

Abstract
Background:
Keratoconus (KC) is a degenerative eye disease which results from thinning of the cornea and causes vision distortion. Oxidative stress damage to KC corneas may be because of the failure of corneas to process reactive oxygen species which leads to corneal thinning and loss of vision. Genetic variants in antioxidant defense genes such as catalase (CAT) and glutathione peroxidase (GPX) can decrease antioxidant capacity or increase oxidative stress and alter the risk of KC in patients. We investigated and evaluated the effects of single nucleotide polymorphisms in CAT, GPX-1 on the risk of KC in an Iranian population sample.
Methods:
This case-control study was performed on 140 patients with KC and 150 healthy control subjects in a sample of Iranian population from Zahedan, southern Iran in 2015. Genotyping of CAT rs7943316 and GPX-1 rs1050450 polymorphisms was done using polymerase chain reaction and restriction fragment length polymorphism (PCR-RFLP) method.
Results:
CAT rs7943316 A/T, AA genotype and A allele have a protective role against disease (OR =0.28, 95% CI =0.13–0.61, P=0.001 and OR = 0.50, 95% CI =0.35–0.72, P=0.0001, respectively) and decreased the risk of KC. Moreover, GPX-1 rs1050450 T allele increased the risk of KC in comparison with C allele (OR = 1.42, 95% CI = 1.01–2.03, P=0.03).
Conclusion:
CAT rs7943316 A/T, AA genotype, and A allele decreased the risk of KC. Moreover, in GPX-1 rs1050450 C/T polymorphism, T allele was associated with an increased risk of KC in our population.
Keywords: CAT, GPX-1, Oxidative stress, Keratoconus, Polymorphism
Introduction
Keratoconus (KC) is a degenerative eye disease which results from thinning of the cornea and causes vision distortion (1). This condition is characterized by the stromal thinning of the cornea which often results in bilateral and asymmetric corneal distortion and anterior corneal protrusion (2). Symptoms are highly variable and depend on the stage of the progression of the disorder. In the initial stage of the disease, there may be no symptoms; in the advanced stage, there is significant distortion of vision accompanied by serious visual loss. Patients with KC never become completely blind from their disease (3). KC is a major suggestion for corneal transplantation in developed countries(4). KC usually occurs in the second decade of life with progress in the next two decades and affects both genders and all ethnicities. The expected prevalence in the general population is 54 per 100000(5, 6). KC is most commonly an isolated disease, while several reports describe an association with Down syndrome, monosomy X (Turner syndrome), Leber’s congenital Ehlers-Danlos syndrome, neurocutaneousangiomatosis, neurofibromatosis, xerodermapigmentosa, collagenosis, retinitis pigmentosa and Marfan syndrome is described (7). The cause and original pathological mechanism are unknown but biochemical, genetic, and environmental factors are possible causes of KC and different etiologies may play a role in the development of this disease(8, 9).
Environmental factors which can influence on eye consist of rub eyeballs, allergic reaction, and solarization (9). An excess of environmental factors, particularly UV exposure, causes oxidative damage to KC corneas because of the failure of KC corneas to process reactive oxygen species (ROS) which leads to corneal thinning and loss of vision (10). The accumulation of ROS can damage cells by reacting with proteins, DNA, and membrane phospholipids (11). The normal cornea’s antioxidant enzymes eliminate the ROS before they damage cells; these consist of superoxide dismutase, glutathione reductase, catalase, and glutathione peroxidase, but in the disease condition, ROS can devastate cellular defenses and promote cell damage (12).
Catalase (CAT) is a common enzyme that found in the peroxisome of many aerobic organisms. This enzyme is a tetramer with molecular weight of about 240000 with four heme groups per tetramer (13, 14). Hydrogen peroxide (H2O2) is converted to H2O and O2 by CAT and thereby mitigates the toxic effects of hydrogen peroxide (15). Location of CAT gene is on chromosome 11p13 and comprise 13 exons; rs7943316 (-21 A/T) settled in nearly to the start site at promoter region (16). Deficiency of catalase may cause elevated concentrations of hydrogen peroxide and increase the risk of the progress of pathologies (17). Glutathione Peroxidase-1(GPX-1) encodes a member of the glutathione peroxidase family. GPX-1 is an intracellular antioxidant enzyme that converts H2O2 to water and protects the organism from oxidative damage(18, 19). GPx-1 is selenium-containing Enzyme, located at chromosome 3p21.3(20). GPx-1 has four SNPs that alter the amino acid produced but only one has been studied extensively in human disease, including rs1050450 (or GPx1 Pro197Leu). This C>T variation changes the amino acid from proline (Pro) to leucine (Leu) at position 197 (21). The GPx-1 variant may be associated with a decreased ability to scavenge ROS (22).
To the best of our knowledge, no studies have investigated the role of antioxidant gene CAT and GPX polymorphisms for KC patients. The present study aimed to evaluate the impact of CAT rs7943316 A/T and GPX-1 rs1050450 C/T polymorphisms on KC patients in a sample of Iranian population. These polymorphisms may alter the enzymes’ antioxidant capacity and may be associated with the risk of KC induced by oxidative damage.
Materials and Methods
Patients
This case-control study was done on 140 patients with KC (61 men and 79 women), and 150 healthy individuals as the control group (65 men and 85 women), who were enrolled from Alzahra Eye Hospital, Zahedan, southern Iran in 2015.
Ethical approvals for recruitment were obtained from the local Ethics Committee of Zahedan University of Medical Sciences and informed consents were obtained from all patients and healthy individuals. Detection of KC was done under criteria previously mentioned (23).
Sampling and extraction of DNA
Venus blood samples were collected from the patients and healthy controls, using EDTA-containing tubes, and DNA was extracted using the salting-out method (24). The quality of the isolated DNA was verified using electrophoresis on 1% agarose gel and quantitated spectrophotometrically and stored, at -20°C until further use.
Genotyping of CAT rs7943316 A/T Polymorphism
The restriction fragment length polymorphism, polymerase chain reaction (RFLP -PCR), was used for genotyping CAT rs7943316 polymorphism using forward and reverse primer sequences as follows: 5′-CTTCCAATCTTGGCCT-GCCTAG -3′ and 5′-CCGCTTTCTAAACGGACCTTCG-3′, respectively(25). PCR was directed using PCR master mix (AmpliqonTaq 2x mastermix, Denmark) according to the manufacturer’s instructions. The following conditions were involved for amplification of SNPs in (20 μL volume): 1 μL template DNA (∼100 ng/ μL), 1 μL of each primer (10 pmol/μL), 10 μL mastermix and 7 μLDNase free water were added. PCR product size was 312 bp. The PCR cycling conditions consisted of an initial denaturing step for 5min at 95°C followed by 30 cycles for 30 sec at 95°C, 40 sec at 60°C, and 30 sec at 72°C, anda final extension step for 5 min at 72°C. The PCR products were visualized on a 2% agarose gel containing 0.5 μg/mL of ethidium bromide. Amplified DNA was digested with restriction enzyme HinfI (BIORON, Germany) at 37 °C for 3 h according to the manufacturer’s instructions. HinfI digestion resulted in two 109bp and 203bp fragments for AA genotype, three fragments of 312, 203 and 109bp for AT genotype, and no digested product is a 312bp for TT genotype(Fig. 1A).
Fig. 1:
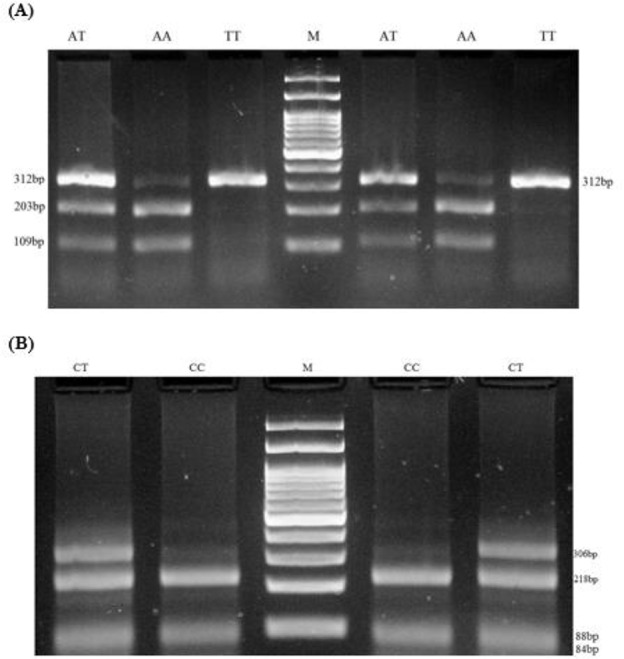
Electrophoresis pattern for detection of SNP in CAT rs7943316 A/T and GPX-1 rs1050450 C/T (A) RFLP -PCR products of CAT rs7943316 A/T, M: DNA marker (100 bp), product size (109, 203, 312bp), demonstrate genotype AT; product size(109, 203bp), demonstrate genotype AA and 312bp demonstrate genotype TT. (B) RFLP -PCR products of GPX-1 rs1050450 C/T, M: DNA marker (100 bp), product size (84/88, 218, 306bp), demonstrate genotype CT; product size(84/88, 218bp), demonstrate genotype CC
Genotyping of GPX-1 rs1050450 C/T Polymorphism
Genotyping of GPX-1 rs1050450 was done by RFLP-PCR method. Briefly, forward and reverse primers were 5′-TTATGACCGACCCCAAGCTC-3′ and 5′-GACACCCGGCACTTTATTAGTG -3, respectively (25). The PCR conditions were justified as follows: 5 min preheating at 95 °C, 30 cycles of 95 °C for 30 sec, 61 °C for 25 sec, and 72 °C for 28 sec followed by a final extension step for 5 min at 72 °C. PCR product size was 312 bp. Ten microliters of PCR product were later digested by ApaI restriction enzyme (Thermo Scientific, Lithuania) at 37 °C for 3 h. C allele produced 84, 88 and 218bp patterns, while T allele produced 84 and 306bp (Fig. 1B).
Statistical analysis
The statistical analysis was performed using SPSS ver. 19.0 software (Chicago IL, USA). Comparison of quantitative variants between two groups was assessed by Student’s t-test. The frequencies of the alleles and genotypes were analyzed using the χ2-test or Fisher’s exact test. The odds ratio(OR) and 95% confidence intervals (CI) were also estimated. P-value < 0.05 was considered statistically significant.
Results
No significant differences were observed between the patients with KC and healthy individual concerning gender and age (patients: 140, aged 10 to 80 yr and mean ± SD = 28 ± 12.5, P-value: 0.2 and controls: 150, aged 8 to 83 yr and mean ± SD = 29.9 ± 15.6, P-value: 0.2).
Table 1, containing the genotypes and allele distribution of CAT rs7943316 and GPX-1 rs1050450 variants in KC patients and control group. In fact, Table 1 shows the relationship between gene variants (SNP) and number of patients with KC and healthy controls was tested by computing the odds ratio (OR) and 95% confidence intervals (95% CI) from logistic regression analyses.
Table 1:
The genotypes and allele distribution of CAT and GPX-1 variants in keratoconus (KC) patients and control group
| Variants | KC Patients n (%) | Controls n (%) | OR(95 % CI) | P-Value |
|---|---|---|---|---|
| rs7943316, CAT | ||||
| TT | 88 (62.9) | 71 (47.3) | Ref. | — |
| TA | 41 (29.2) | 48 (32) | 0.69 (0.41–1.16) | 0.161 |
| AA | 11 (7.9) | 31 (20.7) | 0.28 (0.13–0.61) | 0.001 |
| Allele | ||||
| T | 217 (77.5) | 190 (63.3) | Ref. | — |
| A | 63 (22.5) | 110 (36.7) | 0.50 (0.35–0.72) | 0.0001 |
| rs1050450, GPX-1 | ||||
| CC | 40 (28.6) | 66 (44) | Ref. | — |
| CT | 100 (71.4) | 84 (56) | 1.96 (1.21–3.20) | 0.007 |
| TT | 0 (0) | 0 (0) | — | — |
| Allele | ||||
| C | 180 (64.3) | 216 (72) | Ref. | — |
| T | 100 (35.7) | 84 (28) | 1.42 (1.01–2.03) | 0.03 |
The frequency distributions of CAT rs7943316 A/T genotypes in KC patients were: TT, 62.9%, TA, 29.2% and AA, 7.9%; and the distribution in healthy controls were: TT, 47.3%, TA, 32%, and AA, 20.7%. This SNP was associated with KC. The AA and A allele decreased the risk of KC (OR = 0.28, 95% CI = 0.13–0.61, P-value=0.001 and OR =0.50, 95% CI =0.35–0.72, P-value =0.0001, respectively) (Table 1).
The frequencies distributions of CC and CT genotypes of GPX-1 rs1050450 C/T gene polymorphism in KC patients were: 28.6% and 71.4% and in healthy controls were: 44% and56%, respectively. The T allele was associated with KC. Thers1050450 T allele increased the risk of KC in comparison with C allele (OR =1.42, 95% CI =1.01–2.03, P-value= 0.03) (Table 1).
The possible association between polymorphisms and clinical and pathological characteristics were analyzed. Significant association was observed between GPX-1 rs1050450 polymorphism with level of KC and cross-linking (CXL) (P-value= 0.04 and 0.04, respectively) (Table 2).
Table 2:
Association between the CAT rs7943316 and GPX-1 rs1050450 polymorphisms and clinicopathological characteristics
| Parameters evaluated | Patients n (%) | (rs7943316) P-value | (rs1050450) P-value |
|---|---|---|---|
| KC ocular | |||
| OD | 42(30.0) | ||
| OS | 36(25.7) | 0.95 | 0.05 |
| OU | 62(44.3) | ||
| Level of KC | |||
| KK 1 | 33(23.6) | ||
| KK 2 | 45(32.1) | 0.82 | 0.04 |
| KK 3 | 62(44.3) | ||
| CXL | |||
| OD | 39(27.9) | ||
| OS | 40(28.6) | 0.45 | 0.04 |
| OU | 42(30.0) | ||
| Candidate | 19(13.6) |
KC: keratoconus, OD (right eye), OS (left eye), OU (both eyes), CXL (cross linking surgery)
We analyzed the possible association between polymorphisms and clinical and pathological characteristics (Table 2). KC ocular showed the existence of keratoconus in a patient’s eye and relation with SNPs. Level of KC represents the level of disease and evaluating these criteria with polymorphisms from patients and our findings were not associated with KC. CXL represents number of persons has cross-linking surgery and relation with SNPS.
Discussion
We investigated the impact of CAT rs7943316 A/T and GPX-1 rs1050450C/T polymorphisms on KC risk in a sample of Iranian population because these polymorphisms may alter the enzymes’ antioxidant capacity, leading to synergistic effects with KC induced by oxidative damage.
Our results indicated that CAT rs7943316 A/T, AA genotype, and A allele significantly decreased the risk of KC. Moreover, GPX-1 rs1050450 C/T, T allele was associated with an increased risk of KC in our population. Genetic variations in antioxidant defense may decrease antioxidant capacity or increase oxidative stress and alter the risk of KC in patients. SNPs and gene variants suggest an intricate aetiology or convergence of multiple disease pathways. Elevation of oxidative stress from SNPs in specific antioxidant enzymes maybe associated with disease (21).
Inflammation plays a crucial role in the pathogenesis of KC. Corneal thinning and distortion in KC associated with some features of inflammation in ocular rosacea (26). Further evidence emphasizes the reduced levels of antioxidant defense enzymes in KC to remove ROS associated with inflammatory reactions (27). Oxidative stress results from increased ROS or decreased levels of antioxidants (28). Ultraviolet light (UV) is a source of ROS, and excessive exposure to sunlight leads to oxidative damage to KC corneas, where there is a reduced amount of the enzymes including CAT and GPX-1 necessary to remove the ROS (29). UV-B is damaging to the cornea, and UV-A damages both the cornea and lens (30). The effects of sunlight on the eye may be acute after a latent period of several hours. UV-B can foundation homolytic fission of H2O2 to generate hydroxyl radical (31). Moreover, there is an accumulation of oxidized mitochondrial DNA in KC relative to control corneas (32). The normal cornea’s antioxidant enzymes such as CAT and GPX eliminate the ROS before they damage cells, but in the disease condition, ROS can devastate cellular defenses and promote cell damage (12). KC corneas do not eliminate ROS in a normal way, which may play a major role in the pathogenesis of this disease(33).
Irregular expression of several major antioxidant enzymes including glutathione activities has been reported in KC corneas (34). In KC corneas compared with normal corneas, striking decreased in antioxidant potency and glutathione content (35). The KC corneas were exhibited a 2.2-fold increase of catalase mRNA level and 1.8-fold of enzyme activity. Since H2O2 induces catalase expression, KC corneas had high levels of this ROS, explaining some oxidative damages related with KC corneas (36). The elevated levels of oxidative stress products and decreased anti-oxidant capacity and antioxidant defenses in KC corneas indicate that oxidative stress may be involved in the progress of this pathology (35).
CAT rs7943316 and GPX-1 rs1050450 polymorphisms alter their antioxidant capacity and associated with different kinds of diseases such as Kashin-Beck (37), prostate cancer (38), peripheral neuropathy (39), diabetes mellitus (16) recurrent miscarriage (40), and brain tumors (41). In opposite to our findings, CAT rs7943316 genotype (TT) was associated with an increased risk of T2DM and no evidence was found to support an association between GPx-1 (198 C/T) polymorphism and patients with T2DM (25). Allele and genotype frequencies of the CAT–21A/T and GPX1–198C/T, variations on patients with cataract and significant differences were not observed compare controls (41); both in terms of disease and result obtained did not establish relationship with our results.
The variability of the XRCC1, POLG and FEN1 genes in the base excision repair pathway may play a role in KC pathogenesis and increase the risk of this disease (42, 43). Patients with KC corneas that have lower paraoxonase activity there were sensitive to oxidative stress (44).
The current report is the first to suggest an association between the polymorphisms of antioxidant enzyme genes with KC in the Iranian population. However, the present study has some limitations. For example, we examined just one polymorphism from each gene associated with KC and we did not consider different ethnics.
Conclusion
CAT rs7943316 decreased the risk of KC. Moreover, GPX-1 rs1050450 T allele increased the risk of KC in the study population. Further studies with different ethnicities and nationalities are required to confirm our findings.
Ethical considerations
Ethical issues (Including plagiarism, informed consent, misconduct, data fabrication and/or falsification, double publication and/or submission, redundancy, etc.) have been completely observed by the authors.
Acknowledgements
This paper was funded as research Grant (no.6878) from the Deputy for Research, Zahedan University of Medical Sciences, Zahedan, Iran.
Footnotes
Conflict of interest
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Cuellar-Partida G, Springelkamp H, Lucas SE, et al. (2015). WNT10A exonic variant increases the risk of keratoconus by decreasing corneal thickness. Hum Mol Genet, 24(17):5060–8. [DOI] [PubMed] [Google Scholar]
- 2.Wheeler J, Hauser MA, Afshari NA, et al. (2012). The Genetics of Keratoconus: A Review. Reprod Syst Sex Disord, (Suppl 6):001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Rabinowitz YS. (1998). Keratoconus. Surv Ophthalmol, 42(4):297–319. [DOI] [PubMed] [Google Scholar]
- 4.Kang PC, Klintworth GK, Kim T, et al. (2005). Trends in the indications for penetrating keratoplasty, 1980–2001. Cornea, 24(7):801–3. [DOI] [PubMed] [Google Scholar]
- 5.Romero-Jimenez M, Santodomingo-Rubido J, Wolffsohn JS. (2010). Keratoconus: a review. Cont Lens Anterior Eye, 33(4):157–66. [DOI] [PubMed] [Google Scholar]
- 6.Saee-Rad S, Hashemi H, Miraftab M, et al. (2011). Mutation analysis of VSX1 and SOD1 in Iranian patients with keratoconus. Mol Vis, 17:3128–36. [PMC free article] [PubMed] [Google Scholar]
- 7.Stabuc-Silih M, Strazisar M, Ravnik-Glavac M, et al. (2010). Genetics and clinical characteristics of keratoconus. Acta Dermatovenerol Alp Pannonica Adriat, 19(2):3–10. [PubMed] [Google Scholar]
- 8.Sugar J, Macsai MS. (2012). What causes keratoconus? Cornea, 31(6):716–9. [DOI] [PubMed] [Google Scholar]
- 9.Davidson AE, Hayes S, Hardcastle AJ, et al. (2014). The pathogenesis of keratoconus. Eye (Lond), 28(2):189–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cristina Kenney M, Brown DJ. (2003). The cascade hypothesis of keratoconus. Cont Lens Anterior Eye, 26(3):139–46. [DOI] [PubMed] [Google Scholar]
- 11.Esterbauer H, Schaur RJ, Zollner H. (1991). Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic Biol Med, 11(1):81–128. [DOI] [PubMed] [Google Scholar]
- 12.Squadrito GL, Pryor WA. (1998). Oxidative chemistry of nitric oxide: the roles of superoxide, peroxynitrite, and carbon dioxide. Free Radic Biol Med, 25(4–5):392–403. [DOI] [PubMed] [Google Scholar]
- 13.Kirkman HN, Gaetani GF. (1984). Catalase: a tetrameric enzyme with four tightly bound molecules of NADPH. Proc Natl Acad Sci U S A, 81(14):4343–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sabet EE, Salehi Z, Khodayari S, et al. (2014). Polymorphisms of glutathione peroxidase 1 (GPX1 Pro198Leu) and catalase (CAT C-262T) in women with spontaneous abortion. Syst Biol Reprod Med, 60(5):304–7. [DOI] [PubMed] [Google Scholar]
- 15.Yung LM, Leung FP, Yao X, et al. (2006). Reactive oxygen species in vascular wall. Cardiovasc Hematol Disord Drug Targets, 6(1):1–19. [DOI] [PubMed] [Google Scholar]
- 16.Flekac M, Skrha J, Hilgertova J, et al. (2008). Gene polymorphisms of superoxide dismutases and catalase in diabetes mellitus. BMC Med Genet, 9:30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Vitai M, Fatrai S, Rass P, et al. (2005). Simple PCR heteroduplex, SSCP mutation screening methods for the detection of novel catalase mutations in Hungarian patients with type 2 diabetes mellitus. Clin Chem Lab Med, 43(12):1346–50. [DOI] [PubMed] [Google Scholar]
- 18.Bera S, Weinberg F, Ekoue DN, et al. (2014). Natural allelic variations in glutathione peroxidase-1 affect its subcellular localization and function. Cancer Res, 74(18):5118–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lubos E, Loscalzo J, Handy DE. (2011). Glutathione peroxidase-1 in health and disease: from molecular mechanisms to therapeutic opportunities. Antioxid Redox Signal, 15(7):1957–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Goldberg M, Alberts DS, Buckmeier JA, et al. (2011). Loss of heterozygosity at the glutathione peroxidase 1 locus is not an early event in colon carcinogenesis. Genes Cancer, 2(9):910–913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Crawford A, Fassett RG, Geraghty DP, et al. (2012). Relationships between single nucleotide polymorphisms of antioxidant enzymes and disease. Gene, 501(2):89–103. [DOI] [PubMed] [Google Scholar]
- 22.Nemoto M, Nishimura R, Sasaki T, et al. (2007). Genetic association of glutathione peroxidase-1 with coronary artery calcification in type 2 diabetes: a case control study with multi-slice computed tomography. Cardiovasc Diabetol, 6:23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Saravani R, Hasanian-Langroudi F, Validad MH, et al. (2015). Evaluation of possible relationship between COL4A4 gene polymorphisms and risk of keratoconus. Cornea, 34(3):318–22. [DOI] [PubMed] [Google Scholar]
- 24.Hashemi M, Yousefi J, Hashemi SM, et al. (2015). Association between Programmed Cell Death 6 Interacting Protein Insertion/Deletion Polymorphism and the Risk of Breast Cancer in a Sample of Iranian Population. Dis Markers, 2015:854621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Saravani S, Miri HR, Saravani R, et al. (2015). Association of catalase (rs7943316) and glutathione peroxidase-1 (rs1050450) polymorphisms with the risk of type 2 diabetes (T2DM). Mol Gen Mikrobiol Virusol, 30(4):216–20. [Google Scholar]
- 26.McMonnies CW. (2015). Inflammation and keratoconus. Optom Vis Sci, 92(2):e35–41. [DOI] [PubMed] [Google Scholar]
- 27.Behndig A, Karlsson K, Johansson BO, et al. (2001). Superoxide dismutase isoenzymes in the normal and diseased human cornea. Invest Ophthalmol Vis Sci, 42(10):2293–6. [PubMed] [Google Scholar]
- 28.Matough FA, Budin SB, Hamid ZA, et al. (2012). The role of oxidative stress and antioxidants in diabetic complications. Sultan Qaboos Univ Med J, 12(1):5–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Gordon-Shaag A, Millodot M, Shneor E, et al. (2015). The Genetic and Environmental Factors for Keratoconus. Biomed Res Int, 2015:795738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Mihail S. (1989). [The eye and ultraviolet radiation]. Rev Chir Oncol Radiol O R L Oftalmol Stomatol Ser Oftalmol, 33(4):241–4. [PubMed] [Google Scholar]
- 31.Shoham A, Hadziahmetovic M, Dunaief JL, et al. (2008). Oxidative stress in diseases of the human cornea. Free Radic Biol Med, 45(8):1047–55. [DOI] [PubMed] [Google Scholar]
- 32.Atilano SR, Coskun P, Chwa M, et al. (2005). Accumulation of mitochondrial DNA damage in keratoconus corneas. Invest Ophthalmol Vis Sci, 46(4):1256–63. [DOI] [PubMed] [Google Scholar]
- 33.Buddi R, Lin B, Atilano SR, et al. (2002). Evidence of oxidative stress in human corneal diseases. J Histochem Cytochem, 50(3):341–51. [DOI] [PubMed] [Google Scholar]
- 34.Gondhowiardjo TD, van Haeringen NJ. (1993). Corneal aldehyde dehydrogenase, glutathione reductase, and glutathione S-transferase in pathologic corneas. Cornea, 12(4):310–4. [DOI] [PubMed] [Google Scholar]
- 35.Arnal E, Peris-Martinez C, Menezo JL, et al. (2011). Oxidative stress in keratoconus? Invest Ophthalmol Vis Sci, 52(12):8592–7. [DOI] [PubMed] [Google Scholar]
- 36.Kenney MC, Chwa M, Atilano SR, et al. (2005). Increased levels of catalase and cathepsin V/L2 but decreased TIMP-1 in keratoconus corneas: evidence that oxidative stress plays a role in this disorder. Invest Ophthalmol Vis Sci, 46(3):823–32. [DOI] [PubMed] [Google Scholar]
- 37.Xiong YM, Mo XY, Zou XZ, et al. (2010). Association study between polymorphisms in selenoprotein genes and susceptibility to Kashin-Beck disease. Osteoarthritis Cartilage, 18(6):817–24. [DOI] [PubMed] [Google Scholar]
- 38.Steinbrecher A, Meplan C, Hesketh J, et al. (2010). Effects of selenium status and polymorphisms in selenoprotein genes on prostate cancer risk in a prospective study of European men. Cancer Epidemiol Biomarkers Prev, 19(11):2958–68. [DOI] [PubMed] [Google Scholar]
- 39.Tang TS, Prior SL, Li KW, et al. (2012). Association between the rs1050450 glutathione peroxidase-1 (C > T) gene variant and peripheral neuropathy in two independent samples of subjects with diabetes mellitus. Nutr Metab Cardiovasc Dis, 22(5):417–25. [DOI] [PubMed] [Google Scholar]
- 40.Khadzhieva MB, Lutcenko NN, Volodin IV, et al. (2014). Association of oxidative stress-related genes with idiopathic recurrent miscarriage. Free Radic Res, 48(5):534–41. [DOI] [PubMed] [Google Scholar]
- 41.Zhang Y, Zhang L, Sun D, et al. (2011). Genetic polymorphisms of superoxide dismutases, catalase, and glutathione peroxidase in age-related cataract. Mol Vis, 17:2325–32. [PMC free article] [PubMed] [Google Scholar]
- 42.Wojcik KA, Synowiec E, Sobierajczyk K, et al. (2014). Polymorphism of the DNA base excision repair genes in keratoconus. Int J Mol Sci, 15(11):19682–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wojcik KA, Synowiec E, Polakowski P, et al. (2014). Polymorphism of the flap endonuclease 1 gene in keratoconus and Fuchs endothelial corneal dystrophy. Int J Mol Sci, 15(8):14786–802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Poh R, Tan JA, Deva JP, et al. (2012). Paraoxonase 1 status in keratoconus: a preliminary study of activity and polymorphism. West Indian Med J, 61(6):569–73. [PubMed] [Google Scholar]