

Abstract
Natural language descriptions of organismal phenotypes, a principal object of study in biology, are abundant in the biological literature. Expressing these phenotypes as logical statements using ontologies would enable large-scale analysis on phenotypic information from diverse systems. However, considerable human effort is required to make these phenotype descriptions amenable to machine reasoning. Natural language processing tools have been developed to facilitate this task, and the training and evaluation of these tools depend on the availability of high quality, manually annotated gold standard data sets. We describe the development of an expert-curated gold standard data set of annotated phenotypes for evolutionary biology. The gold standard was developed for the curation of complex comparative phenotypes for the Phenoscape project. It was created by consensus among three curators and consists of entity–quality expressions of varying complexity. We use the gold standard to evaluate annotations created by human curators and those generated by the Semantic CharaParser tool. Using four annotation accuracy metrics that can account for any level of relationship between terms from two phenotype annotations, we found that machine–human consistency, or similarity, was significantly lower than inter-curator (human–human) consistency. Surprisingly, allowing curatorsaccess to external information did not significantly increase the similarity of their annotations to the gold standard or have a significant effect on inter-curator consistency. We found that the similarity of machine annotations to the gold standard increased after new relevant ontology terms had been added. Evaluation by the original authors of the character descriptions indicated that the gold standard annotations came closer to representing their intended meaning than did either the curator or machine annotations. These findings point toward ways to better design software to augment human curators and the use of the gold standard corpus will allow training and assessment of new tools to improve phenotype annotation accuracy at scale.
Introduction
Phenotype descriptions of organisms are documented across nearly all areas of biological research including biomedicine, evolution, developmental biology and paleobiology. The vast majority of such descriptions are expressed in the scientific literature using natural language. While allowing for rich semantics, natural language descriptions can be difficult for nonexperts to understand, are opaque to machine reasoning and thus hinder the integration of phenotypic information across different studies, taxonomic systems and branches of biology (1).
To make phenotype descriptions more amenable to computation, model organism databases employ human curators to convert natural language phenotype descriptions into machine-readable phenotype annotations that use standard ontologies (e.g. 2–5). One format used for phenotype annotations is the ontology-based entity (E)–quality (Q) (EQ) representation, in which an entity represents a biological object such as an anatomical structure, space, behavior or a biological process; a Q represents a trait or property that an E possesses, e.g. shape, color or size; an optional related E (RE) allows for binary relations such as adjacency (6, 7). Among formal representations of phenotype descriptions, EQ is the most widely used, e.g. (8), although other formal representations have been proposed (9–11). Further, to create entities and qualities that adequately represent the often highly detailed phenotype descriptions, curators create complex logical expressions called ‘post-compositions’ by combining ontology terms, relations and spatial properties in different ways. In contrast to EQ expressions with single-term Es and Qs, creating post-composed entities and qualities (Table 1) can be a complex task, due to the flexibility in logic expression and the different semantic interpretations that free-text descriptions often allow. Additionally, the varied ways in which concepts from multiple ontologies can be combined to create post-composed expressions result in a vast set of possible EQ combinations where consistency is difficult to achieve. As a result, it can be expected that EQ annotations involving post-compositions will show variability between different curators.
Table 1.
Examples of EQ annotations of varying complexity from the present study
| Character: state | E | Q | RE |
|---|---|---|---|
| A sclerotic ossicles: greatly enlarged | Uberon: ‘scleral ossicle’ | PATO: ‘increased size’ | |
| B nasal-prefrontal contact: present | Uberon: ‘nasal bone’ | PATO: ‘in contact with’ | Uberon: ‘prefrontal bone’ |
| C lateral pelvic glands: absent in males | Uberon: ‘gland’ and (‘part_of’ some {BSPO: ‘lateral region’ and [‘part_of’ some Uberon: ‘pelvis’ and (‘part_of’ some Uberon: ‘male organism’)]}) | PATO: ‘absent’ |
‘A’ illustrates a simple EQ annotation; ‘B’ shows an EQ annotation in which the Q term relates two entities to each other; and ‘C’ provides an example of an E that does not correspond to a term in an existing ontology but is instead a complex logical expression post-composed from multiple ontology terms
To best resolve the ambiguities inherent in natural language descriptions, human curators will often not only use their domain expertise but also refer to external information for deducing the original author’s intent. Phenotype descriptions found in the literature, however, are typically in a concise format with little or no contextualizing information that would help with disambiguating the intended meaning. The difficulty of disambiguation can be exacerbated when the requisite E and Q domain ontologies do not yet include an obviously appropriate term for a particular annotation (12). As a consequence of this and other challenges, manual curation tends to be extremely labor-intensive and few projects have the resources to comprehensively curate the relevant literature. To help address this bottleneck, text mining and natural language processing (NLP) systems have been developed with the goal of supplementing or augmenting the work of human curators. Facilitating continuous improvement of these systems, tools and algorithms requires means to compare different systems objectively and fairly with each other and with human curators, in particular, with respect to accuracy of generated annotations. This raises several questions. One, what is the reference against which accuracy is best assessed if annotations generated for a given task show variability between different human curators? Two, how consistent is the result of machine annotation with that of a human curator? Three, to what extent is machine annotation performance limited by inherent differences between how a machine and a human expert execute a curation task? In particular, in contrast to human curators who will consult external information, a software tool will normally only use the vocabulary and domain knowledge it is initially provided with in the form of input lexicons and ontologies.
The variability among expert curators can be used to provide a baseline for the performance evaluation of automated systems. Cui et al. (13) conducted an inter-curator consistency experiment to evaluate Semantic CharaParser (SCP), an NLP tool designed for generating EQ annotations from character descriptions in the comparative anatomy literature [specifically, from phylogenetic character matrices (14)]. Characters consist of two or more character states contrasting the variation in phenotype among a set of taxa. Character-by-taxon matrices are used in phylogenetic and comparative analyses to infer the evolutionary relationships among the taxa under study and to reconstruct putative character state evolution on the phylogeny.
To our knowledge, SCP is the first semi-automatic software designed to generate EQ annotations. SCP works by parsing the original character descriptions to identify E and Q terms, matching these terms to ontology concepts and generating logical relations and, where appropriate, post-compositions from the matched concepts based on a set of rules. In the experiment, three curators independently annotated a set of 203 characters, randomly chosen from seven publications representing extant and extinct vertebrates for a variety of anatomical systems with an emphasis on skeletal anatomy, corresponding to the curators’ domain of expertise (Table 2). In the first, or ‘Naïve’, round of annotation, curators were not allowed access to sources of knowledge external to the character description, including the publication from which the matrix originated. In the second, or ‘Knowledge’, round curators were allowed to access external sources of knowledge, such as the full publication from which the character was drawn, related literature and other online sources. The curators were given a set of initial ontologies to use for curation. The new ontology terms created during curation were added to the ‘Initial’ ontologies to create curator-specific ‘Augmented’ ontologies. At the end of the curation rounds, all curator-specific augmented ontologies were merged to create a final ‘Merged’ ontology.
Table 2.
Phylogenetic studies from which characters were selected
| Reference | Taxonomic group | No. of taxa | No. of characters |
|---|---|---|---|
| Hill (31) | Amniotes | 80 | 365 |
| Skutschas and Gubin (32) | Amphibians | 22 | 69 |
| Nesbitt et al. (33) | Birds | 22 | 107 |
| Coates and Sequeira (30) | Cartilaginous fishes | 23 | 86 |
| Chakrabarty (34) | Cichlid fishes | 41 | 89 |
| O’Leary et al. (35) | Mammals | 84 | 4541 |
| Conrad (36) | Squamate reptiles | 223 | 363 |
The Cui et al. (13) study was designed such that SCP was used to annotate the same set of characters as human curators using three sets of ontologies (Initial, Augmented and Merged) with progressively more comprehensive coverage, as described below. The primary findings were as follows. The performance of SCP was significantly lower as compared to human curators. When comparing the performance of SCP to human curators, no statistically significant differences were found between Naïve and Knowledge rounds. Inter-curator recall and precision were also not found to be significantly different between the Naïve and Knowledge rounds. SCP performed significantly better with Augmented versus Initial ontologies. However, there was no significant difference in performance between Augmented and Merged ontologies.
While useful, there were several limitations in the Cui et al. (13) evaluation of SCP, including the lack of a gold standard against which to measure its performance. Manually annotated gold standard data sets are high-quality benchmarks for both evaluation and training of automated NLP systems, e.g. (15–17). Another limitation was the use of performance measures that did not fully account for the continuum of similarity possible between semantic phenotype annotations. While these authors recognized that phenotypes annotated with parent and daughter terms in the ontology bear some partial resemblance, here we introduce semantic similarity measures that can account for any level of relationship between the terms from two phenotype annotations.
The present work describes the development of an expert-curated gold standard data set of annotated phenotypes for evolutionary biology that is the best available given current constraints in semantic representation. The gold standard was developed for the annotation of the complex evolutionary phenotypes described in the systematics literature for the Phenoscape project (14, 18). Unlike many published gold standards for ontology annotation, which frequently focus on E recognition, e.g. (19), the Phenoscape gold standard consists of EQ expressions of varying complexity. We evaluate how well the annotations of individual curators and the machine (SCP) compare to those of the gold standard, using four ontology-aware metrics. Two of these are traditional measures of semantic similarity (20) and two are extensions of precision and recall that account for partial semantic similarity. In addition, we directly assessed the quality of the gold standard with an author survey, in which the original domain experts were invited to rank the accuracy of a subset of the annotations from the gold standard, the individual human curators and SCP.
Related work
Gold standard corpora are collections of articles manually annotated by expert curators and they provide a high-quality comparison against which to test automated text processing systems. Funk et al. (17), for example, used the CRAFT annotation corpus (19, 21) for the evaluation of three concept annotation systems. Within the biomedical sciences, a number of gold standard corpora have been developed (22–24) and these focus on concept recognition. Concepts are annotated at the text-string level, e.g. (19), or, in some cases, annotations are attached at the whole-document level, e.g. (23). Because of the effort and costs required for manual annotation, ‘silver standard’ corpora have also been created, in which automatically generated annotations are grouped into a single corpus (25, 26). As far as we are aware, there are no published gold standard corpora for EQ phenotypes and none for evolutionary phenotypes.
Inter-curator consistency has been used by several studies as a baseline against which to evaluate the performance of automated curation software (27–29). Wiegers et al. (27) measured the performance of text mining software that identifies chemical–gene interactions from the literature by comparing the output against inter-curator consistency on the same task. Sohngen et al. (28) evaluated the performance of the DRENDA text-mining systemthat retrieves enzyme-related information on diseases. Most similar to the work reported here is the study by Camon et al. (29) in which inter-curator consistency was used as a baseline to evaluate performance of text-mining systems to retrieve Gene Ontology (GO) terms from the literature. In their experiment, three curators co-curated 30 papers and extracted GO terms from the text. In inter-curator comparisons, GO term pairs were classified into three categories: exact matches, same lineage (terms related via subsumption relationships) and different lineage (unrelated terms). They found that curators chose exactly the same terms (39%), related terms (43%) and unrelated terms (19%) of the time. Our approach differs in that we evaluate inter-curator consistency at the task of phenotype (EQ) annotation and we employ metrics that can account for partial matches between annotations by taking advantage of both the ontology structure and the information content (IC) from annotation frequencies.
Methods
Source of phenotypes
Twenty-nine characters were randomly selected from each of seven published phylogenetic studies, yielding 203 characters and 463 character states in total (Table 2). The studies were chosen to (i) have a wide taxonomic breadth across vertebrates, (ii) include both extinct and extant taxa and (iii) include characters from several anatomical systems (e.g. skeletal, muscular and nervous systems). These objectives were intended to reduce potential sources of systematic bias. For example, the prevailing style of character descriptions can differ depending on the taxonomic group of interest. Further, the curators had varying expertise across the vertebrate taxa. The characters and character states presented to curators were extracted directly from the character list in each publication [e.g. ‘Pelvic plate semicircular with anterolateral concavity. Absent (0); present (1)’ from character 39 in Coates and Sequeira (30)]. Thus, curators had access to the full-character and state descriptions for each of the selected characters, in addition to taxonomic scope and publication source, but they—and the SCP developers—were blind to the choice of papers and the selection of characters prior to the experiment.
Experimental design
The common set of character states was annotated independently by three curators (W. Dahdul, T. A. Dececchi and N. Ibrahim) and by SCP. The curators were randomly assigned identifiers C1, C2 and C3 at the beginning of the study. Curators used Phenex software (12, 37) for manually generating annotations. The annotations are complex expressions made up of E, Q and, where required, an RE. The E and RE components employ Uberon (38, 39) concepts and may be post-composed with terms from multiple ontologies including Uberon, Phenotype and Trait Ontology [PATO; (40, 41)] and the Biological Spatial Ontology (BSPO) (42) while the Q component uses PATO concepts. Curators were free to create one or multiple EQ annotations per state, and they were encouraged to annotate at a fine level of detail (43). To measure the effect of external knowledge on inter-curator consistency, two rounds of human curation were performed. In the first (‘Naïve’) round, the character and character state text were the only information the curators were allowed to consult. Accessing the source publication or any external information was not permitted. This was intended to simulate the extent of information available to SCP, although curators naturally use their subject-domain expertise when composing annotations. In the second (‘Knowledge’) round, the curators annotated the same set of characters as in the Naïve round, but they were free to consult the full text of the source publication and to access any other external information. In total, this resulted in six sets of human-curated EQ annotations and six augmented ontologies produced by the curators independently during the Naïve and Knowledge rounds.
Several steps were taken to promote consistency among the human curators and between curators and SCP. First, curators developed and were trained on a set of curation guidelines for the annotation of phylogenetic characters [the Phenoscape Guide to Character Annotation (44)]. These guidelines were also made available to SCP developers and are the basis of rules according to which SCP generates EQ expressions. Second, curators took advantage of an interactive consistency review panel available in Phenex, which reports missing or problematic annotations, such as a relational Q used to annotate a character state without also specifying an RE. Further, each curator had at least 1 year of experience with EQ annotation prior to the experiment. Note that each curator still performed their curation tasks in the experiment independently from each other, and thus there was still room for variation. For instance, for a given character state, one curator might choose to use an imperfectly matching E term, while another might aim for a more precise representation by post-composing a new term from existing terms, and yet another might choose to add a new single term to their Initial ontology. To avoid advantaging SCP beyond an initial training data set, SCP developers were not allowed to observe the human curation process during the experiment.
The gold standard
The gold standard corpus, which consists of a unique set of EQ annotations for each character state in the 203 character data set, was created as a consensus data set by the three curators. After completing the Knowledge round, the curators reviewed and discussed all the EQs in their three separate Knowledge round curator data sets for the purpose of developing a single gold standard data set. In assembling this set of EQ annotations for the gold standard, the curators were not limited to choosing among the individual EQs that they had created during the experiment; instead, they were free to modify existing annotations or create entirely new ones. In cases where there was insufficient information to resolve ambiguities, the curators consulted additional published literature and other online resources. In some cases, they also contacted domain experts to clarify terminology or anatomy. Once all three curators were in agreement, they used the Phenex curation software to create the gold standard EQ annotations for the final gold standard data set.
In the course of developing the gold standard, the curators updated the best practices for EQ annotation of characters documented in the Phenoscape Guide to Character Annotation (44). We updated the list of commonly encountered character categories (e.g. presence/absence, position, size) with new categories, examples and EQ conventions. Each phenotype in the gold standard references one or more of the character categories from the guide.
Ontologies
The human curators and SCP were provided with the same initial set of ontologies: Uberon [version phenoscape-ext/15 March 2013, (38, 39)], BSPO [release 17 May 2018, (42)] and PATO [release 03 June 2013, (41)].
In both the Naïve and Knowledge rounds, each curator was free to provisionally add terms that they deemed missing from any of the Initial ontologies, resulting in Augmented ontologies that differed from their Initial versions. New term requests were added as provisional terms by using the ontology request broker in Phenex (12), which provides an interface to the BioPortal’s provisional term API (45). Ontology curators can subsequently resolve these requests as mistakenly overlooked existing terms, new synonyms to existing terms or bona fide new terms. At the end of the experiment, there were six sets of Augmented ontologies, one from each curator in each round (Table 3). These were subsequently combined to produce a Merged set of ontologies for which redundant classes were manually reconciled. To test the effect of ontology coverage on automated EQ annotation, SCP was run with the Initial ontology, the Augmented ontologies and the final Merged ontology. The results in each case were compared to those obtained by the human curators, as reported in Cui et al. (13).
Table 3.
Augmentation of E (UBERON), Q (PATO) and spatial (BSPO) ontologies by the three curators in both rounds of curation (Naïve and Knowledge) and in the final Merged ontology
| Curation | Human | Terms added to | ||
|---|---|---|---|---|
| round | curator | UBERON | PATO | BSPO |
| Naïve | C1 | 109 | 70 | 3 |
| C2 | 49 | 32 | 0 | |
| C3 | 89 | 23 | 2 | |
| Knowledge | C1 | 129 | 74 | 3 |
| C2 | 72 | 52 | 0 | |
| C3 | 108 | 35 | 3 | |
| Merged | 199 | 127 | 7 | |
Measuring similarity between annotation sources
When different ontology terms are chosen to annotate a given character state, the selected terms may nonetheless be semantically similar. Thus, it is desirable to use measures of annotation similarity that allow for varying degrees of relatedness using the background ontology and annotation corpus (20). Here, we use four measures, two of which are semantic similarity metrics with a history of usage in the literature and two of which are modifications of the traditional measures of precision and recall that account for different but semantically similar annotations. All four measures can be applied to both full EQ annotations and to comparisons among E terms alone.
Semantic similarity measures between annotation sources (e.g. different curators) were aggregated at the level of the individual character state and across all character states (Figure 1). Aggregation of pairwise (EQ to EQ) annotations by character state is necessary because a curator may generate more than one EQ annotation for a given character state. This is illustrated by Figure 1 where Curator A generated three EQs and Curator B generated two EQs for State  . To measure the overall similarity between two annotation sources (e.g. Curator A to Curator B in Figure 1, top), we first compute a similarity score between corresponding character state pairs as the best match (maximum score) among all pairwise comparisons between EQs for the same character state (Maximum Character State Similarity in Figure 1). We then compute the similarity between two annotation sources by taking the arithmetic mean of the pairwise character state similarity scores across all character state pairs (Mean Curator Similarity in Figure 1, bottom).
. To measure the overall similarity between two annotation sources (e.g. Curator A to Curator B in Figure 1, top), we first compute a similarity score between corresponding character state pairs as the best match (maximum score) among all pairwise comparisons between EQs for the same character state (Maximum Character State Similarity in Figure 1). We then compute the similarity between two annotation sources by taking the arithmetic mean of the pairwise character state similarity scores across all character state pairs (Mean Curator Similarity in Figure 1, bottom).
Figure 1.
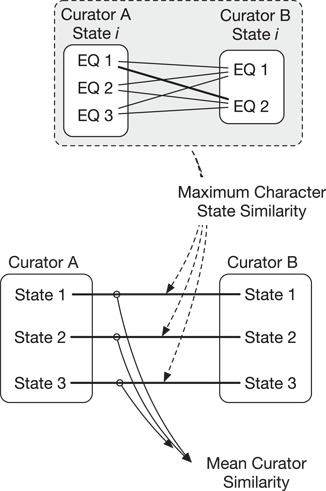
Similarity of annotations between two curators is calculated across multiple character states (e.g. states 1–3, bottom). First, the maximum character state similarity is calculated at the level of a single character state and is the best match (maximum score) in pairwise comparisons across that state’s EQ annotations. Mean curator similarity is then calculated as the mean of the maximum similarities across all character state pairs.
Generating subsumers for EQ annotations
We treat each EQ annotation as a node in an ad hoc EQ ontology. Creating the complete cross product of the component ontologies would necessarily include all possible subsumers but would be prohibitive. As a memory-saving measure, we developed a computationally efficient approach to identify subsumers for EQ annotations on an ad hoc basis, as follows.
A comprehensive ontology was created by taking the union of Uberon, PATO and BSPO ontologies using the ‘-merge-support-ontologies’ command in the owltools software (https://github.com/owlcollab/owltools). In order to enable reasoning on additional dimensions (e.g. ‘part of’) in post-compositions while identifying subsumers, we added additional classes to the comprehensive ontology. For every concept ‘U’ in the Uberon ontology and every object property ‘OP’ used in post-compositions, a class of the form ‘OP some U’ was added to the comprehensive ontology.
First, every EQ annotation is split into individual E, Q and, optionally, RE components (Figure 2, Step 1). Simultaneously, the EQ annotation is transformed into a Web Ontology Language (OWL) class expression of the form ‘Q and inheres in some E and towards some RE’ (Figure 2, Step 1). Next, superclasses of these individual components and the class expression are retrieved using the ELK reasoner on the comprehensive ontology (Figure 2, Step 2). Individual E, Q and RE superclasses are combined to create superclasses of the form E–Q–RE. The combined class expression and combinatorial E–Q–RE superclasses form the subsumers of an EQ annotation (Figure 2, Step 3). While it is possible that additional subsumers could be found in the case that a class in another part of the hierarchy has a logical definition that matches an EQ expression, it is unlikely for these ontologies because subsuming Q terms in the PATO ontology do not have logical definitions that make use of Uberon entities.
Figure 2.
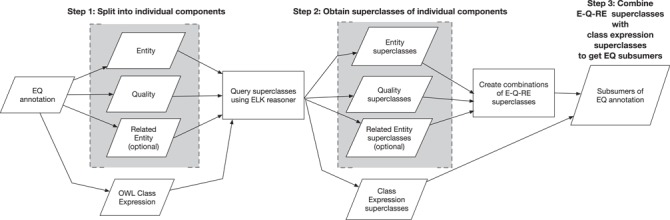
EQ annotations are split into E, Q and RE components and also transformed into an OWL class expression. Superclasses of E, Q and RE and the class expression are queried via ELK. E, Q and RE superclasses are combined in the form E–Q–RE. These E–Q–RE superclasses along with the class expression’s superclasses form the subsumers of the EQ annotation for computation of semantic similarity.
Jaccard similarity
The Jaccard similarity (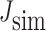 ) between nodes
) between nodes 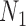 and
and 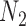 in an ontology graph is defined as the ratio of the number of nodes in the intersection of their subsumers over the number of nodes in the union of their subsumers (46):
in an ontology graph is defined as the ratio of the number of nodes in the intersection of their subsumers over the number of nodes in the union of their subsumers (46):
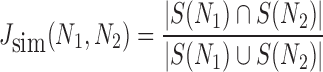 |
where 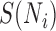 is the set of nodes that subsume
is the set of nodes that subsume 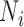 .
. 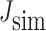 measures the distance between two EQs based on the class structure of the ontology. The range of
measures the distance between two EQs based on the class structure of the ontology. The range of  .
. 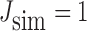 when the two EQs being compared are the same and
when the two EQs being compared are the same and 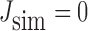 when they have no common subsumers.
when they have no common subsumers.
Information content
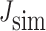 measures the ontology graph distance between two nodes and thus necessarily ignores differences in semantic specificity between parent and child terms in different areas of the ontology graph. ‘IC’ is used to capture the specificity of the annotations. The IC
measures the ontology graph distance between two nodes and thus necessarily ignores differences in semantic specificity between parent and child terms in different areas of the ontology graph. ‘IC’ is used to capture the specificity of the annotations. The IC  of a node
of a node 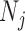 in an ontology is defined as the proportion of annotations to
in an ontology is defined as the proportion of annotations to 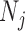 and all nodes subsumed by
and all nodes subsumed by 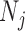 in an annotation corpus (47). Let
in an annotation corpus (47). Let 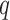 be the number of nodes in the ontology. Define
be the number of nodes in the ontology. Define 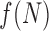 to be the number of annotations directly to
to be the number of annotations directly to 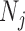 and
and 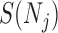 to be the set of nodes subsumed by
to be the set of nodes subsumed by 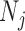 :
:
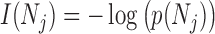 |
where
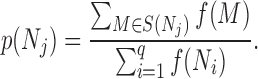 |
The 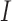 of two nodes is defined as the
of two nodes is defined as the 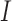 of the least common subsumer (LCS) of the two nodes. If there are multiple LCSs, the node with the highest
of the least common subsumer (LCS) of the two nodes. If there are multiple LCSs, the node with the highest 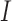 is used (46).
is used (46). 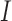 has a minimum of zero at the root and a maximum that is dependent on the size of the corpus
has a minimum of zero at the root and a maximum that is dependent on the size of the corpus
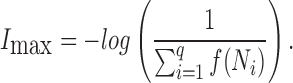 |
To obtain a normalized score 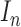 in the range of
in the range of 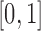 , we use
, we use 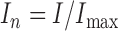 . In our analysis, the corpus for measurement of
. In our analysis, the corpus for measurement of 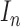 includes all human annotations from both annotation rounds and the annotations from SCP.
includes all human annotations from both annotation rounds and the annotations from SCP.
Partial precision and partial recall
Precision and recall are commonly used to evaluate the performance of information retrieval systems. Traditionally, these two measures do not attempt to account for imperfect matches; information is either retrieved or not. For ontology-based annotations, partial information retrieval is possible because the information to be retrieved is the semantics of the annotated text, rather than a particular term. To account for this, here we use two metrics, partial precision (‘PP’) and partial recall (‘PR’), to measure the success of semantic information retrieval by a test curator (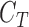 ) relative to a reference curator (
) relative to a reference curator (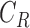 ), where a curator can be understood as either human or software. While other variants of semantic precision and recall are used in the literature (48, 49), the measures we use here specifically use semantic similarity, in this case
), where a curator can be understood as either human or software. While other variants of semantic precision and recall are used in the literature (48, 49), the measures we use here specifically use semantic similarity, in this case 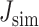 , to quantify partial matches between annotations. In contrast to our approach, (48) and (49) compute semantic precision and recall by examining the superclass sets of two annotations. Depending on the overlap among these sets, each superclass is classified as a true positive, false positive or false negative. These counts are then used to compute semantic precision and recall.
, to quantify partial matches between annotations. In contrast to our approach, (48) and (49) compute semantic precision and recall by examining the superclass sets of two annotations. Depending on the overlap among these sets, each superclass is classified as a true positive, false positive or false negative. These counts are then used to compute semantic precision and recall.
PP measures the proportion of the semantics annotated by 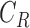 that are retrieved by
that are retrieved by 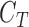 relative to the number of
relative to the number of 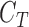 annotations. PR, on the other hand, measures the proportion of semantics that are retrieved by
annotations. PR, on the other hand, measures the proportion of semantics that are retrieved by 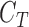 relative to the number of
relative to the number of 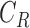 annotations. Thus, both PP and PR have a range of [0,1]. PP will decrease due to extra annotations by
annotations. Thus, both PP and PR have a range of [0,1]. PP will decrease due to extra annotations by 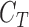 that are dissimilar from those in
that are dissimilar from those in 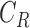 , while PR will decrease due to extra annotations in
, while PR will decrease due to extra annotations in 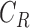 that are lacking from
that are lacking from 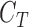 . Both use
. Both use 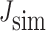 to measure semantic similarity and are computed at the character-state level rather than the individual EQ annotation level. Using
to measure semantic similarity and are computed at the character-state level rather than the individual EQ annotation level. Using 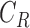 and
and 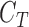 as an example, they are calculated as
as an example, they are calculated as
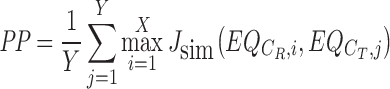 |
(1) |
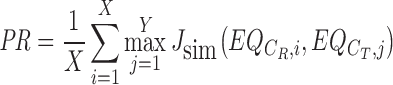 |
(2) |
where 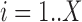 indexes the EQs from
indexes the EQs from 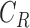 and
and 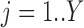 indexes the EQs from
indexes the EQs from 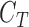 .
.
Author assessment of gold standard, curator and machine annotations
To assess how close EQ annotations created by the different sources came to the intent of the authors of the seven studies from which the characters were drawn, an author from each was invited to evaluate the relative performance of the annotation sources. Using SurveyMonkey (www.surveymonkey.com), we presented one author from each study with 10 randomly selected character states derived from their publication and asked them to rank the five different annotation sources (C1, C2, C3, SCP, and gold standard) for each state [Section1, Supplementary Materials].
Authors were given background material at the beginning of the survey describing the EQ method of character annotation. Authors were then asked to rank annotations in order of preference, with the annotation that best represented the meaning of the character state ranked first. Annotations were presented in random order, and the source of each annotation could not be tracked by the author. All of the EQ annotations for each character state generated by a particular annotation source were presented to the authors.
We used two statistics to test for differences among author preferences for the different annotation sources (50). Anderson’s statistic, 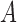 , was used to test whether the overall distribution of ranks was different in the observed (
, was used to test whether the overall distribution of ranks was different in the observed ( ) data than expected (
) data than expected (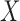 ):
):
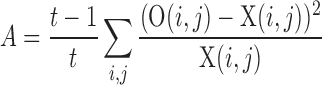 |
where 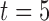 is the number of possible ranks and the expected number of observations
is the number of possible ranks and the expected number of observations 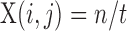 for factor
for factor 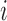 assigned rank
assigned rank 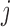 and number of observations
and number of observations 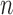 .
. 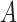 was tested against a
was tested against a 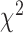 distribution for significance with
distribution for significance with 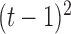 degrees of freedom. The null hypothesis is that all author preferences for all annotation sources will be equally frequent.
degrees of freedom. The null hypothesis is that all author preferences for all annotation sources will be equally frequent.
Friedman’s statistic, 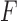 , was used to test if the mean ranks of the different annotation sources differed from chance
, was used to test if the mean ranks of the different annotation sources differed from chance
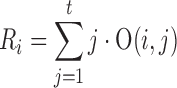 |
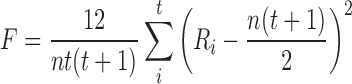 |
where 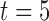 is the number of annotation sources,
is the number of annotation sources, 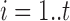 is the annotation source,
is the annotation source, 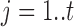 is the number of ranks that can be assigned to an annotation,
is the number of ranks that can be assigned to an annotation, 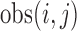 is the number of times rank
is the number of times rank 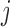 was assigned to factor
was assigned to factor 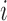 and
and 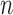 is the number of observations, as before.
is the number of observations, as before. 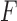 was tested against a
was tested against a 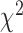 distribution for significance with
distribution for significance with 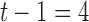 degrees of freedom.
degrees of freedom.
Results
Data sets and source code
The gold standard corpus is available in NeXML (51) (Gold_Standard-final.xml) and spreadsheet formats (Excel: GS-categories.xls; tab-delimited: GS-categories.tsv). The files include the full-text character and character state descriptions, the source study and the associated EQ phenotypes. The spreadsheet format also contains references for each phenotype to the character categories from the Phenoscape Guide to Character Annotation (44). The corpus in the different formats, as well as the ontologies and annotations generated in its production, have been archived at Zenodo (https://doi.org/10.5281/zenodo.1345307). The source code for the analysis of inter-curator and SCP consistency based on semantic similarity metrics, as well as the data and ontologies used as input, have been archived separately, also at Zenodo (https://doi.org/10.5281/zenodo.1218010). The source code used to randomly select characters for the gold standard (52) is available as part of the Phenex software code repository, which has been previously archived at Zenodo (https://doi.org/10.5281/zenodo.838793).
SCP is available in source code from GitHub (https://github.com/phenoscape/phenoscape-nlp/) under the Massachusetts Institute of Technology (MIT) license. The version used for this paper is the 0.1.0-goldstandard release (https://github.com/phenoscape/phenoscape-nlp/releases/tag/v0.1.0-goldstandard), which is also archived at Zenodo (https://doi.org/10.5281/zenodo.1246698).
Gold standard
The gold standard data set consists of 617 EQ phenotypes annotated for 203 characters and 463 character states. In total, these phenotypes are composed of 1096 anatomical terms (312 unique concepts) from Uberon, 698 Q terms (147 unique) from PATO and 148 spatial terms (30 unique) from BSPO. The data set contains 339 post-composed terms (277 anatomical and 62 Q terms) created by relating existing terms from the same or different ontologies.
New anatomy and Q terms were required for the completion of the gold standard annotations. From the full set of terms individually created by the curators during the experiment (Table 3), a total of 111 anatomical terms and 12 synonyms and 20 Q terms and 2 synonyms were added to the public versions of Uberon and PATO, respectively. The remaining subset of terms created by curators in the Merged ontology were not added to the public ontology versions either because a different term was chosen for the gold standard annotation of a particular character, or the term was determined to be invalid after discussion among curators.
Using 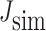 and
and 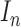 (see Measuring similarity betweenannotation sources) to measure semantic similarity between the four individual annotation sources (C1, C2, C3, and SCP) and the gold standard, we examined (i) whether the human annotations (C1, C2, and C3) showed an increase in similarity to the gold standard between the Naïve and Knowledge rounds and (ii) whether the machine annotations (SCP) showed an increase in similarity to the gold standard as ontologies progressed from the Initial, to Augmented and to the final Merged versions.
(see Measuring similarity betweenannotation sources) to measure semantic similarity between the four individual annotation sources (C1, C2, C3, and SCP) and the gold standard, we examined (i) whether the human annotations (C1, C2, and C3) showed an increase in similarity to the gold standard between the Naïve and Knowledge rounds and (ii) whether the machine annotations (SCP) showed an increase in similarity to the gold standard as ontologies progressed from the Initial, to Augmented and to the final Merged versions.
Figure 3 shows similarity (as measured by PP, PR, 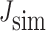 and
and 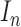 ) between annotations derived from the curators and the gold standard in Naïve and Knowledge curation rounds. Based on two-sided paired Wilcoxon signed rank tests, PR and
) between annotations derived from the curators and the gold standard in Naïve and Knowledge curation rounds. Based on two-sided paired Wilcoxon signed rank tests, PR and 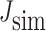 significantly differed for C1 (PR:
significantly differed for C1 (PR: 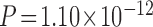 ,
, 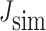 :
: 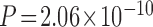 ) and C2 (PR:
) and C2 (PR: 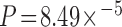 ,
, 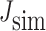 :
: 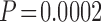 ), PP significantly differed for C1 (
), PP significantly differed for C1 (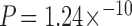 ) while
) while 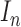 significantly differed for C1 (
significantly differed for C1 (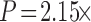
 between the Naïve and Knowledge rounds.
between the Naïve and Knowledge rounds.
Figure 3.
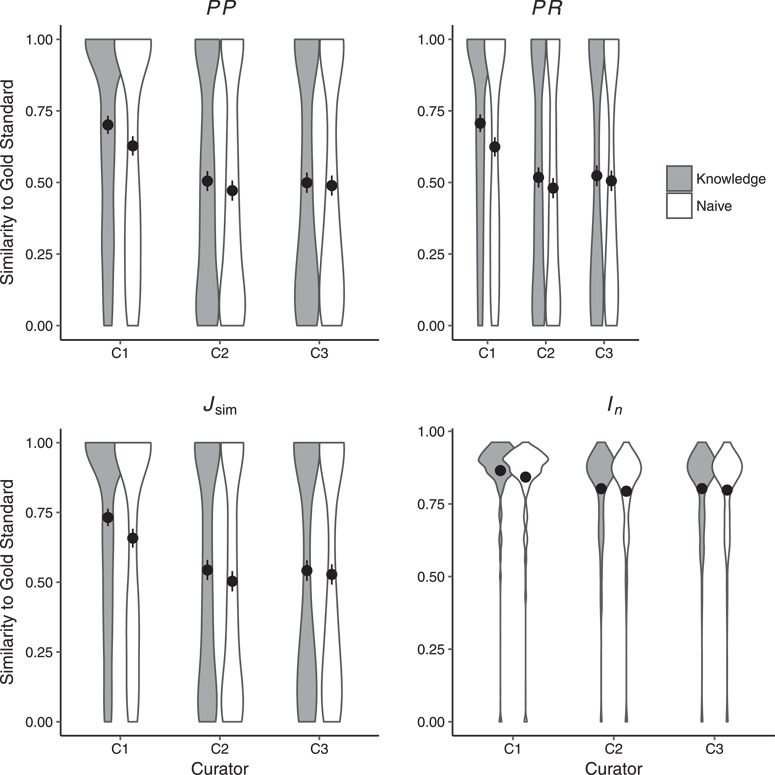
Similarity of human annotations to the gold standard in Naïve and Knowledge rounds. Shown are means across all 463 character states. Error bars represent two standard errors of the mean. Curators C1 (as per PP, PR, 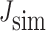 and
and 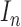 ) and C2 (as per PR,
) and C2 (as per PR, 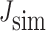 ) were significantly closer to the gold standard in the Knowledge round as compared to the Naïve round. Detailed results are shown in Supplementary Materials, Table 2.
) were significantly closer to the gold standard in the Knowledge round as compared to the Naïve round. Detailed results are shown in Supplementary Materials, Table 2.
Similarity of SCP annotations to the gold standard increased (26% average improvement across the four metrics) after new ontology terms had been added by human curators (detailed results are in Supplementary Materials, Table 2). The majority of statistics were significantly affected between the use of the Augmented and final Merged ontologies in both annotation rounds (Figure 4) with a few exceptions. PP and 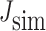 were not affected for C1 in the Knowledge round while PR was not affected in both rounds for C2. For C3,
were not affected for C1 in the Knowledge round while PR was not affected in both rounds for C2. For C3, 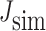 and PP in the Knowledge round and PR in Naïve round were not significantly affected.
and PP in the Knowledge round and PR in Naïve round were not significantly affected. 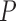 -values for individual comparisons are shown in Supplementary Materials, Table 2.
-values for individual comparisons are shown in Supplementary Materials, Table 2.
Figure 4.
Effect of ontology completeness on SCP performance as measured by similarity to the gold standard. ‘Mean Augmented’ is the mean of similarity scores from the three curator augmented ontologies; error bars show two standard errors of the mean. Significant differences in similarity between SCP and the gold standard were found for the majority of statistics across the two rounds. Detailed results are shown in Supplementary Materials, Table 2.
Consistency among human curators
We computed consistency among curators for the EQ annotations generated for each character state. Figure 5 shows the mean inter-curator consistency scores across three pairwise comparisons in the Naïve and Knowledge rounds, respectively, for PP, PR,  and
and  . The differences between Naïve and Knowledge rounds are not statistically significant (two-sided, paired Wilcoxon signed rank tests,
. The differences between Naïve and Knowledge rounds are not statistically significant (two-sided, paired Wilcoxon signed rank tests, 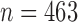 ,
, 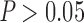 for all comparisons). These results echo those reported by Cui et al. (13) for the same experiment but reflect statistics that account for ontology structure or annotation density.
for all comparisons). These results echo those reported by Cui et al. (13) for the same experiment but reflect statistics that account for ontology structure or annotation density.
Figure 5.
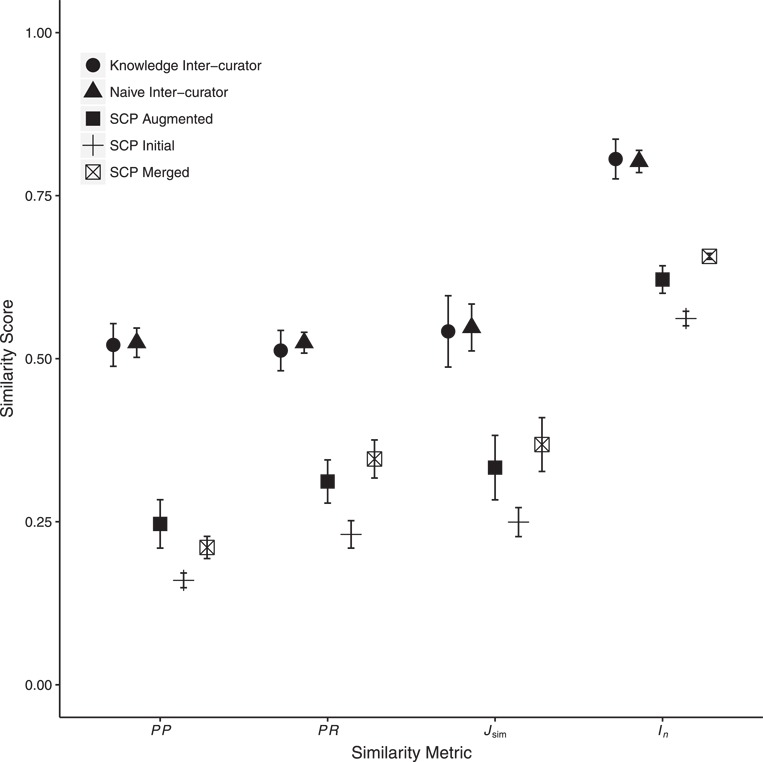
Mean inter-curator consistency and mean similarity between human- and machine (SCP)-generated annotations. Error bars show two standard errors of the mean. Inter-curator consistency results are shown for both the Naïve and Knowledge annotation rounds. SCP runs used either the Initial, C1, C2 or C3 Augmented or the Merged ontologies. Only SCP similarity to human-generated annotations from the Knowledge round are shown. Consistency between SCP annotations to human annotations was significantly lower than human inter-curator consistency. Across all metrics, SCP annotation similarity to human annotations increased significantly between the use of Initial to Augmented ontologies and again from Augmented to Merged ontologies except for PP (decreased from Augmented to Merged). Detailed results are in Supplementary Materials, Tables 4 and 5.
To evaluate whether the absence of a difference in inter-curator consistency between the Naïve and Knowledge rounds was because curators made mostly the same annotations in both rounds, Cui et al. (13) examined the changes in EQ annotations. They found that curators created substantially different EQ annotations in the Knowledge round as compared to the Naïve round. Each curator changed EQ annotations between these rounds for > 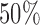 of character states. Among the EQs that were different between the two rounds, 29% were more complex, 33% were less complex and 38% retained the same complexity in the Knowledge round.
of character states. Among the EQs that were different between the two rounds, 29% were more complex, 33% were less complex and 38% retained the same complexity in the Knowledge round.
Because of the lack of significant differences between inter-curator consistency in Naïve and Knowledge rounds (Figure 5), we only report curator results for the Knowledge round in subsequent sections.
Human–machine consistency
Using the same metrics as above, we compared the human-generated annotations to those generated by SCP. To evaluate the effect of the completeness of ontologies on SCP performance, we ran SCP separately with the Initial ontology, each of the three (C1, C2 or C3) Augmented ontologies and the Merged ontology. Approximately 15–20% of character state annotations made by SCP using the different ontologies contained incomplete EQs. Incomplete EQs refer to those statements that are only partially matched to ontology terms, e.g. either E or Q terms are matched. In case of post-compositions, some parts needed in the composition are not matched to an ontology term. Human–machine comparisons involving character states with incomplete EQs were awarded a 0 similarity score.
We found that machine–human consistency was significantly lower than inter-curator consistency by an average of 35% across the four metrics (detailed results are in Supplementary Materials, Tables 3 and 4). The overall averages for the four scores in the human–machine comparison (unfilled square markers in Figure 5) are substantially lower than the averages for the comparisons among the human curators (circle markers in Figure 5). These comparisons are statistically significant for all four metrics (two-sided, paired Wilcoxon signed rank test: PP, 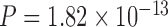 ; PR,
; PR, 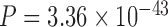 ;
; 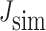 ,
, 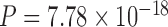 ;
; 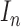 ,
, 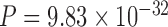 ).
).
Effect of ontology completeness on SCP–human consistency
Figure 5 shows the resulting PP, PR, 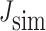 and
and 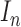 scores comparing SCP annotations generated with the Initial, Merged or Augmented ontologies (plus, unfilled square and filled square markers, respectively) to annotations from the human Knowledge round (as noted above, no statistically significant differences were found in SCP similarity to human annotations between the Naïve versus Knowledge rounds). However, almost universally, the scores among the similarity metrics increased as the ontologies progressed from Initial to Augmented and then from Augmented to Merged. The one exception is PP, which declined from the Augmented to the Merged ontology. All these increases, and the one decrease, were found to be statistically significant with two-sided paired Wilcoxon rank sum tests at the Bonferonni-corrected threshold of
scores comparing SCP annotations generated with the Initial, Merged or Augmented ontologies (plus, unfilled square and filled square markers, respectively) to annotations from the human Knowledge round (as noted above, no statistically significant differences were found in SCP similarity to human annotations between the Naïve versus Knowledge rounds). However, almost universally, the scores among the similarity metrics increased as the ontologies progressed from Initial to Augmented and then from Augmented to Merged. The one exception is PP, which declined from the Augmented to the Merged ontology. All these increases, and the one decrease, were found to be statistically significant with two-sided paired Wilcoxon rank sum tests at the Bonferonni-corrected threshold of 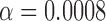 (Table 5).
(Table 5).
Table 5.
Comparison of SCP annotations using Initial, Augmented and Merged ontologies to measure the effect of ontology completeness on SCP–human consistency. Shown are 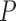 -values from two-sided paired Wilcoxon rank sum tests
-values from two-sided paired Wilcoxon rank sum tests
| Comparison | PP | PR |
|
|
|---|---|---|---|---|
| Initial versus Augmented ontologies |
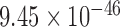
|
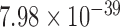
|
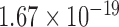
|
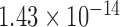
|
| Augmented versus Merged ontologies |
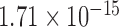
|
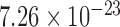
|
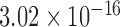
|
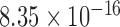
|
Author evaluation
We received responses to survey requests from six of the seven authors of the seven source studies (Table 2). Of the six completed surveys, 3 authors evaluated (ranked) phenotypes for all 10 characters; 1 author ranked 9 characters; and 2 authors ranked 8 characters. Table 4 reports the mean rank assigned to each curation source. The overall distribution of ranks differed significantly among the curation sources (Friedman’s statistic, 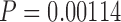 ), and there were significant differences among the mean ranks of each (Anderson’s statistic,
), and there were significant differences among the mean ranks of each (Anderson’s statistic, 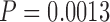 3). The gold standard had the lowest mean rank among the annotation sources, and authors ranked the gold standard annotations first for 21 out of 55 characters, indicating that the gold standard came closest to the meaning of the original authors more frequently than others. SCP had the highest mean rank, indicating that the machine annotations were farthest away from the original authors’ intent more frequently than the individual human curators or the gold standard.
3). The gold standard had the lowest mean rank among the annotation sources, and authors ranked the gold standard annotations first for 21 out of 55 characters, indicating that the gold standard came closest to the meaning of the original authors more frequently than others. SCP had the highest mean rank, indicating that the machine annotations were farthest away from the original authors’ intent more frequently than the individual human curators or the gold standard.
Table 4.
Evaluation of annotations by original authors. Authors ranked the annotations from the gold standard, the three human curators (C1, C2 and C3) and SCP. A lower value corresponds to an annotation deemed to be more accurate or precise
| Annotation source | Mean rank |
|---|---|
| Gold standard | 2.55 |
| C1 | 2.62 |
| C2 | 3.02 |
| C3 | 3.15 |
| SCP | 3.67 |
Discussion
Gold standard
Phenotype curation is typically done manually, without significant assistance from machines. It is difficult and time-consuming, and across a wide variety of fields, from agriculture to medicine, it has been found not to scale to the size of the task at hand (53, 54). Developing effective machine-based methods to aid in this task, however, requires standards against which to measure machine performance. The corpus of annotations developed here as a gold standard is the result of a methodical, multi-step process. Beginning with the choice of seven papers in the field of phylogenetic systematics that represent phenotypic diversity across extinct and extant vertebrates, a set of 203 characters (463 states) were randomly selected. Three experienced curators with training and experience in EQ annotation and research backgrounds in vertebrate anatomy and phylogenetics independently annotated the characters while simultaneously augmenting the initial ontologies. After merging their individual augmented ontologies, the three curators then discussed their annotations for each character state, and in some cases referenced external knowledge and contacted domain experts to clarify concepts, to develop consensus annotations. We then turned to the researchers who conceived of and described the original character states to assess the consensus annotations in relation to the machine-generated and individual curator annotations. Their judgement that the consensus annotations were on average closest in meaning to their original representation in free text validates use of the consensus annotations as a gold standard.
The gold standard presented here is the first of its type for evaluation of progress in machine learning of EQ phenotypes. It differs in a number of other ways from previously published gold standard corpora in the biomedical sciences. Rather than ensuring that every concept in the text of a character state is tagged with an ontology term [as is the case for a concept-based gold standard, such as CRAFT (19)], we focused on generating EQ annotations that best represent the anatomical variation described in a character. Thus, in some cases, the EQ or EQs chosen for a particular character state may not include ontology terms in one-to-one correspondence with concepts described in the character. For example, the character state ‘parietal, entocarotid fossa, absent’ was represented in a single EQ as E: ‘entocarotid fossa’ and Q: ‘absent’. Parietal was not annotated because entocarotid fossa is the focus of the character, not the structure (parietal) that it is a part of. In addition, the domain knowledge that entocarotid fossa is part of the parietal is encoded in the Uberon anatomy ontology.
Similarly, in some cases, character states describing the presence of a structure are not annotated directly in the gold standard. This is because presence can be inferred using machine reasoning on annotations to different attributes (e.g. shape) of the structure (55). In the following character state, for example ‘Hemipenis, horns: present, multi-cusped’ (36), the annotation in the gold standard consists of a single EQ phenotype: E, ‘horn of hemipenis’, and Q, ‘multicuspidate’. The presence of ‘horn of hemipenis’ is inferred by the assertion describing its shape and did not require a separate EQ annotation.
In other cases, ‘coarse’-level annotations were used that did not include every concept in the character state due to limited expressivity in the EQ formalism. For example, take the character ‘Quadrate, proximal portion, lateral condyle separated from the medial condyle by a deep but narrow furrow’. This relates three entities (lateral condyle of quadrate, medial condyle of quadrate, furrow) that cannot be expressed using the current EQ template model in Phenex: (33). Instead, this character state was annotated coarsely as E: ‘lateral condyle of quadrate’, Q: ‘position’ and RE: ‘medial condyle of quadrate’.
More complex annotations can be made using a less restrictive annotation tool (e.g. Protégé) rather than the EQ templates available in Phenex. However, allowing increased complexity when annotating in EQ format is likely to increase inter-curator variability. Pre-composed ontologies, i.e. phenotype ontologies, such as used by the Human Phenotype Ontology (HPO) (56), could, however, potentially decrease inter-curator variability because curators would be more likely to choose among existing terms rather than requesting a new one. Curators would also be aided by having access to existing, vetted annotations when creating new ones. Finally, providing additional context for character descriptions, such as specimen illustrations or images, could greatly aid curators in capturing the original intent of a character. Although most publications do include illustrations or images for some characters, rarely is this done for all characters in a matrix.
Finally, in some cases the gold standard annotations did not fully represent the knowledge (explicit or implicit) of a character due to limitations in the expressivity of OWL. For example, in the character ‘height of the vertebral centrum relative to length of the neural spine’, size is implicitly compared between two structures in the same individual. However, such within-individual comparison cannot be fully represented using an OWL class expression (57).
Inter-curator variation
The goal of evaluating the performance of automated curation tools is to engineer and improve machine-based curation to assist human curation as effectively as possible. Phenotype curation relies on deep domain and ontology knowledge as well as on expert judgement. Semantics in character descriptions can be variably interpreted, creating an inherent inter-curator variability. Thus, to judge the performance of automated curation tools against humans, it is important to first understand the level of variation between human curators as well as the sources of that variation.
As expected, we found considerable variation among human curators in our experiments. We observed that human curators achieved on average 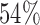 of the maximum possible consistency as measured by
of the maximum possible consistency as measured by 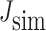 and
and 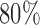 as measured by
as measured by 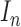 (Figure 5). This variability in inter-curator similarity is within the range reported in previous studies [e.g. (58)] and likely reflects the complexity of annotation tasks requiring domain knowledge, the ability to navigate large ontologies and experience and knowledge of annotation best practices. The inter-curator variability sets a ceiling for the maximum performance of a computational system if we assume that the human variability is primarily a consequence of the inherent ambiguity in how best to capture the semantics of the phenotype statement given the available ontologies.
(Figure 5). This variability in inter-curator similarity is within the range reported in previous studies [e.g. (58)] and likely reflects the complexity of annotation tasks requiring domain knowledge, the ability to navigate large ontologies and experience and knowledge of annotation best practices. The inter-curator variability sets a ceiling for the maximum performance of a computational system if we assume that the human variability is primarily a consequence of the inherent ambiguity in how best to capture the semantics of the phenotype statement given the available ontologies.
Much of the observed inter-curator variation could be assigned to a few general types of sources:
• Curators choose different but related terms. For example, terms may be related through subsumption (e.g. ‘circular’ and ‘subcircular’ in PATO) or sibling relationships (e.g. PATO: ‘unfused from’ and ‘separated from’).
• Curators make different decisions about how to post-compose entities. For example the E for the character ‘lateral pelvic glands, absent in males’ was composed differently by the three curators as ‘gland and [part_of some (lateral region and part_of some pelvis)]’, ‘lateral pelvic gland and (part_of some male organism)’ and ‘male organism and [has_part some (pelvic glands and in_lateral_side_of some multi-cellular organism)]’.
• Curators differ in how they composed an EQ even when choosing the same ontology terms. For example, two differently composed annotations for the character ‘pelvic plate semicircular, present’ were E: ‘pelvic plate and (bearer_of some semicircular)’ + Q: ‘present and E: pelvic plate + Q: semicircular’.
• Curators differ in how they added needed terms to the ontologies. For example, in the phenotype ‘dermal sculpture on skull-roof weak’, one curator created a new term ‘surface sculpting’ and post-composed the E ‘surface sculpting and (part_of some dermatocranium)’ as the ontological translation of the E because ‘dermal sculpture’ did not exist in the Uberon anatomy ontology. Another curator used PATO: ‘sculpted surface’ to create a post-composed E term ‘dermatocranium and (bearer_of some sculpted surface)’ to represent the same E.
Human–machine variation
SCP achieved, on average, 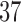 and
and 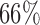 consistency with human curators using the most comprehensive (merged) ontology, as measured by
consistency with human curators using the most comprehensive (merged) ontology, as measured by 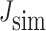 and
and 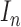 , respectively (Figure 5). This shows that the performance of SCP is significantly lower as compared to human inter-curator performance.
, respectively (Figure 5). This shows that the performance of SCP is significantly lower as compared to human inter-curator performance.
Usefulness of semantic similarity for partial matches
One of the major sources of annotation variation in either human or machine curators stems from choosing terms that are related to each other via subsumption or sibling relationships (see Inter-curator variation section). Comparisons of curator annotations from this experiment show that, on average, only 26% of character-state comparisons are exact matches. Given that the majority of curator annotation pairs are partial matches, the use of semantic similarity metrics that can quantify different degrees of similarity proves to be important.
Effect of external knowledge on inter-curator consistency and accuracy
One of the major differences between human and machine annotation is that humans can access external knowledge during curation, while machines cannot, beyond the encoded knowledge they have access to (here in the form of ontologies). Our measures of semantic similarity agreed with the results of Cui et al. (13) in showing that access to external knowledge had no effect on inter-curator consistency and did not further differentiate them from SCP’s annotations. Further, similarity to the gold standard was not generally increased. This was true despite the fact that curators changed annotations considerably between the Naïve and Knowledge rounds. Interestingly, while we expected a general increase in complexity when curators were at liberty to bring in additional knowledge, this was not borne out by the data.
These results indicate that lack of access to external knowledge is not one of the factors that contributes to SCP’s low performance with respect to human curators. This is encouraging because lack of access to external knowledge during machine curation would be a challenge to remedy.
Machine performance is improved as ontologies become more complete
Our results indicate that using more complete ontologies can significantly improve machine performance (Figures 4 and 5). This is encouraging because ontology completeness is continually improved through the synergistic efforts of the ontology and curator communities.
This finding leads to specific ideas for how the curation workflow could be optimized by alternating execution of steps between human curators and algorithms. For instance, an initial round of machine curation would identify character states in the data set for which good ontology matches were not found. Subsequently, human curators would judge whether the input ontology contains appropriate terms and focus on problem areas to add missing terms accordingly. Machines would then proceed with annotation using the human curator enhanced ontologies. Subsequently, human curators would review machine annotations and then either accept, modify or re-curate them on a per-annotation basis. In such a workflow, machines would valuably augment the work of humans in the annotation process.
Future work
Improving reasoning over EQ annotations
One of the major challenges with EQ annotations is efficiently calculating semantic similarity metrics. Specifically, for virtually all metrics, the first step is to identify common subsuming classes. Although in theory an OWL reasoner can perform this task, it can only identify named classes that already exist in the ontology. A brute-force approach in which a composite ontology is computed as the cross product of 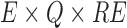 terms (for E, Q, RE; or even only
terms (for E, Q, RE; or even only 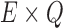 ) (59) would result in a background ontology too large even for efficient reasoners such as ELK, and the vast majority of its compound classes would not be needed as subsumers. Further work is needed to improve this method for efficiency (computational time and memory) of the semantic similarity scoring.
) (59) would result in a background ontology too large even for efficient reasoners such as ELK, and the vast majority of its compound classes would not be needed as subsumers. Further work is needed to improve this method for efficiency (computational time and memory) of the semantic similarity scoring.
Improving semantic charaParser
Cui et al. (13) identified a number of areas of potential improvement for SCP, and the present study further refines our understanding of where the machine curation is encountering obstacles. The observed shortcomings primarily fall in the areas of E post-composition, the handling of relational qualities in annotations and ontology searching in PATO. One way to improve the latter would be to enable the ontology search to locate multiple-word PATO qualities such as ‘posteriorly directed’, which in turn would allow more meaningful post-composed terms to be generated. As mentioned in Machineperformance is improved as ontologies become morecomplete, our results show that more comprehensive input ontologies will lead to improved performance of SCP.
Conclusions
The gold standard data set for EQ phenotype curation developed herein is a high-quality resource that will be of value to the sizable community of biocurators annotating phenotypes using the EQ formalism. As illustrated here, the gold standard enables assessment of how well a machine can perform EQ annotation and the impact of using different ontologies for that task. At present, machine-generated annotations are less similar to the gold standard than those of an expert human curator. The continued use of this corpus as a gold standard will enable training and evaluation of machine curation software in order to ultimately make phenotype annotation accurate at scale.
Supplementary Material
Acknowledgements
We thank M. Haendel and C. Mungall for suggestions on experiment design and ontology usage. We thank P. Chakrabarty, J. Clark, M. Coates, R. Hill, P. Skutschas and J. Wible for completing the author assessments. P. Fernando and L. Jackson provided valuable feedback on the gold standard and Phenoscape Guide to Character Annotation. We thank D. Blackburn for his helpful comments on this manuscript. This manuscript is based on work done by P. Mabee while serving at the US National Science Foundation. The views expressed in this paper do not necessarily reflect those of the National Science Foundation or the US Government.
Funding
National Science Foundation (DBI-1062542, EF-0849982, DBI-1147266).
Conflict of interest. None declared.
Database URL: http://kb.phenoscape.org
References
- 1. Smith B., Ashburner, M., Rosse C.. et al. (2007) The OBO Foundry: coordinated evolution of ontologies to support biomedical data integration. Nat. Biotechnol., 25, 1251–1255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Howe, D.G., Frazer, K., Fashena D.. et al. (2011) Data Extraction, Transformation, and Dissemination through ZFIN. The Zebrafish: Genetics, Genomics and Informatics, 104, 3rd edn, 313–325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Bradford, Y., Conlin, T., Dunn, N. et al. (2011) ZFIN: enhancements and updates to the Zebrafish Model Organism Database. Nucleic Acids Res., 39, D822–D829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Bowes, J.B., Snyder K.A., Segerdell, E.. et al. (2008) Xenbase: a Xenopusbiology and genomics resource. Nucleic Acids Res., 36, D761–D767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Blake, J.A., Bult, C.J., Eppig, J.T.et al. (2009) The Mouse Genome Database genotypes::phenotypes. Nucleic Acids Res., 37, D712–D719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Mungall, C., Gkoutos, G., Washington, N. et al. (2007) Representing phenotypes in OWL. In: Proceedings of the OWLED Workshop on OWL: Experience and Directions, Innsbruck, Austria. [Google Scholar]
- 7. Mungall, C.J., Gkoutos, G.V., Smith C.L. et al. (2010) Integrating phenotype ontologies across multiple species. Genome Biol., 11, R2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Deans, A.R., Lewis S.E., Huala E. et al. (2015) Finding our way through phenotypes. PLoS Biol., 13, e1002033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Loebe F., Stumpf F., Hoehndorf R.. et al. (2012) Towards improving phenotype representation in OWL. J. Biomed. Semantics, 3, 1–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Vogt L., Bartolomaeus T., Giribet G. (2010) The linguistic problem of morphology: structure versus homology and the standardization of morphological data. Cladistics, 26(3), 301–325. [DOI] [PubMed] [Google Scholar]
- 11. Vogt L. (2018) Towards a semantic approach to numerical tree inference in phylogenetics. Cladistics, 34, 200–224. [DOI] [PubMed] [Google Scholar]
- 12. Balhoff J.P., Dahdul W.M., Dececchi T.A. et al. (2014) Annotation of phenotypic diversity: decoupling data curation and ontology curation using Phenex. J. Biomed. Semantics, 5, 45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Cui H., Dahdul W., Dececchi A.. et al. (2015) Charaparser+EQ: performance evaluation without gold standard. In: Proceedings of the Association for Information Science and Technology, St. Louis, Missouri, 52, 1–10. [Google Scholar]
- 14. Mabee P.M., Ashburner M., Cronk Q. et al. (2007) Phenotype ontologies: the bridge between genomics and evolution. Trends Ecol. Evol., 22, 345–350. [DOI] [PubMed] [Google Scholar]
- 15. Campos D., Matos S., Lewin I.. et al. (2012) Harmonization of gene/protein annotations: towards a gold standard MEDLINE. Bioinformatics, 28, 1253–1261. [DOI] [PubMed] [Google Scholar]
- 16. Groza T. Oellrich A. and Collier N. (2013) Using silver and semi-gold standard corpora to compare open named entity recognisers. In: 2013 IEEE International Conference on Bioinformatics and Biomedicine (BIBM), Shanghai, China, 481–485. [Google Scholar]
- 17. Funk C., Baumgartner W., Garcia B. et al. (2014) Large-scale biomedical concept recognition: an evaluation of current automatic annotators and their parameters. BMC Bioinformatics, 15, 59. doi:10.1186/1471-2105-15-59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Mabee P., Balhoff J.P., Dahdul W.M. et al. (2012) 500,000 fish phenotypes: the new informatics landscape for evolutionary and developmental biology of the vertebrate skeleton. J. Appl. Ichthyol., 28, 300–305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Bada M., Eckert M., Evans D. et al. (2012) Concept annotation in the CRAFT corpus. BMC Bioinformatics, 13, 161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Pesquita C., Faria D., Falcão A.O.. et al. (2009) Semantic similarity in biomedical ontologies. PLoS Comput. Biol., 5, e1000443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Bada M., Hunter L. Vasilevsky N., et al. (2016) Gold-standard ontology-based annotation of concepts in biomedical text in the craft corpus: updates and extensions. In: ICBO/BioCreative, CEUR Workshop Proceedings, Corvallis, Oregon, 1747. [Google Scholar]
- 22. Kim J.-D., Ohta T., Tateisi Y. et al. (2003) Genia corpus—a semantically annotated corpus for bio-textmining. Bioinformatics, 19, i180–i182. [DOI] [PubMed] [Google Scholar]
- 23. Lu Z., Kao H.Y., Wei C.H. et al. (2011) The gene normalization task in BioCreative III. BMC Bioinformatics, 12, S2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Kors J.A., Clematide S., Akhondi S.A. et al. (2015) A multilingual gold-standard corpus for biomedical concept recognition: the Mantra GSC. J. Am. Med. Inform. Assoc., 22, 948–956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Oellrich A., Collier N., Smedley D. et al. (2015) Generation of silver standard concept annotations from biomedical texts with special relevance to phenotypes. PLoS One, 10, 1–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Rebholz-Schuhmann D., Yepes A.J.J., Van Mulligen E.M. et al. (2010) CALBC silver standard corpus. J. Bioinform. Comput. Biol., 8, 163–179. [DOI] [PubMed] [Google Scholar]
- 27. Wiegers T.C., Davis A.P., Cohen K.B. et al. (2009) Text mining and manual curation of chemical-gene-disease networks for the Comparative Toxicogenomics Database (CTD). BMC Bioinformatics, 10, 326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Söhngen C. Chang A. and Schomburg D. (2011) Development of a classification scheme for disease-related enzyme information. BMC Bioinformatics, 12, 329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Camon E.B., Barrell D.G., Dimmer E.C. et al. (2005) An evaluation of GO annotation retrieval for BioCreAtIvE and GOA. BMC Bioinformatics, 6, S17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Coates M.I. and Sequeira S.E. (2001) Early sharks and primitive gnathostome interrelationships. In: Ahlberg PE (ed).Major Events in Early Vertebrate Evolution. Taylor & Francis, London, 241–262. [Google Scholar]
- 31. Hill R.V. (2005) Integration of morphological data sets for phylogenetic analysis of Amniota: the importance of integumentary characters and increased taxonomic sampling. Syst. Biol., 54, 530–547. [DOI] [PubMed] [Google Scholar]
- 32. Skutschas P.P. and Gubin Y.M. (2012) A new salamander from the late Paleocene–early Eocene of Ukraine. Acta Palaeontol. Pol., 57, 135–148. [Google Scholar]
- 33. Nesbitt S.J., Ksepka D.T. and Clarke J.A. (2011) Podargiform affinities of the enigmatic Fluvioviridavis platyrhamphusand the early diversification of Strisores (‘Caprimulgiformes’+ Apodiformes). PLoS One, 6, e26350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Chakrabarty P. (2007) A morphological phylogenetic analysis of Middle American cichlids with special emphasis on the section Nandopsis sensu Regan. Museum of Zoology, University of Michigan, 198, 1–30. [Google Scholar]
- 35. O’Leary M.A., Bloch J.I., Flynn J.J. et al. (2013) The placental mammal ancestor and the post-K-Pg radiation of placentals. Science, 339, 662–667. [DOI] [PubMed] [Google Scholar]
- 36. Conrad J.L. (2008) Phylogeny and systematics of Squamata(Reptilia) based on morphology. Bull. Am. Mus. Nat. Hist., 310, 1–182. [Google Scholar]
- 37. Balhoff J.P., Dahdul W.M., Kothari C.R. et al. (2010) Phenex: ontological annotation of phenotypic diversity. PLoS One, 5, e10500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Mungall C.J., Torniai C., Gkoutos G.V. et al. (2012) Uberon, an integrative multi-species anatomy ontology. Genome Biol., 13, R5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Haendel M.A., Balhoff J.P., Bastian F.B. et al. (2014) Unification of multi-species vertebrate anatomy ontologies for comparative biology in Uberon. J. Biomed. Semantics, 5, 21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Gkoutos G.V., Green E.C., Mallon A.M. et al. (2004) Ontologies for the description of mouse phenotypes. Comp. Funct. Genomics, 5, 545–551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Gkoutos G.V., Green E.C.J., Mallon A.M. et al. (2004) Using ontologies to describe mouse phenotypes. Genome Biol., 6, R8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Dahdul W.M., Cui H., Mabee P.M. et al. (2014) Nose to tail, roots to shoots: spatial descriptors for phenotypic diversity in the Biological Spatial Ontology. J. Biomed. Semantics, 5, 34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Dahdul W.M., Balhoff J.P., Engeman J. et al. (2010) Evolutionary characters, phenotypes and ontologies: curating data from the systematic biology literature. PLoS One, 5, e10708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Dahdul W., Balhoff J., Dececchi A. et al. (2018) Phenoscape guide to character annotation. figshare. Fileset. 10.6084/m9.figshare.1210738.v2 10.6084/m9.figshare.1210738.v2. [DOI] [Google Scholar]
- 45. Whetzel P.L., Noy N.F., Shah N.H. et al. (2011) BioPortal: enhanced functionality via new web services from the National Center for Biomedical Ontology to access and use ontologies in software applications. Nucleic Acids Res., 39, W541–W545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Meeta M. and Pavlidis P. (2008) Gene ontology term overlap as a measure of gene functional similarity. BMC Bioinformatics, 9, 327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Resnik P. (1999) Semantic similarity in a taxonomy: an information-based measure and its application to problems of ambiguity in natural language. J. Artif. Intell. Res., 11, 95–130. [Google Scholar]
- 48. Euzenat J. (2007) Semantic precision and recall for ontology alignment evaluation. In: Proceedings of the 20th International Joint Conference on Artificial Intelligence (IJCAI’07), Hyderabad, India, 348–353. [Google Scholar]
- 49. Bada M., Baumgartner W.A. Jr, Funk C.. et al. (2014) Semantic precision and recall for concept annotation of text. In: Proceedings of Bio-Ontologies, Boston, Massachusetts, 30–37.
- 50. Brockhoff P.B., Best D.J. and Rayner J.C.W. (2003) Using Anderson’s statistic to compare distributions of consumer preference rankings. J. Sens. Stud., 18, 77–82. [Google Scholar]
- 51. Vos R.A., Balhoff J.P., Caravas J.A. et al. (2012) Nexml_ rich, extensible, and verifiable representation of comparative data and metadata. Syst. Biol., 61, 675–689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Balhoff J. (2017) https://github.com/phenoscape/Phenex/blob/master/src/main/java/org/phenoscape/main/SelectCharactersForExercise.java. Accession date: August 2, 2017.
- 53. Dahdul W., Dececchi T.A., Ibrahim N.. et al. (2015) Moving the mountain: analysis of the effort required to transform comparative anatomy into computable anatomy. Database (Oxford), 2015, bav040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. International Society for Biocuration (2018) Biocuration: distilling data into knowledge. PLoS Biol., 16, e2002846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Dececchi T.A., Balhoff J.P., Lapp H. et al. (2015) Toward synthesizing our knowledge of morphology: using ontologies and machine reasoning to extract presence/absence evolutionary phenotypes across studies. Syst. Biol., 64, 936–952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Köhler S., Vasilevsky N.A., Engelstad M. et al. (2017) The human phenotype ontology in 2017. Nucleic Acids Res., 45, D865–D876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Motik B., Grau B.C., Horrocks I. et al. (2009) Representing ontologies using description logics, description graphs, and rules. Artif. Intell., 173, 1275–1309. [Google Scholar]
- 58. Arighi C.N., Carterette B., Cohen K.B. et al. (2013) An overview of the BioCreative 2012 Workshop Track III: interactive text mining task. Database (Oxford), 2013, bas056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Washington N.L., Haendel M.A., Mungall C.J. et al. (2009) Linking human diseases to animal models using ontology-based phenotype annotation. PLoS Biol., 7, e1000247. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.