

Abstract
In the history of human medicine, antibiotics represent epochal examples of medical progress. However, with an approaching antibiotic crisis due to the emergence and extensive spread of antimicrobial resistance among bacterial agents, as well as to increasing number of patients with chronic and recalcitrant bacterial biofilm-associated infections, the naturally occurring molecules may become new sources of antibacterial and antibiofilm drugs for clinical usage. Polyphenols represent a class of plant natural products which are important in plant defense against microbial pathogens. The main focus of the review is on the antibiofilm activities of phenolic compounds against bacteria which play an essential role in medical device biofilm-associated infections. The other, not negligible part of the review is devoted to polyphenols’ activity against bacterial agents that cause dental caries and periodontal disease.
Keywords: plant polyphenols, antibacterial activity, bacterial biofilm, medical device-associated infection, dental caries, periodontal disease
1. Introduction
The introduction of antibiotics to clinical practice represents one of the most outstanding contributions to the treatment of life-threatening infectious diseases. However, due to the extensive use of these valuable therapeutics, numerous resistance mechanisms have emerged and rapidly spread among bacterial disease-causative agents. Modern medicine is nowadays facing the threat of returning to the pre-antibiotic era, at least for some types of infectious diseases. The rapid spread of multidrug-resistant (MDR) or extremely drug-resistant (XDR) bacterial strains seems to be the most frightening development. Along with some community strains of Mycobacterium tuberculosis, Streptococcus pneumoniae, or Neisseria gonorrhoeae, these MDR and XDR bacterial strains are represented mainly by nosocomial opportunistic bacterial infectious agents, such as methicillin resistant staphylococci (Staphylococcus aureus and coagulase-negative staphylococci), vancomycin-resistant enterococci, and the Gram-negative XDR strains. Great concern has arisen due to the appearance and spread of MDR and XDR strains of Pseudomonas aeruginosa, Acinetobacter spp., and carbapenem-resistant Enterobacteriaceae (mostly contributed by Klebsiella pneumoniae) [1,2].
In addition the abovementioned resistance caused by mutations or resistance gene acquisition, the second large challenge is represented by the phenomenon of recalcitrant infections in patients with bacteria or fungi growing in biofilms on implanted or inserted medical devices, or in the tissue damaged by various prior pathological processes [3]. A distinct subset of such infections is represented by dental caries and periodontal diseases, caused by members of the normal oral microbiota [4,5].
A biofilm is a sessile form of bacterial existence on solid surfaces or air-liquid interfaces, in which bacteria multiply covered by a self-produced biofilm matrix, composed of bacterial intercellular polysaccharides, proteins, and extracellularly released nucleic acids [6]. The protective effect of bacterial biofilm phenotypes is multifactorial. It includes decreased penetration of antimicrobial agents into the deep layers of biofilms, the capture of positively charged molecules by the extracellular polymeric biofilm matrix, or the ability of biofilm matrices to concentrate bacterial enzymes which inactivate antibiotics [7]. Gradients of nutrients, metabolites, oxygen, pH, redox potential, or antibiotics penetrating to the biofilm produce an environmental stress in the bacteria, resulting in the expression of inducible resistance mechanisms, increased mutability rate, and bacterial adaptive phenotype changes. These changes lead to metabolic suppression in bacteria, which cause increased ability to survive exposure to antibiotics and an increasing rate of persister cell formation. Bacterial persisters survive antimicrobial therapy and may reseed the patient’s infectious focus after discontinuation of antimicrobial therapy, resulting in disease relapse [7,8]. Mechanisms of bacterial adherence, biofilm accumulation, and bacterial dispersion from the mature biofilm, coordinated by quorum-sensing (QS) chemical signals of inter-bacterial communication, reflecting the bacterial population cell density play a vitally important role in the process of biofilm development [9].
Therefore, together with the steps taken to reduce the threat of an antibiotic resistance crisis and the search for new antimicrobial agents [10], research on agents able to modulate some important virulence factors of bacteria, such as adhesivity, biofilm formation and the phenomenon of bacterial persistence, has an exceptional importance for the development of new therapeutics for medical practice.
Many reports on antibacterial activity associated with extracts from an enormous range of plants can be found in the literature. The discovery of novel antibacterial agents in plant extracts most frequently begins with leaves or roots from healthy specimens, even though there is ample evidence that many key components of plant defenses against phytopathogens are induced by infection. Plants respond to microbial attack through a highly coordinated repertoire of molecular, cellular and tissue-based defensive barriers to colonisation and invasion [11,12,13]. Plant secondary metabolites, among them many polyphenols, such as flavonoids, phenolic acids, and tannins, show antibacterial and/or antibiofilm activities. This review focuses on the antibiofilm activities of the abovementioned plant products.
2. Main Antibiofilm Phenolic Compounds
Plant polyphenols represent a large class of biologically active secondary metabolites of plants. They include flavonoids, tannins, anthocyanins, phenolic acids, stilbenes, coumarins, lignans, and lignins [14]. These substances play an important role in resistance against various microbial pathogens and protect against free radicals and toxins [15,16]. Nowadays, plant polyphenols enjoy an ever-increasing recognition not only by the scientific community but also, and most remarkably, by the general public because of their presence and abundance in fruits, seeds, vegetables, and derived foodstuffs and beverages, whose regular consumption has been claimed to be beneficial for human health. They have often been highlighted due to their capacity to scavenge oxidatively generated free radicals that underlies their utility in reducing the risk of certain age-related degenerative processes and diseases [16].
In phenolics, multiple mechanisms of antibacterial activity have been described: they interact with bacterial proteins and cell wall structures, they may cause damage to cytoplasmic membranes, reduce membrane fluidity, inhibit nucleic acid synthesis, cell wall synthesis, or energy metabolism [15,17,18]. On the other hand, antibiofilm activity research on plant phenolics has revealed, besides their destructive activity on bacteria, also “softer” activities leading to biofilm suppression by affecting the bacterial regulatory mechanisms such as quorum sensing or other global regulator systems, without an effect on bacterial growth [19]. A survey of recently published antibiofilm activities of flavonoids, phenolic acids and tannins is provided in Table 1.
Table 1.
Antibiofilm activities of flavonoids, phenolic acids and tannins contained in plant extracts.
| Phenolic Compound | Name of Bacteria | Antibiofilm Activity * | References |
|---|---|---|---|
| ANTHOCYANINS | |||
| Malvidin, Petunidin, Cyanidin | K. pneumoniae | a,b | [20] |
| COUMARINS | |||
| Coumarin | E. coli | a | [21,22] |
| S. aureus | a | ||
| V. anguillarum | a | ||
| E. tarda | a | ||
| Umbelliferone | E. coli | c | [21] |
| Esculetin | S. aureus | a | [23,24] |
| P. aeruginosa | a | ||
| Esculin | P. aeruginosa | a | [23] |
| Psoralen | P. aeruginosa | a | [23] |
| Nodakenetin | P. aeruginosa | a | [25] |
| Coladonin | E. coli | c | [21] |
| FLAVONOIDS | |||
| Chalcone | S. mutans | d | [26] |
| 2′,4′-Dihydroxychalcone | S. aureus | a | [27] |
| 2,2′,4′-Trihydroxychalcone | S. aureus | a | [27] |
| 2′,4′-Dihydroxy-2-methoxychalcone | S. aureus | a | [27] |
| Xanthohumol | S. aureus | a | [28] |
| Naringenin | E. coli | a | [29] |
| Hesperidin | E. coli | a | [29] |
| Neohesperidin | E. coli | a | [29] |
| V. harvey | a | ||
| Neoeriocitrin | E. coli | a | [29] |
| V. harvey | a | ||
| 8-Prenylnaringenin | S. aureus | a | [27,28] |
| Apigenin | E. coli | a | [27,30,31,32] |
| S. aureus | a | ||
| V. harvey | a | ||
| S. mutans | a | ||
| Fisetin | S. aureus | a | [24] |
| Chrysin | E. coli | a | [30,31] |
| S. aureus | a | ||
| Luteolin | E. coli (UPEC) | a | [30,33] |
| S. aureus | a | ||
| Nobiletin | E. coli | a, e | [34] |
| Sinensitin | E. coli | a, e | [29,34] |
| V. harvey | a, e | ||
| Quercitrin | S. mutans | a | [35] |
| Quercetin | E. coli | a | [29,30,36,37] |
| S. aureus | a | ||
| V. harvey | a | ||
| S. mutans | a | ||
| Kaempferol | E. coli | a | [29,30,36] |
| S. aureus | a | ||
| V. harvey | a | ||
| Morin | L. monocytogenes | f | [38] |
| Phloretin | E. coli | g | [31] |
| Rutin | E. coli, V. harvey | a | [29] |
| Daidzein | E. coli (UPEC) | a | [31] |
| Genistein | S. aureus | a | [31,39] |
| E. coli (UPEC) | a | ||
| TANNINS | |||
| Catechin | P. aeruginosa | a | [40] |
| Gallic acid | E. coli | a | [41,42] |
| S. mutans | a | ||
| Methyl gallate | S. aureus | h | [41,43] |
| S. mutans | a | ||
| (−)-Epigallocatechin gallate | S. aureus | a | [44,45,46,47] |
| S. epidermidis | a | ||
| S. mutans | a | ||
| P. gingivalis | a | ||
| E. faecalis | a | ||
| Ellagic acid | E. coli | a | [48] |
| Tannic acid | E. coli | a | [48] |
| Rosmarinic acid | S. aureus | a | [49,50] |
| 1,2,3,4,6-Penta-O-galloyl-b-d-glucopyranose | S. aureus | a | [51] |
* a—inhibited biofilm formation; b—inhibited EPS production; c—reductions in biofilm formation; d—sortase-specific oral biofilm inhibition; e—inhibited motility; f—in vitro and in vivo antibiofilm efficacy; g—reduced pathogenic biofilm; no harm to commensal E. coli K-12 biofilm formation; h—inactivated bacteria in biofilm.
To the most extensively studied bacteria from the point of view of biofilm production belong Staphylococcus aureus and coagulase-negative staphylococci, which play a crucial role in medical device-associated infections [52], and one of the most important dental caries agents—Streptococcus mutans [4].
Tannins represent one of the biggest groups of plant polyphenols. They are subclassified into condensed tannins (proanthocyanidins or catechins) and hydrolysable tannins (gallotannins and ellagitannins) [53]. Gallotannins and ellagitannins derived from the metabolism of the shikimate-derived gallic acid (3,4,5-trihydroxybenzoic acid) result through various esterification and phenolic oxidative coupling reactions in yield numerous monomeric, oligomeric and polyphenolic galloyl ester derivatives of sugar, mainly d-glucose [54].
Tannins possess antibacterial activity both against Gram-positive and Gram-negative bacteria. For example, the catechins are able to penetrate and interact with lipid bilayers [55]. Alternatively, they may cause membrane fusion, a process that results in leakage of intramembranous materials and aggregation [56]. Green tea (Camellia sinensis) rich in catechins has the capacity to reverse methicillin resistance in MRSA isolates at concentrations much lower than those needed to produce inhibition of bacterial growth [57]. Roccaro et al. referred to the modulation effect of catechin gallates to bacterial drug resistance. It has been shown that epigallocatechin gallate (EGCg) (Figure 1) had several antibacterial activities, limiting bacterial growth and invasion and acting in synergy with some antibiotics. Sub-inhibitory concentrations of EGCg were able to reverse tetracycline resistance in staphylococci by inhibition of the Tet(K) efflux pump, in addition to further sensitizing of the susceptible staphylococcal isolates to this antibiotic [58]. Concerning S. aureus biofilm formation, EGCg at subinhibitory concentrations has shown to decrease slime production, therefore inhibiting biofilm formation by this bacterial species [44].
Figure 1.

Chemical structure of (−)-epigallocatechin gallate.
Tannic acid from black tea (Camellia sinensis) inhibited S. aureus biofilm formation without inhibiting bacterial growth via a mechanism dependent upon the putative transglycosylase IsaA, and this acid also inhibited pharyngeal colonization with S. aureus in an in vivo rodent model [59]. Extract of Alnus japonica, with quercetin and tannic acid as the major anti-S. aureus biofilm compounds, was the most active from 498 screened plant extracts. It inhibited biofilm formation by influencing the expression of genes linked to biofilm production, most markedly icaA and icaD [60].
S. aureus antibiofilm activity was described in several phenolic acids, including gallic [61], ellagic [62], ginkgolic [63] and rosmarinic acid [49] at subinhibitory concentrations.
Recent research on Cotinus coggygria leaves rich in gallotannins such as gallic acid and methyl gallate (Figure 2), showed a good activity against S. aureus in planktonic and biofilm growth forms. The 60% methanol extract showed bactericidal activity against all tested S. aureus strains, including polyresistant strains, and eradicated bacteria in already established 24-h biofilm [43].
Figure 2.

Chemical structures of gallic acid (left) and methyl gallate (right).
Ellagic acid and its derivatives from Rubus ulmifolius can limit S. aureus biofilm formation to a degree that can be correlated with increased antibiotic susceptibility [64]. Ellagic acid (Figure 3) and tannic acid were also tested for their ability to inhibit biofilm formation by Escherichia coli. Both compounds reduced biofilm formation significantly. However, no synergistic effect of these two compounds was observed [48]. Methanol extract of pomegranate, rich in ellagic acid, was also shown to inhibit the formation of biofilms of S. aureus, methicillin resistant S. aureus and E. coli as a result of possible damage to the cell membrane [62].
Figure 3.
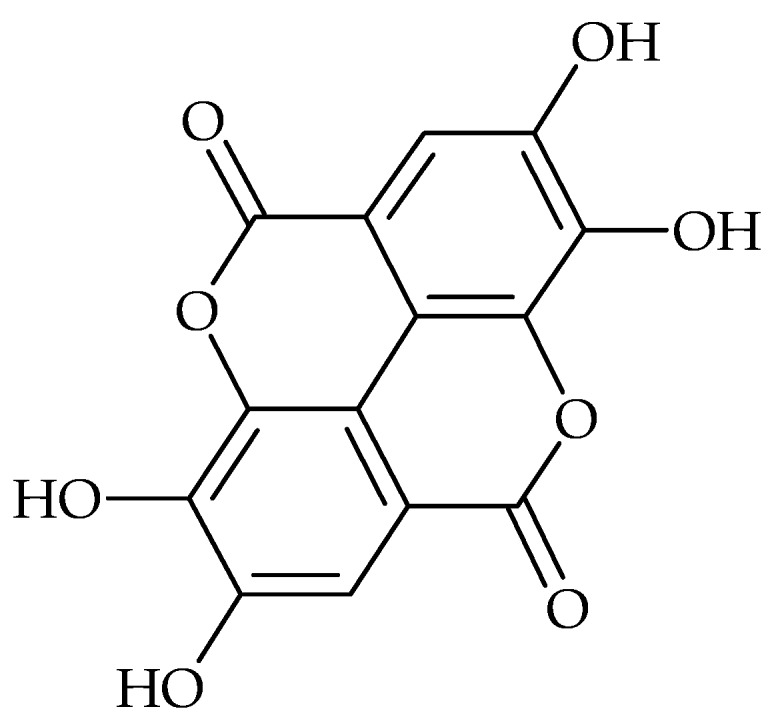
Chemical structure of ellagic acid.
Inhibition of biofilm formation on surfaces covered by plant products may be significant in the future techniques which prevent medical device biofilm-associated infections. Such activity was described in several studies on the antibiofilm activity of plant polyphenols.
Tannic acid (syn. gallotanin) from Eustigma oblongifolium inhibited biofilm formation by S. aureus independently of growth mechanisms. It prevented the initial attachment to solid surfaces and the synthesis of polysaccharide intercellular adhesion compounds. The antibiofilm activity of gallotanin was expressed after application in solution, as well as after coating of the tested surfaces [51]. Similar effect had medical device implant surface coating with hamamelitannin (2′,5-di-O-galloyl-hamamelose, Figure 4) isolated from the bark and leaves of Hamamelis virginiana. Medical device-associated infection in rat model was completely prevented, when sterile collagen-sealed double velour-knitted polyethylene terephthalate (Dacron) grafts were coated with hamamelitannin. No activity of hamamelitanin on bacterial growth was observed and the antibiofilm activity was attributed to the staphylococcal quorum-sensing regulator RNAIII inhibition [65].
Figure 4.

Chemical structure of hamamelitannin.
Trentin et al. reported that B-type linked proanthocyanidin-coated surfaces reduced S. aureus and E. faecalis adhesion. The proposed mechanism of bacterial attachment inhibition is based on electrostatic repulsion, high hydrophilicity and the steric hindrance provided by the coating that blocks bacterium-substratum interactions [66].
Rosmarinic acid (Figure 5), also known as Lamiaceae tanning compound, was identified as a major phenolic compound in many antimicrobially active plants, e.g., in the genera Mentha, Melissa, Lycopus, Origanum, Thymus, Salvia [67]. According to the latest research, rosmarinic acid could be a candidate topical antimicrobial agent with killing activity on planktonic forms of clinical S. aureus strains and suppressing activity in the early stages of biofilm development [49,50]. At subinhibitory concentrations near to MIC this compound suppressed S. aureus biofilm production; however, with further decreases of the rosmarinic acid concentration an increase of biofilm production was observed, which reached its peak at 100-times lower concentrations than MIC [49]. A similar phenomenon of concentration-dependent response in biofilm production was observed in the case of many other antimicrobial agents, as an expression of bacterial stress response modulated by low concentrations of chemical compounds, such as ethanol or antibiotics [68,69,70]. Therefore, the phenomenon described above should be tested and considered when determining the therapeutic concentrations of the potential drugs of plant origin, as underdosing might have counterproductive effects on biofilm-related infections.
Figure 5.

Chemical structure of rosmarinic acid.
Flavonoids are widely distributed phenolics characterized by a phenylbenzopyran chemical structure. In plants, flavonoids have long been known to be synthesized in specific sites and are responsible for the colour and aroma of flowers and fruits to attract pollinators, and consequently fruit dispersion animals; they help in seeding, germination, growth and development of seedlings. Flavonoids protect plants from different biotic and abiotic stresses and act as unique UV-filters. Flavonoids have roles against frost hardiness, drought resistance and may play a functional role in plant heat acclimation and freezing tolerance. They function as signal molecules, allelopathic compounds, phytoalexins, detoxifying agents, and antimicrobial defensive compounds [54,71]. Three different modes of antibacterial activity of flavonoids were described in the literature. The first corresponds to nucleic acid synthesis inhibition [72]. The second way involves damage of the cytoplasmic membrane by a perforation mechanism [73] and a decrease in membrane fluidity [74], and the third, the inhibition of energy metabolism [75]. Flavonoids also exhibit antibiofilm activities. Red wine (from Vitis vinifera) contains, besides tannic acid and trans-resveratrol, plenty of flavonoids such as quercetin, fisetin, kaempeferol, apigenin, chrysin, luteolin (Figure 6) and their derivatives. These red wine compounds were found to be effective in the inhibition of S. aureus biofilm formation, where quercetin was remarkably the most active flavonoid [30]. The seeds of muscadine grape (Vitis rotundifolia) are rich in gallic acid, (+)-catechin and epicatechin, while the skin contains ellagic acid, myricetin, quercetin, kaempferol, and trans-resveratrol. [76]. Polyphenol extract from muscadine grape pomace had antibacterial activity against S. aureus, and at subinhibitory concentrations inhibited its biofilm formation, and at 16 × MIC it eradicated biofilms [77].
Figure 6.

Chemical structures of red wine components: quercetin, fisetin, kaempeferol, apigenin, chrysin and luteolin.
According to the results of a study by Vikram et al. flavonoids found in citrus fruit can modulate bacterial cell–cell communication, E. coli O157:H7 biofilm formation and V. harveyi virulence. Naringenin, quercetin, sinensetin and apigenin were the most active. Among the tested flavonoids, naringenin emerged as potent and possibly nonspecific inhibitor of autoinducer-mediated cell–cell signalling [29].
Sivaranjani et al. explored the in vitro and in vivo antibiofilm efficacy of the flavonol morin (Figure 7) against Listeria monocytogenes, one of the leading foodborne pathogens. They found that morin not only inhibited biofilm production, but also reduced the virulence of L. monocytogenes [38]. Chalcone derivatives can also inhibit biofilm formation. This activity has been demonstrated in 2′,4′-dihydroxychalcone, 2,2′,4′-trihydroxychalcone and 2′,4′-dihydroxy-2-methoxychalcone, which inhibit S. aureus biofilm production [27]. Phloretin (Figure 7), an apple flavonoid, inhibited E. coli O157:H7 biofilm formation without inhibiting the growth of planktonic cells [31].
Figure 7.

Chemical structures of morin (left) and phloretin (right).
Xantohumol (Figure 8), a prenylated chalconoid from Humulus lupulus was found to inhibit S. aureus adhesion and biofilm formation. It also inactivated bacteria in already formed biofilm, most likely by damaging the stability of the bacterial cytoplasmic membrane after inhibition of lipid metabolism [28].
Figure 8.

Chemical structure of xanthohumol.
Naturally-occurring coumarins, derivatives of 5,6-benzo-2-pyrone, display several biological activities, from photosensitizing, vasodilatating, or analgesic properties to excellent anti-inflammatory and antimicrobial activities [78]. Lee et al. examined the antibiofilm abilities of different coumarins, such as coumarin (Figure 9), coumarin-3-carboxylic acid, esculetin, 4-hydroxycoumarin, scopoletin, umbelliferone (Figure 9) and coladonin. They reported that coumarin and umbelliferone exhibited antibiofilm formation activity against enterohaemorrhagic E. coli O157:H7 without inhibiting planktonic cell growth. Furthermore, the biofilm of E. coli was inhibited by coladonin [21]. Inhibition of biofilm formation of P. aeruginosa was detected for esculetin, esculin, psoralen and nodakenetin [23,24,25].
Figure 9.

Chemical structures of coumarin (left) and umbelliferone (right).
Plants polyphenols could be found also in honeys of floral origin, as a result of their natural production. The well characterised Manuka honey contains mainly flavonoids and phenolic acids (for a review see [79]). Their content is closely related to the antioxidant and antimicrobial activity of honey, and they probably cooperate with the other biologically active compounds on the reported antibiofilm activity of Manuka honey [80].
3. Polyphenols in Periodontal Diseases and Caries
Dental biofilm ecological shift contributes to oral diseases affecting a large proportion of the human population [5]. Streptococcus mutans is a bacterium participating at the development of caries, thanks to its acidogenicity, aciduric properties, and an outstanding ability to produce biofilms [81,82]. Periodontal diseases, which are the major cause of tooth loss in humans, are chiefly associated with two anaerobic bacteria—Prevotella spp. and Porphyromonas gingivalis [55].
Numerous studies contain reports on polyphenols’ inhibitory effects on oral biofilm bacteria and on dental biofilm production and accumulation. Many catechin-based polyphenols, flavonoids, proanthocyanidin oligomers and some other plant-derived compounds inhibit S. mutans glycosyltransferase—one of the crucial virulence factors of S. mutans with role in synthesis of glucan polysaccharide, a major biofilm matrix component [83].
Tea polyphenols, especially EGCg, inhibited biofilm formation by S. mutans and reduced viability of bacteria in preformed biofilm. At subinhibitory concentrations EGCg inhibited the acidogenic and aciduric properties of this bacterium, probably by inhibition of the enzymatic activity of F1Fo-ATPase and lactate dehydrogenase, and expressed inhibition of sucrose-dependent initial attachment of S. mutans to surfaces [45,84]. EGCg, derived from green tea, was active also against one of the important periodontal disease agents and destroyed already established P. gingivalis biofilms [46] and completely inhibited the growth and adherence of P. gingivalis onto the buccal epithelial cells [85]. Lee and Tan observed a similar effect of EGCg also against biofilms (and other virulence factors) of Enterococcus faecalis [47], an agent of chronic and refractory dental canal infections [86].
Apigenin showed inhibitory activity to both glucosyltransferase and fructosyltransferase of S. mutans without major impact on bacterial viability and influenced the biomass and polysaccharide content of S. mutans biofilm [32]. Quercitrin inhibited S. mutans biofilm production by reducing the synthesis of both water-soluble and insoluble glucans and several virulence genes suppression [35].
S. mutans saccharide metabolism inhibition by several phenolic acids was detected as well. Gallic acid (and methyl gallate) had inhibitory effects on the growth of cariogenic and periodontopathic bacteria and significantly inhibited the in vitro formation of S. mutans biofilms [41]. Gallic acid and tannic acid at subinhibitory concentrations showed suppressive effect on S. mutans biofilm formation by inhibition of glucosyltransferase and fructosyltransferase [87]. However, the effect of gallic acid on biofilm formation was affected by nutrient levels, temperature, and treatment time [42].
Oligomeric proanthocyanidins, the major secondary metabolites of Vaccinium macrocarpon (cranberry), are further potential anticaries agents that inhibit the production of organic acids and the formation of biofilms by cariogenic bacteria [88]. Cranberry proanthocyanidins, comprised of mostly A-type oligomers of epicatechin, and flavonols (mostly quercetin glycosides) inhibited the activities of glucosyltransferases and F-ATPase, and the acid production by S. mutans cells. Biofilm development and acidogenicity were significantly affected by their topical application [89]. Topical applications of cranberry proanthocyanidins during biofilm formation resulted in less biomass and fewer insoluble polysaccharide formation by S. mutans in vitro and a significant reduction of caries incidence and less severe carious lesions in a rat dental caries model. A-type proanthocyanidin dimers and oligomers effectively diminished the synthesis of insoluble polysaccharides, and also affected bacterial glycolysis [90,91].
Several studies have evaluated the activity of grape, grape wine, grape pomace or grape seeds polyphenol extracts on biofilms produced by oral bacteria. Red wine grape (Vitis vinifera and Vitis interspecies hybrids) and its pomace phenolic extracts remarkably inhibited glucosyltransferase of S. mutans, as well as the glycolytic pH drop without affecting the bacterial viability, even if the anthocyanins and flavan-3-ols content were highly variable [92]. In two studies published by Furiga et al., polyphenols from red wine, grape pomace and grape seed inhibited both the formation of multi-species biofilms composed of oral bacteria (S. mutans, Streptococcus sobrinus, Lactobacillus rhamnosus, Actinomyces viscosus, Porphyromonas gingivalis, and Fusobacterium nucleatum), and the synthesis of insoluble glucan. The most effective was the grape seed extract, containing mainly catechin and epicatechin. Except to a significant antiplaque activity, the extract had synergistic effect with amine fluoride mouthwash, and showed also an important antioxidant capacity in vitro, without any bactericidal effects [93,94]. Antibacterial effect of red wine polyphenols on bacteria in the 5-species biofilm model consisting of Actinomyces oris, F. nucleatum, Streptococcus oralis, S. mutans and Veillonella dispar was detected by Muñoz-Gonzales et al. [95]. The powdered extract of phenolics from the pomace of Japanese wild grape (Vitis coignetiae) with high phenolics and flavanol content reduced adhesion of S. mutans to saliva-coated hydroxyapatite and biofilm formation in a dose-dependent manner, and inhibited water-soluble and water-insoluble glucans synthesis [96].
4. Conclusions
This review is an overview of research articles about antibiofilm activity of selected plant phenolics listed in scientific databases such as SciFinder, Science Direct, PubMed, Scopus, Web of Science, etc. Numerous plant phenolic compounds have already revealed their antimicrobial and antibiofilm activities, but the road to a clinical application form may still be long. It requires further testing—besides antimicrobial and antibiofilm effectivity—the toxicity, pharmacokinetics, pharmacodynamics, drug interactions, including classical antibiotics, and any kind of side-effects should be defined. The most feasible seems to be approval of topical application forms, which are much safer in the case of drugs with higher toxicity, and allow higher, but still safe dosages in comparison with systemic antibiotic therapy, so in the form of solutions, lotions, ointments, tinctures, gels, creams, lozenges, or suppositories, the active phenolics may come relatively soon into the practice. Inhibition of bacterial adherence to skin, mucosal and dental surfaces facilitated by topical application may also have beneficial effects in the prevention of infectious diseases, dental caries and periodontal disease. Plant phenolics-covered medical device surfaces may help in prevention of device-associated biofilm infections.
Acknowledgments
This work was supported by the Grant Agency of Ministry of Education, Science, Research, and Sport of the Slovak Republic (grant No. VEGA-1/0290/16 Comprehensive analysis of natural products with therapeutic potential in human medicine) and the Slovak Research and Development Agency (under the contract No. APVV-15-0123). The authors would also like to thank to Viera Žufková for English emendation.
Abbreviations
| MDR | Multidrug-resistant |
| XDR | Extremely drug-resistant |
| QS | Quorum sensing |
| EGCg | Epigallocatechin gallate |
| EPS | Extracellular polymeric substance |
| UPEC | Uropathogenic Escherichia coli |
Author Contributions
Silvia Fialová, Lívia Slobodníková, Katarína Rendeková and Ján Kováč participated in the study design. Silvia Fialová and Pavel Mučaji provided financial support. The manuscript was written by Silvia Fialová, Lívia Slobodníková and Katarína Rendeková. All authors read and approved the final version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 1.WHO (World Health Organization): Antimicrobial Resistance: Global Report on Surveillance. 2014. [(accessed on 2 November 2016)]. Available online: http://apps.who.int/iris/bitstream/10665/112642/1/9789241564748_eng.pdf.
- 2.Rossolini G.M., Arena F., Pecile P., Pollini S. Update on the antibiotic resistance crisis. Curr. Opin. Pharmacol. 2014;18:56–60. doi: 10.1016/j.coph.2014.09.006. [DOI] [PubMed] [Google Scholar]
- 3.Høiby N., Bjarnsholt T., Moser C., Bassi G.L., Coenye T., Donelli G., Hall-Stoodley L., Holá V., Imbert C., Kirketerp-Møller K., et al. ESCMID Study Group for Biofilms and Consulting External Expert Werner Zimmerli. ESCMID guideline for the diagnosis and treatment of biofilm infections 2014. Clin. Microbiol. Infect. 2015;21:S1–S25. doi: 10.1016/j.cmi.2014.10.024. [DOI] [PubMed] [Google Scholar]
- 4.Do T., Devine D., Marsh P.D. Oral biofilms: Molecular analysis, challenges, and future prospects in dental diagnostics. Clin. Cosmet. Investig. Dent. 2013;5:11–19. doi: 10.2147/CCIDE.S31005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Takahashi N., Nyvad B. The role of bacteria in the caries process: Ecological perspectives. J. Dent. Res. 2011;90:294–303. doi: 10.1177/0022034510379602. [DOI] [PubMed] [Google Scholar]
- 6.Donlan R.M., Costerton J.V. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002;15:167–193. doi: 10.1128/CMR.15.2.167-193.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Høiby N., Bjarnsholt T., Givskov M., Molin S., Ciofu O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents. 2010;35:322–332. doi: 10.1016/j.ijantimicag.2009.12.011. [DOI] [PubMed] [Google Scholar]
- 8.Maisonneuve E., Gerdes K. Molecular mechanisms underlying bacterial persisters. Cell. 2014;157:539–548. doi: 10.1016/j.cell.2014.02.050. [DOI] [PubMed] [Google Scholar]
- 9.Rutherford S.T., Bassler B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2012;2:124–127. doi: 10.1101/cshperspect.a012427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ventola C.L. The antibiotic resistance crisis: Part 2: Management strategies and new agents. Pharm. Ther. 2015;40:344–352. [PMC free article] [PubMed] [Google Scholar]
- 11.Taylor P.W. Alternative natural sources for a new generation of antibacterial agents. Int. J. Antimicrob. Agents. 2013;42:195–201. doi: 10.1016/j.ijantimicag.2013.05.004. [DOI] [PubMed] [Google Scholar]
- 12.Guest D.I., Brown J.F. Plant defences against pathogens. In: Brown J.F., Ogle J.H., editors. Plant Pathogens and Plant Diseases. Rockvale Publications; Armidale, Australia: 1997. pp. 263–286. [Google Scholar]
- 13.Freeman B.C., Beattie G.A. An overview of plant defenses against pathogens and herbivores. Plant Health Instr. 2008 doi: 10.1094/PHI-I-2008-0226-01. [DOI] [Google Scholar]
- 14.Pereira D.M., Valentão P., Pereira J.A., Andrade P.B. Phenolics: From chemistry to biology. Molecules. 2009;14:2202–2211. doi: 10.3390/molecules14062202. [DOI] [Google Scholar]
- 15.Daglia M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012;23:174–181. doi: 10.1016/j.copbio.2011.08.007. [DOI] [PubMed] [Google Scholar]
- 16.Quideau S., Deffieux D., Douat-Casassus C., Pouysegu L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chem. Int. Ed. Engl. 2011;50:586–621. doi: 10.1002/anie.201000044. [DOI] [PubMed] [Google Scholar]
- 17.Cushnie T.P., Lamb A.J. Recent advances in understanding the antibacterial properties of flavonoids. Int. J. Antimicrob. Agents. 2011;38:99–107. doi: 10.1016/j.ijantimicag.2011.02.014. [DOI] [PubMed] [Google Scholar]
- 18.Gyawali R., Ibrahim S.A. Natural products as antimicrobial agents. Food Control. 2014;46:412–429. doi: 10.1016/j.foodcont.2014.05.047. [DOI] [Google Scholar]
- 19.Silva L.N., Zimmer K.R., Macedo A.J., Trentin D.S. Plant natural products targeting bacterial virulence factors. Chem. Rev. 2016;116:9162–9236. doi: 10.1021/acs.chemrev.6b00184. [DOI] [PubMed] [Google Scholar]
- 20.Gopu V., Kothandapani S., Shetty P.H. Quorum quenching activity of Syzygium cumini (L.) Skeels and its anthocyanin malvidin against Klebsiella pneumoniae. Microb. Pathog. 2015;79:61–69. doi: 10.1016/j.micpath.2015.01.010. [DOI] [PubMed] [Google Scholar]
- 21.Lee J.H., Kim Y.G., Cho H.S., Ryu S.Y., Cho M.H., Lee J. Coumarins reduce biofilm formation and the virulence of Escherichia coli O157:H7. Phytomedicine. 2014;21:1037–1042. doi: 10.1016/j.phymed.2014.04.008. [DOI] [PubMed] [Google Scholar]
- 22.Gutierrez-Barranquero J.A., Reen F.J., McCarthy R.R., O’Gara F. Deciphering the role of coumarin as a novel quorum sensing inhibitor suppressing virulence phenotypes in bacterial pathogens. Appl. Microbiol. Biotechnol. 2015;99:3303–3316. doi: 10.1007/s00253-015-6436-1. [DOI] [PubMed] [Google Scholar]
- 23.Zeng Z., Qian L., Cao L., Tan H., Huang Y., Xue X., Shen Y., Zhou S. Virtual screening for novel quorum sensing inhibitors to eradicate biofilm formation of Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2008;79:119–126. doi: 10.1007/s00253-008-1406-5. [DOI] [PubMed] [Google Scholar]
- 24.Durig A., Kouskoumvekaki L., Vejborg R.M., Klemm P. Chemoinformatics-assisted development of new anti-biofilm compounds. Appl. Microbiol. Biotechnol. 2010;87:309–317. doi: 10.1007/s00253-010-2471-0. [DOI] [PubMed] [Google Scholar]
- 25.Ding X., Yin B., Qian L., Zeng Z., Yang Z., Li H., Lu Y., Zhou S. Screening for novel quorum-sensing inhibitors to interfere with the formation of Pseudomonas aeruginosa biofilm. J. Med. Microbiol. 2011;60:1827–1834. doi: 10.1099/jmm.0.024166-0. [DOI] [PubMed] [Google Scholar]
- 26.Wallock-Richards D.J., Marles-Wright J., Clarke D.J., Maitra A., Dodds M., Hanley B., Campopiano D.J. Molecular basis of Streptococcus mutans sortase A inhibition by the flavonoid natural product trans-chalcone. Chem. Commun. 2015;51:10483–10485. doi: 10.1039/C5CC01816A. [DOI] [PubMed] [Google Scholar]
- 27.Manner S., Skogman M., Goeres D., Vuorela P., Fallarero A. Systematic exploration of natural and synthetic flavonoids for the inhibition of Staphylococcus aureus biofilms. Int. J. Mol. Sci. 2013;14:19434–19451. doi: 10.3390/ijms141019434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Rozalski M., Micota B., Sadowska B., Stochmal A., Jedrejek D., Wieckowska-Szakiel M., Rozalska B. Antiadherent and antibiofilm activity of Humulus lupulus L. derived products: New pharmacological properties. BioMed Res. Int. 2013;2013:101089. doi: 10.1155/2013/101089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Vikram A., Jayaprakasha G.K., Jesudhasan P.R., Pillai S.D., Patil B.S. Suppression of bacterial cell-cell signalling, biofilm formation and type III secretion system by citrus flavonoids. J. Appl. Microbiol. 2010;109:515–527. doi: 10.1111/j.1365-2672.2010.04677.x. [DOI] [PubMed] [Google Scholar]
- 30.Cho H.S., Lee J.H., Cho M.H., Lee J. Red wines and flavonoids diminish Staphylococcus aureus virulence with anti-biofilm and anti-hemolytic activities. Biofouling. 2015;31:1–11. doi: 10.1080/08927014.2014.991319. [DOI] [PubMed] [Google Scholar]
- 31.Lee J.H., Regmi S.C., Kim J.A., Cho M.H., Yun H., Lee C.S., Lee J. Apple flavonoid phloretin inhibits Escherichia coli O157:H7 biofilm formation and ameliorates colon inflammation in rats. Infect. Immun. 2011;79:4819–4827. doi: 10.1128/IAI.05580-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Koo H., Hayacibara M.F., Schobel B.D., Cury J.A., Rosalen P.L., Park Y.K., Vacca-Smith A.M., Bowen W.H. Inhibition of Streptococcus mutans biofilm accumulation and polysaccharide production by apigenin and tt-farnesol. J. Antimicrob. Chemother. 2003;52:782–789. doi: 10.1093/jac/dkg449. [DOI] [PubMed] [Google Scholar]
- 33.Shen X.F., Ren L.B., Teng Y., Zheng S., Yang X.L., Guo X.J., Wang X.Y., Sha K.H., Li N., Xu G.Y., et al. Luteolin decreases the attachment, invasion and cytotoxicity of UPEC in bladder epithelial cells and inhibits UPEC biofilm formation. Food Chem. Toxicol. 2014;72:204–211. doi: 10.1016/j.fct.2014.07.019. [DOI] [PubMed] [Google Scholar]
- 34.Vikram A., Jayaprakasha G.K., Uckoo R.M., Patil B.S. Inhibition of Escherichia coli O157:H7 motility and biofilm by β-sitosterol glucoside. Biochim. Biophys. Acta Gen. Subj. 2013;1830:5219–5228. doi: 10.1016/j.bbagen.2013.07.022. [DOI] [PubMed] [Google Scholar]
- 35.Hasan S., Singh K., Danisuddin M., Verma P.K., Khan A.U. Inhibition of major virulence pathways of Streptococcus mutans by quercitrin and deoxynojirimycin: A synergistic approach of infection control. PLoS ONE. 2014;9:e91736. doi: 10.1371/journal.pone.0091736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Zhang J., Rui X., Wang L., Guan Y., Sun X., Dong M. Polyphenolic extract from Rosa rugosa tea inhibits bacterial quorum sensing and biofilm formation. Food Control. 2014;42:125–131. doi: 10.1016/j.foodcont.2014.02.001. [DOI] [Google Scholar]
- 37.Prabu G.R., Gnanamani A., Sadulla S. Guaijaverin—A plant flavonoid as potential antiplaque agent against Streptococcus mutans. J. Appl. Microbiol. 2006;101:487–495. doi: 10.1111/j.1365-2672.2006.02912.x. [DOI] [PubMed] [Google Scholar]
- 38.Sivaranjani M., Gowrishankar S., Kamaladevi A., Pandian S.K., Balamurugan K., Ravi A.V. Morin inhibits biofilm production and reduces the virulence of Listeria monocytogenes—An in vitro and in vivo approach. Inter. J. Food Microb. 2016;237:73–82. doi: 10.1016/j.ijfoodmicro.2016.08.021. [DOI] [PubMed] [Google Scholar]
- 39.Moran A., Gutierrez S., Martinez-Blanco H., Ferrero M.A., Monteagudo-Mera A., Rodriguez-Aparicio L.B. Non-toxic plant metabolites regulate staphylococcus viability and biofilm formation: A natural therapeutic strategy useful in the treatment and prevention of skin infections. Biofouling. 2014;30:1175–1182. doi: 10.1080/08927014.2014.976207. [DOI] [PubMed] [Google Scholar]
- 40.Vandeputte O.M., Kiendrebeogo M., Rajaonson S., Diallo B., Mol A., El Jaziri M., Baucher M. Identification of catechin as one of the flavonoids from Combretum albiflorum bark extract that reduces the production of quorum-sensing-controlled virulence factors in Pseudomonas aeruginosa PAO1. Appl. Environ. Microbiol. 2010;76:243–253. doi: 10.1128/AEM.01059-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kang M.S., Oh J.S., Kang I.C., Hong S.J., Choi C.H. Inhibitory effect of methyl gallate and gallic acid on oral bacteria. J. Microbiol. 2008;46:744–750. doi: 10.1007/s12275-008-0235-7. [DOI] [PubMed] [Google Scholar]
- 42.Shao D., Li J., Li J., Tang R., Liu L., Shi J., Huang Q., Yang H. Inhibition of gallic acid on the growth and biofilm formation of Escherichia coli and Streptococcus mutans. J. Food Sci. 2015;80:1299–1305. doi: 10.1111/1750-3841.12902. [DOI] [PubMed] [Google Scholar]
- 43.Rendeková K., Fialová S., Jánošová L., Mučaji P., Slobodníková L. The activity of Cotinus coggygria Scop. leaves extract on Staphylococcus aureus strains in planktonic and biofilm growth forms. Molecules. 2016;21:50. doi: 10.3390/molecules21010050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Blanco A.R., Sudano-Roccaro A., Spoto G.C., Nostro A., Rusciano D. Epigallocatechin gallate inhibits biofilm formation by ocular staphylococcal isolates. Antimicrob. Agents Chemother. 2005;49:4339–4343. doi: 10.1128/AAC.49.10.4339-4343.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Xu X., Zhou X.D., Wu C.D. The tea catechin epigallocatechin gallate suppresses cariogenic virulence factors of Streptococcus mutans. Antimicrob. Agents Chemother. 2011;55:1229–1236. doi: 10.1128/AAC.01016-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Asahi Y., Noiri Y., Miura J., Maezono H., Yamaguchi M., Yamamoto R., Azakami H., Hayashi M., Ebisu S. Effects of the tea catechin epigallocatechin gallate on Porphyromonas gingivalis biofilms. J. Appl. Microbiol. 2014;116:1164–1171. doi: 10.1111/jam.12458. [DOI] [PubMed] [Google Scholar]
- 47.Lee P., Tan K.S. Effects of epigallocatechin gallate against Enterococcus faecalis biofilm and virulence. Arch. Oral Biol. 2015;60:393–399. doi: 10.1016/j.archoralbio.2014.11.014. [DOI] [PubMed] [Google Scholar]
- 48.Hancock V., Dahl M., Vejborg R.M., Klemm P. Dietary plant components ellagic acid and tannic acid inhibit Escherichia coli biofilm formation. J. Med. Microbiol. 2010;59:496–498. doi: 10.1099/jmm.0.013680-0. [DOI] [PubMed] [Google Scholar]
- 49.Slobodníková L., Fialová S., Hupková H., Grančai D. Rosmarinic acid interaction with planktonic and biofilm Staphylococcus aureus. Nat. Prod. Commun. 2013;8:1747–1750. [PubMed] [Google Scholar]
- 50.Fialová S., Slobodníková L., Veizerová L., Grančai D. Lycopus europaeus: Phenolic fingerprint, antioxidant activity and antimicrobial effect on clinical Staphylococcus aureus strains. Nat. Prod. Res. 2015;29:2271–2274. doi: 10.1080/14786419.2015.1010086. [DOI] [PubMed] [Google Scholar]
- 51.Lin M.-H., Chang R., Hua M.-Y., Wu Y.-C., Liu S.-T. Inhibitory effects of 1,2,3,4,6-penta-O-galloyl-β-d-glucopyranose on biofilm formation by Staphylococcus aureus. Antimicrob. Agents Chemother. 2011;55:1021–1027. doi: 10.1128/AAC.00843-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Donlan R.M. Biofilms and device-associated infections. Emerg. Infect. Dis. 2001;7:277–281. doi: 10.3201/eid0702.010226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Nagy M., Grančai D., Mučaji P. Farmakognózia Biogenéza Prírodných Látok. Osveta; Martin, TN, USA: 2011. [Google Scholar]
- 54.Samanta A., Das G., Das S.K. Roles of flavonoids in plants. Int. J. Pharm. Sci. Technol. 2011;6:12–35. [Google Scholar]
- 55.Taylor P.W., Hamilton-Miller J.M.T., Stapleton P.D. Antimicrobial properties of green tea catechins. Food Sci. Technol. Bull. 2005;2:71–81. doi: 10.1616/1476-2137.14184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ikigai H., Nakae T., Hara Y., Shimamura T. Bactericidal catechins damage the lipid bilayer. Biochim. Biophys. Acta. 1993;1147:132–136. doi: 10.1016/0005-2736(93)90323-R. [DOI] [PubMed] [Google Scholar]
- 57.Yam T.S., Hamilton-Miller J.M.T., Shah S. The effect of a component of tea (Camellia sinensis) on methicillin resistance, PBP2′ synthesis, and β-lactamase production in Staphylococcus aureus. J. Antimicrob. Chemother. 1998;42:211–216. doi: 10.1093/jac/42.2.211. [DOI] [PubMed] [Google Scholar]
- 58.Sudano Roccaro A.L., Blanco A.R., Giuliano F., Rusciano D., Enea V. Epigallocatechin-gallate enhances the activity of tetracycline in staphylococci by inhibiting its efflux from bacterial cells. Antimicrob. Agents Chemother. 2004;48:1968–1973. doi: 10.1128/AAC.48.6.1968-1973.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Payne D.E., Martin N.R., Parzych K.R., Rickard A.H., Underwood A., Boles B.R. Tannic acid inhibits Staphylococcus aureus surface colonization in an IsaA-dependent manner. Infect. Immun. 2013;81:496–504. doi: 10.1128/IAI.00877-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Lee J.-H., Park J.-H., Cho H.S., Joo S.W., Cho M.H., Lee J. Anti-biofilm activities of quercetin and tannic acid against Staphylococcus aureus. Biofouling. 2013;29:491–499. doi: 10.1080/08927014.2013.788692. [DOI] [PubMed] [Google Scholar]
- 61.Luis A., Silva F., Sousa S., Duarte A.P., Domingues F. Antistaphylococcal and biofilm inhibitory activities of gallic, caffeic, and chlorogenic acids. Biofouling. 2014;30:69–79. doi: 10.1080/08927014.2013.845878. [DOI] [PubMed] [Google Scholar]
- 62.Bakkiyaraj D., Nandhini J.R., Malathy B., Pandian S.K. The anti-biofilm potential of pomegranate (Punica granatum L.) extract against human bacterial and fungal pathogens. Biofouling. 2013;29:929–937. doi: 10.1080/08927014.2013.820825. [DOI] [PubMed] [Google Scholar]
- 63.Lee J.-H., Kim Y.-G., Ryu S.Y., Cho M.H., Lee J. Ginkgolic acids and Ginkgo biloba extract inhibit Escherichia coli O157:H7 and Staphylococcus aureus biofilm formation. Int. J. Food Microbiol. 2014;174:47–55. doi: 10.1016/j.ijfoodmicro.2013.12.030. [DOI] [PubMed] [Google Scholar]
- 64.Quave C.L.L., Estévez-Carmona M., Compadre C.M., Hobby G., Hendrickson H., Beenken K.E., Smeltzer M.S. Ellagic acid derivatives from Rubus ulmifolius inhibit Staphylococcus aureus biofilm formation and improve response to antibiotics. PLoS ONE. 2012;7:e28737. doi: 10.1371/journal.pone.0028737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Kiran M.D., Adikesavan N.V., Cirioni O., Giacometti A., Silvestri C., Scalise G., Ghiselli R., Saba V., Orlando F., Shoham M., et al. Discovery of a quorum-sensing inhibitor of drug-resistant staphylococcal infections by structure-based virtual screening. Mol. Pharmacol. 2008;73:1578–1586. doi: 10.1124/mol.107.044164. [DOI] [PubMed] [Google Scholar]
- 66.Trentin D.S., Silva D.B., Frasson A.P., Rzhepishevska O., da Silva M.V., de L Pulcini E., James G., Soares G.V., Tasca T., Ramstedt M., et al. Natural green coating inhibits adhesion of clinically important bacteria. Sci. Rep. 2015;5:82–87. doi: 10.1038/srep08287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Lamaison J.L., Petitjean-Freytet C., Duband F., Carnat A. Rosmarinic acid content and the antioxidant activity in french Lamiaceae. Fitoterapia. 1991;62:166–170. [Google Scholar]
- 68.Knobloch J.K., Bartscht K., Sabottke A., Rohde H., Feucht H.H., Mack D. Biofilm formation by Staphylococcus epidermidis depends on functional RsbU, an activator of the sigB operon: Differential activation mechanisms due to ethanol and salt stress. J. Bacteriol. 2001;183:2624–2633. doi: 10.1128/JB.183.8.2624-2633.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Bernier S.P., Surette M.G. Concentration-dependent activity of antibiotics in natural environments. Front. Microbiol. 2013;4:122–134. doi: 10.3389/fmicb.2013.00020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Arciola C.R., Campoccia D., Ravaioli S., Montanaro L. Polysaccharide intercellular adhesin in biofilm: Structural and regulatory aspects. Front. Cell Infect. Microbiol. 2015;5 doi: 10.3389/fcimb.2015.00007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Middleton E., Jr., Chithan K. The Flavonoids: Advances in Research Since 1986. Chapman and Hall; London, UK: 1993. [Google Scholar]
- 72.Mirzoeva O., Grishanin R., Calder P. Antimicrobial action of propolis and some of its components: The effects on growth, membrane potential and motility of bacteria. Microbiol. Res. 1997;152:239–246. doi: 10.1016/S0944-5013(97)80034-1. [DOI] [PubMed] [Google Scholar]
- 73.Plaper A., Golob M., Hafner I., Oblak M., Solmajer T., Jerala R. Characterization of quercetin binding site on DNA gyrase. Biochem. Biophys. Res. Commun. 2003;306:530–536. doi: 10.1016/S0006-291X(03)01006-4. [DOI] [PubMed] [Google Scholar]
- 74.Wu D., Kong Y., Han C., Chen J., Hu L., Jiang H., Shen X. d-Alanine: d-alanine ligase as a new target for the flavonoids quercetin and apigenin. Int. J. Antimicrob. Agents. 2008;32:421–426. doi: 10.1016/j.ijantimicag.2008.06.010. [DOI] [PubMed] [Google Scholar]
- 75.Ávila H.P., Smânia Ede F., Monache F.D., Smânia A. Structure-activity relationship of antibacterial chalcones. Bioorg. Med. Chem. 2008;16:9790–9794. doi: 10.1016/j.bmc.2008.09.064. [DOI] [PubMed] [Google Scholar]
- 76.Pastrana-Bonilla E., Akoh C.C., Sellappan S., Krewer G. Phenolic content and antioxidant capacity of muscadine grapes. J. Agric. Food Chem. 2003;51:5497–5503. doi: 10.1021/jf030113c. [DOI] [PubMed] [Google Scholar]
- 77.Xu C., Yagiz Y., Hsu W.Y., Simonne A., Lu J., Marshall M.R. Antioxidant, antibacterial, and antibiofilm properties of polyphenols from muscadine grape (Vitis rotundifolia Michx.) pomace against selected foodborne pathogens. J. Agric. Food Chem. 2014;62:6640–6649. doi: 10.1021/jf501073q. [DOI] [PubMed] [Google Scholar]
- 78.Kalkhambkar R.G., Kulkarni G.M., Kamanavalli C.M., Premkumar N., Asdaq S.M.B., Sun C.M. Synthesis and biological activities of some new fluorinated coumarins and 1-aza coumarins. Eur. J. Med. Chem. 2008;43:2178–2188. doi: 10.1016/j.ejmech.2007.08.007. [DOI] [PubMed] [Google Scholar]
- 79.Alvarez-Suarez J.M., Gasparrini M., Forbes-Hernández T.Y., Mazzoni L., Giampieri F. The composition and biological activity of honey: A focus on Manuka honey. Foods. 2014;3:420–432. doi: 10.3390/foods3030420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Lu J., Turnbull L., Burke C.M., Liu M., Carter D.A., Schlothauer R.C., Whitchurch C.B., Harry E.J. Manuka-type honeys can eradicate biofilms produced by Staphylococcus aureus strains with different biofilm-forming abilities. PeerJ. 2014;2:e326. doi: 10.7717/peerj.326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Krzyściak W., Jurczak A., Kościelniak D., Bystrowska B., Skalniak A. The virulence of Streptococcus mutans and the ability to form biofilms. Eur. J. Clin. Microbiol. Infect. Dis. 2014;33:499–515. doi: 10.1007/s10096-013-1993-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Costalonga M., Herzberg M.C. The oral microbiome and the immunobiology of periodontal disease and caries. Immunol. Lett. 2014;162:22–38. doi: 10.1016/j.imlet.2014.08.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Ren Z., Chen L., Li J., Li Y. Inhibition of Streptococcus mutans polysaccharide synthesis by molecules targeting glycosyltransferase activity. J. Oral Microbiol. 2016;8:31095. doi: 10.3402/jom.v8.31095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Xu X., Zhou X.D., Wu C.D. Tea catechin epigallocatechin gallate inhibits Streptococcus mutans biofilm formation by suppressing gtf genes. Arch. Oral Biol. 2012;57:678–683. doi: 10.1016/j.archoralbio.2011.10.021. [DOI] [PubMed] [Google Scholar]
- 85.Sakanaka S.I., Aizawa M., Kim M., Yamamoto T. Inhibitory effects of green tea polyphenols on growth and cellular adherence of an oral bacterium, Porphyromonas gingivalis. Biosci. Biotechnol. Biochem. 1996;60:745–749. doi: 10.1271/bbb.60.745. [DOI] [PubMed] [Google Scholar]
- 86.Kováč J., Kováč D., Slobodníková L., Kotulová D. Enterococcus faecalis and Candida albicans in the dental root canal and periapical infections. Bratisl. Med. J. 2013;114:716–720. [PubMed] [Google Scholar]
- 87.Sendamangalam V., Choi O.K., Kim D., Seo Y. The antibiofouling effect of polyphenols against Streptococcus mutans. Biofouling. 2011;27:13–19. doi: 10.1080/08927014.2010.535897. [DOI] [PubMed] [Google Scholar]
- 88.Bonifait L.L., Grenier D. Cranberry polyphenols: Potential benefits for dental caries and periodontal disease. J. Can. Dent. Assoc. 2010;76:a130. [PubMed] [Google Scholar]
- 89.Duarte S., Gregoire S., Singh A.P., Vorsa N., Schaich K., Bowen W.H., Koo H. Inhibitory effects of cranberry polyphenols on formation and acidogenicity of Streptococcus mutans biofilms. FEMS Microbiol. Lett. 2006;257:50–56. doi: 10.1111/j.1574-6968.2006.00147.x. [DOI] [PubMed] [Google Scholar]
- 90.Koo H., Duarte S., Murata R.M., Scott-Anne K., Gregoire S., Watson G.E., Singh A.P., Vorsa N. Influence of cranberry proanthocyanidins on formation of biofilms by Streptococcus mutans on saliva-coated apatitic surface and on dental caries development in vivo. Caries Res. 2010;44:116–126. doi: 10.1159/000296306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Feng G., Klein M.I., Gregoire S., Singh A.P., Vorsa N., Koo H. The specific degree-of-polymerization of A-type proanthocyanidin oligomers impacts Streptococcus mutans glucan-mediated adhesion and transcriptome responses within biofilms. Biofouling. 2013;29:629–640. doi: 10.1080/08927014.2013.794456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Thimothe J., Bonsi I.A., Padilla-Zakour O.I., Koo H. Chemical characterization of red wine grape (Vitis vinifera and Vitis interspecific hybrids) and pomace phenolic extracts and their biological activity against Streptococcus mutans. J. Agric. Food Chem. 2007;55:10200–10207. doi: 10.1021/jf0722405. [DOI] [PubMed] [Google Scholar]
- 93.Furiga A., Lonvaud-Funel A., Dorignac G., Badet C. In vitro anti-bacterial and anti-adherence effects of natural polyphenolic compounds on oral bacteria. J. Appl. Microbiol. 2008;105:1470–1476. doi: 10.1111/j.1365-2672.2008.03882.x. [DOI] [PubMed] [Google Scholar]
- 94.Furiga A., Roques C., Badet C. Preventive effects of an original combination of grape seed polyphenols with amine fluoride on dental biofilm formation and oxidative damage by oral bacteria. J. Appl. Microbiol. 2013;116:761–771. doi: 10.1111/jam.12395. [DOI] [PubMed] [Google Scholar]
- 95.Muñoz-González I., Thurnheer T., Bartolomé B., Moreno-Arribas M.V. Red wine and oenological extracts display antimicrobial effects in an oral bacteria biofilm model. J. Agric. Food Chem. 2014;62:4731–4737. doi: 10.1021/jf501768p. [DOI] [PubMed] [Google Scholar]
- 96.Yano A., Kikuchi S., Takahashi T., Kohama K., Yoshida Y. Inhibitory effects of the phenolic fraction from the pomace of Vitis coignetiae on biofilm formation by Streptococcus mutans. Arch. Oral Biol. 2012;57:711–719. doi: 10.1016/j.archoralbio.2012.01.001. [DOI] [PubMed] [Google Scholar]