

Abstract
Background
Ruminal microbes are vital to the conversion of lignocellulose-rich plant materials into nutrients for ruminants. Although protozoa play a key role in linking ruminal microbial networks, the contribution of protozoa to rumen fermentation remains controversial; therefore, this meta-analysis was conducted to quantitatively summarize the temporal dynamics of methanogenesis, ruminal volatile fatty acid (VFA) profiles and dietary fiber digestibility in ruminants following the elimination of protozoa (also termed defaunation). A total of 49 studies from 22 publications were evaluated.
Results
The results revealed that defaunation reduced methane production and shifted ruminal VFA profiles to consist of more propionate and less acetate and butyrate, but with a reduced total VFA concentration and decreased dietary fiber digestibility. However, these effects were diminished linearly, at different rates, with time during the first few weeks after defaunation, and eventually reached relative stability. The acetate to propionate ratio and methane production were increased at 7 and 11 wk after defaunation, respectively.
Conclusions
Elimination of protozoa initially shifted the rumen fermentation toward the production of more propionate and less methane, but eventually toward the production of less propionate and more methane over time.
Electronic supplementary material
The online version of this article (10.1186/s40104-018-0305-6) contains supplementary material, which is available to authorized users.
Keywords: Defaunation, Fiber digestibility, Meta-analysis, Methane production, Rumen fermentation
Introduction
The rumen provides an ideal habitat for protozoa, whose concentration can reach 105–106 cells/mL. In return, protozoa serve important functions in the rumen microbial ecosystem, such as predation, competition for nutrients, and involvement in symbiotic relationships with other microorganisms [1, 2]. Protozoa prey on bacteria and fungal spores, but are preferentially retained in the rumen, thus reducing the postruminal microbial protein supply [3]. Protozoa compete with amylolytic bacteria for dietary starch, which is mostly fermented into acetate by protozoa [2] while mostly into propionate by amylolytic bacteria [3, 4]. For host animals, the energy recovery efficiency is reduced by 38% when the substrate (glucose) is fermented into acetate but increased by 9% when fermented into propionate [3, 4]. Protozoa are important ruminal hydrogen (H2) producers, and the produced H2 is mostly converted into methane (CH4) by methanogens situated inside protozoa or on their external surface [5–7]. The CH4 emissions from ruminants represent 2–12% dietary energy loss [8]. Therefore, the presence of protozoa seems to adversely affect animals’ energy efficiency.
Complete removal of ruminal protozoa, termed defaunation, has been suggested as an efficient method for reducing CH4 emissions and enhancing propionate fermentation [9, 10], but these effects have not been consistently observed in studies investigating this 112method [11–13]. Hegarty et al. [12] and Morgavi et al. [13] suggested that the duration of defaunation might be responsible for this inconsistency, but the temporal dynamics of methanogenesis and ruminal volatile fatty acid (VFA) profiles after defaunation are difficult to determine experimentally because of the difficulties in raising defaunated animals over a long-term [14]. Meta-analysis is a statistical method that combines the results from multiple studies to achieve a more precise estimate of treatment effects and to explore the potential sources of between-study heterogeneity [15, 16]. Two prior meta-analyses [14, 17] have summarized the combined responses of rumen fermentation to defaunation; however, the combined effects on ruminal VFA profiles were also inconsistent between them, and neither of them explored the time-dependent effects. Therefore, the current meta-analysis was conducted to quantitatively summarize the temporal dynamics of methanogenesis, ruminal VFA profiles and dietary fiber digestibility in ruminants after defaunation, and to explain the contribution of the defaunated duration to the between-study heterogeneity.
Methods
Literature search, screening and data extraction
A flowchart detailing the process of literature search, screening and data extraction is shown in Fig. 1. The inclusion criteria for the studies were as follows: (1) peer-reviewed and published in the English language; (2) complete defaunation in vivo; (3) inclusion of relevant variables for extraction. The relevant variables for this meta-analysis included the daily CH4 production, ruminal VFA profiles and dietary fiber total-tract digestibility.
Fig. 1.
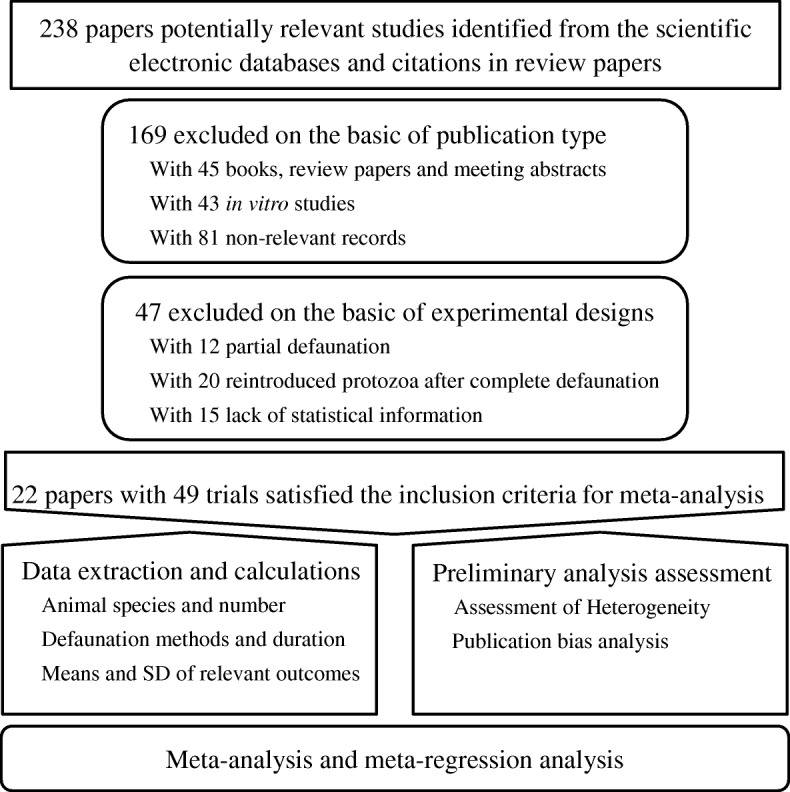
Flow chart of the literature search, screening and data extraction procedures
The faunation states of control animals included the natural ruminal ecosystem without any treatment or reintroduction of protozoa after partial defaunation, which appeared to be restored quickly after withdrawal of the protozoa-inhibiting treatment [18, 19]. The control animals into which protozoa were reintroduced after complete defaunation were excluded from the analysis, because preliminary analysis showed that high heterogeneity existed between the faunation and refaunation subgroups (see Additional file 1). Protozoa-free animals were obtained through either the absence of protozoa from birth (BF) or artificial removal of protozoa from the natural ruminal ecosystem (AF). Artificial defaunation was conducted using chemical agents, such as alkanes and sodium lauryl sulfate, or by applying a rumen washing technique. The defaunation duration was calculated based on the schedule of experimental activities. The defaunation duration of BF animals was calculated as their age shortened by 4 wk, because ruminal protozoa did not appear when newborn calves were fed with milk for 30 d [2], and the concentration of protozoa quickly increased after 5 wk of age [20].
The final database included 22 publications, with 49 in vivo studies that satisfied the inclusion criteria for the meta-analysis. Summary descriptions of the selected studies are provided in Table 1. Briefly, among the 49 selected studies, 38 were conducted in sheep, and 11 were conducted in cattle; 18 measured CH4 production, 29 measured ruminal VFAs, and 15 measured total-tract fiber digestibility. CH4 emissions were measured using the respiration chamber technique or sulfur hexafluoride tracer technique. Methane production was presented in liters per day in most of the studies; thus, values presented in grams or kilojoules (kJ) were converted to liters per day, based on the assumption that one mole of CH4 weighs 16 g or contains 890 kJ of energy and occupies a volume of 24.5 L (under conditions of 25 °C and 1 atmospheric pressure).
Table 1.
Data sources and characteristics of the studies included in the meta-analysis
| Source | Trials | Animal | Defaunation duration | Outcomes |
|---|---|---|---|---|
| Belanche et al. [44] | 2 | Sheep | 23 wk | VFA, digestibility |
| Bird et al. [11] | 2 | Sheep | 11, 26 wk | CH4, VFA, digestibility |
| Chandramoni et al. [45] | 2 | Sheep | 5, 11 wk | CH4, digestibility |
| Chaudhary and Srivastava [46] | 2 | Cattle | 18 wk | Digestibility |
| Eadie and Gill [47] | 2 | Sheep | 22, 55 wk | VFA |
| Eugène et al. [48] | 4 | Sheep | 10, 14, 18, 22 wk | Digestibility |
| Frumholtz [38] | 3 | Sheep | 5, 26, 52 wk | VFA |
| Hegarty et al. [12] | 4 | Sheep | 12, 22, 24, 33 wk | CH4, VFA |
| Kasuya et al. [49] | 1 | Cattle | 21 wk | Digestibility |
| Kreuzer et al. [50] | 3 | Sheep | 9, 10, 11 wk | CH4 |
| Morgavi et al. [13] | 2 | Sheep | 6 wk, 2 yr | CH4, VFA |
| Nagaraja et al. [51] | 2 | Sheep | 14 wk | VFA |
| Nguyen et al. [33] | 2 | Sheep | 9 wk | CH4, VFA, digestibility |
| Ozutsumi et al. [35] | 1 | Cattle | 14 wk | VFA |
| Santra and Karim [52] | 2 | Sheep | 12 wk | Digestibility |
| Santra and Karim [53] | 3 | Sheep | 14 wk | Digestibility |
| Santra et al. [54] | 2 | Sheep | 8 wk | Digestibility |
| Schönhusen et al. [20] | 4 | Cattle | 4, 5, 6, 7 wk | CH4, VFA, digestibility |
| Sultana et al. [55] | 1 | Cattle | 14 wk | VFA |
| Williams and Dinusson [56] | 2 | Cattle | 30, 56 wk | VFA |
| Yáñez-Ruiz et al. [57] | 1 | Sheep | 18 wk | VFA |
| Zhou et al. [58] | 2 | Sheep | 5 wk | CH4, VFA |
Data analysis
The meta-analysis was performed using Stata 14.1 (Stata Corp., Texas, USA).
Assessment of heterogeneity and effect size
Between-study variability was quantified via the I2 statistic, which measures the percentage of variation due to heterogeneity [15, 21]. When the I2 value was less than 50%, indicating low heterogeneity, studies were combined using a fixed effects model, which was based on the assumption that the expected effect from each study was homogeneous. When the value of I2 was over 50%, indicating high heterogeneity, studies were combined using a random effects model, based on the assumption that the expected effect from each study was heterogeneous.
The differences in animal species or ages, daily sampling times or dietary forage percentages across the studies caused that the data of certain relevant variables to vary greatly across the studies (Table 2). To reduce these potential interferences, the effect size in this analysis was estimated via the standardized mean difference (SMD), which was calculated as the raw mean difference between the treatment and control groups divided by their pooled standard deviations [15]. For example, although cattle CH4 production in the study by Schönhusen et al. [20] was higher than in other subgroup studies involving sheep, it was homogeneous with most of them (Additional file 1: Figures S1 and S2). The studies were weighted using the inverse of the variance of the differences in means. Details of the calculations used in the meta-analysis are provided by Lean et al. [15].
Table 2.
Data summary and meta-analysis of relevant variables based on all of the selected studies
| Variables | No. of trials | Defaunation group | Faunation group | Meta-analysis | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| n | Mean | SD | n | Mean | SD | I2, % | SMD | P-value | ||
| CH4, L/d | 18 | 126 | 20.8 | 12.4 | 129 | 23.4 | 12.0 | 71.9 | −0.602 | 0.037 |
| Total VFA, mmol/L | 29 | 211 | 78.8 | 29.7 | 212 | 87.1 | 31.3 | 44.2 | −0.549 | < 0.001 |
| Individual VFA molar proportion, % | ||||||||||
| Acetate | 29 | 210 | 67.7 | 5.6 | 211 | 66.3 | 4.3 | 67.8 | 0.358 | 0.083 |
| Propionate | 29 | 210 | 21.0 | 4.8 | 211 | 20.6 | 3.1 | 73.5 | 0.150 | 0.515 |
| Butyrate | 27 | 202 | 8.2 | 2.0 | 203 | 10.2 | 2.8 | 68.3 | −1.026 | < 0.001 |
| A:P | 18 | 78 | 3.6 | 1.6 | 81 | 3.5 | 0.7 | 77.0 | −0.284 | 0.493 |
| Total-tract fiber digestibility, % | ||||||||||
| NDF | 15 | 109 | 55.0 | 11.6 | 109 | 58.1 | 11.8 | 55.7 | −2.063 | < 0.001 |
| ADF | 11 | 82 | 42.8 | 2.7 | 82 | 45.7 | 3.7 | 69.8 | −3.075 | < 0.001 |
n number of animals, I2 percentage of heterogeneity across studies, SMD standardized mean difference, A:P acetate: propionate ratio, NDF neutral detergent fiber, ADF acid detergent fiber
Meta-regression analysis
The meta-regression analysis was performed using the Knapp-Hartung restricted maximum likelihood method [22], with the SMD of the individual studies used as the response variable and the corresponding standard error of the SMD used as the variance. The percentage of between-study heterogeneity explained by the covariate (defaunation duration) was quantified via the adjusted R2 value.
Preliminary analysis showed that the temporal SMD dynamics after defaunation included a linear phase followed by a plateau phase. The durations of the linear phase and plateau phase for each outcome were dependent on the highest adjusted R2 of the meta-regression analysis and the minimum I2 of the heterogeneity analysis, respectively; therefore, the two phases might overlap over a short duration. The probability levels were set at P < 0.05 for significance and 0.05 ≤ P < 0.10 for a trend.
Results
Effect size and heterogeneity across all the studies
The meta-analysis based on all the selected studies showed that elimination of rumen protozoa reduced (P < 0.05) CH4 production, ruminal VFA concentration, the proportion of butyrate and dietary fiber digestibility, and tended to increase (P = 0.083) the proportion of acetate (Table 2). However, the heterogeneity across the studies was considerable (I2 > 50%) for most of the responses to defaunation, except for the ruminal VFA concentration (I2 = 44.2%).
Methanogenesis dynamics during adaptation to defaunation
Compared with that of faunation, the effect size of defaunation on CH4 emissions presented a linear relationship over time during the first 12 wk (linear phase) after defaunation (Fig. 2 and Table 3): CH4 production was reduced by defaunation (intercept = − 5.484, P = 0.003), and the reduction decreased weekly by 0.486 (P = 0.003) until 12 wk. The defaunation duration explained 76.8% of the between-study heterogeneity (I2 = 76.2%) during the linear phase, and no between-study heterogeneity (I2 = 0.0%) was observed after 11 wk (plateau phase), suggesting that little fluctuation occurred during the plateau phase. Interestingly, the defaunated animals during the plateau phase presented higher CH4 productions (SMD = 0.313, P = 0.039) than the control animals.
Fig. 2.

Methanogenesis dynamics during adaptation to defaunation. The temporal SMD (standardized mean difference) dynamics (a) after defaunation included a linear phase (≤ 12 wk, yellow portion) followed by a plateau phase (≥ 11 wk, blue portion). The overlap of two phases were the green portion. The linear phase was reanalyzed by meta-regression analysis (b), and the plateau phase was reanalyzed by meta-analysis (c). Circles in the graph represent the estimates from each study, and the size of the circles represents the percentage of weight of each study. The blue diamond represent in panel (c) represents the combined effect and its 95% confidence interval
Table 3.
Methane emissions, ruminal VFA profiles and total-tract fiber digestibility dynamics during the linear phase and plateau phase
| Meta-regression analysis for the linear phase | Meta-analysis for the plateau phase | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Duration, wk | No. of trials | I2, % | Intercept | P-value | Coefficient | P-value | Adjusted R2, % | Duration, wk | No. of trials | I2, % | Pooled effect size | P-value | |
| CH4 | ≤ 12 | 13 | 76.2 | −5.484 | 0.001 | 0.486 | 0.003 | 76.8 | ≥ 11 | 9 | 0.0 | 0.313 | 0.039 |
| TVFA | ≤ 12 | 12 | 35.5 | −1.883 | 0.008 | 0.132 | 0.048 | 99.9 | ≥ 11 | 19 | 46.7 | −0.424 | < 0.001 |
| Individual VFA molar proportion | |||||||||||||
| Acetate | ≤ 11 | 11 | 76.8 | −4.086 | 0.004 | 0.563 | 0.004 | 76.7 | ≥ 7 | 22 | 47.5 | 0.748 | < 0.001 |
| Propionate | ≤ 11 | 11 | 84.5 | 7.306 | < 0.001 | −0.898 | < 0.001 | 92.2 | ≥ 7 | 22 | 51.2 | −0.366 | 0.033 |
| Butyrate | ≤ 11 | 11 | 80.6 | −7.059 | < 0.001 | 0.732 | 0.001 | 92.3 | ≥ 7 | 20 | 54.0 | −0.582 | 0.002 |
| A:P | ≤ 11 | 10 | 78.6 | −6.737 | 0.001 | 0.884 | 0.002 | 90.8 | ≥ 7 | 10 | 59.2 | 0.915 | 0.016 |
| Total-tract fiber digestibility | |||||||||||||
| NDF | ≤ 23 | 15 | 55.7 | −4.458 | < 0.001 | 0.153 | 0.005 | 85.0 | |||||
| ADF | ≤ 23 | 11 | 69.8 | −6.276 | 0.001 | 0.213 | 0.030 | 45.1 | |||||
I2, percentage of heterogeneity across studies; Intercept, estimate of the effect size after defaunation; Coefficient, estimate of the change in effect size per week following defaunation; Adjusted R2, percentage of between-study variance explained by the defaunation duration; TVFA, total VFA
Ruminal VFA profiles and total-tract fiber digestibility dynamics during adaptation to defaunation
Consistent with the temporal dynamics of CH4 production after defaunation, the ruminal VFA profiles dynamics also included a linear phase (≤ 11 wk) and a plateau phase (≥ 7 wk) (Table 3). After defaunation, decreases in the acetate proportion, butyrate proportion and A:P (intercept = − 4.086, − 7.059 and − 6.737, respectively), and an increase in the propionate proportion (intercept = 7.306) were estimated (P < 0.01). These alterations decreased linearly (P < 0.01) at different rates over the first 11 wk of defaunation; instead, the ruminal acetate proportion (SMD = 0.748, P < 0.001) and A:P (SMD = 0.915, P = 0.016) were higher, and the propionate proportion (SMD = − 0.366, P = 0.033) was lower in defaunated animals than faunated animals during the plateau phase.
Compared with faunated animals, defaunated animals exhibited a reduced total VFA concentration (intercept = − 1.883 and P = 0.008), and the reduction decreased weekly by 0.132 (P = 0.048) until 12 wk (Table 3). The duration of defaunation could explain 99.9% of the between-study heterogeneity during the linear phase, although the heterogeneity (35.5%) was low. The decrease in total VFA concentration was still observed (SMD = − 0.424, P < 0.001) in defaunated animals during the plateau phase (≥ 11 wk).
Compared with that of faunation, the effect sizes of defaunation on total-tract fiber digestibility were linearly related to defaunation duration (Table 3): total-tract NDF and ADF digestibility (intercept = − 4.458 and − 6.276, respectively) were reduced (P < 0.01) after defaunation, and the reductions decreased weekly (P < 0.05) by 0.153 and 0.213, respectively, until 23 wk, which was the longest studied duration in the available data.
Discussion
Effect size and heterogeneity across all the studies
Based on all the selected studies, this meta-analysis showed that complete elimination of rumen protozoa generated adverse effects on the ruminal VFA concentration, butyrate proportion and dietary fiber digestibility; these findings were consistent with the results of previous meta-analyses [14, 18]. However, the heterogeneity across the studies was considerable for most of the responses to defaunation. Excess between-study variance increases the risk of incorrect average effect sizes when combining studies [15]. For example, the present meta-analysis based on all the studies showed that defaunation tended to increase the proportion of ruminal acetate but had no effect on the proportion of propionate. These findings were consistent with a recent meta-analysis by Newbold et al. [18] but inconsistent with that of Eugène et al. [14], who reported that defaunation induced a reduction in the ruminal acetate proportion and an increase in the propionate proportion. Therefore, the potential source of heterogeneity among the studies needs to be explored to better understand the responses to treatment, and this additional exploration is also one of most important tasks of meta-analysis [15].
The role of protozoa in rumen carbohydrate metabolism
Despite the fact that protozoa make up a large portion of the rumen biomass, their role in ruminal fermentation and their contribution to the metabolism and nutrition of the host are still topics of substantial controversy [2, 14], due to the difficulty of pure cultivation of protozoa in vitro. Rumen protozoa are not essential to the animal for survival, and defaunation has therefore been used to estimate the role of ciliate protozoa in rumen function. However, the adapted alteration of other microbe after defaunation may interfere with such estimations. Hence, the estimated intercept from the meta-regression analysis more accurately reflected the role that protozoa played in rumen fermentation. Reductions in the ruminal acetate proportion, butyrate proportion and CH4 production after complete removal of ruminal protozoa would be expected because protozoa ferment carbohydrates into acetate, butyrate and H2 [2], and the H2 produced is mostly converted to CH4 by methanogens situated inside protozoa or on their external surface [5–7]. A strong correlation between CH4 emissions and protozoa concentration has been reported [23], and protozoa-associated methanogens have been estimated to be responsible for 37% of CH4 production by ruminants [5]. Additionally, ruminal protozoa possess a full complement of hydrolytic enzymes for fermentation of the major components of feedstuffs, and certain ciliates present a wide range of fibrolytic enzyme genes, ingest small plant particles and use cell wall carbohydrates [18, 24]. Moreover, protozoa can indirectly contribute to ruminal degradation kinetics by maintaining a suitable rumen fermentation environment, for example, by scavenging oxygen to maintain anaerobiosis and slowing the rate of starch fermentation to maintain a proper ruminal pH [24], which favors the development and activity of bacteria and fungi [25, 26]. Therefore, the reductions in dietary fiber digestibility and ruminal total VFA concentration observed in this study would be expected after the complete removal of ruminal protozoa.
Temporal dynamics during adaptation to defaunation
Although protozoa are important ruminal H2 producers and exhibit interspecies H2 transfer with methanogens, we found that the effects of short- and long-term defaunation on CH4 production were opposite. This finding supports the conclusion of Morgavi et al. [13], who showed that there was not a simple cause-effect relationship between rumen protozoa and methanogenesis. Bird et al. [11] and Hegarty et al. [12] observed higher CH4 production (although not significantly so) in long-term defaunated ewes (11 and 26 wk) and lambs (12 to 33 wk) than in faunated animals. The significant increasing effect of long-term defaunation on CH4 production detected in this meta-analysis can be attributed to the pooled analysis, in which the number of replicate animals was increased by combining the results of relevant individual studies [15, 16]. The CH4 emissions from ruminants contribute to global greenhouse gas emissions and represent energy loss for the animals [8, 27, 28]. Therefore, the potential environmental protection and energy-saving values following defaunation were gradually lost and eventually became negative.
Acetate production during rumen fermentation is accompanied by reducing equivalents ([H]) production, whereas propionate production is accompanied by [H] consumption [29]; the excess [H] is converted to H2. The shift in the VFA profiles from propionate to acetate following defaunation increased the H2 available for methanogenesis, at least partially explaining the time-related changes in CH4 production observed in this study. When glucose is metabolized into acetate, propionate or butyrate, the energy efficiency relative to glucose for animal is 62%, 109% and 78%, respectively [3, 4]. Propionate fermentation is most energy efficient, due to assimilating energy from H2 and being the main precursor of gluconeogenesis in animals [3, 30]. In ruminants, the VFAs produced in the rumen satisfy up to 70% of energy requirements [30]. Shabat et al. [31] and Weimer et al. [32] observed that the ruminal total VFA concentration and propionate proportion were higher in highly efficient cows than in cows with low efficiency. Therefore, the decreases in the ruminal total VFA concentration and propionate proportion during the plateau phase also suggested that the elimination of rumen protozoa adversely affected the energy supply of animals in a long run.
The time-related variations in CH4, VFA profiles and dietary fiber digestibility implied a series of complex changes in the ruminal ecosystem over the course of defaunation. Nguyen et al. [33] reported that rumen microbes had likely not stabilized after 12 wk of defaunation, which agrees with our results showing that the linear phase for ruminal VFA profiles and CH4 emissions lasted 11 wk and 12 wk, respectively. When sudden major changes are made in the diet, it takes approximately 2 wk for the new microbial population balance to become established [34]; the much longer linear phase associated with defaunation suggests that protozoa play an important role in the ruminal ecosystem. Ruminal methanogens appear to develop more slowly than bacteria following defaunation [13]. Hristov et al. [28] noted that reductions in the population of protozoa-associated methanogens might be compensated by an increase in the population of bacteria- or rumen fluid-associated methanogens, and Mosoni et al. [26] found that long-term defaunation (2 yr) increased the abundance of methanogens. In addition, ruminal protozoa elimination results in increased bacterial abundance and changes in bacterial communities [35, 36]; defaunation has been shown to increase the anaerobic fungal population by two fold [37] and the Ruminococcaceae population by six fold [36]. Frumholtz [38] found that long-term defaunation (6 mo) increased the abundance of cellulolytic bacteria. Similar to protozoa, fungi and cellulolytic bacteria are also the main ruminal cellulolytic and H2-producing microbes that generate acetate, butyrate and/or H2 as primary end products [29, 39]. Therefore, it can be concluded that the increase in the populations of methanogens, fungi and cellulolytic bacteria following defaunation gradually counteracts the defaunation-induced reductions in dietary fiber digestibility, ruminal A:P and CH4 production, which may confirm an earlier theory of Weimer [40] indicating that the multiple microbial taxa in the ruminal community show functional redundancy (overlap of physiological function) and may therefore be substitutable with little impact on ecosystem processes [41, 42]. As noted by Taxis et al. [43] regarding the relationship between ruminal ecosystems and function: the players may change but the game remains. These observations also suggest that defaunation is not a good model for estimating the role of protozoa in rumen function due to the compensation effects of fungi and bacteria. Further animal experiments are required to fully understand the succession of rumen bacterial and archaeal community structure and function following defaunation, and the metabolic characteristics of rumen protozoa need be revealed using their genome and transcriptome data.
Conclusions
The present meta-analysis summarized the temporal dynamics of methanogenesis, ruminal fermentation and dietary fiber digestibility in ruminants after defaunation, and the results showed that defaunation adversely affected dietary fiber digestibility and the ruminal VFAs available to the host animals, although the effects were lessened over time. Furthermore, the energy advantages of defaunation gained by reducing CH4 production and shifting ruminal VFA profiles to more propionate were gradually lost over time, and the effects eventually became disadvantageous. Therefore, elimination of rumen protozoa adversely affects the energy supply of animals over the long-term.
Additional file
Figure S1. Forest plot showing the results of the subgroup meta-analysis of the anti-methanogenic effect size of defaunation, grouped by faunation state and duration of defaunation (11 wk). BF = born and reared protozoa free; AF = artificial defaunation; SMD = standardized mean difference; 95% CI = 95% confidence interval. * I-squared = percentage of heterogeneity across studies; P-value of SMD = 0. Figure S2. Funnel plot for the effect size of defaunation on CH4 production in (A) all studies, (B) short-term defaunation, (C) long-term defaunation, and (D) refaunation. The P-value of publication bias is presented. SMD = standardized mean difference, se = standard error. (DOCX 1373 kb)
Acknowledgments
The authors thank Prof. Chang Xu (Chinese Evidence-Based Medicine Center and Chinese Cochrane Center, West China Hospital, Sichuan University) for help with data analysis.
Funding
This work was supported by the National Key Research and Development Program of China (grant number: 2017YFD0500500).
Availability of data and materials
All the datasets were presented in the main manuscript (reference list in Table 1) and available to readers.
Abbreviations
- ADF
Acid detergent fiber
- AP
Acetate to propionate ratio
- NDF
Neutral detergent fiber
- SMD
Standardized mean difference
- VFA
Volatile fatty acids
Authors’ contributions
Conceived and designed the experiments: ZJL, FL and JHY. Performed the experiments: ZJL, QD and YFL. Analyzed the data: ZJL. Contributed to the writing of the manuscript: ZJL, TY and YCC. All authors reviewed and approved the manuscript.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Contributor Information
Zongjun Li, Email: lizongjun@nwsuaf.edu.cn.
Qi Deng, Email: dengqi17@mails.ucas.edu.cn.
Yangfan Liu, Email: liuyangfan1995@yeah.net.
Tao Yan, Email: 1468535134@qq.com.
Fei Li, Email: xnkiller@126.com.
Yangchun Cao, Email: caoyangchun@126.com.
Junhu Yao, Email: yaojunhu2004@sohu.com.
References
- 1.Ushida K., Jouany J.P., Demeyer D.I. Physiological Aspects of Digestion and Metabolism in Ruminants. 1991. Effects of Presence or Absence of Rumen Protozoa on the Efficiency of Utilization of Concentrate and Fibrous Feeds; pp. 625–654. [Google Scholar]
- 2.Williams AG, Coleman GS. The rumen protozoa. 1. New York: Springer-Verlag; 1992. [Google Scholar]
- 3.Millen DD, Arrigoni MDB, Pacheco RDL. Rumenology. 1. Springer: International Publishing; 2016. [Google Scholar]
- 4.Ryle M, Ørskov ER. Energy nutrition in ruminants. 1. Netherlands: Springer; 1990. [Google Scholar]
- 5.Finlay BJ, Esteban G, Clarke KJ, Williams AG, Embley TM, Hirt RP. Some rumen ciliates have endosymbiotic methanogens. FEMS Microbiol Lett. 1994;117:157–161. doi: 10.1111/j.1574-6968.1994.tb06758.x. [DOI] [PubMed] [Google Scholar]
- 6.Morgavi DP, Forano E, Martin C, Newbold CJ. Microbial ecosystem and methanogenesis in ruminants. Animal. 2010;4:1024–1036. doi: 10.1017/S1751731110000546. [DOI] [PubMed] [Google Scholar]
- 7.Belanche A, de la Fuente G, Newbold CJ. Study of methanogen communities associated with different rumen protozoal populations. FEMS Microbiol Ecol. 2014;90:663–677. doi: 10.1111/1574-6941.12423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Johnson KA, Johnson DE. Methane emissions from cattle. J Anim Sci. 1995;73:2483–2492. doi: 10.2527/1995.7382483x. [DOI] [PubMed] [Google Scholar]
- 9.Whitelaw FG, Eadie JM, Bruce LA, Shand WJ. Methane formation in faunated and ciliate-free cattle and its relationship with rumen volatile fatty acid proportions. Br J Nutr. 1984;52:261–275. doi: 10.1079/BJN19840094. [DOI] [PubMed] [Google Scholar]
- 10.Faichney GJ, Graham NM, Walker DM. Rumen characteristics, methane emissions, and digestion in weaned lambs reared in isolation. Aust J Agric Res. 1999;50:1083–1090. doi: 10.1071/AR98195. [DOI] [Google Scholar]
- 11.Bird SH, Hegarty RS, Woodgate R. Persistence of defaunation effects on digestion and methane production in ewes. Aust J Exp Agric. 2008;48:152–155. doi: 10.1071/EA07298. [DOI] [Google Scholar]
- 12.Hegarty RS, Bird SH, Vanselow BA, Woodgate R. Effects of the absence of protozoa from birth or from weaning on the growth and methane production of lambs. Br J Nutr. 2008;100:1220–1227. doi: 10.1017/S0007114508981435. [DOI] [PubMed] [Google Scholar]
- 13.Morgavi DP, Martin C, Jouany JP, Ranilla MJ. Rumen protozoa and methanogenesis: not a simple cause–effect relationship. Br J Nutr. 2012;107:388–397. doi: 10.1017/S0007114511002935. [DOI] [PubMed] [Google Scholar]
- 14.Eugène M, Archimède H, Sauvant D. Quantitative meta-analysis on the effects of defaunation of the rumen on growth, intake and digestion in ruminants. Livest Prod Sci. 2004;85:81–97. doi: 10.1016/S0301-6226(03)00117-9. [DOI] [Google Scholar]
- 15.Lean IJ, Rabiee AR, Duffield TF, Dohoo IR. Invited review: use of meta-analysis in animal health and reproduction: methods and applications. J Dairy Sci. 2009;92:3545–3565. doi: 10.3168/jds.2009-2140. [DOI] [PubMed] [Google Scholar]
- 16.Viechtbauer W. Learning from the past: refining the way we study treatments. J Clin Epidemiol. 2010;63:980–982. doi: 10.1016/j.jclinepi.2010.04.004. [DOI] [PubMed] [Google Scholar]
- 17.Newbold CJ, de la Fuente G, Belanche A, Ramos-Morales E, McEwan NR. The role of ciliate protozoa in the rumen. Front Microbiol. 2015;6:1313. doi: 10.3389/fmicb.2015.01313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sauer FD, Fellner V, Kinsman R, Kramer JK, Jackson HA, Lee AJ, et al. Methane output and lactation response in Holstein cattle with monensin or unsaturated fat added to the diet. J Anim Sci. 1998;76:906–914. doi: 10.2527/1998.763906x. [DOI] [PubMed] [Google Scholar]
- 19.Guan H, Wittenberg KM, Ominski KH, Krause DO. Efficacy of ionophores in cattle diets for mitigation of enteric methane. J Anim Sci. 2006;84:1896–1906. doi: 10.2527/jas.2005-652. [DOI] [PubMed] [Google Scholar]
- 20.Schönhusen U, Zitnan R, Kuhla S, Jentsch W, Derno M, Voigt J. Effects of protozoa on methane production in rumen and hindgut of calves around time of weaning. Arch Anim Nutr. 2003;57:279–295. doi: 10.1080/00039420310001594423. [DOI] [PubMed] [Google Scholar]
- 21.Higgins JPT, Thompson SG, Deeks JJ, Altman DG. Measuring inconsistency in meta-analyses. BMJ. 2003;327:557–560. doi: 10.1136/bmj.327.7414.557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Knapp G, Hartung J. Improved tests for a random effects meta-regression with a single covariate. Stat Med. 2003;22:2693–2710. doi: 10.1002/sim.1482. [DOI] [PubMed] [Google Scholar]
- 23.Guyader J, Eugène M, Nozière P, Morgavi DP, Doreau M, Martin C. Influence of rumen protozoa on methane emission in ruminants: a meta-analysis approach. Animal. 2014;8:1816–1825. doi: 10.1017/S1751731114001852. [DOI] [PubMed] [Google Scholar]
- 24.Nagaraja TG. Microbiology of the rumen. In: Millen DD, Arrigoni MDB, Pacheco RDL, editors. Rumenology. Cham: Springer; 2016. pp. 39–61. [Google Scholar]
- 25.Jouany JP, Demeyer DI, Grain J. Effect of defaunating the rumen. Anim Feed Sci Technol. 1988;21:229–265. doi: 10.1016/0377-8401(88)90105-8. [DOI] [Google Scholar]
- 26.Mosoni P, Martin C, Forano E, Morgavi DP. Long-term defaunation increases the abundance of cellulolytic ruminococci and methanogens but does not affect the bacterial and methanogen diversity in the rumen of sheep. J Anim Sci. 2011;89:783–791. doi: 10.2527/jas.2010-2947. [DOI] [PubMed] [Google Scholar]
- 27.Karakurt I, Aydin G, Aydiner K. Sources and mitigation of methane emissions by sectors: a critical review. Renew Energy. 2012;39:40–48. doi: 10.1016/j.renene.2011.09.006. [DOI] [Google Scholar]
- 28.Hristov AN, Oh J, Lee C, Meinen R. Mitigation of greenhouse gas emissions in livestock production: a review of technical options for non-CO2 emissions. Food and Agriculture Organization of the United Nations: Rome; 2013. [Google Scholar]
- 29.Janssen PH. Influence of hydrogen on rumen methane formation and fermentation balances through microbial growth kinetics and fermentation thermodynamics. Anim Feed Sci Technol. 2010;160:1–22. doi: 10.1016/j.anifeedsci.2010.07.002. [DOI] [Google Scholar]
- 30.Bergman EN. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol Rev. 1990;70:567–590. doi: 10.1152/physrev.1990.70.2.567. [DOI] [PubMed] [Google Scholar]
- 31.Shabat Sheerli Kruger Ben, Sasson Goor, Doron-Faigenboim Adi, Durman Thomer, Yaacoby Shamay, Berg Miller Margret E, White Bryan A, Shterzer Naama, Mizrahi Itzhak. Specific microbiome-dependent mechanisms underlie the energy harvest efficiency of ruminants. The ISME Journal. 2016;10(12):2958–2972. doi: 10.1038/ismej.2016.62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Weimer PJ, Cox MS, de Paula TV, Lin M, Hall MB, Suen G. Transient changes in milk production efficiency and bacterial community composition resulting from near-total exchange of ruminal contents between high- and low-efficiency Holstein cows. J Dairy Sci. 2017;100:7165–7182. doi: 10.3168/jds.2017-12746. [DOI] [PubMed] [Google Scholar]
- 33.Nguyen SH, Bremner G, Cameron M, Hegarty RS. Methane emissions, ruminal characteristics and nitrogen utilisation changes after refaunation of protozoa-free sheep. Small Rumin Res. 2016;144:48–55. doi: 10.1016/j.smallrumres.2016.08.002. [DOI] [Google Scholar]
- 34.Reece WO. Dukes’ physiology of domestic animals. 12. Comstock Pub. Associates: Ithaca; 2004. [Google Scholar]
- 35.Ozutsumi Y, Tajima K, Takenaka A, Itabashi H. The effect of protozoa on the composition of rumen bacteria in cattle using 16S rRNA gene clone libraries. Biosci Biotechnol Biochem. 2005;69:499–506. doi: 10.1271/bbb.69.499. [DOI] [PubMed] [Google Scholar]
- 36.Morgavi DP, Rathahao-Paris E, Popova M, Boccard J, Nielsen KF, Boudra H. Rumen microbial communities influence metabolic phenotypes in lambs. Front Microbiol. 2015;6:1060. doi: 10.3389/fmicb.2015.01060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Newbold CJ, Hillman K. The effect of ciliate protozoa on the turnover of bacterial and fungal protien in the rumen of sheep. Lett Appl Microbiol. 1990;11:100–102. doi: 10.1111/j.1472-765X.1990.tb01286.x. [DOI] [Google Scholar]
- 38.Frumholtz PP. Manipulation of the rumen fermentation and its effects on digestive physiology: University of Aberdeen; 1991.
- 39.Morvan B, Rieu-Lesme F, Fonty G, Gouet P. In vitro interactions between rumen H2-producing cellulolytic microorganisms and H2-utilizing acetogenic and sulfate-reducing bacteria. Anaerobe. 1996;2:175–180. doi: 10.1006/anae.1996.0023. [DOI] [Google Scholar]
- 40.Weimer PJ. Redundancy, resilience, and host specificity of the ruminal microbiota: implications for engineering improved ruminal fermentations. Front Microbiol. 2015;6:296. doi: 10.3389/fmicb.2015.00296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lawton JH, Brown VK. Redundancy in ecosystems. In: Schulze ED, Mooney HA, editors. Biodiversity and ecosystem function. Berlin: Springer; 1994. pp. 255–270. [Google Scholar]
- 42.Rosenfeld JS. Functional redundancy in ecology and conservation. Oikos. 2002;98:156–162. doi: 10.1034/j.1600-0706.2002.980116.x. [DOI] [Google Scholar]
- 43.Taxis TM, Wolff S, Gregg SJ, Minton NO, Zhang C, Dai J, et al. The players may change but the game remains: network analyses of ruminal microbiomes suggest taxonomic differences mask functional similarity. Nucleic Acids Res. 2015. 10.1093/nar/gkv973. [DOI] [PMC free article] [PubMed]
- 44.Belanche A, Abecia L, Holtrop G, Guada JA, Castrillo C, de la Fuente G, et al. Study of the effect of presence or absence of protozoa on rumen fermentation and microbial protein contribution to the chyme. J Anim Sci. 2011;89:4163–4174. doi: 10.2527/jas.2010-3703. [DOI] [PubMed] [Google Scholar]
- 45.Chandramoni JSB, Tiwari CM, Haque N, Murarilal KMY. Energy metabolism and methane production in faunated and defaunated sheep fed two diets with same concentrate to roughage ratio (70:30) but varying in composition. Asian-Australas J Anim Sci. 2001;14:1238–1244. doi: 10.5713/ajas.2001.1238. [DOI] [Google Scholar]
- 46.Chaudhary LC, Srivastava A. Performance of growing Murrah buffalo calves as affected by treatment with Manoxol and the presence of ciliate protozoa in the rumen. Anim Feed Sci Technol. 1995;51:281–286. doi: 10.1016/0377-8401(94)00689-7. [DOI] [Google Scholar]
- 47.Eadie JM, Gill JC. The effect of the absence of rumen ciliate protozoa on growing lambs fed on a roughage–concentrate diet. Br J Nutr. 1971;26:155–167. doi: 10.1079/BJN19710022. [DOI] [PubMed] [Google Scholar]
- 48.Eugène M, Sauvant D, Weisbecker JL, Archimède H. Effects of defaunation on digestion of fresh Digitaria decumbens grass and growth of lambs. Animal. 2010;4:439–445. doi: 10.1017/S1751731109991194. [DOI] [PubMed] [Google Scholar]
- 49.Kasuya N, Wada I, Shimada M, Kawai H, Itabashi H. Effect of presence of rumen protozoa on degradation of cell wall constituents in gastrointestinal tract of cattle. Anim Sci J. 2007;78:275–280. doi: 10.1111/j.1740-0929.2007.00435.x. [DOI] [Google Scholar]
- 50.Kreuzer M, Kirchgessner M, Müller HL. Effect of defaunation on the loss of energy in wethers fed different quantities of cellulose and normal or steamflaked maize starch. Anim Feed Sci Technol. 1986;16:233–241. doi: 10.1016/0377-8401(86)90114-8. [DOI] [Google Scholar]
- 51.Nagaraja TG, Godfrey SI, Winslow SW, Rowe JB, Kemp KE. Effect of virginiamycin on ruminal fermentation in faunated or ciliate-free sheep overfed with barley grain. Small Rumin Res. 1995;17:1–8. doi: 10.1016/0921-4488(95)00647-4. [DOI] [Google Scholar]
- 52.Santra A, Karim SA. Growth performance of faunated and defaunated Malpura weaner lambs. Anim Feed Sci Technol. 2000;86:251–260. doi: 10.1016/S0377-8401(00)00161-9. [DOI] [Google Scholar]
- 53.Santra A, Karim SA. Nutrient utilization and growth performance of defaunated and faunated lambs maintained on complete diets containing varying proportion of roughage and concentrate. Anim Feed Sci Technol. 2002;101:87–99. doi: 10.1016/S0377-8401(02)00146-3. [DOI] [Google Scholar]
- 54.Santra A, Karim SA, Chaturvedi OH. Rumen enzyme profile and fermentation characteristics in sheep as affected by treatment with sodium lauryl sulfate as defaunating agent and presence of ciliate protozoa. Small Rumin Res. 2007;67:126–137. doi: 10.1016/j.smallrumres.2005.08.028. [DOI] [Google Scholar]
- 55.Sultana H, Miyazawa K, Kanda S, Itabashi H. Fatty acid composition of ruminal bacteria and protozoa, and effect of defaunation on fatty acid profile in the rumen with special reference to conjugated linoleic acid in cattle. Anim Sci J. 2011;82:434–440. doi: 10.1111/j.1740-0929.2010.00854.x. [DOI] [PubMed] [Google Scholar]
- 56.Williams PP, Dinusson WE. Ruminal volatile fatty acid concentrations and weight gains of calves reared with and without ruminal ciliated protozoa. J Anim Sci. 1973;36:588–591. doi: 10.2527/jas1973.363588x. [DOI] [PubMed] [Google Scholar]
- 57.Yáñez-Ruiz DR, Williams S, Newbold CJ. The effect of absence of protozoa on rumen biohydrogenation and the fatty acid composition of lamb muscle. Br J Nutr. 2007;97:938–948. doi: 10.1017/S0007114507675187. [DOI] [PubMed] [Google Scholar]
- 58.Zhou YY, Mao HL, Jiang F, Wang JK, Liu JX, McSweeney CS. Inhibition of rumen methanogenesis by tea saponins with reference to fermentation pattern and microbial communities in Hu sheep. Anim Feed Sci Technol. 2011;166–167:93–100. doi: 10.1016/j.anifeedsci.2011.04.007. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. Forest plot showing the results of the subgroup meta-analysis of the anti-methanogenic effect size of defaunation, grouped by faunation state and duration of defaunation (11 wk). BF = born and reared protozoa free; AF = artificial defaunation; SMD = standardized mean difference; 95% CI = 95% confidence interval. * I-squared = percentage of heterogeneity across studies; P-value of SMD = 0. Figure S2. Funnel plot for the effect size of defaunation on CH4 production in (A) all studies, (B) short-term defaunation, (C) long-term defaunation, and (D) refaunation. The P-value of publication bias is presented. SMD = standardized mean difference, se = standard error. (DOCX 1373 kb)
Data Availability Statement
All the datasets were presented in the main manuscript (reference list in Table 1) and available to readers.