

Abstract
Chromobox (CBX) proteins are important components of epigenetic regulation complexes known to play key roles in hepatocellular carcinoma (HCC). Little is known about the function of distinct CBXs in HCC. To address this issue, the study investigated the roles of CBXs in the prognosis of HCC using ONCOMINE, UALCAN, Human Protein Atlas, Kaplan-Meier Plotter, c-BioPortal databases. Over expressions of 8 CBXs members were found to be significantly associated with clinical cancer stages and pathological tumor grades in HCC patients. Besides, higher mRNA expressions of CBX1/2/3/6/8 were found to be significantly associated with shorter overall survival (OS) in HCC patients, while higher mRNA expression of CBX7 was associated with favorable OS. Multivariate analysis also showed that high mRNA expressions of CBX1/2/3/6/8 were independent prognostic factors for shorter OS of HCC patients. Moreover, high mutation rate of CBXs (51%) was also observed in HCC patients, and genetic alteration in CBXs was associated with shorter OS and disease-free survival (DFS) in HCC patients. Taken together, these results indicated that CBX1/2/3/6/8 could be prognostic biomarkers for survivals of HCC patients.
Keywords: hepatocellular carcinoma, CBX, prognosis, ONCOMINE, Kaplan-Meier plotter
Introduction
Hepatocellular carcinoma (HCC) represents 90% of the primary liver cancer and is the fifth leading cancer that ranked second in cancer-related death worldwide. The incidence has increased rapidly over the years and the mortality rate is unfavorable. Studies suggested patients with HCC could not survive by more than 5 years [1,2]. Efforts have been made in studying the mechanisms of the development, progression and metastasis of HCC; however, the molecular characteristics of HCC, to date, remain unknown. By understanding the underlying pathogenesis and etiology of HCC would show light in discovering the advanced treatment and diagnostic biomarkers.
Certain cancer genetics such as mutations, small nucleotide polymorphism (SNP), translocations, deletions, and insertions could contribute to the genetic regulation of cancers [3]. Moreover, recent studies suggested aberration of epigenetic regulation also played pivotal roles in HCC regulations [4]. Polycomb group (PcG) complexes are epigenetic regulatory complexes, dysregulation of which has been associated with many cancer types [5–7]. Chromobox (CBX) family proteins are canonical components of PcG that regulate tumorigenesis and progression of many cancers including HCC by inhibition of cell differentiation and self-renewal of cancer stem cells [8,9]. A comprehensive study of distinct CBXs family members in HCC will help to uncover the molecular mechanisms involved in the development of HCC and could unveil novel prognostic and therapeutic targets for the devastating disease.
To date, 8 CBXs family proteins have been identified in human genomes. All of them take part in the regulation of heterochromatin, gene expression, and developmental programs. Based on the molecular structure of CBXs family proteins, they can be subdivided into two groups: HP1 group (includes CBX1, CBX3 and CBX5) and Pc group (includes CBX2, CBX4, CBX6, CBX7, and CBX8). HP1 group consists of an N-terminal chromodomain and a C-terminal chromoshadow domain, while Pc group contains only a conserved N-terminal chromodomain [10]. It is worth noting that functions of different CBXs family proteins are correlated with distinct regions of chromatin and are non-overlapped in embryonic stem cells [11–13].
Previous studies have found aberrant expressions and their prognostic values in some members of CBXs family. For instance, CBX4 was over-expressed in clinical tissues and multiple HCC cell lines. High expression of CBX4 was associated with tumor size, pathologic differentiation and poorer survival of patients, while down-regulation of CBX7 was found to be associated with shorter overall survival (OS) of HCC patients [14,15]. Nevertheless, the role of distinct CBXs family members remained unknown in the development and progression of HCC. In the present study, we addressed this problem by analyzing the expression and mutations of different CBXs family members and their relations with clinical parameters in HCC patients. Furthermore, we also analyzed the predicted functions and pathways of the mutations in CBXs as well as their 50 frequently altered neighbor genes.
RESULTS
Over-expression of different CBXs family members in patients with HCC
In order to explore the distinct prognostic and potential therapeutic value of different CBXs members in HCC patients, mRNA expression and protein expression were analyzed by ONCOMINE database (www.oncomine.org), UALCAN (http://ualcan.path.uab.edu), and Human Protein Atlas (https://www.proteinatlas.org). As were shown in Figure 1 and Table 1, mRNA expressions of 8 CBXs family members in 20 types of cancers were first measured and compared to normal tissues by ONCOMINE database. Significantly higher mRNA expressions of CBX1/3/5 were found in HCC tissues in multiple datasets. In Roessler Liver 2 dataset, CBX1 over-expression was found in HCC tissues compared with normal tissues with a fold change of 2.688 (p=4.33E-80) [16], while Wurmbach observed 1.781-fold increase in CBX1 mRNA expression in HCC samples (p=4.38E-8) [17] and Roessler found 2.405-fold increase in CBX1 mRNA expression in HCC tissues (p=1.37E-7) [16]. Significant up-regulation of CBX3 was also found in HCC tissues compared to normal tissues. The result from Roessler dataset showed that there were 1.888-fold (p=7.91E-63) and 1.704 fold (p=2.12E-5) increase in CBX3 mRNA expression in HCC tissues, respectively [16]. Similarly, in Roessler Liver dataset, 1.662-fold increase in CBX5 mRNA expression was found in HCC tissues compared to normal tissues (p=7.47E-6) [16]. Next, the mRNA expression patterns of 8 CBXs family members were further measured by UALCAN whose resources were based on level 3 RNA-seq and clinical data from 31 cancer types of TCGA database, which was different from ONCOMINE database. As was shown in Figure 2, mRNA expressions of 8 CBXs members were all found to be significantly up-regulated in primary HCC tissues compared to normal samples (all p<0.05).
Figure 1.
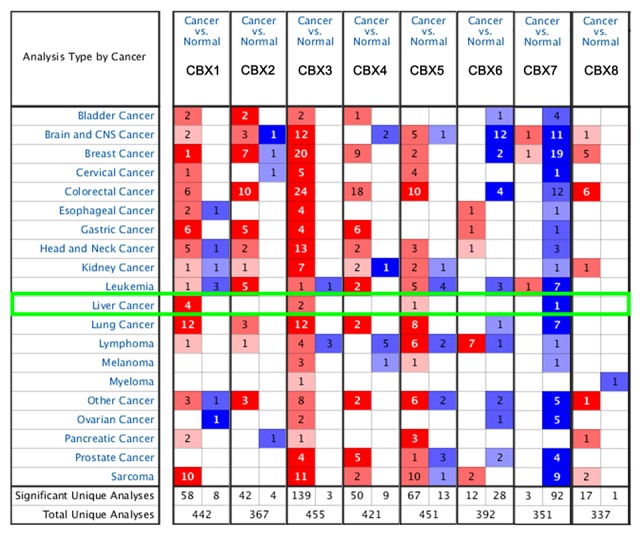
Transcriptional expression of CBXs in 20 different types of cancer diseases (ONCOMINE database). Difference of transcriptional expression was compared by students’ t-test. Cut-off of p value and fold change were as following: p value: 0.01, fold change: 1.5, gene rank: 10%, data type: mRNA.
Table 1. Significant changes of CBXs expression in transcription level between HCC and normal liver tissues (ONCOMINE).
| Types of HCC VS. Liver | Fold Change | P value | t-test | Ref | |
| CBX1 | |||||
| Hepatocellular Carcinoma | 2.688 | 4.33E-80 | 23.586 | Roessler Liver 2 [25] | |
| Hepatocellular Carcinoma | 1.781 | 4.38E-08 | 6.462 | Wurmbach Liver [26] | |
| Hepatocellular Carcinoma | 2.405 | 1.37E-07 | 6.649 | Roessler Liver [25] | |
| CBX3 | |||||
| Hepatocellular Carcinoma | 1.888 | 7.91E-63 | 19.952 | Roessler Liver 2 [25] | |
| Hepatocellular Carcinoma | 1.704 | 2.12E-05 | 4.71 | Roessler Liver [25] | |
| CBX5 | |||||
| Hepatocellular Carcinoma | 1.662 | 7.47E-06 | 5.161 | Roessler Liver [25] | |
HCC: hepatocellular carcinoma; CBX: chromobox
Figure 2.
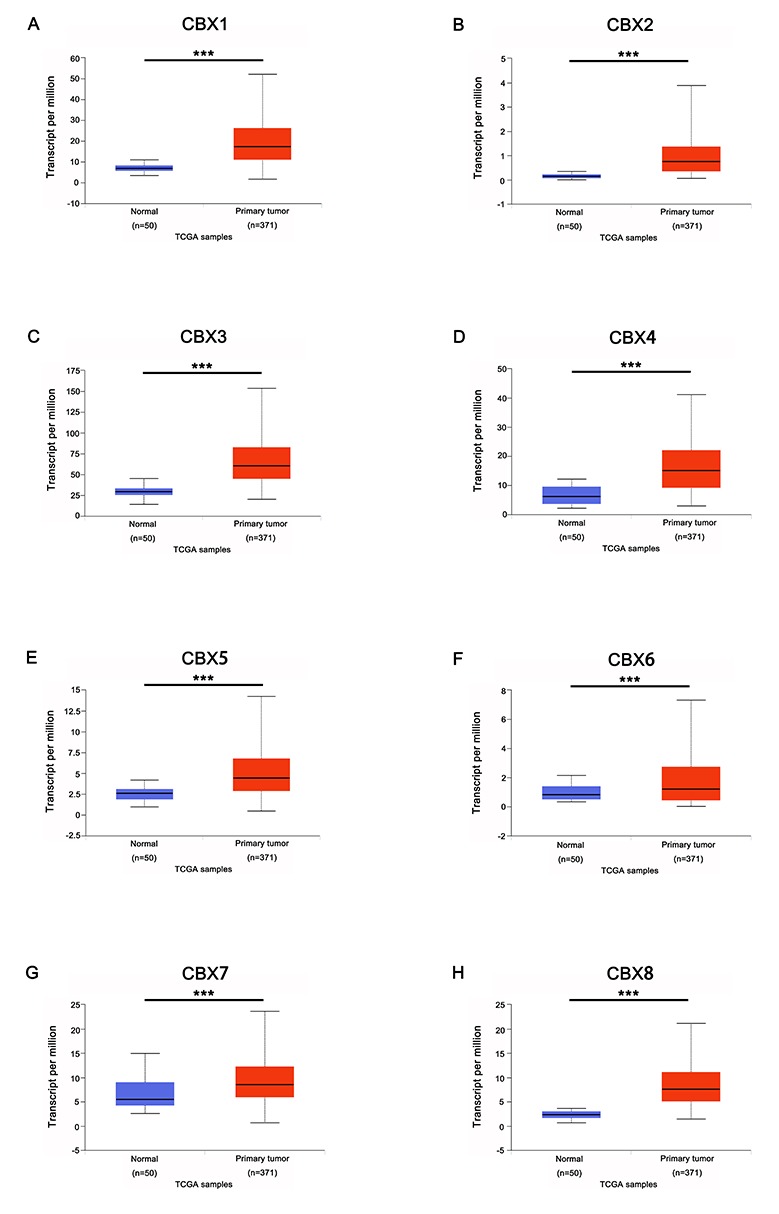
mRNA expression of distinct CBXs family members in HCC tissues and adjacent normal liver tissues (UALCAN). mRNA expressions of 8 CBXs family members were found to be over-expressed in primary HCC tissues compared to normal samples (A-H). *** p<0.001.
After examining the mRNA expression patterns of CBXs in HCC, we tried to explore the protein expression patterns of CBXs in HCC by the Human Protein Atlas. As was shown in Figure 3, CBX2/5/7/8 proteins were not expressed in normal liver tissues, whereas low and medium expressions of them were observed in HCC tissues (Figure 3B, E, G-H). In addition, low protein expressions of CBX1/3/4 were expressed in normal liver tissues, while medium and high protein expressions of them were observed in HCC tissues (Figure 3A, C-D). However, low protein expression of CBX6 was observed both at normal liver tissues and HCC tissues (Figure 3F). Taken together, our results showed that transcriptional and proteinic expressions of CBXs were over-expressed in patients with HCC.
Figure 3.

Representative immunohistochemistry images of distinct CBXs family members in HCC tissues and normal liver tissues (Human Protein Atlas). CBX2/5/7/8 proteins were not expressed in normal liver tissues, whereas their low and medium expressions were observed in HCC tissues (B, E, G-H). Low protein expressions of CBX1/3/4 were found in normal liver tissues, while their medium and high protein expressions were observed in HCC tissues (A, C-D). Low protein expression of CBX6 was observed both at normal liver tissues and HCC tissues (F).
Association of mRNA expression of different CBXs family members with clinicopathological parameters of HCC patients
After mRNA expression and protein expression were found to be over-expressed in HCC patients, we next analyzed the relationship between mRNA expression of different CBXs family members with clinico-pathological parameters of HCC patients by UALCAN (http://ualcan.path.uab.edu), including patients’ individual cancer stages and tumor grades. As was shown in Figure 4, mRNA expressions of 8 CBXs family members were remarkably correlated with patients’ individual cancer stages, and patients who were in more advanced cancer stages tended to express higher mRNA expression of CBXs. The highest mRNA expressions of CBX4/8 were found in stage 4 (Figure 4D, H), while the highest mRNA expressions of CBX1/2/3/5/6/7 were found in stage 3 (Figure 4A-C, E-G). The reason why mRNA expressions of CBX1/2/3/5/6/7 in stage 3 seemed to be higher than that in stage 4 may be due to the small sample size (only 6 HCC patients were at stage 4). Similarly, as was shown in Figure 5, mRNA expressions of 8 CBXs family members were significantly related to tumor grades, and, as tumor grade increased, the mRNA expression of CBXs tended to be higher. The highest mRNA expressions of CBX1/3/4/5/6/8 were found in tumor grade 4 (Figure 5A, C-F, H), while the highest mRNA expression of CBX2 was found in grade 3 (Figure 5B). However, the highest mRNA expression of CBX7 was found in grade 1, and as tumor grade increased, the mRAN expression of CBX7 tended to be lower (Figure 5G). In short, the results above suggested that mRNA expressions of 8 CBXs family members were significantly associated with clinicopathological parameters in HCC patients.
Figure 4.
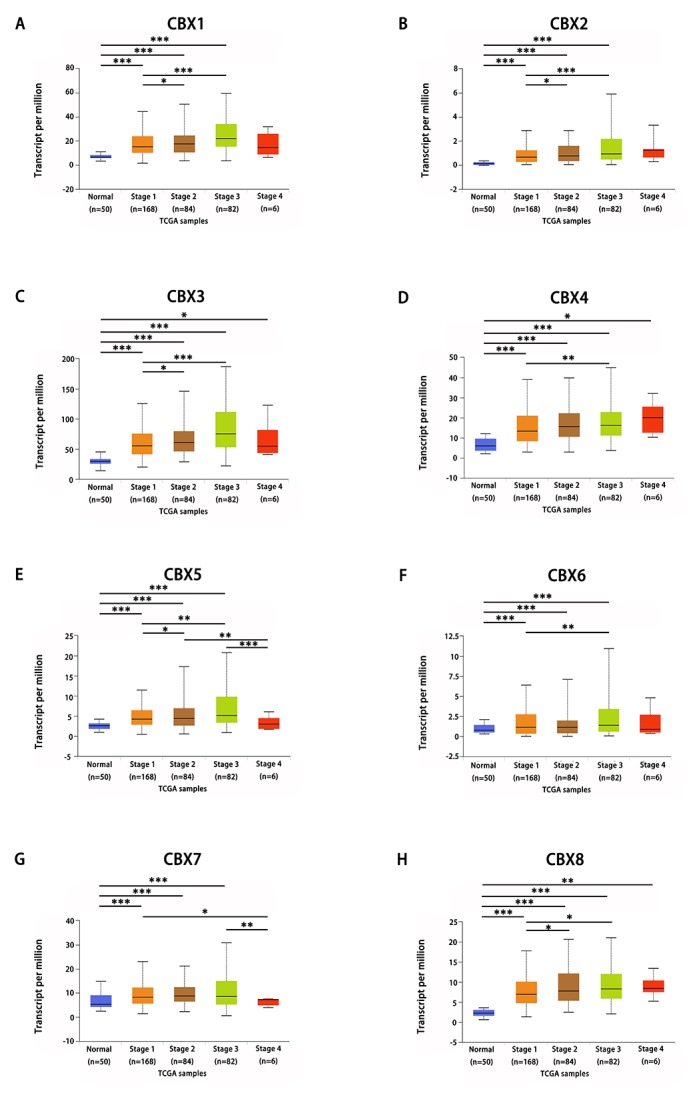
Relationship between mRNA expression of distinct CBXs family members and individual cancer stages of HCC patients. mRNA expressions of 8 CBXs family members were remarkably correlated with patients’ individual cancer stages, patients who were in more advanced stages tended to express higher mRNA expression of CBXs. The highest mRNA expressions of CBX4/8 were found in stage 4 (D, H), while the highest mRNA expressions of CBX1/2/3/5/6/7 were found in stage 3 (A-C, E-G). *p<0.05, **p<0.01, ***p<0.001.
Figure 5.
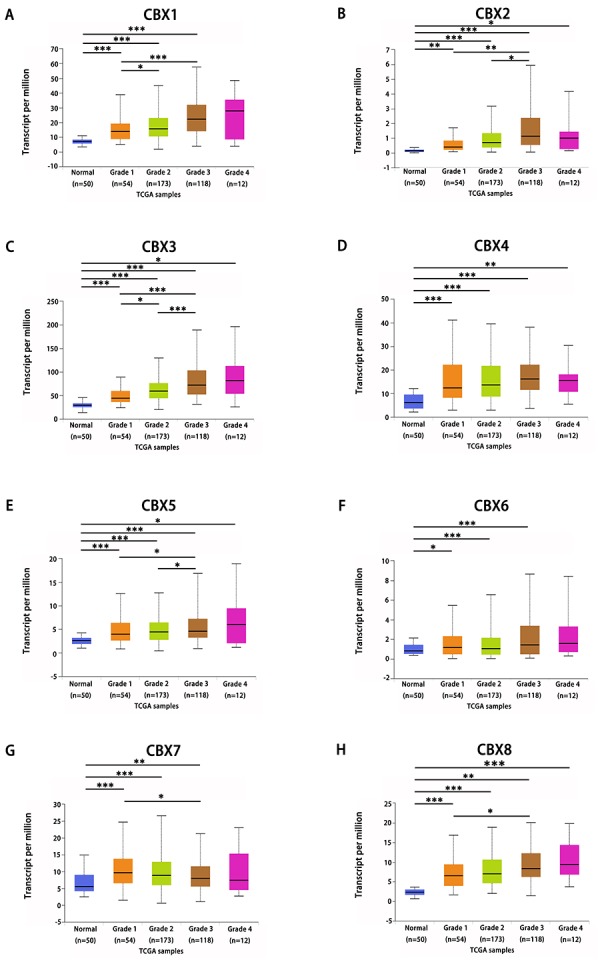
Association of mRNA expression of distinct CBXs family members with tumor grades of HCC patients. mRNA expressions of 8 CBXs family members were significantly related to tumor grades, and as tumor grade increased, the mRAN expressions of CBXs tended to be higher. The highest mRNA expressions of CBX1/3/4/5/6/8 were found in tumor grade 4 (A, C-F, H), while the highest mRNA expression of CBX2 was found in grade 3 (B). However, the highest mRNA expression of CBX7 was found in grade 1, and as tumor grade increased, the mRAN expression of CBX7 tended to be lower (G). *p<0.05, **p<0.01, ***p<0.001.
Prognostic value of mRNA expression of CBXs in liver cancer patients
Further, we used Kaplan-Meier plotter (http://kmplot.com/analysis/) to analyze the prognostic values of the mRNA expression of CBXs in liver cancer patients. As was shown in Figure 6, mRNA expressions of most of the CBXs family members were significantly associated with liver cancer patients’ prognosis. First, the relationship between combinatory mRNA expressions of all 8 CBXs family members and prognosis of liver cancer patients was analyzed (Figure 6A). Our results showed that higher combinatory mRNA expressions of all 8 CBXs family members was associated with poorer OS in liver cancers patients (HR=2.18, 95% CI: 1.54-3.08, and p=6.3E-06). Next, the association between mRNA expression of distinct CBXs family members and prognosis of liver cancer patients were further analyzed. As were shown in Figure 6B-6E, Figure 6G and Figure 6I, higher mRNA expression of CBX1 (HR=2.11, 95% CI: 1.45-3.07, and p=7.2e-05), CBX2 (HR=2.7, 95% CI: 1.89-3.85, and p=1.3E-05), CBX3 (HR=2.37, 95% CI: 1.67-3.36, and p=6.9E-07), CBX4 (HR=1.54, 95% CI: 1.06-2.24, and p=0.023), CBX6 (HR=1.54, 95% CI: 1.05-2.25, and p=0.026), CBX8 (HR=1.54, 95% CI: 1.08-2.19, and p=0.015) were significantly associated with shorter OS of liver cancers patients, while higher mRNA expression of CBX7 was significantly related to favorable OS of liver cancer patients (HR=0.51, 95% CI: 0.36-0.74, and p=0.00022) (Figure 6H). However, CBX5 mRNA expression showed no correlation with prognosis of liver cancer patients (HR=1.29, 95% CI: 0.9-1.86, and p=0.016) (Figure 6F). These results indicated that mRNA expressions of CBX1/2/3/4/6/7/8 were significantly associated with liver cancer patients’ prognosis and they may be exploited as useful biomarkers for prediction of liver cancer patients’ survival.
Figure 6.

Prognostic value of mRNA expression of distinct CBXs family members in liver cancer patients (Kaplan-Meier Plotter). Generally, higher combinatory mRNA expressions of all 8 CBXs family members were associated with poorer OS in liver cancers patients (A). Specifically, higher mRNA expressions of CBX1/2/3/4/6/8 were significantly associated with shorter OS of liver cancers patients (B-E, G, I), while higher mRNA expression of CBX7 was significantly related to favorable OS of liver cancer patients (H). However, CBX5 mRNA expression showed no correlation with prognosis in liver cancer patients (F).
Independent prognostic value of mRNA expression of CBXs in terms of OS in liver cancer patients
After mRNA expressions of CBX1/2/3/4/6/7/8 were found to be significantly associated with liver cancer patients’ prognosis, we then tried to assess the independent prognostic value of mRNA expression of CBXs in terms of OS in liver cancer patients. We downloaded clinical data (Supplementary Table 1) and mRNA expression of CBXs of 364 HCC patients of TCGA database from the Firebrowse website (http://firebrowse.org/api-docs/) for Cox survival regression analysis. In univariate analysis, we found that high pathologic stage (HR=1.586, 95% CI: 1.304-1.929, and p<0.001), high mRNA expressions of CBX1 (HR=1.560, 95% CI: 1.192-2.040, and p=0.001), CBX2 (HR= 1.337, 95% CI: 1.196-1.494, and p<0.001), CBX3 (HR= 1.787, 95% CI: 1.247-2.561, and p=0.002), CBX6 (HR= 1.150, 95% CI: 1.025-1.290, and p=0.017), CBX8 (HR= 1.325, 95% CI: 1.056-1.663, and p=0.015) and low mRNA expression of CBX7 (HR= 0.792, 95% CI: 0.645-0.971, and p=0.025) were related to shorter OS of liver cancer patients (Supplementary Table 2). Multivariate analysis showed that high mRNA expressions of CBX1 (HR= 1.534, 95% CI: 1.167-2.018, and p=0.002), CBX2 (HR= 1.349, 95% CI: 1.198-1.519, and p<0.001), CBX3 (HR= 1.691, 95% CI: 1.175-2.432, and p=0.005), CBX6 (HR= 1.124, 95% CI: 1.001-1.261, and p=0.048), CBX8 (HR= 1.300, 95% CI: 1.036-1.631, and p=0.023) were independently associated with significantly shorter OS of liver cancer patients (Supplementary Tables 3-10). These results showed that transcriptional expressions of CBX1/2/3/6/8 were independent prognostic factors for OS of liver cancer patients.
Genetic mutations in CBXs and their associations with OS and disease-free survival (DFS) of HCC patients
Next, we analyzed genetic alteration in CBXs and their associations with OS and DFS of HCC patients. As was shown in Figure 7A, high mutation rate of CBXs was observed in HCC patients. In the 360 sequenced HCC patients, genetic alteration was found in 183 HCC patients and the mutation rate was 51%. CBX8, CBX1, CBX2 and CBX4 ranked the highest four genes with genetic alterations, and their mutation rates were 18%, 16%, 16% and 15%, respectively. Furthermore, results from Kaplan-Meier plot and log-rank test showed that genetic alteration in CBXs was associated with shorter OS (Figure 7B, p=1.407E-4) and DFS (Figure 7C, p=1.790E-4) of HCC patients. These results implied that genetic alteration of CBXs could also significantly affect HCC patients’ prognosis.
Figure 7.
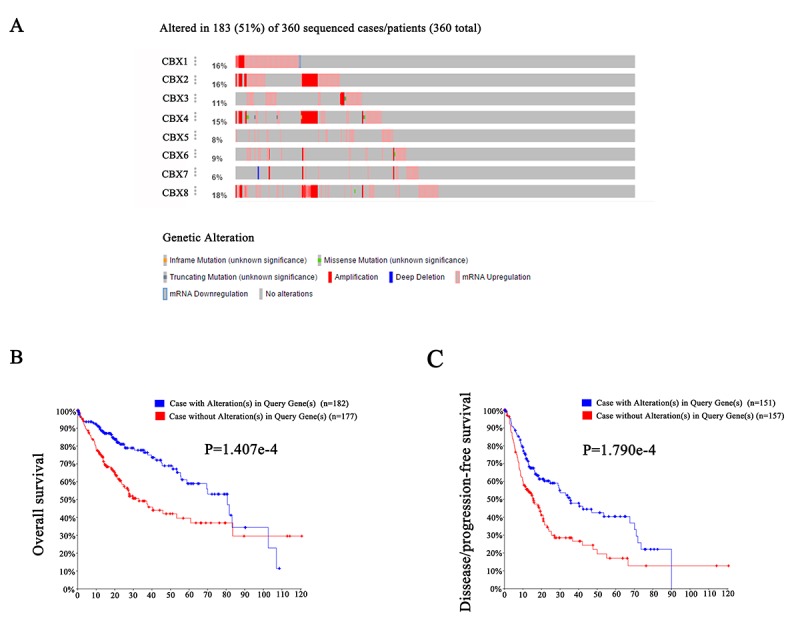
Genetic mutations in CBXs and their association with OS and DFS of HCC patients (cBioPortal). High mutation rate (51%) of CBXs was observed in HCC patients. CBX8, CBX1, CBX2 and CBX4 ranked the highest four genes of genetic alterations, and their mutation rates were 18%, 16%, 16% and 15%, respectively (A). Genetic alterations in CBXs were associated with shorter OS (B) and DFS (C) of HCC patients.
Predicted functions and pathways of the mutations in CBXs and their 50 frequently altered neighbor genes in HCC patients
After analyzing the genetic alterations in CBXs and their prognostic value in HCC patients, we further analyzed 50 neighbor genes which were significantly associated with CBXs mutations and constructed an integrated network by cBioPortal (www.cbioportal.org). As was shown at Figure 8A, the DNA repair and DNA replication related genes including HIST1H3A, HIST1H2AB, H3F3A, HIST2H2AA3, HIST2H3C and HIST3H2BB were significantly related to CBXs mutations. Moreover, functions of CBXs and their 50 frequently altered neighbor genes were analyzed by GO and KEGG in DAVID (https://david.ncifcrf.gov/summary.jsp). As were shown in Figure 8B-8D, biological processes such as GO: 0045815 (positive regulation of gene expression, epigenetic), GO: 0006334 (nucleosome assembly), GO: 0000183 (chromatin silencing at rDNA), GO: 0006362 (transcription elongation from RNA polymerase I promoter) and GO: 0006363 (termination of RNA polymerase I transcription) were remarkably regulated by the CBXs mutations in HCC (Figure 8B). Cellular components, including GO: 0000786 (nucleosome), GO: 0000788 (nuclear nucleosome), GO: 0005654 (nucleoplasm), GO: 0005634 (nucleus) and GO: 0000784 (nuclear chromosome, telomeric region) were significantly associated with the CBXs alterations (Figure 8C). In addition, CBXs mutations also prominently affected the molecular functions, such as GO: 0046982 (protein heterodimerization activity), GO: 0042393 (histone binding), GO: 0031492 (nucleosomal DNA binding), GO: 0003677 (DNA binding) and GO: 0003899 (DNA-directed RNA polymerase activity) (Figure 8D). In KEGG analysis, 6 pathways including has: 05322 (systemic lupus erythematosus), has: 05034 (alcoholism), has: 05203(viral carcinogenesis), has: 05202 (transcriptional misregulation in cancer) and has: 03020 (RNA polymerase) were associated with the functions of CBXs mutations in HCC (Figure 8E).
Figure 8.
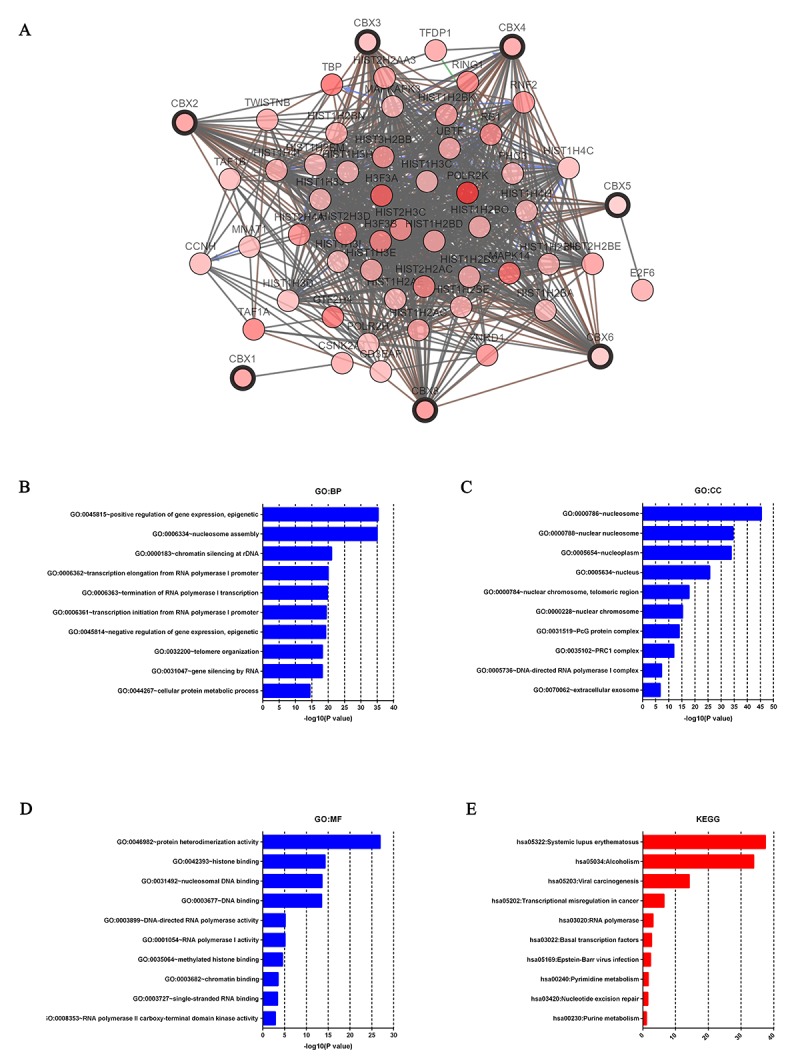
Predicted functions and pathways of the mutations in CBXs and their 50 frequently altered neighbor genes in HCC patients (c-BioPortal and DAVID). Network of CBXs mutations and their 50 frequently altered neighbor genes was constructed. DNA repair and DNA replication related genes including HIST1H3A, HIST1H2AB, H3F3A, HIST2H2AA3, HIST2H3C and HIST3H2BB were significantly related to CBXs mutations (A). GO functional enrichment analysis predicted three main functions of CBXs mutations and their 50 frequently altered neighbor genes, including biological process, cellular components and molecular functions (B-D). KEGG pathway analysis on CBXs and their 50 most frequently altered neighbor genes was shown at figure E.
DISCUSSION
In addition to cancer genetics, aberrant epigenetic regulation has also been found to take part in the development and progression and of HCC [4]. Being important components of epigenetic regulation complexes, CBXs family proteins are implicated in the development of multiple cancers, including HCC [9]. Despite some members of CBXs have been confirmed to play key roles in HCC, distinct roles of CBXs family members in HCC remained to be elucidated. In this study, the expression, mutation and prognostic values of different CBXs family members in HCC were analyzed.
Results from our study showed that over-expressions of mRNA and protein were found in all the 8 CBXs family members, and mRNA expression of CBXs was significantly associated with patients’ individual cancer stages and tumor grades in HCC patients. Besides, higher mRNA expressions of CBX1/2/3/6/8 were significantly associated with shorter OS in liver cancers patients, while higher mRNA expression of CBX7 was significantly related to favorable OS in liver cancer patients. Multivariate analysis showed that high mRNA expressions of CBX1/2/3/6/8 were independent prognostic factors for shorter OS of liver cancer patients. Moreover, high mutation rate (51%) of CBXs was observed in HCC patients and the genetic alteration in CBXs was associated with shorter OS and DFS in HCC patients. Finally, functions and pathways of the mutations in CBXs and their 50 frequently altered neighbor genes in HCC patients were analyzed and our results showed that the DNA repair and DNA replication related genes including HIST1H3A, HIST1H2AB, and HIST3H2BB were significantly related to CBXs mutations. Biological processes such as GO: 0045815 (positive regulation of gene expression, epigenetic), cellular components such as GO: 0000786 (nucleosome), molecular functions such as GO: 0046982 (protein heterodimerization activity), pathways such as, has: 05202 (transcriptional misregulation in cancer) were remarkably regulated by the CBXs mutations in HCC.
Over-expression of CBX1 had been found in castration-resistant prostate cancer (CRPC) and breast cancers (BC) [18,19]. In prostate cancer (PCa) tissues, higher CBX1 expression correlated with Gleason score and tri-methylation levels of histone H3K9. Inhibition of CBX1 suppressed the growth of androgen/androgen receptor-expressing PCa cells via inducing cell-cycle arrest at the G1 phase [18]. Similarly, over-expression of CBX1 was also found to be related with poorly differentiated breast tumors and poorer prognosis of BC patients [19]. Recently, the expression of CBX1 was found to be noticeably over-expressed in HCC tissues and cell lines. High CBX1 expression was associated with larger tumor size, poor tumor differentiation, tumor vascular invasion and unfavorable OS and DFS in HCC cases. In vitro study demonstrated that CBX1 over-expression promoted HCC cells proliferation and migration by interacting with transcription factor HMGA2 to activate the Wnt/β-Catenin signaling pathway, whereas knockdown of CBX1 or suppression of β-Catenin markedly decreased CBX1-mediated cell growth [20]. In our present study, significantly higher mRNA and protein expressions of CBX1 were found in HCC tissues compared to normal tissues, and that the mRNA expression of CBX1 was significantly associated with patients’ individual cancer stages and tumor grades which was in accordance with previous studies. Besides, higher mRNA expression of CBX1 was also significantly related with shorter OS of liver cancers patients and was an independent prognostic factor for shorter OS of liver cancer patients, indicating CBX1 took part in the tumorigenesis of HCC.
Recently, Clermont et al. conducted a genotranscriptomic meta-analysis of CBX2 in human cancers and found that CBX2 mRNA expression in many cancers was higher than that in normal tissues, which was independent of CDKN2A/B silence. Besides, over-expression and amplification of CBX2 were significantly related with metastatic progression and shorter OS in many cancer types, particularly in BC patients [21]. Mechanistically, Di Costanzo et al. found that CBX2 was over-expressed in leukemic cells and knockdown of CBX2 suppressed the tumorigenic properties and self-renewal capability of leukemic cells [22]. Clermont et al. also observed that CBX2 depletion abrogated cell viability and induced caspase3-mediated apoptosis in metastatic PCa cell lines [23]. In our study, significantly higher mRNA and protein expression of CBX2 were found in HCC tissues, and mRNA expression of CBX2 was remarkably correlated with patients’ individual cancer stages and tumor grades, which were similar to the findings of Clermont’ studies [21]. Moreover, higher mRNA expression of CBX2 was also significantly related with poorer OS of liver cancers patients and was an independent prognostic factor for shorter OS of liver cancer patients, indicating an oncogenic role of CBX2 in the HCC.
Significant up-regulation of CBX3 had been found in a variety of cancers, including lung adenocarcinoma (LUAD) and non-small cell lung cancer (nSCLC), tongue squamous cell carcinoma (TSCC), and colorectal cancer [24–27]. Studies from Liu et at showed that CBX3 was over-expressed in human colorectal cancer and it promoted cell proliferation of colorectal cancer cell lines by directly regulating CDKN1A in a manner associated with methylation of histone H3K9 on its promoter. Moreover, miR-30a could target CBX3 in vitro and in vivo to specifically inhibit the growth of colorectal cancer in mouse xenograft models [26]. Alam et al. also revealed that CBX3 was one of the most frequently over-expressed and amplified histone reader proteins in human LUAD and that high CBX3 mRNA level was associated with poor prognosis of LUAD patients. CBX3 promoted the proliferation, colony formation, and migration of LUAD cells by directly repressing NCOR2 and ZBTB7A. In vivo depletion of CBX3 suppressed K-RasG12D-driven LUAD and increased survival of mice bearing K-RasG12D-induced LUAD [25]. In the present study, significantly higher mRNA and protein expressions of CBX3 were found in HCC tissues, and mRNA expression of CBX3 was dramatically associated with patients’ individual cancer stages and tumor grades which was consistent with the studies above. Higher mRNA expression of CBX3 was also significantly related with unfavorable OS of liver cancers patients and was an independent prognostic factor for shorter OS of liver cancer patients, which indicated that CBX3 took part in the tumorigenesis of HCC.
CBX4 is the most well studied member of the CBXs family in HCC. Studies carried out by Jiao et al. and Wang et al. showed that CBX4 was over-expressed in clinical tissues and multiple HCC cell lines. Higher expression ofCBX4 was associated with clinical parameters including α-fetoprotein level, tumor size, pathologic differentiation, shorter OS and RFS [14,28]. Mechanistically, Li et al. found that CBX4 over-expression promoted tumor progression by increasing VEGF production and angiogenesis under hypoxia in subcutaneously and orthotopically transplanted HCC mice, while endogenous knockdown of CBX4 eliminated the oncogenic effect of CBX4 [29]. Moreover, Zhang et al. had revealed that miR-195 could significantly inhibit the proliferative, invasive and migratory capacities of HepG2 cells and HCC growth in vivo experiments by down-regulation of CBX4 [30]. Similar tumorigenic effect of CBX4 in HCC was also found in our present study. Our results showed that higher mRNA and protein expressions of CBX4 were found in HCC tissues, and that mRNA expression of CBX4 was significantly related with patients’ individual cancer stages and tumor grades.
Similar to CBX3, over-expression of CBX5 was found in many kinds of malignancies, such as pancreatic cancers, breast cancers and lung cancers [31]. In BC patients, higher CBX5 expression was correlated with decreased survival and increased occurrence of metastasis over time. Using CBX5 expression to predict disease outcome was a better prognostic biomarker than standard prognostic ones. Besides, down-regulation of CBX5 resulted in mitotic defects of breast cancer cell lines Hs578T [32]. In lung cancer patients, mRNA expression of CBX5 was significantly higher in the tumor samples as well as in the metastatic lesions and was associated with worse OS. Moreover, knockdown of CBX5 significantly inhibited capabilities of sphere and colony formation, migration of CD133+-tumor stem-like cells (TSLCs) in vitro and the tumorigenic engraftment, tumor growth rate, and metastatic tendency to lung caused by lung CD133+-TSLCs in vivo [33]. In our study, mRNA and protein expressions of CBX5 were found to be significantly higher in HCC tissues, and mRNA expression of CBX5 was significantly related with patients’ individual cancer stages and tumor grades. Despite patients with shorter OS tended to express higher mRNA expression of CBX5, the difference was not statistically significant. Further verifications are needed to discuss whether CBX5 plays an oncogenic role in HCC as other CBXs family members.
Frequent up-regulation of CBX6 had been found in HCC tissues and HCC cell lines and that CBX6 expression was significantly related with tumor sizes and multiple tumors. HCC patients with higher CBX6 expression had significantly shorter RFS and OS than those with lower CBX6 expression, and increased CBX6 expression was an independent unfavorable prognostic factor for HCC patients. Moreover, mechanistic study had shown that over-expression of CBX6 profoundly promoted HCC cell growth both in vitro and in vivo by regulating S100A9/NF-κB/MAPK pathway [34]. Similarly, in our study, higher mRNA expression of CBX6 was found in HCC tissues compared to normal tissues, and was significantly related with patients’ individual cancer stages, tumor grades. CBX6 was also significantly related with shorter OS of liver cancers patients and was an independent prognostic factor for shorter OS of liver cancer patients. All these results showed that CBX6 contributed to the development and progression of HCC and it may serve as a novel prognostic biomarker in HCC treatment.
Conflicting roles of CBX7 had been found in different kinds of human cancers [35]. On one hand, decreased expression of CBX7 had been found in most of the human malignant carcinomas, including bladder cancers, thyroid cancers, colorectal cancers, breast cancers and lung carcinomas, and in these cancers, down-regulation of CBX7 had been shown to correlate with cancer aggressiveness and poor prognosis, suggesting an oncosuppressor role of CBX7 in these cancers [36–40]. Mechanistic studies had found that CBX7 was able to counteract the oncogenic function of the HMGA proteins and to inhibit the expression of proliferation related and migration related genes, such as CCNE and SPP1 [35]. On the other hand, over-expression of CBX7 had also been found in some malignancies, such as prostate cancers and ovarian cancers [41,42]. Patients with over-expression of CBX7 exhibited reduced overall and progression-free survival rates compared to those expressing lower CBX7. Accordingly, inhibition of CBX7 decreased cell viability of ovarian carcinoma cell lines by promoting expression of TRAIL [42]. In regard to HCC, down-regulation of CBX7 had been found in HCC tissues and was associated with shorter OS of HCC patients [15]. Moreover, over-expression of miR-18a promoted cell proliferation and migration of HCC cell lines partly through decreasing CBX7 and depletion of CBX7 had the similar effects as miR-18a over-expression on HCC cell lines [43]. In our study, conflicting findings about the role of CBX7 in HCC were observed. On one hand, higher mRNA and protein expressions of CBX7 were found in HCC tissues, and mRNA expression of CBX7 was significantly related with patients’ individual cancer stages and tumor grades. However, on other hand, higher mRNA expression of CBX7 was correlated with better OS in liver cancers patients. Therefore, further studies are still required to assess the exact role of CBX7 in HCC.
Increased expression of CBX8 had been found in HCC tissues and was associated with poor prognosis of HCC patients [44]. Functional study had showed that over-expression of CBX8 promoted tumor growth and metastasis by increasing EGR1 and miR-365-3p to stimulate the AKT/β-catenin pathway, while CBX8 inhibition suppressed these effects [45]. Likewise, in the present study, significantly higher mRNA and protein expression of CBX8 were also found in HCC tissues, mRNA expression of CBX8 was remarkably correlated with patients’ individual cancer stages and tumor grades. Accordingly, higher mRNA expression of CBX8 was also significantly related with shorter OS of liver cancers patients and was an independent prognostic factor for shorter OS of liver cancer patients. Together with other findings discussed above, our results suggested that CBX8 played an oncogenic role in HCC.
There were some limitations in our study. First, although high mRNA expressions of CBX1/2/3/6/8 were independent prognostic factors for shorter OS of liver cancer patients, all the data analyzed in our study was retrieved from the online databases, further studies consist of larger sample sizes are required to validate our findings and to explore the clinical application of the CBXs members in the treatment of HCC. Second, we did not assess the potential diagnostic and therapeutic roles of CBXs in HCC, so future studies are needed to explore whether CBXs could be exploited as diagnostic markers or as therapeutic targets. Finally, we did not explore the potential mechanisms of distinct CBXs in HCC. Future studies worth to investigate the detailed mechanism between distinct CBXs and HCC.
In conclusion, our results showed that over expressions of 8 CBXs members were found to be significantly associated with clinical cancer stages and pathological tumor grades in HCC patients. Besides, higher mRNA expressions of CBX1/2/3/6/8 were found to be significantly associated with OS in HCC patients, while higher mRNA expression of CBX7 was associated with favorable OS. Multivariate analysis also showed that high mRNA expressions of CBX1/2/3/6/8 were independent prognostic factors for shorter OS of liver cancer patients. Moreover, high mutation rate of CBXs (51%) was also observed in HCC patients, and genetic alteration in CBXs was associated with shorter OS and DFS in HCC patients. These results indicated that CBX1/2/3/6/8 could be prognostic biomarkers for survivals of HCC patients.
MATERIALS AND METHODS
Ethics statement
Our study protocol was approved by the Ethics Committee of the Third Affiliated Hospital of Sun Yat-sen University. As all the data were retrieved from the online databases, so it could be confirmed that all written informed consent had already been obtained.
ONCOMINE database
ONCOMINE database (www.oncomine.org) is an integrated online cancer microarray database for DNA or RNA sequences analysis, which aims to facilitate discovery from the gene-wide expression analyses [46]. In our study, transcriptional expressions of 8 different CBXs members between different cancer tissues and their corresponding adjacent normal control samples were got from ONCOMINE database. Difference of transcriptional expression was compared by students’ t-test. Cut-off of p value and fold change were as following: p value: 0.01, fold change: 1.5, gene rank: 10%, data type: mRNA.
UALCAN
UALCAN (http://ualcan.path.uab.edu) is an interactive web resource based on level 3 RNA-seq and clinical data of 31 cancer types from TCGA database. It can be used to analyze relative transcriptional expression of potential genes of interest between tumor and normal samples and association of the transcriptional expression with relative clinicopathologic parameters [47]. In this study, UALCAN was used to analyze the mRNA expressions of 8 CBXs family members in primary HCC tissues and their association with clinicopathologic parameters. Difference of transcriptional expression was compared by students’ t test and p <0.01 was considered as statically significant.
Human Protein Atlas
The Human Protein Atlas (https://www.proteinatlas.org) is a website that contains immunohistochemistry-based expression data for near 20 highly common kinds of cancers and each tumor type includes 12 individual tumors [48]. Users can identify tumor-type specific proteins expression patterns that are differentially expressed in a given tumors of type. In this study, direct comparison of protein expression of different CBXs family members between human normal and HCC tissues was performed by immunohistochemistry image.
Kaplan-Meier plotter
The prognostic value of mRNA expression of distinct CBXs in liver cancers was analyzed by using Kaplan-Meier plotter (http://kmplot.com/analysis/),in which information about association of gene expression with survival of patients of liver cancer, breast cancer, ovarian cancer, lung cancer and gastric cancer could be easily access to [49–52]. In Kaplan-Meier plotter, cancer patients were divided into high and low expression group based on median values of mRNA expression and validated by K-M survival curves. Information about the number-at-risk cases, median values of mRNA expression levels, HRs, 95% CIs and p-values can be found at the K-M plotter webpage. Statically significant difference was considered when a p value < 0.05.
Cancer Genome Atlas (TCGA) database
TCGA is a comprehensive and coordinated project designed to improve diagnosis methods, treatment standards, and ultimately to prevent cancer. Information about sequencing and pathological data of more than 30 kinds of human tumors can be analyzed in TCGA [53]. In our analysis, clinicopathological parameters of 377 HCC patients and mRNA expression of CBXs of 371 HCC patients were downloaded from the Firebrowse website (http://firebrowse.org/api-docs/). 7 of 377 HCC patients were excluded because of the absence of follow-up data. Finally, 364 HCC patients subjected to mRNA expression of CBXs were included in our analysis. Clinical data, including gender, age, weight, PLT, albumin, creatinine, prothrombin time, total bilirubin, AFP, Child-Pugh stage, adjacent tissue inflammation, cirrhosis, histologic grade and pathologic stage were summarized in Supplementary Table 1.
cBioPortal
cBioPortal (www.cbioportal.org) is an online open-access website resource for exploring, visualizing, and analyzing multidimensional cancer genomics data [54]. In this study, we analyzed the genomic profiles of 8 CBXs family members, which contained mutations, putative copy-number alterations from GISTIC and mRNA Expression z-Scores (RNASeq V2 RSEM) with a z-score threshold ±1.8. Genetic mutations in CBXs and their association with OS and DFS of HCC patients were displayed as Kaplan-Meier plots and log-rank test was performed to identify the significance of the difference between the survival curves, and when a p value <0.05, the difference was considered statically significant.
Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis
Functions of CBXs mutations and 50 genes significantly associated with CBX mutations were analyzed by GO and KEGG in the Database for Annotation, Visualization, and Integrated Discovery (DAVID) (https://david.ncifcrf.gov/summary.jsp). GO enrichment analysis can predict the functional roles of CBXs mutations and 50 genes significantly associated with CBX mutations on the basis of three aspects, including biological processes(BP), cellular components (CC), and molecular functions (MF), while KEGG analysis can define the pathways related to the CBXs mutations and 50 frequently neighbor genes associated with CBXs mutations.
Statistical methods
Cox regression analysis was used to evaluate the association of mRNA expression of CBXs with patient survival by SPSS software version 20.0. First, missing covariates were imputed with methods similar to the methods of White [55]. Then, effect of clinical parameters and mRNA expression of CBXs on survival of HCC patients was evaluated by univariate Cox regression, followed by a filter which retained those with p ≤ 0.1 for subsequent analysis. Finally, association of mRNA expression of CBXs with patient survival was further analyzed with multivariate Cox regression, which adjusted for other parameters (e.g. Child-Pugh stage, histologic grade) similar to the methods of Hou [56]. P < 0.05 was considered statistically significant.
SUPPLEMENTARY MATERIAL
ACKNOWLEDGMENTS
We thank Hong-Ye Jiang for helpful writing.
Abbreviations
- HCC
hepatocellular carcinoma
- SNP
small nucleotide polymorphism
- PcG
polycomb group
- CBXs
chromobox
- TCGA
cancer genome atlas
- GO
gene ontology
- KEGG
Kyoto encyclopedia of genes and genomes
- BP
biological processes
- CC
cellular components
- MF
molecular functions
- OS
over survival
- DFS
disease free survival
- CPPC
castration-resistant prostate cancer
- BC
breast cancers
- PCa
prostate cancer
- LUAD
lung adenocarcinoma
- nSCLC
non-small cell lung cancer
- TSCC
tongue squamous cell carcinoma
- TSLCs
tumor stem-like cells
Footnotes
AUTHOR CONTRIBUTIONS: Liang Peng and Chan Xie designed the research; Gang Ning, Yan-Lin Huang and Li-Min Zhen carried out the research; Wen-Xiong Xu, Qian Jiao, Fang-Ji Yang, Li-Na Wu, Yong-Yuan Zheng and Jie Song analyzed the data; Gang Ning, Yan-Lin Huang and Li-Min Zhen wrote the paper, Yen-Sheng Wang helped to write the revised manuscript.
CONFLICTS OF INTEREST: All authors declared that there were no conflicts of interest with the contents of this article.
FUNDING: This study was supported by the National Natural Science Foundation of China (No. 81873572, 81570539 and 81472259), Plan of Science and Technology of Guangdong (No. 2016A020215221, 2015A020212007, and 2014A030313042), Guangzhou Science and Technology Project (No.201508020118, 201510010292 and 2014Y2-00544), and the Sun Yat-Sen University Clinical Research 5010 Program (2015004).
REFERENCES
- 1.Clark T, Maximin S, Meier J, Pokharel S, Bhargava P. Hepatocellular carcinoma: review of epidemiology, screening, imaging diagnosis, response assessment, and treatment. Curr Probl Diagn Radiol. 2015; 44:479–86. 10.1067/j.cpradiol.2015.04.004 [DOI] [PubMed] [Google Scholar]
- 2.Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015; 65:87–108. 10.3322/caac.21262 [DOI] [PubMed] [Google Scholar]
- 3.Vedham V, Verma M. Cancer-associated infectious agents and epigenetic regulation. Methods Mol Biol. 2015; 1238:333–54. 10.1007/978-1-4939-1804-1_18 [DOI] [PubMed] [Google Scholar]
- 4.Ma L, Chua MS, Andrisani O, So S. Epigenetics in hepatocellular carcinoma: an update and future therapy perspectives. World J Gastroenterol. 2014; 20:333–45. 10.3748/wjg.v20.i2.333 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Aloia L, Di Stefano B, Di Croce L. Polycomb complexes in stem cells and embryonic development. Development. 2013; 140:2525–34. 10.1242/dev.091553 [DOI] [PubMed] [Google Scholar]
- 6.Müller J, Verrijzer P. Biochemical mechanisms of gene regulation by polycomb group protein complexes. Curr Opin Genet Dev. 2009; 19:150–58. 10.1016/j.gde.2009.03.001 [DOI] [PubMed] [Google Scholar]
- 7.Wang W, Qin JJ, Voruganti S, Nag S, Zhou J, Zhang R, and Polycomb Group. (PcG) proteins and human cancers: multifaceted functions and therapeutic implications. Med Res Rev. 2015; 35:1220–67. 10.1002/med.21358 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Klauke K, Radulović V, Broekhuis M, Weersing E, Zwart E, Olthof S, Ritsema M, Bruggeman S, Wu X, Helin K, Bystrykh L, de Haan G. Polycomb Cbx family members mediate the balance between haematopoietic stem cell self-renewal and differentiation. Nat Cell Biol. 2013; 15:353–62. 10.1038/ncb2701 [DOI] [PubMed] [Google Scholar]
- 9.Ma RG, Zhang Y, Sun TT, Cheng B. Epigenetic regulation by polycomb group complexes: focus on roles of CBX proteins. J Zhejiang Univ Sci B. 2014; 15:412–28. 10.1631/jzus.B1400077 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wotton D, Merrill JC. Pc2 and SUMOylation. Biochem Soc Trans. 2007; 35:1401–04. 10.1042/BST0351401 [DOI] [PubMed] [Google Scholar]
- 11.Vincenz C, Kerppola TK. Different polycomb group CBX family proteins associate with distinct regions of chromatin using nonhomologous protein sequences. Proc Natl Acad Sci USA. 2008; 105:16572–77. 10.1073/pnas.0805317105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ruddock-D’Cruz NT, Prashadkumar S, Wilson KJ, Heffernan C, Cooney MA, French AJ, Jans DA, Verma PJ, Holland MK. Dynamic changes in localization of Chromobox (Cbx) family members during the maternal to embryonic transition. Mol Reprod Dev. 2008; 75:477–88. 10.1002/mrd.20752 [DOI] [PubMed] [Google Scholar]
- 13.Morey L, Pascual G, Cozzuto L, Roma G, Wutz A, Benitah SA, Di Croce L. Nonoverlapping functions of the Polycomb group Cbx family of proteins in embryonic stem cells. Cell Stem Cell. 2012; 10:47–62. 10.1016/j.stem.2011.12.006 [DOI] [PubMed] [Google Scholar]
- 14.Wang B, Tang J, Liao D, Wang G, Zhang M, Sang Y, Cao J, Wu Y, Zhang R, Li S, Ding W, Zhang G, Kang T. Chromobox homolog 4 is correlated with prognosis and tumor cell growth in hepatocellular carcinoma. Ann Surg Oncol. 2013. (Suppl 3); 20:S684–92. 10.1245/s10434-013-3171-7 [DOI] [PubMed] [Google Scholar]
- 15.Guan ZP, Gu LK, Xing BC, Ji JF, Gu J, Deng DJ. [Downregulation of chromobox protein homolog 7 expression in multiple human cancer tissues]. Zhonghua Yu Fang Yi Xue Za Zhi. 2011; 45:597–600. [PubMed] [Google Scholar]
- 16.Roessler S, Jia HL, Budhu A, Forgues M, Ye QH, Lee JS, Thorgeirsson SS, Sun Z, Tang ZY, Qin LX, Wang XW. A unique metastasis gene signature enables prediction of tumor relapse in early-stage hepatocellular carcinoma patients. Cancer Res. 2010; 70:10202–12. 10.1158/0008-5472.CAN-10-2607 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wurmbach E, Chen YB, Khitrov G, Zhang W, Roayaie S, Schwartz M, Fiel I, Thung S, Mazzaferro V, Bruix J, Bottinger E, Friedman S, Waxman S, Llovet JM. Genome-wide molecular profiles of HCV-induced dysplasia and hepatocellular carcinoma. Hepatology. 2007; 45:938–47. 10.1002/hep.21622 [DOI] [PubMed] [Google Scholar]
- 18.Shiota M, Song Y, Yokomizo A, Tada Y, Kuroiwa K, Eto M, Oda Y, Inokuchi J, Uchiumi T, Fujimoto N, Seki N, Naito S. Human heterochromatin protein 1 isoform HP1beta enhances androgen receptor activity and is implicated in prostate cancer growth. Endocr Relat Cancer. 2010; 17:455–67. 10.1677/ERC-09-0321 [DOI] [PubMed] [Google Scholar]
- 19.Lee YH, Liu X, Qiu F, O’Connor TR, Yen Y, Ann DK. HP1β is a biomarker for breast cancer prognosis and PARP inhibitor therapy. PLoS One. 2015; 10:e0121207. 10.1371/journal.pone.0121207 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Yang YF, Pan YH, Tian QH, Wu DC, Su SG. CBX1 indicates poor outcomes and exerts oncogenic activity in hepatocellular carcinoma. Transl Oncol. 2018; 11:1110–18. 10.1016/j.tranon.2018.07.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Clermont PL, Sun L, Crea F, Thu KL, Zhang A, Parolia A, Lam WL, Helgason CD. Genotranscriptomic meta-analysis of the Polycomb gene CBX2 in human cancers: initial evidence of an oncogenic role. Br J Cancer. 2014; 111:1663–72. 10.1038/bjc.2014.474 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Di Costanzo A, Del Gaudio N, Conte L, Dell’Aversana C, Vermeulen M, de Thé H, Migliaccio A, Nebbioso A, Altucci L. The HDAC inhibitor SAHA regulates CBX2 stability via a SUMO-triggered ubiquitin-mediated pathway in leukemia. Oncogene. 2018; 37:2559–72. 10.1038/s41388-018-0143-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Clermont PL, Crea F, Chiang YT, Lin D, Zhang A, Wang JZ, Parolia A, Wu R, Xue H, Wang Y, Ding J, Thu KL, Lam WL, et al. Identification of the epigenetic reader CBX2 as a potential drug target in advanced prostate cancer. Clin Epigenetics. 2016; 8:16. 10.1186/s13148-016-0182-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Chang SC, Lai YC, Chen YC, Wang NK, Wang WS, Lai JI. CBX3/heterochromatin protein 1 gamma is significantly upregulated in patients with non-small cell lung cancer. Asia Pac J Clin Oncol. 2017. [DOI] [PubMed] [Google Scholar]
- 25.Alam H, Li N, Dhar SS, Wu SJ, Lv J, Chen K, Flores ER, Baseler L, Lee MG. HP1gamma promotes lung adenocarcinoma by downregulating the transcription-repressive regulators NCOR2 and ZBTB7A. Cancer Res. 2018; 78:3834–48. 10.1158/0008-5472.CAN-17-3571 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Liu M, Huang F, Zhang D, Ju J, Wu XB, Wang Y, Wang Y, Wu Y, Nie M, Li Z, Ma C, Chen X, Zhou JY, et al. Heterochromatin protein HP1γ promotes colorectal cancer progression and is regulated by miR-30a. Cancer Res. 2015; 75:4593–604. 10.1158/0008-5472.CAN-14-3735 [DOI] [PubMed] [Google Scholar]
- 27.Zhang H, Fu X, Su X, Yang A. CBX3/HP1γ is upregulated in tongue squamous cell carcinoma and is associated with an unfavorable prognosis. Exp Ther Med. 2018; 15:4271–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jiao HK, Xu Y, Li J, Wang W, Mei Z, Long XD, Chen GQ. Prognostic significance of Cbx4 expression and its beneficial effect for transarterial chemoembolization in hepatocellular carcinoma. Cell Death Dis. 2015; 6:e1689. 10.1038/cddis.2015.57 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Li J, Xu Y, Long XD, Wang W, Jiao HK, Mei Z, Yin QQ, Ma LN, Zhou AW, Wang LS, Yao M, Xia Q, Chen GQ. Cbx4 governs HIF-1α to potentiate angiogenesis of hepatocellular carcinoma by its SUMO E3 ligase activity. Cancer Cell. 2014; 25:118–31. 10.1016/j.ccr.2013.12.008 [DOI] [PubMed] [Google Scholar]
- 30.Zheng C, Li J, Wang Q, Liu W, Zhou J, Liu R, Zeng Q, Peng X, Huang C, Cao P, Cao K. MicroRNA-195 functions as a tumor suppressor by inhibiting CBX4 in hepatocellular carcinoma. Oncol Rep. 2015; 33:1115–22. 10.3892/or.2015.3734 [DOI] [PubMed] [Google Scholar]
- 31.Vad-Nielsen J, Nielsen AL. Beyond the histone tale: HP1α deregulation in breast cancer epigenetics. Cancer Biol Ther. 2015; 16:189–200. 10.1080/15384047.2014.1001277 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.De Koning L, Savignoni A, Boumendil C, Rehman H, Asselain B, Sastre-Garau X, Almouzni G. Heterochromatin protein 1alpha: a hallmark of cell proliferation relevant to clinical oncology. EMBO Mol Med. 2009; 1:178–91. 10.1002/emmm.200900022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yu YH, Chiou GY, Huang PI, Lo WL, Wang CY, Lu KH, Yu CC, Alterovitz G, Huang WC, Lo JF, Hsu HS, Chiou SH. Network biology of tumor stem-like cells identified a regulatory role of CBX5 in lung cancer. Sci Rep. 2012; 2:584. 10.1038/srep00584 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zheng H, Jiang WH, Tian T, Tan HS, Chen Y, Qiao GL, Han J, Huang SY, Yang Y, Li S, Wang ZG, Gao R, Ren H, et al. CBX6 overexpression contributes to tumor progression and is predictive of a poor prognosis in hepatocellular carcinoma. Oncotarget. 2017; 8:18872–84. 10.18632/oncotarget.14770 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Pallante P, Forzati F, Federico A, Arra C, Fusco A. Polycomb protein family member CBX7 plays a critical role in cancer progression. Am J Cancer Res. 2015; 5:1594–601. [PMC free article] [PubMed] [Google Scholar]
- 36.Hinz S, Kempkensteffen C, Christoph F, Krause H, Schrader M, Schostak M, Miller K, Weikert S. Expression parameters of the polycomb group proteins BMI1, SUZ12, RING1 and CBX7 in urothelial carcinoma of the bladder and their prognostic relevance. Tumour Biol. 2008; 29:323–29. 10.1159/000170879 [DOI] [PubMed] [Google Scholar]
- 37.Pallante P, Federico A, Berlingieri MT, Bianco M, Ferraro A, Forzati F, Iaccarino A, Russo M, Pierantoni GM, Leone V, Sacchetti S, Troncone G, Santoro M, Fusco A. Loss of the CBX7 gene expression correlates with a highly malignant phenotype in thyroid cancer. Cancer Res. 2008; 68:6770–78. 10.1158/0008-5472.CAN-08-0695 [DOI] [PubMed] [Google Scholar]
- 38.Mansueto G, Forzati F, Ferraro A, Pallante P, Bianco M, Esposito F, Iaccarino A, Troncone G, Fusco A. Identification of a New pathway for tumor progression: microRNA-181b up-regulation and CBX7 down-regulation by HMGA1 protein. Genes Cancer. 2010; 1:210–24. 10.1177/1947601910366860 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pallante P, Terracciano L, Carafa V, Schneider S, Zlobec I, Lugli A, Bianco M, Ferraro A, Sacchetti S, Troncone G, Fusco A, Tornillo L. The loss of the CBX7 gene expression represents an adverse prognostic marker for survival of colon carcinoma patients. Eur J Cancer. 2010; 46:2304–13. 10.1016/j.ejca.2010.05.011 [DOI] [PubMed] [Google Scholar]
- 40.Forzati F, Federico A, Pallante P, Abbate A, Esposito F, Malapelle U, Sepe R, Palma G, Troncone G, Scarfò M, Arra C, Fedele M, Fusco A. CBX7 is a tumor suppressor in mice and humans. J Clin Invest. 2012; 122:612–23. 10.1172/JCI58620 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bernard D, Martinez-Leal JF, Rizzo S, Martinez D, Hudson D, Visakorpi T, Peters G, Carnero A, Beach D, Gil J. CBX7 controls the growth of normal and tumor-derived prostate cells by repressing the Ink4a/Arf locus. Oncogene. 2005; 24:5543–51. 10.1038/sj.onc.1208735 [DOI] [PubMed] [Google Scholar]
- 42.Shinjo K, Yamashita Y, Yamamoto E, Akatsuka S, Uno N, Kamiya A, Niimi K, Sakaguchi Y, Nagasaka T, Takahashi T, Shibata K, Kajiyama H, Kikkawa F, Toyokuni S. Expression of chromobox homolog 7 (CBX7) is associated with poor prognosis in ovarian clear cell adenocarcinoma via TRAIL-induced apoptotic pathway regulation. Int J Cancer. 2014; 135:308–18. 10.1002/ijc.28692 [DOI] [PubMed] [Google Scholar]
- 43.Yongyu Z, Lewei Y, Jian L, Yuqin S. MicroRNA-18a targets IRF2 and CBX7 to promote cell proliferation in hepatocellular carcinoma. Oncol Res. 2018; 26;:1327–34. 10.3727/096504018X15165493852990 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gao SB, Sun SL, Zheng QL, Zhang L, Zhu Y, Jin GH, Xue LX. Genetic alteration and misexpression of Polycomb group genes in hepatocellular carcinoma. Am J Cancer Res. 2015; 5:2969–79. [PMC free article] [PubMed] [Google Scholar]
- 45.Zhang CZ, Chen SL, Wang CH, He YF, Yang X, Xie D, Yun JP. CBX8 exhibits oncogenic activity via AKT/beta-catenin activation in hepatocellular carcinoma. Cancer Res. 2018; 78:51–63. 10.1158/0008-5472.CAN-17-0700 [DOI] [PubMed] [Google Scholar]
- 46.Rhodes DR, Yu J, Shanker K, Deshpande N, Varambally R, Ghosh D, Barrette T, Pandey A, Chinnaiyan AM. ONCOMINE: a cancer microarray database and integrated data-mining platform. Neoplasia. 2004; 6:1–6. 10.1016/S1476-5586(04)80047-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Chandrashekar DS, Bashel B, Balasubramanya SA, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BV, Varambally S. UALCAN: A portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia. 2017; 19:649–58. 10.1016/j.neo.2017.05.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Asplund A, Edqvist PH, Schwenk JM, Pontén F. Antibodies for profiling the human proteome-The Human Protein Atlas as a resource for cancer research. Proteomics. 2012; 12:2067–77. 10.1002/pmic.201100504 [DOI] [PubMed] [Google Scholar]
- 49.Szász AM, Lánczky A, Nagy Á, Förster S, Hark K, Green JE, Boussioutas A, Busuttil R, Szabó A, Győrffy B. Cross-validation of survival associated biomarkers in gastric cancer using transcriptomic data of 1,065 patients. Oncotarget. 2016; 7:49322–33. 10.18632/oncotarget.10337 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Györffy B, Lanczky A, Eklund AC, Denkert C, Budczies J, Li Q, Szallasi Z. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1,809 patients. Breast Cancer Res Treat. 2010; 123:725–31. 10.1007/s10549-009-0674-9 [DOI] [PubMed] [Google Scholar]
- 51.Gyorffy B, Lánczky A, Szállási Z. Implementing an online tool for genome-wide validation of survival-associated biomarkers in ovarian-cancer using microarray data from 1287 patients. Endocr Relat Cancer. 2012; 19:197–208. 10.1530/ERC-11-0329 [DOI] [PubMed] [Google Scholar]
- 52.Győrffy B, Surowiak P, Budczies J, Lánczky A. Online survival analysis software to assess the prognostic value of biomarkers using transcriptomic data in non-small-cell lung cancer. PLoS One. 2013; 8:e82241. 10.1371/journal.pone.0082241 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Tomczak K, Czerwińska P, Wiznerowicz M. The Cancer Genome Atlas (TCGA): an immeasurable source of knowledge. Contemp Oncol (Pozn). 2015; 19:A68–77. 10.5114/wo.2014.47136 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Gao J, Aksoy BA, Dogrusoz U, Dresdner G, Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, Cerami E, Sander C, Schultz N. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci Signal. 2013; 6:pl1. 10.1126/scisignal.2004088 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.White IR, Royston P. Imputing missing covariate values for the Cox model. Stat Med. 2009; 28:1982–98. 10.1002/sim.3618 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hou X, He X, Wang K, Hou N, Fu J, Jia G, Zuo X, Xiong H, Pang M. Genome-wide network-based analysis of colorectal cancer identifies novel prognostic factors and an integrative prognostic index. Cell Physiol Biochem. 2018; 49:1703–16. 10.1159/000493614 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.