors (also known as INSTIs) have been shown to be potent antiviral compounds. One such compound, Raltegravir (Isentress®), which was developed based on early studies by Hazuda et al., was approved for clinical use in autumn 2007 as the first antiretroviral drug targeting the viral integrase [6]. Other compounds such as Elvitegravir or GSK1349572 and GSK1265744, are currently in advanced clinical trials [7]. Unfortunately drug-resistance variants have emerged [8]. Moreover, INSTIs differ from the other classes of antiretrovirals in that they lead to the accumulation of reverse transcribed genomes in the form of two LTRs DNA circles. Such accumulation may cause immune signals whose effects are not known [9]. INSTIs may also interfere with endogenous retroelements which replication involves an integration step, with poorly evaluated consequences [9]. Altogether, these observations have prompted not only the search for novel INSTIs but also a reassessment of the potential inhibitory activity of 3'-processing inhibitors. From this viewpoint, stryrylquinolines are a family of compounds that were found to be potent inhibitors of 3’-processing with a significant inhibitory activity against viral replication in cell cultures. In this review, we recapitulate the different steps of the development of this series and the data obtained in vitro and ex vivo that shed light on their mechanism of action.
2. Chemistry and Structure-Activity Relationship of Styrylquinolines in vitro
3’-Processing and strand transfer reactions are both essential for the life cycle of the virus. In both cases the key step is the hydrolysis of a phosphodiester bond. However, because of the high stability of the DNA phosphodiester bond, a highly efficient catalytic process is required to provide the critical rate enhancement needed to cleave this bond [10]. Polynucleotidyl transferases, including HIV-1 IN take advantage of the catalytic effect of two-metal-ion operating in concert to achieve the tremendous rate enhancement required. For instance, in the case of the 3’-5’ exonuclease reaction of Escherichia coli DNA polymerase I, Beese and Steitz [11] have suggested that the phosphodiester bond cleavage is promoted by two divalent cations separated by about 3.9 Å, the first one (M1) initiates the formation of the hydroxide ion, whereas the second (M2) directly ligates the leaving group atom, thereby facilitating the bond breakage step by neutralizing the developing negative charge (Figure 2). Although the exact nature of the metal cations is unknown in the case of HIV-1 integrase the crystal structure of Mg(II)-complexed core domain of HIV-1 IN shows that one Mg2+ ion is coordinated by two of the three critical active site residues (Asp-64 and Asp-116) [12]. It is probable that the second ion binds only in the presence of the DNA substrate [13].
Figure 2.

The “two-metal-ion” enzymatic mechanism for the 3'-5' exonuclease reaction of Escherichia coli DNA polymerase I according to the Beese and Steitz hypothesis [11] and general structure of styrylquinoline inhibitors.
On this basis, polyhydroxylated SQLs which embedded in their structure two chelating subunits were designed to chelate divalent metal ion(s) present in the catalytic core domain of HIV-1 IN. SQLs, exemplified by 1, (Figure 1) were found to be potent HIV-1 IN inhibitors in in vitro experiments. On the other hand they block the replication of HIV-1 in cell culture and are devoid of cytotoxicity [14]. Following this initial finding, a program of systematic chemical modification of the hit compounds was undertaken since receptor based design of novel HIV-1 inhibitors was hampered by the lack of complete three-dimensional bioactive structure of HIV-1 integrase. More than 150 new analogues were prepared and biologically investigated in an attempt to delineate the structure-activity relationship of this family. Modifications were initially performed on the quinoline ring since it was recognized that this moiety constituted the main pharmacophore. Further modifications and elaborate substitutions at various sites of the ancillary ring were next studied. Finally some modulations of the spacer were examined.
2.1. Discovery of the 8-hydroxy-quinoline-7-carboxylic acid- pharmacophore
In line with the initial proposal that a metal ion chelator might act as integrase inhibitor, 8-hydroxyquinoline derivatives were screened in both 3'-processing and strand-transfer assays. However, a complete lack of inhibitory potency was observed with the parent 8-hydroxyquinoline (2a) or 8-hydroxyquinaldine (2b) and with their O-alkylated and O-acylated derivatives. Modulation of the quinoline ring was therefore undertaken taking advantage of the 8-hydroxyl group.
Kolbe-Schmitt carbonatation according to a previously reported procedure provided acids 2a, 2b in 20–30% yields (Scheme 1) [15]. Latter on, an improved 60% yield of 2b was obtained according to a modified protocol in which the potassium phenoxide, formed in anhydrous condition, was treated at 125 ºC under 4 bar of carbon dioxide [16]. Functionalization of the C-2 methyl group required protection of the salicylic moiety. Thus, chemoselective methylation of the carboxylic group was achieved by alkylation using methyl iodide and potassium bicarbonate as base [17]. After protection of the C-8 phenol group as a pivalate ester, selenious anhydride oxidation in pyridine of the fully protected quinaldine 6 and acidic deprotection yielded diacid 7 [18]. Chlorosulfonylation of the C-5 position using neat chlorosulfonic acid at room temperature followed by condensation with morpholine provided the C-5 sulphonamide 4 in 50% yield [19]. Amidification reaction of the C-7 carboxyl group was quite challenging in this series. For example, DMAP/DCC-catalyzed condensation of 3b with 4-fluorobenzylamine delivered 5 in only 26% yield [20]. After many unsuccessful attempts, it was finally found that carboxyl activation using N,N'-carbonyl diimidazole followed by primary amine addition gave an improved 60% yield of quinolinecarboxamide 5 [19].
Scheme 1.

Synthesis of various quinoline and quinaldine derivatives.
Looking for alternative scaffolds, Polanski et al. prepared the quinaldine 10 in which the salicylic moiety was shifted to the opposite side of the quinoline ring using Doebner-Miller condensation of 4-aminosalicylic acid with crotonaldehyde. Furthermore, Polanski studied the introduction of a nitro group at C-7 instead of the carboxylic acid. Direct sulfonation of 8-hydroxyquinoline (2a) gave 11 which was further reacted with sulfonitric acid to yield the 8-hydroxy-7-nitro-quinadine-5-sulfonic acid (12) (Scheme 1) [21].
The biological evaluation of these simple quinoline derivatives on both 3'-processing and strand transfer reactions is depicted in Table 1. Thus, 8-hydroxyquinaldine (2b) was found inactive. Adding a carboxylic acid group at C-7 (3a,b) did not improve the inhibitory potency of the quinoline scaffold. Nevertheless a promising observation was made in this series: quinaldine 4 bearing a salicylic function at C-7, C-8 and a sulphonamide at C-5 exhibited a low but undeniable activity [19]. Replacement of the carboxyl by another strong electro-withdrawing group such as a nitro group (compound 12) suppressed completely the activity [21]. When the salicylic system was moved from the C-7, C-8 position to the C-5, C-6 as in 10, a discrete but significant activity was observed making this compound a promising scaffold [21]. Amide 5 derived from 8-hydroxyquinaldine (2b) was found completely devoid of activity. This result was in strong contrast with the Merck finding that 8-hydroxyquinoline and 7-carboxamide[1,6]naphthyridin-8-ols such as 13 and 14, substituted at C-7 with a large lipophilic residue greatly inhibited strand-transfer [22,23].
Table 1.
HIV-1 IN inhibitory potency of the quinoline scaffold.
| Compound | 3’ProcessingIC50 (μM) | Strand Transfer IC50 (μM) | Reference | |
|---|---|---|---|---|
| 2b | 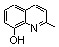 |
>100.000 | >100.000 | 14 |
| 3a |
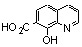
|
>100.000 | >100.000 | 19 |
| 3b |
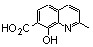
|
>100.000 | 114.000 | 14 |
| 4 |
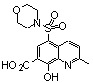
|
77.000 | untested | 19 |
| 7 |
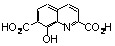
|
>100.000 | untested | 19 |
| 12 |
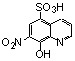
|
>100.000 | untested | 21 |
| 10 |
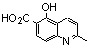
|
47.000 | untested | 21 |
| 5 |

|
>100.000 | untested | 20 |
| 13 |
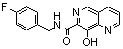
|
0.033 | 23 | |
| 14 |
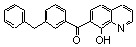
|
0.370 | 22 | |
Thus, despite their similarity, the 2-substituted quinoline derivatives behaved differently than the unsubstituted parent compounds. The origin of this dichotomy is not clear. As a hypothesis, it may be assumed that two different orientations of the oxine in the enzyme core coexist. In the first one, the nitrogen atom of C-2 unsubstituted quinolines such as 13 or 14 fitted in a sterically restricted area, whereas a wide lipophilic space is available for large aromatic groups around the C-7 carbon allowing additional hydrophobic interactions. A simple methyl or carboxyl group at C-2 (as in 5 and 7) impaired such an orientation; hence the quinoline system took an alternative orientation in which a polar group at C-7 such a carboxylic acid greatly improved the binding. These two distinct sites may also account for the different biological properties between the styrylquinoline class which behaved as integrase binding inhibitors (INBIs) and the 8-quinolines and 8-naphthyridines developed by Merck and Shionogi which were found to be highly potent strand-transfer inhibitors (INSTIs).
2.2.Modulation of the C-8 substituent of the styrylquinoline inhibitors
In the quest for more potent inhibitors the introduction of an additional metal-chelating system was thus studied. An aryl nucleus possessing various hydroxy substitution patterns linked by means of an appropriate central spacer was chosen for the second chelating subunit, on the basis of the observation that the most potent HIV-integrase inhibitors known to date generally contained an ortho-dihydroxylated (catechol-type) aromatic ring. All compounds were prepared by condensing the given quinaldine with 3,4-dihydroxybenzaldehyde (16) according to a Perkin-type condensation as depicted in Scheme 2 [14]. Similarly, Lee et al. prepared the corresponding styrylquinazoline 20 in 5% overall yield by condensation of 3,4-dihydroxybenzaldehyde with the 2-methylquinazoline 19 which is easily available from anthranilic acid (18) [24].
Scheme 2.

Synthesis of 8-substituted 3',4'-dihydroxystyrylquinolines.
The influence of the C-8 substituent on the 3',4'-dihydroxystyrylquinoline scaffold is summarized in Table 2.
Table 2.
HIV-1 IN inhibitory potency of various 3',4'-dihydroxystyrylquinolines.
| Compd.Nb |
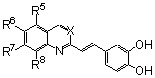
|
3’-Processing IC50 (μM) | References | ||||
|---|---|---|---|---|---|---|---|
| R5 | R6 | R7 | R8 | X | |||
| 21 | H | H | H | H | CH | >100.0 | 14 |
| 22 | H | H | H | OH | CH | 7.4 | 14 |
| 20 | H | H | H | OH | N | 57.0 | 24 |
| 23 | H | H | H | OAc | CH | >100.0 | 14 |
| 24 | H | H | H | NO2 | CH | >100.0 | 14 |
| 25 | H | H | H | NH2 | CH | >100.0 | 14 |
| 26 | H | H | H | CO2H | CH | >100.0 | 25 |
| 27 | CO2H | H | H | H | CH | >100.0 | 25 |
| 28 | H | CO2H | H | H | CH | >100.0 | 25 |
| 29 | H | H | CO2H | H | CH | >100.0 | 25 |
| 30 | CO2H | H | CO2H | H | CH | >100.0 | 25 |
| 31 | OH | CO2H | H | H | CH | >100.0 | 19 |
| 32 | CO2H | H | H | OH | CH | 7.0 | 19 |
Although the parent compound 21 is devoid of activity, the 8-hydroxystyrylquinoline 19 displayed a modest but encouraging in vitro activity. However, the replacement of the free hydroxyl group by an acetoxy substituent completely abolished the activity (compare 22 with 23) [14]. Hence, the inhibitory potency is clearly associated with the presence of “free” phenolic hydroxyl group at C-8 since a nitro or a carboxyl groups at C-8 were unable to assure integrase inhibition. SQLs 23-31 proved to be completely devoid of activity, despite the presence of a catechol moiety in all these compounds [14,25]. Unexpectedly, no noticeable integrase inhibition was observed when an amino group was fixed at C-8, indicating that a simple hydrogen bond donor residue is inefficient in promoting inhibition. The influence of the carboxylic residue in the absence of the 8-hydroxy group was also briefly investigated [25]. Acids 26-29 were found to be completely devoid of integrase inhibition potency, a result indicating that IN affinity seems to be strongly connected with the Mg2+ complexing ability of the oxine moiety. The influence of additional polar group such as carboxyl group at C-5 of the 8-hydroxyquinoline ring as in 32 might be correlated with an increase acidity of the C-8 phenol group which led to higher ion-chelating potency. Finally, modest inhibitory potency was found for the styrylquinazoline (20) compared to SQLs, unfortunately neither the influence of an additional carboxyl group at C-7 nor the antiviral evaluation were reported [24].
2.3.Modulation of the C-7 substituent on the 8,3',4'-trihydroxystyrylquinoline scaffold
The introduction of a polar group at C-7 was the key breakthrough in the styryquinoline class design story. Thus, when an additional nitrile group was introduced at the C-7 position in the quinoline half (compound 34, Table 3) micromolar levels of anti-integrase inhibition were reached. The starting material in the preparation of nitrile 34 was the known 7-carbaldehyde-8-hydroxy-2-methylquinoline which was first converted into oxime 33. Concomitant dehydration of the oxime function of 33 took place during the Perkin condensation with 3,4-dihydroxybenzaldehyde (16), affording carbonitrile 34 in 71% yield. Similarly, styrylquinoline 35 resulted from the condensation between quinaldine 3b and aldehyde 16, followed by hydrolysis of the crude product by using a pyridine-water mixture [14].
Table 3.
HIV-1 IN inhibitory potency of various 7-substituted 8,3’,4’-trihydroxystyrylquinolines.

|
3’-Processing IC50 (μM) | Strand Transfer IC50 (μM) | Reference | |
|---|---|---|---|---|
| 34 | X = CN | 3.0 | Untested | 14 |
| 35 | X = CO2H | 2.4 | 1 | 14 |
| 54 | X = CO2CH3 | >100 | >100 | 26 |
| 42 |
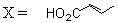
|
2a | 30 | |
| 43 |
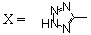
|
Untested | 2.4 | 30 |
| 46 | X = (HO)2PO | Untested | >100 | 30 |
| 47 | X = HO2CCO | 100 | >100 | 32 |
| 53 |
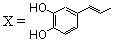
|
2.3 | Untested | 14 |
| 56 |

|
100 | Untested | 30 |
| 55 |
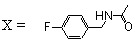
|
>300 | Untested | 19 |
a overall binding assay.
The introduction of a carboxylic acid group at C-7 (compound 35) produced potent HIV-1 IN inhibitors. Furthermore, beside this good in vitro potency, this compound gave rise to a significant antiviral effect as measured by β-gal assay, whereas no toxicity on either infected or non-infected cells was observed. In sharp contrast, compound 54 in which the 7-carboxyl group was replaced by a 7-carbomethoxy group, was completely devoid of biological activity [26]. The latter result clearly indicated that a free carboxylic group greatly improved integrase inhibition, suggesting that a Lewis base containing group at C-7 is need for good inhibition potency. Interestingly enough, an amide function is unable to play such a role, since styrylquinoline carboxamides 55 (Table 3), obtained through direct condensation of the requisite amine with 35,exhibited no noticeable HIV-1 IN inhibition when tested up to 300 μM [19]. The significant loss of HIV-1 IN inhibitory activities of these compounds compared to 35 indicated that the increased size of such compounds seems not suitable to fit into the catalytic pocket of the active site although these compounds possess two pharmacophores that can bind with Mg2+. The importance of the basic nitrogen in the styrylquinoline scaffold has been illustrated by comparison with the corresponding styrylbenzofuran 37 which was found to be 20-fold less potent that 35. The latter was prepared with 20% yield through a Wittig type olefination using phosphonium 36 and 3,4-dihydroxybenzaldehyde (16) [27].
In line with the above results, well known isosters of carboxylic acid such as phosphonic acid and tetrazole group were introduced at the C-7 position of the styrylquinoline scaffold to evaluate their influence (compounds 43 and 46). Furthermore, the distance between the quinoline ring and the acid function introduced at C-7 was modulated by introducing an ethylenic spacer as in 42.
The syntheses of 43 and 46 are depicted in Scheme 4. First, the crucial regioselective bromination of the C-7 position of 8-hydroxyquinaldine 2b was carried out using the tert-BuNH2·Br2 complex [28]. Condensation of 7-bromo-8-hydroxyquinaldine (38) with 3,4-dihydroxybenzaldehyde (16) and protection of the hydroxyl groups gave SQLs 40 and 44 respectively. Stille cross-coupling reaction of bromide 40 with the stannyl tetrazole subunit 41 [29] followed by fluoride deprotection of the silyl groups provided the desired 7-tetrazole-styrylquinoline 43, albeit in modest yield. Similarly the fully protected styrylquinoline 44 was metallated with n-butyllithium and treated with DMF. The resulting aldehyde 45 was condensed with carbomethoxytriphenylmethylenephosphorane and deprotected with BBr3 to provide the unsaturated acid 42 [30]. In turn, phosphonic acid 46 was prepared in 15% overall yield from 44 through sequential lithium bromine exchange with n-butyllithium and treatment of the resulting lithio derivative with diethylchlorophosphate, followed with HBr deprotection [30].
Scheme 4.

Synthesis of C7-modified styrylquinolines 42, 43 and 46.
Unexpectedly, the tetrazole 43 did not show any improved integrase inhibition activity whereas the phosphonic acid 46 was completely inactive. On the other hand, the α,β-unsaturated acid 42 was found equipotent than the parent compound 35, suggesting that the geometric constraints for the additional interaction at the C-7 carbon center were not stringent.
Structurally, compound 47 only differs from lead inhibitor 35 in the presence of an additional keto group at C-7. The factor that has stimulated interest in synthesizing styrylquinoline 47 has been the striking structural analogy between the HOOC-CO-C=C(OH)- system and the enol form of the α,γ-diketo acid pharmacophore found in the potent Merck inhibitor L 731,988 [31].
The introduction of this critical moiety has taken advantage of the one-carbon elongation of esters using the anion of tris(methylthio)methane 49 according to the Scheme 5. In contrast with parent compound 35, this keto acid exhibited a complete lack of in vitro inhibitory potency (IC50 >100 μM) [32]. This result confirmed that structural requirements of INBIs strongly differ from those of diketoacid strand transfer inhibitors. The influence of non-acidic functional groups at the C-7 position of the quinoline half was also studied. Thus, 7-(2-furyl)styrylquinoline 56 available in 24% overall yield by Stille coupling of bromide 38 with 2-(tributylstannyl)furan followed by Perkin-type condensation, was found inactive [30]. This result is in line with the hypothesis of the requirement of a Lewis base atom at the C-7 center. On the other hand the quinoline derivative 53 bearing two similar dihydroxyphenylethenyl subunits at C-2 and C-7 was found equally potent that the lead compound 35 [14], indicating that the catechol group might play the same role that the C-7, C-8 hydroxy acid group and that the exact orientation of the molecule in the active site is still in debate (Table 3).
Scheme 5.

Synthesis of 7-ketoacid 47 and bis-catechol 53.
Since 7-aroyl-8-hydroxyquinoline derivatives such as 14 (Table 1) were found to be highly efficient strand transfer inhibitors by Merck [22], a series of 7-acyl/aroyl SQLs were prepared and evaluated. All these compounds were obtained in 10–40% overall yield from the fully protected 7-bromo-styrylquinoline 44 through sequential lithium/halogen exchange (PhLi, Et2O, -78 ºC), condensation of the resulting 7-lithio derivative with the given aldehyde, MnO2 oxidation and finally demethylation using either HBr at reflux or BBr3 at -78 ºC (Scheme 6) [33].
Scheme 6.

Synthesis of 7-aroyl-styrylquinolines 59.
In sharp contrast with SQLs of type 35 displaying a carboxylic acid group at C-7, most of these compounds proved to be inactive in 3'-processing or overall binding assays as shown in Table 4 probably due to the steric bulk of the C-7 substituent. In line with this hypothesis, the acetyl substituted compound 59d constituted a notable exception. However, this result could not directly be compared since the ancillary half is modified [34]. The acid substituted derivatives 59n-p constituted another exception, reinforcing the previous finding that carboxylic acid residues improved the anti-IN activity [33].
Table 4.
In vitro integrase inhibition activities of the 7-aroyl-styrylquinolines derivatives.

|
3’-Processing IC50 (μM) | Strand Transfer IC50 (μM) | References |
|---|---|---|---|
| 59a, R = H | 82.0 | Untested | 30 |
| 59b, R = Ph | 50.0a | Untested | 33 |
| 59c, R = 1-Naphthyl | 92.0 | Untested | 33 |
| 59da, R = CH3 | 0.2 | 0.2 | 34 |
| 59e, R = C8H17 | >100.0 b | Untested | 33 |
| 59f, R = PhCH2CH2 | >100.0 a | Untested | 33 |
| 59g, R = 4-NO2Ph | 100.0 | >100.0 | 33 |
| 59h, R = 2-NO2Ph | >100.0 | 70.0 | 33 |
| 59i, R = 3,4-F2Ph- | 24.0 | >100.0 | 33 |
| 59j, R = 4-pyridyl | >100.0 a | Untested | 33 |
| 59k, R = 3-pyridyl | >100.0 | >100.0 | 33 |
| 59l, R = 2-OH-Ph | >100.0 | >100.0 | 33 |
| 5m, R = 4-OH-Ph | 65.0 | >100.0 | 33 |
| 59n, R = 4-(HO2C)Ph | 0.2 | Untested | 33 |
| 59o, R = 3-(HO2C)Ph | 0.2 | Untested | 33 |
| 59p, R = 2-(HO2C)Ph | 0.3 | Untested | 33 |
a 1,3,4-trihydroxyphenyl instead of the 3,4-dihydroxyphenyl ancillary ring. b Overall binding assay.
2.4. Functionalization of the C-5 quinoline position
Given the positive influence of a polar group at the C-7 position of the quinoline ring on the anti-integrase potency the introduction of an additional group at the C-5 position was studied. 5,7-Dihalostyrylquinolines 60 and 61 (Table 5) were easily prepared through Perkin condensation of the commercially available 5,7-dihalo-8-hydroxyquinaldines with aldehyde 16 [26]. New structural variants possessing an additional carboxyl group at the C-5 carbon atom, either directly bound to the quinoline ring or through an ethylenic spacer were also prepared. The synthesis of 5,7-quinaldinedicarboxylic acid (69) took at profit the palladium catalyzed alkoxycarbonylation reaction of 7-iodo-quinaldine 67 which is available in two steps from 66. Perkin-type condensation of 69 with 3,4-dihydroxy-5-methoxybenzaldehyde and hydrolysis gave the diacid 63 in 21% overall yield. Heck reaction of bromoquinaldine 70 with methyl acrylate afforded unsaturated ester 71 which was next engaged into Perkin condensation with 16 and hydrolysis to provide the 5-propenoic acid derivative 64 in 32% overall yield [34].
Table 5.
HIV-1 IN inhibitory potency of 5,7-disubstituted -8-hydroxystyrylquinoline derivatives.
| Compd. Numb. |
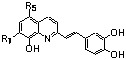
|
In vitro biological activity | Reference | ||
|---|---|---|---|---|---|
| R5 | R7 | 3’-Processing IC50 (μM) | Strand TransferIC50 (μM) | ||
| 60 | Cl | Cl | >100.00 | >100.00 | 26 |
| 61 | Br | Br | 21.00 | 33.00 | 19 |
| 62 | Br | CO2H | 5.00 | Untested | 30 |
| 63a | CO2H | CO2H | 0.20 | 0.07 | 34 |
| 64 |
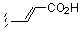
|
CO2H | 1.40 | 1.00 | 34 |
| 65 |
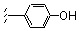
|
CO2H | >100.00c | Untested | 19 |
| 76b | Cl | SO2NHpClPh | >100.00 | Untested | 36 |
a 3,4-dihydroxy-5-methoxyphenyl instead of the 3,4-dihydroxyphenyl ancillary ring b 2,4-dihydroxyphenyl instead of the 3,4-dihydroxyphenyl ancillary ring; c Overall binding assay.
Scheme 7.

Synthesis of C-5 functionalized styrylquinolines 62 and 64.
The 7-arylstyryquinoline 65 was obtained in 22% overall yield using Suzuki cross-coupling of 4-methoxyboronic acid with 5,7-dibromostyrylquinoline 72. The remarkable regioselectivity may have its origin in the low reactivity of the C-7 bromine atom in the oxidative addition step due to the presence of a vicinal methoxy group. Lithium bromine exchange reaction, followed by carbon dioxide condensation with the resulting 7-lithio derivative afforded after HBr deprotection the 5-aryl-styrylquinoline 65 (Scheme 8) [19]. Assuming that a sulfonamide group may engage a H-bonding network as a carboxylic acid, Zen et al. synthetized a series of 2-styrylquinoline-7-sulfonamides exemplified by 76, starting from the 7-chloro-8-hydroxyquinaldine (74), according to a four-step sequence involving chlorosulfonation, amination, Perkin-type condensation with 3,4-dihydroxybenzaldehyde (16) and hydrolysis [36].
Scheme 8.

Synthesis of 5-arylstyrylquinoline 65 and 7-sulfonamide 76.
Not surprisingly, the dichloro and dibromoquinoline derivatives 60 and 61 were found to be either inactive or poorly active. On the other hand, bromoacid 62, which displays a carboxyl group a C-7, showed roughly the same activity that the unsubstitued C-5 derivative 35. The introduction of a carboxyl residue at C-5 either directly (compound 63) or through an ethylenic spacer (compound 64) was highly beneficial to the in vitro activity, since diacid 63 was one of the most potent styrylquinoline prepared so far [34]. On the other hand, sulfonamide 76 prepared by the Zen’s group, turned out to be totally inactive. Clearly the bulky sulfonate group, although it might provide a hydrogen-bond is unable to play the same role that the 7-carboxyl group which is probably ionized at the pH of the integrase assay (Table 5) [36].
2.5. Modulation of the ancillary ring
Following the initial finding that a (3,4-dihydroxyphenyl)ethenyl substituent bound to the C-2 position of the 8-hydroxyquinoline scaffold produced good integrase inhibitor a systematic program of modulation of the ancillary ring was engaged. All compounds were prepared by Perkin-type condensations of the given aldehyde with 8-hydroxy-2-methyl-quinoline-7-carboxylic acid (3b) according to Scheme 2. More than thirty analogues were synthesized and evaluated against integrase.
The results obtained with compounds bearing simple phenyl groups, heteroaromatic and monosubstituted aromatic ancillary groups are depicted in Table 6. Whatever the nature of the ancillary ring, SQLs 77a-i displayed good level of IN inhibition potency. This result seemed indicate that this part of the molecule interacts at least in part, with IN through hydrophobic or pi-type interactions. However, the chlorobenzamide group in 77k, and at a lesser extent the thiomethyl group in 77j were too bulky to fit in the hydrophobic pocket of the enzyme, making these compounds inactive.
Table 6.
Influence of the ancillary ring on the HIV-1 IN inhibitory potency of 8-hydroxy-2-styryl-quinoline-7-carboxylic acids.

|
In vitro Biological activity | Reference | ||
|---|---|---|---|---|
| Compd. nb. | R | 3’-Processing IC50(μM) | Strand Transfer IC50 (μM) | |
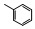
| 77a |
|
5.3 | 2.1 | 14 |
| 77b |
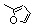
|
1.9 | 5.1 | 26 |
| 77c |

|
3.4 | 3.0 | 26 |
| 77d |

|
4.0 | 11.0 | 26 |
| 77e |
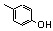
|
1.6 | untested | 26 |
| 77f |
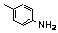
|
2.2 | 3.5 | 26 |
| 77g |
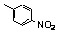
|
1.2 | 1.7 | 26 |
| 77h |
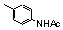
|
3.4 | 31.0 | 26 |
| 77i |
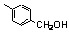
|
5.0 | untested | 25 |
| 77j |
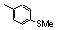
|
39.0 | 30.0 | 30 |
| 77k |
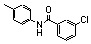
|
>100.0 | untested | 30 |
Results obtained with SQLs di- and trisubstituted on the aromatic ancillary ring are listed in Table 7. Beside the 3,4-dihydroxy pattern, good IN inhibitory activities were also observed with isomers, 77l-n, with a special mention for compound 77n in which the phenyl nucleus is 2,3-dihydroxylated. Replacement of the hydroxyl groups with fluorine atoms reduced the activity significantly but did not suppress it totally, in line with our assumption that hydrophobic interaction is the main factor for in vitro integrase inhibition.
Table 7.
Influence of the ancillary ring on the HIV-1 IN inhibitory potency of 8-hydroxy-2-styryl-quinoline-7-carboxylic acids.
|
In vitro Biological activity | Reference | ||
|---|---|---|---|---|
| Compd. nb. | R | 3’-Processing IC50(μM) | Strand Transfer IC50 (μM) | |
| 35 |
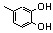
|
2.40 | 1.00 | 14 |
| 77l |
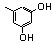
|
3.20 | 3.20 | 26 |
| 77m |
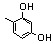
|
3.70 | 2.80 | 14 |
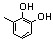
| 77n |
|
0.60 | 0.03 | 26 |
| 77o |
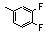
|
10.00 | 30.00 | 25 |
| 77p |
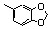
|
5.00 | untested | 25 |
| 77q |
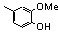
|
2.80 | 3.70 | 26 |
| 77r |
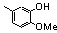
|
1.30 | 3.00 | 26 |
| 77s |
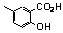
|
2.70 | 0.60 | 14 |
| 77t |
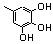
|
0.30 | 0.40 | 14 |
| 77u |
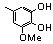
|
0.70 | 2.80 | 26 |
| 77v |
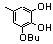
|
5.40 | untested | 26 |
| 77w |
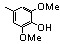
|
4.90 | 4.50 | 26 |
| 77x |
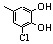
|
0.30 | 0.30 | 37 |
| 77y |
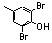
|
1.20 | 1.00 | 26 |
| 77z |
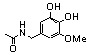
|
2.00 | 2.00 | 19 |
| 77aa |
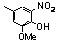
|
7.00 | 4.00 | 19 |
| 77ab |

|
7.00 | 7.00 | 19 |
| 77ac |
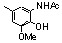
|
10.00 | untested | 19 |
a Overall binding assay.
It is noteworthy that compared with the catechol-containing compound 35, the same level of activity was gained with substrates 77q,r,s in which one of the hydroxyl substituents was replaced by a methoxy or carboxyl group [14,26]. Thus, the latter examples have clearly demonstrated that the design of potent SQLs inhibitors does not necessarily require the presence of an ortho-dihydroxylated (catechol-type) aromatic moiety. An advantageous result, considering the potential stability problems associated with the pharmacological development of catechol-containing drugs.
The 3’,4’,5’-trihydroxyphenyl styrylquinoline 77t was found highly active (3'-processing : 0.3 μM). This confirmed the previously observed trend that groups able to give hydrogen bonding improved the biological activity. Replacement of one hydroxyl by a methoxy group (compound 77u) reduced only slightly the activity. By contrast a large lipophilic chain as in 77v had a more severe effect. The replacement of two of thee hydroxyl groups with bromine atoms (77y) did not reduced drastically the inhibition potency. This result seemed indicate that an increased acidy of the phenol group is beneficial for the IN inhibition [14,26]. A hypothesis reinforced by the good inhibition potency of the chloro derivative 77x [37]. Nitrogen containing derivatives 77aa, ab, ac were found slightly less active.
The screening of a large number of derivatives, have thus demonstrated that the influence of the substitution on the ancillary ring is only moderate. Although hydroxyl groups had clearly a favourable effect, other hydrogen bond donating groups can be equally effective. Thus the choice of a lead compound for further development in the styrylquinoline series will be more surely dictated by the antiviral properties and physico-chemical behaviour than based on the structure-activity relationship of the ancillary ring.
2.6. Modulation of the spacer
In a search for new structural features able to improve the biological profile of this class of inhibitors, the linker between the two aromatic units was modified. First of all the double bond was reduced to increase the conformational liberty between the two aromatic subunits. In an attempt to optimize the binding within the active site of the enzyme, polar functions able to interact through hydrogen bond with near amino acids were intercalated between the two rings, instead of the initial olefinic linker. Thus, compounds 79–82 possessing an amide, a hydrazide or a urea spacer were prepared, keeping constant the 7-CO2H, 8-OH pattern on the quinoline ring (Figure 3) [18]. Synthetically, the reduced quinoline 78 was easily available through catalytic hydrogenation of the corresponding SQL 35 [26]. The others derivatives were prepared using the pivotal acid 84 available in 5 steps from 8-hydroxy-2-methyl-quinoline-7-carboxylic acid (3b). Activation of carboxylic group of 84 using N-hydroxysuccinimide, followed by condensation with the requisite amines 87 provided amides 79 in 15–90% overall yield, after TFA deprotection using guaiacol as tert-butyl cation scavenger.
Figure 3.

Structure of inhibitors exhibiting a nitrogen containing spacer.
Similarly condensation of 85 with hydrazines 88 afforded hydrazides 81 in 55–65% overall yield. On the other hand, Curtius rearrangement of acyl azide 86 and condensation of the intermediate isocyanate with aromatic amines gave the expected urea 82 which were deprotected as previously upon treatment with TFA. Curtius rearrangement in presence of benzyl alcohol, followed by hydrogenolysis of the resulting benzyl carbamate gave the C-2 amino-derivative which was converted to retro-amide 80 by treatment with benzoyl chloride and TFA deprotection (Scheme 9) [18].
Scheme 9.

Synthetic scheme of amides 79, 80, hydrazide 81 and urea 82.
The results of the biological evaluation of these linker-modified compounds keeping constant the 3’,4’-dihydroxyphenyl subunit are depicted in Table 8. The reduction of the central double bond did not affect the integrase inhibition potency (compound 78). In first place, this result demonstrated that the electron circulation from the catechol system towards the quinoline nitrogen, according to a "pull-push effect" was not required for inhibition. Furthermore an increased conformational liberty appeared not to be detrimental to the inhibition potency. Quinoline-2-carboxamide 92, which possessed a two-carbon spacer displayed a slightly improved activity compared to the corresponding styrylquinoline 35, whereas higher the length of the arm lower the activity. In this respect, both ureas 94 and 95 were found inactive (Table 8).
Table 8.
HIV-1 IN inhibitory potency of spacer modified inhibitors.

|
In vitro biological activity | Reference | ||
|---|---|---|---|---|
| Compd nb. | X | 3’-Processing IC50(μM) | Strand Transfer IC50 (μM) | |
| 35 | HC=CH | 2.4 | 1.0 | 14 |
| 78 | H2CCH2 | 2.3 | 1.5 | 26 |
| 92 | CONH | 0.9 | untested | 18 |
| 93 | CONHCH2 | 5.0 | 18 | |
| 94 | HNCONHCH2 | >100.0 | 18 | |
| 95 | HNCONH | >100.0 | 18 | |
To further study the influence of the ancillary ring on spacer-modified compounds, quinolines 96–106 were prepared according to Scheme 9 using various amine and hydrazine derivatives [18]. Dimeric hydrazide 106 was prepared upon treatment of N-hydroxysuccinimide 85 with hydrazine followed by TFA deprotection [19]. The biological evaluation of these new compounds is shown in Table 9. However in no case, were compounds with nitrogen modified spacers found to be more active than the corresponding styrylquinolines. Interestingly enough, the 3,5-dinitrophenylhydrazone derivative 104 was found to me unexpectedly active, although devoid of a hydroxyl group on the right hand. This result confirmed that the catechol moiety is not required for in vitro IN inhibition.
Table 9.
HIV-1 IN inhibitory potency of spacer modified inhibitors.
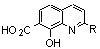
|
In vitro biological activity | ||
|---|---|---|---|
| Compd. Nb. | R | 3’-Processing, IC50 (μM) | 3’-Processing activity of the corresponding SQL IC50 (μM) |
| 96 |
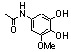
|
6.5 | 0.7 (77u) |
| 97 |
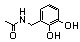
|
5.0 | 0.6 (77n) |
| 98 |
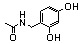
|
5.0 | 3.7 (77m) |
| 99 |
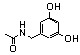
|
>100.0 | 3.2 (77l) |
| 100 |
|
6.5 | 0.7 (77u) |
| 101 |
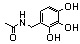
|
1.5 | _ |
| 102 |

|
>100.0 | 5.3 (77a) |
| 103 |

|
>100.0 | 5.3 (77a) |
| 104 |
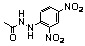
|
3.0 | _ |
| 105 |
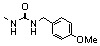
|
>100.0 | _ |
| 106 |
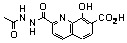
|
2.9 | _ |
3. Physico-Chemical Rational of the Styrylquinoline-Integrase Interaction
3.1. Magnesium binding ability of styrylquinoline integrase inhibitors
Integrase is a 32-kDa protein that consists of three functional domains. The three domains are necessary for all IN activities except the disintegration, an apparent reverse reaction of the strand transfer [38], for which the catalytic core alone or truncated IN are catalytically competent. The core domain and the C-terminal domain display a DNA-binding activity and are therefore potential targets for SQLs. However, early studies have shown that the core domain-mediated disintegration reaction was efficiently inhibited by SQL compounds, with IC50 values similar to those characterizing the 3’-processing reaction, demonstrating that the SQL binding site is within the central domain of IN [14].
As hypothesized for the early development of SQLs, one possible mechanism of action is that they block the polynucleotide binding and catalytic site of HIV-1 IN through functional sequestration of the Mg2+ cofactor by either by the oxine moiety or by their salicylic acid part [26]. With the aim to reinforce this hypothesis, the complexing ability of SQLs for magnesium ions was verified through further experiments including the elaboration and crystallographic characterization of SQL-divalent metal complexes [39]. Unfortunately, attempts to solve the crystal structure of a variety of Mg2+ complexes of styrylquinolines were unsuccessful. Therefore, complexation of 3b (Figure 4) by Mg2+ ion was thus investigated, bearing in mind that previous structure-activity relationship studies [14,26]. and docking procedures [40,41,42]. have identified the salicylic acid moiety of the quinoline half of SQLs as a critical pharmacophore for antiviral activity. The molecular structure of dimeric complex 3b-Mg is depicted in Figure 4. Two tridentate ligands coordinate two Mg2+ cations via the carboxylato O2-atom, the phenoxo O3-atom and the quinoline nitrogen atom. N and O2 atoms are linked to one Mg2+ ion whereas O3 is bonded to the two metal atoms. In order to quantify and further explore the metal-chelating property of 3b a DFT computation of the electronic structure of the complex 3b-Mg was performed [39,43]. The most stable configuration was found for monometallic complex 3b-Mg A where the magnesium cation is linked to both carboxyl and hydroxyl oxygen atoms. Comparatively, the complex 3b-Mg B involving O3 and N atoms, exhibited the smallest binding energy of the Mg cation.
Figure 4.

Structures of 3b and X-ray structures of magnesium complex 3b-Mg. DFT electrostatic potential features of putative magnesium complexes 3b-Mg A, 3b-Mg B. Grey and black isosurfaces correspond to +0.20 and -0.10 a.u., respectively. The arrows indicate the local minimum values of the electrostatic potential.
Although the exact mechanism by which SQL analogues exert their antiviral activity is still a matter of controversy, several studies have showed that such compounds inhibit IN at its interface with viral DNA and divalent metal(s), thereby preventing DNA-IN binding [44]. The ability for progenitor 3b to coordinate magnesium(II) ions is consistent with such a model. We have previously demonstrated through X-ray studies that both drug 77u and its progenitor 3b crystallize in zwitterionic state [43]. Indeed, the carboxylate group of these molecules was found to be ionized in the solid state. This phenomenon, highlighted by the large negative electrostatic potential isosurface surrounding the carboxylate, clearly reinforces the chelating ability of these ligands towards metal cations.
3.2. Comparative molecular field analysis and docking studies
Initial modelling studies were conducted using the Rous sarcoma virus (RSV) integrase core domain [40,41]. Ab initio studies were first performed in order to elucidate the conformation and charge distribution of 77q (Table 7). The s-trans conformation as usually represented in the schemes was found the most stable. The same conformation was observed in the crystal structure [43]. However, the s-cis confirmation was found to be higher in energy by only 0.3 kcal/mol. Thus, we can assume that considerable flexibility around this bond exists in biological medium. Docking of the styrylquinoline 77q shows that the inhibitor binds closely to the Mg2+ cation making contacts with the D64, N122, N149, Q153, N160 residues [40,41]. In a more recent work [44], compound 35 (Scheme 3) was docked on the HIV-1 IN catalytic core. After rigid docking and minimization, the best target-drug complexes were selected according to their interaction energies (coulombic and Van der Waals). Three main cluster sites were identified. The contacting residues of cluster 1 overlap the catalytic triad (Figure 5a). Cluster 2 is actually made of a succession of overlapping sites and is spread over a large region of the protein surface (Figure 5b) whereas a little number of conformations forms the cluster 3 (not shown). As expected, the negatively charged drug bound to lysine-rich regions. Residues K156 and K159 Q148 and S119 in cluster 1 have been previously found to be involved in DNA binding [45,46,47]. In contrast, no residues from cluster 2 are known to be essential for DNA binding. In conclusion, from computational simulations, it seems that cluster 1 is the primary binding site for SQLs as already observed with the RSV core domain. This cluster, which corresponds to one of the two proposed DNA-binding sites, contains the catalytic triad and several key residues for DNA-binding [48].
Scheme 3.

Synthesis of 8-hydroxy-3',4'-dihydroxystyrylquinoline 7-carbonitrile (34), 7-carboxylic acid 35 and benzofuran 37.
Figure 5.

View of one SQ-IN complex typical conformation belonging to each cluster: a, cluster 1; b, cluster 2. The protein is shown in magenta, Mg2+ in green, water in white; drug in red, the catalytic triad (Asp64-Asp116-Glu152) in cyan (sticks and balls) and contacting residues are in yellow.
Using a fluorescence anisotropy-based DNA-binding assay it was confirmed that SQLs prevent recognition of viral by IN, demonstrating that the 3’-processing inhibitory effect is in fact due to the ability to impair DNA binding by competitive inhibition. This mechanism strongly differs from the one of INSTIs which do not bind efficiently to the active site of the free IN but display a strong affinity for the IN/viral DNA complex [31,49].
Furthermore, attempts were made to obtain a Comparative Molecular Field Analysis (CoMFA) model to predict the activity and the rational binding mode of styrylquinoline derivatives to IN [42]. (CoMFA) was performed to analyze three-dimensional quantitative structure-activity relationship (3D-QSAR) of styrylquinoline derivatives. The stability of 3DQSAR models was proved by the analysis of cross-validated and non-cross-validated methods.
4. Antiviral Activity of the Styrylquinoline Integrase Inhibitor Class
All compounds were evaluated for their antiviral activities against HIV-1 replication in either Hela or CEM cells. They were tested for their ability to lower the viral charge in culture supernatants and for their eventual cytoxicity. The results are summarized in Table 10–Table 12.
Table 10.
Antiviral activity and cytotoxicity of the (3’,4’-dihydroxyphenyl)styrylquinoline derivatives.
| Compd. | Antiviral Activity on CEM cells IC50 ( μM) | Cytotoxicity IC50 ( μM) | Reference |
|---|---|---|---|
| 3b | >100.00 | >100.00 | 14 |
| 7 | >100.00 | 500.00 | 19 |
| 14 | 5.00a | 1.25 | 22 |
| 22 | 3.60 | 8.10 | 14 |
| 26 | 12.00 | >100.00 | 25 |
| 27 | 50.00 | >100.00 | 25 |
| 28 | 20.00 | >100.00 | 25 |
| 29 | 20.00 | 20-30.00 | 25 |
| 30 | 15.00 | >100.00 | 25 |
| 32 | 4.00 | 200.00 | 19 |
| 34 | NR | 6.20 | 14 |
| 35 | 1.30 | 120.00 | 14 |
| 42 | 10.00 | 30.00 | 30 |
| 43 | >100.00 | >100.00 | 30 |
| 46 | >100.00 | >100.00 | 30 |
| 47 | 10.00 | 50.00 | 32 |
| 53 | 95.00 | >100.00 | 14 |
| 54 | NR | 9.20 | 26 |
| 59d | NR | 12.50 | 35 |
| 59m | 40.00 | 80.00 | 33 |
| 59n | 63.00 | >100.00 | 33 |
| 59o | 35.00 | >100.00 | 33 |
| 60 | NR | 17.00 | 14 |
| 61 | 0.80 | 10.00 | 19 |
| 62 | 40.00 | 150.00 | 19 |
| 63 | 1.60 | 60.00 | 34 |
| 64 | 35.00 | >100.00 | 34 |
a MT-4 T-lymphoid cells, NR: not reached.
Table 12.
Antiviral activity and cytotoxicity of spacer modified inhibitors.
| Compd. | Antiviral Activity on CEM cells IC50 (μM) | Cytotoxicity IC50 (μM) | Reference | |
|---|---|---|---|---|
| 35 | 1.3 | >100 | 14 | |
| 78 | NR | 61 | 26 | |
| 92 | 30.0 | >100 | 18 | |
| 93 | 2.0 | >100 | 18 | |
| 94 | >100.0 | 70 | 18 | |
| 95 | 55.0 | >100 | 18 | |
| 96 | 2.0 | >100 | 18 | |
| 97 | 40.0 | >100 | 18 | |
| 98 | 10.0 | >100 | 18 | |
| 99 | >100.0 | >100 | 18 | |
| 100 | 25.0 | >100 | 18 | |
| 101 | 4.0 | >100 | 18 | |
| 102 | >100.0 | >100 | 18 | |
| 103 | 45.0 | >100 | 18 | |
| 104 | >100.0 | >100 | 18 | |
| 105 | >100.0 | >100 | 18 | |
| 106 | >100.0a | >100 | 19 |
NR: not reached, a Hela P4.
4.1. Influence of the quinoline substitution of the (3’,4’-dihydroxyphenyl)-substituted styrylquinoline scaffold on the antiviral activity
Simple quinoline derivatives such as compound 3b or 7 were found completely inactive (Table 10). These result markedly contrast with the high antiviral potency of the ketone 14 developed by the Merck Laboratories devoid of any substituent at the C-2 carbon center. Introduction of the (3’,4’-dihydroxyphenyl)ethenyl motive (as in 22) provided some biological activity, although burdened by cytotoxicity. Several compounds in Table 10 displayed moderate antiviral properties while completely devoid of integrase inhibition potency (See for example compounds 26, 28, 29, 30), it may thus be concluded that the styrylquinoline scaffold interacted with other targets within HIV-1. For example the diacid 30 was found to inhibit the HIV-1 RT with IC50 = 10 μM. Such activity may account for the apparent discrepancy observed.
The observation that an additional carboxyl group at the C-7 position of the quinoline nucleus produced efficient antiviral agent was a crucial breakthrough in the design of styryquinoline class. Indeed, styrylquinoline 35 gave rise to a strong antiviral effect as measured by β-gal assay, whereas no toxicity on either infected or noninfected cells was observed. By contrast either a nitrile group (34) or a carbomethoxy group (54) induced strong cytotoxicity masking the antiviral effect. These results confirmed that the 8-hydroxy-quinoline-7-carboxylic acid system is required for both good anti-integrase inhibition and antiviral activity. Since the pKa of the carboxyl group is below 4, it may be assumed that the 7-carboxyl group is ionized under physiological conditions. Accordingly, it appeared that a carboxylate ion at C-7 is required for efficient antiviral activity. Complexation of the magnesium ion within the active site of integrase could tentatively be invoked to explain this result. The favourable influence of an acid group on the activity is evident, however, when the acid is in a position C-5 (compound 32) this effect is less marked. On the other hand, whatever the position of the acid group the cytotoxicity was deeply reduced.
Compounds 43 and 46 (Table 10) bearing at the C-7 position isosters of the carboxyl group were found completely inactive, whereas the acid 42 in which the carboxyl group is bound at C-7 through an ethylenic linker preserved some activity [30]. The α-ketoacid 47 displayed a moderate antiviral activity although devoid of integrase inhibition potency. Finally, the styrylquinoline derivative 53 bearing two similar dihydroxyphenylethenyl subunits was found inactive. This result revealed that contrary to the integrase inhibition structure-activity relationship criteria, ex vivo the available space for the substituent at C-7 is limited.
The dichloro and dibromo derivatives 60 and 61 displayed high cytotoxicity masking the antiviral activity. Curiously, the introduction of a large bromine atom at C-5 (62) induced a dramatic decline of the antiviral potency with respect to the parent compound 35. Similarly, styrylquinolines 64 which displayed an unsaturated acid at C-5 was poorly active. These two results seemed indicate that the available space for a substituent at C-5 is relatively narrow. On the other hand, highly active integrase inhibitor 63 (70 nM on the strand transfer), in which the carboxyl residue was directly bound on the quinoline ring, recovered a good antiviral activity but without significant improvement compared to the level of activity of 35 [38]. Derivatives 59d,m-o which were the only compounds with a significant anti-integrase activity among the 7-keto-styrylquinoline subclass were found either weakly active in cell culture assays (59m,n,o) or cytotoxic (as 59d) [33,35].
4.2. Modulation of the ancillary ring
While all compounds bearing a simple phenyl or a heteroaromatic ring (77a–77d) have proved to be good integrase inhibitors, they were found either toxic or inactive in cell cultures assays. As their unsubstituted counterpart, the monosubstituted styrylquinolines 77e–77j were found inactive, regardless of the nature of the substituent on the ancillary ring (Table 11). As a general rule, only compounds bearing a hydroxyl group at the para position and an other oxygenated function at C-3’ possessed a promising activity. This result confirmed that structural requirement for ex vivo activity are much more stringent that for in vitro integrase inhibition. The higher the number of hydroxyl groups on the ancillary ring, the better the antiviral activity. However, the presence of three hydroxyl groups (77t) induced some cytotoxicity. In this respect, styrylquinoline 77u constituted the best compromise.
Table 11.
Influence of the ancillary ring substitution on antiviral activity.
| Compd. | Antiviral Activity on CEM cells IC50 (μM) | Cytotoxicity IC50 (μM) | Reference |
|---|---|---|---|
| 77e | 95.0 | 100.0 | 26 |
| 77f | 50.0 | 100.0 | 26 |
| 77g | NR | 43.0 | 26 |
| 77h | NR | 47.0 | 26 |
| 77i | 110.0 | >100.0 | 25 |
| 77j | 57.0 | 63.0 | 30 |
| 35 | 1.3 | >100.0 | 14 |
| 77l | >100.0 | >100.0 | 26 |
| 77m | 80.0 | >100.0 | 14 |
| 77n | 30.0 | 110.0 | 19 |
| 77o | 30.0 | 50.0 | 23 |
| 77p | 10.0 | 50.0 | 23 |
| 77q | 3.0 | 95.0 | 26 |
| 77r | 54.0 | 95.0 | 26 |
| 77s | 62.0 | >100.0 | 14 |
| 77t | 1.2 | 10.0 | 14 |
| 77u | 1.5 | 250.0 | 26 |
| 77w | 9.0 | 100.0 | 26 |
| 77x | 36.0 | 22.0 | 37 |
| 77y | 50.0 | >100.0 | 26 |
| 77aa | 6.0a | >100.0 | 19 |
| 77ab | 14.5a | >100.0 | 19 |
| 77ac | 26.0a | 230.0 | 19 |
NR: not reached, a Hela P4 cells.
4.3. Modulation of the spacer
The antiviral evaluation of the linker modified compounds is depicted in Table 12. The reduction of the central double bond which did not affect the integrase inhibition potency dramatically reduced the antiviral activity [26]. Likewise, replacement of the double bond with a simple amide (92), or urea (94,95) drastically reduced activity. In this context, the amide 93 is a noteworthy exception [18]. Modulation of the hydroxyl substituent pattern on the ancillary ring did not modify the picture. Only, 96, 101 and to a lesser extent 98 possessed an antiviral activity.
5. Conclusions
DNA binding assays and computational docking data have highlighted a competitive mechanism between SQLs and the viral DNA end, which leads to the inhibition of the 3’-processing and makes SQLs representative compounds of the INBI group. In cells, the stable IN-DNA interaction within the preintegration complex (PIC) might preclude inhibitors from acting as competitor of viral DNA from being very effective in the cellular context. Together with the dramatic effectiveness of INSTI compounds, this consideration questions the relevance of further developing INBIs.
Yet, the following observations temper this conclusion. First, although most 3'-processing inhibitors actually lack antiviral effect, this is not the case for SQLs, which show a moderate activity ex vivo [44,50,51]. Second, investigation of the mechanism of viral inhibition by compound 77u in cells revealed the possibility that the inability of integrase to bind to the viral DNA impairs the formation of the IN/DNA nucleoprotein complex. Finally, the lead compound 77u proved also to be active against viruses resistant to AZT, lamivudine (3TC), nevirapine, efavirenz and against INSTIs-resistant integrase [50]. In vitro selection of resistant virus identified the two mutants C280Y and (V165I V249I) which differ from those conferring resistance to Raltegravir (N155H, G140S, Q148H, Y143) [52,53]. It is not clear how these mutations promote resistance to SQLs. V165 has been proposed to participate in the nuclear translocation of the integrase [54], a step that might be impaired in cells by SQL as observed in vitro [51]. In agreement with this difference of mechanism of action, SQLs demonstrated an additive to moderate synergic effect with INSTIs [55].
Altogether, these results evidence that INBIs could be used in combination with other antiretroviral drugs. Considering that resistance has already emerged against INSTIs, they justify pursuing the effort to improve SQLs.
References
- 1.Lalezari J.P., Henry K., O'Hearn M., Montaner J.S., Piliero P.J., Trottier B., Walmsley S., Cohen C., Kuritzkes D.R., Eron J.J., Jr., Chung J., DeMasi R., Donatacci L., Drobnes C., Delehanty J., Salgo M. Enfuvirtide, an HIV-1 fusion inhibitor, for drug-resistant HIV infection in North and South America. N. Engl. J. Med. 2003;348:2175–2185. doi: 10.1056/NEJMoa035026. [DOI] [PubMed] [Google Scholar]
- 2.Stellbrink H.-J. Novel compounds for the treatment of HIV type-1 infection. Antivir. Chem. Chemother. 2009;19:189–200. doi: 10.1177/095632020901900502. [DOI] [PubMed] [Google Scholar]
- 3.Hunt J.S., Romanelli F. Maraviroc, a CCR5 coreceptor antagonist that blocks entry of human immunodeficiency virus type 1. Pharmacotherapy. 2009;29:295–304. doi: 10.1592/phco.29.3.295. [DOI] [PubMed] [Google Scholar]
- 4.Pommier Y., Johnson A.A., Marchand C. Integrase inhibitor to treat HIV/AIDS. Nature Rev. Drug Disc. 2005;4:236–247. doi: 10.1038/nrd1660. [DOI] [PubMed] [Google Scholar]
- 5.Lewinski M.K., Bushman F.D. Retroviral DNA integration-mechanism and consequences. Adv. Genet. 2005;55:147–181. doi: 10.1016/S0065-2660(05)55005-3. [DOI] [PubMed] [Google Scholar]
- 6.Chirch L.M., Morrison S., Steigbigel R.T. Treatment of HIV infection with raltegravir. Expert Opin. Pharmaco. 2009;10:1203–1211. doi: 10.1517/14656560902911488. [DOI] [PubMed] [Google Scholar]
- 7.Vandeckerckhove L. GSK-1349572, a novel integrase inhibitor for the treatment of HIV infection. Curr. Opin. Investig. Drugs. 2010;11:203–212. [PubMed] [Google Scholar]
- 8.Katlama C., Murphy R. Emerging role of integrase inhibitors in the management of treatment-experienced patients with HIV infection. Ther. Clin. Risk Manag. 2009;5:331–340. doi: 10.2147/tcrm.s3137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Beck-Engeser G.B., Eilat D., Harrer D., Jäck H.-M., Wabl M. Early onset of autoimmune disease by the retroviral integrase inhibitor raltegravir. Proc. Natl. Acad. Sci. USA. 2009;106:20865–20870. doi: 10.1073/pnas.0908074106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Williams N.H., Takasaki B., Wall M., Chin J. Structure and nuclease activity of simple dinuclear metal complexes: Quantitative dissection of the role of metal ions. Acc. Chem. Res. 1999;32:485–493. [Google Scholar]
- 11.Beese L.S., Steitz T.A. Structural basis for the 3'-5' exonuclease activity of Escherichia coli DNA polymerase I: a two metal ion mechanism. EMBO J. 1991;10:25–33. doi: 10.1002/j.1460-2075.1991.tb07917.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Goldgur Y., Dyda F., Hickman A.B., Jenkins T.M., Craigie R., Davies D.R. Three new structures of the core domain of HIV-1 integrase: an active site that binds magnesium. Proc. Natl. Acad. Sci. USA. 1998;95:9150–9154. doi: 10.1073/pnas.95.16.9150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lins R.D., Adesokan A., Soares T.A., Briggs J.M. Investigations on human immunodeficiency virus type 1 integrase/DNA binding interactions via molecular dynamics and electrostatics calculations. Pharmacol. Therapeut. 2000;85:123–131. doi: 10.1016/s0163-7258(99)00059-5. [DOI] [PubMed] [Google Scholar]
- 14.Mekouar K., Mouscadet J.-F., Desmaële D., Subra F., Savouré D., Auclair C., d'Angelo J. Styrylquinoline derivatives: a new class of potent HIV-1-integrase inhibitors that block HIV-1 replication in CEM cells. J. Med. Chem. 1998;41:2846–2857. doi: 10.1021/jm980043e. [DOI] [PubMed] [Google Scholar]
- 15.Meek W.H., Fuschman C.H. Carboxylation of substituted phenol in N,N-dimethylamide solvents at atmospheric pressure. J. Chem. Eng. Data. 1969;14:388–391. doi: 10.1021/je60042a005. [DOI] [Google Scholar]
- 16.Baret P., Béguin C.G., Boukhala H., Caris C., Laulhère J.-P., Pierre J.-L., Serratrice G. O-TRENSOX: A promising water-soluble iron chelator (Both FeIII and FeII) potentally suitable for plant nutrition and iron chelation therapy. J. Am. Chem. Soc. 1995;117:9760–9761. [Google Scholar]
- 17.Guo W., Li J., Fan N., Wu W., Zhou P., Xia C. A simple and effective method for chemoselective esterification of phenolic acids. Synth. Commun. 2005;35:145–152. [Google Scholar]
- 18.Bénard C., Zouhiri F., Normand-Bayle M., Danet M., Desmaële D., Leh H., Mouscadet J.-F., Mbemba G., Thomas C.-M., Bonnenfant S., Le Bret M., d’Angelo J. Linker-Modified Quinoline Derivatives Targeting HIV-1 Integrase: Synthesis and Biological Activity. Bioorg. Med. Chem. Lett. 2004;14:2473–2476. doi: 10.1016/j.bmcl.2004.03.005. [DOI] [PubMed] [Google Scholar]
- 19.Desmaële D., Zouhiri F. Unpublished Results.
- 20.Polanski J., Niedbala H., Musiol R., Podeszwa B., Tabak D., Palka A., Mencel A., Finster J., Mouscadet J.-F., Le Bret M. Fragment based approach for the investigation of HIV-1 integrase inhibition. Lett. Drug Des. Discovery. 2007;4:99–105. doi: 10.2174/157018007779422532. [DOI] [Google Scholar]
- 21.Polanski J., Niedbala H., Musiol R., Podeszwa B., Tabak D., Palka A., Mencel A., Finster J., Mouscadet J.-F., Le Bret M. 5-hydroxy-6-quinaldic acid as a novel molecular scaffold for HIV-1 integrase inhibitors. Lett. Drug Des. Discovery. 2006;3:175–178. [Google Scholar]
- 22.Zhuang L., Wai J.S., Embrey M.W., Fisher T.E., Egbertson M.S., Payne L.S., Guare J.P., Jr., Vacca J.P, Hazuda D.J., Felock P J., Wolfe A.L., Stillmock K.A., Witmer M.V., Moyer G., Schleif W.A., Gabryelski L.J., Leonard Y.M., Lynch J.J., Jr., Michelson S.R., Young S.D. Design and synthesis of 8-hydroxy-[1,6]naphthyridines as novel inhibitors of HIV-1 integrase in vitro and in infected cells. J. Med. Chem. 2003;46:453–456. doi: 10.1021/jm025553u. [DOI] [PubMed] [Google Scholar]
- 23.Guare J.P., Wai J.S., Gomez R.P., Anthony N.J., Jolly S.M., Cortes A.R., Vacca J.P., Felock P.J., Stillmock K.A., Schleif W.A., Moyer G., Gabryelski L.J., Jin L.X., Chen I.W., Hazuda D.J., Young S.D. A series of 5-aminosubstituted 4-fluorobenzyl-8-hydroxy-[1,6]naphthyridine-7-carboxamide HIV-1 integrase inhibitors. Bioorg. Med. Chem. Lett. 2006;16:2900–2904. doi: 10.1016/j.bmcl.2006.03.003. [DOI] [PubMed] [Google Scholar]
- 24.Lee J.Y., Park J.H., Lee S.J., Park H., Lee Y.S. Styrylquinazoline derivatives as HIV-1 integrase inhibitors. Arch. Pharm. 2002;335:277–282. doi: 10.1002/1521-4184(200208)335:6<277::AID-ARDP277>3.0.CO;2-A. [DOI] [PubMed] [Google Scholar]
- 25.Polanski J., Zouhiri F., Jeanson L., Desmaële D., d'Angelo J., Mouscadet J. F., Gieleciak R., Gasteiger J., Le Bret M. Use of the Kohonen neural network for rapid screening of ex vivo anti-HIV activity of styrylquinolines. J. Med. Chem. 2002;45:4647–4654. doi: 10.1021/jm020845g. [DOI] [PubMed] [Google Scholar]
- 26.Zouhiri F., Mouscadet J.-F., Mekouar K., Desmaële D., Savouré D., Leh H., Subra F., Le Bret M., Auclair C., d'Angelo J. Structure-activity relationships and binding mode of Styrylquinolines as potent inhibitors of HIV-1-integrase and replication of HIV-1 in cell culture. J. Med. Chem. 2000;43:1533–1540. doi: 10.1021/jm990467o. [DOI] [PubMed] [Google Scholar]
- 27.Yoo H., Lee J.Y., Park J.H., Chung B.Y., Lee Y.S. Synthesis of styrylbenzofuran derivatives as styrylquinoline analogues for HIV-1 integrase inhibitors. Farmaco. 2003;58:1243–1250. doi: 10.1016/j.farmac.2003.08.001. [DOI] [PubMed] [Google Scholar]
- 28.Pearson D.E., Wysong R.D., Breder C.V.J. The ortho bromination of phenols. J. Org. Chem. 1967;32:2358–2360. doi: 10.1021/jo01282a063. [DOI] [Google Scholar]
- 29.Bookser B.C. 2-Benzyloxymethyl-5-(tributylstannyl)tetrazole. A reagent for the preparation of 5-aryl- and 5-heteroaryl-1H-tetrazoles via the Stille reaction. Tetrahedron Lett. 2000;41:2805–2809. doi: 10.1016/S0040-4039(00)00268-9. [DOI] [Google Scholar]
- 30.Normand-Bayle M. Synthèse et activité biologique de nouvelles styrylquinoléines inhibitrices de l’intégrase du VIH-1. Ph.D. Dissertation, Université Paris-Sud : Paris, France, 2007. [Google Scholar]
- 31.Espeseth A.S., Felock P., Wolfe A., Witmer M., Grobler J., Anthony N., Egbertson M., Melamed J.Y., Young S., Hamill T., Cole J.L., Hazuda D.J. Proc. Natl. Acad. Sci. USA. 2000;97:11244–11249. doi: 10.1073/pnas.200139397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Zouhiri F., Desmaële D., d'Angelo J., Ourevitch M., Mouscadet J.-F., Leh H., Le Bret M. HIV-1 replication inhibitors of the styrylquinoline class: incorporation of a masked diketo acid pharmacophore. Tetrahedron Lett. 2001;42:8189–8192. [Google Scholar]
- 33.Normand-Bayle M., Bénard C., Zouhiri F., Mouscadet J.-F., Leh H., Thomas C.-M., Mbemba G., Desmaële D., d’Angelo J. New HIV-1 replication inhibitors of the styryquinoline class bearing aroyl/acyl groups at the C-7 position: Synthesis and biological activity. Bioorg. Med. Chem. Lett. 2005;15:4019–4022. doi: 10.1016/j.bmcl.2005.06.036. [DOI] [PubMed] [Google Scholar]
- 34.Zouhiri F., Danet M., Bénard C., Normand-Bayle M., Mouscadet J.-F., Leh H., Thomas C.-M., Mbemba G., d'Angelo J., Desmaële D. HIV-1 replication inhibitors of the styrylquinoline class: Introduction of an additional carboxyl group at the C-5 position of the quinoline. Tetrahedron Lett. 2005;46:2201–2205. [Google Scholar]
- 35.Michaut M., Monneret C., Soma E., Thibault L., Wermuth C. Styrylquinolines, their process of preparartion and their therapeutic uses. WO/2010/010148 A1. PCT Int. Appl. 2010
- 36.Zeng C., Niu L., Ping D., Zhong R. Design and synthesis of 2-styrylquinoline-7-sulfonamide derivatives as potential HIV integrase inhibitors. Chin. J. Org. Chem. 2009;29:1105–1114. [Google Scholar]
- 37.Auclair C., Giethlen B., Michaut M., Monneret C., Soma E., Thibault L., Wermuth C. Styrylquinolines, their process of preparartion and their therapeutic uses. WO/2010/010147 A1. PCT Int. Appl. 2010
- 38.Chow S.A., Vincent K.A., Ellison V., Brown P.O. Reversal of integration and DNA splicing mediated by integrase of human immunodeficiency virus. Science. 1992;255:723–726. doi: 10.1126/science.1738845. [DOI] [PubMed] [Google Scholar]
- 39.Courcot B., Firley D., Fraisse B., Becker P., Gillet J. M., Pattison P., Chernyshov D., Sghaier M., Zouhiri F., Desmaele D., d'Angelo J., Bonhomme F., Geiger S., Ghermani N.E. Crystal and electronic structures of magnesium(II), copper(II), and mixed magnesium(II)-copper(II) complexes of the quinoline half of styrylquinoline-type HIV-1 integrase inhibitors. J. Phys. Chem. 2007;B111:6042–6050. doi: 10.1021/jp0687551. [DOI] [PubMed] [Google Scholar]
- 40.Ouali M., Laboulais C., Leh H., Gill D., Desmaele D., Mekouar K., Zouhiri F., d'Angelo J., Auclair C., Mouscadet J.-F., Le Bret M. Modeling of the inhibition of retroviral integrases by styrylquinoline derivatives. J. Med. Chem. 2000;43:1949–1957. doi: 10.1021/jm9911581. [DOI] [PubMed] [Google Scholar]
- 41.Ouali M., Laboulais C., Leh H., Gill D., Xhuvani E., Zouhiri F., Desmaele D., d'Angelo J., Auclair C., Mouscadet J.-F., Le Bret M. Tautomers of styrylquinoline derivatives containing a methoxy substituent: computation of their population in aqueous solution and their interaction with RSV integrase catalytic core. Acta Biochim. Pol. 2000;47:11–22. [PubMed] [Google Scholar]
- 42.Ma X.-h., Zhang X.-y., Tan J.-j., Chen W;-z, Wang C.-x. Exploring binding mode for styrylquinoline HIV-1 integrase inhibitors using comparative molecular field analysis docking studies. Acta Pharmacol. Sin. 2004;25:950–958. [PubMed] [Google Scholar]
- 43.Firley D., Courcot B., Gillet J.M., Fraisse B., Zouhiri F., Desmaele D., d'Angelo J., Ghermani N.E. Experimental/theoretical electrostatic properties of a styrylquinoline-type HIV-1 integrase inhibitor and its progenitors. J. Phys. Chem. B. 2006;110:537–547. doi: 10.1021/jp0582179. [DOI] [PubMed] [Google Scholar]
- 44.Deprez E., Barbe S., Kolaski M., Leh H., Zouhiri F., Auclair C., Brochon J.-C., Le Bret M., Mouscadet J.-F. Mechanism of HIV-1 integrase inhibition by styrylquinoline derivatives in vitro. Mol. Pharmacol. 2004;65:85–98. doi: 10.1124/mol.65.1.85. [DOI] [PubMed] [Google Scholar]
- 45.Jenkins T.M., Esposito D., Engelman A., Craigie R. Critical contacts between HIV-1 integrase and viral DNA identified by structure-based analysis and photo-crosslinking. EMBO J. 1997;16:6849–6859. doi: 10.1093/emboj/16.22.6849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Esposito D., Craigie R. Sequence specificity of viral end DNA binding by HIV-1 integrase reveals critical regions for protein-DNA interaction. EMBO J. 1998;17:5832–5843. doi: 10.1093/emboj/17.19.5832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Harper A.L., Skinner L.M., Sudol M., Katzman M. Use of patient-derived human immunodeficiency virus type 1 integrases to identify a protein residue that affects target site selection. J. Virol. 2001;75:7756–7762. doi: 10.1128/JVI.75.16.7756-7762.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Perryman A.L., McCammon J.A. AutoDocking dinucleotides to the HIV-1 integrase core domain: exploring possible binding sites for viral and genomic DNA. J. Med. Chem. 2002;45:5624–5627. doi: 10.1021/jm025554m. [DOI] [PubMed] [Google Scholar]
- 49.Hazuda D.J., Felock P., Witmer M., Wolfe A., Stillmock K., Grobler J.A., Espeseth A., Gabryelski L., Schleif W., Blau C., Miller M.D. Inhibitors of strand transfer that prevent integration and inhibit HIV-1 replication in cells. Science. 2000;287:646–650. doi: 10.1126/science.287.5453.646. [DOI] [PubMed] [Google Scholar]
- 50.Bonnenfant S., Thomas C.M., Vita C., Subra F., Deprez E., Zouhiri F., Desmaele D., d'Angelo J., Mouscadet J.-F., Leh H. Styrylquinolines, integrase inhibitors acting prior to integration: a new mechanism of action for anti-integrase agents. J. Virol. 2004;78:5728–5736. doi: 10.1128/JVI.78.11.5728-5736.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Mousnier A., Leh H., Mouscadet J.-F., Dargemont C. Nuclear import of HIV-1 integrase is inhibited in vitro by styrylquinoline derivatives. Mol. Pharmacol. 2004;66:783–788. doi: 10.1124/mol.104.001735. [DOI] [PubMed] [Google Scholar]
- 52.Malet I., Delelis O., Valantin M.-A., Montes B., Soulie C., Wirden M., Tchertanov L., Peytavin G., Reynes J., Mouscadet J.-F., Katlama C., Calvez V., Marcelin A.-G. Mutations associated with failure of raltegravir treatment affect integrase sensitivity to the inhibitor in vitro. Antimicrob. Agents Chemother. 2008;52:1351–1358. doi: 10.1128/AAC.01228-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Delelis O., Thierry S., Subra F., Simon F., Malet I., Alloui C., Sayon S., Calvez V., Deprez E., Marcelin A.-G., Tchertanov L., Mouscadet J.-F. Impact of Y143 HIV-1 integrase mutations on resistance to raltegravir in vitro and in vivo. Antimicrob. Agents Chemother. 2010;54:491–501. doi: 10.1128/AAC.01075-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Dvorin J.D., Bell P., Maul G.G., Yamashita M., Emerman M., Malim M.H. Reassessment of the roles of integrase and the central DNA flap in human immunodeficiency virus type 1 nuclear import. J. Virol. 2002;76:12087–12096. doi: 10.1128/JVI.76.23.12087-12096.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Thibault L., Rochas S., Dourlat J., Monneret C., Carayon K., Deprez E., Mouscadet J.F., Soma E., Lebel-Binay S. New integrase binding inhibitors acting in synergy with raltegravir. Antivir. Ther. 2009;14:A29. abstract 27. [Google Scholar]