

Abstract
The semi-aquatic bugs (Heteroptera, Gerromorpha) conquered water surfaces worldwide and diversified to occupy puddles, ponds, streams, lakes, mangroves and even oceans. Critical to this lifestyle is the evolution of sets of hairs that allow these insects to maintain their body weight on the water surface and protect the animals against wetting and drowning. In addition, the legs of these insects are equipped with various grooming combs that are important for cleaning and tidying the hair layers for optimal functional efficiency. Here we show that the hairs covering the legs of water striders represent innervated bristles. Genomic and transcriptomic analyses revealed that in water striders the achaete–scute complex, known to control bristle development in flies, contains only the achaete–scute homologue (ASH) gene owing to the loss of the gene asense. Using RNA interference, we show that ASH plays a pivotal role in the development of both bristles and grooming combs in water striders. Our data suggest that the ASH locus may have contributed to the adaptation to water surface lifestyle through shaping the hydrophobic bristles that prevent water striders from wetting and allow them to exploit water surface tension.
Keywords: achaete–scute complex, water locomotion, adaptation, bristles, water striders, evodevo
1. Introduction
The semi-aquatic bugs (Hemiptera: Gerromorpha) are freshwater or marine insects that occupy various water surface niches worldwide [1–4]. These insects are thought to have derived from a terrestrial ancestor that evolved the ability to stand and generate efficient movement on the water [5]. Early-diverging lineages occupy transitional zones and walk both on land and water, whereas derived lineages evolved rowing as a novel mode of locomotion on the open-water surface [1,4]. Water surface invasion is directly associated with the ability of these insects to support their body weight on the water–air interface by exploiting surface tension [5–7]. An important adaptive morphological trait that has been critical to this transition is the evolution of hydrophobic leg hairs, whose density, morphology and orientation contribute to water repellency [6,8,9]. In addition, the complex body hair layers confer hydrophobicity to the semi-aquatic bugs and protect the animal from wetting and drowning. The body of the semi-aquatic bugs is generally covered by two hair layers [10]: a dense layer of microtrichiae (short hairs) close to the body surface is thought to play the role of waterproofing in case of submergence, whereas stiff macrotrichiae (long hairs) are thought to play the role of rainproofing [11]. The semi-aquatic bugs maintain the anti-wetting properties of the hair architecture through meticulous care using grooming combs consisting of rows of stiff hairs that are present on the distal tibiae [12]. In spite of their ecological importance, the nature of the hairs covering the leg and those forming the grooming combs remains elusive. These hairs might alternatively be innervated mechanosensory structures (bristles) or non-innervated cuticular projections (trichomes) [13].
In the fly Drosophila, the four genes of the achaete–scute family encode basic helix–loop–helix factors that are master regulators of bristle development [14,15]. Achaete (ac), scute (sc) and lethal of scute (l'sc) have redundant proneural functions and promote the formation of neural precursors, whereas asense (ase) is expressed in neuronal precursors. Loss-of-function mutant flies for ac and sc display a notum devoid of bristles [16]. Similarly, loss of l'sc leads to the loss of body bristles whereas ectopic expression of l'sc in the fly notum produces additional bristles [17]. All four genes are intronless, oriented in the same orientation and clustered in a 100 kb region containing numerously shared and interspersed cis-regulatory elements [18,19]. Genetic re-arrangements within the complex lead to mutant phenotypes owing to disruption of the cis-regulatory organization [20].
The ancestral achaete-scute complex (AS-C) in insects contained two genes: an ase gene and an achaete–scute homologue (ASH) gene which has undergone independent duplications in the Diptera [21]. In the flour beetle Tribolium castaneum, Tc-ASH is required to promote neural precursor formation and could play the role of multiple duplicated proneural ac–sc genes present in species such as Drosophila [22]. Moreover, the expression pattern of Tc-ase is highly conserved between Drosophila and Tribolium suggesting a conserved ancestral function for asense genes [22]. In addition to the conservation of their function, ac–sc genes show a conserved genomic structural organization between Diptera and Coleoptera [21]. It is widely thought that the organization of the complex and the presence of shared cis-regulatory regions prevent separation of the genes.
Given the key role that hydrophobic bristles play in water strider biology, we hypothesized that changes to the ac–sc gene family could have occurred during the evolution of the Gerromorpha lineage. To address this question, we investigated the genomic organization of ac-sc genes in 14 species of the Gerromorpha in comparison with other insect lineages. We generated and compiled a unique dataset of AS-C proteins across insects, including available transcriptomes and genomes of early-diverging lineages. Furthermore using RNA interference (RNAi), we examined the role of this locus in the development of hydrophobic bristles in multiple species of Gerromorpha. Our findings suggest that the gene ase was lost in the Gerromorpha lineage and the only remaining member of the ac–sc family plays a key role in the formation of various bristles in this group of insects.
2. Material and methods
(a). Data collection
Phylogenetic markers were identified in available genomes or transcriptomes by tBLASTn using a set of selected Drosophila melanogaster genes as a probe. Species names are indicated by the following prefixes Aae: Aedes aegypti, Aca: Aplysia californica, Aga: Anopheles gambiae, Ame: Apis mellifera, Apa: Aquarius paludum, Bge: Blattella germanica, Bmo: Bombyx mori, Bta: Bemisia tabaci, Caq: Catajapyx aquilonaris, Cca: Ceratitis capitata, Cle: Cimex lectularius, Cpa: Cylindrostethus palmaris, Csa: Cupiennus salei, Dci: Diaphorina citri, Dma: Daphnia magna, Dme: Drosophila melanogaster, Dpo: Dendroctonus ponderosae, Dpu: Daphnia pulex, Eda: Ephemera danica, Eaf: Eurytemora affinis, Foc: Frankliniella occidentalis, Gbu: Gerris buenoi, Hha: Halyomorpha halys, Hst: Hydrometra stagnorum, Htu: Husseyella turmalis, Hvi: Homalodisca vitripennis, Hvu: Hydra vulgaris, Lde: Leptinotarsa decemlineata, Ldi: Limnoporus dissortis, Lfr: Limnogonus franciscanus, Lfu: Ladona fulva, Llu: Limnephilus lunatus, Lmi: Locusta migratoria, Mam: Microvelia americana, Mdo: Musca domestica, Mex: Medauroidea extradentata, Mfu: Mesovelia furcata, Mhe: Metrobates hesperius, Mlo: Microvelia longipes, Mmu: Mesovelia mulsanti, Mmus: Mus musculus, Nlu: Nilaparvata lugens, Nvi: Nasonia vitripennis, Oci: Orchesella cincta, Ocu: Oiovelia cunucunumana, Ofa: Oncopeltus fasciatus, Pac: Paravelia conata, Pap: Pyrrhocoris apterus, Pba: Pogonomyrmex barbatus, Pbr: Platyvelia brachialis, Pbu: Paravelia bullialata, Pdu: Platynereis dumerilii, Phu: Pediculus humanus, Pve: Pachypsylla venusta, Ran: Rhagovelia antilleana, Rob: Rhagovelia obesa, Rpr: Rhodnius prolixus, Rze: Rhagoletis zephyria, Sma: Strigamia maritima, Smi: Stegodyphus mimosarum, Sst: Stridulivelia strigosa, Ste: Stridulivelia tersa, Tca: Tribolium castaneum, Tlo: Triops longicaudatus, Xla: Xenopus laevis, Zne: Zootermopsis nevadensis. The novel sequences generated for this analysis have been deposited in the EMBL database with specific accession numbers (electronic supplementary material, table S1).
(b). Phylogenetic analysis
Nucleotide sequences were aligned with MUSCLE [23], manually adjusted and selected blocks were used for phylogenetic reconstruction. Maximum-likelihood (ML) searches were performed using RAxML v.8 [24] under the site-homogeneous LG+Γ model. One hundred bootstrap replicates were conducted for support estimation. Bayesian phylogenetic analyses were performed using MrBayes 3.2 [25] under the GTR+Γ model. We ran two chains for at least 1 000 000 generations and removed the first 250 000 generations as burn-in. The different nucleotide sequence alignments and tree files are available from the Dryad Digital Repository at: http://dx.doi.org/10.5061/dryad.rc454pc [26].
(c). Ancestral state reconstruction
Ancestral reconstruction of the number of grooming combs on the different legs was performed using R software. ML methods were adapted to discrete characters (ace, package ape [27]) and the package phytools [28] was used for generating graphical representations. The simplest model ‘ER’, with equal transition rates across all categories, was the best both with Akaike information criterion and likelihood comparisons.
(d). Microscopy
For transmission electron microscopy (TEM), adult tarsi were transversally cut into several pieces and fixed in 2% glutaraldehyde in 75 mM sodium cacodylate buffer (pH = 7.3) overnight at 4°C. Samples were washed in cacodylate buffer and post-fixed in 1% osmium tetroxide for 1 h. Samples were then dehydrated in ethanol and embedded in epoxy resin. Samples were sectioned on a Leica UC7 ultramicrotome, stained with uranyl acetate and lead citrate and imaged with a Philips CM120 TEM at 80 kV. For scanning electron microscopy (SEM), adult bugs were fixed in 4% paraformaldehyde : heptane (ratio 1 : 3) for 20 min at room temperature. Samples were examined by using a Hitachi S800 SEM.
(e). In situ hybridization
Dissected embryos were fixed in 4% paraformaldehyde : heptane (ratio 1 : 3) for 20 min at room temperature, washed several times in cold methanol and then rehydrated through a methanol series to phosphate buffered saline-Tween 20 0.05%. Embryos were prehybridized for an hour at 60°C in hybridization buffer (for composition, see [29]) prior to addition of digoxigenin (DIG)-labelled RNA probe overnight at 60 °C. Blocking step was performed in 1% bovine serum albumin prior to incubation with anti-DIG antibody coupled with alkaline phosphatase for 2 h at room temperature. Embryos were washed several times before revelation with nitro-blue tetrazolium chloride/5-bromo-4-chloro-3′-indolyphosphate in alkaline phosphatase buffer. Embryos were mounted on slides in Hoyer's medium and photographed on a Zeiss Axio observer microscope. The polymerase chain reaction primers we used in this study are listed in the electronic supplementary material, table S2.
(f). Parental RNAi
Gene knockdown of ASH using parental RNAi was conducted following the protocol described in [29]. To obtain stronger phenotypes, Mesovelia mulsanti embryos were placed at 30–31°C which allowed them to develop faster. Technically, this method shortens the delay between the injection of the double-stranded RNA and the late expression of ASH transcripts.
3. Results and discussion
(a). Nature and structure of leg hairs
The legs of water striders are covered by layers of hairs of different sizes, which form a cushion between the leg and the water surface thus preventing the leg from breaking surface tension [6,9,10,30]. Although some authors informally refer to these structures as hairs/setae [5,6,8,9], bristle-like setae [31] or bristles [32], their cellular origin is unknown. In the water strider Gerris buenoi (Gerridae), leg tarsi are covered with at least two types of ‘hairs’ based on their size and that are regularly arranged along the leg (figure 1a). This feature is shared by the microveliid Microvelia americana (figure 1b) but not the mesoveliid Me. mulsanti where only one ‘hair’ type can be detected (figure 1c). Differences in the density of hairs are also observed across species (figure 1a–c). SEM images show a cuticle protuberance at the base of each hair (figure 1f), reminiscent of the socket cell of innervated bristles in insects [33] (figure 1g). TEM imaging on leg hairs revealed the presence of an outer dendritic segment underneath every single hair (figure 1d,e,g and electronic supplementary material, S1). We therefore conclude that the numerous structures found on Gerromorpha legs are innervated bristles.
Figure 1.

Hydrophobic leg hairs are bristles. Three Gerromorpha species have been investigated: Gerris buenoi (a), Microvelia americana (b) and Mesovelia mulsanti (c–f). All species have hydrophobic bristles on legs that are clearly visible using SEM (a–c). In Me. mulsanti, high magnification pictures show the socket cell at the base of the bristle, and the ridges of the bristle resulting from the inner actin organization (d–f). In Me. mulsanti, TEM pictures show that bristles are innervated structures with an outer dendritic segment underneath each single bristle (d,e). (g) Schematic representation of a bristle and associated structures. cu: cuticle, ds: dendritic sheath, EDM: electron-dense materials, od: outer dendritic segment, mt: microtubule, s: shaft.
(b). Diversification of grooming combs in Gerromorpha
Grooming, or active cleaning of body parts with specialized grooming structures, is a critical activity in the life of insects [34]. Grooming allows care of the body surface through the removal of contaminants [35,36], parasitoids [37] and pathogens [38]. Grooming also facilitates the distribution of hydrocarbons and antiseptic secretions on the body surface [39,40], as well as displacement behaviour in stressful conditions [41]. In Gerromorpha, grooming combs are of critical importance as they are used to keep the hair layers tidily arranged to prevent the leg from breaking water surface tension and the bug from drowning [5,12] (electronic supplementary material, videos S1–S3).
We found tibial grooming combs to be present in all semi-aquatic bugs we have investigated in this study. However, the number of grooming combs varies between legs in a given individual, as well as across species (figure 2). First instar nymphs have two grooming combs on the foreleg and one grooming comb in midleg tibiae in all Gerromorpha, except in Hydrometra stagnorum which has one grooming comb in the foreleg and none in the midleg. The reconstruction of the plesiomorphic state predicts that the common ancestor of Gerromorpha had two grooming combs in the forelegs and a single one in the midlegs. This result suggests that the loss of one of the two grooming combs of the forelegs and that of the midlegs occurred in the lineage leading to Hy. stagnorum (figure 2). This species is known to preferentially live on solid substrates [4], which might explain the loss of grooming combs.
Figure 2.

Evolutionary history of grooming combs in Gerromorpha. Presence/absence of grooming combs are mapped onto a phylogenetic tree of representative species. The phylogenetic trees, obtained through maximum-likelihood and Bayesian reconstructions, were conducted under the LG + Γ and the GTR + Γ model, respectively. Support values are shown in the electronic supplementary material, figure S2.
The presence/absence pattern of the hindleg grooming comb is more labile across the species we investigated. The eight Gerromorpha species that lack grooming combs on the hindleg are not clustered in the phylogenetic tree, which argues for independent events during the course of evolution. However, statistical tests did not support either of the two possible hypotheses over the other: the ancestor had a hindleg grooming comb versus the ancestor lacks a hindleg grooming comb. In the event of hindleg grooming comb present in the ancestor, at least three independent losses would have occurred during the diversification of Gerromorpha (figure 2). Conversely, if the ancestor was missing hindleg combs, at least three independent gains took place in semi-aquatic bugs. According to published work in true bugs (Hemiptera: Heteroptera), the presence of tibial grooming combs on all legs is regarded as the most plesiomorphic state [42]. Moreover, the loss of grooming combs on the different legs appears to be a very common trend in terrestrial bugs [42]. It is therefore tempting to envision that the last common ancestor of extant Gerromorpha had a hindleg comb that has been independently lost in several lineages.
(c). Conservation of achaete-scute complex genomic architecture
The structures covering the legs of the Gerromorpha being innervated bristles, it is reasonable to consider bristle specification genes as candidates to explain the specific characteristics of these leg bristles. The analysis of the AS-C gene complex is essential to understanding the contribution that proneural genes have made to the evolution of epidermal structures in insects. Whereas AS-C genes have been extensively investigated in flies, little is known about the molecular evolution of these genes across insects. To evaluate how conserved the extended genomic locus is in other insect genomes, we conducted both content and synteny analyses of this locus in two hemipterans (G. buenoi and Homalodisca vitripennis), one thysanopteran genome (Frankliniella occidentalis) and one blattodean genome (Blattella germanica). Strikingly, we failed to detect asense in the genome of G. buenoi, Ho. vitripennis and F. occidentalis species. To confirm the absence of asense in these genomes, we performed synteny analysis, and we took advantage of the fact that the extended genomic locus of the whole AS-C complex exhibited a set of conserved features across the Holometabola (figure 3). In the fly D. melanogaster, the four AS-C genes are flanked by the genes yellow (y) and Cytochrome P450–4g1 (Cyp4g1) and are present in the same position in many other insects [21] (figure 3). First, the two genes y and Cyp4g1 that constitute the boundary markers in the Holometabola have not been identified in the vicinity of the AS-C in the three non-holometabolan genomes we investigated (figure 3). Conversely, we identified several other markers, especially major facilitator superfamily genes, whose homologues are present within the AS-C locus in most insect species (figure 3). Second, we showed that genes which flank the position of the missing asense gene (ASH in the 5′ region; major facilitator superfamily genes in the 3′ region) are conserved across insects. This result suggests that asense was lost in the lineage leading to the Hemiptera (G. buenoi, Ho. vitripennis) and the Thysanoptera (F. occidentalis), and this loss is not associated with a reshuffling of the surrounding genomic locus.
Figure 3.

Syntenic organization of the achaete–scute locus in representative insects. The D. melanogaster ac–sc locus, which is used as a reference, is arbitrarily limited to 12 genes with specific colours. Single-copy genes depicted with colour combinations in other species mean that duplication(s) of this gene occurred in the lineage leading to the Diptera. Yellow bar: gene duplications specific to D. melanogaster; red star: loss of asense.
(d). Evolution of the achaete–scute complex in the Hemiptera
To further confirm this lineage-specific loss of asense, we searched for putative orthologues of asense in a broader sampling among insects. Using publicly available insect sequence datasets [43,44] along with in-house generated transcriptomes of 14 species of the Gerromorpha, we assembled sequence data of 77 AS-C genes and performed ML and Bayesian phylogenetic reconstruction. This sequence-based phylogenetic reconstruction identified both orthologues of ASH and asense genes in most insect lineages (figure 4, electronic supplementary material, table S3). Furthermore, both ASH and asense clades include sequences from early-diverging lineages of insects and hexapods but exclude non-hexapod sequences. Non-hexapod genes grouping closely to both ASH and asense clades are probable pro-orthologues of the ASH and asense duplicated genes. We conclude with confidence that the last common ancestor of the extant insects possessed at least two AS-C genes, corresponding to the precursors of the ASH and asense clades. This transcriptome analysis, together with the characterization of the AS-C genomic locus, suggests that the asense gene has been lost in the lineage leading to both Hemiptera (true bugs) and Thysanoptera (thrips) (Condylognatha). Both of these groups have sucking mouthparts and their monophyletic relationship was recently confirmed [45]. This finding represents, to our knowledge, the first reported case of asense gene loss in insects.
Figure 4.

Phylogram of the 77 taxon analyses obtained through maximum-likelihood and Bayesian reconstructions were conducted using LG + Γ and GTR + Γ models, respectively. (a) Full tree depicting all insect taxa used in this analysis. (b) Zoom on Gerromorpha clade. Support values are shown in the electronic supplementary material, figure S3. Scale bar indicates a number of changes per site.
(e). Expression of achaete-scute homologue in developing Gerromorpha embryos
To determine the role of the unique AS-C gene during development in the Gerromorpha, we investigated the expression of ASH by in situ hybridization in G. buenoi, Me. mulsanti and Mi. americana. At mid-embryogenesis, Mmu-ASH is expressed in ectodermal cell clusters throughout the central nervous system (CNS), especially in the head and abdominal segments (figure 5a,b). These cell clusters are likely to correspond to the presumptive neural precursors, suggesting that Mmu-ASH acts as a proneural gene in Me. mulsanti. Because we also detected CNS expression for Mam-ASH and Gbu-ASH (data not shown), it is reasonable to conclude that ASH has a proneural function in all Gerromorpha. These expression domains mirror those of ac-sc genes known in Coleoptera [22], Diptera [46,47] and Lepidoptera [48].
Figure 5.
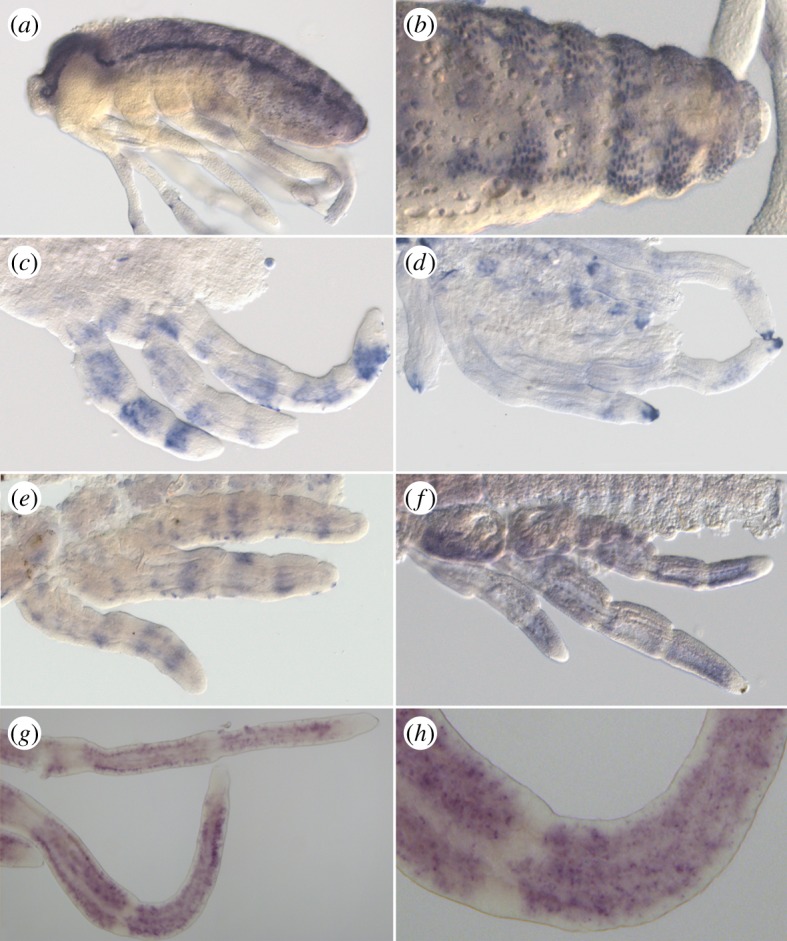
In situ hybridization showing ASH expression pattern. (a,b) Staining of ASH in Mesovelia mulsanti. ASH is expressed during central nervous system development in ectodermal cell clusters (a) that are progressively restricted to neural precursors (b). (c,d) Staining of ASH in Microvelia americana. In post-katatrepsis embryos, ASH is expressed in discrete large transverse bands along the leg (c). This staining is restricted to the distal tip of the tarsus in later stages (d). Staining of ASH in Gerris buenoi (e–h). Similarly to Mi. americana embryos, ASH expression pattern is dynamic in legs in sharp transverse bands (e), in larger bands (f), then in spotty domains (g,h).
We also detected ASH expression in transverse stripes in all legs in post-katatrepsis embryos. In G. buenoi, the stripes are first sharp and narrow (figure 5e) before the domain of expression expands throughout leg segments (figure 5f). The expression of Gbu-ASH eventually becomes restricted to numerous dots in the legs (figure 5g,h), each dot likely to prefigure the position of a future bristle. This expression pattern recapitulates only the early, but not the late, expression of ac and sc found in D. melanogaster legs [49]. In D. melanogaster, the predominant expression from 3 to 5 h after puparium formation is in discontinuous transverse stripes that encircle the tarsal segments. At approximately 5 h ac and sc expression in longitudinal stripes begins to appear in conjunction with the transverse stripes, and by 6 h the longitudinal stripes in each segment replace the transverse stripes [49]. We have not detected any expression in longitudinal stripes either in G. buenoi, Me. mulsanti or Mi. americana.
In late embryos, the expression of ASH is restricted to discrete regions such as the distal tip of the tarsus (Mam-ASH, figure 5d) that prefigures the location of the future claw.
(f). Achaete-scute homologue gene regulates neural precursors in the Gerromorpha
We used RNAi to deplete ASH function in the embryos. In M. mulsanti, we observed altered bristle development in 82% of ASH RNAi-treated embryos (n = 32 out of 39), ranging from a weak to a dramatic reduction in the number of bristles in the thorax and the abdomen (figure 6a–d), as well as in the legs (data not shown) in late embryos. The variability we observe in the knockdown phenotype is a consequence of the RNAi technique as reported in previous studies in the Gerromorpha [29,50,51]. In Mi. americana, only mild alterations of bristle development have been obtained (40% of embryos affected, n = 12 out of 30), showing missing bristles in the first two thoracic segments (figure 6e,f). In G. buenoi, we observed an altered phenotype in 97% of ASH RNAi-treated embryos (n = 208 out of 214), consisting of reduced bristle number on the whole body (figure 6g–j). ASH depletion, even subtle, consistently affected abdominal bristles (figure 6g,h), thoracic and head bristles secondarily and leg bristles (figure 6i,j). The variability in RNAi leg phenotype also ranges from mild to strong (electronic supplementary material, figure S4). The bristles that are missing or altered are not consistently the same ones between RNAi-treated embryos. However, all bristles can be affected independently of their location. It is reasonable to hypothesize that ASH controls the specification of all leg bristles and that the variability we see results from the partial efficiency of knockdown methods to silence the ASH gene.
Figure 6.
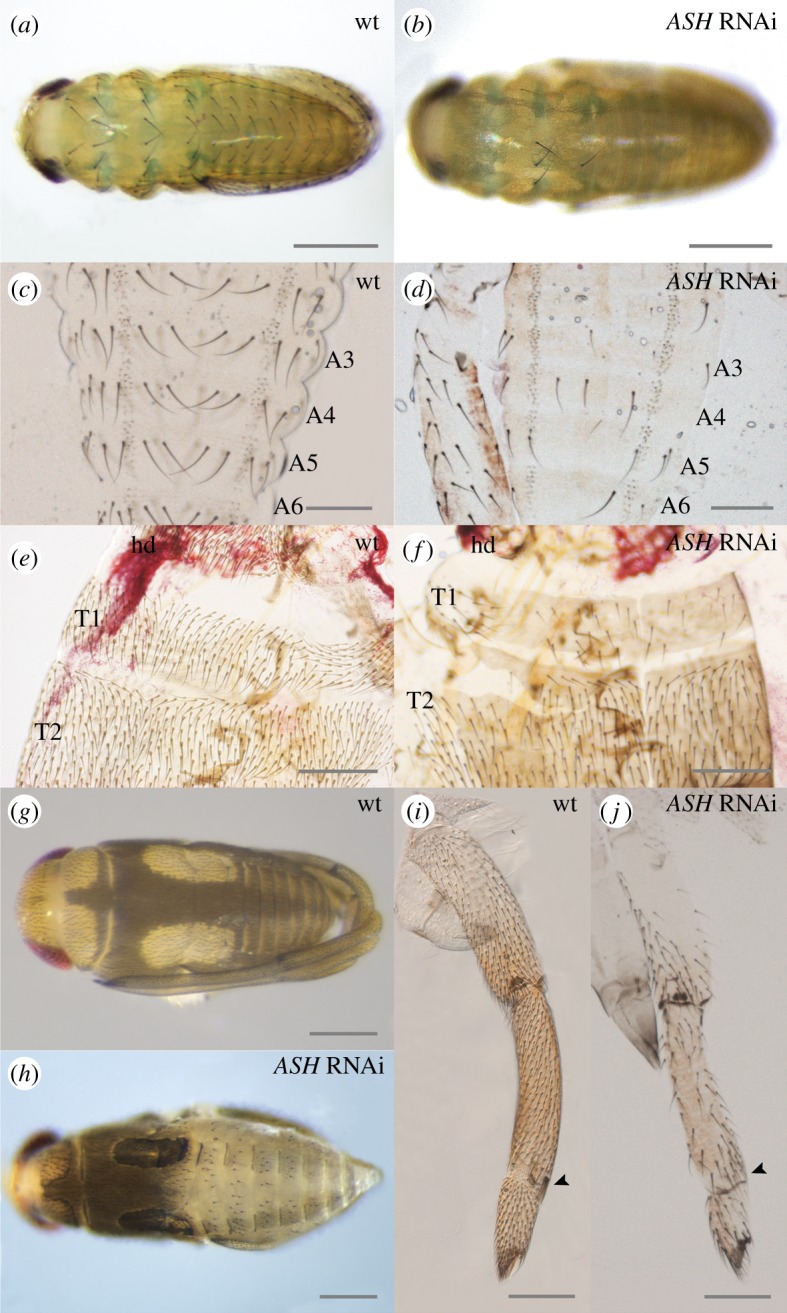
Phenotypes observed in ASH RNAi individuals in Gerromorpha. (a–d) Malformation of bristles after ASH RNAi in Mesovelia mulsanti. (e,f) Malformation of thoracic bristles after ASH RNAi in Microvelia americana. (g–j) Malformation of bristles, grooming combs, and claws after ASH RNAi in Gerris buenoi. Arrowheads show the location of grooming combs; A: abdominal segment; hd: head, T: thoracic segment. The scale bar indicates 200 m.
Another important role of ASH, as revealed by RNAi, is the control of grooming comb development in all legs. Whereas wild-type G. buenoi embryos have two grooming combs on the forelegs, and one comb on mid and hindlegs (figure 2), ASH RNAi-treated embryos are devoid of grooming combs altogether (figure 6j and electronic supplementary material, figure S4). Previous studies have shown that the knockdown of the gene Ultrabithorax (Ubx) does not affect grooming combs on any of the legs in the Gerridae [50,52]. However, Ubx RNAi leads to the development of an ectopic hindleg comb in Microvelia and Mesovelia both of which lack hindleg combs otherwise [50]. This suggests that the absence of hindleg grooming combs in some Gerromorpha is owing to ASH repression by Ubx. Further experiments would be required to test this possible interaction between Ubx and ASH genes.
ASH RNAi-treated embryos also lack claws (figure 6j). This phenotype is observed for the first time in the context of an altered ac–sc complex. In Drosophila, mutation of one or several genes of the ac–sc complex does not affect the tarsal claw. However, the proneural gene amos, whose postembryonic expression prefigures the anlage of the innervated tarsal claw, is thought to be involved in the formation of sensory organ precursors in the tarsal claw [53]. Previous studies have shown that amos prevents bristle formation through the repression of scute function in Drosophila [54]. As a future direction, it might be interesting to test whether the genes ASH and amos act in the same developmental network during tarsal claw development in Gerromorpha.
4. Conclusion
The evolution of a higher density of hydrophobic leg hairs accompanied the invasion of water surface by semi-aquatic bugs. Variation in the density of bristles and the number of grooming combs found among the Gerromorpha might have evolved as an adaptation to the diversity of habitats these bugs live in [4]. We have shown these leg hairs to be bristles representing innervated structures that are known to act as mechanoreceptors. The massive number of leg bristles is associated with the semi-aquatic bugs' lifestyle that involves detection of prey trapped on the water surface, as well as detection of predators. We therefore hypothesize that the increase in leg bristle density during the course of evolution could have played a dual role: exploiting water surface tension for locomotion and developing a higher sensitivity to detect various activities through vibrations of the water–air interface including prey and predator attacks [55]. Finally, we uncovered the pivotal role of the gene ASH in bristle and grooming comb specification in Gerromorpha. We identified unexpected changes at the genomic ASH locus. Our results suggest that changes in the regulation of the gene ASH could have been associated with the adaptation of semi-aquatic bugs to the water surface and their subsequent diversification.
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
Acknowledgements
We thank Aidamalia Vargas Lowman, Antonin Crumière, Augustin le Bouquin and William Toubiana for comment and helpful discussions. We thank Séverine Viala for help with scanning electron microscopy, Antoine Melet and Marie Bernard for insect husbandry. Electron microscopy studies have been done at the ‘Centre Technologique des Microstructures’—Claude Bernard University of Lyon. We thank ERC for providing financial support to the work through the ERC-CoG no. 616346 to A.K.
Data accessibility
Data is available from the Dryad Digital Repository at: http://dx.doi.org/10.5061/dryad.rc454pc [26].
Authors' contributions
A.K. and C.F. conceived the work, C.F. and A.D. performed experiments and D.A. generated genomic and transcriptomic data. A.K. and C.F. wrote the manuscript.
Competing interests
We declare we have no competing interests.
Funding
This work was funded by an ERC-CoG no. 616346 to A.K.
References
- 1.Andersen NM. 1982. The semiaquatic bugs (Hemiptera: Gerromorpha). Klampenborg, Denmark: Scandinavian Science Press Ltd. [Google Scholar]
- 2.Cheng L. 2004. Marine insects and the sea-skater Halobates (Hemiptera: Gerridae). In Encyclopedia of entomology (ed. Capinera JL.), pp. 1346–1349. Dordrecht, The Netherlands: Springer. [Google Scholar]
- 3.Polhemus JT, Polhemus DA. 2007. Global diversity of true bugs (Heteroptera; Insecta) in freshwater. In Freshwater animal diversity assessment. 198 (eds Balian EV, Lévêque C, Segers H, Martens K), pp. 379–391. Dordrecht, The Netherlands: Springer. [Google Scholar]
- 4.Crumiere AJJ, Santos ME, Semon M, Armisen D, Moreira FFF, Khila A. 2016. Diversity in morphology and locomotory behavior is associated with niche expansion in the semi-aquatic bugs. Curr. Biol. 26, 3336–3342. ( 10.1016/j.cub.2016.09.061) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Andersen NM. 1976. A comparative study of locomotion on the water surface in semiaquatic bugs (Insecta, Hemiptera, Gerromorpha). Videnskabelige Meddelelser Dansk Naturhistorisk Forening 139, 337–396. [Google Scholar]
- 6.Gao X, Jiang L. 2004. Biophysics: water-repellent legs of water striders. Nature 432, 36 ( 10.1038/432036a) [DOI] [PubMed] [Google Scholar]
- 7.Hu DL, Bush JWM. 2010. The hydrodynamics of water-walking arthropods. J. Fluid Mech. 644, 5–33. ( 10.1017/S0022112009992205) [DOI] [Google Scholar]
- 8.Xue Y, Yuan H, Su W, Shi Y, Duan H. 2014. Enhanced load-carrying capacity of hairy surfaces floating on water. Proc. R. Soc. A 470, 20130832 ( 10.1098/rspa.2013.0832) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wang Q, Yao X, Liu H, Quere D, Jiang L. 2015. Self-removal of condensed water on the legs of water striders. Proc. Natl Acad. Sci. USA 112, 9247–9252. ( 10.1073/pnas.1506874112) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Andersen NM. 1977. Fine structure of the body hair layers and morphology of the spiracles of semiaquatic bugs in relation to life on the water surface. Videnskabelige Meddelelser Dansk Naturhistorisk Forening. 140, 7–37. [Google Scholar]
- 11.Thorpe W, Crisp D. 1947. Studies on plastron respiration III. The orientation responses of Apelocheirus [Hemiptera, Aphelocheiridae (Naucoridae)] in relation to plastron respiration; together with an account of specialized pressure receptors in aquatic insects. J. Exp. Biol. 24, 310–328. [DOI] [PubMed] [Google Scholar]
- 12.Rensing L. 1962. Beiträge zur vergleichenden Morphologie, Physiologie und Ethologie der Wasserläufer (Gerroidea). Zool Beitr NF. 7, 447–485. [Google Scholar]
- 13.Chapman RF. 2013. Mechanoreception. In The insects: structure and function, 5th edn (eds Simpson SJ, Douglas AE), pp. 738–770. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 14.Calleja M, Renaud O, Usui K, Pistillo D, Morata G, Simpson P. 2002. How to pattern an epithelium: lessons from achaete-scute regulation on the notum of Drosophila. Gene 292, 1–12. ( 10.1016/S0378-1119(02)00628-5) [DOI] [PubMed] [Google Scholar]
- 15.Hartenstein V. 2005. Development of insect sensilla. In Comprehensive molecular insect science. 1 (eds Gilbert L, Iatrou K, Gill S), pp. 379–419. New York, NY: Elsevier. [Google Scholar]
- 16.Campuzano S, et al. 1985. Molecular genetics of the achaete-scute gene complex of D. melanogaster. Cell 40, 327–338. ( 10.1016/0092-8674(85)90147-3) [DOI] [PubMed] [Google Scholar]
- 17.Hinz U, Giebel B, Campos-Ortega JA. 1994. The basic-helix-loop-helix domain of Drosophila lethal of scute protein is sufficient for proneural function and activates neurogenic genes. Cell 76, 77–87. ( 10.1016/0092-8674(94)90174-0) [DOI] [PubMed] [Google Scholar]
- 18.Gomez-Skarmeta JL, Rodriguez I, Martinez C, Culi J, Ferres-Marco D, Beamonte D, Modolell J. 1995. Cis-regulation of achaete and scute: shared enhancer-like elements drive their coexpression in proneural clusters of the imaginal discs. Genes Dev. 9, 1869–1882. ( 10.1101/gad.9.15.1869) [DOI] [PubMed] [Google Scholar]
- 19.Modolell J, Campuzano S. 1998. The achaete-scute complex as an integrating device. Int. J. Dev. Biol. 42, 275–282. [PubMed] [Google Scholar]
- 20.Ruiz-Gomez M, Modolell J. 1987. Deletion analysis of the achaete-scute locus of Drosophila melanogaster. Genes Dev. 1, 1238–1246. ( 10.1101/gad.1.10.1238) [DOI] [PubMed] [Google Scholar]
- 21.Negre B, Simpson P. 2009. Evolution of the achaete-scute complex in insects: convergent duplication of proneural genes. Trends Genet. 25, 147–152. ( 10.1016/j.tig.2009.02.001) [DOI] [PubMed] [Google Scholar]
- 22.Wheeler SR, Carrico ML, Wilson BA, Brown SJ, Skeath JB. 2003. The expression and function of the achaete-scute genes in Tribolium castaneum reveals conservation and variation in neural pattern formation and cell fate specification. Development 130, 4373–4381. ( 10.1242/dev.00646) [DOI] [PubMed] [Google Scholar]
- 23.Edgar RC. 2004. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797. ( 10.1093/nar/gkh340) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Stamatakis A. 2014. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313. ( 10.1093/bioinformatics/btu033) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Huelsenbeck JP, Ronquist F. 2001. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17, 754–755. ( 10.1093/bioinformatics/17.8.754) [DOI] [PubMed] [Google Scholar]
- 26.Finet C, Decaras A, Armisén D, Khila A. 2018. Data from: The achaete-scute complex contains a single gene that controls bristle development in the semi-aquatic bugs Dryad Digital Repository. ( 10.5061/dryad.rc454pc) [DOI] [PMC free article] [PubMed]
- 27.Paradis E, Claude J, Strimmer K. 2004. APE: analyses of phylogenetics and evolution in R language. Bioinformatics 20, 289–290. ( 10.1093/bioinformatics/btg412) [DOI] [PubMed] [Google Scholar]
- 28.Kembel SW, Cowan PD, Helmus MR, Cornwell WK, Morlon H, Ackerly DD, Blomberg SP, Webb CO. 2010. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 26, 1463–1464. ( 10.1093/bioinformatics/btq166) [DOI] [PubMed] [Google Scholar]
- 29.Khila A, Abouheif E, Rowe L. 2009. Evolution of a novel appendage ground plan in water striders is driven by changes in the Hox gene Ultrabithorax. PLoS Genet. 5, e1000583 ( 10.1371/journal.pgen.1000583) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hu DL, Chan B, Bush JW. 2003. The hydrodynamics of water strider locomotion. Nature 424, 663–666. ( 10.1038/nature01793) [DOI] [PubMed] [Google Scholar]
- 31.Westlake KP, Rowe L, Currie DC. 2000. Phylogeny of the water strider genus Rheumatobates (Heteroptera : Gerridae). Syst. Entomol. 25, 125–145. ( 10.1046/j.1365-3113.2000.00098.x) [DOI] [Google Scholar]
- 32.Sun T, Jiang L. 2010. Surface microstructures and nanostructures in natural systems. In Nanotechnology, volume 8: nanostructured surfaces. 8 (ed. Chi L.), pp. 401–426. Weinheim, Germany: WILEY-VCH Verlag GmbH & Co. KGaA. [Google Scholar]
- 33.Keil TA. 1997. Functional morphology of insect mechanoreceptors. Microsc. Res. Tech. 39, 506–531. ( 10.1002/(SICI)1097-0029(19971215)39:6%3C506::AID-JEMT5%3E3.0.CO;2-B) [DOI] [PubMed] [Google Scholar]
- 34.Dawkins R, Dawkins M. 1976. Hierarchical organization and postural facilitation: rules for grooming in flies. Anim. Behav. 24, 739–755. ( 10.1016/S0003-3472(76)80003-6) [DOI] [Google Scholar]
- 35.Elawami IO, Dent DR. 1995. The interaction of surface and dust particle-size on the pick-up and grooming behavior of the German cockroach Blattella germanica. Entomol. Exp. Appl. 77, 81–87. ( 10.1111/j.1570-7458.1995.tb01988.x) [DOI] [Google Scholar]
- 36.Hackmann A, Delacave H, Robinson A, Labonte D, Federle W. 2015. Functional morphology and efficiency of the antenna cleaner in Camponotus rufifemur ants. R. Soc. open sci. 2, 150129 ( 10.1098/rsos.150129) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Vincent CM, Bertram SM. 2010. Crickets groom to avoid lethal parasitoids. Anim. Behav. 79, 51–56. ( 10.1016/j.anbehav.2009.10.001) [DOI] [Google Scholar]
- 38.Reber A, Purcell J, Buechel SD, Buri P, Chapuisat M. 2011. The expression and impact of antifungal grooming in ants. J. Evol. Biol. 24, 954–964. ( 10.1111/j.1420-9101.2011.02230.x) [DOI] [PubMed] [Google Scholar]
- 39.Hefetz A, Soroker V, Dahbi A, Malherbe MC, Fresneau D. 2001. The front basitarsal brush in Pachycondyla apicalis and its role in hydrocarbon circulation. Chemoecology 11, 17–24. ( 10.1007/PL00001827) [DOI] [Google Scholar]
- 40.Böröczky K, Wada-Katsumata A, Batchelor D, Zhukovskaya M, Schal C. 2013. Insects groom their antennae to enhance olfactory acuity. Proc. Natl Acad. Sci. USA 110, 3615–3620. ( 10.1073/pnas.1212466110) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Root-Bernstein M. 2010. Displacement activities during the honeybee transition from waggle dance to foraging. Anim. Behav. 79, 935–938. ( 10.1016/j.anbehav.2010.01.010) [DOI] [Google Scholar]
- 42.Lis JA, Schaefer CW. 2005. Tibial combs in the Cydnidae (Hemiptera: Heteroptera) and their functional, taxonomic and phylogenetic significance. J. Zool. Syst. Evol. Res. 43, 277–283. ( 10.1111/j.1439-0469.2005.00328.x) [DOI] [Google Scholar]
- 43.Poelchau M, Childers C, Moore G, Tsavatapalli V, Evans J, Lee C-Y, Lin H, Lin J-W, Hackett K. 2015. The i5 k Workspace@ NAL—enabling genomic data access, visualization and curation of arthropod genomes. Nucleic Acids Res. 43, D714–D7D9. ( 10.1093/nar/gku983) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Yin C, et al. 2016. InsectBase: a resource for insect genomes and transcriptomes. Nucleic Acids Res. 44, D801–D807. ( 10.1093/nar/gkv1204) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Misof B, et al. 2014. Phylogenomics resolves the timing and pattern of insect evolution. Science 346, 763–767. ( 10.1126/science.1257570) [DOI] [PubMed] [Google Scholar]
- 46.Skeath JB, Carroll SB. 1992. Regulation of proneural gene expression and cell fate during neuroblast segregation in the Drosophila embryo. Development 114, 939–946. [DOI] [PubMed] [Google Scholar]
- 47.Wulbeck C, Simpson P. 2000. Expression of achaete-scute homologues in discrete proneural clusters on the developing notum of the medfly Ceratitis capitata, suggests a common origin for the stereotyped bristle patterns of higher Diptera. Development 127, 1411–1420. [DOI] [PubMed] [Google Scholar]
- 48.Galant R, Skeath JB, Paddock S, Lewis DL, Carroll SB. 1998. Expression pattern of a butterfly achaete-scute homolog reveals the homology of butterfly wing scales and insect sensory bristles. Curr. Biol. 8, 807–813. ( 10.1016/S0960-9822(98)70322-7) [DOI] [PubMed] [Google Scholar]
- 49.Orenic TV, Held LI, Paddock SW, Carroll SB. 1993. The spatial organization of epidermal structures: hairy establishes the geometrical pattern of Drosophila leg bristles by delimiting the domains of achaete expression. Development 118, 9–20. [DOI] [PubMed] [Google Scholar]
- 50.Refki PN, Armisen D, Crumiere AJ, Viala S, Khila A. 2014. Emergence of tissue sensitivity to Hox protein levels underlies the evolution of an adaptive morphological trait. Dev. Biol. 392, 441–453. ( 10.1016/j.ydbio.2014.05.021) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Santos ME, Berger CS, Refki PN, Khila A. 2015. Integrating evo-devo with ecology for a better understanding of phenotypic evolution. Brief Funct. Genomics 14, 384–395. ( 10.1093/bfgp/elv003) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Khila A, Abouheif E, Rowe L. 2014. Comparative functional analyses of ultrabithorax reveal multiple steps and paths to diversification of legs in the adaptive radiation of semi-aquatic insects. Evolution 68, 2159–2170. [DOI] [PubMed] [Google Scholar]
- 53.Goulding SE, zur Lage P, Jarman AP. 2000. Amos, a proneural gene for Drosophila olfactory sense organs that is regulated by lozenge. Neuron 25, 69–78. ( 10.1016/S0896-6273(00)80872-7) [DOI] [PubMed] [Google Scholar]
- 54.zur Lage PI, Prentice DR, Holohan EE, Jarman AP. 2003. The Drosophila proneural gene amos promotes olfactory sensillum formation and suppresses bristle formation. Development 130, 4683–4693. ( 10.1242/dev.00680) [DOI] [PubMed] [Google Scholar]
- 55.Armisen D, Refki PN, Crumiere AJ, Viala S, Toubiana W, Khila A. 2015. Predator strike shapes antipredator phenotype through new genetic interactions in water striders. Nat. Commun. 6, 8153 ( 10.1038/ncomms9153) [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Citations
- Finet C, Decaras A, Armisén D, Khila A. 2018. Data from: The achaete-scute complex contains a single gene that controls bristle development in the semi-aquatic bugs Dryad Digital Repository. ( 10.5061/dryad.rc454pc) [DOI] [PMC free article] [PubMed]
Supplementary Materials
Data Availability Statement
Data is available from the Dryad Digital Repository at: http://dx.doi.org/10.5061/dryad.rc454pc [26].