

Abstract
Alkaloids, a category of natural products with ring structures and nitrogen atoms, include most U.S. Food and Drug Administration approved plant derived anti-cancer agents. Evodiamine is an alkaloid with attractive multitargeting antiproliferative activity. Its high content in the natural source ensures its adequate supply on the market and guarantees further medicinal study. To the best of our knowledge, there is no systematic review about the antiproliferative effects of evodiamine derivatives. Therefore, in this article the review of the antiproliferative activities of evodiamine will be updated. More importantly, the antiproliferative activities of structurally modified new analogues of evodiamine will be summarized for the first time.
Keywords: evodiamine, alkaloid, antiproliferative effect, structural modification
1. Introduction
Nature derived compounds are important for therapeutic leads [1,2,3,4,5,6,7]. Especially for anti-cancer agents, more than half cancer chemotherapy drugs are natural products and their derivatives [8,9,10]. Natural sources often provide privileged skeletons and define novel drug targets [11,12]. Among which, alkaloids, containing extra nitrogen atoms compared to other natural products, enrich structural diversity and some of them show better drug-like properties, such as drug-protein affinity and bioavailability. Most U.S. Food and Drug Administration approved plant derived anti-cancer agents are alkaloids, including vincristine, paclitaxel, and homoharringtonine. Evodiamine (1, Figure 1) is a famous alkaloid with a quinazolinocarboline skeleton. It was isolated from Evodia rutaecarpa, the dried and nearly ripe fruits of Euodia rutaecarpa (Juss.) Benth., E. rutaecarpa (Juss.) Benth var. officinalis (Dode) Huang, and E. rutaecarpa (Juss.) Benth var. bodinieri (Dode) Huang, three plants belonging to the family of Rutaceae. Evodiamine has been traditionally used to cure headache, amenorrhea, postpartum hemorrhage and gastrointestinal disorders. The biological activities of evodiamine have been widely investigated, including anti-obesity [13,14], anti-inflammatory [15,16], anti-atherosclerotic [17], neuroprotective [18,19], anti-gastrointestinal motility [20], and antiproliferative effects [21]. Among them, the antiproliferative activity of the multitargeting molecule evodiamine is attractive.
Figure 1.
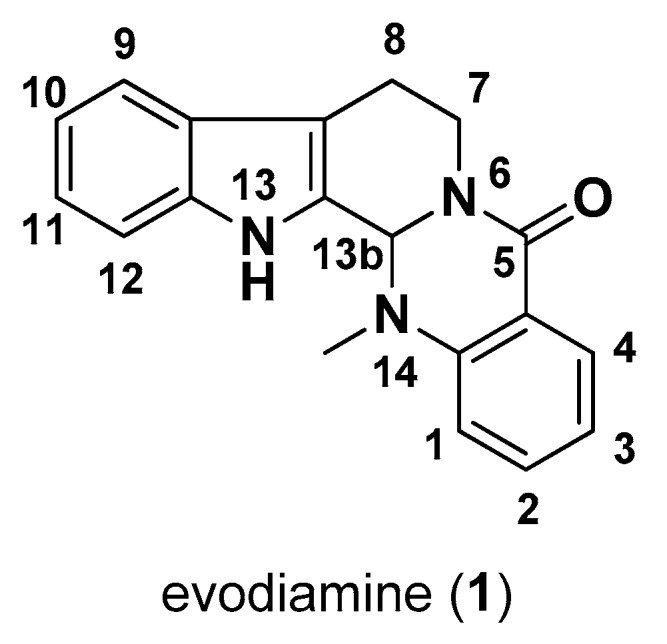
The structure and numbering of evodiamine (1).
In the past eight years, there have been five reviews on the biological activity of evodiamine. Evodiamine focusing on antiproliferative activities and molecular mechanisms have been reviewed by Jiang in 2009 [22], and Lu in 2012 [23]. In 2013, Yu et al. summarized the pharmacokinetics and the detailed exploration of target-binding properties of evodiamine [24]. Moreover, a patent review for therapeutic and cosmetic applications of evodiamine and its derivatives was reported by Gavaraskar in 2015 [25]. In 2016, Tan et al. reviewed the evodiamine functions and mechanisms of action in various chronic diseases [21]. To the best of our knowledge, there is no systematic review about the antiproliferative effects of evodiamine derivatives. This article updated the review of the antiproliferative activities of evodiamine. Moreover, the antiproliferative activities of structurally modified new analogues of evodiamine were summarized for the first time.
2. Pharmacological Activity
Previous studies evaluated the cytostatic effects of evodiamine in a variety of cancer cell lines. The cytotoxic effects of evodiamine in cancer cells were related to the induction of apoptosis, as well as inhibition of proliferation, migration, cell cycle progression, and angiogenesis. In addition, evodiamine was shown to regulate these cellular behaviors by affecting multi-targets. These works will be discussed in details.
2.1. Cytotoxic Effects in Lung Cancer Cells
Less than 15% of all lung cancers are small cell lung cancer (SCLC), and nearly 85% of lung cancer patients were diagnosed as non-small cell lung cancer (NSCLC) [26]. In human SCLC NCI-H1688 and NCI-H446 cells, G2/M arrest and the subsequent apoptosis were induced by evodiamine through mitochondria-dependent intrinsic apoptosis pathway rather than death receptor (DR)-induced extrinsic apoptotic pathway [27]. Moreover, evodiamine was shown to effectively increase the mitochondrial membrane depolarization and Bax/Bcl-2 ratio in human A549 NSCLC cell. The increased release of cytochrome c to cytosol activated intrinsic apoptotic pathway mediated by activation of caspase-9, -3, while increased release of cytochrome c to nuclear activated extrinsic apoptotic pathway mediated through activation of caspase-8. These studies indicated that evodiamine could induce apoptosis in NSCLC via both intrinsic and extrinsic pathways [28,29]. Further, the proliferation inhibition in A549 cells was induced by evodiamine, which was linked with the ability of evodiamine to increase the expression of oncoprotein metadherin, promote oxidative injury, arrest the cell cycle, and regulate the tumor-associated genes expression by controlling protein kinase B/ nuclear factor-κB (AKT/NF-κB) and sonic hedgehog/GLI family zinc finger 1 (SHH/GLI1) pathways [30,31]. Su et al. (2018) reported that evodiamine activate DNA methyltransferase 3A (DNMT3A)-induced neurogenic locus notch homolog protein 3 (NOTCH3) methylation, and subsequently induced growth inhibition in both A549 and H1299 NSCLC cells [32]. Moreover, the cytoprotective autophagy was induced by evodiamine in Lewis lung carcinoma (LLC) cells, and the combination of evodiamine with autophagy inhibitor therapy increased cell chemosensitivity [33]. Invasion activity of LLC cells in vitro and in vivo was shown to be decreased by evodiamine in previous studies [34,35]. As angiogenesis plays an important role in the development of lung carcinogenesis [36], the anti-angiogenesis activity of evodiamine was investigated by Shyu et al. (2006). Evodiamine was shown to decrease the expression of vascular endothelial growth factor (VEGF), a potent endothelial cell mitogen, in human lung adenocarcinoma CL1 cells. The conditioned medium derived from CL1 cells induced the formation of tube-like structures by human umbilical vein endothelial cells (HUVECs), while conditioned medium produced from evodiamine-pretreated CL1 cells exerted low ability of inducing capillary tube formation. Consistently, treatment of evodiamine could also decrease blood vessel density in chick embryo chorioallantoic membrane in vivo [37].
Treatment of erlotinib is not effective in the treatment of NSCLC cells with wild-type epidermal growth factor receptor (EGFR) [38]. By inactivating mammalian target of rapamycin/p70 ribosomal S6 kinase 1/Mcl-1 (mTOR/S6K1/Mcl-1) pathway, evodiamine and erlotinib acted in synergy in inhibiting cell proliferation and increasing apoptosis in NSCLC cells (NCI-H460, NCI-H1299, NCI-H1650, and A549) with wild-type EGFR [39] (Figure 2).
Figure 2.

The schematic representation of cytotoxic effects of evodiamine.
The low water solubility of evodiamine limited its application [40]. To enhance the bioactivity of evodiamine in lung cancer cells, a previous study used the fructus bruceae oil-based emulsive nanosystems to increase the bioavailability of evodiamine in A549 cells [41]. Moreover, the antitumor effects of evodiamine were also enhanced with the emulsive nanosystems in A549-tumor-bearing mice [42].
2.2. Cytotoxic Effects in Liver Cancer Cells
Liver cancer ranks second among causes of cancer death, and about 90% of liver cancers worldwide are hepatocellular carcinoma (HCC) [43]. Yang et al. (2017) reported that evodiamine downregulated cell viability and inhibited cell cycle progression in human HCC HepG2 cells by decreasing p-Akt level and increasing the levels of apoptotic proteins Bax, cleaved-caspase-3 and cleaved-PARP (poly (ADP-ribose) polymerase) [44]. Signal transducer and activator of transcription signaling 3 (STAT3) is involved in the development of HCC, and the impact of STAT3 on evodiamine-induced cell death of HepG2 was determined by Yang et al. (2013). The study revealed that evodiamine suppressed the phosphorylation of Tyr705 of STAT3 leading to the growth inhibition of HepG2 cell line. Moreover, the expression of phosphatase shatterproof 1 (SHP-1) in HepG2 cells was increased by evodiamine treatment, and the inhibition of SHP-1 attenuated evodiamine-suppressed activation of STAT3 [45]. In another study, evodiamine induced the expression of tumor-suppressor WWdomain-containing oxidoreductase (WWOX) in human HepG2 and mouse Hepa1-6 cells. And the inhibition of WWOX reversed the cytotoxic effect of evodiamine. As such, the anti-hepatocellular carcinoma activity of evodiamine was linked with the activation of WWOX [46]. Activator protein 1 (AP-1) is required for tumor promotion [47]. Chao et al. (2011) reported that evodiamine effectively inhibited AP-1 activity and anchorage-independent transformation of human liver cells [48]. Furthermore, the progression of HCC is promoted under hypoxia stress, and a previous study evaluated the impact of evodiamine on growth of HCC cells (HepG2, Hep3B, and Huh-7) subjected to hypoxia. The cytotoxic effects of vorinostat, an anti-HCC drug, were shown to be increased by co-treatment with evodiamine through the downregulation of hypoxia-inducible factor 1α (HIF-1α) [49]. In addition, the combination of evodiamine with berberine acted in synergy in inducing cell death of SMMC-7721 human hepatocellular carcinoma cells [50].
Signaling pathways involved in angiogenesis are promising targets for HCC therapy, and evodiamine was shown to exert cytotoxic effects on HCCs (HepG2, SMMC-7721, H22) by downregulating angiogenesis, which was linked with inhibition of expressions of β-catenin and VEGFa [51]. Besides its role in regulating angiogenesis, the activation of β-catenin induced expressions of target genes that in turn promoted cell proliferation. The results indicated that β-catenin might be one of the factors that are critical to the development of HCC [52]. Yet, it remains unclear whether the downregulation of β-catenin expression contributes to evodiamin-induced cytotoxicity in HCC cells.
Qiu et al. (2016) incorporated evodiamine into hydroxypropyl-β-cyclodextrin (EVO/HP-β-CD) to increase its bioavailability in HCC cells. Compared with evodiamine alone, EVO/HP-β-CD exerted higher cytotoxic activities in human hepatoma HepG2 cells as evidenced by higher levels of apoptosis induced by the activation of caspase-3 [53].
2.3. Cytotoxic Effects in Leukemia Cells
Leukemias, characterized by over-proliferation of abnormal blood cells, represent 31% of all cancers [54]. Evodiamine was shown to exert antiproliferative activity against human leukemia cells THP-1, K562, CCRF-CEM and camptothecin-resistant CCRF-CEM/C1 by inactivating topoisomerase I (topo I) and II (topo II). In addition, the formation of cleavage complexes of topoisomerases with DNA was not affect by evodiamine [55]. Evodiamine was also reported to cause the mitotic arrest and a consequent apoptosis in CCRF-CEM cells. And the study revealed that evodiamine-induced apoptosis was correlated with increased ratio of Bax/Bcl-2 and activity of caspase-3 [56]. However, benzyloxycarbonyl-Val-Ala-Aspfluoromethyl ketone (z-VAD-fmk), pan-caspase inhibitor, only partly attenuated the anti-apoptotic activity of evodiamine in human leukemic U937 cells, and overexpression of Bcl-2 failed to completely block the cytotoxic effect of evodiamine [57]. Further study showed that evodiamine treatment in U937 cells can also activate an apoptosis-inducing factor (AIF)-mediated apoptosis, which is not downregulated by Bcl-2 overexpression and caspase-independent. In addition, evodiamine is not toxic to human normal peripheral blood mononuclear cell (PBMC) [57,58]. In another study, peroxisome proliferator-activated receptor γ (PPARγ) signaling pathway was reported to be involved in the proliferation inhibition of evodiamine on K562 cells via inhibiting cyclin D1 and stimulating of p21, the important factors for the regulation of cell cycle progression [59]. Moreover, evodiamine decreased NF-κB activity and expressions of NF-κB-dependent antiapoptotic, proliferative, and metastatic genes in human myeloid leukemia KBM-5 cells, which indicated NF-κB might be another potential target of evodiamine for anti-leukemia therapy [60] (Figure 3).
Figure 3.

Cytotoxic effects of evodiamine in leukemia cells.
2.4. Cytotoxic Effects in Breast Cancer Cells
Breast cancer accounts for 23% of all cancers, and is highly prevalent in females [61]. Evodiamine was reported to downregulate migration and upregulate apoptosis by inactivating phosphorylation of extracellular signal-regulated kinase (p-ERK) and activating p38 mitogen-activated protein kinase (MAPK) in human breast cancer MDA-MB-231 cells [62]. By arresting the cell cycle at G0/G1 phase and inducing apoptosis, evodiamine could also effectively reduce the viability of human breast cancer MCF-7 cells. Further, the combination of evodiamine with berberine possesses synergistic cytotoxic activity [63]. The regulation of expressions of ERα, ERβ, and PPARγ, as well as inactivation of Ras/MEK/ERK pathway, may also contribute to the growth inhibitory activity of evodiamine in breast cancer cell lines, such as T47D, MDA-MB-468, MCF-7, and MDA-MB-231 [64,65]. Moreover, evodiamine has been reported to inhibit topo I activity in MCF-7 cells by inducing stabilization of the complex of topo I and DNA, and subsequently inhibit the replication and transcription of DNA [66]. However, the data are controversial to the results published by Pan et al. (2012) showing that evodiamine interferes with the enzyme catalytic cycle rather than affecting the stabilization of the covalent complexes of enzyme-DNA [55]. Cancer stem-like cells (CSLCs) play an important role in tumor progression [67]. Han et al. (2016) enriched the CSLCs in the mammosphere-forming condition during culture of MDA-MB-231 or MCF-7 cells. Compared with bulk carcinoma cells, CSLCs were more sensitive to cytotoxic effects of evodiamine treatment. The selective activation of p53 and p21, and inactivation of Rb in CSLCs led to cell death at the G1/S checkpoint [68].
As the occurrence of multidrug resistance is a major problem for the therapy of breast cancer, the inhibitory activity of evodiamine against doxorubicin- and adriamycin-resistant human breast cancer NCI/ADRRES cells was determined in previous studies [69,70]. Evodiamine reversed phenomenon of apoptosis resistance through inhibition of expression of IAPs, upregulation of Bcl-2, and activation of Ras/MEK/ERK pathway, which subsequently disrupted the microtubule network and induced apoptosis of NCI/ADRRES cells.
Zou et al. (2016) developed a nanoparticulate delivery system based on PLGA nanoparticles (EVO-PLGA NPs) to enhance the uptake of evodiamine by breast cancer. Compared with evodiamine alone, the antiproliferation effects of EVO-PLGA NPs in MCF-7 cells were even more obvious. However, EVO-PLGA NPs failed to enhance the impact of evodiamine on G2/M cell cycle arrest, suggesting that the nanoparticles sustained evodiamine release [71].
2.5. Cytotoxic Effects in Ovarian Cancer Cells
Ovarian cancer is another common gynecological tumor, and >70% of patients ultimately die from the disease [72,73]. Chen et al. (2016) evaluate the cytotoxic activities of evodiamine in human epithelial ovarian cancer cell lines ES-2, SKOV-3, A2780, and A2780CP. Evodiamine induced apoptosis of these cancer cells via activation of double-stranded RNA-activated protein kinase-like ER kinase (PERK) and c-Jun N-terminal kinase (JNK), as well as disruption of mitochondrial membrane potential. The structure-activity relationships (SARs) study demonstrated that the alkyl group at position 14 of evodiamine plays an important role in inducing apoptosis in ovarian cancer cells [74]. Further, the suppression of PI3K/Akt pathway and downregulation of p38 MAPK and ERK1/2 expressions might also be correlated with evodiamine-induced cell death of human ovarian cancer cell line HO8910PM [75]. As chemoresistance is associated with poor prognosis in patients with ovarian cancer, Zhong et al. (2015) evaluated the impact of evodiamine on cell viability of chemoresistant epithelial ovarian cancer cells (paclitaxel-resistant A2780/PTXR cells). Evodiamine was shown to enhance the chemosensitivity potential of paclitaxel-resistant A2780/PTXR cells by downregulating MDR-1 expression, which indicated the potential of evodiamine in the treatment of chemoresistant ovarian cancer [76]. In addition, evodiamine resulted in G2/M arrest in HER-2/neu-overexpressing SKOV3 cells (taxol resistant ovarian cancer cell line), while the underlying mechanism is still unclear [77].
2.6. Cytotoxic Effects in Cervical Cancer Cells
The number of cervical cancer deaths is higher than any other gynecological tumor, and the development of cervical cancer is highly correlated with infections with carcinogenic human papilloma virus [78]. By arresting cell cycle at G2/M, evodiamine was shown to induce apoptosis in human cervical cancer HeLa cells with caspase-3, -8-dependent pathway [79]. However, a controversy study reported that evodiamine activated caspase-3 without affecting the activity of caspase-8, -1 in HeLa cells [80]. In addition, the increased ratios of Bax/Bcl-2 and Bax/Bcl-xL are responsible for apoptosis triggered by evodiamine-induced productions of reactive oxygen species (ROS) and nitric oxide (NO). Autophagy was shown to be activated by evodiamine at an early period and reduced at late stages, and the blockage of autophagy resulted in apoptosis in HeLa cells [81]. Further study also suggested that G2/M cell cycle arrest in HeLa cells was triggered by ROS/NO productions, which could be attenuated by the inhibition of protein tyrosine kinase (PTK) [82]. Moreover, DNA topo I was shown to be associated with poor prognosis in ovarian cancers, and evodiamine exerted inhibitory effect against topo I catalytic activity. Accordantly, evodiamine effectively decreased the tumor volume in the subcutaneous xenografts of the human ovarian cancer cell line, A2780R2000, in a mouse model [83].
2.7. Cytotoxic Effects in Glioma Cells
Glioblastomas is the most frequent type of gliomas with highly invasive behavior [84]. In human glioblastoma U87 cells, evodiamine was reported to increase the intracellular calcium level, and activate autophagy and apoptosis, while the induction of autophagy blocked the apoptosis. In another study, evodiamine increased cytosolic calcium level, autophagy and apoptosis in U87 cells, which was attenuated by the inhibition of endoplasmic reticulum (ER) calcium channel. The study indicated an important role of calcium in mediating autophagy and apoptosis pathways in U87 cells [85,86]. Consistently, Wu et al. (2017) found that evodiamine induced G2/M arrest and decreased the viability of both human glioblastoma U87 cells and rat glioblastoma C6 cells, while incubation with JNK inhibitor attenuated the effects of evodiamine [87]. In addition, the inactivation of PI3K/AKT pathway and upregulation of phosphorylation of p38 MAPK were also shown to be linked with the cytotoxic activity of evodiamine in human glioblastoma U251 and LN229 cells [88]. Moreover, evodiamine did not reduce cell viability of primary rat astrocytes. A study of SARs further showed that methyl at position 14 may be important for the cytotoxic effect of evodiamine against glioblastoma cells [87].
The tumor necrosis factor-α-related apoptosis-inducing ligand (TRAIL) could promote the apoptosis in cancer cells, while the resistance to TRAIL limits the effects of chemotherapy [89]. TRAIL alone failed to exert cytotoxic effect in human glioblastoma U87 cells. However, evodiamine increased the apoptosis promoted by TRAIL through the upregulation of expressions of DR4, DR5, and caspase-8, -3 [90].
2.8. Cytotoxic Effects in Colorectal Carcinoma Cells
Colorectal cancer is a common cancer in the elderly [91]. Evodiamine induced apoptosis of human colorectal carcinoma cells COLO-205 and HCT-116 via the upregulation of p53 and Bax/Bcl-2 ratio, as well as decreasing mitochondrial transmembrane potential [92]. In another study, evodiamine exerted a prominent effect on expression of DNMTs and microRNAs (miRNAs) during early tumorigenesis in rat malignant colonic tissue stimulated by transforming growth factor-β1 (TGF-β1). Among these evodiamine-regulated miRNAs, the increase of expression of miR-29a (targets DNMT3A/3B) was most obvious [93]. Liu et al. (2016) found that the level of miR-429 was upregulated in malignant human colorectal tissues compared with normal tissues, which was attenuated by evodiamine administration [94]. These studies indicated that different miRNA plays different roles in mediating anti-tumor effect of evodiamine. In addition, evodiamine exerted cytotoxic effect in human colon carcinoma HT-29 cells, which was attributed to the inhibition of Wnt/β-catenin pathway and telomerase activity [95,96,97]. Another study reported that the increased JNK activity also contributed to G2/M cell cycle arrest and apoptosis stimulated by evodiamine in human colon carcinoma cells COLO-205 and HT-29 [98]. Moreover, S-phase arrest, autophagy and caspase-dependent apoptosis were increased under the stimulation of evodiamine in human colon carcinoma Lovo cells, and the evodiamine-induced cell cycle arrest was attributed to the downregulation of the cyclin A and cyclin A-dependent kinase 2 (CDK2) expressions [99,100]. Further, the alkylation at position 14 was shown to play an important role in the cytotoxicity of evodiamine in colonic cancer cells [97], which is accordant with the previous study on ovarian cancer cells [74].
Previous studies showed that evodiamine decreased the invasion activity of colon 26-L5 cells in vitro and suppressed lung-specific metastasis of 26-L5 cells in mice in vivo [34,35,101]. The downregulated migration activity was revealed to be linked with the inactivation of Janus kinase/STAT3/matrix metalloproteinase 3 (JAK2/STAT3/MMP3) pathway in human colorectal cancer HCT-116 cells [102,103]. Moreover, the expression of midkine, an important growth factor for carcinogenesis, was inhibited by evodiamine in human colon cancer SW620 cells, which might be involved in the inhibitory activities of evodiamine against both invasion ability and anchorage independence growth of SW620 cells [104].
Breast cancer resistance protein, also called ABCG2, is crucial for multidrug resistance (MDR) mechanism in colorectal carcinoma. Evodiamine was shown to suppress ABCG2-mediated drug resistance against oxaliplatin (L-OHP)-resistant HCT-116/L-OHP cells via inactivation of NF-κB in vitro. And the activation of NF-κB and colorectal tumor growth in a MDR xenograft model were inhibited in vivo [105] (Figure 4). In contrast, evodiamine was shown to upregulate NF-κB and p53 levels by down-regulating histone deacetylase 3 in HCT-116 xenografts in another study [106].
Figure 4.

Sites of action of evodiamine against colorectal carcinoma cells.
2.9. Cytotoxic Effects in Gastric Cancer Cells
Gastric cancer, a leading cause of cancer death, is frequently diagnosed at advanced stages [107]. Yue et al. (2013) revealed that evodiamine inhibited the viability of cells from gastric tumor tissues, which was correlated with downregulation of thymidylate synthase (TS) and excision repair cross-complementing 1 (ERCC1) [108]. Besides, evodiamine increased the beneficial effect of radiation treatment for human gastric carcinoma BGC-823 cells. The inactivation of Her-2/AKT/Bcl-2 pathway was further revealed to be correlated with the cytostatic activity of evodiamine [109]. In human gastric carcinoma SGC-7901 cells, evodiamine promoted proliferation and arrested the cell progression at G1 and G2/M phases, which further contributed to the cytotoxic and carcinogenic mechanisms of evodiamine [110,111,112,113,114]. Shen et al. (2015) reported that evodiamine also upregulated apoptosis through the upregulation of caspase-3 and survivin in SGC-7901 cells [115]. However, z-VAD-fmk only partially blocked evodiamin-promoted apoptosis in SGC-7901 cells [116]. The upregulation of acid sphingomyelinase (aSMase) and the downregulation of sphingomyelin were found to be the caspase-independent mechanism for evodiamine-induced apoptosis in SGC-7901 cells [117]. Moreover, evodiamine exerted inhibitory effect against the invasion activity of SGC-7901 cells through downregulation of midkine and polo-like kinase 1 (PLK1) expressions [118,119]. Gastric cancer stem cells were shown to promote tumorigenesis, metastasis and drug resistance [120]. Wen et al. (2015) reported that evodiamine could inactivate the Wnt/β-catenin pathway and subsequently suppress stem cell potential and proliferation of gastric cancer stem cells and repress the epithelial-to-mesenchymal transition [121]. However, evodiamine increased the secretion of IL-8, the cytokine promotes cell invasion, from human gastric cancer AGS cells, while berberine could counteract this side-effect of evodiamine. The study provides a safe strategy implementing combination therapy with evodiamine and berberine [122].
2.10. Cytotoxic Effects in Melanoma Cells
Melanoma is the primary skin cancer with ~132,000 cases worldwide annually [123]. The impact of evodiamine on invasion activity of melanoma was determined in previous studies using B16-F10 melanoma cells [34,35]. And these studies showed that the invasion potential of B16-F10 cells in transwell migration assay was effectively inhibited by evodiamine treatment.
In A375-S2 melanoma cells, the downregulation of protein kinase C (PKC) and silent information regulator of transcription (SIRT1), as well as the upregulation of Bax/Bcl-2 ratio, were correlated with evodiamine-induced increase of apoptosis. Evodiamine could also induce necrosis by activating p38 MAPK and inactivating ERK in A375-S2 cells [79,124,125,126,127]. During the course of evodiamine-induced apoptosis, the upregulated intracellular ROS followed by an onset of mitochondrial depolarization contributes to both Fas-mediated extrinsic and mitochondria-mediated intrinsic apoptosis pathways [128]. Evodiamine exerted a strong inductive effect on the production NO in A375-S2 cells, while the inactivation of p38 MAPK and NF-kB blocked evodiamine induced NO synthesis and cell damage [129]. Wang et al. (2005) found that the inhibition of interleukin 1 (IL-1) receptor in A375-S2 cells effectively attenuated the increased evodiamine-induced expressions of Fas-ligand (Fas-L), caspas-8, -3, and the phosphorylated p38 MAPK, which indicated IL-1-induced death cascade might be one of the targets for evodiamine [130]. Moreover, MG132, the inhibitor of ubiquitin-proteasome, further increased the sensitivity of A375-S2 cell to evodiamine induced cell death through the upregulation of the expressions of Fas-L, Bcl-2, and caspase-3 [131].
2.11. Cytotoxic Effects in Thyroid Cancer Cells
Thyroid carcinoma accounts for 1.3–1.5% of all cancers [132]. Lv et al. (2016) reported that evodiamine-treated human papillary thyroid cancer K1 cells underwent apoptosis that was correlated with inactivation of PI3K/Akt pathway [133]. Moreover, increased levels of G2/M arrest and caspase cascade-induced apoptosis contributed to evodiamin-induced cytotoxicity in human anaplastic thyroid cancer ARO cells [134]. Accordantly, the tumorigenesis of ARO in a xenograft mice model was inhibited by evodiamine in vivo [135]. The changes in protein expression patterns in ARO after evodiamine stimulation were further determined by Yu et al. (2018). And 77 proteins involved in transcriptional control, protein folding, and lipid metabolism contributed to evodiamine-induced cytotoxicity in ARO cells [136].
2.12. Cytotoxic Effects in Nasopharyngeal Carcinoma Cells
Nasopharyngeal carcinoma, the poorly understood cancer, arises from the nasopharynx epithelium [137]. Peng et al. (2015) reported an inhibitory role of evodiamine on the migration and invasion of human nasopharyngeal carcinoma HONE1 and CNE1 cells without affecting cell proliferation. Further investigation revealed that evodiamine downregulated cancer invasion- and metastasis-associated MMP-2 activity, which was controlled by NF-кB p65 nuclear translocation and the ERK1/2 phosphorylation [138].
2.13. Cytotoxic Effects in Renal Cancer Cells
Renal cell carcinoma usually develops from the lining of kidney tubules, and it accounts for ~90% of all renal cancer [139]. Evodiamine was shown to induce apoptosis in human renal cell carcinoma Caki-1, ACHN, A498, and 786-O cells, which was attenuated by stimulation with inhibitors of PERK or JNK. A methyl at position 14 was shown to be important for evodiamine’s cytotoxic action in renal carcinoma cells in a structure activity study [140], which is accordant with the previous study on glioblastoma cells [87]. Moreover, a transcriptome analysis using Caki-1 cells indicated that evodiamine-regulated genes in Caki-1 cells were mainly involved in apoptosis and cell cycle [141].
2.14. Cytotoxic Effects in Bladder Cancer Cells
Bladder cancer is a common genitourinary cancer with 70% recurrence rate [142,143]. It has been shown that human bladder cancer cells 253J and T24 underwent apoptosis in response to the stimulation of evodiamine. In addition, evodiamine enhanced TRAIL-induced apoptosis through the regulation of mTOR/S6K1/Mcl-1 pathway [144]. On the other hand, evodiamine arrested cell cycle progression in G2/M phase and increased caspase-dependent apoptosis in bladder tumor cell lines 5637 and HT1197, and the effects of evodiamine was linked with the activations of p38 MAPK and JNK [145].
2.15. Cytotoxic Effects in Pancreatic Cancer Cells
Pancreatic cancer is a leading cause of death from cancer with limited effective therapies [146]. Evodiamine was shown to induce apoptosis via inhibition of NF-κB activation, and subsequently upregulated Bax/Bcl-2 ratio and ultimately induced apoptosis in human pancreatic tumor cell PANC-1 [147]. Accordantly, evodiamine inactivated NF-κB in human pancreatic tumor cell SW1990, while gemcitabine was shown to induce the activation of NF-κB. These findings were consistent with a previous study researching the impact of evodiamine on activation of NF-κB in various carcinoma cell lines [60]. As the activation of NF-κB is one of the reasons for the development of chemoresistance during cancer therapy, the inactivation of NF-κB by evodaimine may sensitize pancreatic cancer cells to anti-tumor therapies [148].
2.16. Cytotoxic Effects in Prostate Cancer Cells
Prostate cancer is one of the leading causes of cancer-related death in males [149]. Through inhibition of microtubule spindle formation, evodiamine was shown to effectively arrest cell cycle progression in G2/M phase and subsequent induce apoptosis in human prostate cancer PC-3 and DU145 cells [150,151]. And the cytotoxic activities of evodiamine may attributed to the activation of cyclin B1/p34cdc2 complex and caspase-3, -9 [152].
2.17. Cytotoxic Effects in Osteosarcoma Cells
Osteosarcoma is the most common bone sarcoma in children and young adults [153]. Evodiamine was shown to inhibit the proliferation of human osteosarcoma 143B02 cells through inactivation of PTEN/PI3K/Akt pathway [154]. By inactivating Raf/MEK/ERK signaling pathway, evodiamine also induced growth inhibition and inactivated the activities of migration and invasion of U2OS cells [155,156]. Moreover, the inactivation of Wnt/β-catenin signaling pathway contributed to the cytotoxic activity of evodiamine in human osteosarcoma MG-63 cells [157].
2.18. Cytotoxic Effects in Oral Cancer Cells
Oral cancer is common in men in developing countries [158]. The apoptosis was induced in human oral cancer cells (MC3 and HSC4) by evodiamine though activation of caspase-3 and PARP cleavage. Further studies indicated the AKT/myeloid cell leukemia 1 (Mcl-1) signaling pathway is responsible for the observed cytotoxicty of evodiamine in oral cancer cells [159].
2.19. Cytotoxic Effects in Salivary Adenoid Cystic Carcinoma Cells
Salivary adenoid cystic carcinoma, the cancer with varied morphologic and clinical manifestations, is an uncommon salivary gland malignancy [160]. The effect of evodiamine on salivary adenoid cystic carcinoma SACC-M cell growth regulation was determined by Chen et al. (2012). And the study revealed that evodiamine significantly decreased SACC-M cell viability through downregulation of proliferation and upregulation of apoptosis [161].
3. Advances in Antiproliferative Evodiamine Derivatives
Naturally selected chemical scaffolds with bioactivity can be regarded as privileged structures. However, the probability for direct use of natural product as therapeutic drug is very low. The most common method is to generate analogues by structural modification approaches [162,163,164,165,166,167]. Although evodiamine has been well studied for its antiproliferative effects, it is still unqualified to be approved for clinical use. The physicochemical properties and antitumor potency remain to be significantly improved. For better design derivatives and understand the SARs, the existing medicinal chemistry work of evodiamine is summarized as follows.
In 2010, Dong et al. found evodiamine as human Topo I inhibitor by structure-based virtual screening and lead optimization [168]. Subsequently, by introducing alkyl, benzoyl and benzyl groups and ester at N-13, evodiamine derivatives were synthesized as Topo I inhibitors (Scheme 1). The results showed that the substituted benzoyl groups were favorable for the antiproliferative activity and spectrum. Among them, N-benzoyl analogue 7u showed the strongest antiproliferative effects against A549, HCT-116 and MDA-MB-435 human cancer cell lines with IC50 values of 0.86, 2.6 and 0.049 μM, respectively. In addition, in vitro Topo I inhibition assay also demonstrated that 7u was stronger than evodiamine and was active (at 100 μM) against the relaxation of supercoiled DNA, which was consistent with their docking results.
Scheme 1.

Synthetic route of evodiamine 1 and evodiamine derivatives 7a–x. Reagents and conditions: (a) HCOOEt, reflux, 12 h; (b) POCl3, 0 °C 2 h, then rt, 2 h; (c) ClCOOEt, reflux, 12 h; (d) 4, 30–60 °C, 6 h; (e) RX, NaH, 80 °C, 24 h.
Two years later, Dong et al. [169] synthesized a library of evodiamine A-ring (8–16), E-ring (17–28), A, E-ring disubstituted (29–36), and D-ring derivatives (37–40) (Figure 5). Taking the asymmetric center C-13b into account, they investigated the effects of C-13b chirality on the antiproliferative activity. Both R- and S-isomers of a highly active derivative 31 were synthesized (Scheme 2). In vitro results indicated compounds 8, 11, 18, 19 and 31 exerted substantially increased cytostatic activity against HCT-116, MDA-MB-435 and A549 cell lines, with GI50 values lower than 3 nM. The C13b chirality showed distinct effects toward HCT-116 and A549 cells, (S)-31 was slightly more active. Furthermore, these derivatives could induce apoptosis in A549 cells. Together with further computational calculations, these evodiamine derivatives could be used as dual inhibitors of topo I and topo II.
Figure 5.

Chemical structures of evodiamine A-ring (8–16), E-ring (17–28), A, E-ring disubstituted (29–36), and D-ring derivatives (37–40).
Scheme 2.

Synthetic route of (R)- and (S)-31. Reagents and conditions: (a) NaNO2, hydrochloric acid, 0–5 °C, 2 h; (b) KOH, rt, 12 h; (c) rt, 10 h; (d) HCOOH, reflux, 0.5 h; (e) POCl3, reflux, 3 days; (f) cat. A, HCOOH:Et3N (5:2), 0 °C, 8 h; (g) cat. B, HCOOH:Et3N (5:2), 0 °C, 8 h; (h) H2, 10% Pd/C, rt, 12 h.
The mechanism of Topo I and Topo II inhibition suggested that test compounds (11, 33, 35, and 36, Figure 5) acted as Topo II catalytic inhibitors. Moreover, at the dose of 1 mg/kg or 2 mg/kg, compounds 11 and 36 showed good antitumor efficacy and low toxicity in vivo. The SARs could be concluded as follows. Firstly, the 1-, 3-, 10- and 12-positions were favorable to be substituted, chlorine was more suitable than fluorine for both 3- and 10-positions. Free hydroxyl group was important for strong antiproliferative potency. Secondly, for the D-ring, changing the carbonyl group to the thiocarbonyl group at C-5 and the transformation of methyl group to an oxygen atom at N-14 showed positive effects of antiproliferative activity. Thirdly, some compounds of C-3, C-10 disubstituted possess synergistic effects. Compounds 31–33 and 36 with hydrophilic groups showed good antiproliferative activity and water solubility.
In 2013, Song’s group designed and synthesized a series of evodiamine derivatives at N-13 (Scheme 3) [170]. They tested the cytotoxic activities of all derivatives against prostate cancer DU-145, PC-3, lung cancer H460, colon cancer HCT-5, glioblastoma SF-268 and MCF-7 cells. The results showed that 50b, 50c, 50i, 50p and 51b exhibited stronger antiproliferative potency and broader spectra. Among them, 50p was the strongest with IC50 values below 2 µM. The SARs indicated that the appropriate N-13 substitution of evodiamine resulted in higher cytotoxicity. The alkyl N-13 substituted groups decreased antiproliferative potency, and the derivatives with alkylamino-alkyl groups showed improved water solubility, but only moderate antiproliferative potency. The dimers exhibited moderate to good activity, and 50m with a benzoylmethyl group was inactive. The apoptosis related mechanisms of compounds 50c, 50p and 51b were also disclosed. Compounds 50c and 51b drastically induced apoptosis in MCF-7 cells at the early stage, while 50p and 51b induced apoptosis in HCT-15 cells at the late stage. Moreover, 50p and 51b induced late apoptosis in NCl-H460 cells with moderate increasing to 40–50%.
Scheme 3.

Synthetic route of evodiamine derivatives 50a–p and 51a–c. Reagents and conditions: (a) NaH, R1X, 8 h; (b) NaH, Br-R1-Br, 8 h.
In the same year, Nguyen’s group successfully isolated evodiamine enantiomers from 13 Evodia rutaecarpa samples by chiral high-performance liquid chromatographic method [171]. The results showed that S-(+)-evodiamine was present in higher concentration than R-(−)-evodiamine. Soon afterwards, Christodoulou et al. prepared the evodiamine enantiomers by chemical synthesis methods (Scheme 4), and synthesized a series of evodiamine C-ring modified derivatives (Scheme 5) [172]. In vitro antiproliferative assay showed that all compounds had low potency against H460, MCF-7 and HepG2 cells. However, (S)-evodiamine 1a was the most active compound among them. Furthermore, in vitro Topo I inhibition of 1a and 1b showed effects only a high concentration of 500 μM. The human sirtuins SIRT1, SIRT2 and SIRT3 assays indicated S configuration was favorable.
Scheme 4.

Synthetic route of evodiamine enantiomers 1a and 1b. Reagents and conditions: (a) (1S)-10-camphorsulfonyl chloride, NaH, 60 °C, 10 h; (b) NaOH, MeOH/THF 1:1, rt, 18 h.
Scheme 5.

Synthetic route of C-ring modified (S)-evodiamine. Reagents and conditions: (a) HCOOH, (Ac)2O, rt; (b) HCl, dioxane, rt, 2 h; (c) pyridine, reflux, 2 h; (d) LiOH, reflux, 1.5 h; (e) various amines, HATU, DIPEA, rt.
Compounds 1a, 1b, 56, ent-56, 58 and 59 showed better inhibitory effects on SIRT2 (60–80%) than SIRT1. In docking studies, they were well-fit to the binding pocket. Due to the steric hindrance, bulky substituents at position 5 had unfavorable poses, such as 60, 61, 62 and 63. COOH, CONHCH3 and COOCH3 moieties at position 5 could increase the affinity towards sirtuins by additional hydrogen bonds. 1a, 56 and ent-56 also showed selectivity of SIRT2 over SIRT3. With this information in hand, evodiamine can act as a potential scaffold of sirtuins inhibitors.
In late 2013, a series of synthetic evodiamine enantiomer derivatives were synthesized by De Petrocellis et al. (Figure 6) [173]. The results showed the derivatives of S-(+)-evodiamine were more potent than R-(−)-evodiamine derivatives at human and rat transient receptor potential vanilloid type-1 (TRPV1) in transfected HEK-293 cells, with the corresponding EC50 values of 5 μM and 2 nM. Especially, a new lead analogue (72) was identified as the most potent TRPV1 agonist and better than the reference capsaicin. Previously, evodiamine was also reported as an agonist of TRPV1 in vitro and in vivo [174]. This work further confirmed evodiamine would qualify as potent TRPV1 agonists.
Figure 6.

Chemical structures of synthetic evodiamine enantiomer analogues 64–83.
In 2014, sulfonic acid esters and 5-methylene derivatives of evodiamine were designed and synthesized by Liang et al. (Scheme 6) [175]. 85e exhibited more potent antiproliferative activity than evodiamine against A549, HepG2, U251, MCF-7 and HeLa cells, and 86a–g showed higher IC50 values than the parallel compounds 85a–g. The preliminary SARs suggested that N-para benzenesulfonylation was favorable for antiproliferative activity. The conversion from carbonyl group to methylene at C-5 would decrease the activities, and aromatic sulfonic acid esters were better than alkyl sulfonic acid esters for the antiproliferative activity. Moreover, when R was nitryl group, the activity decreased dramatically. In vivo antitumor assessment in the HepS xenograft showed 85e could improve index of thymus and spleen and possessed less side-effect. Finally, according to in silico molecular docking, compounds 1 and 85a–g showed high binding affinity with the target. Although 84 and 86a–g had no binding affinity with the important residues of Topo I, they still exhibited antiproliferative activities. Between the ligand and the active site of Topo I, besides intermolecular H-bonds, other interactions might exist.
Scheme 6.

Synthetic route of evodiamine and 5-methylene evodiamine derivatives 85a–g and 86a–g. Reagents and conditions: (a) LiAlH4, rt, 8 h; (b) NaH, substituted benzenesulfonyl chloride or cyclopropanesulfonyl chloride, rt, 8–12 h.
In 2015, a library of 11 evodiamine-inspired scaffolds and their derivatives were obtained (Figure 7) [176]. Most of them showed antiproliferative activities against selected cancer cell lines. Among them, 104, 3-chloro-10-hydroxyl thio-evodiamine, showed good antitumor efficacy and low toxicity. The mechanism and target studies indicated that 104 was the first-in-class Topo I/Topo II/tubulin triple inhibitor. The significance of this work could be concluded, that it provided the in-depth SAR of evodiamine inspired scaffolds, a series of potent antitumor molecules and the first-in-class Topo I/Topo II/tubulin inhibitors.
Figure 7.

Chemical structures of evodiamine-inspired novel scaffold derivatives 87–106.
In 2016, Hua’s group published their first work in the field of the structural modification of evodiamine [177]. A series of nitric oxide donating evodiamine derivatives were designed and synthesized (Scheme 7). Of which, compound 112 showed good antiproliferative activity against A549, BGC-823 and human hepatoma Bel-7402 cells with IC50 values of 2.31, 0.07 and 2.10 µM, respectively. More importantly, 112 exhibited no cytotoxicity (>100 µM) against L-02 human normal liver cells. In Bel-7402 and BGC-823 cells, all the derivatives could release more than 75 µM/L NO at 1 h, and below 30 µM/L in L-02 cells. The mechanism study showed that 112 induced apoptosis and caused S phase cell-cycle arrest in Bel-7402 cells through caspase-dependent mitochondria related pathways. Western blot analysis revealed 112 could up-regulate proapoptotic Bax, FAS, cytochrome c and caspase-3, -8, -9, and down-regulate the Bcl-2 expression.
Scheme 7.

Synthetic route of nitric oxide donating evodiamine derivative 112. Reagents and conditions: (a) ClCH2COOH, NaOH, 140 °C, 2 h; (b) H2O2, AcOH, rt, 3 h; (c) fuming HNO3, 90 °C, 4 h; (d) HO(CH2)6Br, NaH, rt, 3 h; (e) 110, DBU, −15 °C, 3 h.
In 2017, this group continued structural modifications of evodiamine, and synthesized a series of nitrogen mustard derivatives of evodiamine (Scheme 8) [178]. The antiproliferative activities of 114a–d, 115a–d and 116a–d against PC-3, HepG2, THP-1, HL-60 and PBMCs were tested. The results showed all the derivatives exhibited antiproliferative activities against tumor cells and no (>200 μM) antiproliferative activities against PBMCs. Among them, 116c showed the strongest cytotoxicity against HL-60 and THP-1 cells with IC50 values of 0.50 and 4.05 μM, respectively. Further mechanism study manifested that 116c could induce apoptosis and arrest cell cycle at G2/M phase via mitochondria-related pathways in HL-60 cells.
Scheme 8.

Synthetic route of nitrogen mustard derivatives (114–116) of evodiamine. Reagents and conditions: (a) NaH, rt, 3 h; (b) benzoic acid nitrogen mustard, methyl melphalan butyric acid or chlorambucil, EDCI, DMAP, rt, 12–24 h.
Recently, H2S donating derivatives of evodiamine (121–125) were further designed and synthesized (Scheme 9) [179]. Resulting hybrids were evaluated against Bel-7402, MCF-7, SGC-7901, human epithelial colorectal adenocarcinoma Caco-2 and HL-60 cells, and PBMCs for antiproliferative activities. Among these derivatives, compound 125c showed the strongest potency against Caco-2 and HL-60 cells with IC50 values of 2.02 and 0.58 μM, respectively. Furthermore, compound 125c possessed high selectivity between human normal PBMCs and HL-60 cells. The mechanism studies showed that 125c could arrest HL-60 cell cycle at the G2/M phase and induce apoptosis. Western blot results confirmed the intrinsic apoptotic pathways.
Scheme 9.

Synthetic route of H2S-donating derivatives (121–125) of evodiamine. Reagents and conditions: (a) Br(CH2)nBr, potassium carbonate, reflux, 8 h; (b) Br(CH2)nCOOC2H5, NaH, 80 °C, 12 h; (c) potassium hydroxide, rt, 2 h; (d) α-thioctic acid, 117, EDCI, DMAP, rt, 8–12 h; (e) 118a–c, TEA, 80 °C, 8 h.
4. Conclusions
The cytotoxic effects of evodiamine were correlated with its inhibitory activities against cell cycle progression, proliferation, invasion and angiogenesis, as well as apoptosis-induced effect. A variety of evodiamine derivatives have been synthesized to increase the antiproliferative effects of evodiamine. However, most of the studies were taken in in vitro and animal models, more clinical evidences regarding the effectiveness of evodiamine in the treatment of various types of human cancers are urgently needed. In addition, the low bioavailability limits the anti-cancer efficacy of evodiamine, novel evodiamine derivatives with enhanced bioavailability are necessary to be designed and synthesized. Further, development of new evodiamine delivery systems, such as a supermolecular nanoemulsion [180], or a phospholipid complex [40], might also be the appropriate strategies to increase the bioavailability of evodiamine in clinical practice.
Acknowledgments
This paper was financially supported by Key Laboratory of Quality Control of TCM of Liaoning Province (17-137-1-00), Career Development Support Plan for Young and Middle-aged Teachers in Shenyang Pharmaceutical University, General Scientific Research Projects of Department of Education in Liaoning Province (2017LQN05), and China Postdoctoral Science Foundation (2017M621161; 2018T110462).
Abbreviations
| AP-1 | Activator protein 1 |
| AIF | Apoptosis-inducing factor |
| AKT | Protein kinase B |
| aSMase | Acid sphingomyelinase |
| CSLCs | Cancer stem-like cells |
| DR | Death receptor |
| DNMT | DNA methyltransferase |
| EGFR | Epidermal growth factor receptor |
| EVO/HP-β-CD | Evodiamine into hydroxypropyl-β-cyclodextrin |
| ERK | Extracellular signal-regulated kinase |
| ER | Endoplasmic reticulum |
| ERCC1 | Excision repair cross-complementing 1 |
| Fas-L | Fas-ligand |
| HUVECs | Human umbilical vein endothelial cells |
| HCC | Hepatocellular carcinoma |
| HIF-1α | Hypoxia-inducible factor 1α |
| IL-1 | Interleukin 1 |
| JNK | c-Jun N-terminal kinase |
| JAK2 | Janus kinase 2 |
| LLC | Lewis lung carcinoma |
| L-OHP | Oxaliplatin |
| mTOR/S6K1 | Mammalian target of rapamycin/p70 ribosomal S6 kinase 1 |
| MAPK | Mitogen-activated protein kinase |
| miRNAs | microRNAs |
| MMP3 | Matrix metalloproteinase 3 |
| MDR | Multidrug resistance |
| Mcl-1 | Myeloid cell leukemia 1 |
| NF-κB | Nuclear factor-κB |
| NPs | Natural products |
| NSCLC | Non-small cell lung cancer |
| NO | Nitric oxide |
| NOTCH3 | Neurogenic locus notch homolog protein 3 |
| PARP | Poly (ADP-ribose) polymerase |
| PBMC | Peripheral blood mononuclear cell |
| PERK | Double-stranded RNA-activated protein kinase-like ER kinase |
| PTK | Protein tyrosine kinase |
| PLK1 | Polo-like kinase 1 |
| PKC | Protein kinase C |
| PPAR-γ | Peroxisome proliferator-activated receptor γ |
| ROS | Reactive oxygen species |
| SCLC | Small cell lung cancer |
| SHH/GLI1 | Sonic hedgehog/GLI family zinc finger 1 |
| STAT3 | Signal transducer and activator of transcription signaling 3 |
| SHP-1 | Shatterproof 1 |
| SARs | Structure-activity relationships |
| SIRT1 | Silent information regulator of transcription |
| Topo | Topoisomerase |
| TRAIL | Tumor necrosis factor-α-related apoptosis‑inducing ligand |
| TGF-β1 | Transforming growth factor-β1 |
| TS | Thymidylate synthase |
| TRPV1 | Transient receptor potential vanilloid type-1 |
| VEGF | Vascular endothelial growth factor |
| WWOX | WWdomain-containing oxidoreductase |
| z-VAD-fmk | Benzyloxycarbonyl-Val-Ala-Aspfluoromethyl ketone |
Author Contributions
H.H., D.L., and F.X. conceived the study; X.H., C.C., X.L., and X.W. performed the review of literature; X.H. and D.L. wrote the first draft of the manuscript; Y.J., H.H., and F.X. supervised the whole process; all authors approved the final draft.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 1.Rodrigues T., Reker D., Schneider P. Counting on natural products for drug design. Nat. Chem. 2016;8:531–541. doi: 10.1038/nchem.2479. [DOI] [PubMed] [Google Scholar]
- 2.Thomford N.E., Senthebane D.A., Rowe A., Munro D., Seele P., Maroyi A., Dzobo K. Natural products for drug discovery in the 21st century: Innovations for novel drug discovery. Int. J. Mol. Sci. 2018;19:1578. doi: 10.3390/ijms19061578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Mullard A. Bracing for the biosimilar wave. Nat. Rev. Drug Discov. 2017;16:152–154. doi: 10.1038/nrd.2017.36. [DOI] [PubMed] [Google Scholar]
- 4.Davis A.M., Plowright A.T., Valeur E. Directing evolution: The next revolution in drug discovery? Nat. Rev. Drug Discov. 2017;16:681–698. doi: 10.1038/nrd.2017.146. [DOI] [PubMed] [Google Scholar]
- 5.Singh S.P., Sashidhara K.V. Lipid lowering agents of natural origin: An account of some promising chemotypes. Eur. J. Med. Chem. 2017;140:331–348. doi: 10.1016/j.ejmech.2017.09.020. [DOI] [PubMed] [Google Scholar]
- 6.Wright G.D. Opportunities for natural products in 21st century antibiotic discovery. Nat. Prod. Rep. 2017;34:694–701. doi: 10.1039/C7NP00019G. [DOI] [PubMed] [Google Scholar]
- 7.Zhang H., Bai L., He J., Zhong L., Duan X., Ouyang L., Zhu Y., Wang T., Zhang Y., Shi J. Recent advances in discovery and development of natural products as source for anti-Parkinson’s disease lead compounds. Eur. J. Med. Chem. 2017;141:257–272. doi: 10.1016/j.ejmech.2017.09.068. [DOI] [PubMed] [Google Scholar]
- 8.Newman D.J., Cragg G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016;79:629–661. doi: 10.1021/acs.jnatprod.5b01055. [DOI] [PubMed] [Google Scholar]
- 9.Joshi P., Vishwakarma R.A., Bharate S.B. Natural alkaloids as P-gp inhibitors for multidrug resistance reversal in cancer. Eur. J. Med. Chem. 2017;138:273–292. doi: 10.1016/j.ejmech.2017.06.047. [DOI] [PubMed] [Google Scholar]
- 10.Seca A.M.L., Pinto D.C.G.A. Plant secondary metabolites as anticancer agents: Successes in clinical trials and therapeutic application. Int. J. Mol. Sci. 2018;19:263. doi: 10.3390/ijms19010263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rodrigues T., Sieglitz F., Bernardes G.J. Natural product modulators of transient receptor potential (TRP) channels as potential anti-cancer agents. Chem. Soc. Rev. 2016;45:6130–6137. doi: 10.1039/C5CS00916B. [DOI] [PubMed] [Google Scholar]
- 12.Pham D., Koide K. Discoveries, target identifications, and biological applications of natural products that inhibit splicing factor 3B subunit 1. Nat. Prod. Rep. 2016;33:637–647. doi: 10.1039/C5NP00110B. [DOI] [PubMed] [Google Scholar]
- 13.Wang T., Wang Y., Kontani Y., Kobayashi Y., Sato Y., Mori N., Yamashita H. Evodiamine improves diet-induced obesity in a uncoupling protein-1-independent manner: Involvement of antiadipogenic mechanism and extracellularly regulated kinase/mitogen-activated protein kinase signaling. Endocrinology. 2008;149:358–366. doi: 10.1210/en.2007-0467. [DOI] [PubMed] [Google Scholar]
- 14.Wang T., Wang Y., Yamashita H. Evodiamine inhibits adipogenesis via the EGFR-PKCα-ERK signaling pathway. FEBS Lett. 2009;583:3655–3659. doi: 10.1016/j.febslet.2009.10.046. [DOI] [PubMed] [Google Scholar]
- 15.Chiou W.F., Sung Y.J., Liao J.F., Shum A.Y., Chen C.F. Inhibitory effect of dehydroevodiamine and evodiamine on nitric oxide production in cultured murine macrophages. J. Nat. Prod. 1997;60:708–711. doi: 10.1021/np960495z. [DOI] [PubMed] [Google Scholar]
- 16.Fan X., Zhu J.Y., Sun Y., Luo L., Yan J., Yang X., Yu J., Tang W.Q., Ma W., Liang H.P. Evodiamine inhibits zymosan-induced inflammation in vitro and in vivo: Inactivation of NF-κB by inhibiting IκBα phosphorylation. Inflammation. 2017;40:1012–1027. doi: 10.1007/s10753-017-0546-0. [DOI] [PubMed] [Google Scholar]
- 17.Wang L., Eftekhari P., Schachner D., Ignatova I.D., Palme V., Schilcher N., Ladurner A., Heiss E.H., Stangl H., Dirsch V.M., et al. Novel interactomics approach identifies abca1 as direct target of evodiamine, which increases macrophage cholesterol efflux. Sci. Rep. 2018;8:11061. doi: 10.1038/s41598-018-29281-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Zhang Y.N., Yang Y.F., Yang X.W. Blood-brain barrier permeability and neuroprotective effects of three main alkaloids from the fruits of Euodia rutaecarpa with MDCK-pHaMDR cell monolayer and PC12 cell line. Biomed. Pharmacother. 2018;98:82–87. doi: 10.1016/j.biopha.2017.12.017. [DOI] [PubMed] [Google Scholar]
- 19.Zhang Y., Wang J., Wang C., Li Z., Liu X., Zhang J., Lu J., Wang D. Pharmacological basis for the use of evodiamine in Alzheimer’s disease: Antioxidation and antiapoptosis. Int. J. Mol. Sci. 2018;19:1527. doi: 10.3390/ijms19051527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ren H.X., Tang Q.C., Yan L., Xia H., Luo H.S. Evodiamine inhibits gastrointestinal motility via CCK and CCK1 receptor in water-avoidence stress rat model. Life Sci. 2018;209:210–216. doi: 10.1016/j.lfs.2018.08.003. [DOI] [PubMed] [Google Scholar]
- 21.Tan Q., Zhang J. Evodiamine and its role in chronic diseases. Adv. Exp. Med. Biol. 2016;929:315–328. doi: 10.1007/978-3-319-41342-6_14. [DOI] [PubMed] [Google Scholar]
- 22.Jiang J., Hu C. Evodiamine: A novel anti-cancer alkaloid from Evodia rutaecarpa. Molecules. 2009;14:1852–1859. doi: 10.3390/molecules14051852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lu J.J., Bao J.L., Chen X.P., Huang M., Wang Y.T. Alkaloids isolated from natural herbs as the anticancer agents. Evid. Based Complement. Altern. Med. 2012;2012:485042. doi: 10.1155/2012/485042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Yu H., Jin H., Gong W., Wang Z., Liang H. Pharmacological actions of multi-target-directed evodiamine. Molecules. 2013;18:1826–1843. doi: 10.3390/molecules18021826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gavaraskar K., Dhulap S., Hirwani R.R. Therapeutic and cosmetic applications of evodiamine and its derivatives—A patent review. Fitoterapia. 2015;106:22–35. doi: 10.1016/j.fitote.2015.07.019. [DOI] [PubMed] [Google Scholar]
- 26.Zhou C., Yao L.D. Strategies to improve outcomes of patients with egrf-mutant non-small cell lung cancer: Review of the literature. J. Thorac. Oncol. 2016;11:174–186. doi: 10.1016/j.jtho.2015.10.002. [DOI] [PubMed] [Google Scholar]
- 27.Fang C., Zhang J., Qi D., Fan X., Luo J., Liu L., Tan Q. Evodiamine induces G2/M arrest and apoptosis via mitochondrial and endoplasmic reticulum pathways in H446 and H1688 human small-cell lung cancer cells. PLoS ONE. 2014;9:e115204. doi: 10.1371/journal.pone.0115204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Mohan V., Agarwal R., Singh R.P. A novel alkaloid, evodiamine causes nuclear localization of cytochrome-c and induces apoptosis independent of p53 in human lung cancer cells. Biochem. Biophys. Res. Commun. 2016;477:1065–1071. doi: 10.1016/j.bbrc.2016.07.037. [DOI] [PubMed] [Google Scholar]
- 29.Hong J.Y., Park S.H., Min H.Y., Park H.J., Lee S.K. Anti-proliferative effects of evodiamine in human lung cancer cells. J. Cancer Prev. 2014;19:7–13. doi: 10.15430/JCP.2014.19.1.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Zou Y., Qin X., Xiong H., Zhu F., Chen T., Wu H. Apoptosis of human non-small-cell lung cancer A549 cells triggered by evodiamine through MTDH-dependent signaling pathway. Tumour Biol. 2015;36:5187–5193. doi: 10.1007/s13277-015-3174-z. [DOI] [PubMed] [Google Scholar]
- 31.Lin L., Ren L., Wen L., Wang Y., Qi J. Effect of evodiamine on the proliferation and apoptosis of A549 human lung cancer cells. Mol. Med. Rep. 2016;14:2832–2838. doi: 10.3892/mmr.2016.5575. [DOI] [PubMed] [Google Scholar]
- 32.Su T., Yang X., Deng J.H., Huang Q.J., Huang S.C., Zhang Y.M., Zheng H.M., Wang Y., Lu L.L., Liu Z.Q. Evodiamine, a novel NOTCH3 methylation stimulator, significantly suppresses lung carcinogenesis in vitro and in vivo. Front. Pharmacol. 2018;9:434. doi: 10.3389/fphar.2018.00434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Tu Y.J., Fan X., Yang X., Zhang C., Liang H.P. Evodiamine activates autophagy as a cytoprotective response in murine lewis lung carcinoma cells. Oncol. Rep. 2013;29:481–490. doi: 10.3892/or.2012.2125. [DOI] [PubMed] [Google Scholar]
- 34.Ogasawara M., Matsunaga T., Takahashi S., Saiki I., Suzuki H. Anti-invasive and metastatic activities of evodiamine. Biol. Pharm. Bull. 2002;25:1491–1493. doi: 10.1248/bpb.25.1491. [DOI] [PubMed] [Google Scholar]
- 35.Ogasawara M., Suzuki H. Inhibition by evodiamine of hepatocyte growth factor-induced invasion and migration of tumor cells. Biol. Pharm. Bull. 2004;27:578–582. doi: 10.1248/bpb.27.578. [DOI] [PubMed] [Google Scholar]
- 36.Sozzi G., Conte D., Leon M., Ciricione R., Roz L., Ratcliffe C., Roz E., Cirenei N., Bellomi M., Pelosi G., et al. Quantification of free circulating DNA as a diagnostic marker in lung cancer. J. Clin. Oncol. 2003;21:3902–3908. doi: 10.1200/JCO.2003.02.006. [DOI] [PubMed] [Google Scholar]
- 37.Shyu K.G., Lin S., Lee C.C., Chen E., Lin L.C., Wang B.W., Tsai S.C. Evodiamine inhibits in vitro angiogenesis: Implication for antitumorgenicity. Life Sci. 2006;78:2234–2243. doi: 10.1016/j.lfs.2005.09.027. [DOI] [PubMed] [Google Scholar]
- 38.Bianco R., Troiani T., Tortora G., Ciardiello F. Intrinsic and acquired resistance to EGFR inhibitors in human cancer therapy. Endocr. Relat. Cancer. 2005;12(Suppl. 1):S159–S171. doi: 10.1677/erc.1.00999. [DOI] [PubMed] [Google Scholar]
- 39.Li Y.L., Pan Y.N., Wu W.J., Mao S.Y., Sun J., Zhao Y.M., Dong J.Y., Zhang D.Y., Pan J.P., Zhang C., et al. Evodiamine induces apoptosis and enhances apoptotic effects of erlotinib in wild-type EGFR NSCLC cells via S6K1-mediated Mcl-1 inhibition. Med. Oncol. 2016;33:16. doi: 10.1007/s12032-015-0726-4. [DOI] [PubMed] [Google Scholar]
- 40.Tan Q., Shan L., Chen X., Wu M., Hong W., Yin H., Dan H., Xiong H., Zhang J. Design and evaluation of a novel evodiamine-phospholipid complex for improved oral bioavailability. AAPS PharmSciTech. 2012;13:534–547. doi: 10.1208/s12249-012-9772-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Aboalnaja K.O., Yaghmoor S., Kumosani T.A., Mcclements D.J. Utilization of nanoemulsions to enhance bioactivity of pharmaceuticals, supplements, and nutraceuticals: Nanoemulsion delivery systems and nanoemulsion excipient systems. Expert Opin. Drug Deliv. 2016;13:1327–1336. doi: 10.1517/17425247.2016.1162154. [DOI] [PubMed] [Google Scholar]
- 42.Zhao J., Liu S., Hu X., Zhang Y., Yan S., Zhao H., Zeng M., Li Y., Yang L., Zhang J. Improved delivery of natural alkaloids into lung cancer through woody oil-based emulsive nanosystems. Drug Deliv. 2018;25:1426–1437. doi: 10.1080/10717544.2018.1474970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ferlay J., Soerjomataram I., Dikshit R., Eser S., Mathers C., Rebelo M., Parkin D.M., Forman D., Bray F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer. 2015;136:E359–E386. doi: 10.1002/ijc.29210. [DOI] [PubMed] [Google Scholar]
- 44.Yang F., Shi L., Liang T., Ji L., Zhang G., Shen Y., Zhu F., Xu L. Anti-tumor effect of evodiamine by inducing Akt-mediated apoptosis in hepatocellular carcinoma. Biochem. Biophys. Res. Commun. 2017;485:54–61. doi: 10.1016/j.bbrc.2017.02.017. [DOI] [PubMed] [Google Scholar]
- 45.Yang J., Cai X., Lu W., Hu C., Xu X., Yu Q., Cao P. Evodiamine inhibits STAT3 signaling by inducing phosphatase shatterproof 1 in hepatocellular carcinoma cells. Cancer Lett. 2013;328:243–251. doi: 10.1016/j.canlet.2012.09.019. [DOI] [PubMed] [Google Scholar]
- 46.Hu C.Y., Wu H.T., Su Y.C., Lin C.H., Chang C.J., Wu C.L. Evodiamine exerts an anti-hepatocellular carcinoma activity through a wwox-dependent pathway. Molecules. 2017;22:1175. doi: 10.3390/molecules22071175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hsiang C.Y., Wu S.L., Ho T.Y. Activation of activator protein 1 and extracellular signal-regulated kinases in human hepatocellular transformation. Tumour Biol. 2004;25:313–320. doi: 10.1159/000081400. [DOI] [PubMed] [Google Scholar]
- 48.Chao D.C., Lin L.J., Hsiang C.Y., Li C.C., Lo H.Y., Liang J.A., Kao S.T., Wu S.L., Ho T.Y. Evodiamine inhibits 12-o-tetradecanoylphorbol-13-acetate-induced activator protein 1 transactivation and cell transformation in human hepatocytes. Phytother. Res. 2011;25:1018–1023. doi: 10.1002/ptr.3392. [DOI] [PubMed] [Google Scholar]
- 49.Li Y.L., Zhang N.Y., Hu X., Chen J.L., Rao M.J., Wu L.W., Li Q.Y., Zhang B., Yan W., Zhang C. Evodiamine induces apoptosis and promotes hepatocellular carcinoma cell death induced by vorinostat via downregulating HIF-1α under hypoxia. Biochem. Biophys. Res. Commun. 2018;498:481–486. doi: 10.1016/j.bbrc.2018.03.004. [DOI] [PubMed] [Google Scholar]
- 50.Wang X.N., Han X., Xu L.N., Yin L.H., Xu Y.W., Qi Y., Peng J.Y. Enhancement of apoptosis of human hepatocellular carcinoma SMMC-7721 cells through synergy of berberine and evodiamine. Phytomedicine. 2008;15:1062–1068. doi: 10.1016/j.phymed.2008.05.002. [DOI] [PubMed] [Google Scholar]
- 51.Shi L., Yang F., Luo F., Liu Y., Zhang F., Zou M., Liu Q. Evodiamine exerts anti-tumor effects against hepatocellular carcinoma through inhibiting β-catenin-mediated angiogenesis. Tumour Biol. 2016;37:12791–12803. doi: 10.1007/s13277-016-5251-3. [DOI] [PubMed] [Google Scholar]
- 52.Nejak-Bowen K.N., Monga S.P.S. Beta-catenin signaling, liver regeneration and hepatocellular cancer: Sorting the good from the bad. Semin. Cancer Biol. 2011;21:44–58. doi: 10.1016/j.semcancer.2010.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Qiu C., Gao L.N., Yan K., Cui Y.L., Zhang Y. A promising antitumor activity of evodiamine incorporated in hydroxypropyl-β-cyclodextrin: Pro-apoptotic activity in human hepatoma HepG2 cells. Chem. Cent. J. 2016;10:46. doi: 10.1186/s13065-016-0191-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Morra M.E., Kien N.D., Elmaraezy A., Abdelaziz O.A.M., Elsayed A.L., Halhouli O., Montasr A.M., Vu T.L., Ho C., Foly A.S., et al. Early vaccination protects against childhood leukemia: A systematic review and meta-analysis. Sci. Rep. 2017;7:15986. doi: 10.1038/s41598-017-16067-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Pan X., Hartley J.M., Hartley J.A., White K.N., Wang Z., Bligh S.W.A. Evodiamine, a dual catalytic inhibitor of type I and II topoisomerases, exhibits enhanced inhibition against camptothecin resistant cells. Phytomedicine. 2012;19:618–624. doi: 10.1016/j.phymed.2012.02.003. [DOI] [PubMed] [Google Scholar]
- 56.Huang Y.C., Guh J.H., Teng C.M. Induction of mitotic arrest and apoptosis by evodiamine in human leukemic T-lymphocytes. Life Sci. 2004;75:35–49. doi: 10.1016/j.lfs.2003.11.025. [DOI] [PubMed] [Google Scholar]
- 57.Lee T.J., Kim E.J., Kim S., Jung E.M., Park J.W., Jeong S.H., Park S.E., Yoo Y.H., Kwon T.K. Caspase-dependent and caspase-independent apoptosis induced by evodiamine in human leukemic U937 cells. Mol. Cancer Ther. 2006;5:2398–2407. doi: 10.1158/1535-7163.MCT-06-0167. [DOI] [PubMed] [Google Scholar]
- 58.Zhang X.D., Gillespie S.K., Hersey P. Staurosporine induces apoptosis of melanoma by both caspase-dependent and -independent apoptotic pathways. Mol. Cancer Ther. 2004;3:187–197. [PubMed] [Google Scholar]
- 59.Sun C., Zhang G., Luan S., Luan C., Shao H., Dong F., Liu X. Evodiamine inhibits the proliferation of leukemia cell line K562 by regulating peroxisome proliferators-activated receptor gamma (PPARγ) pathway. J. Recept. Signal Transduct. Res. 2016;36:422–428. doi: 10.3109/10799893.2015.1122040. [DOI] [PubMed] [Google Scholar]
- 60.Takada Y., Kobayashi Y., Aggarwal B.B. Evodiamine abolishes constitutive and inducible NF-κB activation by inhibiting IκBα kinase activation, thereby suppressing NF-κB-regulated antiapoptotic and metastatic gene expression, up-regulating apoptosis, and inhibiting invasion. J. Biol. Chem. 2005;280:17203–17212. doi: 10.1074/jbc.M500077200. [DOI] [PubMed] [Google Scholar]
- 61.Mittal S., Kaur H., Gautam N., Mantha A.K. Biosensors for breast cancer diagnosis: A review of bioreceptors, biotransducers and signal amplification strategies. Biosens. Bioelectron. 2017;88:217–231. doi: 10.1016/j.bios.2016.08.028. [DOI] [PubMed] [Google Scholar]
- 62.Du J., Wang X.F., Zhou Q.M., Zhang T.L., Lu Y.Y., Zhang H., Su S.B. Evodiamine induces apoptosis and inhibits metastasis in MDA-MB-231 human breast cancer cells in vitro and in vivo. Oncol. Rep. 2013;30:685–694. doi: 10.3892/or.2013.2498. [DOI] [PubMed] [Google Scholar]
- 63.Du J., Sun Y., Lu Y.Y., Lau E., Zhao M., Zhou Q.M., Su S.B. Berberine and evodiamine act synergistically against human breast cancer MCF-7 cells by inducing cell cycle arrest and apoptosis. Anticancer Res. 2017;37:6141–6151. doi: 10.21873/anticanres.12063. [DOI] [PubMed] [Google Scholar]
- 64.Wang K.L., Hsia S.M., Yeh J.Y., Cheng S.C., Wang P.S., Wang S.W. Anti-proliferative effects of evodiamine on human breast cancer cells. PLoS ONE. 2013;8:e67297. doi: 10.1371/journal.pone.0067297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Wang S., Chen M., Wang Y. Tumor suppressive activity of evodiamine in breast cancer cells via inhibition of Ras/MEK/ERK pathway and activation of PPARγ. Planta Med. 2012;78:P_79. doi: 10.1055/s-0032-1307587. [DOI] [Google Scholar]
- 66.Chan A.L., Chang W.S., Chen L.M., Lee C.M., Chen C.E., Lin C.M., Hwang J.L. Evodiamine stabilizes topoisomerase I-DNA cleavable complex to inhibit topoisomerase I activity. Molecules. 2009;14:1342–1352. doi: 10.3390/molecules14041342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Plaks V., Kong N., Werb Z. The cancer stem cell niche: How essential is the niche in regulating stemness of tumor cells? Cell Stem Cell. 2015;16:225–238. doi: 10.1016/j.stem.2015.02.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Han S., Woo J.K., Jung Y., Jeong D., Kang M., Yoo Y.J., Lee H., Oh S.H., Ryu J.H., Kim W.Y. Evodiamine selectively targets cancer stem-like cells through the p53-p21-Rb pathway. Biochem. Biophys. Res. Commun. 2016;469:1153–1158. doi: 10.1016/j.bbrc.2015.12.066. [DOI] [PubMed] [Google Scholar]
- 69.Liao C.H., Pan S.L., Guh J.H., Chang Y.L., Pai H.C., Lin C.H., Teng C.M. Antitumor mechanism of evodiamine, a constituent from chinese herb Evodiae fructus, in human multiple-drug resistant breast cancer NCI/ADR-RES cells in vitro and in vivo. Carcinogenesis. 2005;26:968–975. doi: 10.1093/carcin/bgi041. [DOI] [PubMed] [Google Scholar]
- 70.Wang S., Wang L., Shi Z., Zhong Z., Chen M., Wang Y. Evodiamine synergizes with doxorubicin in the treatment of chemoresistant human breast cancer without inhibiting p-glycoprotein. PLoS ONE. 2014;9:e97512. doi: 10.1371/journal.pone.0097512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Zou L., Chen F., Bao J., Wang S., Wang L., Chen M., He C., Wang Y. Preparation, characterization, and anticancer efficacy of evodiamine-loaded PLGA nanoparticles. Drug Deliv. 2016;23:908–916. doi: 10.3109/10717544.2014.920936. [DOI] [PubMed] [Google Scholar]
- 72.Ramser M., Eichelberger S., Däster S., Weixler B., Kraljević M., Mechera R., Tampakis A., Delko T., Güth U., Stadlmann S., et al. High OX40 expression in recurrent ovarian carcinoma is indicative for response to repeated chemotherapy. BMC Cancer. 2018;18:425. doi: 10.1186/s12885-018-4339-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Rauh-Hain J.A., Krivak T.C., Carmen M.G.D., Olawaiye A.B. Ovarian cancer screening and early detection in the general population. Rev. Obstet. Gynecol. 2011;4:15–21. [PMC free article] [PubMed] [Google Scholar]
- 74.Chen T.C., Chien C.C., Wu M.S., Chen Y.C. Evodiamine from Evodia rutaecarpa induces apoptosis via activation of JNK and PERK in human ovarian cancer cells. Phytomedicine. 2016;23:68–78. doi: 10.1016/j.phymed.2015.12.003. [DOI] [PubMed] [Google Scholar]
- 75.Wei L., Jin X., Cao Z., Li W. Evodiamine induces extrinsic and intrinsic apoptosis of ovarian cancer cells via the mitogen-activated protein kinase/phosphatidylinositol-3-kinase/protein kinase B signaling pathways. J. Tradit. Chin. Med. 2016;36:353–359. doi: 10.1016/s0254-6272(16)30049-8. [DOI] [PubMed] [Google Scholar]
- 76.Zhong Z.F., Tan W., Wang S.P., Qiang W.A., Wang Y.T. Anti-proliferative activity and cell cycle arrest induced by evodiamine on paclitaxel-sensitive and -resistant human ovarian cancer cells. Sci. Rep. 2015;5:16415. doi: 10.1038/srep16415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Yu C.-H., Lin R.-C., Wang P.S. Anti-proliferative effects of evodiamine and rutaecarpine on human ovarian cancer cell line SKOV3. Biol. Reprod. 2010;83(Suppl. 1):134. doi: 10.1093/biolreprod/83.s1.134. [DOI] [Google Scholar]
- 78.Cancer Genome Atlas Research Network Integrated genomic and molecular characterization of cervical cancer. Nature. 2017;543:378–384. doi: 10.1038/nature21386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Zhang Y., Wu L.J., Tashiro S., Onodera S., Ikejima T. Evodiamine induces tumor cell death through different pathways: Apoptosis and necrosis. Acta Pharmacol. Sin. 2004;25:83–89. [PubMed] [Google Scholar]
- 80.Fei X.F., Wang B.X., Li T.J., Tashiro S.I., Minami M., Xing D.J., Ikejima T. Evodiamine, a constituent of Evodiae fructus, induces anti-proliferating effects in tumor cells. Cancer Sci. 2003;94:92–98. doi: 10.1111/j.1349-7006.2003.tb01358.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Yang J., Wu L.J., Tashino S., Onodera S., Ikejima T. Reactive oxygen species and nitric oxide regulate mitochondria-dependent apoptosis and autophagy in evodiamine-treated human cervix carcinoma HeLa cells. Free Radic. Res. 2008;42:492–504. doi: 10.1080/10715760802112791. [DOI] [PubMed] [Google Scholar]
- 82.Yang J., Wu L.J., Tashino S.I., Onodera S., Ikejima T. Protein tyrosine kinase pathway-derived ROS/NO productions contribute to G2/M cell cycle arrest in evodiamine-treated human cervix carcinoma HeLa cells. Free Radic. Res. 2010;44:792–802. doi: 10.3109/10715762.2010.481302. [DOI] [PubMed] [Google Scholar]
- 83.Lee Y.C., Lee C.H., Tsai H.P., An H.W., Lee C.M., Wu J.C., Chen C.S., Huang S.H., Hwang J., Cheng K.T., et al. Targeting of topoisomerase I for prognoses and therapeutics of camptothecin-resistant ovarian cancer. PLoS ONE. 2015;10:e0132579. doi: 10.1371/journal.pone.0132579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Joseph J.V., Conroy S., Pavlov K., Sontakke P., Tomar T., Eggens-Meijer E., Balasubramaniyan V., Wagemakers M., den Dunnen W.F., Kruyt F.A. Hypoxia enhances migration and invasion in glioblastoma by promoting a mesenchymal shift mediated by the HIF1α-ZEB1 axis. Cancer Lett. 2015;359:107–116. doi: 10.1016/j.canlet.2015.01.010. [DOI] [PubMed] [Google Scholar]
- 85.Liu A.J., Wang S.H., Chen K.C., Kuei H.P., Shih Y.L., Hou S.Y., Chiu W.T., Hsiao S.H., Shih C.M. Evodiamine, a plant alkaloid, induces calcium/JNK-mediated autophagy and calcium/mitochondria-mediated apoptosis in human glioblastoma cells. Chem. Biol. Interact. 2013;205:20–28. doi: 10.1016/j.cbi.2013.06.004. [DOI] [PubMed] [Google Scholar]
- 86.Liu A.J., Wang S.H., Hou S.Y., Lin C.J., Chiu W.T., Hsiao S.H., Chen T.H., Shih C.M. Evodiamine induces transient receptor potential vanilloid-1-mediated protective autophagy in U87-MG astrocytes. Evid. Based Complement. Altern. Med. 2013;2013:354840. doi: 10.1155/2013/354840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Wu W.S., Chien C.C., Liu K.H., Chen Y.C., Chiu W.T. Evodiamine prevents glioma growth, induces glioblastoma cell apoptosis and cell cycle arrest through JNK activation. Am. J. Chin. Med. 2017;45:879–899. doi: 10.1142/S0192415X17500471. [DOI] [PubMed] [Google Scholar]
- 88.Wang R., Deng D., Shao N., Xu Y., Xue L., Peng Y., Liu Y., Zhi F. Evodiamine activates cellular apoptosis through suppressing PI3K/AKT and activating MAPK in glioma. OncoTargets Ther. 2018;11:1183–1192. doi: 10.2147/OTT.S155275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Ashkenazi A., Dixit V.M. Intracellular mechanisms of TRAIL: Apoptosis through mitochondrial-dependent and -independent pathways. Oncogene. 2001;20:2122–2133. doi: 10.1038/sj.onc.1204282. [DOI] [PubMed] [Google Scholar]
- 90.Khan M., Bi Y., Qazi J.I., Fan L., Gao H. Evodiamine sensitizes U87 glioblastoma cells to trail via the death receptor pathway. Mol. Med. Rep. 2015;11:257–262. doi: 10.3892/mmr.2014.2705. [DOI] [PubMed] [Google Scholar]
- 91.Feng Q., Liang S., Jia H., Stadlmayr A., Tang L., Lan Z., Zhang D., Xia H., Xu X., Jie Z. Gut microbiome development along the colorectal adenoma–carcinoma sequence. Nat. Commun. 2015;6:6528. doi: 10.1038/ncomms7528. [DOI] [PubMed] [Google Scholar]
- 92.Yang Z.G., Chen A., Liu B. Antiproliferation and apoptosis induced by evodiamine in human colorectal carcinoma cells (COLO-205) Chem. Biodivers. 2010;6:924–933. doi: 10.1002/cbdv.200800256. [DOI] [PubMed] [Google Scholar]
- 93.Huang C., Liu H., Gong X.L., Wu L.Y., Wen B. Effect of evodiamine and berberine on the interaction between dnmts and target micrornas during malignant transformation of the colon by tgf-beta1. Oncol. Rep. 2017;37:1637–1645. doi: 10.3892/or.2017.5379. [DOI] [PubMed] [Google Scholar]
- 94.Liu H., Huang C., Wu L., Wen B. Effect of evodiamine and berberine on miR-429 as an oncogene in human colorectal cancer. OncoTargets Ther. 2016;9:4121–4127. doi: 10.2147/OTT.S104729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Chang J., Wen B., Wang R., Chen W., Wang J. Effects of evodiamine and berberine hydrochloride on cell apoptosis and expression of hTERT-mRNA of human colorectal cancer cell line HT-29. Pharmacol. Clin. Chin. Mater. Med. 2008;19:191–194. [Google Scholar]
- 96.Chang J.R., Chen W.W., Wang J.H. Effects of evodiamine and berberine hydrochloride on telomerase activity of human colorectal cancer cell line HT29. Liaoning J. Tradit. Chin. Med. 2011;38:1326–1329. [Google Scholar]
- 97.Wang Y.H., Zhou Z.H. Regulatory effect of evodiamine on the malignant biological behaviors and wnt/β-catenin signaling pathway of colorectal cancer cell lines HT29. J. Hainan Med. Univ. 2016;22:729–731. [Google Scholar]
- 98.Chien C.C., Wu M.S., Shen S.C., Ko C.H., Chen C.H., Yang L.L., Chen Y.C. Activation of JNK contributes to evodiamine-induced apoptosis and G2/M arrest in human colorectal carcinoma cells: A structure-activity study of evodiamine. PLoS ONE. 2014;9:e99729. doi: 10.1371/journal.pone.0099729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Zhang C., Fan X., Xu X., Yang X., Wang X., Liang H.P. Evodiamine induces caspase-dependent apoptosis and S phase arrest in human colon lovo cells. Anti-Cancer Drugs. 2010;21:766–776. doi: 10.1097/CAD.0b013e32833d26a9. [DOI] [PubMed] [Google Scholar]
- 100.Li J.L., Han S., Fan X., Wang X., Zhang C., Liang H.P., Yang W.J. The effects of evodiamine on autophagy in human colon adenocarcinoma lovo cells. Chin. J. Gen. Surg. 2011;26:41–44. [Google Scholar]
- 101.Ogasawara M., Matsubara T., Suzuki H. Inhibitory effects of evodiamine on in vitro invasion and experimental lung metastasis of murine colon cancer cells. Biol. Pharm. Bull. 2001;24:917–920. doi: 10.1248/bpb.24.917. [DOI] [PubMed] [Google Scholar]
- 102.Zhao L., Liao K., Li K., Li J. Modulatory effect of evodiamine on JAK2/STAT3 signal pathway in HCT-116 cells. Chin. Pharm. Bull. 2015;1398:1394–1397. [Google Scholar]
- 103.Zhao L.C., Li J., Liao K., Luo N., Shi Q.Q., Feng Z.Q., Chen D.L. Evodiamine induces apoptosis and inhibits migration of HCT-116 human colorectal cancer cells. Int. J. Mol. Sci. 2015;16:27411–27421. doi: 10.3390/ijms161126031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Zhou Y., Fan Y., Zhang Y. Effects of evodiamine on invasion and midkine expression of human colon cancer cell. Int. J. Surg. 2009;36:742–744. [Google Scholar]
- 105.Sui H., Zhou L.H., Zhang Y.L., Huang J.P., Liu X., Ji Q., Fu X.L., Wen H.T., Chen Z.S., Deng W.L., et al. Evodiamine suppresses ABCG2 mediated drug resistance by inhibiting p50/p65 NF-κB pathway in colorectal cancer. J. Cell. Biochem. 2016;117:1471–1481. doi: 10.1002/jcb.25451. [DOI] [PubMed] [Google Scholar]
- 106.Shi X.P., Peng L.X., Xiong W., Xing L.H., Guo P., Wang F., Chen D.L., Jing L.I. Evodiamine suppresses proliferation of colon cancer HCT-116 cells in mice. Basic Clin. Med. 2017;37:1373–1377. [Google Scholar]
- 107.Rugge M., Fassan M., Graham D.Y. Gastric Cancer. Springer; Berlin, Germany: 2015. Epidemiology of gastric cancer; pp. 23–34. [Google Scholar]
- 108.Yue G., Wei J., Qian X., Yu L., Zou Z., Guan W., Wang H., Shen J., Liu B. Synergistic anticancer effects of polyphyllin I and evodiamine on freshly-removed human gastric tumors. PLoS ONE. 2013;8:e65164. doi: 10.1371/journal.pone.0065164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Hu C., Gao X., Han Y., Guo Q.I., Zhang K., Liu M., Wang Y., Wang J. Evodiamine sensitizes BGC-823 gastric cancer cells to radiotherapy in vitro and in vivo. Mol. Med. Rep. 2016;14:413–419. doi: 10.3892/mmr.2016.5237. [DOI] [PubMed] [Google Scholar]
- 110.Rasul A., Yu B., Zhong L., Khan M., Yang H., Ma T. Cytotoxic effect of evodiamine in SGC-7901 human gastric adenocarcinoma cells via simultaneous induction of apoptosis and autophagy. Oncol. Rep. 2012;27:1481–1487. doi: 10.3892/or.2012.1694. [DOI] [PubMed] [Google Scholar]
- 111.Liu S.P., Qian W.G., Mao W.D. Effect of evodiamine on inducing apoptosis of human gastric cancer cell line SGC-7901 in vitro. J. Int. Transl. Med. 2014;2:380–384. [Google Scholar]
- 112.Tian X.L., Zhang J., Wang X.L., Wang S., Shi Q.Y., Zhao G.H., Quan L.Q., Song Y. Effect of evodiamine on human gastric adenocarcinoma cell line (SGC-7901) J. Tradit. Chin. Med. 2011;34:115–118. [Google Scholar]
- 113.Yang L., Liu X., Wu D., Zhang M., Ran G., Bi Y., Huang H. Growth inhibition and induction of apoptosis in SGC7901 human gastric cancer cells by evodiamine. Mol. Med. Rep. 2014;9:1147–1152. doi: 10.3892/mmr.2014.1924. [DOI] [PubMed] [Google Scholar]
- 114.Zhang Y.X., Ya-Kun G.E. Study on synergetic anti-tumor effect of evodiamine and paclitaxel in SGC-7901 cells. J. Jilin Agric. Univ. 2014;36:314–318. [Google Scholar]
- 115.Shen H., Zhao S., Xu Z., Zhu L., Han Y., Ye J. Evodiamine inhibits proliferation and induces apoptosis in gastric cancer cells. Oncol. Lett. 2015;10:367–371. doi: 10.3892/ol.2015.3153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Huang Y.Z., Huang H., Zhang Y.Y., Liu X., Bi Y., Liu L. Study of evodiamine on proliferation inhibition and apoptosis induction in human gastric cancer cell line SGC-7901. Lab. Med. 2010;25:952–955. [Google Scholar]
- 117.Huang H., Zhang Y., Liu X., Li Z., Xu W., He S., Huang Y., Zhang H. Acid sphingomyelinase contributes to evodiamine-induced apoptosis in human gastric cancer SGC-7901 cells. DNA Cell Biol. 2011;30:407–412. doi: 10.1089/dna.2010.1122. [DOI] [PubMed] [Google Scholar]
- 118.Cheng Z.M., Zhou Y.J., Zhang Y.L., Qi W.D., Fan Y. Effects of evodiamine on invasion and polo-like kinase 1 expression of human gastric cancer cell. J. Jiangsu Univ. 2011;21:69–72. [Google Scholar]
- 119.Qi W.D., Zhou Y.J., Fan Y. Effects of evodiamine on invasion and proliferation in human gastric cancer cell. Shandong Med. J. 2010;34:016. [Google Scholar]
- 120.Singh S.R. Gastric cancer stem cells: A novel therapeutic target. Cancer Lett. 2013;338:110–119. doi: 10.1016/j.canlet.2013.03.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Wen Z., Feng S., Wei L., Wang Z., Hong D., Wang Q. Evodiamine, a novel inhibitor of the wnt pathway, inhibits the self-renewal of gastric cancer stem cells. Int. J. Mol. Med. 2015;36:1657–1663. doi: 10.3892/ijmm.2015.2383. [DOI] [PubMed] [Google Scholar]
- 122.Shi H.L., Wu X.J., Liu Y., Xie J.Q. Berberine counteracts enhanced IL-8 expression of AGS cells induced by evodiamine. Life Sci. 2013;93:830–839. doi: 10.1016/j.lfs.2013.09.010. [DOI] [PubMed] [Google Scholar]
- 123.Michael H., Day C.P., Yang H., Michalowski A., Lee M., Merlino G., Michael H., Day C.P., Yang H., Michalowski A. Abstract 1037: Progression from melanocytic nevi to melanoma is associated with increased genomic mutations in a uv-induced mouse model of human melanoma. Cancer Res. 2017;77:1037. doi: 10.1158/1538-7445.AM2017-1037. [DOI] [Google Scholar]
- 124.Wang C., Wang M., Tashiro S.-I., Onodera S., Ikejima T. Effect of protein kinase C on human melanoma A375-S2 cell death induced by evodiamine. Acta Pharm. Sin. 2005;40:1033–1036. [PubMed] [Google Scholar]
- 125.Wang C., Wang M.S., Onodera S., Ikejima T. Roles of SIRT1 and phosphoinositide 3-OH kinase/protein kinase C pathways in evodiamine-induced human melanoma A375-S2 cell death. J. Pharmacol. Sci. 2005;97:494–500. doi: 10.1254/jphs.FPJ04055X. [DOI] [PubMed] [Google Scholar]
- 126.Zhang Y., Wu L., Tashiro S.I., Onodera S., Ikejima T. Evodiamine induces A375-S2 cell death through two different pathways. Acta Pharm. Sin. 2003;38:650–653. [PubMed] [Google Scholar]
- 127.Zhang Y., Wu L.J., Tashiro S., Onodera S., Ikejima T. Intracellular regulation of evodiamine-induced A375-S2 cell death. Biol. Pharm. Bull. 2003;26:1543–1547. doi: 10.1248/bpb.26.1543. [DOI] [PubMed] [Google Scholar]
- 128.Yang J., Wu L.J., Tashino S., Onodera S., Ikejima T. Critical roles of reactive oxygen species in mitochondrial permeability transition in mediating evodiamine-induced human melanoma A375-S2 cell apoptosis. Free Radic. Res. 2007;41:1099–1108. doi: 10.1080/10715760701499356. [DOI] [PubMed] [Google Scholar]
- 129.Yang J., Wu L.J., Tashiro S., Onodera S., Ikejima T. Nitric oxide activated by p38 and NF-κB facilitates apoptosis and cell cycle arrest under oxidative stress in evodiamine-treated human melanoma A375-S2 cells. Free Radic. Res. 2008;42:1–11. doi: 10.1080/10715760701762407. [DOI] [PubMed] [Google Scholar]
- 130.Wang C., Wang M.S., Onodera S., Ikejima T. Evodiamine induced human melanoma A375-S2 cell death partially through interleukin 1 mediated pathway. Chin. Pharmacol. Bull. 2005;28:984–989. doi: 10.1248/bpb.28.984. [DOI] [PubMed] [Google Scholar]
- 131.Wang C., Li S., Wang M.W. Evodiamine-induced human melanoma A375-S2 cell death was mediated by PI3K/Akt/caspase and Fas-L/NF-κB signaling pathways and augmented by ubiquitin-proteasome inhibition. Toxicol. In Vitro. 2010;24:898–904. doi: 10.1016/j.tiv.2009.11.019. [DOI] [PubMed] [Google Scholar]
- 132.Liu C., Yin L., Chen J., Chen J. The apoptotic effect of shikonin on human papillary thyroid carcinoma cells through mitochondrial pathway. Tumor Biol. 2014;35:1791–1798. doi: 10.1007/s13277-013-1238-5. [DOI] [PubMed] [Google Scholar]
- 133.Lv Z., Zhao D., Liu R., Guo J., Lin Y., Zhang M. Evodiamine inhibits proliferation of human papillary thyroid cancer cell line k1 by regulating of PI3K/Akt signaling pathway. Int. J. Clin. Exp. Med. 2016;9:15216–15225. [Google Scholar]
- 134.Chen M.C., Yu C.H., Wang S.W., Pu H.F., Kan S.F., Lin L.C., Chi C.W., Ho L.L., Lee C.H., Wang P.S. Anti-proliferative effects of evodiamine on human thyroid cancer cell line ARO. J. Cell Biochem. 2010;110:1495–1503. doi: 10.1002/jcb.22716. [DOI] [PubMed] [Google Scholar]
- 135.Lee Y.R., Lu C.H., Chang Y.S., Chen S.H., Liu Y.W. Evodiamine inhibits human thyroid cancer cells in vitro and in vivo. Endocrine. 2015;37:EP902. doi: 10.1530/endoabs.37.EP902. [DOI] [Google Scholar]
- 136.Yu H.I., Chou H.C., Su Y.C., Lin L.H., Lu C.H., Chuang H.H., Tsai Y.T., Liao E.C., Wei Y.S., Yang Y.T., et al. Proteomic analysis of evodiamine-induced cytotoxicity in thyroid cancer cells. J. Pharm. Biomed. Anal. 2018;160:344–350. doi: 10.1016/j.jpba.2018.08.008. [DOI] [PubMed] [Google Scholar]
- 137.Chua M.L.K., Wee J.T.S., Hui E.P., Chan A.T.C. Nasopharyngeal carcinoma. Lancet. 2016;387:1012–1024. doi: 10.1016/S0140-6736(15)00055-0. [DOI] [PubMed] [Google Scholar]
- 138.Peng X., Zhang Q., Zeng Y., Li J., Wang L., Ai P. Evodiamine inhibits the migration and invasion of nasopharyngeal carcinoma cells in vitro via repressing MMP-2 expression. Cancer Chemother. Pharmacol. 2015;76:1173–1184. doi: 10.1007/s00280-015-2902-9. [DOI] [PubMed] [Google Scholar]
- 139.Xu Q., Krause M., Samoylenko A., Vainio S. Wnt signaling in renal cell carcinoma. Cancers. 2016;8:57. doi: 10.3390/cancers8060057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Wu W.S., Chien C.C., Chen Y.C., Chiu W.T. Protein kinase RNA-like endoplasmic reticulum kinase-mediated Bcl-2 protein phosphorylation contributes to evodiamine-induced apoptosis of human renal cell carcinoma cells. PLoS ONE. 2016;11:e0160484. doi: 10.1371/journal.pone.0160484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Yuan X.L., Zhang P., Liu X.M., Du Y.M., Hou X.D., Cheng S., Zhang Z.F. Cytological assessments and transcriptome profiling demonstrate that evodiamine inhibits growth and induces apoptosis in a renal carcinoma cell line. Sci. Rep. 2017;7:12572. doi: 10.1038/s41598-017-12918-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Siegel R.L., Miller K.D., Jemal A. Cancer statistics, 2015. CA Cancer J. Clin. 2015;65:5–29. doi: 10.3322/caac.21254. [DOI] [PubMed] [Google Scholar]
- 143.Lopez-Beltran A. Bladder cancer: Clinical and pathological profile. Scand. J. Urol. Nephrol. Suppl. 2008;42:95–109. doi: 10.1080/03008880802325226. [DOI] [PubMed] [Google Scholar]
- 144.Zhang T., Qu S., Shi Q., He D., Jin X. Evodiamine induces apoptosis and enhances TRAIL-induced apoptosis in human bladder cancer cells through mTOR/S6K1-mediated downregulation of Mcl-1. Int. J. Mol. Sci. 2014;15:3154–3171. doi: 10.3390/ijms15023154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Shi C.S., Li J.M., Chin C.C., Kuo Y.H., Lee Y.R., Huang Y.C. Evodiamine induces cell growth arrest, apoptosis and suppresses tumorigenesis in human urothelial cell carcinoma cells. Anticancer Res. 2017;37:1149–1159. doi: 10.21873/anticanres.11428. [DOI] [PubMed] [Google Scholar]
- 146.Maisonneuve P., Lowenfels A. Epidemiology and Prospects for Prevention of Pancreatic Cancer. 2nd ed. Springer; New York, NY, USA: 2016. pp. 3–25. [Google Scholar]
- 147.Khan M., Qazi J.I., Rasul A., Zheng Y., Ma T. Evodiamine induces apoptosis in pancreatic carcinoma PANC-1 cells via NFκB inhibition. Bangladesh J. Pharmacol. 2013;8 doi: 10.3329/bjp.v8i1.12802. [DOI] [Google Scholar]
- 148.Wei W.T., Chen H., Wang Z.H., Ni Z.L., Liu H.B., Tong H.F., Guo H.C., Liu D.L., Lin S.Z. Enhanced antitumor efficacy of gemcitabine by evodiamine on pancreatic cancer via regulating PI3K/Akt pathway. Int. J. Biol. Sci. 2012;8:1–14. doi: 10.7150/ijbs.8.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Varambally S., Dhanasekaran S.M., Zhou M., Barrette T.R., Kumar-Sinha C., Sanda M.G., Ghosh D., Pienta K.J., Sewalt R.G., Otte A.P. The polycomb group protein ezh2 is involved in progression of prostate cancer. Nature. 2002;419:624–629. doi: 10.1038/nature01075. [DOI] [PubMed] [Google Scholar]
- 150.Huang D.M., Guh J.H., Huang Y.T., Chueh S.C., Chiang P.C., Teng C.M. Induction of mitotic arrest and apoptosis in human prostate cancer PC-3 cells by evodiamine. J. Urol. 2005;173:256–261. doi: 10.1097/01.ju.0000141587.72429.e3. [DOI] [PubMed] [Google Scholar]
- 151.Kan S.F., Yu C.H., Pu H.F., Hsu J.M., Chen M.J., Wang P.S. Anti-proliferative effects of evodiamine on human prostate cancer cell lines DU145 and PC-3. J. Cell Biochem. 2007;101:44–56. doi: 10.1002/jcb.21036. [DOI] [PubMed] [Google Scholar]
- 152.Kan S.F., Huang W.J., Lin L.C., Wang P.S. Inhibitory effects of evodiamine on the growth of human prostate cancer cell line LNCaP. Int. J. Cancer. 2004;110:641–651. doi: 10.1002/ijc.20138. [DOI] [PubMed] [Google Scholar]
- 153.Poon V.I., Roth M., Piperdi S., Geller D., Gill J., Rudzinski E.R., Hawkins D.S., Gorlick R. Ganglioside GD2 expression is maintained upon recurrence in patients with osteosarcoma. Clin. Sarcoma Res. 2015;5:4. doi: 10.1186/s13569-014-0020-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Meng Z.J., Wu N., Liu Y., Shu K.J., Zou X., Zhang R.X., Pi C.J., He B.C., Ke Z.Y., Chen L. Evodiamine inhibits the proliferation of human osteosarcoma cells by blocking PI3K/Akt signaling. Oncol. Rep. 2015;34:1388–1396. doi: 10.3892/or.2015.4084. [DOI] [PubMed] [Google Scholar]
- 155.Bai X., Meng H., Ma L., Guo A. Inhibitory effects of evodiamine on human osteosarcoma cell proliferation and apoptosis. Oncol. Lett. 2015;9:801–805. doi: 10.3892/ol.2014.2791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Zhou Y., Hu J. Evodiamine induces apoptosis, G2/M cell cycle arrest, and inhibition of cell migration and invasion in human osteosarcoma cells via Raf/MEK/ERK signalling pathway. Med. Sci. Monit. 2018;24:5874–5880. doi: 10.12659/MSM.909682. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 157.Yuan Y., Li S., Wang Z., Shen H., Li W., Wang W. Evodiamine inhibits apoptosis of human osteosarcoma MG-63 cells by blocking wnt/β-catenin signaling. Int. J. Oncol. 2017;44:86–90. [Google Scholar]
- 158.Sankaranarayanan R., Ramadas K., Thomas G., Muwonge R., Thara S., Mathew B., Rajan B. Effect of screening on oral cancer mortality in kerala, india: A cluster-randomised controlled trial. Lancet. 2005;365:1927–1933. doi: 10.1016/S0140-6736(05)66658-5. [DOI] [PubMed] [Google Scholar]
- 159.Sachita K., Kim Y., Yu H.J., Cho S.D., Lee J.S. In vitro assessment of the anticancer potential of evodiamine in human oral cancer cell lines. Phytother. Res. 2015;29:1145–1151. doi: 10.1002/ptr.5359. [DOI] [PubMed] [Google Scholar]
- 160.Liu B., Mitani Y., Rao X., Zafereo M., Zhang J., Zhang J., Futreal P.A., Lozano G., El-Naggar A.K. Spatio-temporal genomic heterogeneity, phylogeny, and metastatic evolution in salivary adenoid cystic carcinoma. J. Natl. Cancer Inst. 2017;109 doi: 10.1093/jnci/djx033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Chen J.Y., Di Y.U., Zeng D.S., Hong H.P. Regulatory impact of evodiamine on salivary adenoid cystic carcinoma SACC-M cell growth. Chin. Arch. Tradit. Chin. Med. 2012;5:1127–1129. [Google Scholar]
- 162.Wang Z.X., Xiang J.C., Wang M., Ma J.T., Wu Y.D., Wu A.X. One-pot total synthesis of evodiamine and its analogues through a continuous biscyclization reaction. Org. Lett. 2018;20:6380–6383. doi: 10.1021/acs.orglett.8b02667. [DOI] [PubMed] [Google Scholar]
- 163.Tian K., Xu F., Gao X., Han T., Li J., Pan H., Zang L., Li D., Li Z., Uchita T., et al. Nitric oxide-releasing derivatives of brefeldin A as potent and highly selective anticancer agents. Eur. J. Med. Chem. 2017;136:131–143. doi: 10.1016/j.ejmech.2017.05.018. [DOI] [PubMed] [Google Scholar]
- 164.Han T., Li J., Xue J., Li H., Xu F., Cheng K., Li D., Li Z., Gao M., Hua H. Scutellarin derivatives as apoptosis inducers: Design, synthesis and biological evaluation. Eur. J. Med. Chem. 2017;135:270–281. doi: 10.1016/j.ejmech.2017.03.020. [DOI] [PubMed] [Google Scholar]
- 165.Yao H., Liu J., Xu S., Zhu Z., Xu J. The structural modification of natural products for novel drug discovery. Expert Opin. Drug Discov. 2017;12:121–140. doi: 10.1080/17460441.2016.1272757. [DOI] [PubMed] [Google Scholar]
- 166.Gao X., Li J., Wang M., Xu S., Liu W., Zang L., Li Z., Hua H., Xu J., Li D. Novel enmein-type diterpenoid hybrids coupled with nitrogen mustards: Synthesis of promising candidates for anticancer therapeutics. Eur. J. Med. Chem. 2018;146:588–598. doi: 10.1016/j.ejmech.2018.01.069. [DOI] [PubMed] [Google Scholar]
- 167.Han T., Tian K., Pan H., Liu Y., Xu F., Li Z., Uchita T., Gao M., Hua H., Li D. Novel hybrids of brefeldin A and nitrogen mustards with improved antiproliferative selectivity: Design, synthesis and antitumor biological evaluation. Eur. J. Med. Chem. 2018;150:53–63. doi: 10.1016/j.ejmech.2018.02.088. [DOI] [PubMed] [Google Scholar]
- 168.Dong G., Sheng C., Wang S., Miao Z., Yao J., Zhang W. Selection of evodiamine as a novel topoisomerase I inhibitor by structure-based virtual screening and hit optimization of evodiamine derivatives as antitumor agents. J. Med. Chem. 2010;53:7521–7531. doi: 10.1021/jm100387d. [DOI] [PubMed] [Google Scholar]
- 169.Dong G., Wang S., Miao Z., Yao J., Zhang Y., Guo Z., Zhang W., Sheng C. New tricks for an old natural product: Discovery of highly potent evodiamine derivatives as novel antitumor agents by systemic structure-activity relationship analysis and biological evaluations. J. Med. Chem. 2012;55:7593–7613. doi: 10.1021/jm300605m. [DOI] [PubMed] [Google Scholar]
- 170.Song S., Chen Z., Li S., Huang Y., Wan Y., Song H. Design, synthesis and evaluation of N13-substituted evodiamine derivatives against human cancer cell lines. Molecules. 2013;18:15750–15768. doi: 10.3390/molecules181215750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Nguyen N.V., Lee K.R., Lee Y.J., Choi S., Kang J.S., Mar W., Kim K.H. Chiral high-performance liquid chromatographic separation of evodiamine enantiomers and rutaecarpine, isolated from Evodiae fructus. J. Pharm. Biomed. Anal. 2013;81–82:151–159. doi: 10.1016/j.jpba.2013.04.018. [DOI] [PubMed] [Google Scholar]
- 172.Christodoulou M.S., Sacchetti A., Ronchetti V., Caufin S., Silvani A., Lesma G., Fontana G., Minicone F., Riva B., Ventura M., et al. Quinazolinecarboline alkaloid evodiamine as scaffold for targeting topoisomerase I and sirtuins. Bioorg. Med. Chem. 2013;21:6920–6928. doi: 10.1016/j.bmc.2013.09.030. [DOI] [PubMed] [Google Scholar]
- 173.De Petrocellis L., Schiano Moriello A., Fontana G., Sacchetti A., Passarella D., Appendino G., Di Marzo V. Effect of chirality and lipophilicity in the functional activity of evodiamine and its analogues at TRPV1 channels. Br. J. Pharmacol. 2014;171:2608–2620. doi: 10.1111/bph.12320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Ivanova B., Spiteller M. Evodiamine and rutaecarpine alkaloids as highly selective transient receptor potential vanilloid 1 agonists. Int. J. Biol. Macromol. 2014;65:314–324. doi: 10.1016/j.ijbiomac.2014.01.059. [DOI] [PubMed] [Google Scholar]
- 175.Liang C., Xia J., Song H., Zhou Z., Xue Y., Yao Q. Synthesis, in vitro and in vivo antitumor activity and docking studies of new evodiamine derivatives. J. Chem. Pharm. Res. 2014;6:1161–1171. [Google Scholar]
- 176.Wang S., Fang K., Dong G., Chen S., Liu N., Miao Z., Yao J., Li J., Zhang W., Sheng C. Scaffold diversity inspired by the natural product evodiamine: Discovery of highly potent and multitargeting antitumor agents. J. Med. Chem. 2015;58:6678–6696. doi: 10.1021/acs.jmedchem.5b00910. [DOI] [PubMed] [Google Scholar]
- 177.Zhao N., Tian K.T., Cheng K.G., Han T., Hu X., Li D.H., Li Z.L., Hua H.M. Antiproliferative activity and apoptosis inducing effects of nitric oxide donating derivatives of evodiamine. Bioorg. Med. Chem. 2016;24:2971–2978. doi: 10.1016/j.bmc.2016.05.001. [DOI] [PubMed] [Google Scholar]
- 178.Hu X., Wang Y., Xue J., Han T., Jiao R., Li Z., Liu W., Xu F., Hua H., Li D. Design and synthesis of novel nitrogen mustard-evodiamine hybrids with selective antiproliferative activity. Bioorg. Med. Chem. Lett. 2017;27:4989–4993. doi: 10.1016/j.bmcl.2017.10.014. [DOI] [PubMed] [Google Scholar]
- 179.Hu X., Jiao R., Li H., Wang X., Lyu H., Gao X., Xu F., Li Z., Hua H., Li D. Antiproliferative hydrogen sulfide releasing evodiamine derivatives and their apoptosis inducing properties. Eur. J. Med. Chem. 2018;151:376–388. doi: 10.1016/j.ejmech.2018.04.009. [DOI] [PubMed] [Google Scholar]
- 180.Hu J., Sun L., Zhao D., Zhang L., Ye M., Tan Q., Fang C., Wang H., Zhang J. Supermolecular evodiamine loaded water-in-oil nanoemulsions: Enhanced physicochemical and biological characteristics. Eur. J. Pharm. Biopharm. 2014;88:556–564. doi: 10.1016/j.ejpb.2014.06.007. [DOI] [PubMed] [Google Scholar]