

Abstract
Nature endowed snakes with a lethal secretion known as venom, which has been fine-tuned over millions of years of evolution. Snakes utilize venom to subdue their prey and to survive in their natural habitat. Venom is known to be a very poisonous mixture, consisting of a variety of molecules, such as carbohydrates, nucleosides, amino acids, lipids, proteins and peptides. Proteins and peptides are the major constituents of the dry weight of snake venoms and are of main interest for scientific investigations as well as for various pharmacological applications. Snake venoms contain enzymatic and non-enzymatic proteins and peptides, which are grouped into different families based on their structure and function. Members of a single family display significant similarities in their primary, secondary and tertiary structures, but in many cases have distinct pharmacological functions and different bioactivities. The functional specificity of peptides belonging to the same family can be attributed to subtle variations in their amino acid sequences. Currently, complementary tools and techniques are utilized to isolate and characterize the peptides, and study their potential applications as molecular probes, and possible templates for drug discovery and design investigations.
Keywords: polypeptide structure and function, therapeutic peptides, X-ray crystallography, three-finger toxins, bradykinin potentiating peptides, crotamine, kunitz-type inhibitor
1. Introduction
Nature has always been an intriguing source for drug design investigations. From among the animal kingdom, insects and reptilians are interesting and valuable sources for identifying molecules with potential pharmaceutical applications [1]. To date, various bioactive proteins and peptides have been reported from the venom of different species of snakes, conus, scorpions, centipedes, lizards, spiders, sea anemones, bees and octopus [1,2,3,4,5,6,7,8,9]. In this context, venomous snakes are a focus of investigations, and a large number of pharmacologically useful molecules have already been isolated and characterized from snake venoms [10,11]. Among the approximately 3000 known snake species, only a relatively small number are venomous, mainly belonging to Viperidae and Elapidae families. Some members of Colubridae and Atractaspididae are also reported to be venomous [12].
Snake venom is a glandular secretion which snakes use to immobilize and digest their prey. It is also used as a defensive and a survival tool [13]. This lethal mixture is composed of amino acids, nucleic acids, carbohydrates, lipids, proteins and peptides [14]. Figure 1 (adapted from our own reference) illustrates the composition of snake venom [15,16].
Figure 1.

Snake venom composition.
In the present review, we focus our discussion on snake venom peptides, their classification and bioactivities. We define snake venom peptides as the group of non-enzymatic polypeptides in the venom which fold into monomeric domains and are smaller than 80–100 residues. However, variations can be found. For example, three-finger toxins and disintegrins [17,18,19] can exist as dimers. Among three-finger toxins, haditoxin [Protein Data Bank (PDB) code: 3HH7] and ҝ-bungarotoxins (PDB code: 2ABX) are non-covalent dimers, and irditoxin (PDB code: 2H7Z) and α-cobratoxin (PDB code: 2CTX) are covalent dimers [20,21]. Nonetheless, we shall not classify these dimers separately, because they bear structural and functional similarities to their monomeric counter parts.
Peptides are an interesting component of the snake venom. Although otherwise poisonous, when used in the right proportions or structurally engineered, several venom peptides can be used directly as therapeutic drugs or as drug leads, Table 1 [22,23,24,25,26]. These peptides are of great value, due to their diversified and distinct pharmacological activity, and high affinity and selectivity towards their receptors. For example, snake venom toxins have proved to be invaluable tools in determining the structure and functions of receptors. An excellent example in this regard are α-neurotoxins [27]. Snake venom peptides are mostly stable molecules, able to survive the harsh proteolytic environment of the venom gland itself. Also, it is the inherent stability of these peptides that helps them reach their target receptors, inside their prey (upon envenomation). The stability of these molecules is attained when they are recruited into the venom gland through disulfide bonds formation and/or post-translational modifications [28].
Table 1.
| Peptide | Mechanism of Action | Biological Significance/Therapeutics | Reference |
|---|---|---|---|
| 3FTX (neurotoxin) | Selective inhibition of nAChRs at neuromuscular junction and interfere with nerve transmission. | Tool to decipher structural and functional details of nAChRs. α-cobratoxin is under clinical trial for drug-resistant HIV strains, treatment of multiple sclerosis, muscular dystrophy, myasthenia gravis and amyotrophic lateral sclerosis. | [31] |
| 3FTX (cardiotoxin) | Membrane perturbation by electrostatic and hydrophobic interactions with the cell membranes. | Under scientific investigation for cancer inhibitory studies and potential use as anti-microbial agent. | [32,33] |
| Disintegrin | Selectively bind to integrin receptors present at the surface of platelet and other cells. | Tirofiban and Eptifibatide are under clinical use as antithrombotic agents. These compounds were developed from the snake venom disintegrns echistatin and barbourin. Contortrostatin is in preclinical studies for the inhibition of platelet aggregation and prostate cancer. | [34,35,36,37,38] |
| Kunitz-type inhibitor | Inhibition of serine proteases (e.g., plasmin, kallikrein, trypsin). Interferes with the blood coagulation cascade and fibrinolysis. | A plasmin inhibitor Textilinin-1 is in preclinical studies as antibleeding agent. | [39,40] |
| Natriuretic peptide | Interaction of Nps with guanylyl cyclase receptors leads to an increase of cylic guanosine monophosphate (cGMP), and affects subsequent signalling cascade. Nps can interfere the renin-angiotensin system by inhibiting the angiotensin converting enzyme. | These peptides serve as tool to understand NP biology. Cenderitide was under clinic studies for cardiovascular disease. However, its clinical development was terminated by Capiricor (US pharmaceutical company) in 2017. | [41,42,43] |
| BPPs | Inhibit the function of angiotensin converting enzyme, and raise the level of bradykinin. | Captopril and its analogue are under clinical use for the treatment of hypotension. These compounds were developed from the snake venom BPP. | [44] |
| Crotamine | Interacts electrostatically with DNA. Penetrates membranes via heparan sulphate proteoglycans binding. | Carrier for biomolecules, tool for cancer studies. | [45] |
| Sarafotoxin | Vasoconstriction via endothelin receptors. | Molecular probe to better understand endothelial system and related diseases. | [46] |
| Waglerin | nAChR antagonist. | Anti-wrinkle cosmetic cream SYN-AKE is available in the market. The active ingredient of this cream is a peptide mimic, which was designed using waglerin as a template. | [47] |
Based on the available data, snake venom peptides can be grouped into several families, discussed in Section 2. Table 1 and Table 2 present a brief structural and functional summary of these peptides.
Table 2.
Snake venom peptides with well-defined three-dimensional structure.
| Toxin Family | Representative Structure | Representative Sequences |
|---|---|---|
| 3FTX | 1UG4
|

|
| Kunitz-type Inhibitor | 3BYB
|
|
| Disintegrin | 1J2L
|

|
| Crotamine | 4GV5
|
|
| Sarafotoxin | 2LDE
|

|
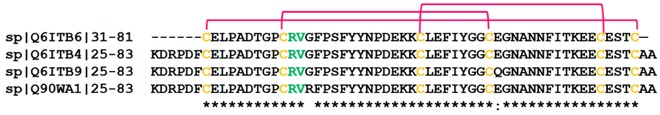
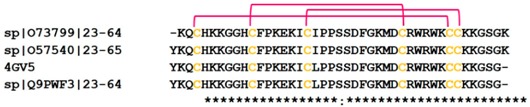
Protein Data Bank ID’s of snake venom peptide are given, along with their three-dimensional structure, in the second column. The third column presents sequence alignment prepared in Clustal W. Sequence alignment of the toxins having high homology with that of the sequence of the toxin in the second column were selected for alignment. Cysteines are colored yellow. Reactive bond residues are shown in green. The pink lines illustrate disulfide bond connectivity.
Applying advanced analytical tools and methodologies is important to investigate and unravel the complexities of these venomous mixtures. Today, several integral approaches and complementary techniques are applied to isolate, characterize and explore the structure-function relationships of snake venom peptides. Advancements in proteomics, multidimensional chromatographic and mass spectrometric methods, requiring only small amounts of sample, have paved the way towards the discovery and characterization of these peptides. High-resolution X-ray crystallographic techniques provide insights into the three-dimensional structures of snake venom peptides and their complexes with proteins. Also, chemical and bioassays are of utmost importance in leading towards the exploration of novel functions of these biomolecules. In addition to the above-mentioned experimental methods, bioinformatics and molecular modeling approaches allow scientists to study the interaction of snake venom peptides with proteins and other complex environments and are therefore an important tool for rational drug design investigations.
2. Snake Venom Peptides and Their Potential Pharmacological Applications
Technological advancement and continuous interest in venomous molecules have supported the discovery of a large number of snake venom peptides. The snake venom peptides are classified into distinct families based on the structural and functional similarities in the organization of these molecules.
2.1. Three-Finger Toxins (3FTxs)
The 3FTxs family of polypeptides is comprised of 60–74 amino acid residues. These peptides show diverse functionalities, however, having a conserved structure. A distinct structural feature of 3FTxs is the unique fold, consisting of three loops (β-stranded), emerging from a hydrophobic globular core, Table 2. Four to five disulfide bonds are present in these 3FTxs, thereby stabilizing the three-dimensional structure. Subtle variations in the length of their loops, conformations and amino acid residues are responsible for their distinct biological functions [48]. Based on their function, these snake venom peptides can be classified into the groups described in Section 2.1.1, Section 2.1.2, Section 2.1.3, Section 2.1.4, Section 2.1.5 and Section 2.1.6.
Proteomic and transcriptomic analysis has shown that the ratio of 3FTxs can be higher relative to other toxins in the Elapidae venoms, as shown, for example, in the venom of Naja ashei, Micrurus pyrrhocryptus and Micrurus t. tschudii [49,50,51,52,53]. 3FTxs constitute more than 60% of the venom composition of cobra snakes [54,55,56,57], while there is a wide distribution of relative abundance of 3FTxs among different krait species, ranging from as low as 1.3% in B. fasciatus [58,59] to as high as 60% in Laticauda colubrina (a sea krait) [60,61]. The proteomic study of mamba venoms revealed that 3FTxs are also one of the most abundant toxins in their venom [50,62,63,64,65]. Proteomic studies have also shown that a combination of different 3FTxs can be present in the venom of the same species [58]. Although these peptides primarily exist in the venom of the Elapinae subfamily, however, they have also been reported for venoms of hydrophiids, colubrids, viperids and crotalids, either at the transcriptome or protein level [66,67,68].
Evolutionary studies on 3FTxs showed that these peptides evolved from genes encoding non-toxic ancestral proteins, as the three-finger scaffold can be found in other non-venom proteins [69,70]. It was reported by Fry and colleagues that three-finger neurotoxins are evolving under positive selection in line with the receptors of prey species, while the evolution of three-finger cytotoxins is limited by negative selection, as they interact nonspecifically with the cell membranes [71].
2.1.1. Neurotoxins
The primary target of these toxins is the cholinergic system, and they display selectivity towards different receptor subtypes. Based on their selective binding with cholinergic receptors, they can be further classified as:
Curaremimetic Toxins (α-Neurotoxins or Postsynaptic Neurotoxins)
These peptides show very high selectivity, specificity and affinity towards postsynaptic nicotinic acetylcholine receptors (nAChRs). Therefore, they impede the acetylcholine neurotransmission at the skeletal muscle neuromuscular junction. The α-neurotoxins are of two types: short-chain neurotoxins (60–62 amino acids, four disulfide bridges) and long-chain neurotoxins (66–74 amino acids, five disulfide bridges). The long-chain peptide binds with higher affinity with neuronal α7 nAChR. This specificity of the long-chain peptide towards its target is attributed to the presence of the fifth disulfide bond.
The crystal structure of α-Bungarotoxin in complex with nAChR α1 revealed an unprecedented view of the interactions between the receptor and the toxin. It was determined that Arg36 and Phe32 (present in the finger II of the toxin) are the two key residues which block the aromatic cage of the receptor. The toxin also disrupts the interaction between the glycan chain on the Cys-loop and loop C, which breaks the communication between the ligand binding site and the membrane pore [72].
The α-elapitoxin, isolated from Dendroaspis p. polylepis, was found to have an amidated C-terminal arginine. This post-translational modification is rarely observed for snake venom three-finger toxins. Assay binding studies showed that α-elapitoxin strongly inhibits α7 nAChR; however, it was suggested that the C-terminal modification does not significantly affect toxin selectivity [73]. The three-finger toxin peptides have served as an excellent molecular probe in deciphering the structural and functional details of nAChR since the discovery of the iconic peptide α-bungarotoxin from the venom of a krait, Bungarus multicinctus [74,75]. This discovery inspired further studies in the field, and as a result, it was possible to understand various medical problems like myasthenia gravis [76]. Bungarotoxin is still a molecule of choice in delineating underlying mechanisms of many neuronal disorders. Mutation of the invariant Cys-loop Asp residue, in the acetylcholine receptor δ-subunit was identified in a patient suffering from congenital myasthenia syndrome [77]. A recent study employing the use of radioactive derivative of α-bungarotoxin suggested the possible function of neuronal nAChR in the development of Parkinson’s disease [78]. Apart from serving as molecular probes in various medical disorders, three-finger neurotoxins are also gaining attention as potential analgesics, anti-inflammatory molecules, and immunosuppressant [79]. A neurotoxin characterized from Naja n. atra venom in rats suppressed skin allograft rejection by inhibiting the T-cell-mediated immune response [79]. A new group of three-finger toxins (namely, Ω-neurotoxins) were also recently isolated from the venom of the Ophiophagus hannah snake. Interestingly, the characteristic functional residues of α-neurotoxin Oh9-1 are not conserved and interact with nAChR via distinct residues. Furthermore, unlike other α-neurotoxins, the tip of loop II is not involved; rather, the β-strand of loop II interacts with the receptor [80].
Muscarinic Toxins
These toxins bind to muscarinic acetylcholine receptors (mAChRs) and act as either antagonists or agonists towards various subsets of muscarinic AChR (M1–M5). An interesting example in this regard is that of MT1 (a muscarinic toxin from Naja kaouthia), which acts as an agonist at M1 and an antagonist at M4. This selective activity of muscarinic toxins has made them an invaluable tool for distinct biomedical investigations [81]. Other muscarinic toxins (namely MT3 and MT7) isolated from the venom of Dendroaspis angusticeps illustrates high specificity for M4 and M1 mAChRs, respectively [82].
k-Neurotoxins
The structures of these peptides are similar to long chain α-neurotoxins. They exist as dimers in contrast to other members of the 3FTxs family. They recognize only α3β2 and α4β2 nAChR subtypes and not the α1 type.
An interesting peptide, namely haditoxin, was isolated from O. hannah. Its crystal structure revealed it to be a homodimer-like kappa-neurotoxin. However, the monomeric subunits of haditoxin are structurally homologous to curaremimetic short-chain α-neurotoxins. This peptide is reported to interact with neuronal alpha7 nAChRs, as well as with muscle nAChRs [83].
Chemical- and site-directed mutagenesis studies have shown that different types of neurotoxins utilize a common set of amino acid residues, in addition to some other residues, such as that found at the tip of loop I in erabutoxin (Laticauda semifasciata) and the carboxy terminal in α-cobratoxin (Naja kaouthia), to distinctly interact with their target molecules [84].
2.1.2. Cardiotoxins (CTXs)
Cardiotoxins are the second-largest group of three-finger toxins [49]. CTXs are also known as cytotoxins because of their ability to cause lysis in many different cells [85]. They are hydrophobic, basic and, on the whole, relatively short polypeptides (60–62 amino acids). Their structure is similar to short-chain neurotoxins. In comparison to α-neurotoxins, the functional site of cardiotoxins is present at the tip of the three loops. These are generally hydrophobic amino acids, which impart amphiphilic properties to the molecule. A row of positively charged amino acids borders the hydrophobic extremities of loops I–III. This pattern of hydrophobic and cationic regions is commonly found among these cytolytic peptides. The shapes of these CTXs loops are also different to other common neurotoxins. A superposition of CTX and α-neurotoxin shows that the middle loop in the α-neurotoxin is more extended than the other two loops, as shown in Figure 2. The upper parts of both types of toxins have disulfide bridges (termed cysteine-knots) and are homologous and well superimposable [86].
Figure 2.

(A) Snake venom neurotoxin showing the extended central finger (PDB Code; 1NTN). (B) Snake venom cardiotoxin (PDB Code; 1ug4). Both three-finger toxins show a core of disulfide bonds from which the three-finger-like loops extend.
Cardiotoxins are known for their membrane perturbation ability, and the extent to which these peptides can perturb the membrane is known to depend on the presence of Ser30 or Pro28 residues in loop II. Cardiotoxins are therefore classified as S-type and P-type CTXs, respectively. The P-type CTXs bind more strongly to the membranes [87]. CTXs insert into membranes containing anionic lipids and demonstrate certain characteristic features, summarized by Dubovskii et al., which differentiates them from other membrane active molecules [86]. Although until now there has been no clear evidence of whether and how CTXs interact with proteins, some indications have been suggested [88,89]. Hemachatoxin is a P-type CT isolated and crystallized from Hemacatus hemacatus venom. It has been suggested that loop II of this toxin is highly flexible [90].
Cytotoxins are thermostable and also show high stability towards various denaturing agents [88]. Another class of cardiotoxins known as β-cardiotoxins is known to bind to β-adrenergic receptors. Studies showed that these peptides decrease the heart rate, unlike CTXs, which increase the heart rate [91].
Snake venom cardiotoxins are presently also the focus of several cancer-inhibiting studies, and promising results have already been obtained in this regard [86,92,93]. Some studies report cytotoxins to be necrotic, while others as apoptotic agents. Cytotoxin-1 and cytotoxin-II isolated from Naja oxiana venom showed anticancer activity better than cisplatin (anti-cancer drug). It was also demonstrated that these toxins had a minimal effect on normal cells (i.e., MDCK), and induced their apoptotic effect in various cancer cell lines (i.e., HepG2, MCF-7, HL-60 and DU-145) via lysosomal pathway and cytosolic release of cathepsins [32]. Cardiotoxins are also known to possess antimicrobial activity. The tip and β-strand of the first finger of cardiotoxin-1 was utilized as a template to design 20 amino acid residue peptides. Interestingly, these peptides demonstrated a broad range of antimicrobial activity with low cytotoxicity and hemolytic activity towards eukaryotic cells [33].
2.1.3. Acetylcholinesterase Inhibitors
These toxins are structurally similar to short-chain neurotoxins and inhibit the enzyme acetylcholinesterase at the neuromuscular junction, thereby inducing fasciculation of the muscle, and hence are named fasciculins. They block the peripheral site of the enzyme; thus, the enzyme is not able to degrade its substrate (acetylcholine) any more. Earlier studies have shown that amino acid residues present on the first and second loop of the toxin are involved in the inhibition of enzyme activity [69].
2.1.4. Non-Conventional 3FTxs
These polypeptides have an additional fifth disulfide bond either in loop I or loop II. Kini and Doley described these peptides as non-conventional 3FTxs, while earlier these peptides were classified as “weak” neurotoxins [69]. Site-directed mutagenesis and NMR (nuclear magnetic resonance) studies of WTX from Naja Kauthia revealed that loop II of the toxin plays a key role in binding to mAChRs [82]. Candoxin, isolated from B. candidus, shows nanomolar-binding affinity towards both muscle and neuronal AChR. Binding to muscle AChR is easily reversible [94]. Denmotoxin and Irditoxin from colubrid venom show a relatively high binding affinity towards avian neuromuscular junction compared to that of mammals [66,69]. Mambaglins obtained from the venom of eastern green mamba demonstrated analgesic properties. In addition, these peptides also have the ability to inhibit acid-sensing ion channels [95].
2.1.5. Ion Channel Blockers/Modulators
Calciseptine and FS2 isolated from the venom of black mamba are potent L-type calcium channel blockers. They bind to the 1,4-dihydropyridine binding site of the L-type calcium channel and block the passage of calcium through the channel. Structurally, they are similar to short-chain neurotoxins. It has been predicted by some studies that the functional site of these peptides is located on the III loop, between residue 42 and 47 [96,97,98].
Mambalgins (I–III), belong structurally to the non-conventional 3FTxs and specifically inhibit acid-sensing ion channels (ASICs), producing analgesic effects as strong as morphine, but with fewer side effects. Their functional domain is present in loop II, and it has been proposed that mambalgins inhibit ASICs by a pH sensor-trapping mechanism [99].
Recently, a short-chain neurotoxin designated Calliotoxin was isolated from the venom of a coral snake, Calliophis bivirgatus. It bears 53% sequence homology to Rho-elapitoxin-Da1b (UniProt ID: P86419). It was shown to modulate the voltage-gated sodium channel. Further studies of this novel class of 3FTxs could provide insight into the physiology and pharmacology of voltage-gated sodium channels [100]. Another recent study reported a new toxin, namely Tx7335, from Dendroaspis augusticeps, which activates the bacterial potassium channel KcsA. This new identified toxin has high sequence similarity to long chain neurotoxins, but greatly differs with regard to the number and positions of cysteine [101].
2.1.6. Platelet Aggregation Inhibitor
Dendroaspin is known as a potent inhibitor of platelet aggregation. It contains a tripeptide sequence RGD, responsible for the adhesive function of some proteins. It renders its inhibitory activity by preventing the interaction of fibrinogen and its receptor glycoprotein. The functional site of dendroaspin is located at the tip of loop III [102,103].
2.2. Kunitz-Type Serine Protease Inhibitor
Kunitz-type inhibitors are a family of serine protease inhibitors found in the venoms of Elapidae and Viperidae snakes [62]. It has been suggested that they play a role in disturbing the prey’s homeostasis, mainly by interfering with the blood coagulation cascade [26]. Proteome analysis of different snake venoms has shown the relative abundance of snake venom Kunitz-type inhibitors. The Kunitz-type inhibitors represent approximately 28% of the venom proteome of the snake Daboia russelii russelii, found also in Pakistan [104], and approximately 16% of Dendroaspis angusticeps [50]. Furthermore, it is known that more than one type of Kunitz-type inhibitor can be found in the venom of a single snake species [105,106].
Evolutionary studies explain the diversification of snake venom Kunitz-type inhibitors by a positive Darwinian selection, and it was suggested that the driving force for the variability of the inhibiting loop was caused by selective pressure, driven by the presence of diverse prey proteases. With respect to functional activities, snake venom Kunitz-type inhibitors are divided into two major groups: non-neurotoxin (trypsin and chymotrypsin inhibitors) and neurotoxin (potassium and calcium blockers) snake venom Kunitz-type inhibitors [107].
Snake venom Kunitz-type inhibitors bear a structural similarity to aprotinin and consist of approximately 60 amino acids. They have three conserved disulfide bridges responsible for the stability of the molecule and two antiparallel β-strands, which are linked by a β-hairpin in the central part of the molecule. There are two helical regions, one 310-helix near the N-terminus and an α-helix near the C-terminus (Table 2). Kunitz-type inhibitors interact with some serine proteases through an exposed loop in a canonical confirmation. The P1 amino acid residue is considered to be the primary reactive site, determining the specificity and extent of reactivity of the Kunitz-type inhibitor towards the serine protease. X-ray structures of Kunitz-type inhibitors in complex with serine protease revealed both the canonical binding (P3-P3′) and secondary binding loops, which represent important regions of the peptide interacting with the protease [39]. Despite the overall conserved structural scaffold, the amino acid residues in the canonical and secondary binding loop possess subtle sequence variations in these regions and thus responsible for functional diversity in snake venom Kunitz-type inhibitors [108].
The neurotoxic snake venom Kunitz-type inhibitors do not usually have serine protease inhibitory activity. However, they act as potassium and calcium channel inhibitors. A well-known example of a snake venom potassium channel blocker with Kunitz domain is dendrotoxin, isolated from mamba snakes [109]. A recent study demonstrated that dendrotoxin binds and stabilizes eelAChE and even enhances its activity [110], a property of dendrotoxins not described before.
The crystal structure of α-dendrotoxin (α-DTX) and its comparison with the structure of Bovine pancreatic trypsin inhibitor (BPTI) revealed subtle structural differences, which explain the inability of these toxins to form a complex with serine proteases. Differences in the amino acid sequence of the anti-protease loop of this toxin and BPTI, also indicate that required side chain interactions are different. The P1 residue of BPTI is Lys15, which in α-DTX is Tyr17, which cannot provide the same electrostatic interactions as lysine. Another residue adjacent to Tyr17 in α-DTX is aspartate (Asp18), which seems to play an important role in reducing its inhibitory properties. In BPTI and its homologues either alanine or glycine are present at this position. The Asp18 side chain is negatively charged, and it would be placed in an energetically uncomfortable environment, as explained by a model complex of α-DTX with trypsin. Secondly, another important residue in α-DTX is Pro21 (the corresponding residue in BPTI is Ile19), which plays an important role in preventing the binding of α-DTX with trypsin. In contrast, BPTI Ile19 forms a hydrophobic interaction with Tyr39. However, Pro21 of α-DTX would force a different conformation of Tyr39 in trypsin in order to avoid overlap considering Van der Waals distances, thus prohibiting complex formation [111].
Studies have shown that these toxins bind to the potassium channel via 310-helix and β-turn, and a subtle variation in these residues allows the toxins to develop specificity for various subtypes of potassium channels [112,113,114]. Furthermore, a study reported a bifunctional peptide (i.e., BF9) possessing both Kunitz-type protease and potassium channel inhibition properties. It was suggested that the peptide uses basic residues located at its N- and C-terminus to interact with potassium channel subtype Kv1.3, and Asn17 (P1 site) to interact with chymotrypsin and elastase [115]. These potassium channel inhibitors are being employed to understand various physiological disorders, as painful diabetic neuropathy and thereby propose a possible treatment [116]. These Kunitz-type potassium channel inhibitors can also help to understand the structure and function of potassium channels in more detail and provide useful information for drug design investigations, targeting problems associated with the malfunctions of these channels.
Snake venom Kunitz-type serine protease inhibitors also offer promising pharmaceutical applications [117]. For example Textilinin-1 has been shown to be more effective and more specific anti-bleeding agent as compared to Trasylol [aprotinin (Bovine pancreatic trypsin inhibitor)], and demonstrate fewer side effects [118].
Textilinin-1 was isolated from the venom of Pseudonaja textilis [119,120]. It is a slow and tight binding inhibitor of plasmin. The peptide (TP1) isolated from Micrurus tener tener venom is also an inhibitor of plasmin [39,121,122]. TP1 has a molecular mass similar to Kunitz-type peptides, but its sequence is not reported till now. It was suggested that TP1 has potential therapeutic applications as antifibrinolytic agent. The inhibitory activity of TSPI (a recombinant form of a Kunitz-type inhibitor isolated from Oxyuranus scutellats) was investigated comparatively against 12 selected enzymes involved in hemostasis. It showed the highest inhibitory activity towards plasma kallikrein. Rusvikunin (II) and its protein complex, isolated from Daboia r. russelii venom inhibited the activity of trypsin, plasmin and FXa. The peptide and its complex also inhibited the fibrinogen-clotting activity of thrombin. Another Kunitz-type inhibitor (namely PIVL) isolated from Macrovipera lebetina venom, displayed potent anti-tumor activity through interference with integrin receptor function [26,122,123,124,125].
2.3. Disintegrins
Disintegrins are cysteine-rich peptides that result from the post-translational cleavage of snake venom metalloproteases (SVMPs), which are phylogenetically related with ADAMS (a disintegrin and metalloprotease). The possible function and activity of disintegrins in snake venoms is assigned to support the distribution of other toxins throughout prey tissues by binding integrins and inhibiting platelet aggregation direct upon envenomation [126]. Disintegrins are found in the venoms of mainly Crotalidae and Viperidae snakes, and constitute approximately 17% and 18% of total venom proteins, respectively [62].
Calvete and colleagues addressed the evolutionary pathways of disintegrins in detail, and concluded that different disintegrin subfamilies evolved from a common ADAM scaffold, and that structural diversification of disintegrins occurred through disulfide bond engineering and minimization of genomic and protein structures [17,127]. Furthermore, Kini has recently described that exonization and intronization plays an important role in the evolution of disintegrin/metalloprotease genes [128].
Studies have shown that disintegrins are elongated peptides, consisting of several loops and turns, stabilized by disulfide bonds, as exemplified by Trimestatin shown in Figure 3A and Table 2. Disintegrins can exist as monomers, homodimers and/or heterodimers [129]. The monomers include short (49–51 amino acid residues; four disulfide bonds), medium (70 amino acids; six disulfide bonds) and long disintegrins (84 amino acids; seven disulfide bonds). Most of the disintegrins belong to the monomeric type.
Figure 3.

(A) Ribbon diagram of the crystal structure of Trimestatin (a snake venom disintegrin). The figure illustrates cysteine residues forming disulfide bonds in yellow, while the tripeptide sequence (RGD) binding motif is shown in pink. (B) The tripeptide sequence motifs of various snake venom disintegrins having different intermolecular interactions are shown with reference to their specificity.
Disintegrins bind to integrins, which play a fundamental role in many pathological and physiological processes. Disintegrin binding specificity with integrin is determined by the presence of a tri-peptide motif located in the hairpin loop. Therefore, three functional classes containing RGD, MLD or R/KTS motifs have been recognized. The interaction of disintegrins containing either of these motifs, with specific types of integrins is mapped in Figure 2B. Most disintegrins studied to date belong to the RGD-containing class of motifs and are monomers. Some dimeric disintegrins having a RGD subunit are also known. The MLD motif is present only in heterodimeric disintegrins. KTS/RTS-containing disintegrins selectively bind to the collagen receptor α1β1 integrin [23]. It has been suggested by different studies that the conserved aspartate residue in the tripeptide sequence of many disintegrins binds to a specific β subunit, whereas the other two amino acids determine binding affinity to the specific α integrin subunit [130].
In vivo studies showed that disintegrins containing MLD and KTF motifs are not toxic in avian and rodent models at various therapeutic doses. Further studies have shown that amino acid residues other than the tripeptide domain interact with integrin receptors. For example, any change at the C-terminus region of echistatin and eristostatin can modify their receptor binding ability. Replacement of Met28 with Leu selectively reduced echistatin’s ability to recognize α5β1 only. While replacement of Met28 with Asn completely abolished its ability to recognize integrin [131]. Recently, Urra and Araya-Maturana reviewed the anti-metastatic mechanism of snake venom toxins in detail and summarized that snake venom toxins (particularly snake venom cystatins, disintegrins, lectins, and cardiotoxins) inhibit pro-migratory and pro-invasive signals stimulated by extracellular matrix proteins and growth factors [29].
The modulatory properties of disintegrins were observed in investigations targeting various disorders, such as asthma, insulin-dependent diabetes mellitus, neurodegenerative illness, cell apoptosis, as well as, in this context, cancer angiogenesis and metastasis [132]. Saxatilin isolated and cloned from the venom of Gloydius saxatilis, showed thrombolytic effects in mice model. It was shown that saxatilin has no fibrinolytic activity; however, it can inhibit multiple integrins by acting on platelets [133]. A growing interest in the development of recombinant and/or chimeric disintegrins can be recognize. For example, recombinant r-mojastin and r-viridistatin (RGD-disintegrins) showed inhibition of pancreatic tumor cells. These disintegrins possess strong angiogenesis properties, and bind to ανβ3 and ανβ5 receptors. Potential applications of DisBa01 (RGD-disintegrin) have been identified, for example, also in the treatment of cancer, hernia and fibroproliferative diseases. Vicrostatin is a chimeric disintegrin which can significantly inhibit breast cancer cells and tubulogenesis [134]. Finally, disintegrins have already served as novel lead compounds supporting the design of various approved drugs, for example Aggarast and Integrilin [7,135,136].
2.4. Natriuretic Peptides (NPs)
Natriuretic peptides found in vertebrates play a vital role in natriuresis. Homologous peptides have also been reported in plants and bacteria. Three mammalian NPs are known: atrial natriuretic peptide (ANP), B-type natriuretic peptide (BNP), and C-type natriuretic peptide (CNP). These NPs regulate the functions of cardiovascular and renal systems, and render their function by forming a complex and binding to the natriuretic peptide receptors (NPR-A, NPR-B) [137]. For example, ANP and BNP act in an endocrine manner to maintain blood pressure and volume. Both ANP and BNP are released by cardiomyocytes in response to elevated blood pressure and hypervolemia [138,139], and CNP is produced by endothelial cells [140]. NPs are of great interest today, as they support the development of therapeutics and medical procedures for the treatment and diagnosis of unfortunate physiological conditions, such as heart failure and hypertension [7]. Further details about the structures, receptors and physiological functions can be found in recent reviews published by Volpe M and Potter L. et al. [141,142].
Snake venom NPs are similar to mammalian natriuretic peptides in terms of structure and function, but also possess certain distinct characteristics. All NPs have a conserved 17-residue disulfide ring and variable N- and C-termini. Five amino acid residues (F, D, R, I and L/I) within the ring structure of the NPs are crucial for receptor binding. Different NPs recognize different specific receptors by a slight sequence variation in the N- and C-terminal region [143]. Sequence alignment of different snake venom and human natriuretic peptides is shown in Figure 4. The structure of NPs is rather flexible. However, a natriuretic peptide isolated from Krait venom (known as KNP), was reported to have a putative α-helix, formed by its extended C-terminus [144]. Another study reported an O-glycosylated natriuretic peptide (TcNPa), from the venom of Tropichedis carinatus, and NMR analysis of TcNPa confirmed the presence of an α-helix as well [145].
Figure 4.

Sequence comparison of human natriuretic peptides with those of different snake venoms, aligned by Clustal W. Conserved cysteine residues are highlighted in yellow, while relatively conserved amino acid residues known for NPR binding [143] are shaded blue.
Snake venom NPs have been reported to produce a strong hypotensive effect upon envenomation, and the prey rapidly loses consciousness [146,147]. Proteomics analysis has shown that snake venom NPs are more extensively found in Viperidae than the Elapidae. NPs constitute only 3% of the venom proteome of Dendroaspis polyepis and 37% of that of Bothriechis nigroviridis [62].
The Elapidae snake venom contains ANP/BNP natriuretic peptides and CNP natriuretic peptides are found in Viperidae. Evolutionary studies suggested that CNP natriuretic peptides were recruited independently after the divergence of vipers from more advanced snakes [148]. However, later studies reported the presence of ANP-like NPs in the venom of C. durissus casacavella and C. oreganus abyssus.
Dendroaspis natriuretic peptide (DNP) was the first natriuretic peptide isolated from the venom of Dendroaspis angusticeps. DNP was found to have greater stability to neutral endopeptidase than three mammalian NPs. The relatively high stability of DNP was attributed to the presence of elongated N- and C-termini [149]. The discovery of DNP led to the isolation and characterization of many novel snake venom NPs, which possess greater stability and novel mechanism of actions, as compared to their mammalian counterparts. A novel 45 amino acid mature natriuretic peptide (Na-Np) was isolated from Chinese cobra venom. This NP showed similar but relatively weak activity compared to human ANP. Also, Na-Np did not inhibit human platelet aggregation stimulated by thrombin or stejnulxin [150]. Bochra Tourki et al. studied the cardioprotective effect of Lebetin 2 (L2, isolated from the venom of Macrovipera lebetina) against regional and/or global ischemia-reperfusion (IR) in isolated rat hearts. Lebetin 2 is a structural homologue of BNP. In this study, it was also established that L2 utilizes identical mechanism for cardioprotection to that of BNP. The mechanism involves the activation of NP receptors and mitoKATP channels. Subsequently, mPTP (mitochondrial permeability transition pore) opening at the time of reperfusion is delayed. Consequently, IR-induced damage is attenuated. However, lebetin 2 was found to be more potent than BNP in enhancing the coronary flow, improving contractile dysfunction of myocardium and surviving global IR. It was also suggested that L2 could be a strong drug candidate for acute myocardial ischemia [151].
Recently, the development of chimeric natriuretic peptides has been the subject of interest, to combine the benefits of several peptides and avoid unwanted side effects. An interesting example in this context is Cenderitide (CD-NP), which was designed by fusing 22 amino acids of CNP with 15 amino acid residues of the carboxyl termini of DNP. Thus, CD-NP was designed to co-activate the two particulate guanylyl cyclase (pGC) receptors namely pGC-A and pGC-B. In vivo studies revealed that CD-NP, like ANP, BNP, and DNP (but unlike CNP) possesses renal-enhancing actions through pGC-A/cGMP activation. CD-NP was under clinical trials to treat cardiorenal syndrome [41]. However, Capricor Beverly Hills, California, recently completed an additional study with patients having stable heart failure in combination with moderate renal impairment. These clinical investigations performed by Capricor were terminated only recently [6].
A structure-function analysis was carried out on a NP (namely KNP) isolated from Krait venom. It was shown that KNP has two pharmacophores, a K-ring and the helix extension at the C-terminus. Both segments induce vasodilation through orthogonal pathways. Furthermore, this study also indicated the amino acids that act as molecular switches, controlling the activity of NP on smooth muscle and kidney. Thus, it can be concluded that engineered NP analogues may serve as key therapeutic leads that could help to regulate either blood pressure or volume in distinct cohorts of heart failure (HF) patients [143].
2.5. Bradykinin Potentiating Peptides (BPPs)
BPPs are small proline-rich hypotensive peptides consisting of 5–14 amino acid residues found in snake venom glands. Studies have also reported their presence in the brain, as part of C-type natriuretic peptide. These small peptides are well-known natural inhibitors of the angiotensin-converting enzyme (ACE). ACE is a zinc-containing metalloenzyme and an important component of the renin angiotensin system (RAS), a key regulator of blood pressure. ACE raises the blood pressure by converting angiotensin I to II (a potent vasoconstrictor) and by catalyzing the degradation of bradykinin (a natural vasodilator) [105,106].
BPPs were described first in the venom of B. jararaca as inhibitors of ACE [152]. Further investigations resulted in the approval and marketing of Captopril, which is an orally active peptidomimetic of the BPP [153]. This is the best and most well-known example of snake venom-based drug discovery investigation. Later similar compounds have been developed with fewer side effects, like enalapril, lisinopril, perindopril, etc., popularly known as ACE inhibitors [154]. Recently, we conducted a study on synthetic analogues of four BPPs isolated from Agkistrodon bilineatus and suggested that a net negative charge on the peptide is an important aspect to be considered for drug design towards ACE inhibition [155]. Nonetheless, further studies are required in this context.
Snake venom BPPs and orally active ACE inhibitors lower the blood pressure by blocking the ACE activity; as a result, the level of bradykinin in the blood increases. A large number of BPPs have been isolated from various snake venoms to date, all resembling one another in terms of structural motif, and having a proline-rich C-terminus, as shown in Figure 5. In the mature BPPs, pyroglutamate modification is usually present at the amino terminus. The modification of the N-terminus and the presence of a high percentage of proline residues are important factors in providing stability to these peptides. Structural studies of ACE in complex with a BPPb revealed that the C-terminal proline binds with the primary binding pocket of the enzyme through the formation of hydrogen bonds (Figure 5). Thus, nature has endowed these peptides with a structural motif that is highly resistant to hydrolytic degradation and that is able to penetrate the narrow channel-like active site of ACE [155].
Figure 5.

(A) Crystal structure of the angiotensin-converting enzyme complexed with BPP (PDB Code: 4APJ). The figure shows BPP in the channel-like cavity (i.e., active site) of the enzyme. (B) Sequence alignment showing the comparison of various snake venom BPPs. All peptides have a common IPP motif at the C-terminus (marked by asterisk). Left, uniprot IDs of the peptides are also provided.
Although all BPPs revealed high sequence similarities, their mechanism of action is not restricted to ACE inhibition only. Studies have shown that one BPP (namely, Bj-PRO-10c) is a potent inhibitor of ACE in vitro, but evokes a strong antihypertensive effect in an ACE-independent manner. This peptide was found to activate kidney arginosuccinate synthetase (ASS). Upon activation of ASS, the level of l-arginine and nitric oxide increases, and blood pressure decreases. Bj-PRO-10c was also reported to induce the release of gamma-aminobutyric acid (GABA) and glutamate by mobilizing calcium fluxes in neurons, which are also involved in controlling the blood pressure. Two other BPPs (namely, Bj-PRO-5a and Bj-PRO-10c) were demonstrated to be the modulators of bradykinin-B2 and M1 muscarinic acetylcholine receptors. These peptides induced vasodilatation in vivo. Therefore, these peptides can serve as pharmacological tools to determine new pathways and to better understand the mechanisms of blood pressure regulation. They can also help to identify novel therapeutic targets for the treatment of hypertension and related diseases [156,157].
2.6. Sarafotoxins (SRTXs)
Snake venom SRTXs are structurally and functionally related to vertebrate endothelin (ETs). Both families of peptides are potent vasoconstrictors and interact with endothelin receptors (ETA and ETB), which can modulate the contraction of cardiac and smooth muscles in different tissues. SRTXs are highly toxic venomous peptides, whereas ETs are hormones produced by the mammalian vascular system [46,158]. SRTXs are exclusively and highly expressed in various isoforms in the venom gland of Atractaspis genus [159]. These peptides range in length from 15 to 30 amino acids as established by mass spectrometric experiments [160]. The most abundant isoform has 25 amino acids, while all other isoforms of these peptides contain a common core of 21 amino acids and adopt a highly conserved three-dimensional scaffold stabilized by an α-helical structure (Table 2). The amino acid Trp21 is invariably present and plays a crucial role in endothelin receptor binding [161]. It was also described in a previous study that the amino acids at positions 4–7 present a variable region of these peptides [162]. On the other hand, C-terminal extension determines the affinity and selectivity towards endothelin receptors. It was demonstrated experimentally by an in vitro study applying cloned human ET receptors that the C-terminus extension reduces the binding affinity of SRTXs to both ETA and ETB receptors. Furthermore, it was also suggested that steric hindrance associated with this extension was responsible for a reduced interaction between SRTX and ET receptor [161]. The respiratory effects of sarafotoxin-b (short isomer) and sarafotoxin-m (long isoform) were studied in vivo, revealing that both peptides acted differently on the respiratory mechanics. This difference of action of both peptides was also attributed to a C-terminus extension for the long isoforms. C-terminal extension can result in different spatial conformation of the two peptides [163]. SRTXs can serve to better understand the endothelin system and related diseases, such as pulmonary hypertension, asthma, and/or heart failure [163,164,165]. SRTX was used to uncover the communication pathways between the peptide and ET receptor. It was suggested that phosphoinositide second messenger system and Ca2+ signal-transduction mechanisms are involved in this communication [46,166,167].
2.7. Tripeptides
These are also very small peptides in snake venoms, consisting of only three amino acids, with pyroglutamate or Glutamate at the N-terminus and tryptophan at the C-termini [105]. They are also known to be snake venom metalloprotease inhibitors, and to date have been reported only for Viperidae snake venoms. These inhibitory peptides protect the venom gland from autodigestion by its own proteinases and also prevent the hydrolysis of other venom proteins during storage in the venom gland [168,169,170,171]. A recent study reported a synthetic tripeptide (Glu-Val-Trp) as a promising lead compound for drug design investigation with respect to restoring axonal connectivity in neurodegenerative processes [172]. Two tripeptides, namely Pt-A (Glu-Asn-Trp) and Pt-B (Glu-Gln-Trp), were purified from the venom of Deinakistrodon acutus. These peptides showed antiplatelet aggregation and antithrombotic effects. It was further demonstrated that both peptides protected mice from ADP-induced paralysis, but only Pt-A exerted antiplatelet aggregation activity in vivo, and not Pt-B. Therefore, it was proposed that Pt-B approaches a different mechanistic pathway to carry out its antithrombosis function from Pt-A [173].
2.8. Crotamine
Crotamine (myoneurotoxin) is a major component of the venom of the South American snake Crotalus durissus terrificus [174]. This polypeptide is composed of 42 amino acids, folded into a three-dimensional structure stabilized by three disulfide bonds, having two α-helices and two β-sheets (Table 2). The crystal structure of crotamine shows that almost all hydrophobic and charged residues are exposed to the solvent, providing a unique interaction pattern. This property of crotamine, along with a very stable structural scaffold, is considered to be responsible for its complex interaction with target proteins [175]. As a result, crotamine has demonstrated very versatile pharmacological activities. It is highly positively charged, and thus has the ability to penetrate cells, bind DNA, and can even cross the blood–brain barrier (BBB). It has also shown a particular anticancer activity against melanoma cells [176]. Crotamine is strongly attracted to cancer cells, because the surfaces of cancer cells have more negatively charged molecules compared to normal cells [177]. Studies have shown that crotamine displays analgesic properties 30 times more potent than morphine [178], and additionally possesses antimicrobial activities. Further, it can interact with different molecular targets, involved in platelet aggregation and mitochondria [179]. Crotamine also has the ability to transport drugs in mammalian cells without requiring any specific receptors. A recent study has also suggested the use of a synthetic analogue of crotamine to probe tumor cells, thus acting as a selective tool for the delivery of anticancer compounds [180]. A review published by Kerkis et al. provides details of the characteristics, properties, mechanisms of cell penetration and other biotechnological applications of crotamine [174].
2.9. Waprin Family
Nawaprin is the first member of the ‘snake waprin family’, purified from the venom of Naja nigricolis. Nawaprin and omwaprin-a (from the venom of Oxyuranus microlepidotus) have similar functions and a distinct structural fold, resembling a flat disc-like structure with a single WAP (whey acidic protein) domain containing four disulfide bonds, short antiparallel β-sheets, a 310 helix and a spiral back bone [181,182]. Figure 6A shows the structure of a synthesized omwaprin. The high sequence homology between the waprins known so far indicates that their structure is highly conserved (Figure 6B). Nawaprin bears structural similarity to elafin (a human leukocyte elastase-specific inhibitor) [182]. It has also been shown that omwaprin displays dose-dependent and strain-specific antibacterial activity and is devoid of any hemolytic activity and toxicity to mice. In the same study, the authors also concluded that the unique three-dimensional scaffold is important for the function of the peptide. Scanning electron microscopy revealed that omwaprin elicits its function through membrane disruption [183,184]. Furthermore, St. Pierre et al. proposed that Kunitz-type peptides and Waprin evolved from a common ancestral gene [185], and the transcripts of venom toxins with fused Kunitz-Wap domains were identified in the venom of Sistrurus catenatus edwardsii and Suta fasciata [186,187].
Figure 6.

(A) Crystal structure of the synthesized Omwaprin peptide (PDB Code: 3NGG). (B) Multiple sequence alignment of the waprin-like peptides shows highly homologous primary structures. Cysteine residues are highlighted in yellow. Uniprot ID of the sequences are also provided.
2.10. Waglerins
Waglerins are lethal toxins found in the venom of Tropidolaemus wagleri. These peptides consist of 22–24 amino acids, and four distinct isoforms have been reported to date [47,188]. These isoforms bear high sequence homology with each other and differ only by a few amino acid residues (Figure 7). A recent proteomic study revealed that Waglerin is the major toxin in the venom of Tropidolaemus wagleri [189], representing almost 40% of the total venom. In contrast, in a separate study, it was reported that waglerin constitutes 15% of the total composition of Tropidolaemus wagleri venom [190].
Figure 7.

Sequence comparison of the four distinct isoforms of waglerin.
According to an evolutionary study conducted by Debono et al., these neurotoxic peptides are most probably the result of a de novo evolution within the pre-pro region of the C-type natriuretic peptide gene in Tropidolaemus wagleri venoms, a site distinct from the domain encoding for the NP [191]. These peptides mediate their neurotoxic activity by binding to the muscle nAChR. Human, mouse and rat nAChR have different specificities for Waglerin-1 (Wtx-1). It imparts the highest toxicity towards mouse and causes death by respiratory failure [192]. An anti-wrinkle cream, SYN-AKETM, has been developed by Pentapharm Ltd. (Basel, Switzerland). It is a peptide mimic (H-b-Ala-Pro-DabNHB) of Wtx-1 [6]. According to Pentapharm, the Syn-Ake invokes its action by blocking the muscle nAChR in a reversible manner by blocking the ion channel. As a result, the Na+ uptake is substantially disturbed, and the muscles remain relaxed (http://articles.latimes.com/2009/may/03/image/ig-beauty3).
2.11. Antimicrobial Peptides
Cathelicidins are small cationic antimicrobial peptides (cAMP) which were reported for elapid snake venoms [193]. Some of the cathelicidins (e.g., C-BF) were shown to be able to suppress inflammation [194]. Vipercidin was reported in the venom of four South American pit vipers, and due to its excellent bactericidal profile, it could serve as promising lead molecule for the development of future peptidyl antibiotics [195].
3. Research Methods
Several advanced and complementary bioanalytical methods and techniques are applied to isolate, characterize and evaluate the pharmacological properties of snake venom proteins and peptides in terms of drug discovery investigations. A thorough discussion of all the methodologies is beyond the scope of this review. In brief, for investigations, snake venom is milked first, filtered through 0.45 µm filters to remove any cellular debris, lyophilized and stored at −20 °C until further use [196]. Bioassay-guided purification of the peptide is one of the most utilized methods for screening the venom in high-throughput assays against targets of therapeutic interest. This requires a combination of chromatographic methods for purifying the peptide of interest. In terms of our investigations analyzing snake venom peptides, we optimized this procedure first, as well as the further fractionation of crude venom, by applying size exclusion chromatography (SEC FPLC). We performed our purification while keeping sample solutions at pH 5.0 (0.1 M volatile buffer), which is a good choice, as most snake venoms are naturally present at pH 5.0. The bioactive fractions can then be further purified by other chromatographic modes, such as ion exchange, reversed phase and/or affinity chromatography. Recently, a new online microfluidic high-resolution screening method was employed to identify neurotoxic components from 47 snake venoms. This method provides a new analytical platform to rapidly analyze neurotoxic components of snake venom in vitro [57]. Complementary mass spectrometric methods are applied to determine molecular mass and sequence of the bioactive peptides. Since the advent of proteomics, the opportunities to discover novel bioactive peptides have accelerated substantially. During the last few years, there has been enormous development in both multidimensional chromatographic and mass spectrometric techniques. State-of-the-art mass spectrometers with high resolution and sensitivity are at the disposal of scientists, also enabling novel fragmentation techniques. Modern mass spectrometers (MS) have dual-mode ionization sources and hybrid mass analyzers to maximize performance. These days, a holistic approach, known as venomics, is employed to discover new proteins and peptides. It is an integrated approach exploiting genomics, transcriptomics and proteomics to increase the availability of information, applying only a limited amount of sample. The venom gland preparations are investigated with next-generation sequencers. These technologies have revolutionized DNA sequencing, due to the parallelization and miniaturization of the reactions. The peptides are analyzed with ESI or MALDI TOF mass spectrometers. MALDI is a very versatile source of ionization and is used in MS imaging of even the venom gland or tissue. There are two different types of MS-based sequencing approaches in use today: true de novo sequencing and data search-based sequencing. Search-based sequencing is more common, using bioinformatics tools, like Mascot (Matrix Science) or Protein Pilot (ABSciex). Different MS methods are utilized to perform MS/MS or MSn or to cleave the peptide backbone, including post-source decay (PSD), collision-induced dissociation (CID) and higher collision energy dissociation (HCD). The peptides with labile posttranslational modification are analyzed with techniques based on radical fragmentation, such as electron capture dissociation (ECD) and electron transfer dissociation (ETD) [197].
Crystallographic studies of venom proteins and peptides provide insights into the three-dimensional structure and function of the individual components and its complex formation with other receptors or proteins. These studies are of utmost importance in structure-based drug discovery investigations. Many important details have already been presented to date. For example, crystallographic studies, in particular, have revealed that snake venom BPPs bind to ACE in a zinc-independent manner, while the currently available synthetic ACE inhibitor binds at the active site via a zinc metal ion [155]. Our labs at DESY, Hamburg, have the most advanced protein crystallization setups and X-ray crystallography infrastructure. We routinely perform crystallization experiments of peptides and proteins to apply crystallography in combination with highly intensive synchrotron radiation. In this context, we also developed new methods to improve protein crystallization [198,199,200]. Although realizing the highly flexible nature of some venom peptides and difficulties in crystallization, multidirectional NMR spectroscopy can be applied [201].
Apart from structural studies, different groups are working to generate large libraries of recombinant peptides by high-throughput expression of venom peptides in E. coli for drug discovery investigations [202,203]. Different metabolomics approaches are also used to analyze low-molecular-weight components of animal venoms [204]. In addition to these experimental approaches, bioinformatics and molecular modeling studies provide guidance for rational drug design, in understanding the complex formation of venom peptides and their receptors, and in delineating novel functions of venom peptides [205,206].
4. Conclusions
Snake venoms have, until now, been considered a mostly untapped resource of unique peptides and proteins. Snake venom peptides have been identified to be mostly very stable, as they have to reach the site of action before the internal mechanisms of their prey degrades, neutralizes or excretes them. Nature has met this requirement through the recruitment of highly stable molecular scaffolds, which are mostly resistant to protease degradation and can bypass other immune mechanisms of prey. Several investigations have shown that these venom peptides influence important physiological systems, such as blood pressure regulation, homeostasis, the nervous system, etc. The latest analytical technologies have made it possible to investigate several unknown snake venom peptides. These peptides can serve as molecular probes. Also, they can be used directly or as lead compounds for drug discovery and drug design investigations, because they are easy to synthesize and less prone to inducing an immune response.
Acknowledgments
SA Ali would like to thank the financial support from Higher Education Commission (HEC) of Pakistan (HEC-No:20-3891/NRPU/R&D/HEC/14/1098).
Author Contributions
Concept; C.B. and A.M., write up; C.B., A.M, A.A. and S.A.A, Editing; C.B., A.M and S.A.A., supervision; C.B.
Funding
No funding was received to draft this review.
Conflicts of Interest
The authors declare no conflict of interest.
Key Contribution
The article highlights the types and classification of snake venom peptides and their pharmacological importance and applications in biodiscovery. Moreover, an overview of structure, function and evolution of these peptides is summarized and research methodologies involved are discussed briefly.
References
- 1.Utkin Y.N. Animal venom studies: Current benefits and future developments. World J. Biol. Chem. 2015;6:28–33. doi: 10.4331/wjbc.v6.i2.28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Undheim E.A., Georgieva D.N., Thoen H.H., Norman J.A., Mork J., Betzel C., Fry B.G. Venom on ice: First insights into antarctic octopus venoms. Toxicon. 2010;56:897–913. doi: 10.1016/j.toxicon.2010.06.013. [DOI] [PubMed] [Google Scholar]
- 3.King G.F. Venoms as a platform for human drugs: Translating toxins into therapeutics. Expert Opin. Biol. Ther. 2011;11:1469–1484. doi: 10.1517/14712598.2011.621940. [DOI] [PubMed] [Google Scholar]
- 4.Ruder T., Ali S.A., Ormerod K., Brust A., Roymanchadi M.L., Ventura S., Undheim E.A., Jackson T.N., Mercier A.J., King G.F., et al. Functional characterization on invertebrate and vertebrate tissues of tachykinin peptides from octopus venoms. Peptides. 2013;47:71–76. doi: 10.1016/j.peptides.2013.07.002. [DOI] [PubMed] [Google Scholar]
- 5.Dutertre S., Jin A.H., Vetter I., Hamilton B., Sunagar K., Lavergne V., Dutertre V., Fry B.G., Antunes A., Venter D.J., et al. Evolution of separate predation- and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014;5:3521. doi: 10.1038/ncomms4521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Pennington M.W., Czerwinski A., Norton R.S. Peptide therapeutics from venom: Current status and potential. Bioorg. Med. Chem. 2018;26:2738–2758. doi: 10.1016/j.bmc.2017.09.029. [DOI] [PubMed] [Google Scholar]
- 7.Koh C.Y., Kini R.M. From snake venom toxins to therapeutics—Cardiovascular examples. Toxicon. 2012;59:497–506. doi: 10.1016/j.toxicon.2011.03.017. [DOI] [PubMed] [Google Scholar]
- 8.Waheed H., Moin S.F., Choudhary M.I. Snake venom: From deadly toxins to life-saving therapeutics. Curr. Med. Chem. 2017;24:1874–1891. doi: 10.2174/0929867324666170605091546. [DOI] [PubMed] [Google Scholar]
- 9.Mirshafiey A. Venom therapy in multiple sclerosis. Neuropharmacology. 2007;53:353–361. doi: 10.1016/j.neuropharm.2007.05.002. [DOI] [PubMed] [Google Scholar]
- 10.Otvos R.A., Heus F., Vonk F.J., Halff J., Bruyneel B., Paliukhovich I., Smit A.B., Niessen W.M., Kool J. Analytical workflow for rapid screening and purification of bioactives from venom proteomes. Toxicon. 2013;76:270–281. doi: 10.1016/j.toxicon.2013.10.013. [DOI] [PubMed] [Google Scholar]
- 11.Simoes-Silva R., Alfonso J., Gomez A., Holanda R.J., Sobrinho J.C., Zaqueo K.D., Moreira-Dill L.S., Kayano A.M., Grabner F.P., da Silva S.L., et al. Snake venom, a natural library of new potential therapeutic molecules: Challenges and current perspectives. Curr. Pharm. Biotechnol. 2018;19:308–335. doi: 10.2174/1389201019666180620111025. [DOI] [PubMed] [Google Scholar]
- 12.Del Brutto O.H., Del Brutto V.J. Neurological complications of venomous snake bites: A review. Acta Neurol. Scand. 2012;125:363–372. doi: 10.1111/j.1600-0404.2011.01593.x. [DOI] [PubMed] [Google Scholar]
- 13.Liu S., Yang F., Zhang Q., Sun M.Z., Gao Y., Shao S. “Anatomical” view of the protein composition and protein characteristics for gloydius shedaoensis snake venom via proteomics approach. Anat. Rec. 2011;294:273–282. doi: 10.1002/ar.21322. [DOI] [PubMed] [Google Scholar]
- 14.Meier J., Stocker K.F. Biology and distributuion of venomous snakes of medical importance and the composition of snake venom. In: Meier J., White J., editors. Handbook of Clinical Toxicology of Animal Venoms and Proteins. CRC Press; Boca Raton, FL, USA: 1995. pp. 367–412. [Google Scholar]
- 15.Georgieva D., Arni R.K., Betzel C. Proteome analysis of snake venom toxins: Pharmacological insights. Expert Rev. Proteom. 2008;5:787–797. doi: 10.1586/14789450.5.6.787. [DOI] [PubMed] [Google Scholar]
- 16.Munawar A. Analysis of the Low Molecular Weight Peptides of Selected Snake Venoms. University of Hamburg; Hamburg, Germany: 2012. [Google Scholar]
- 17.Calvete J.J., Moreno-Murciano M.P., Theakston R.D., Kisiel D.G., Marcinkiewicz C. Snake venom disintegrins: Novel dimeric disintegrins and structural diversification by disulphide bond engineering. Biochem. J. 2003;372:725–734. doi: 10.1042/bj20021739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lucena S., Rodriguez-Acosta A., Grilli E., Alfonso A., Goins A., Ogbata I., Walls R., Suntravat M., Uzcategui N.L., Guerrero B., et al. The characterization of trans-pecos copperhead (agkistrodon contortrix pictigaster) venom and isolation of two new dimeric disintegrins. Biologicals. 2016;44:191–197. doi: 10.1016/j.biologicals.2016.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Calvete J.J., Marcinkiewicz C., Sanz L. Snake venomics of bitis gabonica gabonica. Protein family composition, subunit organization of venom toxins, and characterization of dimeric disintegrins bitisgabonin-1 and bitisgabonin-2. J. Proteome Res. 2007;6:326–336. doi: 10.1021/pr060494k. [DOI] [PubMed] [Google Scholar]
- 20.Kini R.M. Evolution of three-finger toxins—A versatile mini protein scaffold. Acta Chim. Slov. 2011;58:693–701. [PubMed] [Google Scholar]
- 21.Osipov A.V., Rucktooa P., Kasheverov I.E., Filkin S.Y., Starkov V.G., Andreeva T.V., Sixma T.K., Bertrand D., Utkin Y.N., Tsetlin V.I. Dimeric alpha-cobratoxin X-ray structure: Localization of intermolecular disulfides and possible mode of binding to nicotinic acetylcholine receptors. J. Biol. Chem. 2012;287:6725–6734. doi: 10.1074/jbc.M111.322313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Koh D.C., Armugam A., Jeyaseelan K. Snake venom components and their applications in biomedicine. Cell. Mol. Life Sci. 2006;63:3030–3041. doi: 10.1007/s00018-006-6315-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Arruda Macedo J.K., Fox J.W., de Souza Castro M. Disintegrins from snake venoms and their applications in cancer research and therapy. Curr. Prot. Pept. Sci. 2015;16:532–548. doi: 10.2174/1389203716666150515125002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kong Y., Wang Y., Yang W., Xie Z., Li Z. Lx0702, a novel snake venom peptide derivative, inhibits thrombus formation via affecting the binding of fibrinogen with gpiib/iiia. J. Pharmacol. Sci. 2015;127:462–466. doi: 10.1016/j.jphs.2015.03.010. [DOI] [PubMed] [Google Scholar]
- 25.Uzair B., Atlas N., Malik S.B., Jamil N., Salaam T.O., Rehman M.U., Khan B.A. Snake venom as an effective tool against colorectal cancer. Protein Pept. Lett. 2018;25:626–632. doi: 10.2174/0929866525666180614112935. [DOI] [PubMed] [Google Scholar]
- 26.Mukherjee A.K., Mackessy S.P. Pharmacological properties and pathophysiological significance of a kunitz-type protease inhibitor (rusvikunin-ii) and its protein complex (rusvikunin complex) purified from daboia russelii russelii venom. Toxicon. 2014;89:55–66. doi: 10.1016/j.toxicon.2014.06.016. [DOI] [PubMed] [Google Scholar]
- 27.Dutertre S., Nicke A., Tsetlin V.I. Nicotinic acetylcholine receptor inhibitors derived from snake and snail venoms. Neuropharmacology. 2017;127:196–223. doi: 10.1016/j.neuropharm.2017.06.011. [DOI] [PubMed] [Google Scholar]
- 28.Lewis R.J., Garcia M.L. Therapeutic potential of venom peptides. Nat. Rev. Drug Discov. 2003;2:790–802. doi: 10.1038/nrd1197. [DOI] [PubMed] [Google Scholar]
- 29.Urra F.A., Araya-Maturana R. Targeting metastasis with snake toxins: Molecular mechanisms. Toxins. 2017;9:390. doi: 10.3390/toxins9120390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Sanhajariya S., Duffull S.B., Isbister G.K. Pharmacokinetics of snake venom. Toxins. 2018;10:73. doi: 10.3390/toxins10020073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Utsintong M., Talley T.T., Taylor P.W., Olson A.J., Vajragupta O. Virtual screening against alpha-cobratoxin. J. Biomol. Screen. 2009;14:1109–1118. doi: 10.1177/1087057109344617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ebrahim K., Shirazi F.H., Mirakabadi A.Z., Vatanpour H. Cobra venom cytotoxins; apoptotic or necrotic agents? Toxicon. 2015;108:134–140. doi: 10.1016/j.toxicon.2015.09.017. [DOI] [PubMed] [Google Scholar]
- 33.Sala A., Cabassi C.S., Santospirito D., Polverini E., Flisi S., Cavirani S., Taddei S. Novel naja atra cardiotoxin 1 (ctx-1) derived antimicrobial peptides with broad spectrum activity. PLoS ONE. 2018;13:e0190778. doi: 10.1371/journal.pone.0190778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Gan Z.R., Gould R.J., Jacobs J.W., Friedman P.A., Polokoff M.A. Echistatin. A potent platelet aggregation inhibitor from the venom of the viper, echis carinatus. J. Biol. Chem. 1988;263:19827–19832. [PubMed] [Google Scholar]
- 35.Scarborough R.M., Rose J.W., Hsu M.A., Phillips D.R., Fried V.A., Campbell A.M., Nannizzi L., Charo I.F. Barbourin. A gpiib-iiia-specific integrin antagonist from the venom of sistrurus m. Barbouri. J. Biol. Chem. 1991;266:9359–9362. [PubMed] [Google Scholar]
- 36.Moiseeva N., Swenson S.D., Markland F.S., Jr., Bau R. Purification, crystallization and preliminary X-ray analysis of the disintegrin contortrostatin from agkistrodon contortrix contortrix snake venom. Acta Crystallogr. D Biol. Crystallogr. 2002;58:2122–2124. doi: 10.1107/S0907444902011204. [DOI] [PubMed] [Google Scholar]
- 37.Juan-Rivera M.C., Martinez-Ferrer M. Integrin inhibitors in prostate cancer. Cancers. 2018;10:44. doi: 10.3390/cancers10020044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zhou Q., Sherwin R.P., Parrish C., Richters V., Groshen S.G., Tsao-Wei D., Markland F.S. Contortrostatin, a dimeric disintegrin from agkistrodon contortrix contortrix, inhibits breast cancer progression. Breast Cancer Res. Treat. 2000;61:249–260. doi: 10.1023/A:1006457903545. [DOI] [PubMed] [Google Scholar]
- 39.Millers E.K., Johnson L.A., Birrell G.W., Masci P.P., Lavin M.F., de Jersey J., Guddat L.W. The structure of human microplasmin in complex with textilinin-1, an aprotinin-like inhibitor from the Australian brown snake. PLoS ONE. 2013;8:e54104. doi: 10.1371/journal.pone.0054104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Flight S., Mirtschin P., Masci P.P. Comparison of active venom components between eastern brown snakes collected from south australia and queensland. Ecotoxicology. 2006;15:133–141. doi: 10.1007/s10646-005-0047-z. [DOI] [PubMed] [Google Scholar]
- 41.Lee C.Y., Huntley B.K., McCormick D.J., Ichiki T., Sangaralingham S.J., Lisy O., Burnett J.C., Jr. Cenderitide: Structural requirements for the creation of a novel dual particulate guanylyl cyclase receptor agonist with renal-enhancing in vivo and ex vivo actions. Eur. Heart J. Cardiovasc. Pharmacother. 2016;2:98–105. doi: 10.1093/ehjcvp/pvv040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Levin E.R., Gardner D.G., Samson W.K. Natriuretic peptides. N. Engl. J. Med. 1998;339:321–328. doi: 10.1056/NEJM199807303390507. [DOI] [PubMed] [Google Scholar]
- 43.St Pierre L., Flight S., Masci P.P., Hanchard K.J., Lewis R.J., Alewood P.F., de Jersey J., Lavin M.F. Cloning and characterisation of natriuretic peptides from the venom glands of australian elapids. Biochimie. 2006;88:1923–1931. doi: 10.1016/j.biochi.2006.06.014. [DOI] [PubMed] [Google Scholar]
- 44.Ferreira S.H., Rocha e Silva M. Potentiation of bradykinin and eledoisin by bpf (bradykinin potentiating factor) from bothrops jararaca venom. Experientia. 1965;21:347–349. doi: 10.1007/BF02144709. [DOI] [PubMed] [Google Scholar]
- 45.Lima S.C., Porta L.C., Lima A.D.C., Campeiro J.D., Meurer Y., Teixeira N.B., Duarte T., Oliveira E.B., Picolo G., Godinho R.O., et al. Pharmacological characterization of crotamine effects on mice hind limb paralysis employing both ex vivo and in vivo assays: Insights into the involvement of voltage-gated ion channels in the crotamine action on skeletal muscles. PLoS Negl. Trop. Dis. 2018;12:e0006700. doi: 10.1371/journal.pntd.0006700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kloog Y., Ambar I., Sokolovsky M., Kochva E., Wollberg Z., Bdolah A. Sarafotoxin, a novel vasoconstrictor peptide: Phosphoinositide hydrolysis in rat heart and brain. Science. 1988;242:268–270. doi: 10.1126/science.2845579. [DOI] [PubMed] [Google Scholar]
- 47.Weinstein S.A., Schmidt J.J., Bernheimer A.W., Smith L.A. Characterization and amino acid sequences of two lethal peptides isolated from venom of wagler′s pit viper, trimeresurus wagleri. Toxicon. 1991;29:227–236. doi: 10.1016/0041-0101(91)90107-3. [DOI] [PubMed] [Google Scholar]
- 48.Nirthanan S., Gwee M.C. Three-finger alpha-neurotoxins and the nicotinic acetylcholine receptor, forty years on. J. Pharmacol. Sci. 2004;94:1–17. doi: 10.1254/jphs.94.1. [DOI] [PubMed] [Google Scholar]
- 49.Kessler P., Marchot P., Silva M., Servent D. The three-finger toxin fold: A multifunctional structural scaffold able to modulate cholinergic functions. J. Neurochem. 2017;142:7–18. doi: 10.1111/jnc.13975. [DOI] [PubMed] [Google Scholar]
- 50.Lauridsen L.P., Laustsen A.H., Lomonte B., Gutierrez J.M. Toxicovenomics and antivenom profiling of the eastern green mamba snake (dendroaspis angusticeps) J. Proteom. 2016;136:248–261. doi: 10.1016/j.jprot.2016.02.003. [DOI] [PubMed] [Google Scholar]
- 51.Sanz L., Pla D., Perez A., Rodriguez Y., Zavaleta A., Salas M., Lomonte B., Calvete J.J. Venomic analysis of the poorly studied desert coral snake, micrurus tschudii tschudii, supports the 3ftx/pla(2) dichotomy across micrurus venoms. Toxins. 2016;8:178. doi: 10.3390/toxins8060178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Hus K.K., Buczkowicz J., Petrilla V., Petrillova M., Lyskowski A., Legath J., Bocian A. First look at the venom of naja ashei. Molecules. 2018;23:609. doi: 10.3390/molecules23030609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Olamendi-Portugal T., Batista C.V.F., Pedraza-Escalona M., Restano-Cassulini R., Zamudio F.Z., Benard-Valle M., Rafael de Roodt A., Possani L.D. New insights into the proteomic characterization of the coral snake micrurus pyrrhocryptus venom. Toxicon. 2018;153:23–31. doi: 10.1016/j.toxicon.2018.08.003. [DOI] [PubMed] [Google Scholar]
- 54.Das D., Sharma M., Kumar Das H., Pratim Sahu P., Doley R. Purification and characterization of nk-3ftx: A three finger toxin from the venom of north east indian monocled cobra. J. Biochem. Mol. Toxicol. 2016;30:59–70. doi: 10.1002/jbt.21734. [DOI] [PubMed] [Google Scholar]
- 55.Shan L.L., Gao J.F., Zhang Y.X., Shen S.S., He Y., Wang J., Ma X.M., Ji X. Proteomic characterization and comparison of venoms from two elapid snakes (bungarus multicinctus and naja atra) from china. J. Proteom. 2016;138:83–94. doi: 10.1016/j.jprot.2016.02.028. [DOI] [PubMed] [Google Scholar]
- 56.Dutta S., Chanda A., Kalita B., Islam T., Patra A., Mukherjee A.K. Proteomic analysis to unravel the complex venom proteome of eastern india naja naja: Correlation of venom composition with its biochemical and pharmacological properties. J. Proteom. 2017;156:29–39. doi: 10.1016/j.jprot.2016.12.018. [DOI] [PubMed] [Google Scholar]
- 57.Slagboom J., Otvos R.A., Cardoso F.C., Iyer J., Visser J.C., van Doodewaerd B.R., McCleary R.J.R., Niessen W.M.A., Somsen G.W., Lewis R.J., et al. Neurotoxicity fingerprinting of venoms using on-line microfluidic achbp profiling. Toxicon. 2018;148:213–222. doi: 10.1016/j.toxicon.2018.04.022. [DOI] [PubMed] [Google Scholar]
- 58.Ziganshin R.H., Kovalchuk S.I., Arapidi G.P., Starkov V.G., Hoang A.N., Thi Nguyen T.T., Nguyen K.C., Shoibonov B.B., Tsetlin V.I., Utkin Y.N. Quantitative proteomic analysis of vietnamese krait venoms: Neurotoxins are the major components in bungarus multicinctus and phospholipases a2 in bungarus fasciatus. Toxicon. 2015;107:197–209. doi: 10.1016/j.toxicon.2015.08.026. [DOI] [PubMed] [Google Scholar]
- 59.Rusmili M.R., Yee T.T., Mustafa M.R., Hodgson W.C., Othman I. Proteomic characterization and comparison of malaysian bungarus candidus and bungarus fasciatus venoms. J. Proteom. 2014;110:129–144. doi: 10.1016/j.jprot.2014.08.001. [DOI] [PubMed] [Google Scholar]
- 60.Oh A.M.F., Tan C.H., Ariaranee G.C., Quraishi N., Tan N.H. Venomics of bungarus caeruleus (indian krait): Comparable venom profiles, variable immunoreactivities among specimens from Sri Lanka, India and Pakistan. J. Proteom. 2017;164:1–18. doi: 10.1016/j.jprot.2017.04.018. [DOI] [PubMed] [Google Scholar]
- 61.Tan C.H., Wong K.Y., Tan K.Y., Tan N.H. Venom proteome of the yellow-lipped sea krait, laticauda colubrina from Bali: Insights into subvenomic diversity, venom antigenicity and cross-neutralization by antivenom. J. Proteom. 2017;166:48–58. doi: 10.1016/j.jprot.2017.07.002. [DOI] [PubMed] [Google Scholar]
- 62.Tasoulis T., Isbister G.K. A review and database of snake venom proteomes. Toxins. 2017;9:290. doi: 10.3390/toxins9090290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ainsworth S., Petras D., Engmark M., Sussmuth R.D., Whiteley G., Albulescu L.O., Kazandjian T.D., Wagstaff S.C., Rowley P., Wuster W., et al. The medical threat of mamba envenoming in sub-Saharan Africa revealed by genus-wide analysis of venom composition, toxicity and antivenomics profiling of available antivenoms. J. Proteom. 2017;172:173–189. doi: 10.1016/j.jprot.2017.08.016. [DOI] [PubMed] [Google Scholar]
- 64.Petras D., Heiss P., Harrison R.A., Sussmuth R.D., Calvete J.J. Top-down venomics of the east african green mamba, dendroaspis angusticeps, and the black mamba, dendroaspis polylepis, highlight the complexity of their toxin arsenals. J. Proteom. 2016;146:148–164. doi: 10.1016/j.jprot.2016.06.018. [DOI] [PubMed] [Google Scholar]
- 65.Laustsen A.H., Lomonte B., Lohse B., Fernandez J., Gutierrez J.M. Unveiling the nature of black mamba (dendroaspis polylepis) venom through venomics and antivenom immunoprofiling: Identification of key toxin targets for antivenom development. J. Proteom. 2015;119:126–142. doi: 10.1016/j.jprot.2015.02.002. [DOI] [PubMed] [Google Scholar]
- 66.Pawlak J., Mackessy S.P., Sixberry N.M., Stura E.A., Le Du M.H., Menez R., Foo C.S., Menez A., Nirthanan S., Kini R.M. Irditoxin, a novel covalently linked heterodimeric three-finger toxin with high taxon-specific neurotoxicity. FASEB J. 2009;23:534–545. doi: 10.1096/fj.08-113555. [DOI] [PubMed] [Google Scholar]
- 67.Pahari S., Mackessy S.P., Kini R.M. The venom gland transcriptome of the desert massasauga rattlesnake (sistrurus catenatus edwardsii): Towards an understanding of venom composition among advanced snakes (superfamily colubroidea) BMC Mol. Biol. 2007;8:115. doi: 10.1186/1471-2199-8-115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Lomonte B., Sasa M., Rey-Suarez P., Bryan W., Gutierrez J.M. Venom of the coral snake micrurus clarki: Proteomic profile, toxicity, immunological cross-neutralization, and characterization of a three-finger toxin. Toxins. 2016;8:138. doi: 10.3390/toxins8050138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Kini R.M., Doley R. Structure, function and evolution of three-finger toxins: Mini proteins with multiple targets. Toxicon. 2010;56:855–867. doi: 10.1016/j.toxicon.2010.07.010. [DOI] [PubMed] [Google Scholar]
- 70.Fry B.G. From genome to “venome”: Molecular origin and evolution of the snake venom proteome inferred from phylogenetic analysis of toxin sequences and related body proteins. Genome Res. 2005;15:403–420. doi: 10.1101/gr.3228405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Sunagar K., Jackson T.N., Undheim E.A., Ali S.A., Antunes A., Fry B.G. Three-fingered ravers: Rapid accumulation of variations in exposed residues of snake venom toxins. Toxins. 2013;5:2172–2208. doi: 10.3390/toxins5112172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Utkin Y.N. Three-finger toxins, a deadly weapon of elapid venom--milestones of discovery. Toxicon. 2013;62:50–55. doi: 10.1016/j.toxicon.2012.09.007. [DOI] [PubMed] [Google Scholar]
- 73.Wang C.I., Reeks T., Vetter I., Vergara I., Kovtun O., Lewis R.J., Alewood P.F., Durek T. Isolation and structural and pharmacological characterization of alpha-elapitoxin-dpp2d, an amidated three finger toxin from black mamba venom. Biochemistry. 2014;53:3758–3766. doi: 10.1021/bi5004475. [DOI] [PubMed] [Google Scholar]
- 74.Chang C.C., Lee C.Y. Isolation of neurotoxins from the venom of bungarus multicinctus and their modes of neuromuscular blocking action. Arch. Int. Pharmacodyn. Ther. 1963;144:241–257. [PubMed] [Google Scholar]
- 75.Changeux J.P., Kasai M., Lee C.Y. Use of a snake venom toxin to characterize the cholinergic receptor protein. Proc. Nat. Acad. Sci. USA. 1970;67:1241–1247. doi: 10.1073/pnas.67.3.1241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Chu N.S. Contribution of a snake venom toxin to myasthenia gravis: The discovery of alpha-bungarotoxin in Taiwan. J. Hist. Neurosci. 2005;14:138–148. doi: 10.1080/096470490881770. [DOI] [PubMed] [Google Scholar]
- 77.Shen X.M., Brengman J., Neubauer D., Sine S.M., Engel A.G. Investigation of congenital myasthenia reveals functional asymmetry of invariant acetylcholine receptor (achr) cys-loop aspartates. J. Biol. Chem. 2016;291:3291–3301. doi: 10.1074/jbc.M115.683995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Kryukova E.V., Shelukhina I.V., Kolacheva A.A., Alieva A.K., Shadrina M.I., Slominsky P.A., Kasheverov I.E., Utkin Y.N., Ugrumov M.V., Tsetlin V.I. Possible involvement of neuronal nicotinic acetylcholine receptors in compensatory brain mechanisms at early stages of Parkinson’s disease. Biomeditsinskaia Khimiia. 2017;63:241–247. doi: 10.18097/pbmc20176303241. [DOI] [PubMed] [Google Scholar]
- 79.Xu Y.L., Kou J.Q., Wang S.Z., Chen C.X., Qin Z.H. Neurotoxin from naja naja atra venom inhibits skin allograft rejection in rats. Int. Immunopharmacol. 2015;28:188–198. doi: 10.1016/j.intimp.2015.05.040. [DOI] [PubMed] [Google Scholar]
- 80.Hassan-Puttaswamy V., Adams D.J., Kini R.M. A distinct functional site in omega-neurotoxins: Novel antagonists of nicotinic acetylcholine receptors from snake venom. ACS Chem. Biol. 2015;10:2805–2815. doi: 10.1021/acschembio.5b00492. [DOI] [PubMed] [Google Scholar]
- 81.Jerusalinsky D., Kornisiuk E., Alfaro P., Quillfeldt J., Ferreira A., Rial V.E., Duran R., Cervenansky C. Muscarinic toxins: Novel pharmacological tools for the muscarinic cholinergic system. Toxicon. 2000;38:747–761. doi: 10.1016/S0041-0101(99)00196-8. [DOI] [PubMed] [Google Scholar]
- 82.Lyukmanova E.N., Shenkarev Z.O., Shulepko M.A., Paramonov A.S., Chugunov A.O., Janickova H., Dolejsi E., Dolezal V., Utkin Y.N., Tsetlin V.I., et al. Structural insight into specificity of interactions between nonconventional three-finger weak toxin from naja kaouthia (wtx) and muscarinic acetylcholine receptors. J. Biol. Chem. 2015;290:23616–23630. doi: 10.1074/jbc.M115.656595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Roy A., Zhou X., Chong M.Z., D′Hoedt D., Foo C.S., Rajagopalan N., Nirthanan S., Bertrand D., Sivaraman J., Kini R.M. Structural and functional characterization of a novel homodimeric three-finger neurotoxin from the venom of ophiophagus hannah (king cobra) J. Biol. Chem. 2010;285:8302–8315. doi: 10.1074/jbc.M109.074161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Antil-Delbeke S., Gaillard C., Tamiya T., Corringer P.J., Changeux J.P., Servent D., Menez A. Molecular determinants by which a long chain toxin from snake venom interacts with the neuronal alpha 7-nicotinic acetylcholine receptor. J. Biol. Chem. 2000;275:29594–29601. doi: 10.1074/jbc.M909746199. [DOI] [PubMed] [Google Scholar]
- 85.Chien K.Y., Chiang C.M., Hseu Y.C., Vyas A.A., Rule G.S., Wu W. Two distinct types of cardiotoxin as revealed by the structure and activity relationship of their interaction with zwitterionic phospholipid dispersions. J. Biol. Chem. 1994;269:14473–14483. [PubMed] [Google Scholar]
- 86.Dubovskii P.V., Konshina A.G., Efremov R.G. Cobra cardiotoxins: Membrane interactions and pharmacological potential. Curr. Med. Chem. 2014;21:270–287. doi: 10.2174/09298673113206660315. [DOI] [PubMed] [Google Scholar]
- 87.Dubovskii P.V., Lesovoy D.M., Dubinnyi M.A., Konshina A.G., Utkin Y.N., Efremov R.G., Arseniev A.S. Interaction of three-finger toxins with phospholipid membranes: Comparison of s- and p-type cytotoxins. Biochem. J. 2005;387:807–815. doi: 10.1042/BJ20041814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Konshina A.G., Dubovskii P.V., Efremov R.G. Structure and dynamics of cardiotoxins. Curr. Prot. Pept. Sci. 2012;13:570–584. doi: 10.2174/138920312803582960. [DOI] [PubMed] [Google Scholar]
- 89.Munawar A., Akrem A., Hussain A., Spencer P., Betzel C. Molecular model of cytotoxin-1 from naja mossambica mossambica venom in complex with chymotrypsin. Theor. Biol. Forum. 2015;108:89–99. [PubMed] [Google Scholar]
- 90.Girish V.M., Kumar S., Joseph L., Jobichen C., Kini R.M., Sivaraman J. Identification and structural characterization of a new three-finger toxin hemachatoxin from hemachatus haemachatus venom. PLoS ONE. 2012;7:e48112. doi: 10.1371/journal.pone.0048112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Rajagopalan N., Pung Y.F., Zhu Y.Z., Wong P.T., Kumar P.P., Kini R.M. Beta-cardiotoxin: A new three-finger toxin from ophiophagus hannah (king cobra) venom with beta-blocker activity. FASEB J. 2007;21:3685–3695. doi: 10.1096/fj.07-8658com. [DOI] [PubMed] [Google Scholar]
- 92.Tsai P.C., Fu Y.S., Chang L.S., Lin S.R. Cardiotoxin iii inhibits hepatocyte growth factor-induced epithelial-mesenchymal transition and suppresses invasion of mda-mb-231 cells. J. Biochem. Mol. Toxicol. 2016;30:12–21. doi: 10.1002/jbt.21735. [DOI] [PubMed] [Google Scholar]
- 93.Bhowmik T., Saha P.P., Sarkar A., Gomes A. Evaluation of cytotoxicity of a purified venom protein from naja kaouthia (nkct1) using gold nanoparticles for targeted delivery to cancer cell. Chem.-Biol. Interact. 2017;261:35–49. doi: 10.1016/j.cbi.2016.11.007. [DOI] [PubMed] [Google Scholar]
- 94.Nirthanan S., Charpantier E., Gopalakrishnakone P., Gwee M.C., Khoo H.E., Cheah L.S., Kini R.M., Bertrand D. Neuromuscular effects of candoxin, a novel toxin from the venom of the malayan krait (bungarus candidus) Br. J. Pharmacol. 2003;139:832–844. doi: 10.1038/sj.bjp.0705299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Diochot S., Baron A., Salinas M., Douguet D., Scarzello S., Dabert-Gay A.S., Debayle D., Friend V., Alloui A., Lazdunski M., et al. Black mamba venom peptides target acid-sensing ion channels to abolish pain. Nature. 2012;490:552–555. doi: 10.1038/nature11494. [DOI] [PubMed] [Google Scholar]
- 96.De Weille J.R., Schweitz H., Maes P., Tartar A., Lazdunski M. Calciseptine, a peptide isolated from black mamba venom, is a specific blocker of the l-type calcium channel. Proc. Nat. Acad. Sci. USA. 1991;88:2437–2440. doi: 10.1073/pnas.88.6.2437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Imanishi T., Matsushima K., Kawaguchi A., Wada T., Yoshida S., Ichida S. Increased response to high kcl-induced elevation in the intracellular-ca(2+) concentration in differentiated ng108-15 cell and the inhibitory effect of the l-type ca(2+) channel blocker, calciseptine. Neurochem. Res. 2006;31:33–40. doi: 10.1007/s11064-005-9003-9. [DOI] [PubMed] [Google Scholar]
- 98.Albrand J.P., Blackledge M.J., Pascaud F., Hollecker M., Marion D. Nmr and restrained molecular dynamics study of the three-dimensional solution structure of toxin fs2, a specific blocker of the l-type calcium channel, isolated from black mamba venom. Biochemistry. 1995;34:5923–5937. doi: 10.1021/bi00017a022. [DOI] [PubMed] [Google Scholar]
- 99.Mourier G., Salinas M., Kessler P., Stura E.A., Leblanc M., Tepshi L., Besson T., Diochot S., Baron A., Douguet D., et al. Mambalgin-1 pain-relieving peptide, stepwise solid-phase synthesis, crystal structure, and functional domain for acid-sensing ion channel 1a inhibition. J. Biol. Chem. 2016;291:2616–2629. doi: 10.1074/jbc.M115.702373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Yang D.C., Deuis J.R., Dashevsky D., Dobson J., Jackson T.N., Brust A., Xie B., Koludarov I., Debono J., Hendrikx I., et al. The snake with the scorpion′s sting: Novel three-finger toxin sodium channel activators from the venom of the long-glanded blue coral snake (calliophis bivirgatus) Toxins. 2016;8:303. doi: 10.3390/toxins8100303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Rivera-Torres I.O., Jin T.B., Cadene M., Chait B.T., Poget S.F. Discovery and characterisation of a novel toxin from dendroaspis angusticeps, named tx7335, that activates the potassium channel kcsa. Sci. Rep. 2016;6:23904. doi: 10.1038/srep23904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.McDowell R.S., Dennis M.S., Louie A., Shuster M., Mulkerrin M.G., Lazarus R.A. Mambin, a potent glycoprotein iib-iiia antagonist and platelet aggregation inhibitor structurally related to the short neurotoxins. Biochemistry. 1992;31:4766–4772. doi: 10.1021/bi00135a004. [DOI] [PubMed] [Google Scholar]
- 103.Lu X., Davies J., Lu D., Xia M., Wattam B., Shang D., Sun Y., Scully M., Kakkar V. The effect of the single substitution of arginine within the rgd tripeptide motif of a modified neurotoxin dendroaspin on its activity of platelet aggregation and cell adhesion. Cell Commun. Adhes. 2006;13:171–183. doi: 10.1080/15419060600726183. [DOI] [PubMed] [Google Scholar]
- 104.Mukherjee A.K., Kalita B., Mackessy S.P. A proteomic analysis of pakistan daboia russelii russelii venom and assessment of potency of indian polyvalent and monovalent antivenom. J. Proteom. 2016;144:73–86. doi: 10.1016/j.jprot.2016.06.001. [DOI] [PubMed] [Google Scholar]
- 105.Munawar A., Trusch M., Georgieva D., Spencer P., Frochaux V., Harder S., Arni R.K., Duhalov D., Genov N., Schluter H., et al. Venom peptide analysis of vipera ammodytes meridionalis (viperinae) and bothrops jararacussu (crotalinae) demonstrates subfamily-specificity of the peptidome in the family viperidae. Mol. Biosyst. 2011;7:3298–3307. doi: 10.1039/c1mb05309d. [DOI] [PubMed] [Google Scholar]
- 106.Munawar A., Trusch M., Georgieva D., Hildebrand D., Kwiatkowski M., Behnken H., Harder S., Arni R., Spencer P., Schluter H., et al. Elapid snake venom analyses show the specificity of the peptide composition at the level of genera naja and notechis. Toxins. 2014;6:850–868. doi: 10.3390/toxins6030850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Zupunski V., Kordis D., Gubensek F. Adaptive evolution in the snake venom kunitz/bpti protein family. FEBS Lett. 2003;547:131–136. doi: 10.1016/S0014-5793(03)00693-8. [DOI] [PubMed] [Google Scholar]
- 108.Millers E.K., Trabi M., Masci P.P., Lavin M.F., de Jersey J., Guddat L.W. Crystal structure of textilinin-1, a kunitz-type serine protease inhibitor from the venom of the australian common brown snake (pseudonaja textilis) FEBS J. 2009;276:3163–3175. doi: 10.1111/j.1742-4658.2009.07034.x. [DOI] [PubMed] [Google Scholar]
- 109.Norton R.S., Chandy K.G. Venom-derived peptide inhibitors of voltage-gated potassium channels. Neuropharmacology. 2017;127:124–138. doi: 10.1016/j.neuropharm.2017.07.002. [DOI] [PubMed] [Google Scholar]
- 110.Vanzolini K.L., Ainsworth S., Bruyneel B., Herzig V., Seraus M.G.L., Somsen G.W., Casewell N.R., Cass Q.B., Kool J. Rapid ligand fishing for identification of acetylcholinesterase-binding peptides in snake venom reveals new properties of dendrotoxins. Toxicon. 2018;152:1–8. doi: 10.1016/j.toxicon.2018.06.080. [DOI] [PubMed] [Google Scholar]
- 111.Skarzynski T. Crystal structure of alpha-dendrotoxin from the green mamba venom and its comparison with the structure of bovine pancreatic trypsin inhibitor. J. Mol. Biol. 1992;224:671–683. doi: 10.1016/0022-2836(92)90552-U. [DOI] [PubMed] [Google Scholar]
- 112.Jin L., Wu Y. Molecular mechanism of delta-dendrotoxin-potassium channel recognition explored by docking and molecular dynamic simulations. J. Mol. Recognit. 2011;24:101–107. doi: 10.1002/jmr.1031. [DOI] [PubMed] [Google Scholar]
- 113.Katoh E., Nishio H., Inui T., Nishiuchi Y., Kimura T., Sakakibara S., Yamazaki T. Structural basis for the biological activity of dendrotoxin-i, a potent potassium channel blocker. Biopolymers. 2000;54:44–57. doi: 10.1002/(SICI)1097-0282(200007)54:1<44::AID-BIP50>3.0.CO;2-Z. [DOI] [PubMed] [Google Scholar]
- 114.Wang F.C., Bell N., Reid P., Smith L.A., McIntosh P., Robertson B., Dolly J.O. Identification of residues in dendrotoxin k responsible for its discrimination between neuronal k+ channels containing kv1.1 and 1.2 alpha subunits. Eur. J. Biochem. 1999;263:222–229. doi: 10.1046/j.1432-1327.1999.00494.x. [DOI] [PubMed] [Google Scholar]
- 115.Yang W., Feng J., Wang B., Cao Z., Li W., Wu Y., Chen Z. Bf9, the first functionally characterized snake toxin peptide with kunitz-type protease and potassium channel inhibiting properties. J. Biochem. Mol. Toxicol. 2014;28:76–83. doi: 10.1002/jbt.21538. [DOI] [PubMed] [Google Scholar]
- 116.Wang X.C., Wang S., Zhang M., Gao F., Yin C., Li H., Zhang Y., Hu S.J., Duan J.H. Alpha-dendrotoxin-sensitive kv1 channels contribute to conduction failure of polymodal nociceptive c-fibers from rat coccygeal nerve. J. Neurophysiol. 2016;115:947–957. doi: 10.1152/jn.00786.2014. [DOI] [PubMed] [Google Scholar]
- 117.Thakur R., Mukherjee A.K. Pathophysiological significance and therapeutic applications of snake venom protease inhibitors. Toxicon. 2017;131:37–47. doi: 10.1016/j.toxicon.2017.03.011. [DOI] [PubMed] [Google Scholar]
- 118.Earl S.T., Masci P.P., de Jersey J., Lavin M.F., Dixon J. Drug development from australian elapid snake venoms and the venomics pipeline of candidates for haemostasis: Textilinin-1 (q8008), haempatch (q8009) and covase (v0801) Toxicon. 2012;59:456–463. doi: 10.1016/j.toxicon.2010.12.010. [DOI] [PubMed] [Google Scholar]
- 119.Flight S., Johnson L., Trabi M., Gaffney P., Lavin M., de Jersey J., Masci P. Comparison of textilinin-1 with aprotinin as serine protease inhibitors and as antifibrinolytic agents. Pathophysiol. Haemost. Thromb. 2005;34:188–193. doi: 10.1159/000092421. [DOI] [PubMed] [Google Scholar]
- 120.Masci P.P., Whitaker A.N., Sparrow L.G., de Jersey J., Winzor D.J., Watters D.J., Lavin M.F., Gaffney P.J. Textilinins from pseudonaja textilis textilis. Characterization of two plasmin inhibitors that reduce bleeding in an animal model. Blood Coagul. Fibrinol. 2000;11:385–393. doi: 10.1097/00001721-200006000-00011. [DOI] [PubMed] [Google Scholar]
- 121.Flight S.M., Johnson L.A., Du Q.S., Warner R.L., Trabi M., Gaffney P.J., Lavin M.F., de Jersey J., Masci P.P. Textilinin-1, an alternative anti-bleeding agent to aprotinin: Importance of plasmin inhibition in controlling blood loss. Br. J. Haematol. 2009;145:207–211. doi: 10.1111/j.1365-2141.2009.07605.x. [DOI] [PubMed] [Google Scholar]
- 122.Vivas J., Ibarra C., Salazar A.M., Neves-Ferreira A.G., Sanchez E.E., Perales J., Rodriguez-Acosta A., Guerrero B. Purification and characterization of tenerplasminin-1, a serine peptidase inhibitor with antiplasmin activity from the coral snake (micrurus tener tener) venom. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2016;179:107–115. doi: 10.1016/j.cbpc.2015.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Morjen M., Honore S., Bazaa A., Abdelkafi-Koubaa Z., Ellafi A., Mabrouk K., Kovacic H., El Ayeb M., Marrakchi N., Luis J. Pivl, a snake venom kunitz-type serine protease inhibitor, inhibits in vitro and in vivo angiogenesis. Microvasc. Res. 2014;95:149–156. doi: 10.1016/j.mvr.2014.08.006. [DOI] [PubMed] [Google Scholar]
- 124.Morjen M., Kallech-Ziri O., Bazaa A., Othman H., Mabrouk K., Zouari-Kessentini R., Sanz L., Calvete J.J., Srairi-Abid N., El Ayeb M., et al. Pivl, a new serine protease inhibitor from macrovipera lebetina transmediterranea venom, impairs motility of human glioblastoma cells. Matrix Biol. 2013;32:52–62. doi: 10.1016/j.matbio.2012.11.015. [DOI] [PubMed] [Google Scholar]
- 125.Earl S.T., Richards R., Johnson L.A., Flight S., Anderson S., Liao A., de Jersey J., Masci P.P., Lavin M.F. Identification and characterisation of kunitz-type plasma kallikrein inhibitors unique to oxyuranus sp. Snake venoms. Biochimie. 2012;94:365–373. doi: 10.1016/j.biochi.2011.08.003. [DOI] [PubMed] [Google Scholar]
- 126.Saviola A.J., Modahl C.M., Mackessy S.P. Disintegrins of crotalus simus tzabcan venom: Isolation, characterization and evaluation of the cytotoxic and anti-adhesion activities of tzabcanin, a new rgd disintegrin. Biochimie. 2015;116:92–102. doi: 10.1016/j.biochi.2015.07.005. [DOI] [PubMed] [Google Scholar]
- 127.Calvete J.J. The continuing saga of snake venom disintegrins. Toxicon. 2013;62:40–49. doi: 10.1016/j.toxicon.2012.09.005. [DOI] [PubMed] [Google Scholar]
- 128.Kini R.M. Accelerated evolution of toxin genes: Exonization and intronization in snake venom disintegrin/metalloprotease genes. Toxicon. 2018;148:16–25. doi: 10.1016/j.toxicon.2018.04.005. [DOI] [PubMed] [Google Scholar]
- 129.Bilgrami S., Yadav S., Kaur P., Sharma S., Perbandt M., Betzel C., Singh T.P. Crystal structure of the disintegrin heterodimer from saw-scaled viper (echis carinatus) at 1.9 a resolution. Biochemistry. 2005;44:11058–11066. doi: 10.1021/bi050849y. [DOI] [PubMed] [Google Scholar]
- 130.Juarez P., Comas I., Gonzalez-Candelas F., Calvete J.J. Evolution of snake venom disintegrins by positive darwinian selection. Mol. Biol. Evol. 2008;25:2391–2407. doi: 10.1093/molbev/msn179. [DOI] [PubMed] [Google Scholar]
- 131.Wierzbicka-Patynowski I., Niewiarowski S., Marcinkiewicz C., Calvete J.J., Marcinkiewicz M.M., McLane M.A. Structural requirements of echistatin for the recognition of alpha(v)beta(3) and alpha(5)beta(1) integrins. J. Biol. Chem. 1999;274:37809–37814. doi: 10.1074/jbc.274.53.37809. [DOI] [PubMed] [Google Scholar]
- 132.Walsh E.M., Marcinkiewicz C. Non-rgd-containing snake venom disintegrins, functional and structural relations. Toxicon. 2011;58:355–362. doi: 10.1016/j.toxicon.2011.07.004. [DOI] [PubMed] [Google Scholar]
- 133.Kwon I., Hong S.Y., Kim Y.D., Nam H.S., Kang S., Yang S.H., Heo J.H. Thrombolytic effects of the snake venom disintegrin saxatilin determined by novel assessment methods: A fecl3-induced thrombosis model in mice. PLoS ONE. 2013;8:e81165. doi: 10.1371/journal.pone.0081165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.David V., Succar B.B., de Moraes J.A., Saldanha-Gama R.F.G., Barja-Fidalgo C., Zingali R.B. Recombinant and chimeric disintegrins in preclinical research. Toxins. 2018;10:321. doi: 10.3390/toxins10080321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Peerlinck K., De Lepeleire I., Goldberg M., Farrell D., Barrett J., Hand E., Panebianco D., Deckmyn H., Vermylen J., Arnout J. Mk-383 (l-700,462), a selective nonpeptide platelet glycoprotein iib/iiia antagonist, is active in man. Circulation. 1993;88:1512–1517. doi: 10.1161/01.CIR.88.4.1512. [DOI] [PubMed] [Google Scholar]
- 136.Scarborough R.M., Naughton M.A., Teng W., Rose J.W., Phillips D.R., Nannizzi L., Arfsten A., Campbell A.M., Charo I.F. Design of potent and specific integrin antagonists. Peptide antagonists with high specificity for glycoprotein iib-iiia. J. Biol. Chem. 1993;268:1066–1073. [PubMed] [Google Scholar]
- 137.Vink S., Jin A.H., Poth K.J., Head G.A., Alewood P.F. Natriuretic peptide drug leads from snake venom. Toxicon. 2012;59:434–445. doi: 10.1016/j.toxicon.2010.12.001. [DOI] [PubMed] [Google Scholar]
- 138.Yoshimura M., Yasue H., Morita E., Sakaino N., Jougasaki M., Kurose M., Mukoyama M., Saito Y., Nakao K., Imura H. Hemodynamic, renal, and hormonal responses to brain natriuretic peptide infusion in patients with congestive heart failure. Circulation. 1991;84:1581–1588. doi: 10.1161/01.CIR.84.4.1581. [DOI] [PubMed] [Google Scholar]
- 139.Kinnunen P., Vuolteenaho O., Ruskoaho H. Mechanisms of atrial and brain natriuretic peptide release from rat ventricular myocardium: Effect of stretching. Endocrinology. 1993;132:1961–1970. doi: 10.1210/endo.132.5.8477647. [DOI] [PubMed] [Google Scholar]
- 140.Suga S., Itoh H., Komatsu Y., Ogawa Y., Hama N., Yoshimasa T., Nakao K. Cytokine-induced c-type natriuretic peptide (cnp) secretion from vascular endothelial cells--evidence for cnp as a novel autocrine/paracrine regulator from endothelial cells. Endocrinology. 1993;133:3038–3041. doi: 10.1210/endo.133.6.8243333. [DOI] [PubMed] [Google Scholar]
- 141.Potter L.R., Yoder A.R., Flora D.R., Antos L.K., Dickey D.M. Natriuretic peptides: Their structures, receptors, physiologic functions and therapeutic applications. Handb. Exp. Pharmacol. 2009;191:341–366. doi: 10.1007/978-3-540-68964-5_15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Volpe M. Natriuretic peptides and cardio-renal disease. Int. J. Cardiol. 2014;176:630–639. doi: 10.1016/j.ijcard.2014.08.032. [DOI] [PubMed] [Google Scholar]
- 143.Sridharan S., Kini R.M. Decoding the molecular switches of natriuretic peptides which differentiate its vascular and renal functions. Biochem. J. 2018;475:399–413. doi: 10.1042/BCJ20170690. [DOI] [PubMed] [Google Scholar]
- 144.Sridharan S., Kini R.M. Tail wags the dog: Activity of krait natriuretic peptide is determined by its c-terminal tail in a natriuretic peptide receptor-independent manner. Biochem. J. 2015;469:255–266. doi: 10.1042/BJ20150281. [DOI] [PubMed] [Google Scholar]
- 145.Reeks T., Jones A., Brust A., Sridharan S., Corcilius L., Wilkinson B.L., Thaysen-Andersen M., Payne R.J., Kini R.M., Daly N.L., et al. A defined alpha-helix in the bifunctional o-glycosylated natriuretic peptide tcnpa from the venom of tropidechis carinatus. Angew. Chem. Int. Ed. Engl. 2015;54:4828–4831. doi: 10.1002/anie.201411914. [DOI] [PubMed] [Google Scholar]
- 146.Fry B.G., Vidal N., Norman J.A., Vonk F.J., Scheib H., Ramjan S.F., Kuruppu S., Fung K., Hedges S.B., Richardson M.K., et al. Early evolution of the venom system in lizards and snakes. Nature. 2006;439:584–588. doi: 10.1038/nature04328. [DOI] [PubMed] [Google Scholar]
- 147.Gutierrez J.M., Lomonte B., Leon G., Rucavado A., Chaves F., Angulo Y. Trends in snakebite envenomation therapy: Scientific, technological and public health considerations. Curr. Pharm. Des. 2007;13:2935–2950. doi: 10.2174/138161207782023784. [DOI] [PubMed] [Google Scholar]
- 148.Fry B.G., Wuster W. Assembling an arsenal: Origin and evolution of the snake venom proteome inferred from phylogenetic analysis of toxin sequences. Mol. Biol. Evol. 2004;21:870–883. doi: 10.1093/molbev/msh091. [DOI] [PubMed] [Google Scholar]
- 149.Chen H.H., Lainchbury J.G., Burnett J.C., Jr. Natriuretic peptide receptors and neutral endopeptidase in mediating the renal actions of a new therapeutic synthetic natriuretic peptide dendroaspis natriuretic peptide. J. Am. Coll. Cardiol. 2002;40:1186–1191. doi: 10.1016/S0735-1097(02)02127-7. [DOI] [PubMed] [Google Scholar]
- 150.Zhang Y., Wu J., Yu G., Chen Z., Zhou X., Zhu S., Li R., Zhang Y., Lu Q. A novel natriuretic peptide from the cobra venom. Toxicon. 2011;57:134–140. doi: 10.1016/j.toxicon.2010.10.014. [DOI] [PubMed] [Google Scholar]
- 151.Tourki B., Mateo P., Morand J., Elayeb M., Godin-Ribuot D., Marrakchi N., Belaidi E., Messadi E. Lebetin 2, a snake venom-derived natriuretic peptide, attenuates acute myocardial ischemic injury through the modulation of mitochondrial permeability transition pore at the time of reperfusion. PLoS ONE. 2016;11:e0162632. doi: 10.1371/journal.pone.0162632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Ondetti M.A., Williams N.J., Sabo E.F., Pluscec J., Weaver E.R., Kocy O. Angiotensin-converting enzyme inhibitors from the venom of bothrops jararaca. Isolation, elucidation of structure, and synthesis. Biochemistry. 1971;10:4033–4039. doi: 10.1021/bi00798a004. [DOI] [PubMed] [Google Scholar]
- 153.Cushman D.W., Ondetti M.A. Design of angiotensin converting enzyme inhibitors. Nat. Med. 1999;5:1110–1113. doi: 10.1038/13423. [DOI] [PubMed] [Google Scholar]
- 154.Van Vark L.C., Bertrand M., Akkerhuis K.M., Brugts J.J., Fox K., Mourad J.J., Boersma E. Angiotensin-converting enzyme inhibitors reduce mortality in hypertension: A meta-analysis of randomized clinical trials of renin–angiotensin–aldosterone system inhibitors involving 158 998 patients. Eur. Heart J. 2012;33:2088–2097. doi: 10.1093/eurheartj/ehs075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Munawar A., Zahid A., Negm A., Akrem A., Spencer P., Betzel C. Isolation and characterization of bradykinin potentiating peptides from agkistrodon bilineatus venom. Proteome Sci. 2016;14:1. doi: 10.1186/s12953-016-0090-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Morais K.L., Ianzer D., Miranda J.R., Melo R.L., Guerreiro J.R., Santos R.A., Ulrich H., Lameu C. Proline rich-oligopeptides: Diverse mechanisms for antihypertensive action. Peptides. 2013;48:124–133. doi: 10.1016/j.peptides.2013.07.016. [DOI] [PubMed] [Google Scholar]
- 157.Almeida J.R., Resende L.M., Watanabe R.K., Carregari V.C., Huancahuire-Vega S., Coutinho-Neto A., Soares A.M., Vale N., Marangoni S., Da S.S. Snake venom peptides and low mass proteins: Molecular tools and therapeutic agents. Curr. Med. Chem. 2017;24:3254–3282. doi: 10.2174/0929867323666161028155611. [DOI] [PubMed] [Google Scholar]
- 158.Hayashi M.A., Ligny-Lemaire C., Wollberg Z., Wery M., Galat A., Ogawa T., Muller B.H., Lamthanh H., Doljansky Y., Bdolah A., et al. Long-sarafotoxins: Characterization of a new family of endothelin-like peptides. Peptides. 2004;25:1243–1251. doi: 10.1016/j.peptides.2004.05.010. [DOI] [PubMed] [Google Scholar]
- 159.Ducancel F. Endothelin-like peptides. Cell. Mol. Life Sci. 2005;62:2828–2839. doi: 10.1007/s00018-005-5286-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Quinton L., Le Caer J.P., Phan G., Ligny-Lemaire C., Bourdais-Jomaron J., Ducancel F., Chamot-Rooke J. Characterization of toxins within crude venoms by combined use of fourier transform mass spectrometry and cloning. Anal. Chem. 2005;77:6630–6639. doi: 10.1021/ac050575k. [DOI] [PubMed] [Google Scholar]
- 161.Mourier G., Hajj M., Cordier F., Zorba A., Gao X., Coskun T., Herbet A., Marcon E., Beau F., Delepierre M., et al. Pharmacological and structural characterization of long-sarafotoxins, a new family of endothelin-like peptides: Role of the c-terminus extension. Biochimie. 2012;94:461–470. doi: 10.1016/j.biochi.2011.08.014. [DOI] [PubMed] [Google Scholar]
- 162.Kloog Y., Sokolovsky M. Similarities in mode and sites of action of sarafotoxins and endothelins. Trends Pharmacol. Sci. 1989;10:212–214. doi: 10.1016/0165-6147(89)90261-7. [DOI] [PubMed] [Google Scholar]
- 163.Malaquin S., Bayat S., Abou Arab O., Mourier G., Lorne E., Kamel S., Dupont H., Ducancel F., Mahjoub Y. Respiratory effects of sarafotoxins from the venom of different atractaspis genus snake species. Toxins. 2016;8:215. doi: 10.3390/toxins8070215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Mahjoub Y., Malaquin S., Mourier G., Lorne E., Abou Arab O., Massy Z.A., Dupont H., Ducancel F. Short- versus long-sarafotoxins: Two structurally related snake toxins with very different in vivo haemodynamic effects. PLoS ONE. 2015;10:e0132864. doi: 10.1371/journal.pone.0132864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Xu X., Li B., Zhu S., Rong R. Hypotensive peptides from snake venoms: Structure, function and mechanism. Curr. Top. Med. Chem. 2015;15:658–669. doi: 10.2174/1568026615666150217113835. [DOI] [PubMed] [Google Scholar]
- 166.Lal H., Woodward B., Williams K.I. Actions of endothelins and sarafotoxin 6c in the rat isolated perfused lung. Br. J. Pharmacol. 1995;115:653–659. doi: 10.1111/j.1476-5381.1995.tb14982.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Lal H., Woodward B., Williams K.I. Investigation of the contributions of nitric oxide and prostaglandins to the actions of endothelins and sarafotoxin 6c in rat isolated perfused lungs. Br. J. Pharmacol. 1996;118:1931–1938. doi: 10.1111/j.1476-5381.1996.tb15627.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Francis B., Kaiser I.I. Inhibition of metalloproteinases in bothrops asper venom by endogenous peptides. Toxicon. 1993;31:889–899. doi: 10.1016/0041-0101(93)90224-7. [DOI] [PubMed] [Google Scholar]
- 169.Huang K.F., Hung C.C., Wu S.H., Chiou S.H. Characterization of three endogenous peptide inhibitors for multiple metalloproteinases with fibrinogenolytic activity from the venom of taiwan habu (trimeresurus mucrosquamatus) Biochem. Biophys. Res. Commun. 1998;248:562–568. doi: 10.1006/bbrc.1998.9017. [DOI] [PubMed] [Google Scholar]
- 170.Wagstaff S.C., Favreau P., Cheneval O., Laing G.D., Wilkinson M.C., Miller R.L., Stocklin R., Harrison R.A. Molecular characterisation of endogenous snake venom metalloproteinase inhibitors. Biochem. Biophys. Res. Commun. 2008;365:650–656. doi: 10.1016/j.bbrc.2007.11.027. [DOI] [PubMed] [Google Scholar]
- 171.Yee K.T., Pitts M., Tongyoo P., Rojnuckarin P., Wilkinson M.C. Snake venom metalloproteinases and their peptide inhibitors from myanmar russell′s viper venom. Toxins. 2016;9:15. doi: 10.3390/toxins9010015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Bernardes C.P., Santos N.A.G., Sisti F.M., Ferreira R.S., Santos-Filho N.A., Cintra A.C.O., Cilli E.M., Sampaio S.V., Santos A.C. A synthetic snake-venom-based tripeptide (glu-val-trp) protects pc12 cells from mpp(+) toxicity by activating the ngf-signaling pathway. Peptides. 2018;104:24–34. doi: 10.1016/j.peptides.2018.04.012. [DOI] [PubMed] [Google Scholar]
- 173.Ding B., Xu Z., Qian C., Jiang F., Ding X., Ruan Y., Ding Z., Fan Y. Antiplatelet aggregation and antithrombosis efficiency of peptides in the snake venom of deinagkistrodon acutus: Isolation, identification, and evaluation. Evid. Based Complement. Altern. Med. 2015;2015:412841. doi: 10.1155/2015/412841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Kerkis I., Hayashi M.A., Prieto da Silva A.R., Pereira A., De Sa Junior P.L., Zaharenko A.J., Radis-Baptista G., Kerkis A., Yamane T. State of the art in the studies on crotamine, a cell penetrating peptide from south american rattlesnake. Biomed. Res. Int. 2014;2014:675985. doi: 10.1155/2014/675985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Coronado M.A., Gabdulkhakov A., Georgieva D., Sankaran B., Murakami M.T., Arni R.K., Betzel C. Structure of the polypeptide crotamine from the brazilian rattlesnake crotalus durissus terrificus. Acta Crystallogr. Sect. D Biol. Crystallogr. 2013;69:1958–1964. doi: 10.1107/S0907444913018003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Pereira A., Kerkis A., Hayashi M.A., Pereira A.S., Silva F.S., Oliveira E.B., Prieto da Silva A.R., Yamane T., Radis-Baptista G., Kerkis I. Crotamine toxicity and efficacy in mouse models of melanoma. Expert Opin. Investig. Drugs. 2011;20:1189–1200. doi: 10.1517/13543784.2011.602064. [DOI] [PubMed] [Google Scholar]
- 177.Papo N., Shai Y. Host defense peptides as new weapons in cancer treatment. Cell. Mol. Life Sci. 2005;62:784–790. doi: 10.1007/s00018-005-4560-2. [DOI] [PubMed] [Google Scholar]
- 178.Giorgi R., Bernardi M.M., Cury Y. Analgesic effect evoked by low molecular weight substances extracted from crotalus durissus terrificus venom. Toxicon. 1993;31:1257–1265. doi: 10.1016/0041-0101(93)90399-4. [DOI] [PubMed] [Google Scholar]
- 179.Batista da Cunha D., Pupo Silvestrini A.V., Gomes da Silva A.C., Maria de Paula Estevam D., Pollettini F.L., de Oliveira Navarro J., Alves A.A., Remedio Zeni Beretta A.L., Annichino Bizzacchi J.M., Pereira L.C., et al. Mechanistic insights into functional characteristics of native crotamine. Toxicon. 2018;146:1–12. doi: 10.1016/j.toxicon.2018.03.007. [DOI] [PubMed] [Google Scholar]
- 180.Mambelli-Lisboa N.C., Mozer Sciani J., Rossan Brandao Prieto da Silva A., Kerkis I. Co-localization of crotamine with internal membranes and accentuated accumulation in tumor cells. Molecules. 2018;23:968. doi: 10.3390/molecules23040968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Banigan J.R., Mandal K., Sawaya M.R., Thammavongsa V., Hendrickx A.P., Schneewind O., Yeates T.O., Kent S.B. Determination of the X-ray structure of the snake venom protein omwaprin by total chemical synthesis and racemic protein crystallography. Prot. Sci. 2010;19:1840–1849. doi: 10.1002/pro.468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Torres A.M., Wong H.Y., Desai M., Moochhala S., Kuchel P.W., Kini R.M. Identification of a novel family of proteins in snake venoms. Purification and structural characterization of nawaprin from naja nigricollis snake venom. J. Biol. Chem. 2003;278:40097–40104. doi: 10.1074/jbc.M305322200. [DOI] [PubMed] [Google Scholar]
- 183.Nair D.G., Fry B.G., Alewood P., Kumar P.P., Kini R.M. Antimicrobial activity of omwaprin, a new member of the waprin family of snake venom proteins. Biochem. J. 2007;402:93–104. doi: 10.1042/BJ20060318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Lavergne V., Alewood P.F., Mobli M., King G.F. The structural universe of disulfide-rich venom peptides. In: King G., editor. Venoms to Drugs: Venom as a Source for the Development of Human Therapeutics. Royal Society of Chemistry; Cambridge, UK: 2015. pp. 57–58. [Google Scholar]
- 185.St Pierre L., Earl S.T., Filippovich I., Sorokina N., Masci P.P., De Jersey J., Lavin M.F. Common evolution of waprin and kunitz-like toxin families in australian venomous snakes. Cell. Mol. Life Sci. 2008;65:4039–4054. doi: 10.1007/s00018-008-8573-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Viala V.L., Hildebrand D., Trusch M., Fucase T.M., Sciani J.M., Pimenta D.C., Arni R.K., Schluter H., Betzel C., Mirtschin P., et al. Venomics of the australian eastern brown snake (pseudonaja textilis): Detection of new venom proteins and splicing variants. Toxicon. 2015;107:252–265. doi: 10.1016/j.toxicon.2015.06.005. [DOI] [PubMed] [Google Scholar]
- 187.Campos P.F., Andrade-Silva D., Zelanis A., Paes Leme A.F., Rocha M.M., Menezes M.C., Serrano S.M., Junqueira-de-Azevedo Ide L. Trends in the evolution of snake toxins underscored by an integrative omics approach to profile the venom of the colubrid phalotris mertensi. Genome Biol. Evol. 2016;8:2266–2287. doi: 10.1093/gbe/evw149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Schmidt J.J., Weinstein S.A., Smith L.A. Molecular properties and structure-function relationships of lethal peptides from venom of wagler′s pit viper, trimeresurus wagleri. Toxicon. 1992;30:1027–1036. doi: 10.1016/0041-0101(92)90047-9. [DOI] [PubMed] [Google Scholar]
- 189.Tan C.H., Tan K.Y., Yap M.K., Tan N.H. Venomics of tropidolaemus wagleri, the sexually dimorphic temple pit viper: Unveiling a deeply conserved atypical toxin arsenal. Sci. Rep. 2017;7:43237. doi: 10.1038/srep43237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Zainal Abidin S.A., Rajadurai P., Chowdhury M.E., Ahmad Rusmili M.R., Othman I., Naidu R. Proteomic characterization and comparison of malaysian tropidolaemus wagleri and cryptelytrops purpureomaculatus venom using shotgun-proteomics. Toxins. 2016;8:299. doi: 10.3390/toxins8100299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Debono J., Xie B., Violette A., Fourmy R., Jaeger M., Fry B.G. Viper venom botox: The molecular origin and evolution of the waglerin peptides used in anti-wrinkle skin cream. J. Mol. Evol. 2017;84:8–11. doi: 10.1007/s00239-016-9764-6. [DOI] [PubMed] [Google Scholar]
- 192.Molles B.E., Rezai P., Kline E.F., McArdle J.J., Sine S.M., Taylor P. Identification of residues at the alpha and epsilon subunit interfaces mediating species selectivity of waglerin-1 for nicotinic acetylcholine receptors. J. Biol. Chem. 2002;277:5433–5440. doi: 10.1074/jbc.M109232200. [DOI] [PubMed] [Google Scholar]
- 193.Tajbakhsh M., Akhavan M.M., Fallah F., Karimi A. A recombinant snake cathelicidin derivative peptide: Antibiofilm properties and expression in escherichia coli. Biomolecules. 2018;8:118. doi: 10.3390/biom8040118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Perumal Samy R., Stiles B.G., Franco O.L., Sethi G., Lim L.H.K. Animal venoms as antimicrobial agents. Biochem. Pharmacol. 2017;134:127–138. doi: 10.1016/j.bcp.2017.03.005. [DOI] [PubMed] [Google Scholar]
- 195.Falcao C.B., de La Torre B.G., Perez-Peinado C., Barron A.E., Andreu D., Radis-Baptista G. Vipericidins: A novel family of cathelicidin-related peptides from the venom gland of south american pit vipers. Amino Acids. 2014;46:2561–2571. doi: 10.1007/s00726-014-1801-4. [DOI] [PubMed] [Google Scholar]
- 196.Jesupret C., Baumann K., Jackson T.N., Ali S.A., Yang D.C., Greisman L., Kern L., Steuten J., Jouiaei M., Casewell N.R., et al. Vintage venoms: Proteomic and pharmacological stability of snake venoms stored for up to eight decades. J. Proteom. 2014;105:285–294. doi: 10.1016/j.jprot.2014.01.004. [DOI] [PubMed] [Google Scholar]
- 197.Dutertre S., Undheim E.A.B., Pineda S.S., Jin A.H., Lavergne V., Fry B.G., Lewis R.J., Alewood P.F., King G.F. Venoms-based drug discovery: Proteomic and transcriptomic approaches. RSC Drug Discov. 2015;11:80–96. [Google Scholar]
- 198.Meyer A., Dierks K., Hilterhaus D., Klupsch T., Muhlig P., Kleesiek J., Schopflin R., Einspahr H., Hilgenfeld R., Betzel C. Single-drop optimization of protein crystallization. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2012;68:994–998. doi: 10.1107/S1744309112024074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Oberthuer D., Melero-Garcia E., Dierks K., Meyer A., Betzel C., Garcia-Caballero A., Gavira J.A. Monitoring and scoring counter-diffusion protein crystallization experiments in capillaries by in situ dynamic light scattering. PLoS ONE. 2012;7:e33545. doi: 10.1371/journal.pone.0033545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Guo Y.Z., Sun L.H., Oberthuer D., Zhang C.Y., Shi J.Y., Di J.L., Zhang B.L., Cao H.L., Liu Y.M., Li J., et al. Utilisation of adsorption and desorption for simultaneously improving protein crystallisation success rate and crystal quality. Sci. Rep. 2014;4:7308. doi: 10.1038/srep07308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Fry B.G., Undheim E.A.B., Jakson T.N.W., Roelants K., Grorgieva D., Vetter I., Calvete J.J., Scheib H., Cribb B.W., Yang D.C., et al. Research methods. In: Fry B.G., editor. Venomous Reptiles and their Toxins: Evolution, Pathophysiology, and Biodiscovery. Oxford University Press; New York, NY, USA: 2015. p. 169. [Google Scholar]
- 202.Sequeira A.F., Turchetto J., Saez N.J., Peysson F., Ramond L., Duhoo Y., Blemont M., Fernandes V.O., Gama L.T., Ferreira L.M., et al. Gene design, fusion technology and tev cleavage conditions influence the purification of oxidized disulphide-rich venom peptides in escherichia coli. Microb. Cell Fact. 2017;16:4. doi: 10.1186/s12934-016-0618-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Turchetto J., Sequeira A.F., Ramond L., Peysson F., Bras J.L., Saez N.J., Duhoo Y., Blemont M., Guerreiro C.I., Quinton L., et al. High-throughput expression of animal venom toxins in escherichia coli to generate a large library of oxidized disulphide-reticulated peptides for drug discovery. Microb. Cell Fact. 2017;16:6. doi: 10.1186/s12934-016-0617-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Klupczynska A., Pawlak M., Kokot Z.J., Matysiak J. Application of metabolomic tools for studying low molecular-weight fraction of animal venoms and poisons. Toxins. 2018;10:306. doi: 10.3390/toxins10080306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Gorai B., Sivaraman T. Delineating residues for haemolytic activities of snake venom cardiotoxin 1 from naja naja as probed by molecular dynamics simulations and in vitro validations. Int. J. Biol. Macromol. 2017;95:1022–1036. doi: 10.1016/j.ijbiomac.2016.10.091. [DOI] [PubMed] [Google Scholar]
- 206.Khan N., Niazi Z.R., Khan N.R., Shah K., Rehman K., Wahab A., Khan S. Simulation studies and dynamic interaction of venom peptides with ion channels. Prot. Pept. Lett. 2018;25:652–662. doi: 10.2174/0929866525666180619095245. [DOI] [PubMed] [Google Scholar]