

Abstract
Hinokinin is a lignan isolated from several plant species that has been recently investigated in order to establish its biological activities. So far, its cytotoxicity, its anti-inflammatory and antimicrobial activities have been studied. Particularly interesting is its notable anti-trypanosomal activity.
Keywords: cubebinolide, cytotoxicity, Trypanosoma, Chagas disease, antigenotoxic activity
1. Introduction
Lignans are important components of foods and medicines biosynthetically deriving from the radical coupling of two molecules of coniferyl alcohol at C-8/C-8′ positions (Figure 1). They are classified in different groups—dibenzylfuran, dihydroxybenzylbutane, dibenzylbutyrolactol, dibenzylbutyrolactone, aryltetraline lactone and arylnaphtalene derivatives—on the basis of the skeleton oxidation [1] and of the way in which oxygen is incorporated into the skeleton [2] (Figure 1). Podophyllotoxin and deoxypodophyllotoxin are, perhaps, the most important biologically active lignans, and their properties have been broadly reviewed [3,4].
Figure 1.

General classes of lignans.
In these last years, the biological activities of several lignans have been studied in depth [5,6,7] and among them hinokinin (1) is emerging as a new interesting compound. The aim of this review is to examine hinokinin (1) from a phytochemical and biological point of view. Peer-reviewed articles on hinokinin were acquired via the Scopus, SciFinder, and PubMed databases.
2. Phytochemistry
Hinokinin (1, Figure 2) was isolated for the first time by Yoshiki and Ishiguro in 1933 from the ether extract of hinoki wood (Chamecyparis obtusa Sieb. et Zucc.) as a colorless crystalline compound [8] and later Mameli, Briggs and Keimatsu established the identity of hinokinin with cubebinolide [9,10,11]. Haworth and Woodcock determined the trans configuration of the lactone ring by synthesis [12]. Biosynthesis of (−)-hinokinin was recently studied in Linum corymbulosum Reichenb by Bayindir et al. [13]. Starting from the observation that callus cultures of L. corymbulosum accumulate 1 [14], and according to the lignan composition found in Chamaecyparis obtusa by Takaku [15], the authors proposed two different pathways for the biosynthesis of hinokinin starting from (+)-pinoresinol (Scheme 1).
Figure 2.
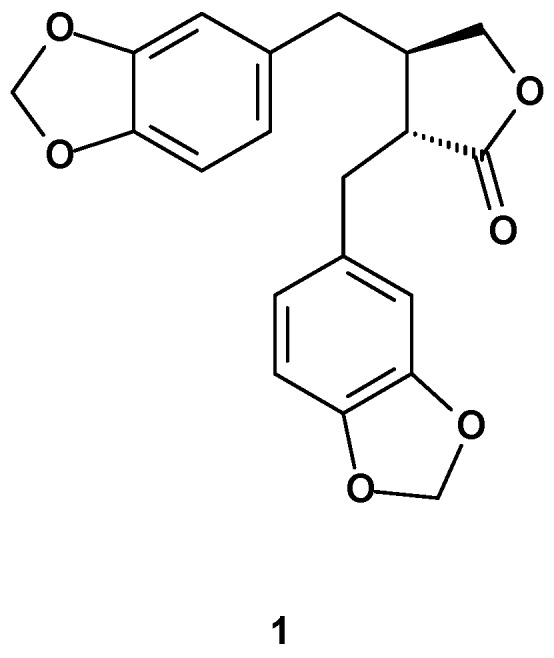
Hinokinin’s structure.
Scheme 1.

Proposed biosynthetic pathways for hinokinin (1). PS, pinoresinol synthase; PLR, pinoresino-lariciresinol reductase; SDH, secoisolariciresinol dehydrogenase; PLS, pluviatolide synthase; HS, hinokinin synthase; PSS, piperitol-sesamin synthase; SDR, sesamin-dihydrosesamin synthase; DDH, dihydrocubebin dehydrogenase [13].
In the first pathway, pinoresinol is reduced to secoisolariciresinol by a pinoresinol-lariciresinol reductase (PLR-Lc1), followed by the formation of the methylenedioxy bridges. In the second pathway, there is the formation of the methylenedioxy bridges on pinoresinol to give sesamin and the latter is then converted into dihydrocubebin and hinokinin. By the isolation of (PLR-Lc1), the enzyme responsible of the enantiospecific conversion of (+)-pinoresinol to (−)-secoisolariciresinol, they established that the first pathway is operative in hinokinin biosynthesis.
3. Distribution
After the first isolation from C. obtusa, hinokinin was isolated from C. formonensis [16] and from several other plants [17,18,19,20,21,22,23], such as for example Zanthoxylum simulans [24], Z. naranjillo [25], Z. lemairie [26], Z. monophyllum [27], Z. pistaciiflorum [28], Z. ailanthoides [29]. It was also found in different species of Phyllanthus [30,31,32], Aristolochia [33,34,35,36,37,38,39,40,41,42,43], Piper [44,45,46,47,48,49,50,51,52,53], Virola [54,55,56,57,58], Linum [59,60,61,62,63]. Another genus that produces hinokinin is Bursera. Compound 1 was found in B. cuneata and B. citronella by Koulman [64] and in B. simaruba by Maldini et al. [65].
4. Biological acitivities
4.1. Cytotoxic Activity
Hinokinin (1) was found to be a component of several cytotoxic extracts such as the petroleum ether fraction of a 75% ethanol extract of Zanthoxylum ailanthoides Sieb. & Zucc. stem bark [66]. The cytotoxicity of hinokinin (1) has been investigated by several authors against different cancer lines: P-388 (murine lymphocytic leukemia), HT-29 (human colon adenocarcinoma), A-549 (human lung adenocarcinoma) and MCF-7 (human breast adenocarcinoma) [18,67].
Ikeda et al. tested hinokinin (1) isolated from Anthriscus sylvestris [68] against B16F10 (murine metastatic melanoma), HeLa (human cervical cancer) and MK-1 (murine gastric adenocarcinoma) cell lines using the 3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyltetrazolium bromide (MTT)-microculture assay [69]. Results are reported in Table 1.
Table 1.
Cytotoxicity of hinokinin (1) against selected tumor cell lines a.
| P-388 | HT-29 | A-549 | MCF-7 | B16F10 | HeLa | MK-1 | |
|---|---|---|---|---|---|---|---|
| Hinokinin (1) | 1.54 b | 4.61 b | 8.01 b | 2.72 c | 2.58 c | 1.67 c | |
| 11.4 d | 26.1 d | 13.8 d | |||||
| 5.87 e | 3.52 e | 6.61 e | |||||
| Mithramycin | 0.08 b | 0.07 b | 0.06 b | ||||
| 0.06 e | 0.08 e | 0.07 e | |||||
| Adriamycin | 0.1 d | 0.02 d | 0.1 c | ||||
| Podophyllotoxin e | 0.001 | 0.0025 | 0.006 |
Hinokinin was also examined as antitumor promoter agent in a human cytomegalovirus (HCMV) immediate early (IE) antigen expression in human lung adenocarcinoma (A-549) cells [70]. After establishing the ID50 (dose causing 50% inhibition) in A-549 cell line (40.72 μg/mL), the authors found that 1 was able to reduce the IE antigen expression in HCMV-infected lung cancer cells in a dose-dependent manner (ID1: 81, ID10: 57% reduction, respectively).
According to Suffness and Pezzuto pure compounds are considered to have antitumor activity if they show ED50 values less than 4 μg/mL [71]. From this point of view, hinokinin (1) could be regarded as an antitumoral compound against P-388, HT-29, B16F10, HeLa and MK-1 cell lines. Mansoor et al. evaluated the apoptosis induction of hinokinin in human hepatoma HuH-7 cells [72]. Hinokinin significantly reduced viability of HuH-7 cells and it showed to be a strong inducer of apoptosis, inducing 2.4- and 2.5-fold increases in apoptotic cells as compared to controls. Furthermore, hinokinin was found to be highly toxic using the brine shrimp letality test (BST) [73].
Recently Awale et al. studied the cytotoxicity of several lignans isolated from W. indica, against Panc-1 cancer cell line (human pancreatic cancer) [74]. They found that (8S,8′S)-(+)-hinokinin as well as other lignans, such as (+)-arctigenin, with the same stereochemistry, were inactive against Panc-1 cell line, whereas the (−) enantiomers were cytotoxic. These results indicate that the absolute configuration of (−)-enantiomers is required for the cytotoxicity. Hinokinin resulted ineffective against HONE-1 (nasopharyngeal carcinoma) and UGC-3 (gastric adenocarcinoma) cell lines [75].
4.2. Anti-Inflammatory Activity
It is well known that inflammation is a key event in cancer development [76] and for this reason nowadays the anti-inflammatory activity of natural compounds is broadly studied. Hinokinin (1) was shown to be a potent inhibitory compound on human neutrophil superoxide generation and elastase release by neutrophils with an IC50 of 0.06 ± 0.12 μg/mL and an inhibitory percentage of 24.7 ± 6.2 at 10 μg/mL, respectively (diphenyleneiodonium: IC50 0.54 ± 0.21, phenylmethylsulfonyl fluoride: 35.24 ± 5.62% of inhibition) [77].
Furthermore, it was able to inhibit LPS-induced nitric oxide generation in RAW264.7 macrophages (IC50 21.56 ± 1.19 μM; aminoguanidine: 6.51 ± 1.15 μM) [78]. da Silva et al. studied the in vivo anti-inflammatory activity of hinokinin in the rat paw oedema reduction assay. Hinokinin (1) was shown to possess a good anti-oedema activity (in terms of efficacy) in a dose dependent manner (at the dose of 30 mg/kg it induced 63% of reduction, similar to indomethacin at the dose of 5 μg/mL) [79]. This anti-inflammatory activity was accompanied by an analgesic effect as demonstrated by the same authors in the acetic acid-induced writhing test in mice. Compound 1 produced high inhibition levels of the algogenic process (97%).
Immunosuppressive activity can play an important role in managing and resolving inflammation. Regarding the immunosuppressive activity of hinokinin (1), it has no activity against NFAT transcription factor [80], but it was found active in the lipopolysaccharide (LPS) induced cytokine production assay for IL-10, IL-12, and TNF-α [81] and remarkably active in a lymphocyte transformation assay [82] (Table 2). Recently, Desal et al. studied the anti-inflammatory effects of hinokinin against IL-6 and TNF-α, establishing that 1 exerts its anti-inflammatory effects via an NFκB-dependent mechanism [83].
Table 2.
Immunosuppressive activity of hinokinin (1).
| Cytokine Production Ratio a | LTI d | ||||
|---|---|---|---|---|---|
| TNF-α | IL-12 | IL-10 | IL-6c | ||
| Hinokinin (1) | 0.36 b | 0.44 b | 0.37 b | 25.94 ± 1.02 | |
| 77.5 c | 20.5 | ||||
| LPS b | 1 | 1 | 1 | ||
| Prednisolone b | 0.6 | 0.2 | 0.41 | ||
| Dexamethasone | 9.17 ± 0.53 | ||||
Lima et al. evaluated the anti-inflammatory and analgesic activities of bark crude dichloromethane extract (BCED) of Z. riedelianum [84]. They found that BCED was able to reduce carrageenan-induced rat paw oedema after 4 h at the dose 100 mg/Kg (% inhibition: 57.4; indometacin 43.2% at 10 mg/Kg). One of the components of the active extract was hinokinin. The authors suggested that the extract could display anti-inflammatory activity associated with COX inhibition. Moreover, BCED displayed a central analgesic activity too.
4.3. Anti-Parasitic Activities
4.3.1. Activity against Trypanosoma cruzi
Hinokinin (1) showed an interesting activity against Trypanosoma cruzi, the responsible of Chagas’ disease, a neglected protozoan disease that affects some 8 million people in Latin America [85,86]. Currently, there are only two effective drugs for Chagas’ disease treatment, namely nifurtimox and benznidazole (BZN), which both cause serious side effects, therefore, there is an urgent demand for the discovery of safer and more effective new therapeutic compounds. T. cruzi has a complex life cycle characterized by several developmental forms present in vertebrate and invertebrate hosts. This parasite exists in at least three morphologically distinct forms: infective (metacyclic or blood trypomastigotes), insect borne (epimastigotes) which replicate in the vector, and intracellular replicative (amastigotes) [87]. Hinokinin (1) in these last years has been studied as an interesting antitripanosomal compound [86]. In 2005 de Souza et al. testedhinokinin (1) in vitro against free amastigotes forms of Y strain of T. cruzi [88]. They found that 1 had an IC50 of 0.7 μM compared to BZN (IC50 0.8 μM) (Table 3).
Table 3.
In vitro anti-trypanosomal activity of hinokinin (1).a
| Free Amastigotes Y Strain b | Intracellular Amastigotes CL Strain c | Epimastigotes Forms of CL Strain c | % of Parasitaemia Reduction c | Trypomastigotes d | Intracellular Amastigotes d | |
|---|---|---|---|---|---|---|
| Hinokinin (1) | 0.7 | 18.36 | 0.67 | 70.8 | 94.49 | >141.24 |
| BZN | 0.8 | 20.00 | 30.89 | 29.0 | 146.02 | >190.83 |
In view of its anti-trypanosomal activity, hinokinin (1) was later selected to be assayed against epimastigote and intracellular amastigote forms of T. cruzi, both in vitro and in vivo assays [91] (see Table 3). In the in vivo assays obtained results showed that the treatment with hinokinin (1)promoted 70.8% of parasitaemia reduction in the parasitaemic peak, while benznidazole displayed approximately 29.0% of parasite reduction.
The antitrypanosomal activity of hinokinin was determined using the MTT assay by Sartorelli and coworkers [90]. They evaluated 1 against trypomastigotes and intracellular amastigotes of T. cruzi. Results are shown in Table 3. In order to study the toxicity of hinokinin (1) in mammalian cells, Sartorelli also studied hinokinin’s hemolytic activity and cytotoxicity. Hinokinin was shown to be effective on trypomastigotes, but it resulted toxic to mammalian cells and with a low parasite selectivity (selectivity index <1) [90].
To obtain better efficacy of this promising lead compound towards the intracellular forms of the parasite, Saraiva et al. prepared and investigated the effect of a new formulation using biodegradable polymers, such as poly(D,L-lactide-co-glycolic acid; PLGA), for the controlled release of hinokinin. The treatment of infected mice with hinokinin-loaded microparticles was able to provoke significant decrease in parasitemia levels compared with those observed in untreated controls [91]. Furthermore, Saraiva et al. showed that the administration of hinokinin-loaded microparticles was able to reduce the number of parasites more than hinokinin itself, in the course of the overall infection.
The reduction of tissue parasitism upon treatment with hinokinin (1), was evaluated in vivo by Esperandim and coworkers by quantifying the enzyme β-galactosidase expressed by the CLB5 clone strain of T. cruzi [92,93]. Treatment of mice infected with T. cruzi CLB5 with hinokinin (1) promoted significant reduction of tissue parasitism (liver, spleen and heart) compared with data recorded for untreated controls. Treatment with hinokinin (1) or benznidazole at a drug concentration of 50 mg/Kg a day, furnished a parasitism reduction of 50.5% or 41.7% in the liver; 71% or 16% in the spleen; and 41.4%, or 30.4% in the heart, respectively. The authors noted that there were some differences between the oral and intraperitoneal administration routes, being the former more effective for all evaluated organs, while BZN administered intraperitonealy was more effective for spleen and heart parasitism reduction [92]. Later, Esperandim evaluated in detail the in vivo therapeutic properties of oral administered hinokinin (1) against CLB5 strain of T. cruzi [93]. Hinokinin was assayed at concentration of 20 and 50 mg/kg. The authors observed that hinokinin at 20 mg/kg reduced the number of circulating forms at peak parasitemia of 51%, while at 50 mg/kg of 34.2%. The karyometry analysis once again showed a better behavior of 20 mg/kg dose (Table 4).
Table 4.
Karyometry analysis. Mean values of the nuclear area from cells of the spleen, liver, and heart of control groups and mice inoculated with the CL Brener clone strain of Trypanosoma cruzi B5.
| Groups | Area (μm2) | ||
|---|---|---|---|
| Spleen | Heart | Liver | |
| CINF a | 10.86 ± 2.45 | 18.20 ± 8.81 | 32.99 ± 7.78 |
| C b | 8.12 ± 2.04 | 15.05 ± 8.64 | 28.56 ± 5.69 |
| Hinokinin 20 c | 9.32 ± 2.22 | 17.48 ± 8.53 | 30.15 ± 7.90 |
| Hinokinin 50 d | 10.00 ± 2.68 | 18.56 ± 7.74 | 30.50 ± 7.49 |
| BZN 20 c | 9.69 ± 2.50 | 17.59 ± 7.08 | 29.46 ± 8.03 |
| BZN 50 d | 9.62 ± 2.37 | 20.42 ± 10.75 | 28.56 ± 6.45 |
a CINF: infected not treated animals; b C: control, uninfected animals; c Tested dose: 20 mg/kg; d Tested dose: 50 mg/kg.
The non-linear behavior between the two doses, with the 20 mg/Kg dose being more effective than the other, has been explained by an immunomodulatory response that hinokinin (1) can exert. It is well known that the immunosuppression of chronically infected patients can lead to disease reactivation, with high parasitemia and it has been already reported that hinokinin (1) can act as an immunosuppressive compound (see above).
4.3.2. Antiplasmodial Activity
Hinokinin was tested for its antiplasmodial activity against 3D7-chloroquine sensitive and Dd2-chloroquine resistant strains of Plasmodium falciparum. The IC50 of hinokinin (90.7 ± 1.4 μg/mL and 54.4 ± 8.5 μg/mL, respectively; chloroquine IC50 0.094 μg/mL) showed that 1 doesn’t possess significant antimalarial activity against either strain [94].
4.4. Antimicrobial Activity
Hinokinin (1) has been studied for its bioactivity against several other microorganisms. For example, Silva et al. examined the activity of this compound against oral pathogens such as Enterococcus faecalis, Candida albicans and several Streptococcus strains (see Table 5). It can be pointed out from data reported in Table 5 that, although chlorhexidine is much more active than hinokinin, 1 nevertheless showed a discrete antimicrobial activity [95]. Considering this antibacterial activity of hinokinin, Silva et al. evaluated the anti-mycobacterial activity of 1 and others lignans [96]. Hinokinin showed to be moderately active against M. tuberculosis, with a MIC value equal to 62.5 μg/mL and inactive against M. kansasii and M. avium (MIC 2000 μg/mL and 500 μg/mL, respectively).
Table 5.
Minimum inhibitory concentrations (MIC; mM) of hinokinin against oral pathogens.
| E. faecalis | S. salivarius | S. sanguinis | S. mitis | S. mutans | S. sobrinus | C. albicans | |
|---|---|---|---|---|---|---|---|
| Hinokinin (1) | 0.38 | 0.25 | 0.25 | 0.25 | 0.32 | 0.28 | 0.28 a |
| Chlorhexidine b | 5.9 | 1.7 | 3.9 | 5.9 | 5.9 | 1.5 | 7.9 |
a Fungicidal concentration; b MIC: μM.
4.5. Antiviral Activity
Several research groups studied the antiviral properties of hinokinin against human hepatitis B virus (HBV) [97], human immunodeficiency virus (HIV) [29], SARS-virus (SARS-CoV) [98], and in all cases 1 showed significant antiviral activity.
4.6. Genotoxic and Antigenotoxic Activities
In light of the interesting biological activities of hinokinin (1) and its potential use as therapeutic agent, it is important to investigate its mutagenic and genotoxic activities. Recently Resende et al. used the Ames and Comet assays, to assess the safety of using hinokinin as a drug [99]. In the Comet assay, on Chinese hamster lung fibroblasts (V79), hinokinin was shown to not be genotoxic. In the treatments with hinokinin associated with the known mutagen doxorubicin (DXR), the lower concentrations of 1 (0.5; 1.0 and 2.0 μM) significantly reduce DXR-induced DNA damage. The reduction in the DNA damage frequency ranged from 60.8% to 76.0% and it is not dose dependent.
Resende also showed that hinokinin has a protective effect in preventing clastogenic damage caused by methyl methanesulfonate (MMS), with the percent reduction ranging from 37.4% to 57.6% [100]. Mutagenic activity was evaluated by the Ames test, using the Salmonella typhimurium tester strains TA98, TA100, TA97a and TA102, using five different concentrations of hinokinin (9.75–78.0 μg/plate) selected on the basis of a preliminary toxicity test. The mutagenicity assays show that 1 did not induce any increase in the number of revertant colonies relative to the negative control, indicating the absence of any mutagenic activity.
Medola and coworkers studied the mutagenic and/or antimutagenic effects of hinokinin (1) in vivo using the rat peripheral blood micronucleus test. The differences of micronucleated cells between treated animals and control were not significative, demonstrating no genotoxic effect, while co-exposition of the animals to hinokinin and DXR showed a significant reduction in the frequencies of MNPCEs (micronucleated polychromatic erythrocytes). However, this protective effect of hinokinin was not dose dependent [101].
4.7. Target-Based Studies
Hinokinin (1) was tested for several other biological activities, such as antispasmodic effect on electrically induced (ECI), acetylcholine induced (AChI) and histamine induced contractions in isolated guinea-pig ileum, using the Ca2+ channel blocker verapamil as a positive control [102]. Hinokinin (1) significantly inhibited ECI and AChI contractions.
Neurite outgrowth-promoting activity in PC12 cells of hinokinin (1) isolated from C. obtusa in the presence or absence of Nerve Growth Factor (NGF, 2 ng/mL) was studied [103]. Hinokinin showed potent neurite outgrowth-promoting activities: 76.0% ± 6.0% at 10 μg/mL, and 50.9% ± 2.6% at 5 μg/mL when cultured with NGF, and 33.2% ± 5.4% at 10 μg/mL and 16.5% ± 2.6% at 5 μg/mL without NGF.
Nowadays, it is well established that neurons and glia development is regulated by neurotransmitters. Monoamine neurotransmitters such as dopamine, norepinephrine and serotonine have a positive action as classical growth factors, while glutamate and GABA (γ-aminobutyric acid) are down-regulating proliferation agents [104]. Hinokinin (1) showed neuroprotective activity against glutamate induced neurotoxicity in primary cultures of rat cortical cells (at 1.0 μM percentage of protection 42.6 ± 2.4, at 10.0 μM 56.9 ± 3.4; dizocipline maleate, a non-competitive antagonist of NMDA (N-methyl-d-aspartate) receptor (one of the glutamate receptors) showed at 1.0 μM 71.7 ± 1.2 and at 10.0 μM 77.4 ± 2.1 percentage of protection) [105]. Furthermore, Timple et al. demonstrated that hinokinin is a selective inhibitor of human dopamine and norepinephrine transporters in a noncompetitive manner with a low affinity for the serotonine transporter [106].
Cytochrome P450 (CYP) enzymes play an important role in phase I oxidation metabolism of a widw range of xenobiotics. In humans, 57 isoforms of CYP were identified, CYP3A4, CYP1A2, CYP2A6, CYP2D6, CYP2C8 and CYP2E1 among others.
Methylenedioxyphenyl compounds were well known to inhibit cytochrome P (CYP) reaction because they form stable complexes with CYP enzymes [107]. For this reason, several natural compounds incorporating this structural feature have been studied for their inhibitory activity of CYP enzymes. Hinokinin (1) containing two methylenedioxyphenyl rings in the molecules, showed potent CYP inhibition [108]. Later Usia et al. showed that hinokinin is active on CYP3A4 but not on CYP2D6 [109] and that CYP3A4 is inhibited in a time-, concentration- and NADPH-dependent manners via the formation of a metabolite intermediate complex [110], therefore, attention should be paid to a probable drug-drug interaction between hinokinin-containing preparations and molecules that are substrates of CYP3A4.
5. Conclusions
Lignans represent an important biologically active class of secondary metabolites. The most studied biological activities of these compounds are their antioxidant and anticancer properties. However, in recent years the importance of such metabolites, especially hinokinin, as potential antichagasic agents has been pointed out. In addition, hinokinin was shown to be non-genotoxic and to possess a neuroprotective effects. For all these reasons, hinokinin is emerging as a promising compound with broad and interesting biological activity.
Author Contributions
M.C.M. conceived the work. M.C.M. and A.P. collected and organized bibliographic data. M.C.M. and M.C. wrote the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 1.Moss G.P. Nomenclature of lignans and neolignans (IUPAC Recommendations 2000) Pure Appl.Chem. 2000;72:1493–1523. [Google Scholar]
- 2.Suzuki S., Umezawa T. Biosynthesis of lignans and norlignans. J. Wood Sci. 2007;53:273–284. [Google Scholar]
- 3.Khaled M., Jiang Z.Z., Zhang L.Y. Deoxypodophyllotoxin: A promising therapeutic agent from herbal medicine. J. Ethnopharmacol. 2013;149:24–34. doi: 10.1016/j.jep.2013.06.021. [DOI] [PubMed] [Google Scholar]
- 4.Liu Y.Q., Yang L., Tian X. Podophyllotoxin: current perspectives. Curr. Bioact. Compd. 2007;3:37–66. [Google Scholar]
- 5.Gordaliza M., Castro M.A., del Corral J.M., Feliciano A.S. Antitumor properties of podophyllotoxin and related compounds. Curr. Pharm. Des. 2000;6:1811–1839. doi: 10.2174/1381612003398582. [DOI] [PubMed] [Google Scholar]
- 6.Ríos J.L., Giner R.M., Prieto J.M. New Findings on the Bioactivity of Lignans. In: Atta ur R., editor. Studies in Natural Products Chemistry. Volume 26. Elsevier; Amsterdam, The Netherlands: 2002. pp. 183–292. [Google Scholar]
- 7.Cunha W.R., Andrade e Silva M.L., Sola Veneziani R.C., Ambrósio S.R., Kenupp Bastos J. Lignans: Chemical and Biological Properties. In: Rao V., editor. Phytochemicals-A Global Perspective of their Role in Nutrition and Health. InTech; Rijeka, Croatia: 2012. [Google Scholar]
- 8.Yoshiki Y., Ishiguro T. Crystalline constituents of hinoki oil. Yakugaku Zasshi. 1933;53:73–151. (in German 112–130) [Google Scholar]
- 9.Mameli E. Cubebin.VIII. Identity of cubebinolide with hinokinin. Gazz. Chim. Ital. 1935;65:886–888. [Google Scholar]
- 10.Keimatsu S., Ishiguro T. Constituents of hinokiol.II. A comparison of hinokinin and cubebinolide. Yakugaku Zasshi. 1935;55:96–99. [Google Scholar]
- 11.Briggs L.H. Hinokinin the enantiomorph of cubebinolide. J. Am. Chem. Soc. 1935;57:1383–1384. [Google Scholar]
- 12.Haworth R.D., Woodcock D. Constituents of natural phenolic resins. XIII. Synthesis of dl-, d- and l-hinokinin. J. Chem. Soc. 1938:1985–1989. doi: 10.1039/jr9380001985. [DOI] [Google Scholar]
- 13.Bayindir Ü., Alfermann A.W., Fuss E. Hinokinin biosynthesis in Linum corymbulosum Reichenb. Plant J. 2008;55:810–820. doi: 10.1111/j.1365-313X.2008.03558.x. [DOI] [PubMed] [Google Scholar]
- 14.Mohagheghzadeh A., Schmidt T.J., Bayindir R.N., Fuss E., Mehregan I., Alfermann A.W. Diarylbutyrolactone lignans from Linum corymbulosum in vitro cultures. Planta Med. 2006;72:1165–1167. doi: 10.1055/s-2006-947238. [DOI] [PubMed] [Google Scholar]
- 15.Takaku N., Okunishi T., Mikame K., Suzuki S., Sakakibara N., Umezawa T. Lignans from Chamaecyparis obtusa cv. Breviramea and cell suspension cultures of Daphne odora. Wood Res. 2001;88:44–45. [Google Scholar]
- 16.Lin T.C., Fang J.M., Cheng Y.S. Terpenes and lignans from leaves of Chamaecyparis formosensis. Phytochemistry. 1999;51:793–801. doi: 10.1016/S0031-9422(99)00074-6. [DOI] [Google Scholar]
- 17.Ingert N., Bombarda I., Herbette G., Faure R., Moretti C., Raharivelomanana P. Oleodaphnoic acid and coriaceol, two new natural products from the stem bark of Wikstroemia coriacea. Molecules. 2013;18:2988–2996. doi: 10.3390/molecules18032988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lin R.W., Tsai I.L., Duh C.Y., Lee K.H., Chen I.S. New lignans and cytotoxic constituents from Wikstroemia lanceolata. Planta Med. 2004;70:234–238. doi: 10.1055/s-2004-815540. [DOI] [PubMed] [Google Scholar]
- 19.Kato M., He Y.M., Dibwe D.F., Li F., Awale S., Kadota S., Tezuka T. New Guaian-type sesquiterpene from Wikstroemia indica. Nat. Prod. Commun. 2014;9:1–2. [PubMed] [Google Scholar]
- 20.Sousa de Lucena H.F., Madeiro S.A.L., Siqueira C.D., Filho J.M.B., de Fátima Agra M., da Silva M.S., Fechine Tavares J. Hypenol, a new lignan from Hypenia salzmannii. Helv. Chim. Acta. 2013;96:1121–1125. [Google Scholar]
- 21.Yamamoto S., Cox R.E., Simoneit B.R.T. Gas Chromatography/Mass Spectrometry of the lignans in resin of Callitris preissii. J. Mass Spectrom. Soc. Jpn. 2010;58:195–209. [Google Scholar]
- 22.Salmoun M., Braekman J.C., Ranarivelo Y., Rasamoelisendra R., Ralambomanana D., Dewelle J., Darro F., Kiss R. New calamenene sesquiterpenes from Tarenna madagascariensis. Nat. Prod. Res. 2007;21:111–120. doi: 10.1080/14786410600899084. [DOI] [PubMed] [Google Scholar]
- 23.Rios M.Y., Aguilar-Guadarrama A.B., Gutierrez Mdel C. Analgesic activity of affinin, an alkamide from Heliopsis longipes (Compositae) J. Ethnopharmacol. 2007;110:364–367. doi: 10.1016/j.jep.2006.09.041. [DOI] [PubMed] [Google Scholar]
- 24.Chen I.S., Wu S.J., Tsai I.L., Wu T.S., Pezzuto J.M., Lu M.C., Chai H., Suh N., Teng C.M. Chemical and bioactive constituents from Zanthoxylum simulans. J. Nat. Prod. 1994;57:1206–1211. doi: 10.1021/np50111a003. [DOI] [PubMed] [Google Scholar]
- 25.Bastos J.K., Albuquerque S., Silva M.L.A. Evaluation of the trypanocidal activity of lignans isolated from the leaves of Zanthoxylum naranjillo. Planta Med. 1999;65:541–544. doi: 10.1055/s-1999-14012. [DOI] [PubMed] [Google Scholar]
- 26.Adesina S.K., Olugbade T.A., Akinwusi D.D., Bergenthal D. Extractives from Zanthoxylum lemairie root and stem. Pharmazie. 1997;52:720–724. [Google Scholar]
- 27.Cuca S L.E., Martinez V J.C., Delle Monache F. Chemical constituents of Zanthoxylum monophyllum. Rev. Colomb. Quim. 1998;27:17–27. [Google Scholar]
- 28.Chen J.J., Huang H.Y., Duh C.Y., Chen I.S. Cytotoxic constituents from the stem bark of Zanthoxylum pistaciiflorum. J. Chin. Chem. Soc. 2004;51:659–663. [Google Scholar]
- 29.Cheng M.J., Lee K.H., Tsai I.L., Chen I.S. Two new sesquiterpenoids and anti-HIV principles from the root bark of Zanthoxylum ailanthoides. Bioorg. Med. Chem. 2005;13:5915–5920. doi: 10.1016/j.bmc.2005.07.050. [DOI] [PubMed] [Google Scholar]
- 30.Huang Y.L., Chen C.C., Ou J.C. Isolintetralin: A New Lignan from Phyllanthus niruri. Planta Med. 1992;58:473–474. doi: 10.1055/s-2006-961520. [DOI] [PubMed] [Google Scholar]
- 31.Huang Y.L., Chen C.C., Hsu F.L., Chen C.F. A New Lignan from Phyllanthus virgatus. J. Nat. Prod. 1996;59:520–521. doi: 10.1021/np970336v. [DOI] [PubMed] [Google Scholar]
- 32.Chang C.C., Lien Y.C., Liu K.C.S.C., Lee S.S. Lignans from Phyllanthus urinaria. Phytochemistry. 2003;63:825–833. doi: 10.1016/s0031-9422(03)00371-6. [DOI] [PubMed] [Google Scholar]
- 33.Achari B., Bandyopadhyay S., Saha C.R., Pakrashi S.C. A phenanthroid lactone, steroid and lignans from Aristolochia indica. Heterocycles. 1983;20:771–774. [Google Scholar]
- 34.Lopes L.M.X., Bolzani V.D.S., Trevusan L.M.V. Lignans from Brazilian Aristolochiaceae. Rev. Latinoam. Quim. 1988;19:113–117. [Google Scholar]
- 35.Leitào G.G., Kaplan M.A.C., Galeffi C. Epi-populifolic acid from Aristolochia cymbifera. Phytochemistry. 1992;31:3277–3279. [Google Scholar]
- 36.Kuo P.C., Li Y.C., Wu T.S. Chemical constituents and pharmacology of the Aristolochia species. J. Trad. Compl. Med. 2012;2:249–266. doi: 10.1016/s2225-4110(16)30111-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Marchesini A.M., Prado G.G., Messiano G.B., Machado M.B., Lopes L.M.X. Chemical constituents of Aristolochia giberti. J. Braz. Chem. Soc. 2009;20:1598–1608. [Google Scholar]
- 38.De Pascoli I.C., Nascimento I.R., Lopes L.M.X. Configurational analysis of cubebins and bicubebin from Aristolochia lagesiana and Aristolochia pubescens. Phytochemistry. 2006;67:735–742. doi: 10.1016/j.phytochem.2006.01.019. [DOI] [PubMed] [Google Scholar]
- 39.Shi L.S., Kuo P.C., Tsai Y.L., Damu A.G., Wu T.S. The alkaloids and other constituents from the root and stem of Aristolochia elegans. Bioorg. Med. Chem. 2004;12:439–446. doi: 10.1016/j.bmc.2003.10.031. [DOI] [PubMed] [Google Scholar]
- 40.Nascimento I.R., Lopes L.M. Diterpene esters of aristolochic acids from Aristolochia pubescens. Phytochemistry. 2003;63:953–957. doi: 10.1016/s0031-9422(03)00335-2. [DOI] [PubMed] [Google Scholar]
- 41.Navickiene H.M.D., Lopes L.M.X. Alkamides and phenethyl derivatives from Aristolochia gehrtii. J. Braz. Chem. Soc. 2001;12:467–472. [Google Scholar]
- 42.Wu T.S., Chan Y.Y., Leu Y.L. The constituents of the root and stem of Aristolochia cucurbitifolia Hayata and their biological activity. Chem. Pharm. Bull. 2000;48:1006–1009. doi: 10.1248/cpb.48.1006. [DOI] [PubMed] [Google Scholar]
- 43.Bomm M.D., Zukerman-Schpector J., Lopes L.M.X. Rearranged (4→2)-abeo-clerodane and clerodane diterpenes from Aristolochia chamissonis. Phytochemistry. 1999;50:455–461. [Google Scholar]
- 44.Koul S.K., Taneja S.C., Dhar K.L., Atal C.K. Lignans of Piper clusii. Phytochemistry. 1983;22:999–1000. [Google Scholar]
- 45.Elfahmi, Ruslan K., Batterman S., Bos R., Kayser O., Woerdenbag H.J., Quax W.J. Lignan profile of Piper cubeba, an Indonesian medicinal plant. Biochem. Syst. Ecol. 2007;35:397–402. [Google Scholar]
- 46.Parmar V.S., Jain S.C., Bisht K.S., Jain R., Taneja P., Jha A., Tyagi O.D., Prasad A.K., Wengel J., Olsen C.E., et al. Phytochemistry of the genus Piper. Phytochemistry. 1997;46:597–673. [Google Scholar]
- 47.Prabhu B.R., Mulchandani N.B. Lignans from Piper cubeba. Phytochemistry. 1985;24:329–331. [Google Scholar]
- 48.Koul S.K., Taneja S.C., Pushpangadan P., Dhar K.L. Lignans of Piper trichostachyon. Phytochemistry. 1988;27:1479–1482. [Google Scholar]
- 49.Sumathykutty M.A., Rao J.M. Lignans from leaves of Piper nigrum Linn. Indian J. Chem. Sect. B. 1988;27B:388–389. [Google Scholar]
- 50.Gangan V.D., Hussain S.S. Alkaloids from Piper hookeri: Revision of NMR assignments by the application of 2D NMR spectroscopy. J. Pharm. Res. 2011;4:4265–4267. [Google Scholar]
- 51.Bodiwala H., Singh G., Singh R., Dey C., Sharma S., Bhutani K., Singh I. Antileishmanial amides and lignans from Piper cubeba and Piper retrofractum. J. Nat. Med. 2007;61:418–421. [Google Scholar]
- 52.Chen Y.C., Liao C.H., Chen I.S. Lignans, an amide and anti-platelet activities from Piper philippinum. Phytochemistry. 2007;68:2101–2111. doi: 10.1016/j.phytochem.2007.05.003. [DOI] [PubMed] [Google Scholar]
- 53.Kijjoa A., Pinto M.M.M., Tantisewie B., Herz W. A new linalool derivative and other constituents from Piper ribesoides. Planta Med. 1989;55:193–194. doi: 10.1055/s-2006-961923. [DOI] [PubMed] [Google Scholar]
- 54.Cavalcante S.H., Yoshida M., Gottlieb O.R. The chemistry of Brazilian Myristicaceae.XXV. Neolignans from Virola carinata fruit. Phytochemistry. 1985;24:1051–1055. [Google Scholar]
- 55.Kato M.J., Yoshida M., Gottlieb O.R. The chemistry of Brazilian Myristicaceae. Part 34. Flavones and lignans in flowers, fruits and seedlings of Virola venosa. Phytochemistry. 1991;31:283–287. [Google Scholar]
- 56.Vidigal M.C.S., Cavalheiro A.J., Kato M.J., Yoshida M. Lignans from kernels of Virola michellii. Phytochemistry. 1995;40:1259–1261. [Google Scholar]
- 57.De Almeida Blumenthal E.E., Da Silva M.S., Yoshida M. Lignoids, flavonoids and polyketides of Virola surinamensis. Phytochemistry. 1997;46:745–749. [Google Scholar]
- 58.Nunomura S.M., Yoshida M. Lignans and benzoic acid derivatives from pericarps of Virola multinervia (Myristicaceae) Biochem. Syst. Ecol. 2002;30:985–987. [Google Scholar]
- 59.Koulman A., Konuklugil B. Lignan profile of Linum meletonis. Biochem. Syst. Ecol. 2004;32:91–93. [Google Scholar]
- 60.Schmidt T.J., Hemmati S., Fuss E., Alfermann A.W. A combined HPLC-UV and HPLC-MS method for the identification of lignans and its application to the lignans of Linum usitatissimum L. and L. bienne Mill. Phytochem. Anal. 2006;17:299–311. doi: 10.1002/pca.918. [DOI] [PubMed] [Google Scholar]
- 61.Schmidt T.J., Alfermann A.W., Fuss E. High-performance liquid chromatography/mass spectrometric identification of dibenzylbutyrolactone-type lignans: insights into electrospray ionization tandem mass spectrometric fragmentation of lign-7-eno-9,9'-lactones and application to the lignans of Linum usitatissimum L. (Common Flax) Rapid. Commun. Mass Spectrom. 2008;22:3642–3650. doi: 10.1002/rcm.3783. [DOI] [PubMed] [Google Scholar]
- 62.Meagher L.P., Beecher G.R., Flanagan V.P., Li B.W. Isolation and characterization of the lignans, isolariciresinol and pinoresinol, in flaxseed meal. J. Agric. Food Chem. 1999;47:3173–3180. doi: 10.1021/jf981359y. [DOI] [PubMed] [Google Scholar]
- 63.Schmidt T.J., Hemmati S., Klaes M., Konuklugil B., Mohagheghzadeh A., Ionkova I., Fuss E., Wilhelm Alfermann A. Lignans in flowering aerial parts of Linum species—Chemodiversity in the light of systematics and phylogeny. Phytochemistry. 2010;71:1714–1728. doi: 10.1016/j.phytochem.2010.06.015. [DOI] [PubMed] [Google Scholar]
- 64.Koulman A. Podophyllotoxin: A Study of the Biosynthesis, Evolution, Function and Use of Podophyllotoxin and Related Lignans; Ph.D. Thesis; 18 September 2003.University of Groningen, Groningen; [Google Scholar]
- 65.Maldini M., Montoro P., Piacente S., Pizza C. Phenolic compounds from Bursera simaruba Sarg. bark: Phytochemical investigation and quantitative analysis by tandem mass spectrometry. Phytochemistry. 2009;70:641–649. doi: 10.1016/j.phytochem.2009.02.009. [DOI] [PubMed] [Google Scholar]
- 66.Cao X.L., Xu J., Bai G., Zhang H., Liu Y., Xiang J.F., Tang Y.L. Isolation of anti-tumor compounds from the stem bark of Zanthoxylum ailanthoides Sieb. & Zucc. by silica gel column and counter-current chromatography. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2013;929:6–10. doi: 10.1016/j.jchromb.2013.04.006. [DOI] [PubMed] [Google Scholar]
- 67.Chang S.T., Wang D.S.Y., Wu C.L., Shiah S.G., Kuo Y.H., Chang C.J. Cytotoxicity of extractives from Taiwania cryptomerioides heartwood. Phytochemistry. 2000;55:227–232. doi: 10.1016/s0031-9422(00)00275-2. [DOI] [PubMed] [Google Scholar]
- 68.Ikeda R., Nagao T., Okabe H., Nakano Y., Matsunaga H., Katano M., Mori M. Antiproliferative constituents in Umbelliferae plants.IV. Constituents in the fruits of Anthriscus sylvestris Hoffm. Chem. Pharm. Bull. 1998;46:875–878. doi: 10.1248/cpb.46.875. [DOI] [PubMed] [Google Scholar]
- 69.Mosmann T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J. Immunol. Met. 1983;65:55–63. doi: 10.1016/0022-1759(83)90303-4. [DOI] [PubMed] [Google Scholar]
- 70.Pusztai R., Abrantes M., Sherly J., Duarte N., Molnar J., Ferreira M.J.U. Antitumor-promoting activity of lignans: Inhibition of human cytomegalovirus IE gene expression. Anticancer Res. 2010;30:451–454. [PubMed] [Google Scholar]
- 71.Stuffness M.S., Pezzuto J.M. Assays related to cancer drug discovery. In: Hostettmann K., editor. Methods in Plant Biochemistry. Volume 6. Academic Press Ltd.; London, UK: 1991. Assays for bioactivity. [Google Scholar]
- 72.Mansoor T.A., Ramalho R.M., Rodrigues C.M., Ferreira M.J. Dibenzylbutane- and butyrolactone-type lignans as apoptosis inducers in human hepatoma HuH-7 cells. Phytother. Res. 2012;26:692–696. doi: 10.1002/ptr.3629. [DOI] [PubMed] [Google Scholar]
- 73.Huang J.M., Nakade K., Kondo M., Yang C.S., Fukuyama Y. Brine shrimp lethality test active constituents and new highly oxygenated seco-prezizaane-type sesquiterpenes from Illicium merrillianum. Chem. Pharm. Bull. 2002;50:133–136. doi: 10.1248/cpb.50.133. [DOI] [PubMed] [Google Scholar]
- 74.Awale S., Kato M., Dibwe D.F., Li F., Miyoshi C., Esumi H., Kadota S., Tezuka Y. Antiausterity activity of arctigenin enantiomers: importance of (2R,3R)-absolute configuration. Nat. Prod. Commun. 2014;9:79–82. [PubMed] [Google Scholar]
- 75.Cheng M.J., Lee S.J., Chang Y.Y., Wu S.H., Tsai I.L., Jayaprakasam B., Chen I.S. Chemical and cytotoxic constituents from Peperomia sui. Phytochemistry. 2003;63:603–608. doi: 10.1016/s0031-9422(03)00183-3. [DOI] [PubMed] [Google Scholar]
- 76.Elinav E., Nowarski R., Thaiss C.A., Hu B., Jin C., Flavell R.A. Inflammation-induced cancer: Crosstalk between tumours, immune cells and microorganisms. Nat. Rev. Cancer. 2013;13:759–771. doi: 10.1038/nrc3611. [DOI] [PubMed] [Google Scholar]
- 77.Chen J.J., Chung C.Y., Hwang T.L., Chen J.F. Amides and benzenoids from Zanthoxylum ailanthoides with inhibitory activity on superoxide generation and elastase release by neutrophils. J. Nat. Prod. 2009;72:107–111. doi: 10.1021/np800689b. [DOI] [PubMed] [Google Scholar]
- 78.Lee D.Y., Seo K.H., Jeong R.H., Lee S.M., Kim G.S., Noh H.J., Kim S.Y., Kim G.W., Kim J.Y., Baek N.I. Anti-inflammatory lignans from the fruits of Acanthopanax sessiliflorus. Molecules. 2013;18:41–49. doi: 10.3390/molecules18010041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Da Silva R., de Souza G.H.B., da Silva A.A., de Souza V.A., Pereira A.C., Royo V.D. A., e Silva M.L.A., Donate P.M., de Matos Araujo A.L.S., Carvalho J.C.T., et al. Synthesis and biological activity evaluation of lignan lactones derived from (−)-cubebin. Bioorg. Med. Chem. Lett. 2005;15:1033–1037. doi: 10.1016/j.bmcl.2004.12.035. [DOI] [PubMed] [Google Scholar]
- 80.Cai X.F., Lee I.S., Dat N.T., Shen G., Kang J.S., Kim D.H., Kim Y.H. Inhibitory lignans against NFAT transcription factor from Acanthopanax koreanum. Arch. Pharmacal Res. 2004;27:738–741. doi: 10.1007/BF02980142. [DOI] [PubMed] [Google Scholar]
- 81.Ramos F., Takaishi Y., Kawazoe K., Osorio C., Duque C., Acuna R., Fujimoto Y., Sato M., Okamoto M., Oshikawa T., et al. Immunosuppressive diacetylenes, ceramides and cerebrosides from Hydrocotyle leucocephala. Phytochemistry. 2006;67:1143–1150. doi: 10.1016/j.phytochem.2006.03.004. [DOI] [PubMed] [Google Scholar]
- 82.Zhang W., Yao Z., Zhang Y.W., Zhang X.X., Takaishi Y., Duan H.Q. Immunosuppressive sesquiterpenes from Buddleja daviddi. Planta Med. 2010;76:1882–1887. doi: 10.1055/s-0030-1249974. [DOI] [PubMed] [Google Scholar]
- 83.Desai D.C., Jacob J., Almeida A., Kshirsagar R., Manju S.L. Isolation, structural elucidation and anti-inflammatory activity of astragalin, (−)-hinokinin, aristolactam I and aristolochic acids (I & II) from Aristolochia indica. Nat. Prod. Res. 2014:1–5. doi: 10.1080/14786419.2014.905563. [DOI] [PubMed] [Google Scholar]
- 84.Lima L.M., Perazzo F.F., Carvalho J.C.T., Bastos J.K. Anti-inflammatory and analgesic activities of the ethanolic extracts from Zanthoxylum riedelianum (Rutaceae) leaves and stem bark. J. Pharm. Pharmacol. 2007;59:1151–1158. doi: 10.1211/jpp.59.8.0014. [DOI] [PubMed] [Google Scholar]
- 85.Rassi A., Jr., Rassi A., Marin-Neto J.A. Chagas disease. Lancet. 2010;375:1388–1402. doi: 10.1016/S0140-6736(10)60061-X. [DOI] [PubMed] [Google Scholar]
- 86.Schmidt J.T., Khalid A.S., Romanha J.A., Alves M.T., Biavatti W.M., Brun R., Da Costa B.F., de Castro L.S., Ferreira F.V., de Lacerda V.G.M., et al. The potential of secondary metabolites from plants as drugs or leads against protozoan neglected diseases—Part II. Curr. Med. Chem. 2012;19:2176–2228. [PubMed] [Google Scholar]
- 87.Burleigh B.A., Andrews N.W. A 120-kDa alkaline peptidase from Trypanosoma cruzi is involved in the generation of a novel Ca2+-signaling factor for mammalian cells. J. Biol. Chem. 1995;270:5172–5180. doi: 10.1074/jbc.270.10.5172. [DOI] [PubMed] [Google Scholar]
- 88.De Souza V.A., da Silva R., Pereira A.C., Royo Vde A., Saraiva J., Montanheiro M., de Souza G.H., da Silva Filho A.A., Grando M.D., Donate P.M., et al. Trypanocidal activity of (−)-cubebin derivatives against free amastigote forms of Trypanosoma cruzi . Bioorg. Med. Chem. Lett. 2005;15:303–307. doi: 10.1016/j.bmcl.2004.10.079. [DOI] [PubMed] [Google Scholar]
- 89.Saraiva J., Vega C., Rolon M., da S.R., Andrade E.S.M.L., Donate P.M., Bastos J.K., Gomez-Barrio A., de A.S. In vitro and in vivo activity of lignan lactones derivatives against Trypanosoma cruzi. Parasitol. Res. 2007;100:791–795. doi: 10.1007/s00436-006-0327-4. [DOI] [PubMed] [Google Scholar]
- 90.Sartorelli P., Carvalho C.S., Reimao J.Q., Lorenzi H., Tempone A.G. Antitrypanosomal activity of a diterpene and lignans isolated from Aristolochia cymbifera. Planta Med. 2010;76:1454–1456. doi: 10.1055/s-0029-1240952. [DOI] [PubMed] [Google Scholar]
- 91.Saraiva J., Lira A.A.M., Esperandim V.R., da S.F.D., Ferraudo A.S., Bastos J.K., Andrade E.S.M.L., de G.C.M., de A.S., Marchetti J.M. Hinokinin-loaded poly(d,l-lactide-co-glycolide) microparticles for Chagas disease. Parasitol. Res. 2010;106:703–708. doi: 10.1007/s00436-010-1725-1. [DOI] [PubMed] [Google Scholar]
- 92.Esperandim V.R., da S.F.D., Saraiva J., Silva M.L.A., Costa E.S., Pereira A.C., Bastos J.K., de A.S. Reduction of parasitism tissue by treatment of mice chronically infected with Trypanosoma cruzi with lignano lactones. Parasitol. Res. 2010;107:525–530. doi: 10.1007/s00436-010-1885-z. [DOI] [PubMed] [Google Scholar]
- 93.Esperandim V.R., da Silva Ferreira D., Rezende K.C., Cunha W.R., Saraiva J., Bastos J.K., e Silva M.L., de Albuquerque S. Evaluation of the in vivo therapeutic properties of (−)-cubebin and (−)-hinokinin against Trypanosoma cruzi. Experim. Parasitol. 2013;133:442–446. doi: 10.1016/j.exppara.2012.12.005. [DOI] [PubMed] [Google Scholar]
- 94.Abrantes M., Mil-Homens T., Duarte N., Lopes D., Cravo P., Madureira M.D.C., Ferreira M.J.U. Antiplasmodial activity of lignans and extracts from Pycnanthus angolensis. Planta Med. 2008;74:1408–1412. doi: 10.1055/s-2008-1081317. [DOI] [PubMed] [Google Scholar]
- 95.Silva M.L.A., Coimbra H.S., Pereira A.C., Almeida V.A., Lima T.C., Costa E.S., Vinholis A.H.C., Royo V.A., Silva R., Filho A.A.S., et al. Evaluation of Piper cubeba extract, (−)-cubebin and its semi-synthetic derivatives against oral pathogens. Phytoth. Res. 2007;21:420–422. doi: 10.1002/ptr.2088. [DOI] [PubMed] [Google Scholar]
- 96.Silva M.L.A., Martins C.H.G., Lucarini R., Sato D.N., Pavanb F.R., Freitas N.H.A., Andrade L.N., Pereira A.C., Bianco T.N.C., Vinholis A.H.C., et al. Antimycobacterial activity of natural and semi-synthetic lignans. Z. Naturforsch. C. 2009;64:779–784. doi: 10.1515/znc-2009-11-1204. [DOI] [PubMed] [Google Scholar]
- 97.Huang R.L., Huang Y.L., Ou J.C., Chen C.C., Hsu F.L., Chang C. Screening of 25 compounds isolated from Phyllanthus species for anti-human Hepatitis B virus in vitro. Phytother. Res. 2003;17:449–453. doi: 10.1002/ptr.1167. [DOI] [PubMed] [Google Scholar]
- 98.Wen C.C., Kuo Y.H., Jan J.T., Liang P.H., Wang S.Y., Liu H.G., Lee C.K., Chang S.T., Kuo C.J., Lee S.S., et al. Specific plant terpenoids and lignoids possess potent antiviral activities against severe acute respiratory syndrome coronavirus. J. Med. Chem. 2007;50:4087–4095. doi: 10.1021/jm070295s. [DOI] [PubMed] [Google Scholar]
- 99.Resende F.A., Barbosa L.C., Tavares D.C., de Camargo M.S., de Souza Rezende K.C., e Silva M.L., Varanda E.A. Mutagenicity and antimutagenicity of (−)-hinokinin a trypanosomicidalcompound measured by Salmonella microsome and comet assays. BMC Complement. Altern. Med. 2012;12:203. doi: 10.1186/1472-6882-12-203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Resende F.A., Tomazella I.M., Barbosa L.C., Ponce M., Furtado R.A., Pereira A.C., Bastos J.K., Andrade e Silva, M.L., Tavares D.C. Effect of the dibenzylbutyrolactone lignan (−)-hinokinin on doxorubicin and methyl methanesulfonate clastogenicity in V79 Chinese hamster lung fibroblasts. Mutat. Res. 2010;700:62–66. doi: 10.1016/j.mrgentox.2010.04.023. [DOI] [PubMed] [Google Scholar]
- 101.Medola J.F., Cintra V.P., Pesqueira e Silva É.P.C., de Andrade Royo V., da Silva R., Saraiva J., Albuquerque S., Bastos J.K., e Silva M.L., Tavares D.C. (−)-Hinokinin causes antigenotoxicity but not genotoxicity in peripheral blood of Wistar rats. Food Chem. Toxicol. 2007;45:638–642. doi: 10.1016/j.fct.2006.10.012. [DOI] [PubMed] [Google Scholar]
- 102.Zhang G., Shimokawa S., Mochizuki M., Kumamoto T., Nakanishi W., Watanabe T., Ishikawa T., Matsumoto K., Tashima K., Horie S., et al. Chemical constituents of Aristolochia constricta: Antispasmodic effects of its constituents in guinea-pig ileum and isolation of a diterpeno-lignan hybrid. J. Nat. Prod. 2008;71:1167–1172. doi: 10.1021/np800041t. [DOI] [PubMed] [Google Scholar]
- 103.Kuroyanagi M., Ikeda R., Gao H.Y., Muto N., Otaki K., Sano T., Kawahara N., Nakane T. Neurite outgrowth-promoting active constituents of the Japanese cypress (Chamaecyparis obtusa) Chem. Pharm. Bull. 2008;56:60–63. doi: 10.1248/cpb.56.60. [DOI] [PubMed] [Google Scholar]
- 104.Cameron H.A., Hazel T.G., McKay R.D. Regulation of neurogenesis by growth factors and neurotransmitters. J. Neurobiol. 1998;36:287–306. [PubMed] [Google Scholar]
- 105.Yoon J.S., Koo K.A., Ma C.J., Sung S.H., Kim Y.C. Neuroprotective lignans from Biota orientalis leaves. Nat. Prod. Sci. 2008;14:167–170. [Google Scholar]
- 106.Timple J.M., Magalhaes L.G., Souza Rezende K.C., Pereira A.C., Cunha W.R., e Silva M.L., Mortensen O.V., Fontana A.C. The lignan (−)-hinokinin displays modulatory effects on human monoamine and GABA transporter activities. J. Nat. Prod. 2013;76:1889–1895. doi: 10.1021/np400452n. [DOI] [PubMed] [Google Scholar]
- 107.Wilkinson C.F., Murray M., Marcus C.B. Interactions of methylenedioxyphenyl compounds with cytochrome P-450 and effects on microsomal oxidation. Rev. Biochem. Toxicol. 1984;6:27–63. [Google Scholar]
- 108.Tsukamoto S., Tomise K., Miyakawa K., Cha B.C., Abe T., Hamada T., Hirota H., Ohta T. CYP3A4 Inhibitory Activity of New Bisalkaloids, Dipiperamides D and E, and Cognates from White Pepper. Bioorg. Med. Chem. 2002;10:2981–2985. doi: 10.1016/s0968-0896(02)00130-x. [DOI] [PubMed] [Google Scholar]
- 109.Usia T., Watabe T., Kadota S., Tezuka Y. Potent CYP3A4 inhibitory constituents of Piper cubeba. J. Nat. Prod. 2005;68:64–68. doi: 10.1021/np0401765. [DOI] [PubMed] [Google Scholar]
- 110.Usia T., Watabe T., Kadota S., Tezuka Y. Metabolite-cytochrome P450 complex formation by methylenedioxyphenyl lignans of Piper cubeba: Mechanism-based inhibition. Life Sci. 2005;76:2381–2391. doi: 10.1016/j.lfs.2004.12.005. [DOI] [PubMed] [Google Scholar]