

Abstract
An ability to visualize HCN in mitochondria in real time may permit additional insights into the critical toxicological and physiological roles this classic toxin plays in living organisms. Herein, we report a mitochondria-specific coumarin pyrrolidinium-derived fluorescence probe (MRP1) that permits the real-time ratiometric imaging of HCN in living cells. The response is specific, sensitive (detection limit is ca. 65.6 nM), rapid (within 1 s), and reversible. Probe MRP1 contains a benzyl chloride subunit designed to enhance retention within the mitochondria under conditions where the mitochondria membrane potential is eliminated. It has proved effective in visualizing different concentrations of exogenous HCN in the mitochondria of HepG2 cells, as well as the imaging of endogenous HCN in the mitochondria of PC12 cells and within neurons. Fluctuations in HCN levels arising from the intracellular generation of HCN could be readily detected.
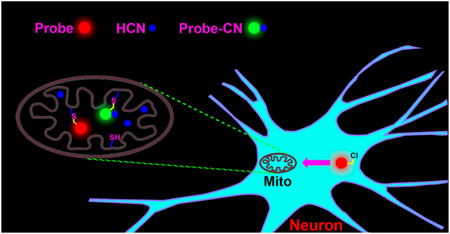
Graphical Abstract
INTRODUCTION
The cyanide anion (CN−) is a recognized toxin.1 For humans, blood cyanide levels higher than 20 μM are considered toxic.2 However, cyanide is widespread in the modern world, and cyanide exposure from gold mining, electroplating, metallurgy,3a consumption of certain foods,3b bacterial cyanogenesis,3c uncontrolled fires,3d cigarette smoke,3e vehicle exhaust,3f and from pharmaceutical byproducts3g are potential concerns. At the subcellular level, the mitochondria are recognized as being the primary site of cyanide toxicity. This is because cyanide inactivates mitochondrial cytochrome C oxidase and halts cellular respiration.4 It also induces production of reactive oxygen species, including hydroxyl radicals (•OH) within mitochondria.5 This leads to the oxidative damage to mitochondrial DNA and further results in mitochondrial dysfunction. This dysfunction is linked to a variety of diseases, including Alzheimer’s disease,6a Huntington’s disease,6b Parkinson’s disease,6c diabetes,6d and cancer.6e
On the other hand, despite its high toxicity, growing evidence suggests that the cyanide anion is generated in specific cells and plays important physiological roles within the human body.7–9 At the physiological pH of 7.4, cyanide exists predominantly as hydrogen cyanide (HCN) due to its pKa value of 9.3. In leukocytes, HCN is produced by the myeloperoxidase-mediated oxidative chlorination of glycine or N-terminal glycyl peptides during phagocytosis, and is used as a defense against bacteria, fungi, and other pathogens.7 It has also been suggested that HCN is generated within neurons upon the activation of opioid receptors by endogenous or exogenous opioids.8 The HCN generated in this way has been proposed as a novel neuromodulator within the central and peripheral nervous system in loose analogy to nitric monoxide (NO), carbon monoxide (CO), and hydrogen sulfide (H2S).9 Unfortunately, experimental support for the putative role of HCN in neurons remains elusive. Mitochondria have been suggested as being the main locus for the production of HCN within the neurons;8c however, this is far from established. Effective HCN probes that operate within mitochondria may allow deeper insights into the critical toxicological and physiological roles of HCN within living organisms. As detailed below, we have now prepared a probe that appears promising in this regard.
Fluorescent probes have received considerable attention due to the high sensitivity, fast response time, technical simplicity, and applicability to biological milieus.10 In combination with laser scanning confocal microscope (LSCM), appropriately designed fluorescent probes can be used to sense important species in subcellular organelles in a noninvasive manner.11 Currently, a large number of fluorescent probes for cyanide are known. As a general rule, these systems rely on the unique nucleophilic reactivity of the cyanide anion to trigger a change in the fluorescent response.12 Fluorescent probes for the cyanide anion that are based on its interactions with metal ions,13 H+,14 and boronic acid derivatives15 are also known. To date, cyanide anion probes have been used to sense endogenous cyanide in plant samples.13a,16 However, to our knowledge, no cyanide anion probe has been used successfully to monitor endogenous HCN levels in mitochondria.
To be effective at the mitochondrial level, we suggest that a fluorescent HCN probe needs to meet the following prerequisites: (a) It should display a fast response upon exposure to HCN. (b) It should give rise to a so-called ratiometric output so as to reduce potential artifacts associated with, for example, photobleaching, probe distribution, or changes in excitation intensity.17 (c) The induced changes should be reversible so as to allow fluctuations in HCN levels in cells and organelles to be monitored in real time. (d) It should possess a high inherent fluorescence quantum yield (Φf) and be effective at low concentrations (25−500 nM). (e) It should be retained in the mitochondria when the mitochondria membrane potential is eliminated. The latter requirement is particularly germane to the design of an HCN probe. The negative mitochondrial membrane potential (ΔΨ) typically allows the targeting of positively charged species to the mitochondria.11 However, high concentrations of HCN can lead to a loss in the mitochondrial membrane potential,18 leading to diffusion of erstwhile targeted species into the cytoplasm. In such instances, the further study of mitochondria-related changes in HCN concentrations would be precluded.
RESULTS AND DISCUSSION
Rational Design of the Probe.
Bearing the above considerations in mind, a ratiometric fluorescent probe for visualizing HCN within the mitochondria of living cells was designed as shown in Scheme 1a. Full synthetic details are provided in the Supporting Information. A 7-diethylaminocoumarin dye (green color) was selected as the fluorophore, while a 1-methylenepyrrolidinium subunit (pink color) was used as the reactivity-based recognition site for HCN. In preliminary studies, we found that the 1-methylenepyrrolidinium group can rapidly, specifically, and reversibly react with HCN. By tethering a 7-diethylamino-coumarin moiety to the 1-methylenepyrrolidinium subunit through an ethylene linker, we expected to have a system that would give rise to a ratiometric fluorescence response to HCN as the result of modulating the intramolecular charge-transfer (ICT)19 interactions between the electron-donating coumarin and electron-withdrawing pyrrolidinium groups (referred to as EDG and EWG, respectively). Perturbing the ICT interactions between linked EDG and EWGs can have a significant effect on the fluorescence emission.20,21 In an effort to study this effect in the present instance and optimize the fluorescence quantum yield of the probe, a 4-substituted phenyl group (blue color) was tethered to the electron-deficient 1-methylenepyrrolidinium group. We hypothesized that changing the substituents at the 4-position of the phenyl group might allow the extent of the ICT interaction to be fine-tuned. Thus, compounds 1a−1e with different electron-donating or electron-withdrawing substituents at the 4-position of the appended phenyl group were designed and synthesized (Scheme 1a). Predicative studies of these HCN probes, discussed below, then allowed for the preparation of a mitochondrial probe, MRP1, suitable for use in vitro.
Scheme 1.

(a) Components Making Up the Present Series of HCN Probes;a and (b) Proposed Basis for the Sensing Response of MRP1 and Related HCN Probes
aNote that different substituents are present at the 4-position of the phenyl group.
As a first step toward understanding the HCN response function of compounds 1a−1e, their optical properties were investigated (Figure S1 and Table S1). Compound 1a exhibited a dominant absorption feature centered at 521 nm, which is ascribed to an ICT transition. In the case of the systems bearing electron-withdrawing substituents (Cl, CN) at the 4-position of the phenyl group, that is, 1d and 1e, slightly red-shifted absorption bands were observed at 524 and 530 nm, respectively. In contrast, more substantial blue shifts were seen upon the introduction of electron-donating groups (i.e., OH, diethylamino); in fact, absorption features at 515 and 513 nm were seen for compounds 1b and 1c, respectively.
Upon excitation at 521 nm, compound 1a displayed a fluorescence emission band centered at 602 nm with an associated quantum yield (Φf) of 0.184 (Figure S1b and Table S1). As compared to 1a, the fluorescence quantum yields of 1d (Φf = 0.166) and 1e (Φf = 0.062) were reduced. In contrast, compound 1b had a higher fluorescence quantum yield (Φf =0.285). On the other hand, compound 1c displayed little appreciable fluorescence intensity (Φf = 0.002).
To obtain insights into the optical properties of compounds 1a−1e, time-dependent density functional theory (TD-DFT) calculations were carried out. The electron densities in the HOMO and LUMO of these compounds in the ground state and first excited singlet state are shown in Figure S2. On this basis, it is concluded that the HOMO → LUMO transitions for compounds 1a, 1b, 1d, and 1e involve electron density redistributions from the EDG to EWG. In other words, ICT is predicated upon excitation. In stark contrast, in the first excited singlet state of compound 1c, the HOMO → LUMO transition involves an electron density redistribution from the 4-diethylamino-phenyl group to the 7-diethylaminocoumarin moiety, which acts as an electron acceptor, not donor.
The dihedral angles between the 7-diethylamino-coumarin ring and the 4-substituted phenyl ring of compounds 1a−1e in the optimized geometry are given in Table S2 and Figure S3. In the ground state (S0), the dihedral angles between the coumarin ring and the phenyl ring in compounds 1a, 1b, 1d, and 1e were approximately 60°. In the first excited singlet state (S1), these dihedral angles decrease to around 50°. The dihedral angle of compound 1c in its ground (S0) state is 57°, which is similar to that calculated for 1a, 1b, 1d, and 1e, whereas in the S1 state, the corresponding dihedral angle for 1c increases to 88°. It was also found that the fluorescence intensity of 1c increases with increasing solvent viscosity (Figure S4). Taken in concert, these findings lead us to suggest that the fluorescence quenching of compound 1c is best ascribed to a twisted intramolecular charge transfer (TICT)22 from the 4-diethylamino phenyl group to the 7-diethylaminocoumarin.
The charge-transfer process underlying the presumed ICT may be described in terms of the charge-transfer parameter fCT = ΔqD − ΔqA, where ΔqD and ΔqA denote the changes in the charge of the donor and the acceptor, respectively, upon excitation from the ground state to the lowest excited state.23 As shown in Table S3, in the S1 state, compound 1a has an fCT around 0.45. By varying the substituents, the fCT value could be increased to 0.57 in the case of 1e. Likewise, it decreases to about 0.39 in 1b. Thus, the ICT strength can be fine-tuned by varying the substituents at the 4-position of the phenyl group.
Compound 1b displayed the highest fluorescence quantum yield within the series 1a−1e. The key para-oxygen atom was thus retained in designing a putative ratiometric fluorescent probe for HCN, MRP1 (Scheme 1a). Probe MRP1 differs from 1b in that an additional benzyl chloride group (orange color) is incorporated into the framework. This group can react with free thiol groups present in mitochondrial proteins and thus serve potentially to immobilize the probe within mitochondria, even in instances when the mitochondria membrane potential is eliminated.24
Spectral Response of Probe MRP1 to HCN.
The ability of probe MRP1 to react with HCN was evaluated in 20 mM potassium phosphate buffer/acetonitrile (pH 7.4, 1:3 v/v) at room temperature. In its free form, probe MRP1 exhibited a fluorescence emission feature centered at 599 nm (Φf = 0.261) under these conditions. Upon addition of cyanide (as its KCN salt), the fluorescence emission of MRP1 at 599 nm gradually decreased, while a new emission feature at 490 nm was seen to grow in (Figure 1a). An isoemission point was clearly visible at 552 nm during this addition process. The emission ratio (I490/I599), an indicator of a ratiometric response, was found to be linearly proportional to the concentration of cyanide anion over the 0−35 μM concentration range (Figure S5). On the basis of a signal-to-noise (S N1−) ratio of 3, a detection limit of 65.6 nM was calculated; this is much lower than the concentration of HCN typically found in the brain of healthy individuals (2−10 μM).9b The absorption maximum of probe MRP1 was also found to undergo a blue shift (from 516 to 411 nm) upon treatment with cyanide (Figure S6). The ratiometric fluorescence response of MRP1 to cyanide was also evaluated at different pH values. On the basis of the results obtained (cf., Figure S7), it is concluded that MRP1 can be used to detect cyanide over the pH 5−8 range. The stability of MRP1 in aqueous solution was tested by means of fluorescence spectral analyses. In various mixtures of acetonitrile and buffer, no change in the fluorescence signature was seen over the course of 24 h, leading us to infer that MRP1 is stable over at least such a time scale (Figure S8). The effect of the relative concentration of CH3CN was also investigated (Figure S9); on this basis, it was determined that MRP1 can be used to sense cyanide even in aqueous solutions containing only 30% acetonitrile.
Figure 1.

(a) Changes in the fluorescence emission spectra of MRP1 (1 μM) seen upon the addition of increasing quantities of cyanide (0− 60 μM as its KCN salt) at room temperature (λex = 467 nm). (b) Time-dependent changes in the fluorescence emission intensities recorded at 490 and 599 nm observed upon treatment of MRP1 (1 μM) with cyanide (60 μM). (c) Changes in the fluorescence intensity of MRP1 (1 μM) seen upon the sequential addition of first cyanide (60 μM) and then AgNO3 (60 μM) (λex = 467 nm, λem = 599 nm); (■) MRP1, (blue ▲) MRP1 + CN−, and (red ●) MRP1 + CN− + AgNO3. (d) Ratiometric fluorescence response of MRP1 (1 μM) seen upon exposure to 60 μM of various putative analytes. Inset: Photos of MRP1 (1 μM) taken in the presence of various species (60 μM) under the illumination of a hand-held UV lamp. All studies were carried out in 20 mM potassium phosphate buffer/acetonitrile, pH 7.4, 1:3 v/v.
Time-dependent studies of the fluorescence changes produced when MRP1 is treated with cyanide revealed a response that occurs within ≤1 s (Figure 1b). The fast response time is considered beneficial for a probe designed to monitor HCN concentrations in real time. After reacting with cyanide, probe MRP1 was treated with AgNO3 (Figure 1c). Sequential additions of cyanide and silver(I) led to a turning off and on of the 599 nm fluorescent feature. On this basis, we conclude that MRP1 may allow for the reversible monitoring of HCN fluctuations in living cells.
The response of MRP1 toward other putative analytes was also explored. When MRP1 (1 μM) was treated with either of the other physiologically relevant anions (e.g., F−, Cl−, Br−, I−, CH3COO−, HSO3−, NO3−, ClO4−, and N3−; studied as their Na+, Li+, or K+ salts) or typical biological molecules (glucose, Gly, and Cys; all at 60 μM), no significant change was observed in the (I490/I599) emission ratio (Figure 1d). Only when cyanide was added was a noticeable response observed. This response toward cyanide was retained in the presence of other relevant species, most of which produced no appreciable or only minimal interference (Figure S10). This selectivity was apparent to the naked eye. Under conditions of illumination using a hand-held UV lamp, cyanide produced a distinctive color signature with MRP1 not seen in the case of other putative analytes (Figure 1d).
Compound 1a displayed a similar ratiometric fluorescence response to cyanide (Figure S11). However, its structure is much simpler than that of MRP1. Therefore, for clarity, compound 1a was used to investigate the sensing reaction mechanism. The reaction product of 1a with cyanide was isolated and characterized. As inferred from 1H NMR and 13C NMR spectral studies and mass spectrometric analyses (Figures S12−S14), the optical response seen in the case of 1a and by extension MRP1 reflects the nucleophilic addition of cyanide anion to the 1-methylenepyrrolidinium moiety present in these two congeneric probes. This addition interferes with an efficient ICT from the EDG to the EWG, which, in turn, results in a blue shift in both the emission and the absorption spectral features (Scheme 1b). Support for this conclusion came from TD-DFT calculations (Figures S15 and S16).
Fluorescence Imaging of HCN in the Mitochondria of Living Cells.
To assess the mitochondrial staining ability of MRP1, colocalization experiments were carried out using Mitotracker Deep Red (MTDR),25 a commercially available mitochondrial dye. In these studies, HepG2 cells were coincubated with MRP1 (0.5 μM) and MTDR (0.5 μM) at 37 °C for 15 min. As can be seen by inspection of Figure 2c, a clear mitochondria profile with strong near-infrared (NIR) fluorescence was observed in the NIR channel, which was ascribed to MTDR. A similar mitochondria profile was observed in the red channel, which is largely reflective of probe MRP1 (Figure 2b). The MRP1 response coincides well with that of MTDR (Figure 2d) as reflected in a Pearson’s colocalization coefficient of 0.94. Compound 1b was also studied and likewise found to display mitochondria-specific staining (Figure 2e–h). Considered in concert, these results provide support for the notion that both MRP1 and 1b act as organelle-specific probes that target the mitochondria.
Figure 2.

Confocal image of HepG2 cells costained with either MRP1 and MTDR or 1b and MTDR. (a−c) Bright field, red (λem = 575−650 nm), and NIR (λem = 660−740 nm) images of HepG2 cells costained with MRP1 and MTDR. (d) Intensity profile of regions of interest (ROIs) in the costained HepG2 cells as indicated by the white arrows in (b) and (c). (e−g) Bright field, red, and NIR images of HepG2 cells costained with 1b and MTDR. (h) Intensity profile of the ROIs indicated in (f) and (g).
To test whether MRP1 would remain in the mitochondria after the mitochondrial membrane potential was eliminated, the HepG2 cells were stained with MRP1 and then further incubated with CCCP (carbonyl cyanide m-chlorophenyl hydrazone). CCCP is a reagent that can collapse the mitochondrial membrane potential.26 As inferred from the results summarized in Figure 3, MRP1 is retained in the mitochondria after coincubating with CCCP for 80 min. However, after treatment with CCCP, compound 1b was found to diffuse from the mitochondria into the cytoplasm. The difference between these two dye systems is ascribed to the presence of the benzyl chloride moiety in MRP1 that, per the design expectations, acts as an immobilizing group.
Figure 3.

Confocal images of HepG2 cells stained with MRP1 (0.5 μM) or 1b (0.5 μM) in the presence of CCCP (10 μM) and recorded at different time intervals. The images were produced by monitoring the red channel (λem = 575−650 nm).
Prior to testing MRP1 as a sensor for endogenous cyanide, its ability to function as a ratiometric fluorescence probe for exogenous HCN was examined. It was found that HepG2 cells loaded with MRP1 alone showed negligible fluorescence in the green channel but a strong fluorescence in the red channel (Figure 4a–c). On this basis, it is concluded that negligible quantities of HCN are present in HepG2 cells. After the addition of 15 μM KCN, which is largely transformed to HCN under physiological conditions, the fluorescence seen in the red channel decreased, while that in the green channel increased (Figure 4d–f). In the presence of 40 μM cyanide, the fluorescence in the red channel was almost fully quenched, while that of the green channel became commensurately brighter (Figure 4g–i). The subsequent addition of cyanide led to an increase in the green-to-red fluorescence ratio (Fgreen/Fred) (Figure 4m). The increase was found to be linearly proportional to the cyanide concentration, leading us to suggest that probe MRP1 is potentially suitable for quantitative studies (Figure 4n).
Figure 4.

Confocal images of live HepG2 cells stained with MRP1 (0.5 μM) in the presence of different concentrations of exogenous cyanide. (a−c) Images recorded over the green (λem = 460−530 nm) and red (λem = 575−650 nm) channels, and the corresponding (Fgreen/Fred) ratio for HepG2 cells stained with MRP1 cells and incubated with 0 μM cyanide. (d−f) Green and red channel and ratiometric imaging of HepG2 cells stained with MRP1 and incubated with 15 μM cyanide. (g−i) Green and red channels and ratiometric imaging of MRP1 stained HepG2 cells incubated with 40 μM cyanide. (j−l) Green and red channels and ratiometric imaging of MRP1 stained HepG2 cells pretreated with 40 μM MHb, and then incubated with 40 μM cyanide. (m) Average intensity ratios from images (c), (f), (i), and (l). (n) Linear relationship between the average intensity ratio and cyanide concentration. Error bars represent the standard deviation.
To verify that the fluorescence response was due to HCN, the HepG2 cells loaded with cyanide were treated with methemoglobin (MHb; a scavenger of cyanide)27 to reduce the intracellular levels of HCN. In the presence of MHb, the fluorescence response was significantly inhibited (Figure 4j–l), as expected for a fluorescence response that is directly correlated to the HCN levels.
Next, the potential utility of MRP1 as a probe for endogenously generated HCN was evaluated. This was done using rat pheochromocytoma cells (PC12 cells), neuron-like cells that are known to release HCN.8a Upon incubation with MRP1 for 15 min, a strong fluorescence in the green channel (Figure 5a) and weak fluorescence in the red channel (Figure 5b) were seen. The fluorescence ratio (Figure 5c) was enhanced in the case of the PC12 cells as compared to what was seen with HepG2 cells (cf., Figure 4c). On the basis of the fluorescence ratio and the equation given in Figure 4n, the response in the PC12 cells was equivalent to that seen when the HepG2 cells were treated with 8.1 μM cyanide.
Figure 5.

Confocal image of endogenous mitochondrial HCN in PC12 cells. (a−c) Green (λem = 460−530 nm), red (λem = 575−650 nm), and ratiometric (Fgreen/Fred) imaging of PC12 cells incubated with MRP1 (0.5 μM). (d−f) Green, red, and ratiometric imaging of PC12 cells incubated with MRP1 (0.5 μM), and then incubated with 10 μM HYDROM. (g−i) Green, red, and ratiometric imaging of PC12 cells pretreated with 40 μM MHb and then incubated with MRP1 (0.5 μM). (j) Average intensity ratios from images (c), (f), and (i). Error bars represent the standard deviations.
Hydromorphine (HYDROM) is a type of μ-opiate agonist that can stimulate nerve cells to produce HCN.8a After treatment with 10 μM HYDROM for 15 min, the fluorescence ratio seen in the case of the PC12 cells was further enhanced. In this case, the endogenous HCN concentration was calculated to be 23.0 μM. On the other hand, pretreatment of the PC12 cells with 40 μM MHb (to remove the endogenous HCN) followed by incubatation with MRP1 for 15 min led to little if any fluorescence being observed in the green channel (Figure 5g). However, under these conditions, a strong fluorescence response was seen in the red channel (Figure 5h). The fluorescence ratio was reduced significantly (Figure 5i,j), and the HCN concentration was calculated to be only 0.2 μM.
The above results provide support for the suggestion that probe MRP1 may be used to detect different concentrations of endogenous HCN in PC12 cells. Because of the fast response time seen in the initial predicative studies discussed above, a further effort was made to test whether MRP1 could be applied to visualize fluctuations in HCN levels in neurons. The neurons were differentiated from the neural stem cells as detailed in the Supporting Information. After incubating the neurons with MRP1 for 15 min, the fluorescence ratio (Fgreen/Fred) was determined to be 0.89 (Figure 6b). On this basis, the concentration of the endogenous HCN was calculated to be 8.4 μM. The cells were also stimulated with HYDROM (10 μM) for 15 min. Under these conditions, the fluorescence ratio increased to 2.09 (Figure 6c). This value corresponds to an HCN concentration of 21.7 μM. When the HCN scavenger MHb (40 μM) was added to these cells, the fluorescence ratio immediately decreased to 0.18 (Figure 6d). Thus, we suggest that MRP1 can be used to monitor in real time the variations in endogenous HCN concentrations within the mitochondria of neurons. To our knowledge, this represents the first instance wherein a fluorescent probe is used to visualize endogenous HCN in nerve cells.
Figure 6.

Confocal image of endogenous mitochondrial HCN in neurons. (a) Bright field image. (b) Ratiometric (Fgreen/Fred) imaging of the neuron cells after incubation with MRP1 (0.5 μM) for 10 min. (c) Ratiometric (Fgreen/Fred) imaging of the cells in panel (b) and then further stimulated with HYDROM (10 μM) for 10 min. (d) Ratiometric imaging of the cells in panel (c) that were then treated with MHb (40 μM).
The cytotoxicity of MRP1 in HepG2 cells was determined using a standard 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay. On the basis of the results obtained, we infer that the low cytotoxicity of MRP1 in live cells is relatively low (≥95% cell viability at a concentration of 5 μM; cf., Figure S17).
CONCLUSION
In summary, we have prepared and characterized a mitochondria-specific ratiometric fluorescence probe for HCN. This probe relies on 7-diethylamino-coumarin as the fluorophore, a 1-methylenepyrrolidinium group as the reactive cyanide recognition and mitochondria targeting subunit, and a benzyl chloride group to provide immobilization within mitochondria. Photophysical and spectral characterization studies revealed that a lead probe (MRP1) reacts rapidly (within 1 s) and responds reversibly to HCN with both high selectivity and sensitivity (the detection limit is 65.6 nM). Notably, probe MRP1 is retained in the mitochondria even when the mitochondria membrane potential is decreased. It can be used to visualize different concentrations of exogenous HCN within the mitochondria of HepG2 cells and proved effective in monitoring fluctuations in endogenous HCN concentrations within the mitochondria of PC12 Cells and neurons. It is thus believed that probe MRP1 and related systems could emerge as useful tools that might allow the critical toxicological and physiological roles of HCN in the human body to be understood in greater detail.
Supplementary Material
ACKNOWLEDGMENTS
The work in Austin was supported by the National Institutes of Health (RGM 103790A to J.L.S.) and the Robert A. Welch Foundation (F-0018). L.L. gratefully acknowledges the program of China Scholarships Council (201608320096 to L.L.) and the National Sciences Foundation of China (21202063 to L.L.) for financial support.
Footnotes
Supporting Information
The Supporting Information is available free of charge on the ACS Publications website at DOI: 10.1021/jacs.7b12545.
Synthetic details, the method of cell culture and fluorescence imaging, and other spectroscopic data (PDF)
The authors declare no competing financial interest.
REFERENCES
- (1).Baskin SI; Brewer TG Cyanide poisoning In Medical Aspects of Chemical and Biological Warfare; Sidell F, Takafuji ET, Franz DR, Eds.; TMM Publication: Washington, DC, 1997; p 271. [Google Scholar]
- (2).Mokhlesi B; Leikin JB; Murray P; Corbridge TC Chest 2003, 123, 897. [DOI] [PubMed] [Google Scholar]
- (3).(a) Young C; Tidwell L; Anderson C Cyanide: Social, Industrial, and Economic Aspects; Minerals, Metals, and Materials Society: Warrendale, 2001. [Google Scholar]; (b) Long L; Wang L; Wu Y; Gong A; Da Z; Zhang C; Han Z Chem. - Asian J 2014, 9, 3291. [DOI] [PubMed] [Google Scholar]; (c) Zdor RE J. Appl. Microbiol 2015, 118, 267. [DOI] [PubMed] [Google Scholar]; (d) Ferrari LA; Giannuzzi L Forensic Sci. Int 2015, 256, 46. [DOI] [PubMed] [Google Scholar]; (e) Surleva AR; Drochioiu G J. Chem. Educ 2013, 90, 1654. [Google Scholar]; (f) Moussa SG; Leithead A; Li S-M; Chan TW; Wentzell JJB; Stroud C; Zhang J; Lee P; Lu G; Brook JR; Hayden K; Narayan J; Liggio J Atmos. Environ 2016, 131, 185. [Google Scholar]; (g) Reade MC; Davies SR; Morley PT; Dennett J; Jacobs IC; Australian Resuscitation, C. Emerg. Med. Australas 2012, 24, 225. [DOI] [PubMed] [Google Scholar]
- (4).Way JL Annu. Rev. Pharmacol. Toxicol 1984, 24, 451. [DOI] [PubMed] [Google Scholar]
- (5).Yamamoto H.-a.; Mohanan PV Toxicology 2002, 179, 29. [DOI] [PubMed] [Google Scholar]
- (6).(a) Prabhakaran K; Li L; Mills EM; Borowitz JL; Isom GE J. Pharmacol. Exp. Ther 2005, 314, 1338. [DOI] [PubMed] [Google Scholar]; (b) Sawa A; Wiegand GW; Cooper J; Margolis RL; Sharp AH; Lawler JF; Greenamyre JT; Snyder SH; Ross CA Nat. Med 1999, 5, 1194. [DOI] [PubMed] [Google Scholar]; (c) Dauer W; Przedborski S Neuron 2003, 39, 889. [DOI] [PubMed] [Google Scholar]; (d) McMillan DE; Geevarghese PJ Diabetes Care 1979, 2, 202. [DOI] [PubMed] [Google Scholar]; (e) Boland ML; Chourasia AH; Macleod KF Front. Oncol 2013, 3, 292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (7).(a) Stelmaszynska T; Zgliczynski J The role of myeloperoxidase in phagocytosis, with special regard to HCN formation In Cyanide in Biology; Vennesland B, Conn E, Knowles C, Westley J, Wissing F, Eds.; Academic Press: New York, 1981; p 371. [Google Scholar]; (b) Zgliczyński JM; Stelmaszyńska T Biochim. Biophys. Acta, Enzymology 1979, 567, 309. [DOI] [PubMed] [Google Scholar]; (c) DeChatelet LR; McPhail LC; Shirley PS Blood 1977, 49, 445. [PubMed] [Google Scholar]
- (8).(a) Borowitz JL; Gunasekar PG; Isom GE Brain Res 1997, 768, 294. [DOI] [PubMed] [Google Scholar]; (b) Gunasekar P; Prabhakaran K; Li L; Zhang L; Isom G; Borowitz J Neurosci. Res 2004, 49, 13. [DOI] [PubMed] [Google Scholar]; (c) Gunasekar PG; Borowitz JL; Turek JJ; Van Horn DA; Isom GE J. Neurosci. Res 2000, 61, 570. [DOI] [PubMed] [Google Scholar]; (d) Mills EM; Gunasekar PG; Pavlakovic G; Isom GE J. Neurochem 1996, 67, 1039. [DOI] [PubMed] [Google Scholar]
- (9).(a) Cipollone R; Visca P IUBMB Life 2007, 59, 187. [DOI] [PubMed] [Google Scholar]; (b) Cooper CE; Brown GC J. Bioenerg. Biomembr 2008, 40, 533. [DOI] [PubMed] [Google Scholar]
- (10).(a) Yang Y; Zhao Q; Feng W; Li F Chem. Rev 2013, 113, 192. [DOI] [PubMed] [Google Scholar]; (b) Chan J; Dodani SC; Chang CJ Nat. Chem 2012, 4, 973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (11).Xu Z; Xu L Chem. Commun 2016, 52, 1094. [DOI] [PubMed] [Google Scholar]
- (12).(a) Dong YM; Peng Y; Dong M; Wang YW J. Org. Chem 2011, 76, 6962. [DOI] [PubMed] [Google Scholar]; (b) Gomez T; Moreno D; Diaz de Grenu B; Fernandez AC; Rodriguez T; Rojo J; Cuevas JV; Torroba T Chem. - Asian J 2013, 8, 1271. [DOI] [PubMed] [Google Scholar]; (c) Huang X; Gu X; Zhang G; Zhang D Chem. Commun 2012, 48, 12195. [DOI] [PubMed] [Google Scholar]; (d) Kim HJ; Ko KC; Lee JH; Lee JY; Kim JS Chem. Commun 2011, 47, 2886. [DOI] [PubMed] [Google Scholar]; (e) Kumar S; Singh P; Hundal G; Hundal MS; Kumar S Chem. Commun 2013, 49, 2667. [DOI] [PubMed] [Google Scholar]; (f) Li H; Wen Z; Jin L; Kan Y; Yin B Chem. Commun 2012, 48, 11659. [DOI] [PubMed] [Google Scholar]; (g) Lin YD; Pen YS; Su W; Liau KL; Wen YS; Tu CH; Sun CH; Chow TJ Chem. -Asian J 2012, 7, 2864. [DOI] [PubMed] [Google Scholar]; (h) Lv X; Liu J; Liu Y; Zhao Y; Sun YQ; Wang P; Guo W Chem. Commun 2011, 47, 12843. [DOI] [PubMed] [Google Scholar]; (i) Shankar BH; Jayaram DT; Ramaiah D Chem. - Asian J 2014, 9, 1636. [DOI] [PubMed] [Google Scholar]; (j) Yang L; Li X; Yang J; Qu Y; Hua J ACS Appl. Mater. Interfaces 2013, 5, 1317. [DOI] [PubMed] [Google Scholar]; (k) Zhang P; Shi B; You X; Zhang Y; Lin Q; Yao H; Wei T Tetrahedron 2014, 70, 1889. [Google Scholar]; (l) Cho DG; Kim JH; Sessler JL J. Am. Chem. Soc 2008, 130, 12163. [DOI] [PubMed] [Google Scholar]; (m) Shiraishi Y; Nakamura M; Hayashi N; Hirai T Anal. Chem 2016, 88, 6805. [DOI] [PubMed] [Google Scholar]; (n) Bejoymohandas KS; Kumar A; Sreenadh S; Varathan E; Varughese S; Subramanian V; Reddy ML Inorg. Chem 2016, 55, 3448. [DOI] [PubMed] [Google Scholar]
- (13).(a) Divya KP; Sreejith S; Balakrishna B; Jayamurthy P; Anees P; Ajayaghosh A Chem. Commun 2010, 46, 6069. [DOI] [PubMed] [Google Scholar]; (b) Lee JH; Jeong AR; Shin IS; Kim HJ; Hong JI Org. Lett 2010, 12, 764. [DOI] [PubMed] [Google Scholar]; (c) Xie Y; Ding Y; Li X; Wang C; Hill JP; Ariga K; Zhang W; Zhu W Chem. Commun 2012, 48, 11513. [DOI] [PubMed] [Google Scholar]; (d) Shojaeifard Z; Hemmateenejad B; Shamsipur M ACS Appl. Mater. Interfaces 2016, 8, 15177. [DOI] [PubMed] [Google Scholar]; (e) Kaushik R; Ghosh A; Singh A; Gupta P; Mittal A; Jose DA ACS Sensors 2016, 1, 1265. [Google Scholar]
- (14).(a) Liu J; Lin Q; Yao H; Wang M; Zhang Y-M; Wei T-B Chin. Chem. Lett 2014, 25, 35. [Google Scholar]; (b) Saha S; Ghosh A; Mahato P; Mishra S; Mishra SK; Suresh E; Das S; Das A Org. Lett 2010, 12, 3406. [DOI] [PubMed] [Google Scholar]; (c) Sharma S; Hundal MS; Hundal G Org. Biomol. Chem 2013, 11, 654. [DOI] [PubMed] [Google Scholar]
- (15).(a) Badugu R; Lakowicz JR; Geddes CD J. Am. Chem. Soc 2005, 127, 3635. [DOI] [PMC free article] [PubMed] [Google Scholar]; (b) Jamkratoke M; Ruangpornvisuti V; Tumcharern G; Tuntulani T; Tomapatanaget B J. Org. Chem 2009, 74, 3919. [DOI] [PubMed] [Google Scholar]; (c) Kim Y; Huh HS; Lee MH; Lenov IL; Zhao H; Gabbai FP Chem. - Eur. J 2011, 17, 2057. [DOI] [PubMed] [Google Scholar]; (d) Varlan M; Blight BA; Wang S Chem. Commun 2012, 48, 12059. [DOI] [PubMed] [Google Scholar]
- (16).(a) Wang L; Zheng J; Yang S; Wu C; Liu C; Xiao Y; Li Y; Qing Z; Yang R ACS Appl. Mater. Interfaces 2015, 7, 19509. [DOI] [PubMed] [Google Scholar]; (b) Mannel-Croise C; Probst B; Zelder F Anal. Chem 2009, 81, 9493. [DOI] [PubMed] [Google Scholar]; (c) Mannel-Croise C; Zelder F ACS Appl. Mater. Interfaces 2012, 4, 725. [DOI] [PubMed] [Google Scholar]
- (17).(a) Mello JV; Finney NS Angew. Chem., Int. Ed 2001, 40, 1536. [PubMed] [Google Scholar]; (b) Kikuchi K; Takakusa H; Nagano T TrAC, Trends Anal. Chem 2004, 23, 407. [Google Scholar]
- (18).(a) Johnson LV; Walsh ML; Bockus BJ; Chen LB J. Cell Biol 1981, 88, 526. [DOI] [PMC free article] [PubMed] [Google Scholar]; (b) Lemasters JJ; Qian T; Bradham CA; Brenner DA; Cascio WE; Trost LC; Nishimura Y; Nieminen A-L; Herman BJ Bioenerg. Biomembr 1999, 31, 305. [DOI] [PubMed] [Google Scholar]
- (19).Long LL; Li XF; Zhang DD; Meng SC; Zhang JF; Sun XL; Zhang C; Zhou LP; Wang L RSC Adv 2013, 3, 12204. [Google Scholar]
- (20).Chen Y; Zhu C; Yang Z; Li J; Jiao Y; He W; Chen J; Guo Z Chem. Commun 2012, 48, 5094. [DOI] [PubMed] [Google Scholar]
- (21).Bozdemir OA; Sozmen F; Buyukcakir O; Guliyev R; Cakmak Y; Akkaya EU Org. Lett 2010, 12, 1400. [DOI] [PubMed] [Google Scholar]
- (22).Nag A; Chakrabarty T; Bhattacharyya K J. Phys. Chem 1990, 94, 4203. [Google Scholar]
- (23).(a) Guido CA; Mennucci B; Jacquemin D; Adamo C Phys. Chem. Chem. Phys 2010, 12, 8016. [DOI] [PubMed] [Google Scholar]; (b) Jacquemin D; Le Bahers T; Adamo C; Ciofini I Phys. Chem. Chem. Phys 2012, 14, 5383. [DOI] [PubMed] [Google Scholar]
- (24).Presley AD; Fuller KM; Arriaga EA J. Chromatogr. B: Anal. Technol. Biomed. Life Sci 2003, 793, 141. [DOI] [PubMed] [Google Scholar]
- (25).(a) Dickinson BC; Chang CJ J. Am. Chem. Soc 2008, 130, 9638. [DOI] [PMC free article] [PubMed] [Google Scholar]; (b) Zhou R; Yazdi AS; Menu P; Tschopp J Nature 2011, 469, 221. [DOI] [PubMed] [Google Scholar]
- (26).Caporale T; Ciavardelli D; Di Ilio C; Lanuti P; Drago D; Sensi SL Exp. Neurol 2009, 218, 228. [DOI] [PubMed] [Google Scholar]
- (27).Moriya F; Hashimoto Y Leg. Med 2003, 5, S113. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.