

Abstract
Major improvements in sow prolificacy have resulted in larger litters but, at the same time, increased the proportion of piglets born light weight. Different management strategies aim to enhance the performance of, and limit light-weight piglet contribution to, BW variation within a batch; however, consequences on heavy-weight littermates are often neglected. This study investigated the effects of different litter compositions, created through cross-fostering, and the provision of creep feed on preweaning behavior and short- and long-term performance of piglets born either light weight (≤1.25 kg) or heavy weight (1.50–2.00 kg). Piglets were cross-fostered at birth to create litters with only similar-sized piglets (light weight or heavy weight; UNIFORM litters) and litters with equal numbers of light-weight and heavy-weight piglets (MIXED litters); half of the litters were offered creep feed and the remaining were not. Piglet behavior during a suckling bout and at the creep feeder was assessed; a green dye was used to discern between consumers and nonconsumers of creep feed. The interaction between litter composition and birth weight (BiW) class influenced piglet BW at weaning (P < 0.001): piglets born light weight were lighter at weaning in MIXED litters than those in UNIFORM litters (6.93 vs. 7.37 kg); however, piglets born heavy weight performed considerably better in MIXED litters (8.93 vs. 7.96 kg). Total litter gain to weaning was not affected (P = 0.565) by litter composition. Teat position affected heavy-weight piglet performance by d 10 (P < 0.001), with heavy-weight piglets in UNIFORM litters being disadvantaged when suckling the middle and posterior teats. Creep feed provision did not affect BW at weaning (P > 0.05) for either BiW class. However, litter composition significantly affected daily creep feed consumption (P = 0.046) and fecal color (P = 0.022), with heavy-weight piglets in UNIFORM litters consuming the highest amount of creep feed and having the greenest feces. In addition, a lower number of heavy-weight piglets in UNIFORM litters were classified as nonconsumers (P = 0.002). The weight advantage heavy-weight and light-weight piglets had at weaning when reared in MIXED and UNIFORM litters, respectively, was sustained throughout the productive period. In conclusion, reducing BW variation within litter (UNIFORM litters) was beneficial for piglets born light weight but not for piglets born heavy weight; the latter were disadvantaged up to slaughter. Although heavy-weight piglets in UNIFORM litters consumed the greatest amount of creep feed, this was not able to overcome their growth disadvantage compared with heavy-weight piglets in MIXED litters.
Keywords: birth weight, creep feed, cross-fostering, growth, swine, teat position
INTRODUCTION
The continuous improvement in sow prolificacy has increased litter size while leading to a considerable decrease in average birth weight and an increase in the number of piglets born light weight (Beaulieu et al., 2010; Rutherford et al., 2013). Light piglets, usually less than 1 kg at birth, are at a greater risk of dying before weaning (Hales et al., 2013; Ferrari et al., 2014), remain light throughout production (Beaulieu et al., 2010; Paredes et al., 2012), and need more time to reach slaughter weight (Quiniou et al., 2002; Paredes et al., 2012); therefore, they significantly contribute to batch inefficiency (Douglas et al., 2014). To reduce batch BW variation, it is essential to develop strategies to improve the performance of light-weight piglets.
Creation of uniform litters through cross-fostering reduces BW variation within litter consequently decreasing preweaning mortality (Milligan et al., 2001; Deen and Bilkei, 2004) and resulting in heavier weaning weight for lightweight piglets while weaned heavier (Deen and Bilkei, 2004; Douglas et al., 2014). On the other hand, the effect of uniform litters on the performance of piglets born heavy weight is unknown, because they may face competition from similar-sized pigs (Arnott and Elwood, 2009). Offering creep feed during lactation may reduce any potential negative effects on heavy-weight piglets and maintain litter uniformity. It is notoriously difficult to predict the consequences of creep feed provision, which can be low and variable within and between litters (Bøe and Jensen, 1995; Bruininx et al., 2004; Collins et al., 2013). The objective of this study was to investigate the effect of litter composition and creep feed provision on the lifetime performance of piglets born light weight or heavy weight. It was hypothesized that piglets born light weight would benefit from being in litters with less weight variability, that is, litters comprising only light-weight pigs. On the other hand, it was assumed that litter composition would not affect piglets born heavy weight; any potential adverse effects on them would be counterbalanced by creep feed provision.
MATERIALS AND METHODS
Experimental Design
The experiment was a 2 × 2 × 2 factorial design; treatments involved birth weight (BiW) class (light weight or heavy weight), litter composition (litters with only similar-sized piglets [light weight or heavy weight; UNIFORM litters] or litters with equal numbers of light-weight and heavy-weight piglets [MIXED litters]) and creep feed provision (yes or no). Piglets with a BiW of ≤1.25 kg (600 g minimum BiW) were considered light-weight, and piglets weighing between 1.5 and 2.0 kg were considered heavy-weight, in accordance with the methodology of Douglas et al. (2013, 2014). They were cross-fostered into litters of different compositions within 24 h from birth (see below). The experimental design was implemented on 37 sows and 442 piglets; 12 litters consisted of only light-weight piglets, 12 litters consisted of only heavy-weight piglets, and 13 litters consisted of both light-weight and heavy-weight piglets, with 6 light-weight and 6 heavy-weight piglets. Half of the UNIFORM litters and half of the MIXED litters were offered creep feed, whereas the other halves were not, resulting in at least 6 replicates per treatment. The litter was the experimental unit from birth to the grower stage (approximately 9 wk of age). The experiment was conducted at Cockle Park Farm (Newcastle University, Morpeth, Northumberland, UK) and was approved by the Animal Welfare and Ethical Review Body of Newcastle University.
Animals, Housing, and Management
Multiparous sows farrowed on a 3-wk cycle and were housed in conventional, partially slatted farrowing crates. All sows were Large White × Landrace, inseminated with Hylean boar semen (Hermitage Seaborough, Ltd., Devon, UK). Sows were placed in crates on Monday, and those that had not farrowed by Thursday were induced with a prostaglandin analog (Planate; Intervet UK, Walton, UK). The average number of piglets born alive was 12.1 (range 3 to 17), with an average BiW of 1.47 kg (SD 0.37), based on 121 sows that farrowed over the experimental period including experimental and nonexperimental sows; the average litter size is consistent with the average seen in United Kingdom farms but lower that what is seen in other European herds (AHDB Pork, 2016). All sows were fed a home-milled meal twice a day, and water was available ad libitum throughout lactation. The temperature in the farrowing unit was maintained at 21°C (20.8°C, range 17.3 to 25.4°C).
During the first 2 d postpartum, piglets were locked into the creep while the sow was eating, to minimize crushing. An infrared heat lamp was located in the covered creep area, and wood shavings were provided as bedding. Piglets had unlimited access to a water nipple drinker, which was cleaned daily. Within the first 12 h after birth, piglets had their teeth clipped. At approximately 3 d of age, piglets were tail docked and received an intramuscular iron injection. At 7 d of age, piglets were vaccinated against Mycoplasma hyopneumoniae (M+PAC; Intervet UK). The general health of piglets was examined on a daily basis, and any interventions were monitored.
Piglets were weaned at approximately 28 d of age and vaccinated for M. hyopneumoniae (M+PAC) and porcine circovirus type 2 (Inglevac Mycoflex; Boehringer Ingelheim GmbH, Ingelheim, Germany). Littermates remained together when moved to a fully slatted nursery accommodation. Each pen (183 by 170 cm) had nipple drinkers and a multiple-space feeder that allowed 3 piglets to feed simultaneously. All pigs had ad libitum access to a standard 3-stage pellet feeding regime (Primary Diets, AB Agri Ltd., Ripon, North Yorkshire, UK). Diet 1 was fed until 2 kg was consumed per pig and diet 2 was fed until 3 kg was consumed per pig, which was followed by the weaner feed, which was fed ad libitum up to 9 wk of age (grower stage; Table 1). The initial room temperature in the nursery accommodation was set at 26°C (26.1°C, range 25.0 to 26.8°C) and reduced by approximately 0.2°C each day to a minimum of 22°C (22.7°C, range 19.9 to 23.5°C).
Table 1.
Ingredient composition on an as-fed basis and chemical analysis of the creep feed and the postweaner feeds used1
| Creep feed2 | Postweaning feeds | |||
|---|---|---|---|---|
| Item | Diet 1 | Diet 2 | Weaner | |
| Ingredient, g/kg | ||||
| Barley | 128.7 | 75.0 | 75.0 | 150.0 |
| Wheat | – | 234.1 | 438.1 | 487.5 |
| Micronized wheat | 100.0 | 50.0 | 25.0 | – |
| Micronized maize | – | 25.0 | – | – |
| Porridge oat | – | 75.0 | 25.0 | – |
| Oats | 194.6 | – | – | – |
| Wheat feed | – | – | 12.5 | 25.0 |
| Herring meal | 100.0 | 75.0 | 60.0 | 25.0 |
| Soya bean meal | – | 145.2 | 223.3 | 250.0 |
| Full fat soya bean | – | 25.0 | 25.0 | – |
| Pig weaner vitamin/trace element supplement3 | 5.0 | 5.0 | 5.0 | 5.0 |
| Dried skim milk powder | 140.7 | 61.1 | – | – |
| Whey | 266.4 | 173.2 | 69.4 | – |
| Potato protein | – | 12.5 | – | – |
| L-Lysine HCL | 3.34 | 1.68 | 2.45 | 3.74 |
| DL-Methionine | 2.27 | 1.45 | 1.31 | 1.56 |
| L-Threonine | 2.51 | 1.15 | 1.19 | 1.57 |
| L-Tryptophan | 0.85 | 0.22 | 0.01 | 0.18 |
| L-Valine | 0.72 | |||
| Vitamin E | 0.15 | 0.41 | 0.21 | 0.10 |
| Benzoic acid | – | 5.00 | 5.00 | 5.00 |
| Limestone flour | – | 0.80 | 0.00 | 1.10 |
| Dicalcium phosphate | – | 0.00 | 5.10 | 8.90 |
| Salt | – | 0.00 | 1.15 | 4.10 |
| Binder (LignoBond DD)4 | – | 0.00 | 0.00 | 6.25 |
| Soya oil | 54.6 | 33.2 | 25.2 | 25.0 |
| Analyzed composition, %, as-fed basis | ||||
| CP | 20.0 | 23.1 | 22.1 | 20.8 |
| Crude fiber | 1.8 | 2.0 | 2.4 | 3.2 |
| Moisture | 8.8 | 8.9 | 10.2 | 10.9 |
| Ash | 6.7 | 5.8 | 5.1 | 5.1 |
| Calculated composition, %, as-fed basis, or as specified5 | ||||
| DE, MJ/kg | 16.50 | 16.00 | 15.30 | 14.80 |
| NE, MJ/kg | 11.55 | 10.99 | 10.66 | 10.37 |
| Calcium | 0.75 | 0.59 | 0.54 | 0.59 |
| Phosphorus | 0.71 | 0.59 | 0.60 | 0.59 |
| Lactose | 25.00 | 15.00 | 5.00 | 0.00 |
| Lys | 1.60 | 1.60 | 1.50 | 1.40 |
| SID6 Lys | 1.48 | 1.44 | 1.33 | 1.26 |
| Met | 0.67 | 0.60 | 0.54 | 0.50 |
| SID Met | 0.64 | 0.56 | 0.48 | 0.45 |
| SID Thr | 1.00 | 0.95 | 0.86 | 0.81 |
| SID Trp | 0.29 | 0.28 | 0.25 | 0.24 |
Diets were supplied by Primary Diets, AB Agri Ltd., Ripon, North Yorkshire, UK.
Additional ingredient: 0.10 g/kg chromic oxide.
It provided, per kilogram of complete diet, 11,500 IU vitamin A, 2,000 IU vitamin D3, 100 IU vitamin E, 4 mg vitamin K, 27.5 µg vitamin B12, 15 mg pantothenic acid, 25 mg nicotinic acid, 150 µg biotin, 1.0 mg folic acid, 160 mg Cu (CuSO4), 1.0 mg iodine (Ca (IO3)2), 150 mg Fe (FeSO4), 40 mg Mn (MnO), 0.25 mg Se (bone morphogenetic protein), and 110 mg Zn (ZnSO4).
Borregaard LignoTech, Sarpsborg, Norway.
Values estimated from the values in the Premier Atlas ingredients matrix (Hazzledine, 2008).
SID = standardized ileal digestible.
When moved to the on-site grower accommodation, pigs were fed a home-milled meal (20.4% CP, 9.83 MJ NE/kg diet, and 1.17% total lysine). Upon moving to the grower building, pigs were randomly mixed to create groups of 15 to 20 similarly sized pigs/pen. The pigs were kept in the same group up to slaughter. At approximately 12 to 13 wk of age (approximately 88 d), pigs were again moved to a fully slatted finisher building and were fed a commercial “finisher” pelleted diet (16.1% CP, 9.69 MJ NE/kg diet, and 1.00% total lysine). Pigs had ad libitum access to feed and water during the grower and finisher stages. Pigs reached slaughter weight of 90 to 100 kg at approximately 165 d of age and were sent to slaughter in 2 groups/batch, irrespective of treatment.
Experimental Procedures
Piglets were weighed to the nearest 1 g within 12 h postpartum. Neonates that did not meet the BiW criteria or those that had physical abnormalities were cross-fostered onto nonexperimental sows. Cross-fostering (d 0) was applied to create litters with 12 piglets per sow, including litters with only light-weight or heavy-weight piglets; MIXED litters consisted of equal numbers of light-weight (6) and heavy-weight (6) piglets. Piglets were randomly allocated to one of the treatment groups balanced for sex and litter of origin. Only healthy multiparous sows (>2 parities) with a sufficient number of functional teats were used to create experimental litters. Depending on the number of piglets available per batch, each litter composition was performed in duplicate. In order not to deprive piglets of access to colostrum from their biological mother, piglets were selected and individually identified by ear tagging and cross-fostered according to their BiW class within 24 h after birth.
During the first 4 d postpartum, all litters were given access to a commercial supplementary milk (Farmate; Volac, Royston, UK; 22% protein, 0% fiber, 14% oil and fats, 7.5% ash, and 2% lysine) in a small metal bowl. The milk was refreshed daily by mixing 150 g milk powder with 1 L warm water. Piglets were trained by dipping their snout in the milk bowl twice a day during the first 2 d postpartum. During early lactation (before d 10), individual piglets were weighed daily; piglets that lost weight during 2 consecutive days were removed from the experimental litter and cross-fostered onto a nonexperimental sow.
All piglets were weighed at 10 d of age. From then on and up to weaning, half of the litters were randomly assigned to having access to creep feed and the other half not. The creep feed (Primary Diets, AB Agri Ltd.), provided as pellets, was supplemented with 1.0% chromic oxide as a marker (approved by the United Kingdom Food Standards Agency, York, UK). A feed hopper with 2 feeding spaces was fixed to the wooden board of the pen close to the creep area. To ensure that any spillage was accounted for, a wooden tray that partly covered the slats was attached to the hopper. The amount of creep feed offered and refused was measured on a daily basis and was checked throughout the day to ensure ad libitum creep feed consumption.
Behavioral Observations
Teat Pair and Teat Consistency
Piglet position at the udder during at least 2 successful suckling bouts was assessed on d 2, 5, and 10 of lactation. Position at the udder was classified according to teat pair locations 1 through 7, from anterior to posterior. The start of a successful suckling bout was defined when more than half of the piglets gathered at the sow udder and began massaging. A suckling bout was considered complete when more than half of the litter had ceased massaging, by physically leaving the udder or falling asleep at the udder or when the sow changed position. The position of each individual piglet and whether piglets used more than 1 teat during a milk let down was recorded. If a piglet visited more than 1 teat pair with the same intensity per observation day, the teat numbers were averaged. Piglet teat pair on d 2, 5, and 10 was used to determine teat fidelity. A piglet was given a consistency score (Ci) of 1 when it used the same teat pair during the suckling bouts assessed throughout the day (d 2, 5, and 10). The number of piglets that scored 1 within a litter was expressed relative to the total number of piglets in the litter. A fixed teat position has been established by d 10 of age (Skok and Škorjanc, 2014); therefore, suckling position at d 10 was used to analyze its effect on subsequent performance. The preferred teat pair was grouped classifying the first 2 teat rows as anterior, teat pair 3 to 5 as middle, and teat pair ≥6 as posterior (Kim et al., 2000).
Feeding Behavior
Time spend at the creep feeder by individual piglets was monitored using video recordings on d 19, 21, and 25, because creep feed intake intensifies during the last week of lactation (Barnett et al., 1989; Bruininx et al., 2002); this was also confirmed by preliminary observations. Piglets were marked with a dark spray marker applying a unique mark on the back of each individual enabling identification different combinations of marks. At approximately 0900 h, cameras were turned on and left on for a period of 24 h. From 1600 h, artificial lights in the farrowing house were switched off. During a 7-h period from 0900 to 1600 h, continuous records were taken using CowLog (CowLog 2.0 desktop; Hänninen and Pastell, 2009). The total time (s) a piglet spent at the feeder was expressed relative to the time recorded. A successful feeding bout was defined from the point the piglet placed its snout in the feeder/tray for more than 5 s. A feeding bout was considered to end when the piglet removed its head for at least 15 s (adapted from Pajor et al., 1991). As piglets spilled creep feed on the tray, behaviors directed toward the tray were also assessed.
Individual Creep Feed Intake
In addition to the behavioral observation, the presence of the dye in the feces was used for the assessment of individual creep feed intake. Fecal samples were obtained at 3-d intervals during the first 1 1/2 wk of creep feed provision (d 13, 16, and 19) and 2-d intervals during the last week before weaning (d 21, 23, 25, and 27). For collecting purposes, piglets were placed on a weighing scale for a maximum of 4 min, stimulating voluntary defecation; fecal consistency was recorded, and samples with watery feces were excluded from subsequent analysis. Piglets of both creep-fed and non-creep-fed litters were sampled and, in total, fecal material was obtained from 87% of them. Data collected on d 27 was not used because some litters were weaned prior to this.
The presence of the inert dye in the fecal samples was used in 2 ways to classify creep feed consumers and nonconsumers through 1) the subjective observation of visually green feces and 2) a color reader (Color reader CR-10; Konica Minolta Sensing Inc., Sunderland, UK), objectively measuring fecal appearance.
Classification of Consumers
Total creep feed intake significantly increased from d 19 onward; therefore, piglets showing visibly green feces (dye present) at 19 d of age were considered early consumers. In addition, piglets were defined as consumers according to the number of fecal samples that appeared to be visually green. They were grouped in different consumer classes (i.e., low, moderate, and high), following the methodology of Bruininx et al. (2004) and Collins et al. (2013): piglets having visually green feces on 3 occasions (d 19, 21, and 25) were classified “high consumers.” Piglets that scored positive on 2 out of 3 sampling moments were classed “moderate consumers.” “Low consumers” were piglets having green feces on 1 occasion, and “nonconsumers” never showed green feces.
Colorimetric Assessment of Piglet Feces
The color space used was CIE L*a*b* (Konica Minolta, 2007), resulting in numerical color data: L*, a*, and b*. Measurements of interest were the chromaticity coordinate a*, which when negative indicates greener feces, and hue angle (H*), which defines how a color is perceived. Angles can be calculated from a* and b*, from which 0° represented red, 90° represented yellow, 180° represented green, and 270° represented blue; numbers in between represent intermediate hues. Negative values of b* indicate colors toward blue and positive values indicate colors toward yellow. The higher the value (either + or −), the more saturated a color is. Each reading began with a white tile to calibrate the instrument. The measurements taken were expressed in delta L*, delta a*, delta b*, and delta E, representing the color differences (i.e., + or −) between the white tile and the individual sample. Delta E is the value that indicates the size of the color difference considering L*, a*, and b* in a single measurement but does not indicate in what way the color is different. At least 5 measurements were taken from each fecal sample, because not all samples were uniform in color. The starting point of the target (i.e., L*, a*, and b* of the white tile) with which the samples were compared was recorded enabling true color estimation. The latter was done using the following formula:
A similar formula was used for calculating the true directions of b*. Hue angle was calculated using the following formula (Konica Minolta, 2007):
which is expressed in degrees.
The greenness represented by true a* and H* of creep feed was −7.38 (SD 0.150) and 121 (SD 0.812), respectively. Feces appeared to be greener as pigs matured. Both true a* and H* values of piglets with no access to creep feed were significantly affected by experimental day (P < 0.001), and a tendency was seen for an interaction between litter composition and experimental day (P = 0.067 and P = 0.046 respectively). To ensure that differences in true a* and H* were a result of the presence of chromic oxide, they were both corrected for day and treatment effects seen in noncreep litters, resulting in adjusted a* and adjusted H* (ab). Appendix I describes the methodology used.
Pre- and Postweaning Performance
Piglets were individually weighed at weaning (27.3 d of age [SD 0.9]), and those that had not reached a weaning weight of 4 kg were removed from the trial (Table 2). Additional weights were taken when pigs were moved to the grower facility (d 61.3 [SD 1.2]) and the finisher facility (d 88.0 [SD 2.9]) and the day before slaughter (d 164 [SD 13]), to which most pigs (75%) were followed. To account for pigs that were of different size, ADG was scaled to BW (scaled ADG; g/(d∙kg BW)). Up to 61 d of age, pigs remained in the same litter group, enabling the estimation of feed intake (FI)/litter throughout the nursery phase (d 28–61).
Table 2.
The total number of pigs allocated and the number of pigs removed from the trial, with the reasons for their removal, according to litter composition and birth weight class: light (less than 1.25 kg) or heavy (1.50 to 2.00 kg) were in litters with only similar-sized piglets (light weight or heavy weight; UNIFORM litters) or in litters with equal numbers of light-weight and heavy-weight piglets (MIXED litters). The number of pigs removed are expressed in absolute values and relative (%) to the the total number of pigs
| Litter composition | ||||||||
|---|---|---|---|---|---|---|---|---|
| UNIFORM | MIXED | |||||||
| Birth weight class | ||||||||
| Item | Light | Heavy | Light | Heavy | Total | Significance1 | ||
| Number of pigs on trial2 | ||||||||
| Day 0 | 144 | 144 | 77 | 77 | 154 | |||
| Day 28 | 117 | 129 | 59 | 74 | 133 | |||
| Day 61 | 116 | 129 | 59 | 74 | 133 | |||
| Day 88 | 115 | 129 | 59 | 74 | 133 | |||
| Day 165 | 98 | 109 | 54 | 64 | 118 | |||
| Number of pigs removed | ||||||||
| Found dead at <2 d of age | 6 (4.2%) | 0 (0.0%) | 3 (3.9%) | 0 (0.0%) | 3 (2.1%) | 0.027 | ||
| Lost weight; removed at <10 d of age | 14 (9.7%) | 10 (6.9%)a | 6 (7.8%) | 1 (1.3%)b | 7 (4.9%) | 0.136 | ||
| Found dead at >2 to <28 d of age | 5 (3.5%) | 3 (2.1%) | 5 (6.5%) | 2 (2.6%) | 7 (4.9%) | 0.367 | ||
| Under 4 kg at d 28 | 2 (1.4%)a | 2 (1.4%) | 4 (5.2%)b | 0 (0.0%) | 4 (2.8%) | 0.083 | ||
| Total | 27 (18.8%) | 15 (10.4%)a | 18 (23.4%) | 3 (3.9%)b | 21 (14.6%) | 0.001 | ||
Numbers within a row with different superscripts tended to statistically differ (P < 0.10).
Data were analyzed with a χ2 test.
Pigs were weighed within 12 h after birth (d 0), at weaning (d 27.3 [SD 0.9]), and when moved to the grower (d 61.3 [SD 1.2]) and finisher (d 88.0 [SD 2.9]) facilities. Not all pigs were followed to slaughter (d 164.2 [SD 13]), due to farm practices.
Statistical Analysis
The residual variances of the data were tested for normality using the UNIVARIATE procedure of SAS version 9.4 (SAS Inst. Inc., Cary, NC). Testing for normality showed skewed data for part of the data set, so data were normalized (either by square root, log, cube root, or inverse) and results were back-transformed for presentation using a 95% confidence level. The homogeneity of variance was tested using the Levene's test and graphical diagnostics using PROC GLM. Data were expressed as least squares means, with approximate SED unless otherwise stated. Statistical significance was assessed at the 5% level and tendencies were set at 10%.
A χ2 test was performed to test whether the reason for removing pigs from the experiment or the number of piglets per creep consumer class was affected by litter composition (UNIFORM or MIXED) and BiW class (light weight or heavy weight). Additionally, χ2 was used to determine the effect of creep feed provision (yes or no) and sex on the number of piglets removed.
The effect of BiW class (light weight or heavy weight) on the effect of litter composition on creep feed intake/piglet was estimated using 1-way ANOVA, using PROC GLM in SAS version 9.4, in which litter was the experimental unit. All other data were analyzed using the PROC MIXED procedure and were blocked by farrowing batch.
Main effects of interest were litter composition, BiW class, and their interaction for all models. Except for postweaning FI, creep feed provision did not significantly affect pre- and postweaning performance nor did it significantly interact with any other variable; therefore, it was omitted from subsequent analysis and is not presented in the Results. Sex significantly affected only individual creep feed intake (i.e., feeding behavior and the colorimetric method) and, therefore, was omitted from all other analyses. Initially, foster parity and preweaning litter size (adjusted litter size = [(Σ all the piglet hours piglets were suckling)/24 h]/weaning age in d) were added to model assessing preweaning performance but were not significant and were, therefore, excluded from the final model. Experimental day was an independent factor in the models assessing individual and daily creep feed intake. The factors teat pair class and “creep consumer” class were added to the models assessing their effect on pre- and postweaning performance. Age was added to the model at d 88 and slaughter due to the variability in timing of transfer between stages. Several covariance structures (i.e., first-order auto regression, compound symmetry, and variance components) were tested. For the RANDOM effects the variance components resulted in the lowest Akaike information criteria.
The experimental unit for daily creep feed intake in grams per day per piglet was litter average; average FI was calculated using the following formula: FI (g/d∙piglet) = [(total amount consumed in g)/total time (h) piglets spent with their foster sow] × 24 h. Piglet nested within litter was the experimental unit when assessing individual creep feed intake (i.e., feeding behavior, consumer class, or the colorimetric method) and teat pair class at d 10. Because measurements for feeding behavior and the colorimetric method were taken on subsequent days, repeated measures were used. The covariance structure first-order regression was used in the REPEATED statement. In addition, the PDIFF option in the LSMEANS statement was used to separate means for testing the effect of different variables (i.e., consumer class and teat pair class) on subsequent performance.
The experimental unit for the pre- and postweaning (d 0–61) performance was litter mean for light-weight or heavy-weight piglets. For UNIFORM litters, this was based on approximately 12 piglets born either light weight or heavy weight, and in the MIXED litters, this was the mean of approximately 6 piglets separately for each BiW class. Litter mean was blocked by sow nested within farrowing batch to account for light-weight and heavy-weight piglets in MIXED litters coming from the same litter. As the number of light-weight and heavy-weight piglets between the different litter compositions varied (MIXED versus UNIFORM), a WEIGHT statement was added to the model using the actual number of piglets that were classified light weight or heavy weight. From d 61, pigs were mixed according to their size; therefore, the experimental unit became the pen mean, based on the number of light-weight or heavy-weight piglets within each group, taking preweaning treatments (e.g., litter composition and creep feed provision [yes or no]) into consideration. Again, a WEIGHT statement was used to account for differences in the number of pigs and pen was nested within farrowing batch. The CV was calculated only from weaning up to d 61, because after that, pigs were mixed.
The Pearson correlation coefficient (r) was used to investigate whether creep feed intake was correlated with adjusted a*, adjusted H*ab, and feeding behavior and whether color reader measurements and preweaning performance were similarly correlated.
RESULTS
There was no difference in the average parity number of sows between the different treatments (P > 0.05). Although cross-fostering created litters of 12 piglets/sow, litter size decreased over time due to mortality and/or the removal of piglets. Nevertheless, litter size at weaning was not influenced by litter composition (P > 0.05). Average litter size at weaning was 10.5 (SD 1.7) for MIXED litters, 9.92 (SD 1.31) for light-weight piglets in UNIFORM litters, and 10.9 (SD 1.1) for heavy-weight piglets in UNIFORM litters. Piglet sex was evenly distributed across treatments (i.e., litter composition, BiW class, and creep feed provision), with 46.9% being females and 53.1% being males (P > 0.05).
Table 2 shows the total numbers used and the number of piglets removed from the trial, with the reasons for their removal, according to litter composition and BiW class. Overall preweaning mortality was 5.4% from the time piglets were cross-fostered, excluding removals. There was no effect of creep feed provision on piglet mortality or the removal of piglets before weaning (P > 0.05). Piglet mortality up to 2 d postpartum was significantly affected by BiW class (P = 0.027), with piglets born light weight having a higher mortality rate (3.9 or 4.2% for MIXED and UNIFORM litters, respectively) than piglets born heavy weight (0%). Litter composition and BiW class did not affect the number of piglets removed during early lactation (before d 10) as a result of losing weight or the number of piglets that died between >2 d postpartum and weaning. However, litter composition tended to affect (P = 0.066) the number of heavy-weight piglets that had to be removed as a result of weight loss (before d 10), with heavy-weight piglets in UNIFORM litters being removed in higher numbers (6.9%) than heavy-weight piglets in MIXED litters (1.3%). The number of piglets removed from the trial weighing less than 4 kg at weaning tended to be influenced by litter composition and BiW class (P = 0.083), with light-weight piglets being removed in higher numbers. In addition, light-weight piglets in MIXED litters tended (P = 0.097) to be removed at a higher rate (5.2%) than light-weight piglets in UNIFORM litters (1.4%). Lastly, the total number of piglets removed from birth to weaning was significantly (P = 0.001) affected by litter composition and BiW class. A higher number of light-weight piglets was removed compared with heavy-weight piglets. Nevertheless, litter composition affected only the total number of heavy-weight piglets, because heavy-weight piglets in UNIFORM litters tended (P = 0.091) to be removed at a higher rate (10.4%) than heavy-weight piglets in MIXED litters (3.9%).
Behavioral Observations
Teat Pair and Teat Consistency
The teat Ci was affected by experimental day (P = 0.006), as the percentage of piglets achieving a fixed teat pair (Ci = 1) increased over time, being 71.9% on d 2 (SD 21.5), 79.0% on d 5 (SD 22.7), and 87.2% on d 10 (SD 17.5). In addition, teat consistency was affected by litter composition (P = 0.030) on d 2, with piglets in UNIFORM litters having a significantly lower teat consistency (65.9% [SD 20.6]) than piglets in MIXED litters (82.9% [SD 19.4]).
Table 3 shows the effect of piglet-preferred teat pair class (i.e., anterior, middle, and posterior teat pair), litter composition, and BiW class on preweaning performance. The 3-way interaction among litter composition, BiW class, and teat pair class significantly affected piglet BW on d 10 (P = 0.001) and at weaning (P = 0.046). Heavy piglets in UNIFORM litters were >650 and >1,500 g lighter on d 10 and at weaning, respectively, when suckling a posterior (P < 0.001) or middle teat pair (P < 0.001) than those suckling an anterior teat pair. In contrast, heavy-weight piglets in MIXED litters were >400 g lighter at 10 d of age when suckling a posterior teat than their similar-sized littermates suckling an anterior (P = 0.012) or middle teat pair (P = 0.038). The latter difference was sustained throughout lactation, with heavy-weight piglets suckling an anterior teat pair being 1,000 g heavier at weaning than piglets suckling a posterior teat pair (P = 0.046). Teat pair preference did not influence (P > 0.05) light-weight piglet BW at 10 d of age in either litter composition. On the other hand, light-weight piglets in UNIFORM litters suckling an anterior (P = 0.005) or middle teat pair (P = 0.007) were >1,000 g heavier at weaning than piglets suckling a posterior teat pair. Similarly, light-weight piglets in MIXED litters suckling an anterior teat pair were >700 g heavier at weaning than piglets suckling a middle (P = 0.066) or posterior teat pair (P = 0.075). The 3-way interaction of litter composition × BiW class × teat pair class also significantly affected ADG (g/d) between birth and 10 d of age (P = 0.001) and tended to affect ADG between birth and weaning (P = 0.056).
Table 3.
The effect of piglet preferred teat pair class, litter composition, and birth weight class on preweaning performance: light (less than 1.25 kg) or heavy (1.50 to 2.00 kg) piglets were in litters with only similar-sized piglets (light weight or heavy weight; UNIFORM litters) or in litters with equal numbers of light-weight and heavy-weight piglets (MIXED litters)1
| Litter composition | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| UNIFORM | MIXED | Significance2,3 | |||||||||||||||||||
| Birth weight class | |||||||||||||||||||||
| Light | Heavy | Light | Heavy | ||||||||||||||||||
| Teat pair class4 | |||||||||||||||||||||
| Item | Anterior(n = 44) | Middle(n = 55) | Posterior(n = 20) | Anterior(n = 45) | Middle(n = 60) | Posterior(n = 26) | Anterior(n = 22) | Middle(n = 34) | Posterior(n = 7) | Anterior(n = 29) | Middle(n = 34) | Posterior(n = 11) | SED | Teat pair class | Litter composition × birth weight class × teat pair class | ||||||
| BW, kg | |||||||||||||||||||||
| Day 0 | 1.06 | 1.09 | 1.05 | 1.75 | 1.71 | 1.73 | 1.08 | 1.12 | 1.010 | 1.72 | 1.71 | 1.69 | 0.028 | 0.803 | 0.657 | ||||||
| Day 10 | 2.84 | 2.92 | 2.68 | 3.95a | 3.31b | 3.17b | 3.04 | 2.80 | 2.74 | 4.21a | 4.12a | 3.70b | 0.120 | <0.001 | 0.001 | ||||||
| Day 28 | 7.48a | 7.39a | 6.35b | 8.88a | 7.35b | 7.00b | 7.33c | 6.59d | 6.17d | 9.35a | 8.93 | 8.30b | 0.310 | <0.001 | 0.046 | ||||||
| ADG, g/d | |||||||||||||||||||||
| Day 0–10 | 179 | 183 | 164 | 218a | 159b | 143b | 197c | 170d | 164 | 250a | 241a | 202b | 11.2 | <0.001 | 0.001 | ||||||
| Day 0–28 | 234a | 229a | 194b | 259a | 204b | 194b | 233c | 204d | 189d | 284c | 267 | 250d | 11.1 | <0.001 | 0.056 | ||||||
Within litter composition and birth weight class main treatment comparison (teat pair class), means with different superscripts significantly differed (P < 0.05).
Within litter composition and birth weight class main treatment comparison (teat pair class), means with different superscripts tended to differ (P < 0.10).
Teat pair class was classified according to anatomical location of the teats (i.e., anterior, middle, and posterior) and was assessed at 10 d of age. Data are expressed as least squares means.
The experimental unit was piglet nested within litter; all data were blocked by farrowing batch. Data were analyses with PROC MIXED in SAS (SAS Inst. Inc., Cary, NC); the statistical model used was y = birth weight class + litter composition + teat pair class + birth weight class × litter composition + birth weight class × litter composition × teat pair class + farrowing batch (litter ) + ε. The covariance structure variance components was used in the random statement. The PDIFF option of SAS was used to separate the means.
In addition to the significant effect shown here, birth weight and the interaction between birth weight and litter composition significantly affected BW at d 10 and 28. Similarly, ADG between birth and d 10 and between birth and d 28 was affected by birth weight and birth weight × litter composition.
The position at the udder was classified according to teat pair location: anterior (1–2), middle (3–5), and posterior (≥6).
Teat pair class also significantly (P < 0.001) affected BW at d 10 and at weaning. Piglets suckling an anterior teat pair (3.51 [SD 0.66] and 8.26 kg [SD 1.67]) were considerably heavier at d 10 and at weaning, respectively, than piglets suckling a middle teat pair (3.29 [SD 0.69] and 7.57 kg [SD 1.73], respectively) or a posterior teat pair (3.06 [SD 0.58] and 6.97 kg [SD 1.58], respectively). In addition, teat pair significantly influenced ADG from birth to 10 d of age (P < 0.001) and from birth to weaning (P < 0.001). Piglets suckling an anterior teat pair had a higher ADG between birth and 10 d of age and between birth and weaning (211 [SD 62] and 252 g/d [SD 59], respectively) than those suckling a middle (188 [SD 65] and 226 g/d [SD 61], respectively) or posterior teat pair (169 [SD 57] and 207 g/d [SD 56], respectively).
Feeding Behavior
Feeding behavior assessed at d 19, 21, and 25 was not affected by the interaction between litter composition and BiW class (P > 0.05), litter composition (P > 0.05), or BiW class (P > 0.05). Experimental day (P < 0.001) and sex (P < 0.001) significantly contributed to differences in feeding behavior. Time spent at the feeder, expressed as total time spend at feeder/piglet relative to the time recorded, increased over time, being 0.133% (95% confidence interval 0.049–0.215) at d 19, 0.183% [0.090, 0.325] at d 21, and 0.307% [0.174, 0.495] at d 25. Females (0.262% [0.146, 0.429]) spent more time at the feeder than males (0.133% [0.063, 0.242]).
Creep Feed Intake
Litter Level
The interaction between litter composition and experimental day did not affect (P > 0.05) creep feed consumption in grams per day per piglet. Experimental day affected (P < 0.001) creep feed consumption (g/(d∙piglet)) as shown in Fig. 1. From d 18 onward, creep feed significantly (P < 0.05) increased over time. Most creep feed (85.1% [SD 13.7]) was eaten during the last week (d 20–27) before weaning. Litter composition also significantly (P = 0.046) influenced creep feed consumption; heavy-weight piglets in UNIFORM litters consumed more feed over the total period (6.51 g/(d∙piglet) [3.50, 11.54]) than light-weight piglets (P = 0.015) raised with similar-sized littermates (2.00 g/(d∙piglet) [0.78, 4.08]) or piglets of any BiW class in MIXED litters (P = 0.096; 3.14 g/(d∙piglet) [1.56, 5.69]).
Figure 1.
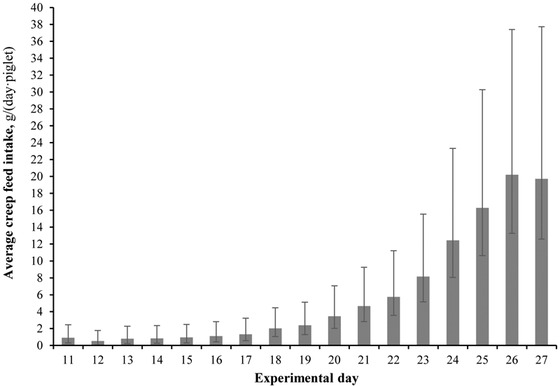
The effect of experimental day on creep feed intake (g/(d∙piglet)) across litters. Pigs had access to ad libitum creep feed from d 10 of lactation up to weaning (d 27.3 [SD 0.9]). Data are expressed as back-transformed (log) least squares means, with error bars representing the 95% confidence interval.
Creep feed consumption correlated with adjusted a* (r = −0.59, P < 0.001), adjusted H*ab (r = 0.64, P < 0.001), and feeding behavior (r = 0.69, P < 0.001). This indicates that litters consuming higher amounts of creep feed had lower adjusted a* and higher adjusted H*ab values, suggesting greener feces, and spent more time at the feeder.
Individual Piglet
Experimental day (P < 0.001) affected the number of visually color-positive fecal samples collected, ranging from approximately 3% on d 13 and 16 to approximately 30% on d 21 and 23. More than half of the piglets had visibly green feces at d 25 of age.
The number of piglets classified as early consumers on d 19 was significantly (P < 0.001) affected by the interaction between litter composition and BiW class. No differences (P > 0.05) were seen between the number of piglets born light weight that were classified as consumers and reared in either MIXED (18.5%) or UNIFORM litters (16.1%). However, more heavy-weight piglets in UNIFORM litters (P = 0.014) were classified as consumers (34.3%) than those reared in MIXED litters (13.5%). Piglets showing visibly green feces on d 19 tended (P = 0.053) to be lighter (5.07 kg [SD 0.16]) than piglets that did not show green feces (5.53 kg [SD 0.17]). However, the effect of being classified as a consumer on d 19 did not significantly affected BW at weaning (P > 0.05) nor did it affect (P > 0.05) ADG between d 19 and weaning (273 g/d [SD 81] for consumers versus 277 g/d [SD 99] for nonconsumers).
Table 4 summarizes the total number of piglets classified as either nonconsumers or consumers (low, moderate, or high) for piglets born light weight and heavy weight and reared either with similar-sized piglets or in MIXED litters. The number of piglets classified as nonconsumers was significantly affected (P = 0.002) by the interaction between litter composition and BiW class. A lower proportion (P = 0.006) of nonconsumers was seen for piglets born heavy weight in UNIFORM litters (27.3%) compared with heavy-weight piglets in MIXED (53.6%) litters. However, the fraction of light-weight piglets classified as nonconsumers was generally high, irrespective of litter composition. Furthermore, the number of piglets classified as high consumers tended to be different (P = 0.064), with piglets born heavy weight and reared in UNIFORM litters having the highest number of piglets classified as consumers (18.2%) and light-weight piglets in UNIFORM litters the lowest (4.7%).
Table 4.
Total number of piglets classified as either nonconsumers or consumers (low, moderate, or high) of creep feed for piglets born light (less than 1.25 kg) and heavy (1.50 to 2.00 kg) and either reared in litters with only similar-sized piglets (light weight or heavy weight; UNIFORM litters) or in litters with equal numbers of light-weight and heavy-weight piglets (MIXED litters)1
| Litter composition | ||||||
|---|---|---|---|---|---|---|
| UNIFORM | MIXED | |||||
| Birth weight class | ||||||
| Item | Light (n = 64) | Heavy (n = 66) | Light (n = 33) | Heavy (n = 41) | Significance2 | |
| High consumer | 3 (4.7%) | 12 (18.2%) | 2 (6.1%) | 4 (9.8%) | 0.064 | |
| Moderate consumer | 12 (18.7%) | 10 (15.1%) | 3 (9.1%) | 4 (9.8%) | 0.474 | |
| Low consumer | 16 (25.0%) | 26 (39.4%) | 7 (21.2%) | 11 (26.8%) | 0.174 | |
| Nonconsumer | 33 (51.6%) | 18 (27.3%)a | 21 (63.6%) | 22 (53.6%)b | 0.002 | |
Within main treatment comparison (litter composition or birth weight category), counts with different superscripts tended to differ (P = 0.006).
Piglets scoring positive (visually green feces) for all 3 sampling days (i.e., d 19, 21, and 25) were classified as “high consumers.” Piglets having green feces at 2 out of the 3 occasions were categorized as “moderate consumers,” and “low consumers” had green feces at 1 occasion. Nonconsumers were piglets that never scored positive on the sampling days. Data are expressed in absolute numbers and relative (%) to the total number of piglets in each colum.
Data were analyzed with a χ2 test.
The effect of consumer class on pre- and postweaning performance is summarized in Table 5. Consumer class significantly affected piglet BW on d 19 (P = 0.039). Animals classified as moderate and high consumers were >750 g lighter on d 19 than those classified as low consumers (P < 0.05) or nonconsumers (P < 0.05). In addition, consumer class tended (P = 0.089) to affect BW at weaning. Piglets classified as moderate consumers (7.01 kg [SD 1.85]) were >800 g lighter at weaning than piglets classified as nonconsumers (7.85 kg [SD 1.61]; P = 0.029) or low consumers (7.79 kg [SD 1.70]; P = 0.058). Average daily gain from birth to 19 d of age was significantly affected (P = 0.023) by consumer class. Piglets classified as moderate and high consumers gained significantly less than those classified as low consumers (P < 0.10) and nonconsumers (P < 0.05). However, consumer class did not affect (P > 0.05) ADG from d 19 to weaning; piglets classified as moderate (245 g/d [SD 90]) and high consumers (262 g/d [SD 99]) performed similar to those classified as nonconsumers (273 g/d [SD 95]) or low consumers (276 g/d [SD 93]). In addition, consumer class tended (P = 0.089) to affect ADG between birth and weaning, in which only moderate consumers seemed to be affected by gaining less than low consumers (P = 0.064) and nonconsumers (P = 0.027).
Table 5.
The effect of “consumer” class on pre- and postweaning performance. Piglets were classified as either nonconsumers or consumers (low, moderate, or high) of creep feed, and those scoring positive (visually green feces) for all 3 sampling days were classified as high consumers.1
| Consumer class | Significance2 | |||||
|---|---|---|---|---|---|---|
| Item | Nonconsumer | Low consumer | Moderate consumer | High consumer | SED | Consumer class |
| BW, kg | ||||||
| Day 0 | 1.38 | 1.41 | 1.40 | 1.41 | 0.013 | 0.896 |
| Day 10 | 3.31 | 3.34 | 3.13 | 3.15 | 0.053 | 0.573 |
| Day 19 | 5.58ac | 5.54ad | 4.98b | 4.82b | 0.089 | 0.039 |
| Day 28 | 7.85a | 7.79c | 7.01bd | 7.08 | 0.123 | 0.089 |
| Day 613 | 22.1 | 22.3 | 21.5 | 23.0 | 0.335 | 0.597 |
| ADG, g/d | ||||||
| Day 0–19 | 221a | 218ac | 188bd | 179b | 4.44 | 0.023 |
| Day 19–28 | 273 | 276c | 245d | 262 | 5.93 | 0.353 |
| –Day 0 28 | 238a | 235c | 208bd | 210 | 4.36 | 0.084 |
| Day 28–61 | 407a | 420a | 411a | 484b | 10.5 | 0.051 |
Within main treatment comparison (creep feed eater class), counts with different superscripts significantly differ (P < 0.05).
Within main treatment comparison (creep feed eater class), counts with different superscripts tended to differ (P < 0.10).
All piglets reared in creep feed litters had access to creep feed containing chromic oxide from 10 d of age up to weaning at approximately d 28 (d 27.3 [SD 0.9]). Visibly green feces indicated that the piglet had eaten creep feed. Fecal samples were taken and visually assessed during 3 d (d 19, 21, and 25). Piglets having green feces at 2 out of the 3 occasions were categorized as moderate consumers, and low consumers had green feces at 1 occasion. Nonconsumers were piglets that never scored positive on the sampling days. Data are expressed as least squares means.
In addition to the significant effect shown here, birth weight and the interaction between birth weight and litter composition significantly affected BW and ADG at the different stages of production. The experimental unit was piglet nested within litter; all data were blocked by farrowing batch. Data were analyses with PROC MIXED in SAS (SAS Inst. Inc., Cary, NC); the statistical model used was y = birth weight class + litter composition + consumer class + birth weight class × litter composition + farrowing batch (litter) + ε. The covariance structure variance components was used in the random statement. The PDIFF option of SAS was used to separate the means.
Piglets remained in the same litter from birth and weaning (d 27.3 [SD 0.9]) to d 61 (d 61.3 [SD 1.2]).
Although consumer class did not significantly affect (P > 0.05) BW once piglets reached grower age (d 61), piglets classified as high consumers were numerically heavier (Table 5). On the other hand, consumer class tended to affect ADG between weaning and 61 d of age (P = 0.051), with piglets classified as high consumers gaining significantly more than piglets classified as non- (P = 0.006), low (P = 0.029), or moderate consumers (P = 0.025).
Creep Estimate through the Colorimetric Method
The average fecal color (95% confidence interval) of piglets that had no access to creep feed was 0.00 [−0.12 to 0.12] for adjusted a* and 1.57 [1.49, 1.65] for adjusted H*ab. The interaction between litter composition and BiW class (P = 0.022) significantly affected adjusted a*. Piglets born heavy weight and reared in UNIFORM litters had significantly (P = 0.018) greener feces (−1.76 [SD 2.37]) than similar-sized piglets in MIXED litters (−0.60 [SD 2.15]). In contrast, adjusted a* for light-weight piglets reared together with heavier littermates was only numerically lower (−0.58 [SD 2.33]) than for piglets born light weight but reared in UNIFORM litters (−0.33 [SD 2.42]), implying that light-weight piglets in MIXED litters did not have greener feces than those in UNIFORM litters. Sex significantly interacted (P = 0.031) with BiW class, whereby light-weight females (−0.84 [SD 2.55]) had significantly (P < 0.001) greener feces than light-weight males (−0.07 [SD 2.14]). Females born heavy weight had only numerically greener feces (−1.27 [SD 2.43]) than their similar-sized males (−1.08 [SD 2.21]). The interaction between sex and experimental day also significantly affected adjusted a* (P < 0.001); females started to have significantly greener feces from d 19 onward (P < 0.05). Also, experimental day as a main effect significantly affected the greenness of feces (P < 0.001) of creep-fed piglets, in which adjusted a* became more negative over time, with 0.38 (SD 2.43) at d 13, −0.14 (SD 2.23) at d 16, −0.63 (SD 2.27) at d 19, −1.23 (SD 3.29) at d 21, −1.42 (SD 2.20) at d 23, and −1.85 (SD 2.38) at d 25. In addition, BiW class significantly contributed to differences in adjusted a* (P = 0.017), where heavy-weight piglets had greener feces (−1.18 [SD 2.32]) than piglets born light weight (−0.46 [SD 2.40]). Sex significantly (P < 0.001) affected adjusted a*, as females had greener feces (−1.06 [SD 2.51) than males (−0.57 [SD 2.22]). Adjusted a* was not affected by teat pair class (P > 0.05).
Adjusted H*ab was not affected by the interaction between litter composition and BiW class (P > 0.05), litter composition (P > 0.05), or BiW class (P > 0.05). Adjusted H*ab was significantly affected by experimental day (P < 0.001), increasing over time from 1.73 [1.37, 2.13] at d 13 to 2.15 [1.76, 2.59] at 25 d of age. In addition, sex significantly (P = 0.030) contributed to differences in adjusted H*ab; feces of females (1.94 [1.60, 2.30]) were greener than those of males (1.71 [1.40, 2.06]). Teat pair class significantly (P = 0.021) affected adjusted H*ab; piglets suckling the anterior teats had less green feces (1.62 [1.31, 1.96]) than those suckling the middle (1.88 [1.56, 2.24]) or posterior teat pair (1.98 [1.57, 2.44]).
Although significant (P < 0.05), correlations between the color readings (i.e., adjusted a* and adjusted H*ab) and preweaning performance were generally weak (r < 0.30). Scaled ADG (g/(d∙kg BW)) between birth and 19 d of age positively correlated (P < 0.05) with adjusted a* on subsequent sampling days (i.e., d 21, 23, and 25), ranging between r = +0.20 and r = +0.35. The opposite was true for adjusted H*ab, resulting in negative correlations (P < 0.05) ranging between r = −0.28 and r = −0.19. On the other hand, during the last week before weaning, from d 19 to 28, piglets that had green feces, represented by lower adjusted a* at d 21, 23, or 25, gained more (g/(d∙kg BW); r ranged between −0.21 and −0.24; P < 0.05). A similar effect (P < 0.05) was seen for adjusted H*ab, resulting in positive correlations at d 21, 23, and 25 between adjusted H*ab and scaled ADG from d 19 to weaning, ranging between r = +0.17 and r = +0.29.
Pre- and Postweaning Performance
Because creep feed provision did not influence performance at any stage of production or interact with litter composition or BiW class, creep feed treatment was removed from subsequent analyses. When all piglets weaned were included, the interaction between litter composition and BiW class (P < 0.001) significantly influenced weaning weight. Piglets born light weight and reared in UNIFORM litters were 600 g heavier at weaning than similar BiW piglets in MIXED litters (7.29 [SD 0.60] versus 6.67 kg [SD 0.85]). The opposite was true for heavy-weight piglets, which weighed more than 1 kg heavier when reared in MIXED litters compared with those in UNIFORM litters (8.93 [SD 0.79] versus 7.86 kg [SD 0.57]).
Table 6 shows the effect of litter composition, BiW class, and their interaction on the performance of piglets born light weight and heavy weight from birth to slaughter at different stages of production; these results include only piglets weaned heavier than 4 kg. The interaction between litter composition and BiW class (P < 0.001) significantly influenced piglet BW at approximately 28 d of age. Piglets born light weight were 400 g heavier at weaning when reared in UNIFORM litters than when reared in MIXED litters. When considering the effect of littermate weight on piglets born heavy weight, piglets from MIXED litters were almost 1 kg heavier at weaning than those reared in UNIFORM litters. Similarly, ADG (P < 0.001) and scaled ADG (P < 0.001) from birth to weaning was significantly affected by the interaction between litter composition and BiW class. Piglets born light weight and reared in UNIFORM litters gained more than those reared in MIXED litters; the opposite was true for piglets born heavy weight. Furthermore, BiW class affected BW at weaning (P < 0.001), with piglets born light weight being 1.3 kg lighter than piglets born heavy weight (7.11 [SD 0.64] vs. 8.40 kg [SD 0.59]). Total litter gain between birth and weaning was not affected by litter composition (P = 0.565); UNIFORM litters gained 64.6 kg (SD 12.9) and MIXED litters 66.7 kg (SD 11.0).
Table 6.
The effect of litter composition (litters with only similar-sized piglets [light weight or heavy weight; UNIFORM litters] vs. litters with equal numbers of light-weight and heavy-weight piglets [MIXED litters]) and birth weight class and their interaction on the performance of piglets born light (less than 1.25 kg) and heavy (1.50 to 2.00 kg) from birth to slaughter at different stages of production (weaner, grower, finisher, and slaughter)1
| Litter composition | ||||||||
|---|---|---|---|---|---|---|---|---|
| UNIFORM | MIXED | Significance2 | ||||||
| Birth weight class | Birthweight class | Littercomposition | Litter composition × birth weight class | |||||
| Item | Light | Heavy | Light | Heavy | SED | |||
| BW, kg | ||||||||
| Day 03 | 1.06 [1.04, 1.09] | 1.72 [1.69, 1.76] | 1.10 [1.06, 1.13] | 1.70 [1.66, 1.75] | <0.001 | 0.411 | 0.093 | |
| Day 28 | 7.37 | 7.96 | 6.93 | 8.93 | 0.099 | <0.001 | 0.137 | <0.001 |
| Day 61 | 20.9 | 23.7 | 20.5 | 25.2 | 0.337 | <0.001 | 0.282 | 0.072 |
| Day 88 | 36.2 | 39.5 | 35.1 | 42.3 | 0.595 | <0.001 | 0.326 | 0.020 |
| Day 165 | 97.1 | 98.7 | 93.4 | 101 | 1.05 | 0.001 | 0.684 | 0.018 |
| ADG, g/d | ||||||||
| Day 0–28 | 264 | 280 | 252 | 324 | 3.97 | <0.001 | 0.032 | <0.001 |
| Day 28–61 | 393 | 454 | 393 | 470 | 7.79 | <0.001 | 0.596 | 0.570 |
| Day 61–88 | 575 | 592 | 541 | 634 | 17.6 | 0.009 | 0.841 | 0.064 |
| Day 88–165 | 777 | 778 | 754 | 780 | 11.0 | 0.299 | 0.415 | 0.327 |
| Scaled ADG, g/(d∙kg BW) | ||||||||
| Day 0–28 | 245 | 162 | 229 | 188 | 5.20 | <0.001 | 0.402 | <0.001 |
| Day 28–61 | 55.8 | 57.7 | 52.9 | 51.4 | 2.61 | 0.940 | 0.140 | 0.561 |
| Day 61–884 | 25.8 [22.0, 31.3] | 23.8 [20.5, 28.4] | 24.6 [20.8, 30.0] | 23.6 [20.2, 28.3] | 0.115 | 0.459 | 0.600 | |
| Day 88–165 | 21.7 | 20.8 | 22.6 | 19.6 | 0.540 | <0.001 | 0.724 | 0.048 |
Light-weight and heavy-weight piglets were in UNIFORM litters or in MIXED litters. Data are expressed as least squares means or as otherwise stated. Piglets remained in the same litter from birth to d 61, after which they were randomly mixed according to their size. Pigs were weighed at birth d 0, when weaned (d 27.3 [SD 0.9]), when moved to the grower facility (d 61.3 [SD 1.2]), when moved to finisher accommodation (d 88.0 [SD 2.9]), and when reaching slaughter weight (d 164.2 [SD 13]).
The experimental unit for performance up to d 61 was litter mean for light-weight or heavy-weight piglets; all data were blocked by litter nested within farrowing batch. Data were analyses with PROC MIXED in SAS (SAS Inst. Inc., Cary, NC); the statistical model used was y = birth weight class + litter composition + birth weight class × litter composition + farrowing batch (litter) + ε. Due to variability in the timing of transfer, age was added to the model from d 88 onward. The experimental unit for performance from d 61 to slaughter was pen mean nested within farrowing batch; the statistical model used was y = birth weight class + litter composition + birth weight class × litter composition + age + farrowing batch (pen) + ε. For both models, the variance components was used as covariance structure in the random statement. In addition, a weight statement was used to account for differences in the number of light-weight and heavy-weight piglets where the litter or pen mean was based on.
3Data are expressed as back-transformed (log) least squares means with the 95% confidence interval.
Data are expressed as back-transformed (inverse) least squares means with the 95% confidence interval.
Body weight at d 61 and 88 and the day before slaughter was significantly (P < 0.05 or P < 0.10) affected by the interaction between litter composition and BiW class. The weight advantage heavy-weight piglets had at weaning when reared in MIXED litters increased to 1.5 kg when they reached grower weight (approximately d 61), 2.8 kg when they reached finisher weight (approximately d 88), and almost 2.5 kg by the day before slaughter. Also, for piglets born light weight and reared in different litter compositions, the effect of litter composition on BW was sustained throughout production, with a 400-g difference at d 61 and a 1.1-kg difference at d 88, and piglets were almost 3.7 kg heavier when they reached slaughter age when reared in UNIFORM litters. The interaction between litter composition and BiW class did not (P > 0.05) influence ADG or scaled ADG during the nursery (d 28–61) or grower phase (d 61–88). Total group gain between weaning and the grower phase, during which pigs remained in the same preweaning group, was not affected by litter composition (P = 0.570); pens of UNIFORM litters had a total group gain of 143 kg (SD 45) and pens of MIXED litters had a total group gain of 136 kg (SD 37). Teat pair class affected postweaning performance (P = 0.002). Piglets suckling an anterior teat pair class had a significantly lower scaled ADG (53.7 g/(d∙kg BW) [SD 18.2]) between weaning and the grower phase than those suckling a middle (61.4 g/(d∙kg BW) [SD 18.1]) or posterior teat pair (64.6 g/(d∙kg BW) [SD 18.2]).
Birth weight class significantly affected BW at the different weighing points (i.e., d 61, 88, and 165) and ADG from weaning to 9 wk of age (P < 0.001) and from d 61 to 88 (P = 0.009). Piglets born light weight were 4 kg lighter when they reached the grower stage (20.9 [SD 2.2] vs. 24.8 kg [SD 2.0]), 5 kg lighter when they reached the finisher stage (35.7 [SD 5.9] vs. 40.9 kg [SD 5.8]), and 5 kg lighter on the day before slaughter than piglets born heavy weight (95.3 [SD 10.8] vs. 100 kg [SD 10.0]). In addition, BiW class significantly affected (P = 0.031) slaughter age, whereby light-weight piglets were generally older at slaughter (166 [158, 177] vs. 162 d of age [155, 170]) than piglets born heavy weight.
Table 7 presents the effect of litter composition and BiW class and their interaction on the CV from the weaning to the grower phase. There was no main effect of litter composition on CV. Only at weaning was the CV significantly influenced by an interaction between BiW class and litter composition (P = 0.007). Piglets born light weight and reared together with heavier littermates (MIXED litters) had a numerically higher CV than those in UNIFORM litters; the opposite was true for heavy-weight piglets (P = 0.009). Birth weight class tended to affect CV at d 28 (P = 0.082) and 61 (P = 0.077). Piglets born light weight had a higher CV, at weaning and when they reached the grower stage (d 61). Furthermore, litter CV of heavy-weight piglets at weaning was affected (P = 0.051) by creep feed provision. Heavy piglets having access to creep feed had a lower CV (15.2 [SD 5.0]) than heavy-weight piglets without creep feed (19.4 [SD 5.2]).
Table 7.
The effect of litter composition (litters with only similar-sized piglets [light weight or heavy weight; UNIFORM litters] vs. litters with equal numbers of light-weight and heavy-weight piglets [MIXED litters]), birth weight class (light vs. heavy) and their interaction on the CV from weaning to the grower phase1
| Litter composition | ||||||||
|---|---|---|---|---|---|---|---|---|
| UNIFORM | MIXED | Significance | ||||||
| Birth weight class | Birthweight class | Littercomposition | Litter composition × birth weight class | |||||
| Item | Light | Heavy | Light | Heavy | SED | |||
| CV | ||||||||
| Day 28 | 18.2 | 19.9 | 21.3 | 14.2 | 0.847 | 0.082 | 0.409 | 0.007 |
| Day 61 | 13.1 | 11.8 | 12.9 | 9.7 | 0.927 | 0.077 | 0.359 | 0.442 |
UNIFORM litters consisted of only piglets born light (less than 1.25 kg) or heavy (1.50 to 2.00 kg), whereas MIXED litters contained mixed weights (both light-weight and heavy-weight piglets). Data are expressed as least squares means. Piglets remained in the same litter from birth and weaning (d 27.3 [SD 0.9]) to d 61 (d 61.3 [SD 1.2]). The experimental unit was litter CV for light-weight or heavy-weight piglets; all data was blocked by litter nested within farrowing batch. Data was analyses with PROC MIXED in SAS (SAS Inst. Inc., Cary, NC); the statistical model used was y = birth weight class + litter composition + birth weight class × litter composition + farrowing batch (litter) + ε. A weight statement was used using the actual number of light-weight and heavy-weight piglets where the litter CV was based on. The covariance structure used in the random statement was variance components.
During the nursery stage (d 28–61), piglets stayed in the same preweaning litter group, enabling the estimation of FI/litter. Litter composition (P = 0.002) significantly influenced daily FI when expressed per piglet. Heavy pigs in UNIFORM litters consumed the highest amount of feed followed by MIXED litters. The lowest amount of weaner feed was consumed by light-weight pigs in UNIFORM litters. Furthermore, numerical differences were found for creep feed provision on postweaning daily intake (P > 0.05). Pigs that had access to creep feed before weaning ate more of the weaner feed (647 g/(d∙piglet) [SD 55]) than pigs raised without it (616 g/(d∙piglet) [SD 53]).
DISCUSSION
The high-level objective of this work was to develop strategies to deal with the challenge of piglets born light weight. Piglets born light can either be born small for gestational age or have experienced intrauterine growth restriction (Rutherford et al., 2013). Although different definitions are considered in the literature, the consistent view is that light-weight piglets have a significantly lower preweaning survival rate (58 vs. 92%; Jourquin et al., 2016); need 7 to 14 d more to reach slaughter weight (105 kg) compared with piglets weighing 1.50 or 2.00 kg, respectively, at birth (Quiniou et al., 2002); and have a poorer feed efficiency (Gondret et al., 2006). Schinckel et al. (2007) predicted that for piglets with a BiW of 1 kg, a 0.1-kg increase in BiW reduced the days to reach 105 kg by 2.86, implying that the extra days needed to reach slaughter weight can be even more. Taking into consideration that around 15% of the newly born piglets weigh less than 1.11 kg at birth (Feldpausch et al., 2016) and that pigs are mostly sold on weight specifications rather than age, this results in batch inefficiency. This might be even more detrimental in very highly prolific sows, because the number of small piglets increases with increasing litter size, with small pigs (<1.00 kg) representing <10% of the population in litter sizes of ≤13 piglets and 23% in litter sizes of >15 piglets (Quiniou et al., 2002; Quesnel et al., 2008; Beaulieu et al., 2010). In our herd, 10% of the piglets weighed less than 1 kg at birth and 25% weighed less than 1.25 kg. There is now consistent evidence to suggest that light-weight piglets benefit from cross-fostering that creates uniform litters through improved preweaning performance (English and Bilkei, 2004; Douglas et al., 2014) and reduced preweaning mortality (Milligan et al., 2001; Deen and Bilkei, 2004). However, it is currently unknown what the consequence of this practice is for the performance of normal- or heavy-weight piglets and, therefore, its effectiveness in reducing batch variation. One can hypothesize that uniform litters comprising only heavy-weight piglets would result in high competition for the more productive teats and an increased indirect competition, stimulating teats essential for subsequent milk withdrawal. Therefore, piglets' short- or long-term performance may be penalized. One way of overcoming this may be through the provision of creep feed. The specific objectives of this experiment were based on exactly this thought process; we focused on light-weight and heavy-weight pigs to exaggerate the contrast for the effects of cross-fostering. We aimed to investigate the effect of litter composition and creep feed availability on lifetime performance of piglets born light weight and heavy weight. It was further hypothesized that creep feed provision would convey some benefits on the light-weight piglets but to a lesser extent than on heavy-weight piglets, because the consumption of creep feed seems to be dependent on whether milk consumption is sufficient to support piglets growth. Lastly, we expected that these benefits on the performance as a result of cross-fostering and creep feed provision would be seen in the long term, that is, to slaughter.
There has been some doubt about the beneficial effects of cross-fostering on piglets born light weight (Milligan et al., 2001). Our results, consistent with those of others (Deen and Bilkei 2004; Douglas et al., 2014), suggest that UNIFORM litters benefit piglets born light weight, which exhibit higher weaning weights than similar-sized piglets in MIXED litters. In addition, light-weight piglets in MIXED litters tended to be removed in greater numbers for being too light (<4 kg) at weaning than when reared in UNIFORM litters. It has to be noted, however, that in our study, litter sizes were relatively small (approximately 12 piglets) and that in large litter sizes of the very highly prolific sows (>15 piglets), the positive effect litter uniformity had on light-weight piglets performance, as shown here, might be less apparent. There are several possible explanations for the weight disadvantage light-weight piglets exhibit when reared together with heavy-weight piglets and its effect on weaning weight. First, rearing light-weight piglets in MIXED litters would negatively influence their ability to directly compete for the more productive anterior teats (Scheel et al., 1977; Mason et al., 2003; Drake et al., 2008). Generally, teat position affects preweaning performance, with piglets suckling a posterior teat having a lower milk intake than those suckling an anterior or middle teat (Skok et al., 2007). The latter seems especially apparent in multiparous sows rather than primiparous sows, where neither differences in teat development nor piglet performance were observed (Nielsen et al., 2001). Second, light-weight piglets could be indirectly disadvantaged through their size in the stimulation of teats essential for subsequent milk let down (King et al., 1997; Drake et al., 2008), which depends on the intensity and duration of massaging (Gill and Thomson, 1956). The absence of indirect competition in UNIFORM litters might have resulted in a greater share of the available milk and improved performance of light-weight piglets. In our study, however, teat pair preference was not affected by BiW. Although we did not look at total milk intake, it is unlikely that direct competition contributed to the impaired performance of light-weight piglets reared in MIXED litters. The weight advantage light-weight piglets had at weaning when reared with similar-sized piglets was sustained throughout production, as suggested by Klindt (2003) and Douglas et al. (2014).
Piglets gaining less during early lactation, for example, by suckling the posterior teats or retrieving an unequal share of the available milk, may be expected to eat larger amounts of creep feed (Algers et al., 1990; Appleby et al., 1992). In addition, Sulabo et al. (2007) suggested that the probability to become a nonconsumer increased with increasing BiW. Also, in our work, piglets classified as moderate and high consumers were generally the lightest at 19 d of age, suggesting that creep feed consumption is dependent on whether the amount of milk consumed is sufficient to support requirements for growth. Therefore, it was expected that light-weight piglets in MIXED litters would consume higher amounts of creep feed to compensate for their insufficient milk intake. However, our findings suggest that piglets born light weight, irrespective of litter composition, consumed hardly any creep feed, represented by a high proportion of piglets classified as nonconsumers and having less green feces. Their (low) milk intake might have been sufficient (Pajor et al., 1991) for their reduced growth capacity, as a result of nutrient restriction in utero (Foxcroft et al., 2006). Another explanation for their low creep feed consumption could be their less mature digestive system, represented by a lower trypsin (Cranwell et al., 1997; Pluske et al., 2003) and lipase activity (Pluske et al., 2003) per gram of pancreas compared with heavier piglets. In addition, heavy-weight piglets in MIXED litters might have had a competitive advantage for the access to the creep feeder (Pajor et al., 1991; Bøe and Jensen, 1995), all of which could have contributed to the absence of substantial creep feed consumption by light-weight piglets demonstrated here.
Cross-fostering has been reported to decrease preweaning mortality of piglets born light weight (Milligan et al., 2001; Deen and Bilkei, 2004; Cecchinato et al., 2008) by limiting competition for teat accessibility and, therefore, essential resources. This suggestion was not confirmed here or in some other trials (Douglas et al., 2014). Previous studies suggesting a beneficial effect of litter uniformity on mortality (Milligan et al., 2001; Deen and Bilkei, 2004) have classified light-weight piglets as those weighing less than 1 kg and have applied cross-fostering within 12 h after birth. Survivability, however, decreases with decreasing BiW; piglets weighing less than 1.10 kg have a significantly lower survivability (Feldpausch et al., 2016; Jourquin et al., 2016). Furthermore, performance and BiW are negatively related (Paredes et al., 2012; Douglas et al., 2013), and moving piglets too early might have deprived piglets of access to colostrum (Baxter et al., 2013), which is important for survivability (Devillers et al., 2011). In addition, Deen and Bilkei (2004) suggested that survivability of light-weight piglets might not solely be dependent on littermate weight but that litter size also plays a prominent role in preweaning mortality. Low litter sizes, however, are hard to maintain in herds with very highly prolific sows. Our protocol involving milk supplementation and creep training during the most critical period postpartum (before d 4) has most likely contributed to the absence of a littermate weight effect on preweaning mortality.
The consequences of creating litters with less weight variability on the performance of piglets born heavy weight have often been neglected. This is surprising, because one needs to know the consequences on the performance of all pigs in a system to assess the effectiveness of a management strategy. Although it was expected that litter composition would not affect preweaning performance of piglets born heavy weight or that any disadvantages would be compensated by the provision of creep feed, piglets born heavy weight and reared in UNIFORM litters were weaned almost 1 kg lighter than similar-sized piglets in MIXED litters, irrespective of creep feed provision.
The negative effect littermate weight had on preweaning performance of piglets born heavy weight could have been a result of 1) increased direct competition for the more productive teats in litters with less weight variability, decreasing teat consistency (Baxter et al., 2013; Hales et al., 2013), and 2) the positive association between BiW and piglet efficiency of massaging and draining teats (King et al., 1997), which could give heavy-weight piglets a weight advantage when reared with light-weight piglets (MIXED litters). Sizing piglets for BW may have led to more aggression (Arnott and Elwood, 2009) and, consequently, more disputes and missed suckling bouts (Milligan et al., 2001). In our study, teat consistency was affected by litter composition at 2 d of age. Litters with less weight variability (UNIFORM), irrespective of BiW, had a lower teat consistency. Although piglets generally explore the entire udder during early lactation (Skok and Škorjanc, 2014), decreasing weight variability may have intensified competition, thus decreasing teat consistency (Baxter et al., 2013; Hales et al., 2013). However, teat ownership is often established shortly after birth as delaying teat cohesion compromises survival (Skok and Škorjanc, 2014). This most likely explains why the effect of litter composition on teat consistency was not sustained in the long term.
In addition, our results demonstrated that heavy-weight piglets in UNIFORM litters tended to be removed in higher numbers during the first 10 d of life as a result of subsequent weight loss compared with heavy-weight piglets in MIXED litters. These pigs most likely were involved in teat disputes or unable to access an adequate teat and therefore lost weight. Although heavy-weight piglets in MIXED litters were significantly disadvantaged only when suckling the posterior teat pair, heavy-weight piglets in UNIFORM litters were disadvantaged when suckling both the posterior and middle teats. Milk yield varies with parity; the highest milk yield is seen for sows of parity 2 to 4, after which it decreased (Dourmad et al., 2012). In addition, differences between performance of piglets suckling the anterior teats and piglets suckling posterior teats seem to increase with increasing parity (parity 2 versus parities 3–4; Dyck et al., 1987). Given that this study used older sows, milk yield and preferred teat position could have substantially limited their performance. Nevertheless, this suggests that 1) the weight advantage heavy-weight piglets had in MIXED litters resulted in an unequal milk distribution across teats favoring the heavier piglets and 2) the increase in indirect competition for heavy-weight piglets in UNIFORM litters resulted in less milk intake per piglet.
Furthermore, it was observed that the weight advantage heavy-weight piglets had in MIXED litters was sustained during the different phases of production. Although it could have been argued that piglets are able to compensate growth once restrictions are eliminated, it has been suggested that nutrient intake during suckling “sets” animals' appetite during later life (Hales and Barker, 2001). In addition, keeping littermates together during nursery (d 28–61) could have given heavy-weight piglets in MIXED litters a competitive advantage for the feeder, whereas the relatively lower space allowance for heavy-weight pigs in UNIFORM litters could have restricted their growth (Vermeer et al., 2014).
To our knowledge, this is the first study that has investigated the effect of different litter compositions on creep feed disappearance. Although chromic oxide is commonly used to discriminate between consumers and nonconsumers of creep feed, the absence of the dye does not necessarily rule out that the piglet has eaten creep feed. Small amounts of creep feed could have been diluted by a high amount of milk (Barnett et al., 1989; Kuller et al., 2007c) and, when consumed for only 1 d, be difficult to detect (Kuller et al., 2007c); therefore, in this study, a color reader was used to objectively assess fecal color. Creep feed consumption is believed to be influenced by littermates, as individuals that start eating creep feed could motivate unexperienced piglets within the same litter (Oostindjer et al., 2014), and by teat position, with piglets suckling the posterior teats eating larger amounts of creep feed (Algers et al., 1990). The latter was supported by our data, as fecal color was significantly affected by teat pair class: feces of piglets suckling the anterior teats were perceived to be less green. The results presented here demonstrated that litter composition influenced creep feed consumption, with heavy-weight piglets in UNIFORM litters consuming the highest amount of creep feed and having the greenest feces. In addition, a significantly higher number of heavy-weight piglets reared in UNIFORM litters were already consuming creep feed by d 19 as well as a lower proportion of heavy-weight piglets being classified as nonconsumers compared with heavy-weight piglets in MIXED litters and light-weight piglets in UNIFORM and MIXED litters. The increased competition for heavy-weight piglets in UNIFORM litters, leading to insufficient milk intake, might have driven these piglets to consume more creep feed.
Consistent with the results of Sulabo et al. (2007), Collins et al. (2013), and Blavi et al. (2015), creep feed provision did not influence weaning weight nor did it contribute to an improved litter CV at weaning. However, not every piglet consumed creep feed, and more than half of the creep feed was eaten during the last week before weaning, as shown in our work and that of others (Barnett et al., 1989; Bruininx et al., 2002). Although high and moderate consumers gained less weight during most of the suckling period and were lighter 1 wk prior to weaning, consumer class did not affect piglets' weaning weight or ADG between d 19 and weaning. The latter suggests that high consumers were able to catch up in growth. In addition, once creep feed was consumed in sufficient quantities, it tended to decrease variation at weaning, as illustrated by the lower CV for heavy-weight piglets. Appleby et al. (1992) found a negative correlation between ADG during the initial 3 wk of lactation and feeding score and a positive correlation between feeding score and ADG between d 21 and 28. The latter was supported by the colorimetric results of the present study and of Kuller et al. (2007a). Being a high consumer before weaning is also believed to positively affect piglets' postweaning performance. Various studies have shown that piglets classified as high consumers performed better during the most critical period after weaning by starting to eat sooner (Bruininx et al., 2002), gaining weight faster (Collins et al., 2013; Blavi et al., 2015), and having a decreased risk for postweaning diarrhea (Kuller et al., 2007b) compared with those classified as nonconsumers. Also, in the current study, ADG between weaning and the grower phase of piglets classified as high consumers was significantly higher than for piglets classified as non-, low, or moderate consumers.
Conclusions
The present study tested the effectiveness of cross-fostering as a management strategy by offsetting the effect cross-fostering had on piglets born light weight to that on piglets born heavy weight. The results presented here demonstrate that light-weight piglets in UNIFORM litters were weaned 6% heavier and removed in lower numbers (−4.6%) than light-weight piglets in MIXED litters. This weight advantage was evident to slaughter. Although litter uniformity successfully improved pre- and postweaning performance of piglets born light weight, BiW played a greater role in subsequent performance (Douglas et al., 2014), with light-weight piglets needing 4 extra days to reach slaughter weight than piglets born heavy weight.
On the other hand, heavy-weight piglets reared in UNIFORM litters were 12% lighter at weaning than those reared in MIXED litters. The overall removal of heavy-weight piglets in UNIFORM litters was considerably higher (+6.5%) than that in MIXED litters. Although preweaning litter gain was not affected by litter composition, also here, weight differences were sustained throughout production, with heavy-weight piglets from UNIFORM litters being 6.0, 6.6, and 2.3% lighter at the grower, finisher, and slaughter stages, respectively. The results imply that the positive effect of cross-fostering on piglets born light weight did not outweigh its negative effect on piglets born heavy weight and also did not contribute to an increase in pre- and postweaning litter gain. However, more information is needed to confirm our results evaluating the effect of littermate weight on mortality and subsequent performance of all piglets in the very prolific sows. In addition, a bioeconomic analysis is necessary to assess what is the best strategy.
Furthermore, our results suggest that heavy-weight piglets in UNIFORM litters tried to compensate for their insufficient milk intake by increasing creep feed consumption. However, this was not sufficient to overcome their growth disadvantage compared with heavy-weight piglets in MIXED litters. Piglets classified as high consumers were generally the lightest in the week prior to weaning; however, they were able to show catch-up growth. Furthermore, being a high consumer before weaning contributed to improved growth after weaning.
APPENDIX I: Methodology for correcting colorimetric readings of creep-fed piglets
True a* from non-creep-fed piglets was averaged per experimental day and litter composition. The latter was subtracted from individual true a* observations of piglets reared in creep-fed litters, resulting in adjusted a*, using the following equation:
The non-creep-fed reference is based on the average for each experimental day (d 13, 19, 21, 23, and 25) and for each litter composition (litters with only similar-sized piglets [light weight or heavy weight; UNIFORM litters] and litters with equal numbers of light-weight and heavy-weight piglets [MIXED litters]) separately.
Adjusted a* obliterated the effect of experimental day (P = 0.989) and the interaction between experimental day and litter composition (P = 0.939), as previously seen in non-creep-fed litters. Therefore, it is likely that any difference in adjusted a* found between piglets reared in creep-fed litters was the result of differences in creep feed consumption. Correcting hue angle (H*) required adjusted a*, adjusted b*, and adjusted Chroma (C*ab). Adjusted C*ab represent the difference in chroma, in which positive numbers indicate that the sample is brighter than the reference sample and negative values indicate that the sample is duller. The following equations were used (Konica Minolta, 2007):
in which H*ab is adjusted H*.
True b* was necessary for calculating adjusted C*ab and, therefore, adjusted H*ab but appeared to be significantly affected by experimental day (P < 0.001), birth weight (BiW) class (P = 0.0024), and the interaction between litter composition and BiW class (P = 0.0487). Therefore, true b* values had to be corrected using the following equation:
The non-creep-fed reference was based on the average of each experimental day (d 13, 19, 21, 23, and 25), each litter composition (UNIFORM and MIXED), and each BiW class (light and heavy) separately.
For example, true b* values of creep feed samples that were collected on d 21 from piglets born light weight and reared in UNIFORM litters were corrected by subtracting the average non-creep-fed reference of feces collected on d 21 of light-weight piglets reared in UNIFORM litters. Using the obtained adjusted b* for calculating adjusted H*ab successfully diminished the effect of experimental day (P = 0.217) and the interaction experimental day and litter composition (P = 0.700) previously seen.
Footnotes
This project was sponsored by AHDB (Agriculture and Horticulture Development Board) Pork and Primary Diets
LITERATURE CITED
- Pork AHDB. 2016. 2015 Pig cost of production in selected countries. https://pork.ahdb.org.uk/media/272651/2015-pig-cost-of-production-in-selected-countries.pdf. (Accessed 20 August 2017.)
- Algers B., Jensen P., Steinwall L. 1990. Behaviour and weight changes at weaning and regrouping of pigs in relation to teat quality. Appl. Anim. Behav. Sci. 26:143–155. doi: 10.1016/0168-1591(90)90094-T [DOI] [Google Scholar]
- Appleby M. C., Pajor E. A., Fraser D. 1992. Individual variation in feeding and growth of piglets: Effect of increased access to creep food. Anim. Sci. 55:147–152. [Google Scholar]
- Arnott G., Elwood R. W. 2009. Assessment of fighting ability in animal contests. Anim. Behav. 77:991–1004. doi: 10.1016/j.anbehav.2009.02.010 [DOI] [Google Scholar]
- Barnett K. L., Kornegay E. T., Risley C. R., Lindemann M. D., Schurig G. G. 1989. Characterization of creep feed consumption and its subsequent effect on immune response, scouring index and performance of weanling pigs. J. Anim. Sci. 67:2698–2708. doi: 10.2527/jas1989.67102698x [DOI] [PubMed] [Google Scholar]
- Baxter E. M., Rutherford K. M. D., D'Eath R. B., Arnott G., Turner S. P., Sandøe P., Moustsen V. A., Thorup F., Edwards S. A., Lawrence A. B. 2013. The welfare implications of large litter size in the domestic pig II: Management factors. Anim. Welf. 22:219–238. doi: 10.7120/09627286.22.2.219 [DOI] [Google Scholar]
- Beaulieu A. D., Aalhus J. L., Williams N. H., Patience J. F. 2010. Impact of piglet birth weight, birth order, and litter size on subsequent growth performance, carcass quality, muscle composition, and eating quality of pork. J. Anim. Sci. 88:2767–2778. doi: 10.2527/jas.2009-2222 [DOI] [PubMed] [Google Scholar]
- Blavi L., Solà-Oriol D., Pérez J. 2015. Effect of supplementary feeding strategies during the suckling period to improve weanling performance. In: Proc. 13th Dig. Physiol. Pigs Symp., Kliczków, Poland. p. 49. [Google Scholar]
- Bøe K., Jensen P. 1995. Individual differences in suckling and solid food intake by piglets. Appl. Anim. Behav. Sci. 42:183–192. doi: 10.1016/0168-1591(94)00528-M [DOI] [Google Scholar]
- Bruininx E. M. A. M., Binnendijk G. P., van der Peet-Schwering C. M. C., Schrama J. W., den Hartog L. A., Everts H, Beynen A. C. 2002. Effect of creep feed consumption on individual feed intake characteristics and performance of group-housed weanling pigs. J. Anim. Sci. 80:1413–1418. doi: 10.2527/2002.8061413x [DOI] [PubMed] [Google Scholar]
- Bruininx E. M. A. M., Schellingerhout A. B., Binnendijk G. P., van der Peet-Schwering C. M. C., Schrama J. W., den Hartog L. A., Everts H, Beynen A. C. 2004. Individually assessed creep food consumption by suckled piglets: Influence of post-weaning food intake characteristics and indicators of gut structure and hind-gut fermentation. Anim. Sci. 78:67–75. doi: 10.1017/S1357729800053856 [DOI] [Google Scholar]
- Cecchinato A., Bonfatti V., Gallo L., Carnier P. 2008. Survival analysis of preweaning piglet survival in a dry-cured ham-producing crossbred line. J. Anim. Sci. 86:2486–2495. doi: 10.2527/jas.2007-0825 [DOI] [PubMed] [Google Scholar]
- Collins C. L., Morrison R. S., Smits R. J., Henman D. J., Dunshea F. R., Pluske J. R. 2013. Interactions between piglet weaning age and dietary creep feed composition on lifetime growth performance. Anim. Prod. Sci. 53:1025–1032. doi: 10.1071/AN12009 [DOI] [Google Scholar]
- Cranwell P. D., Pierzynowski S. G., Rippe C., Pluske J. R., Power G. N., Campbell R. G., Kerton D. J., King R. H., Dunshea F. R. 1997. Weight and age at weaning influence pancreatic size and enzymatic capacity. In: Manipulating Pig Production VI. Proc. 6th Biennial Congr. Australas. Pig Sci. Assoc., Canberra, Australia: p. 66. [Google Scholar]
- Deen M. G. H., Bilkei G. 2004. Cross fostering of low-birthweight piglets. Livest. Prod. Sci. 90:279–284. doi: 10.1016/j.livprodsci.2004.02.012 [DOI] [Google Scholar]
- Devillers N., Le Dividich J., Prunier A. 2011. Influence of colostrum intake on piglet survival and immunity. Animal 5(10):1605–1612. doi: 10.1017/S175173111100067X [DOI] [PubMed] [Google Scholar]
- Douglas S. L., Edwards S. A., Kyriazakis I. 2014. Management strategies to improve the performance of low birth pigs to weaning and their long-term consequences. J. Anim. Sci. 92:2280–2288. doi: 10.2527/jas.2013-7388 [DOI] [PubMed] [Google Scholar]
- Douglas S. L., Edwards S. A., Sutcliffe E., Knap P. W., Kyriazakis I. 2013. Identification of risk factors associated with poor lifetime growth performance in pigs. J. Anim. Sci. 91:4123–4132. doi: 10.2527/jas.2012-5915 [DOI] [PubMed] [Google Scholar]
- Dourmad J. Y., Quiniou N., Heugebaert S., Paboeuf F., Ngo T. T. 2012. Effect of parity and number of suckling piglets on milk production of sows. In: Book of Abstracts of the 63rd Annual Meeting of the European Federation of Animal Science. Wageningen Academic Publishers, Wageningen, the Netherlands: p. 44. [Google Scholar]
- Drake A., Fraser D., Weary D. M. 2008. Parent-offspring resource allocation in domestic pigs. Behav. Ecol. Sociobiol. 62:309–319. doi: 10.1007/s00265-007-0418-y [DOI] [Google Scholar]
- Dyck G. W., Swierstra E. E., McKay R. M., Mount K. 1987. Effect of location of the teat suckled, breed and parity on piglet growth. Can. J. Anim. Sci. 67:929–939. doi: 10.4141/cjas87-098 [DOI] [Google Scholar]
- English J. G. H., Bilkei G. 2004. The effect of litter size and littermate weight on pre-weaning performance of low-birth-weight piglets that have been cross-fostered. Anim. Sci. 79:439–443. doi: 10.1017/S1357729800090305 [DOI] [Google Scholar]
- Feldpausch J. A., Jourquin J., Bergstrom J. R., Bokenkroger C. D., Nelssen J. L., Ritter M. J., Davis D. L., Gonzalez J. M. 2016. Birth weight threshold for identifying piglets at-risk for preweaning mortality. J. Anim. Sci. 94(Suppl. 2):34 (Abstr.) doi:10.2527/msasas2016-074 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrari C. V., Sbardella P. E., Bernardi M. L., Coutinho M. L., Vaz L. S., Jr, Wentz I., Bortolozzo F. P. 2014. Effect of birth weight and colostrum intake on mortality and performance of piglets after cross-fostering in sows of different parities. Prev. Vet. Med. 114:259–266. doi: 10.1016/j.prevetmed.2014.02.013 [DOI] [PubMed] [Google Scholar]
- Foxcroft G. R., Dixon W. T., Novak S., Putman C. T., Town S. C., Vinsky M. D. A. 2006. The biological basis for prenatal programming of postnatal performance in pigs. J. Anim. Sci. 84(Suppl. 13):E105–E112. doi: 10.2527/2006.8413_supplE105x [DOI] [PubMed] [Google Scholar]
- Gill J. C., Thomson W. 1956. Observations on the behaviour of suckling pigs. Anim. Behav. 4(2):46–51. doi: 10.1016/S0950-5601(56)80022-1 [DOI] [Google Scholar]
- Gondret F., Lefaucheur L., Juin H., Louveau I., Lebret B. 2006. Low birth weight is associated with enlarged muscle fiber area and impaired meat tenderness of the longissimus muscle in pigs. J. Anim. Sci. 84:93–103. [DOI] [PubMed] [Google Scholar]
- Hales C. N., Barker D. J. P. 2001. The thrifty phenotype hypothesis. Br. Med. Bull. 60:5–20. doi: 10.1093/bmb/60.1.5 [DOI] [PubMed] [Google Scholar]
- Hales J., Moustsen V. A., Nielsen M. B. F., Hansen C. F. 2013. Individual physical characteristics of neonatal piglets affect preweaning survival of piglets born in a noncrated system. J. Anim. Sci. 91:4991–5003. doi: 10.2527/jas.2012-5740 [DOI] [PubMed] [Google Scholar]
- Hänninen L., Pastell M. 2009. CowLog: Open-source software for coding behaviors from digital video. Behav. Res. Methods 41(2):472–476. doi: 10.3758/BRM.41.2.472 [DOI] [PubMed] [Google Scholar]
- Hazzledine M. 2008. Premier Atlas ingredients matrix. Premier Nutrition Products Ltd., Rugeley, UK. [Google Scholar]
- Jourquin J., Morales J., Bokenkroger C. D. 2016. Impact of piglet birth weight increase on survivability and days to market, a simulation model. J. Anim. Sci. 94(Suppl. 2):34. doi: 10.2527/msasas2016-073 [DOI] [Google Scholar]
- Kim S. W., Hurley W. L., Han I. K., Easter R. A. 2000. Growth of nursing pigs related to the characteristics of nursed mammary glands. J. Anim. Sci. 78:1313–1318. doi: 10.2527/2000.7851313x [DOI] [PubMed] [Google Scholar]
- King R. H., Mullan B. P., Dunshea F. R., Dove H. 1997. The influence of piglet body weight on milk production of sows. Livest. Prod. Sci. 47:169–174. doi: 10.1016/S0301-6226(96)01404-2 [DOI] [Google Scholar]
- Klindt J. 2003. Influence of litter size and creep feeding on preweaning gain and influence of preweaning growth on growth to slaughter in barrow. J. Anim. Sci. 81:2434–2439. doi: 10.2527/2003.81102434x [DOI] [PubMed] [Google Scholar]
- Konica Minolta 2007. Precise color communication. https://www.konicaminolta.com/instruments/knowledge/color/pdf/color_communication.pdf. (Accessed 10 December 2015.)
- Kuller W. I., Soede N. M., van Beers-Schreurs H. M. G., Langendijk P., Taverne M. A. M., Kemp B., Verheijden J. H. M. 2007a. Effect of intermittent suckling and creep feed intake on pig performance from birth to slaughter. J. Anim. Sci. 85:1295–1301. doi: 10.2527/jas.2006-177 [DOI] [PubMed] [Google Scholar]
- Kuller W. I., van Beers-Schreurs H. M. G., Soede N. M., Langendijk P., Taverne M. A. M., Kemp B., Verheijden J. H. M. 2007b. Creep feed intake during lactation enhances net absorption in the small intestine after weaning. Livest. Sci. 108:99–101. doi: 10.1016/j.livsci.2007.01.003 [DOI] [Google Scholar]
- Kuller W. I., van Beers-Schreurs H. M. G., Soede N. M., Taverne M. A. M., Kemp B., Verheijden J. H. M. 2007c. Addition of chromic oxide to creep feed as a fecal marker for selection of creep feed-eating suckling pigs. Am. J. Vet. Res. 68(7):748–752. doi: 10.2460/ajvr.68.7.748 [DOI] [PubMed] [Google Scholar]
- Mason S. P., Jarvis S., Lawrence A. B. 2003. Individual differences in responses of piglets to weaning at different ages. Appl. Anim. Behav. Sci. 80:117–132. doi: 10.1016/S0168-1591(02)00209-5 [DOI] [Google Scholar]
- Milligan B. N., Fraser D., Kramer D. L. 2001. The effect of littermate weight on survival, weight gain, and suckling behavior of low-birth-weight piglets on cross-fostered litters. J. Swine Health Prod. 9(4):161–166. [Google Scholar]
- Nielsen O. L., Pedersen A. R., Sørensen M. T. 2001. Relationship between piglet growth rate and mammary gland size of the sow. Livest. Prod. Sci. 67:273–279. doi: 10.1016/S0301-6226(00)00197-4 [DOI] [Google Scholar]
- Oostindjer M., Kemp B., van den Brand H., Bolhuis J. E. 2014. Facilitating ‘learning from mom how to eat like a pig’ to improve welfare of piglets around weaning. Appl. Anim. Behav. Sci. 160:19–30. doi: 10.1016/j.applanim.2014.09.006 [DOI] [Google Scholar]
- Pajor E. A., Fraser D., Kramer D. L. 1991. Consumption of solid food by suckling pigs: Individual variation and relation to weight gain. Appl. Anim. Behav. Sci. 32:139–155. doi: 10.1016/S0168-1591(05)80038-3 [DOI] [Google Scholar]
- Paredes S. P., Jansman A. J., Verstegen M. W., Awati A., Buist W., den Hartog L. A., van Hees H. M., Quiniou N., Hendriks W. H., Gerrits W. J. 2012. Analysis of factors to predict piglet body weight at the end of the nursery phase. J. Anim. Sci. 90:3243–3251. doi: 10.2527/jas.2011-4574 [DOI] [PubMed] [Google Scholar]
- Pluske J. R., Kerton D. J., Cranwell P. D., Campbell R. G., Mullan B. P., King R. H., Power G. N., Pierzynowski S. G., Westrom B., Rippe C., Peulen O., Dunshea F. R. 2003. Age, sex, and weight at weaning influence organ weight and gastrointestinal development of weanling pigs. Aust. J. Agric. Res. 54:515–527. doi: 10.1071/AR02156 [DOI] [Google Scholar]
- Quesnel H., Brossard L., Valancogne A., Quiniou N. 2008. Influence of some sow characteristics on within-litter variation of piglet birth weight. Animal 2:1842–1849. doi: 10.1017/S175173110800308X [DOI] [PubMed] [Google Scholar]
- Quiniou N., Dagorn J., Gaudré D. 2002. Variation of piglets' birth weight and consequences on subsequent performance. Livest. Prod. Sci. 78:63–70. doi: 10.1016/S0301-6226(02)00181-1 [DOI] [Google Scholar]
- Rutherford K. M. D., Baxter E. M., D'Eath R. B., Turner S. P., Arnott G, Roehe R., Ask B., Sandøe P., Moustsen V. A., Thorup F., Edwards S. A., Berg P., Lawrence A. B. 2013. The welfare implications of large litter size in the domestic pig I: Biological factors. Anim. Welf. 22:199–218. doi: 10.7120/09627286.22.2.199 [DOI] [Google Scholar]
- Scheel D. E., Graves H. B., Sherritt G. W. 1977. Nursing order, social dominance and growth in swine. J. Anim. Sci. 45(2):219–229. doi: 10.2527/jas1977.452219x [DOI] [Google Scholar]
- Schinckel A. P., Cabrera R., Boyd R. D., Jungst S., Booher C., Johnston M., Einstein M. E. 2007. Impact of birth and body weight at twenty days on the postweaning growth of pigs with different weaning management. Prof. Anim. Sci. 23:197–210. [Google Scholar]
- Skok J., Brus M., Škorjanc D. 2007. Growth of piglets in relation to milk intake and anatomical location of mammary glands. Acta Agric. Scand., Sect. A 57:129–135. doi: 10.1080/09064700801907089 [DOI] [Google Scholar]
- Skok J., Škorjanc D. 2014. Group suckling cohesion as a prelude to the formation of teat order in piglets. Appl. Anim. Behav. Sci. 154:15–21. doi: 10.1016/j.applanim.2014.02.003 [DOI] [Google Scholar]
- Sulabo R. C., Tokach M. D., Jacela J. Y., Wiedemann E. J., Nelssen J. L., Dritz S. S., DeRouchey J. M., Goodband R. D. 2007. Effects of lactation feed intake and creep feeding on sow and piglet performance. In: Swine day. Kansas State University, Manhattan, KS: p. 24–37. doi: 10.4148/2378-5977.6975 [DOI] [PubMed] [Google Scholar]
- Vermeer H. M., de Greef K. H., Houwers H. W. J. 2014. Space allowance and pen size affect welfare indicators and performance of growing pigs under comfort class conditions. Livest. Sci. 159:79–86. doi: 10.1016/j.livsci.2013.10.021 [DOI] [Google Scholar]