

Abstract
Cholesterol synthesis is a ubiquitous and housekeeping metabolic pathway that leads to cholesterol, an essential structural component of mammalian cell membranes, required for proper membrane permeability and fluidity. The last part of the pathway involves steroidal triterpenes with cholestane ring structures. It starts by conversion of acyclic squalene into lanosterol, the first sterol intermediate of the pathway, followed by production of 20 structurally very similar steroidal triterpene molecules in over 11 complex enzyme reactions. Due to the structural similarities of sterol intermediates and the broad substrate specificity of the enzymes involved (especially sterol-Δ24-reductase; DHCR24) the exact sequence of the reactions between lanosterol and cholesterol remains undefined. This article reviews all hitherto known structures of post-squalene steroidal triterpenes of cholesterol synthesis, their biological roles and the enzymes responsible for their synthesis. Furthermore, it summarises kinetic parameters of enzymes (Vmax and Km) and sterol intermediate concentrations from various tissues. Due to the complexity of the post-squalene cholesterol synthesis pathway, future studies will require a comprehensive meta-analysis of the pathway to elucidate the exact reaction sequence in different tissues, physiological or disease conditions. A major reason for the standstill of detailed late cholesterol synthesis research was the lack of several steroidal triterpene standards. We aid to this efforts by summarizing commercial and laboratory standards, referring also to chemical syntheses of meiosis-activating sterols.
Keywords: sterols, steroidal triterpenes, cholesterol synthesis, post-squalene pathway
1. Introduction
Cholesterol synthesis is a housekeeping pathway believed to take place in virtually all cells in mammals where cholesterol molecules are synthesized from C2 acetyl residues in more than 20 reactions. The synthesis starts with the acetylated coenzyme A (acetyl CoA) molecule from which all sterol carbon atoms are derived. Acetoacetyl-CoA is formed from two molecules of acetyl-CoA by acetoacetyl-CoA-thiolase and is further combined with one molecule of acetyl-CoA to form 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA) by HMG-CoA synthase. In the next stage HMG-CoA reductase (HMGCR) which is the rate-limiting enzyme of cholesterol synthesis and thus also the target of cholesterol lowering drugs (statins) converts HMG-CoA to mevalonate (Scheme 1). The latter is further metabolised to farnesyl pyrophosphate (farnesyl-PP) by several enzymes. This point presents the first branching of the pathway. One branch serves for further synthesis of cholesterol while the other ends in farnesyl, dolichol, heme A and ubiquinone (Q10). The early part of cholesterol synthesis is completed by the combination of two molecules of farnesyl-PP to squalene by the enzyme squalene synthase 1 (FDFT1) [1]. The post-squalene cholesterol synthesis consists of steroidal triterpenes with cholestane ring structures. It starts with lanosterol, the first cyclic intermediate, which is formed from squalene by squalene epoxidase/monooxigenase (SQLE) and lanosterol synthase (LSS). Then follows a series of demethylation, dehydrogenation and isomerisation reactions of lanosterol leading towards the final product cholesterol (Scheme 1). A detailed metabolic reaction network using the information available from the literature [2,3,4,5,6] and from the pathways databases (Kyoto Encyclopedia of Genes and Genomes—KEGG, BioCyc database collection, LIPID MAPS Consortium) is shown in Scheme 1, where the complexity of the post-squalene phase is presented. Initially, the post-squalene pathway has been divided into the Bloch and Kandutsch-Russell branches [5]. In the Bloch branch, the final reaction is the conversion of desmosterol to cholesterol by sterol-Δ24-reductase (DHCR24); thus, all intermediates from lanosterol to desmosterol contain Δ24 double bonds. In contrast, in the Kandutsch-Russell branch, DHCR24 acts already on lanosterol; thus all intermediates from 24,25-dihydrolanosterol to 7-dehydrocholesterol contain a saturated side chain. Since DHCR24 can, in principle, metabolise any cholesterol synthesis intermediate from lanosterol on, the two branches cannot be treated separately. Study of DHCR24 substrate specificity in vitro [7] showed 24-dehydrolathosterol as the most reactive substrate, suggesting that cholesterol synthesis preferentially starts with the Bloch branch from lanosterol to 24-dehydrolathosterol and is then shifted to Kandutsch-Russell branch via lathosterol. In this case 7-dehydrocholesterol presents the last intermediate before cholesterol. In the present work we attempted to summarise all hitherto known structures of post-squalene steroidal triterpenes of cholesterol synthesis, their biological roles and the enzymes responsible for their synthesis. The article also summarises commercial and laboratory standards together with syntheses of meiosis-activating sterols (MAS), and the kinetic parameters of enzymes and sterol concentrations from various tissues.
Scheme 1.

Cholesterol synthesis scheme. For abbreviations see Table 1.
2. Nomenclature and Numbering
The conventional definition for triterpenes is any isoprenoid having thirty carbon atoms formed from six isoprene (C5-) units. They can be acyclic, as in squalene, or cyclic, as in the tetracycles with a side chain typified by lanosterol [8]. The steroidal triterpenes possess the steroid ring structure and in this article we will refer to the steroidal triterpenes as sterols. The IUPAC-IUB recommendation for numbering of tetracyclic triterpenoid skeleton is shown in Figure 1. 5α-Cholestane is used as a basis for the IUPAC chemical nomenclature in Table 2 [9].
Figure 1.

Chemical structure of 5α-cholestane (20R) and the IUPAC-IUB recommendations for the atom numbering.
Table 2.
Sterols of the post-squalene cholesterol synthesis: trivial and chemical names, structures, formulas and molecular weights (Mw).
| Trivial name | Chemical name | Structure | Formula | Mw |
|---|---|---|---|---|
| Lanosterol | 4,4,14α-trimethyl-5α-cholesta-8(9),24-dien-3β-ol |
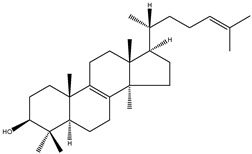
|
C30H50O | 426.72 |
| 24,25-dihydrolanosterol | 4,4,14α-trimethyl-5α-cholest-8(9)en-3β-ol |
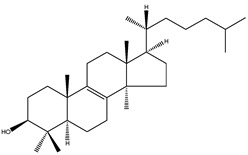
|
C30H52O | 428.73 |
| FF-MAS | 4,4-dimethyl-5α-cholesta-8(9),14,24-trien-3β-ol |
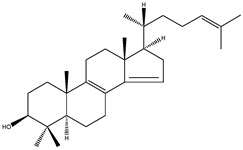
|
C29H46O | 410.67 |
| Dihydro-FF-MAS | 4,4-dimethyl-5α-cholesta-8(9),14-dien-3β-ol |
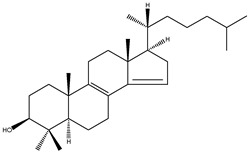
|
C29H48O | 412.69 |
| T-MAS | 4,4-dimethy-5α-cholesta-8(9),24-dien-3β-ol |
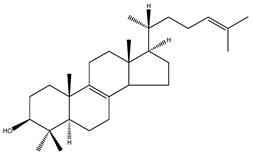
|
C29H48O | 412.69 |
| Dihydro-T-MAS | 4,4-dimethy-5α-cholest-8(9)-en-3β-ol |
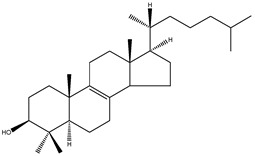
|
C29H50O | 414.69 |
| Zymosterol | 5α-cholesta-8(9),24-dien-3β-ol |
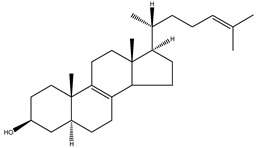
|
C27H44O | 384.64 |
| Zymostenol | 5α-cholest-8(9)-en-3β-ol |
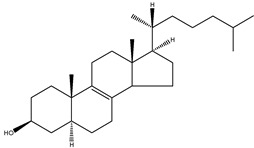
|
C27H46O | 386.65 |
| 24-dehydrolathosterol | 5α-cholesta-7,24-dien-3β-ol |
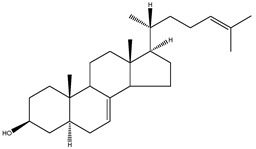
|
C27H44O | 384.64 |
| Lathosterol | 5α-cholest-7-en-3β-ol |
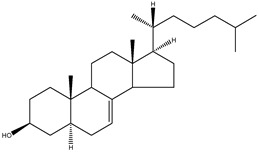
|
C27H46O | 386.66 |
| 7-dehydrodesmosterol | 5α-cholesta-5,7,24-trien-3β-ol |
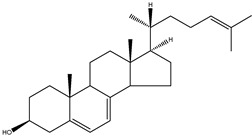
|
C27H46O | 386.66 |
| 7-dehydrocholesterol | 5α-cholesta-5,7-dien-3β-ol |
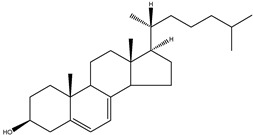
|
C27H44O | 384.64 |
| Desmosterol | 5α-cholesta-5,24-dien-3β-ol |
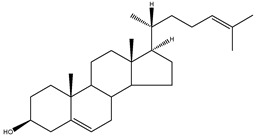
|
C27H44O | 384.64 |
| Cholesterol | 5α-cholest-5-en-3β-ol |
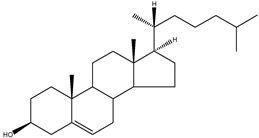
|
C27H46O | 386.66 |
3. Commercial and Laboratory Standards Together with Some Chemical Syntheses
Availability of sterol standards is crucial for cholesterol synthesis pathway studies. Table 3 summarises the hitherto commercially available sterol standards, including their providers. Lanosterol is readily available from different providers (Table 3) since it is a major constituent (about 15%) of the unsaponifiable portion of wool fat (lanoline) and is thus easily chemically isolated. The first isolation from wool fat was achieved in 1956 by Bloch and co-workers [10]. In 1960 Kandutsch and Russell identified 24,25-dihydrolanosterol, another sterol previously found only in wool fat, in preputial gland tumors inoculated subcutaneously into the axial region of C57BL/g mice. In this manner they obtained relatively large amounts of sterols for characterisation [11]. This study lead to further experiments and ultimately resulted in the discovery of Kandutsh-Russell branch of cholesterol synthesis [5]. 24,25-Dihydrolanosterol standard is also available from some providers (Table 3). 7-Dehydrocholesterol, lathosterol and desmosterol are also commercially available (Table 3), but are more expensive than lanosterol. The first problematic standard is zymosterol. It is listed on the Steraloids web page but upon inquiry you realise that it is actually not available. This situation has existed since 2007 when the chemist responsible for the synthesis of zymosterol retired. Luckily, zymosterol and also zymostenol recently became available from Avanti Polar Lipids (Table 3). This was confirmed by our own inquiry.
Table 3.
Commercial availability of unlabelled and labelled sterol standards.
| Sterol | Providers | ||
|---|---|---|---|
| Unlabelled | Deuterium labelled | Tritium labelled | |
| Lanosterol | APL a, SA b, S c, ARC f | APL | ARC |
| 24,25-dihydrolanosterol | APL, S | APL | ARC |
| FF-MAS | APL | ||
| T-MAS | APL | APL | |
| Zymosterol | APL | APL, MI d | |
| Zymostenol | APL | ||
| Lathosterol | APL, SA, S | APL, MI | |
| 7-dehydrodesmosterol | MI | MI | |
| 7-dehydrocholesterol | APL, SA, S, MI | APL, CIL e | |
| Desmosterol | APL, SA, S, MI | APL, MI | ARC |
a APL, Avanti Polar Lipids (Alabaster, AL, USA; http://avantilipids.com); b SA, Sigma-Aldrich (St. Louis, MO, USA; http://www.sigmaaldrich.com); c S, Steraloids (Newport, RI, USA; http://steraloids.com); d MI, Medical Isotopes (Pelham, NH, USA; http://www.medicalisotopes.com); e CIL, Cambridge Isotope Laboratories (Andover, MA, USA; http://www.isotope.com); f ARC, American Radiolabelled Chemicals (St. Louis, MO, USA; http://www.arc-inc.com)
Availability of follicular fluid meiosis-activating sterol (FF-MAS) and testis meiosis-activating sterol (T-MAS) standards presented a problem for the broader research community in the past. FF-MAS and T-MAS were discovered in the laboratory of Prof. Dr. Anne Grete Byskov (The Juliane Marie Center for Children, Women and Reproduction, University Hospital of Copenhagen, Copenhagen, Denmark) who first reported their isolation from follicular fluid and bull semen [12]. In collaboration with Prof. Dr. Ingemar Björkhem (Karolinska Institute, Stockholm, Sweden) we developed a powerful gas chromatography/mass spectrometry method (GC-MS) allowing quantitative analysis of several structurally similar cholesterol precursors (desmosterol, 7-dehydrocholesterol, lathosterol, zymosterol, 24,25-dihydrolanosterol, lanosterol, FF-MAS, T-MAS) in a single chromatographic run. At that time we used the Byskov standards since this was the only resource available to us. We now recommend application and extention of the aforementioned GC-MS method with FF-MAS and T-MAS standards that were recently made commercially available by Avanti Polar Lipids (Table 3). Table 3 also summarises commercially available labelled sterol standards which are necessary for method validation and precise extraction procedures of biological samples. The purity of sterol standards in Table 3 depends on the price. Most of the Avanti Polar Lipids standards are available in more than 99% purity.
In the case that FF-MAS and T-MAS standards would no longer be available from commercial providers they can be extracted from biological samples as described previously [12]. Several chemical syntheses of FF-MAS [13,14,15] have also been reported. A relatively simple, six-step reaction synthesis of FF-MAS from 3β-acetoxy-4,4-dimethyl-5α-cholest-8(14)-en-15-one was published by Ruan et al. [13,15] while Murray et al. [14] described an alternative synthesis of FF-MAS from lithocholic acid (3α-hydroxy-5β-cholan-24-oic acid) in a ten-step reaction. A six-step chemical synthesis reaction of T-MAS from 3β-acetoxy-4,4-dimethyl-5α-cholest-8(14)-en-15-one was also reported by Ruan et al. [13]. Chemical synthesis of dihydro-FF-MAS in a five-step reaction from cholest-4-en-3-one was described by Schroepfer et al. [16].
4. Enzymes of Post-Squalene Cholesterol Synthesis Pathway and Their Biological Significance
For nearly half a century the remarkable enzymatic cyclization of 2,3-oxidosqualene to form lanosterol (Figure 1) has both fascinated and challenged the molecular sciences. The reaction is catalysed by LSS and a crystal structure of the human membrane protein has been reported [17]. The 2,3-oxidosqualene is stereoselectively cyclised and skeletally rearranged in a single enzyme reaction to form the chair-boat-chair conformation of lanosterol. For a detailed mechanism see Wendt et al. [18]. The subsequent enzyme of the post-squalene cholesterol synthesis pathway, lanosterol-14α-demethylase (CYP51), catalyses the oxidative removal of 14α-methyl group of lanosterol and 24,25-dihydrolanosterol to produce FF-MAS or its derivative without the Δ24 bond, dihydro-FF-MAS (Figure 1) [19]. The next step is Δ14-reduction of FF-MAS or dihydro-FF-MAS into T-MAS or its Δ24-reduced derivative dihydro-T-MAS. Two enzymes encoded by two different genes [sterol-Δ14-reductase (DHCR14) and lamin B receptor (LBR)] can perform the same enzymatic reaction [20,21]. Formation of zymosterol or its Δ24-reduced derivative zymostenol is catalysed by the sterol 4,4-dimethylase complex. First sterol-C4-methyl oxidase (SC4MOL) carboxylates one of the 4-methyl groups of T-MAS or its Δ24-reduced derivative and 3β-hydroxy-Δ5-steroid-dehydrogenase (NSDHL) coverts the 3β-hydroxy group to a 3-keto one by removal of CO2. Finally 3β-keto-reductase (HSD17B7) restores the 3β-hydroxy group to yield 4-methylzymosterol or its Δ24-reduced derivative 4-methylzymostenol [6]. The procedure is repeated once more to synthesise zymosterol or zymostenol which are further transformed to 24-dehydrolathosterol and lathosterol by sterol-Δ8-7-isomerase (EBP) which catalyses the shift of a Δ8(9) to a Δ7 double bond. Sterol-C5-desaturase/lathosterol oxidase (SC5DL) catalyses the formation of the C5-double bond by converting aforementioned sterols to 7-dehydrodesmosterol and 7-dehydro-cholesterol, respectively. Reduction of the Δ7 double bond is catalysed by sterol-Δ7-reductase (DHCR7) to convert 7-dehydrodesmosterol to desmosterol and 7-dehyrocholesterol to the final product cholesterol [22,23]. Cholesterol is formed from desmosterol by DHCR24 catalysing the reduction of Δ24 double bond. As aforementioned (see Introduction) all intermediates of the Bloch pathway which contain an unsaturated Δ24 bond, can act as substrates for DHCR24 with 24-dehydrolathosterol having the highest DHCR24 affinity (Table 4) [7]. All enzymes of the post-squalene cholesterol synthesis pathway reside in endoplasmic reticulum being membrane bound [24]. Enzymes and gene abbreviations are listed in Table 1. Enzyme kinetic parameters are listed in Table 4.
Table 4.
Kinetic parameters of cholesterol synthesis enzymes with specific sterols as substrates.
| Enzyme | Substrate sterol | Vmax [nmol/min per mg of protein] | Km [µM] | kcat [Vmax/Km] | Relative kcat (fold) | Reference |
|---|---|---|---|---|---|---|
| CYP51A1 | Lanosterol | 262.3 | 10.5 | 24.1 | [25] | |
| 24,25-dihydrolanosterol | 377.4 | 20.0 | 18.87 | |||
| DHCR14 | 4,4-dimethy-5α-cholesta-7,24-dien-3β-ol | 1.13 | 29.4 | [26] | ||
| EBP | Zymosterol | 0.325 | 25 | [27] | ||
| SC5DL | Lathosterol | 0.017 | 15.4a | [28] | ||
| DHCR7 | 7-dehydrocholesterol | 0.85 | 30 | [22] | ||
| DHCR24 | Lanosterol | 0.361 | 109 | 0.0033 | 1.0 | [7] |
| Zymosterol | 3.550 | 176 | 0.0201 | 6.1 | ||
| 7-dehydrodesmosterol | 2.176 | 37 | 0.0586 | 17.8 | ||
| Desmosterol | 2.945 | 163 | 0.0180 | 5.5 |
a nmol/mg of membrane protein.
Table 1.
Enzymes/gene symbols, names and Enzyme Commission numbers.
| Enzyme/Gene symbol | Enzyme/Gene name | Enzyme Commission number |
|---|---|---|
| HMGCR | 3-hydroxy-3-methylglutaryl-CoA reductase | EC:1.1.1.34 |
| FDFT1 | farnesyl-diphosphate farnesyl transferase 1 | EC:2.5.1.21 |
| SQLE | squalene epoxidase/monooxygenase | EC:1.14.13.132 |
| LSS | lanosterol synthase | EC:5.4.99.7 |
| CYP51A1 | lanosterol-14α-demethylase | EC:1.14.13.70 |
| DHCR14 (TM7SF2) | sterol-Δ14-reductase | EC:1.3.1.70 |
| LBR | lamin B receptor | 3930 a |
| SC4MOL | sterol-C4-methyl oxidase | EC:1.14.13.72 |
| NSDHL | 3β-hydroxy-Δ5-steroid-dehydrogenase | EC:1.1.1.170 |
| HSD17B7 | 3β-keto-reductase | EC:1.1.1.270 |
| EBP | sterol-Δ8-7-isomerase | EC:5.3.3.5 |
| SC5DL | sterol-C5-desaturase/lathosterol oxidase | EC:1.14.21.6 |
| DHCR7 | sterol-Δ7-reductase | EC:1.3.1.21 |
| DHCR24 | sterol-Δ24-reductase | EC:1.3.1.72 |
a KEGG Entry number (not listed as an enzyme).
The human malformations and the mouse knock-out models of enzymes from cholesterol synthesis show their high biological importance. In humans, there are six inherited disorders linked to the post-squalene cholesterol synthesis. The Smith-Lemli-Opitz syndrome (SLOS) is the most common in humans. Patients with this syndrome are characterized by significantly reduced plasma, cell and tissue cholesterol levels and elevated concentrations of 7-dehydrocholesterol and its isomer 8-dehydrocholesterol [29], which are a consequence of DHCR7 defects, as DHCR7 is responsible for conversion of 7-dehydrocholesterol to cholesterol. Desmosterolosis, a rare autosomal recessive disease, is characterized by a number of developmental abnormalities and increased desmosterol concentration in serum and tissues as a consequence of DHCR24 gene mutations [30]. Humans with the mutations in the gene for SC5DL exhibit reduced cholesterol and increased lathosterol levels in plasma, tissues and cell cultures, therefore the disease name lathosterolosis [30,31,32]. HEM dysplasia, the last autosomal recessive disease is caused by the defects of LBR gene, which serves as a sterol Δ14-reductase of post-squalene cholesterol synthesis. It is very rare, but when expressed it is lethal in an early stage of embryonic development [33]. CHILD and Conradi-Hunermann-Happle syndrome are inherited in an X-linked dominant fashion. Patients with CHILD syndrome have a defective gene for NSDHL [34] and the Conradi-Hunermann-Happle syndrome is characterized by the defect of Δ8-7 sterol isomerase enzyme, due to the mutations in EBP gene [35]. The biological significance of the previously mentioned cholesterol synthesis enzymes is confirmed by different mouse knock-out models as defects are generally lethal in mice. The disorders in humans and corresponding knock-out mouse models are summarised in a recent review [4].
5. Biological Role and Toxicity
Sterols are essential components of eukaryote membranes. Their incorporation enhances the packing of acyl chains of phospholipids in the hydrophobic bilayer, increases its mechanical strength, and reduces its permeability [36]. Cholesterol is the sterol found in animal cell membranes. It is also a precursor of bile acids and different steroid hormones such as cortisol, aldosterone, progesterone, estrogen and testosterone. Approximately one quarter of total cholesterol is of dietary origin, while three-quarters of it comes from endogenous synthesis [37]. Using skin fibroblasts obtained from SLOS [3] patients, Tulenko et al. [38] demonstrated that the SLOS membrane has increased 7-dehydrochlesterol and reduced cholesterol content and abnormal membrane fluidity. X-ray diffraction analyses of synthetic membranes prepared to mimic SLOS membranes revealed atypical membrane organization and thus have increased membrane fluidity.
Another biological role of post-squalene cholesterol synthesis intermediates is that abnormal concentrations accelerate the degradation of HMGCR. Song and Javitt [39] reported that lanosterol and 24,25-dihydrolanosterol stimulate ubiquitination of HMGCR, although with reduced efficiency compared to 25-hydroxycholesterol (approximately 3- and 10-fold for 24,25-dihydrolanosterol and lanosterol, respectively). Thus, the HMGCR-ubiquitinating activity of the methylated sterols coincides with their ability to diminish steady-state levels of the enzyme. The mechanism is reviewed in more detail by Goldstein et al. [40]. Fitzky et al. [41] have shown that 7-dehydocholesterol also inhibits cholesterol synthesis by accelerating the degradation of HMGCR protein.
In 1995 two biologically occurring meiosis-activating sterols (MAS) were discovered, FF-MAS and T-MAS [12]. T-MAS concentration in mammalian testes is above 30 µg/g, whereas FF-MAS is present only in trace amounts. FF-MAS is found at high concentrations in the mammalian ovary and T-MAS is also present at half of the FF-MAS concentrations [12,42]. Experiments demonstrated the ability of MAS (FF-MAS, T-MAS, dihydro-FF-MAS and dihydro-T-MAS) to trigger the resumption of meiosis in cultured mouse oocytes in vitro [12] but not in vivo [43]. In addition, MAS compounds were demonstrated to have positive influence on cytoplasmic maturation, an important process occurring during the final step of oocyte development that enables completion of nuclear maturation, fertilization, and proper early embryo development. In contrast to the initially proposed role of MAS in the resumption of meiosis, it was suggested that they might have another role in meiotic nuclear processes and cytoplasmic maturation; however, the exact mechanism of MAS in female reproduction remains controversial [44].
Spermatogenesis and sperm maturation represents another major biological role of sterols [45]. The development of highly specialised spermatozoa requires extensive remodelling of membrane structure, involving stage specific changes in membrane permeability and fluidity. Desmosterol is the predominant sterol in monkey and rabbit spermatozoa and was found to accumulate preferentially in the sperm tail. The difference in membrane desmosterol composition between sperm heads and tails might underline the different functions of these two structures. Desmosterol has an additional double bond between carbon atoms 24 and 25 compared to cholesterol, and might contribute to membrane fluidity, which is necessary for the motility of flagella [46]. A review of Keber et al. [45] gives a more detailed description of sterol roles in spermatogenesis and sperm maturation. Sterol concentrations in various tissues of rats and mice are presented in Table 5.
Table 5.
Total sterol concentrations (free + esterified) in various tissues of rats and mice.
| Organism | Sterol | Blood plasma [µg/mL] | Liver [µg/g] | Brain [µg/g] | Testis [µg/g] |
|---|---|---|---|---|---|
| Mus musculus a | Lanosterol | 0.08–0.86 | 0.92–7.12 | 3.2–4.7 | 4.3–17.8 |
| 24,25-dihydrolanosterol | 0.08–1.17 | 0.10–0.13 | |||
| FF-MAS | 1.10–2.15 | ||||
| T-MAS | 0.61–5.65 | 4.70–8.50 | 31–110 | ||
| Zymosterol | |||||
| Lathosterol | 25.3–30.8 | 7.6–11.6 | |||
| 7-dehydrocholesterol | 0.22–2.39 | 0.94–9.31 | 3.31–7.70 | 2.6–4.8 | |
| Desmosterol | 0.43–2.27 | 0.78–4.14 | 29.4–97.8 | 39–91 | |
| Cholesterol | 405–1,997 | 1,965–6,700 | 12,500–15,500 | 1,337–2,829 | |
| Rattus | Lanosterol | 0.75–1.38 b | ~4 c | ||
| norvegicus | 24,25-dihydrolanosterol | ||||
| FF-MAS | |||||
| T-MAS | ~2 c | ~49 c | |||
| Zymosterol | |||||
| Lathosterol | 15.5–10.4 b | ||||
| 7-dehydrocholesterol | 2.63–3.31 b | ||||
| Desmosterol | 4.55–5.39 b | ~2 c | |||
| Cholesterol | 589–856 b | 2,041–2,618 b | ~1544 c |
a Liver values from Acimovic et al. [47], brain values from Ali et al. [48], blood plasma and testis values from our unpublished data (liver data as mass per wet tissue weight); b Values from Bjorkhem-Bergman et al. [49] (data as mass per wet tissue weight); c Values from Tacer et al. [50] (data as mass per dry tissue weight).
The toxic effects of sterols have been reported by Xu et al. [51]. CHO-7 cells which are a sub-line of CHO-K1 cells were grown in medium containing physiological concentrations of desmosterol, lathosterol and lanosterol. Lanosterol supplemented cells not only failed to support growth but also killed the cells whereas lathosterol only failed to support growth and desmosterol showed no toxic effect, which was confirmed by Rodriguez-Acebes et al. [52], who showed that desmosterol can replace cholesterol in cell proliferation. Accumulation of lanosterol is therefore considered as toxic. Another toxic effect was reported by accumulation of desmosterol in humans in 1960’s. Triparanol (MER-29), a cholesterol-lowering agent, was marketed in United States from 1959 onwards, but was withdrawn by the U.S. Food and Drug Administration in 1962 because the patients receiving triparanol developed cataracts as a result of notable accumulations of desmosterol [53,54,55].
6. Conclusions
Triterpene steroid metabolomics is an important part of lipid metabolomics. In recent years we have learned about many more biological roles of these molecules than had been previously anticipated. For example, the post-squalene portion of cholesterol synthesis was believed to have only one goal: to produce cholesterol. However, we now know of some other biological roles of these intermediates that are not dedicated to cholesterol, such as activation of oocyte meiosis, at least in vitro. The cholesterol synthesis pathway seems to hide additional secrets. Recent data show that sterol intermediates can serve as direct substrates for enzymes of steroid hormone synthesis which is stimulating further research in this area. From our own experience, the quantitative sterol metabolomics is not trivial and requires specialized protocols and of course standards that are frequently difficult to obtain. The aim of this manuscript was thus to review the current state of the art and standard resources in triterpene steroid metabolomics with focus on the cholesterol synthesis to facilitate research in this important area.
Acknowledgments
This work was supported by the Slovenian Research Agency programme grant P1-0104 and project J7-4053.
Conflicts of Interest
The authors declare no conflict of interest.
Footnotes
Sample Availability: Not available.
References
- 1.Goldstein J.L., Brown M.S. Regulation of the Mevalonate Pathway. Nature. 1990;343:425–430. doi: 10.1038/343425a0. [DOI] [PubMed] [Google Scholar]
- 2.Griffiths W.J., Wang Y. Analysis of neurosterols by GC-MS and LC-MS/MS. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2009;877:2778–2805. doi: 10.1016/j.jchromb.2009.05.017. [DOI] [PubMed] [Google Scholar]
- 3.Porter F.D. Malformation syndromes due to inborn errors of cholesterol synthesis. J. Clin. Invest. 2002;110:715–724. doi: 10.1172/JCI16386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Horvat S., McWhir J., Rozman D. Defects in cholesterol synthesis genes in mouse and in humans: lessons for drug development and safer treatments. Drug Metab. Rev. 2011;43:69–90. doi: 10.3109/03602532.2010.540580. [DOI] [PubMed] [Google Scholar]
- 5.Kandutsch A.A., Russell A.E. Preputial gland tumor sterols. 3. A metabolic pathway from lanosterol to cholesterol. J. Biol. Chem. 1960;235:2256–2261. [PubMed] [Google Scholar]
- 6.Marijanovic Z., Laubner D., Moller G., Gege C., Husen B., Adamski J., Breitling R. Closing the gap: identification of human 3-ketosteroid reductase, the last unknown enzyme of mammalian cholesterol biosynthesis. Mol. Endocrinol. 2003;17:1715–1725. doi: 10.1210/me.2002-0436. [DOI] [PubMed] [Google Scholar]
- 7.Bae S.H., Paik Y.K. Cholesterol biosynthesis from lanosterol: development of a novel assay method and characterization of rat liver microsomal lanosterol delta 24-reductase. Biochem. J. 1997;326:609–616. doi: 10.1042/bj3260609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Liu J., Nes W.D. Steroidal triterpenes: design of substrate-based inhibitors of ergosterol and sitosterol synthesis. Molecules. 2009;14:4690–4706. doi: 10.3390/molecules14114690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Goad L.J., Akishisa T. Analysis of Sterols. Blackie Academic & Professional; London, UK: 1997. p. 20. Chapter 1. [Google Scholar]
- 10.Bloch K., Gautschi F., Johnston J.D. Isolation of lanosterol from isocholesterol. J. Biol. Chem. 1957;224:185–190. [PubMed] [Google Scholar]
- 11.Kandutsch A.A., Russell A.E. Preputial gland tumor sterols. I. The occurrence of 24,25-dihydrolanosterol and a comparison with liver and the normal gland. J. Biol. Chem. 1959;234:2037–2042. [PubMed] [Google Scholar]
- 12.Byskov A.G., Andersen C.Y., Nordholm L., Thogersen H., Xia G., Wassmann O., Andersen J.V., Guddal E., Roed T. Chemical structure of sterols that activate oocyte meiosis. Nature. 1995;374:559–562. doi: 10.1038/374559a0. [DOI] [PubMed] [Google Scholar]
- 13.Ruan B., Watanabe S., Eppig J.J., Kwoh C., Dzidic N., Pang J., Wilson W.K., Schroepfer G.J., Jr. Sterols affecting meiosis: novel chemical syntheses and the biological activity and spectral properties of the synthetic sterols. J. Lipid Res. 1998;39:2005–2020. [PubMed] [Google Scholar]
- 14.Murray A., Gronvald F.C., Nielsen J.K., Faarup P. Synthesis of FF-MAS from lithocholic acid. Bioorg. Med. Chem. Lett. 2000;10:1067–1068. doi: 10.1016/S0960-894X(00)00160-8. [DOI] [PubMed] [Google Scholar]
- 15.Ruan B., Wilson W.K., Schroepfer G.J., Jr. An alternative synthesis of 4,4-dimethyl-5 alpha-cholesta-8,14,24-trien-3 beta-ol, an intermediate in sterol biosynthesis and a reported activator of meiosis and of nuclear orphan receptor LXR alph. Bioorg. Med. Chem. Lett. 1998;8:233–236. doi: 10.1016/S0960-894X(98)00016-X. [DOI] [PubMed] [Google Scholar]
- 16.Schroepfer G.J., Jr., Parish E.J., Tsuda M., Kandutsch A.A. Inhibitors of sterol synthesis. Chemical syntheses, properties and effects of 4,4-dimethyl-15-oxygenated sterols on sterol synthesis and on 3-hydroxy-3-methylglutaryl coenzyme A reductase activity in cultured mammalian cells. Chem. Phys. Lipids. 1998;47:187–207. doi: 10.1016/0009-3084(88)90012-6. [DOI] [PubMed] [Google Scholar]
- 17.Thoma R., Schulz-Gasch T., D’Arcy B., Benz J., Aebi J., Dehmlow H., Hennig M., Stihle M., Ruf A. Insight into steroid scaffold formation from the structure of human oxidosqualene cyclase. Nature. 2004;432:118–122. doi: 10.1038/nature02993. [DOI] [PubMed] [Google Scholar]
- 18.Wendt K.U., Schulz G.E., Corey E.J., Liu D.R. Enzyme Mechanisms for Polycyclic Triterpene Formation. Angew. Chem. Int. Ed. Engl. 2000;39:2812–2833. doi: 10.1002/1521-3773(20000818)39:16<2812::AID-ANIE2812>3.0.CO;2-#. [DOI] [PubMed] [Google Scholar]
- 19.Debeljak N., Fink M., Rozman D. Many facets of mammalian lanosterol 14alpha-demethylase from the evolutionarily conserved cytochrome P450 family CYP51. Arch. Biochem. Biophys. 2003;409:159–171. doi: 10.1016/S0003-9861(02)00418-6. [DOI] [PubMed] [Google Scholar]
- 20.Bennati A.M., Schiavoni G., Franken S., Piobbico D., Della Fazia M.A., Caruso D., de Fabiani E., Benedetti L., Cusella De Angelis M.G., Gieselmann V., et al. Disruption of the gene encoding 3beta-hydroxysterol Delta-reductase (Tm7sf2) in mice does not impair cholesterol biosynthesis. FEBS J. 2008;275:5034–5047. doi: 10.1111/j.1742-4658.2008.06637.x. [DOI] [PubMed] [Google Scholar]
- 21.Wassif C.A., Brownson K.E., Sterner A.L., Forlino A., Zerfas P.M., Wilson W.K., Starost M.F., Porter F.D. HEM dysplasia and ichthyosis are likely laminopathies and not due to 3beta-hydroxysterol Delta14-reductase deficiency. Hum. Mol. Genet. 2007;16:1176–1187. doi: 10.1093/hmg/ddm065. [DOI] [PubMed] [Google Scholar]
- 22.Moebius F.F., Fitzky B.U., Lee J.N., Paik Y.K., Glossmann H. Molecular cloning and expression of the human delta7-sterol reductase. Proc. Natl. Acad. Sci. USA. 1998;95:1899–1902. doi: 10.1073/pnas.95.4.1899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bae S.H., Lee J.N., Fitzky B.U., Seong J., Paik Y.K. Cholesterol biosynthesis from lanosterol. Molecular cloning, tissue distribution, expression, chromosomal localization, and regulation of rat 7-dehydrocholesterol reductase, a Smith-Lemli-Opitz syndrome-related protein. J. Biol. Chem. 1999;274:14624–14631. doi: 10.1074/jbc.274.21.14624. [DOI] [PubMed] [Google Scholar]
- 24.Rozman D., Monostory K. Perspectives of the non-statin hypolipidemic agents. Pharmacol. Ther. 2010;127:19–40. doi: 10.1016/j.pharmthera.2010.03.007. [DOI] [PubMed] [Google Scholar]
- 25.Nitahara Y., Aoyama Y., Horiuchi T., Noshiro M., Yoshida Y. Purification and characterization of rat sterol 14-demethylase P450 (CYP51) expressed in Escherichia coli. J. Biochem. 1999;126:927–933. doi: 10.1093/oxfordjournals.jbchem.a022536. [DOI] [PubMed] [Google Scholar]
- 26.Paik Y.K., Trzaskos J.M., Shafiee A., Gaylor J.L. Microsomal enzymes of cholesterol biosynthesis from lanosterol. Characterization, solubilization, and partial purification of NADPH-dependent delta 8,14-steroid 14-reductase. J. Biol. Chem. 1984;259:13413–13423. [PubMed] [Google Scholar]
- 27.Moebius F.F., Reiter R.J., Bermoser K., Glossmann H., Cho S.Y., Paik Y.K. Pharmacological analysis of sterol delta8-delta7 isomerase proteins with [3H]ifenprodil. Mol. Pharmacol. 1998;54:591–598. doi: 10.1124/mol.54.3.591. [DOI] [PubMed] [Google Scholar]
- 28.Takakuwa Y., Nishino H., Ishibe Y., Ishibashi T. Properties and kinetics of membrane-bound enzymes when both the enzyme and substrate are components of the same microsomal membrane. Studies on lathosterol 5-desaturase. J. Biol. Chem. 1994;269:27889–27893. [PubMed] [Google Scholar]
- 29.Kelley R.I., Hennekam R.C. The Smith-Lemli-Opitz syndrome. J. Med. Genet. 2000;37:321–335. doi: 10.1136/jmg.37.5.321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Waterham H.R., Koster J., Romeijn G.J., Hennekam R.C., Vreken P., Andersson H.C., FitzPatrick D.R., Kelley R.I., Wanders R.J. Mutations in the 3beta-hydroxysterol Delta24-reductase gene cause desmosterolosis, an autosomal recessive disorder of cholesterol biosynthesis. Am. J. Hum. Genet. 2001;69:685–694. doi: 10.1086/323473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Brunetti-Pierri N., Corso G., Rossi M., Ferrari P., Balli F., Rivasi F., Annunziata I., Ballabio A., Russo A.D., Andria G., et al. Lathosterolosis, a novel multiple-malformation/mental retardation syndrome due to deficiency of 3beta-hydroxysteroid-delta5-desaturase. Am. J. Hum. Genet. 2002;71:952–958. doi: 10.1086/342668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Krakowiak P.A., Wassif C.A., Kratz L., Cozma D., Kovarova M., Harris G., Grinberg A., Yang Y., Hunter A.G., Tsokos M., et al. Lathosterolosis: An inborn error of human and murine cholesterol synthesis due to lathosterol 5-desaturase deficiency. Hum. Mol. Genet. 2003;12:1631–1641. doi: 10.1093/hmg/ddg172. [DOI] [PubMed] [Google Scholar]
- 33.Waterham H.R., Koster J., Mooyer P., Noort Gv G., Kelley R.I., Wilcox W.R., Wanders R.J., Hennekam R.C., Oosterwijk J.C. Autosomal recessive HEM/Greenberg skeletal dysplasia is caused by 3 beta-hydroxysterol delta 14-reductase deficiency due to mutations in the lamin B receptor gene. Am. J. Hum. Genet. 2003;72:1013–1017. doi: 10.1086/373938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Konig A., Happle R., Bornholdt D., Engel H., Grzeschik K.H. Mutations in the NSDHL gene, encoding a 3beta-hydroxysteroid dehydrogenase, cause CHILD syndrom. Am. J. Med. Genet. 2000;90:339–346. doi: 10.1002/(SICI)1096-8628(20000214)90:4<339::AID-AJMG15>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- 35.Braverman N., Lin P., Moebius F.F., Obie C., Moser A., Glossmann H., Wilcox W.R., Rimoin D.L., Smith M., Kratz L., et al. Mutations in the gene encoding 3 beta-hydroxysteroid-delta 8, delta 7-isomerase cause X-linked dominant Conradi-Hunermann syndrome. Nat. Genet. 1999;22:291–294. doi: 10.1038/10357. [DOI] [PubMed] [Google Scholar]
- 36.Bloch K.E. Sterol structure and membrane function. CRC Crit. Rev. Biochem. 1983;14:47–92. doi: 10.3109/10409238309102790. [DOI] [PubMed] [Google Scholar]
- 37.Voet D., Voet J.G. Biochemistry. 4th. John Wiley & Sons; Hoboken, NJ, USA: 2011. pp. 951–954. Chapter 25. [Google Scholar]
- 38.Tulenko T.N., Boeze-Battaglia K., Mason R.P., Tint G.S., Steiner R.D., Connor W.E., Labelle E.F. A membrane defect in the pathogenesis of the Smith-Lemli-Opitz syndrome. J. Lipid Res. 2006;47:134–143. doi: 10.1194/jlr.M500306-JLR200. [DOI] [PubMed] [Google Scholar]
- 39.Song B.L., Javitt N.B., DeBose-Boyd R.A. Insig-mediated degradation of HMG CoA reductase stimulated by lanosterol, an intermediate in the synthesis of cholesterol. Cell Metab. 2005;1:179–189. doi: 10.1016/j.cmet.2005.01.001. [DOI] [PubMed] [Google Scholar]
- 40.Goldstein J.L., DeBose-Boyd R.A., Brown M.S. Protein sensors for membrane sterols. Cell. 2006;124:35–46. doi: 10.1016/j.cell.2005.12.022. [DOI] [PubMed] [Google Scholar]
- 41.Fitzky B.U., Moebius F.F., Asaoka H., Waage-Baudet H., Xu L., Xu G., Maeda N., Kluckman K., Hiller S., Yu H., et al. 7-Dehydrocholesterol-dependent proteolysis of HMG-CoA reductase suppresses sterol biosynthesis in a mouse model of Smith-Lemli-Opitz/RSH syndrome. J. Clin. Invest. 2001;108:905–915. doi: 10.1172/JCI12103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Byskov A.G., Andersen C.Y., Leonardsen L., Baltsen M. Meiosis activating sterols (MAS) and fertility in mammals and man. J. Exp. Zool. 1999;285:237–242. doi: 10.1002/(SICI)1097-010X(19991015)285:3<237::AID-JEZ6>3.0.CO;2-S. [DOI] [PubMed] [Google Scholar]
- 43.Vaknin K.M., Lazar S., Popliker M., Tsafriri A. Role of meiosis-activating sterols in rat oocyte maturation: effects of specific inhibitors and changes in the expression of lanosterol 14alpha-demethylase during the preovulatory period. Biol. Reprod. 2001;64:299–309. doi: 10.1095/biolreprod64.1.299. [DOI] [PubMed] [Google Scholar]
- 44.Grondahl C. Oocyte maturation. Basic and clinical aspects of in vitro maturation (IVM) with special emphasis of the role of FF-MAS. Dan. Med. Bull. 2008;55:1–16. [PubMed] [Google Scholar]
- 45.Keber R., Rozman D., Horvat S. Sterols in spermatogenesis and sperm maturation. J. Lipid Res. 2013;54:20–33. doi: 10.1194/jlr.R032326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Connor W.E., Lin D.S., Neuringer M. Biochemical markers for puberty in the monkey testis: desmosterol and docosahexaenoic acid. J. Clin. Endocrinol. Metab. 1997;82:1911–1916. doi: 10.1210/jc.82.6.1911. [DOI] [PubMed] [Google Scholar]
- 47.Acimovic J., Kosir R., Kastelec D., Perse M., Majdic G., Rozman D., Kosmelj K., Golicnik M. Circadian rhythm of cholesterol synthesis in mouse liver: A statistical analysis of the post-squalene metabolites in wild-type and Crem-knock-out mice. Biochem. Biophys. Res. Commun. 2011;408:635–641. doi: 10.1016/j.bbrc.2011.04.076. [DOI] [PubMed] [Google Scholar]
- 48.Ali Z., Heverin M., Olin M., Acimovic J., Lovgren-Sandblom A., Shafaati M., Bavner A., Meiner V., Leitersdorf E., Bjorkhem I. On the regulatory role of side-chain hydroxylated oxysterols in the brain. Lessons from CYP27A1 transgenic and cyp27a1−/− mice. J. Lipid Res. 2013;54:1033–1043. doi: 10.1194/jlr.M034124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Bjorkhem-Bergman L., Acimovic J., Torndal U.B., Parini P., Eriksson L.C. Lovastatin prevents carcinogenesis in a rat model for liver cancer. Effects of ubiquinone supplementation. Anticancer Res. 2010;30:1105–1112. [PubMed] [Google Scholar]
- 50.Tacer K.F., Haugen T.B., Baltsen M., Debeljak N., Rozman D. Tissue-specific transcriptional regulation of the cholesterol biosynthetic pathway leads to accumulation of testis meiosis-activating sterol (T-MAS) J. Lipid Res. 2002;43:82–89. [PubMed] [Google Scholar]
- 51.Xu F., Rychnovsky S.D., Belani J.D., Hobbs H.H., Cohen J.C., Rawson R.B. Dual roles for cholesterol in mammalian cells. Proc. Natl. Acad. Sci. USA. 2005;102:14551–14556. doi: 10.1073/pnas.0503590102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Rodriguez-Acebes S., de la Cueva P., Fernandez-Hernando C., Ferruelo A.J., Lasuncion M.A., Rawson R.B., Martinez-Botas J., Gomez-Coronado D. Desmosterol can replace cholesterol in sustaining cell proliferation and regulating the SREBP pathway in a sterol-Delta24-reductase-deficient cell line. Biochem. J. 2009;420:305–315. doi: 10.1042/BJ20081909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Flaxman N. Mer-29 (Triparanol) and Cataract. Med. Trial Tech. Q. 1963;10:11–32. [PubMed] [Google Scholar]
- 54.Kirby T.J. Cataracts produced by triparanol. (MER-29) Trans. Am. Ophthalmol. Soc. 1967;65:494–543. [PMC free article] [PubMed] [Google Scholar]
- 55.Perdriel G. Cataract and Triparanol. Bull. Soc. Ophtalmol. Fr. 1964;64:699–701. [PubMed] [Google Scholar]