

Abstract
Salicylic acid (SA) is responsible for certain plant defence responses and NON EXPRESSER OF PATHOGENESIS RELATED 1 (NPR1) is the master regulator of SA perception. In Arabidopsis thaliana there are five paralogs of NPR1. In this work we tested the role of these paralogs in SA perception by generating combinations of mutants and transgenics. NPR2 was the only paralog able to partially complement an npr1 mutant. The null npr2 reduces SA perception in combination with npr1 or other paralogs. NPR2 and NPR1 interacted in all the conditions tested, and NPR2 also interacted with other SA-related proteins as NPR1 does. The remaining paralogs behaved differently in SA perception, depending on the genetic background, and the expression of some of the genes induced by SA in an npr1 background was affected by the presence of the paralogs. NPR2 fits all the requirements of an SA receptor while the remaining paralogs also work as SA receptors with a strong hierarchy. According to the data presented here, the closer the gene is to NPR1, the more relevant its role in SA perception.
Introduction
Plants use several pathways for their defence against pathogens. Broadly speaking, necrotrophic pathogens are dealt with by the methyl jasmonate pathway (MeJA, [1]), while biotrophic pathogens are resisted by means of the salicylic acid pathway (SA, [2]), with examples of positive and negative interactions between both pathways [3].
NON EXPRESSER OF PATHOGENESIS RELATED 1 (NPR1) is an important SA perception gene. When mutants of Arabidopsis thaliana (Arabidopsis) were found that did not recognize SA or its analogues, the results were three alleles of NO RESPONSE TO BTH 4 (NRB4, [4]), and more than fifty alleles of NPR1 [5–9]. NPR1 is a protein with several domains: a BTB/POZ (broad-complex, tramtrack, and bric-a-brac/poxvirus and zinc-finger) domain, and an ankyrin repeat domain. It also contains a nuclear localization sequence [10], and a transactivation domain in the C terminal [11].
NPR1 interactions have been described profusely. Among these interactors, there are some that negatively regulate its function, such as NIMIN1 (NIM1-INTERACTING 1, [12]), others that bind SA, such as βCAs (β CARBONIC ANHYDRASES, [13]), and others that bind to the promoters of defence genes, such as TGAs (members of the basic/leucine zipper-type transcription factors family, [14]). NPR1 binds SA and has been proposed as the SA receptor [15], although there alternative proteins proposed (see below). NPR1 is localized mainly in the cytoplasm in oligomeric form, and when SA is present, NPR1 is monomerized and concentrates in the nucleus [16]. NPR1 then activates TGAs and is regulated via degradation [17].
NPR1 has five paralogs in Arabidopsis (NPR2, NPR3, NPR4, BLADE ON PETIOLE 1 (BOP1), and BOP2). NPR3 and NPR4 (NPR3/4) have been shown to negatively regulate defences [18], perhaps through NPR1 [19] or independently [20].
BOP1 and BOP2 (BOP1/2) function redundantly, and the double mutant has a phenotype of ectopic blades along the petioles and some alterations in the flowers [21]. bop1 bop2 behaves similarly to wild type (wt) plants for SA perception [22], but is impaired in MeJA perception in a specific phenotype related to defence against Pseudomonas syringae pv. tomato DC3000 (Pto) [23]. NPR1 orthologs have been described in numerous species such as rice, apple, and tobacco ([24–26], respectively). No other paralogs or orthologs have been described that concentrate in the nucleus upon SA treatment like the Arabidopsis NPR1.
Plants with a deficient npr1 allele such as npr1-1 do not respond to SA or to its analogue benzothiadizole (BTH, [27]) in terms of defence or plant growth [22]. However, SA is able to induce the expression of some genes in an npr1 background [28]. Plants with NPR1 null alleles have a small but measurable response to SA in terms of defence and plant growth [9], which suggests that there may be other SA receptors.
We reasoned that the most likely candidates were the NPR1 paralogs (henceforth abbreviated to NPRs), an idea that had already been suggested in a previous study [9]. NPR3/4 have been proposed to act as SA receptors by regulating the NPR1 protein [19], in disagreement with NPR1 being the SA receptor [15]. In this paper we searched for the NPRs’ role in SA perception using several approaches and found that the NPRs, especially NPR2, do play a role in SA perception.
Material and methods
Plant growth and inoculation
Arabidopsis (Arabidopsis thaliana (L.) Heynh.) and Nicotiana benthamiana were sown and grown as described [22], in controlled environment rooms with days of 8 h at 21°C, 150 μmol m-2 s-1 of light intensity and nights of 16 h at 19°C. The treatments, inoculations, and sampling started 30 minutes after the initiation of the artificial day to ensure reproducibility. The following genotypes were used: npr1-1 [7], npr3 npr4 [18], bop1 bop2 [29], npr2 (N622643 from NASC, www.arabidopsis.info). 35S:GFP-NPR1 in npr1-70 [23], 35S:GFP-NPR1 in npr1-1 [23], 35S:NPR1-GFP in npr1-1 [30], 35S:NPR1 [31], and 35S:GFP-NRB4 [4].
Pseudomonas syringae pv. tomato DC3000 (Pto) was grown, inoculated and measured as described [32]. Briefly, 14-day old plants were inoculated by spray with Pto at 5*107 colony forming units (cfu) per mL with 0.02% Silwet L-77 (Crompton Europe Ltd, Evesham, UK). Three days later the amount of cfu per plant was quantified and represented on a logarithmic scale. In the eds-like experiment, twelve seven-week-old plants were hand inoculated with a needleless syringe containing Pto at 5*104 cfu/mL. Three leaves of each plant were completely infiltrated, and three days later the inoculated leaves were collected, weighed, and the amount of bacteria measured. For all the experiments, at least three independent treatments were performed (three independent sets of plants sown and treated on different dates). The statistical analyses were performed with Excel 2007 (Microsoft, Redmond, WA, USA), and R [33].
Chemical treatments
Primers and chemical products were purchased from SIGMA (St. Louis, MO, USA) unless otherwise stated. Benzothiadiazole (BTH, CGA 245704), in the form of a commercial product (Bion 50 WG, a gift from Syngenta Agro S.A., Madrid, Spain) was prepared in water for each treatment and applied with a household sprayer. The response to BTH in terms of fresh weight was done as reported [22]. In short, plants were treated with mock or 350 μM BTH four times over three weeks, after which the fresh weight of the plants was recorded and expressed as the ratio between BTH and mock treated plants. SA (in the form of sodium salicylate), and 4-hydroxybenzoic acid (HBA) were applied at 1 mM unless otherwise is stated.
SA in plates and in planta
Arabidopsis seeds were surface-sterilized for 10 min in ethanol and for 10 min in 1% formaldehyde and then washed five times with distilled water before distributing the seeds on agar plates. The medium contained 0.5x Murashige and Skoog salts (Duchefa BV, Haarlem, the Netherlands), 0.6% (w/v) Phyto Agar (Duchefa), 2% (w/v) sucrose, with 0, 200 or 300 μM SA (final concentration, in the form of sodium salicylate). The results were evaluated 7 days after transfer to growing conditions.
Yeast experiments
The cDNAs of the NPR1 paralogs were cloned in pDONR222 (Invitrogen, Barcelona, Spain) from RT-PCR, and then transferred to pDEST22 and pDEST32 (Invitrogen) for expression in yeast. Additionally, the pARC352 vector [34] was used for the triple interaction. Yeast n-hybrid analyses were carried out as described in [35]. Briefly, yeast was transformed with two cDNAs, one in pDEST22 and one in pDEST32. If the yeast was able to grow on a plate with no histidine, it was considered as an interaction. Since some proteins have a small background (autoactivation [4], as NPR2 (this work) or NRB4 [4]), the plates were supplemented with 5 mM or less of 3-Amino-1,2,4-triazole (3AT, an inhibitor of histidine biosynthesis) when indicated. Also, the yeast plates were supplemented with 100 μM SA (in the form of sodium salicylate), or 100 μM HBA, as specified in the figures.
Expression in planta and microscopy
The cDNAs of the NPR1 paralogs were transferred to pMDC43 [36] for expression of GFP fused proteins in planta. The plasmid pMDC-MBP was also used (a gift from Drs. Carrasco and Vera, IBMCP) for expression of Maltose Binding Protein fused proteins. For the BiFC experiments, the cDNAs were cloned in pYFC43 and pNFC43 [37]. N. benthamiana leaf tissue was mounted in water under a coverslip 4 days after infiltration with Agrobacterium tumefaciens containing the constructs. When indicated, plants were treated with 1 mM SA by spray and collected or visualized one day later. The Arabidopsis transgenic plants were three weeks old when photographed. A Leica TCS SL confocal laser scanning microscope (Leica, Heidelberg, Germany) using an HCX PL APO CS 40X/1.25 water objective was used to study the subcellular localization of the fluorescence-tagged proteins. Green fluorescent protein was visualized by 488-nm excitation with an Ar laser, and its emissions were examined with a band-pass filter for 500 to 530 nm.
RT-qPCR
Total RNA was extracted with RNAzole RT (SIGMA), following the manufacturer’s instructions. cDNA was synthesized with RevertAid First Strand cDNA Synthesis Kit (Fermentas, Madrid, Spain), and the quantitative PCR performed with LuminoCt Sybr Green qPCR Ready Mix (SIGMA) in a 7500 Fast RT-PCR Systems machine (Applied Biosystems, Madrid, Spain), following the manufacturer’s instructions. Three biological replicates were performed for each measurement. The obtained values were referred to the geometric average of three reference genes (At3G18780, At1G49240, and At5G60390), as described [38], and normalized, the value of Col-0 in mock (or HBA when indicated) being equal to one. The list of primers used is provided in S1 Table.
Immunoblots and co-sedimentation assays
Extracts and co-sedimentation assays were done essentially as published [13] by using an amylose resin (New England Biolabs, Schwalbach, Germany). For immunoblots analysis the following antibodies were used: rabbit polyclonal anti-GFP N-terminal antibody (SIGMA), rabbit polyclonal anti-maltose binding protein antibody (Abcam, Cambridge, UK), and rabbit polyclonal anti-PR1 from tobacco [39] kindly provided by S. Kauffmann (IBMP, Strasbourg, France). The secondary antibody was anti-rabbit IgG-peroxidase conjugate (SIGMA) and we used Amersham ECL Plus Western Blotting detection reagents (GE HealthCare, Little Chalfont, UK) detected with a LA-3000 Luminescent Image Analyzer (Fujifilm Life Science, Stamford, CT, USA).
Protein expression in E. coli and SA binding activity
NPR1 paralogs cDNAs were cloned into pHMGWA and expressed as described [40]; [13]. The proteins were purified by amylose affinity chromatography (New England Biolabs) and eluted with maltose. The SA binding activity assay was done as described [41]; [13]. In short, proteins were incubated 50 μM 4-AzSA (Santa Cruz Biotechnology, Dallas, TX, USA), followed by UV irradiation. Reaction mixtures were subjected to SDS–PAGE, and 4-AzSA crosslinked proteins were detected by immunoblot analyses using a sheep anti-SA antibody (Fitzgerald Industries International, Acton, MA, USA).
Accession numbers
Sequence data from this article can be found in the Arabidopsis Genome Initiative or GenBank/EMBL databases under the following accession numbers: NRB4, At1g15780; NPR1, At1g64280; NPR2, At4g26120; NPR3, At5g45110; NPR4, At4g19660; BOP1, At3g57130; BOP2, At2g41370; αDOX1, At3g01420; GRX480, At1g28480; OPR1, At1g76680; PR1, At2g14610; ANAC102, At5g63790; UGT1, At1g05560; NIMIN1, At1g02450; TGA2, At5g06950; TGA5, At5g06960; TGA6, At3g12250; TGA7, At1g77920; CA1, At3g01500; CA2, At5g14740; CA3, At1g23730; CA4, At1g70410.
Results
NPR1 like function in NPR1 paralogs
Our first approach was to transiently express the NPRs of Arabidopsis in Nicotiana benthamiana under the control of the promoter 35S. In order to check if there was any NPR1-like activity, our readout was the expression of the PR1 protein [42]. S1a Fig shows the immunodetection of PR1 after the transient expression of the Green Fluorescence Protein (GFP) fused to the NPRs, while the bottom shows the level of the GFP-NPRs as a control. The same experiment was repeated (S1b Fig) after an SA spray one day prior to the sampling. Without SA, NPR2 produced a stronger signal than NPR1, although the rest of the paralogs also induced PR1 expression to some degree. The expression of NPR1 along with exogenous SA produced an increase in PR1 with respect to the empty vector control. S2 Fig shows the images of the GFP experiment checked by confocal microscopy. NPR1 has been observed to concentrate in the nucleus with SA treatment [30]. The rest of the paralogs did not change their localization with SA, NPR2, NPR3, and NPR4 being in the nucleus, and BOP1 and BOP2 in the cytosol and nucleus.
We then transformed Arabidopsis npr1-1 plants [7] with all the 35S:GFP-NPRs. The plants were tested in T2 by applying SA, and then by inoculating with Pto. Out of 33 lines transformed with 35S:GFP-NPR1, 5 lines showed a reduction of the symptoms caused by Pto (i.e. complementation), with a clearly dominant segregation. When lines transformed with 35S:GFP-NPR2 were tested, 12 of 33 lines showed complementation. There was no complementation when lines transformed with 35S:GFP fused to the empty vector, NPR3, NPR4, BOP1, or BOP2 were tested (33 lines for each).
Three independent 35S:GFP-NPR2 lines that complemented the npr1-1 background were randomly selected and taken to homozygosis. Fig 1a shows the response of these three lines to BTH in terms of plant weight [22]. We included a 35S:GFP-NPR1 in npr1-1 line generated with the same vector as a control. The three 35S:GFP-NPR2 lines partially complemented the npr1-1 mutant, not very different from the control 35S:GFP-NPR1. The second phenotype tested was the induction of defence against a pathogen; the same genotypes were treated with water, SA, and BTH, and one day later were inoculated with Pto (Fig 1b). Clearly the 35S:GFP-NPR2 lines, while responding to SA, were intermediate in this phenotype. The third phenotype tested was the growth of the plants on a plate with SA. In this condition, npr1-1 plants do not develop true leaves, and the cotyledons are bleached [7]. Fig 1c shows that all the genotypes tested grew normally on plates without SA, while in 200 μM SA (Fig 1d) and 300 μM SA (Fig 1e), npr1-1 was affected. The 35S:GFP-NPR2 lines did not complement the npr1-1 mutation, although the control 35S:GFP-NPR1 was not completely wt. S3 Fig shows the images of the transgenic lines used, visualized by confocal microscopy after SA or mock treatment. We also included two NPR1 additional controls for reference (S3c–S3f Fig). As with N. benthamiana, NPR2 was localized in the nucleus whether or not SA had been added. The third line, 35S:GFP-NPR2c had a level of expression below our detection limit.
Fig 1. NPR2 complements npr1-1 in Arabidopsis.

(a) Homozygous transgenic plants 35S:GFP-NPR2 of Arabidopsis in an npr1-1 background were tested for their response to benzothiadizole (BTH, an analogue of SA), along with control genotypes. The response to BTH was measured as weight, and plants were treated with either mock or 350 μM BTH. The ratio is expressed as percentage of fresh weight (%FW). The letters above the bars indicate different homogeneous groups with statistically significant differences (Fisher’s LSD Test, P < 0.05). In all the figures that give numerical information, the data represent the average, with the error bars plotting the standard deviation. All the experiments were repeated at least three times with similar results. (b) The indicated genotypes (14-day-old plants) were treated with either 1 mM SA, 350 μM BTH or a mock solution. One day later the plants were inoculated with Pseudomonas syringae pv. tomato isolate DC3000 (Pto) at 5*107 colony forming units (cfu) per mL. Three days after inoculation, the growth of Pto was evaluated as the Logarithm of cfu per plant. Asterisks indicate statistically significant differences from the mock treatment in each genotype (P < 0.05 one asterisk, P < 0.01 two) using the Student’s t test (one tail). (c) The indicated genotypes were grown on MS plates, and the picture was taken at day 7 after germination. (d) The same experiment as in (a), with 200 μM SA. (e) The same experiment as in (a), with 300 μM SA.
Phenotype of knock out NPRs
We then constructed the sextuple mutant that lacks all the NPRs, and several combinations of KOs in the paralogs. Fig 2a shows these KO combinations upon BTH treatment. Adding npr2 to npr1-1 did not change anything, but surprisingly, the sextuple mutant grew even better than npr1-1. npr3 npr4 and bop1 bop2 both responded as wt to BTH, as reported [22], but the combination npr3 npr4 bop1 bop2 had an even stronger response than their parents (Fig 2a). The experiment was repeated with 35 μM BTH (ten times less). Fig 2b shows that again npr3 npr4 bop1 bop2 had a stronger response than their parents, in fact even stronger than the transgenic lines that overexpressed NPR1 or NRB4 [4].
Fig 2. Phenotypes of the NPR1 paralogs knockouts.

(a) SA perception of combinations of KOs, measured by fresh weight after BTH treatment, as in Fig 1a. The abbreviations used are: n1 = npr1-1; n2 = npr2; n3 = npr3; n4 = npr4; n5 = bop2; n6 = bop1. The combinations of mutants are indicated by the letter “n” and the corresponding numbers, e.g. n1234 correspond to the quadruple npr1-1 npr2 npr3 npr4. (b) Enhanced SA perception of the quadruple npr3 npr4 bop1 bop2. Plants were treated and measured as in (a), but with a ten-fold reduction in the BTH, to visualize increased sensitivity to SA. 35S:NPR1 and 35S:GFP-NRB4 are included as controls of enhanced perception ([31] and [4] respectively). (c) SA perception, measured as pathogen growth as in Fig 1b. (d) eds-like phenotype. Seven-week-old plants were hand infiltrated with Pto at 5*104 cfu/mL. Three days after inoculation, the growth of Pto was evaluated as the Logarithm of cfus per g of fresh weight.
Another phenotype tested was the growth of Pto after mock, SA, or BTH treatment (Fig 2c). In these experiments, npr2 plants behaved as wt and npr3 npr4 had a constitutive expression of defence [18]. But the combination npr2 npr3 npr4 produced plants closer to wt. A similar effect happened in npr1-1 npr3 npr4 and npr1-1 npr3 npr4 bop1 bop2. These lines did not respond to SA or BTH, but there was a certain level of resistance already in the mock treatment. However, when npr2 was introduced (npr1-1 npr2 npr3 npr4, and npr1-1 npr2 npr3 npr4 bop1 bop2, respectively) there was less resistance and they were closer to npr1-1 alone.
The genotype npr1-1 npr2 was more susceptible than npr1-1 alone by a narrow margin. This difference can be seen more clearly with the eds-like phenotype (Fig 2d). In this experiment, adult plants are inoculated by hand with a small amount of Pto [6], and the result is that npr2 in an npr1-1 background produced plants more susceptible to Pto.
There were no significant phenotypes of these genotypes on plates with SA (S4 Fig), nor was there any significant difference between bop1 bop2 and the sextuple mutant, regarding the “blade-on-petiole” phenotype (S5 Fig).
Gene expression in mutant combinations
One phenotype frequently measured in SA perception is the expression of well-known marker genes. In our case, we choose genes induced by SA in wt, especially those also induced in an npr1 mutant (or SA dependent, NPR1 independent (SdNi), [43];[44]). Fig 3a shows the expression of α dioxygenase1 (αDOX1, [45]) in several genotypes treated with mock or SA, one day before the sampling. In these conditions, the npr3 npr4 bop1 bop2 plants expressed much more αDOX1 upon SA treatment than the wt. A similar effect was detected in the case of one glutaredoxin (GRX480 [46], Fig 3b). The third gene studied was PR1, a gene strictly dependent on SA and NPR1, included as a control (Fig 3c). The results are expressed on a logarithmic scale, since the range expands by several orders of magnitude. Again, the induction of this gene in npr3 npr4 bop1 bop2 was stronger than the wt. The fourth gene in this set was 12-Oxophytodienoate Reductase 1 (OPR1 [47]). Fig 3d shows that this gene was repressed in wt and in npr3 npr4 bop1 bop2. OPR1 was also induced in npr1-1, and in all the genotypes that contained this mutation. Surprisingly, the induction of OPR1 in the sextuple mutant is stronger than in npr1-1 alone. In fact, the behaviour of the sextuple mutant in these four genes was quite different from npr1-1 or npr1-1 npr2, especially in the presence of SA. Since the SdNi genes have been described at very short times after SA treatment, we carried out a new experiment, with three genotypes and samples taken 2.5 h after SA spray. It is theoretically possible that the genes were induced by the chemical characteristics of SA, regardless of the specific recognition of SA in defence. To check this point, we used 4-hydroxybenzoic acid (HBA, an isomer of SA with no biological activity) as a mock control. We repeated two genes from the previous experiment, GRX480 (Fig 3e) and OPR1 (Fig 3f), and the results were quite different. For GRX480 with SA, the sextuple phenocopies npr1-1, while for OPR1 with SA the expression of the sextuple is quite different from npr1-1. Two more genes described as SdNi, Nac Domain Containing Protein 102 (ANAC102 [44]), and Udp-Glucose Transferase 1 (UGT1 [48]), were tested in our conditions (Fig 3g and 3h, respectively). In both cases, the levels of expression in the sextuple were lower than npr1-1, as with OPR1, indicating that part of their regulation is independent of NPR1, but dependent on the rest of NPRs. In these four genes at 2.5 h (Fig 3e–3h), the levels of expression for HBA treated plants in the sextuple were quite low. Clearly, HBA did not induce these genes in a similar fashion to SA, so the SdNi induction is quite specific to SA. Our data seem to show a paradox. npr3 npr4 bop1 bop2 have an enhanced response to SA, but the same mutations introduced in an npr1-1 npr2 background enhanced the phenotypes of insensibility to SA, in five of the genes and conditions tested. And in two genes and conditions the sextuple is closer to the wt than to npr1-1 npr2 (Fig 3a and 3b).
Fig 3. Gene expression depending on NPR1 paralogs.

RNA was extracted from two week old plants of the indicated genotypes. In the (a) to (d) panels, the samples were taken one day after treatments of mock or 1 mM SA, while in the (e) to (h) panels, the samples were taken 2.5 h after treatment of 1 mM 4-hydroxybenzoic (HBA, a isomer of SA with no biological activity) or 1 mM SA. (a) Fold enrichment of α -Dioxygenase 1, one day after treatment. (b) Fold enrichment of Glutaredoxin 480, one day after treatment. (c) Fold enrichment of PR1 one day after treatment, in logarithmic scale. (d) Fold enrichment of 12-Oxophytodienoate reductase 11, one day after treatment. (e) Fold enrichment of Glutaredoxin 480, 2.5 h after treatment. (f) Fold enrichment of 12-Oxophytodienoate reductase 11, 2.5 h after treatment. (g) Fold enrichment of NAC domain containing protein 102, 2.5 h after treatment. (h) Fold enrichment of UDP-glucose transferase 1, 2.5 h after treatment. The transcript levels of the indicated genes were measured by RT-qPCR, and the levels of expression are normalized to three reference genes and to the level of Col-0 in mock or HBA treatment.
Protein interactions among the NPRs
NPR2 has a function in SA perception, but does NPR2 work like NPR1? We tested whether NPR2 interacted with some of the proteins described as NPR1 interactors in the yeast two-hybrid system (Fig 4a and 4b). NPR2 did not interact with the repressor NIMIN1, but it did interact with some members of TGAs. While NPR2 interacted in our conditions with TGA2, TGA6, and TGA7 (Fig 4a), NPR1 interacted with all four tested (the same constructs were used in [4]). Fig 4b shows that NPR2 only interacts with one of the four βCAs proteins tested, whereas NPR1 was able to interact with all of them [13]. In all the cases tested, there was interaction with no SA in the media. Only in the case of βCA1f, SA increased the interaction with NPR2, while with NPR1 it was necessary [13].
Fig 4. NPR2 interactions.

(a) Interactions of NPR2 with proteins that interact with NPR1. Yeast transformed with the indicated plasmids and inserts were grown on three different sets of plates, by depositing a 0.5 μL drop of OD600s 1, 0.1, and 0.01 (indicated in the top of the first plate). The first plate contained minimal media supplemented with histidine (+His). The second had the same minimal media with no histidine (-His), 100 μM SA, and 5 mM 3-Amino-1,2,4-triazole (3AT), while the third plate is -His, 100 μM HBA, and 5 mM 3AT. (b) Interactions of NPR2 with β carbonic anhydrases (βCAs), as in (a) but without 3AT.
NPR1 has also been found to interact with NPR2, NPR3, and NPR4 [19]. In our conditions and among the paralogs, NPR1 interacted strongly in yeast with NPR2 (Fig 5a), with SA improving the interaction. When no 3AT was present, NPR1 also interacted with itself and with NPR4 in the absence of SA (Fig 5a). When the vectors were switched, NPR2 interacted strongly with NPR1 and less so with the BOPs (Fig 5b). The same interaction between NPR4 and NPR1 depending of the absence of SA could be found when the vectors were switched. Thus, NPR4 interacted with NPR1 in the presence of 0 to 5 mM 3AT. In the presence of SA (Fig 5d), the interaction between NPR4 and NPR1 is disrupted consistent with the data published [19]. Both NPR2 and NPR3 interacted with NPR4 in the presence of SA (Fig 5d), while NPR4 interacted with the BOPs regardless of the presence of SA (Fig 5d).
Fig 5. NPR1 paralogs in yeast two hybrid.

(a) Interactions of NPR1 with the paralogs. (b) Interactions of NPR2 with the paralogs. (c) Interactions of NPR3 with the paralogs. (d) Interactions of NPR4 with the paralogs. (e) Controls of empty plasmids.
As mentioned, the interaction between NPR1 and βCA1f is dependent on SA. However, if NRB4 (an interactor of βCA1f) is introduced, the interaction is independent of SA [13]. We tested if any of the NPRs could have a similar effect, and only NPR2 had some effect if we grew the yeast long enough to see the colonies in the control (S6a Fig). Perhaps the interaction of NPR1 or NPR2 with NPR3 and NPR4 would be improved with NPR3 and NPR4 both present. We tried several combinations in yeast, but did not detect any interaction (S6b Fig).
After the yeast, we tested the interactions among the NPRs in N. benthamiana with transient expression. We checked by Bimolecular Fluorescence Complementation (BiFC [49]), and Fig 6 shows the results of the interactions between NPR1 or NPR2 with the NPRs. In general, we detected interactions in all the cases, with NPR2 giving a stronger signal than NPR1.
Fig 6. In planta interactions among NPR1 paralogs.

Bimolecular fluorescence complementation (BiFC) among NPR1 paralogs. The constructs were agroinfiltrated with 1mM SA treatment one day before visualization. The first protein is fused with the N-terminal part of GFP, and the second with the C-terminal. (a) NPR1 and NPR1. (b) NPR2 and NPR1. (c) NPR1 and NPR2. (d) NPR2 and NPR2. (e) NPR1 and NPR3. (f) NPR2 and NPR3. (g) NPR1 and NPR4. (h) NPR2 and NPR4. (i) NPR1 and BOP1. (j) NPR2 and BOP1. (k) NPR1 and BOP2. (l) NPR2 and BOP2. (m) NPR1 and AKIN10, as a negative control[37]. (n) NPR2 and AKIN10, as a negative control. The bars in these pictures represent 20 μm.
A better way to determine interactions between two proteins is to coprecipitate them. Fig 7 shows such an experiment. We transiently expressed GFP-NPR2 in N. benthamiana (Fig 7a) and MBP fused to several NPRs (Fig 7b). After the NPRs were pulled down by affinity, NPR2 was identified in the precipitate. In these conditions, NPR2 interacts with itself, NPR1, NPR3, and with NPR4 (Fig 7c). Fig 7d shows the purified MBP-NPRs as controls. We used MBP-AKIN10 [37], an unrelated protein, as a control. Note that the amount of MBP-AKIN10 recovered is less than the MBP-NPRs, so that this control needs to be taken with some caution.
Fig 7. Co-sedimentation among paralogs.
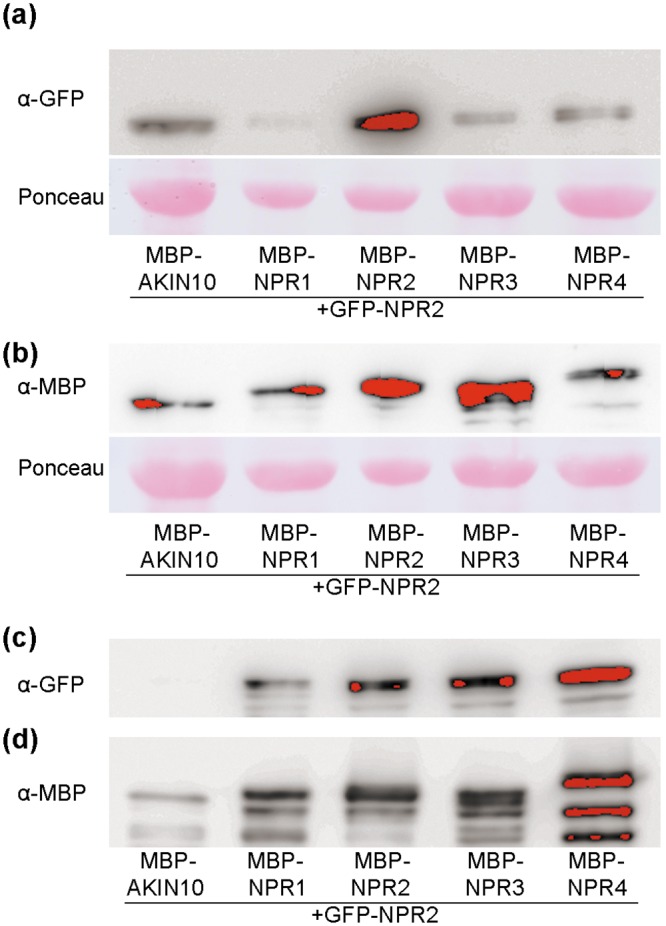
(a) GFP-NPR2 and the MBP fused to some NPR1 paralogs were transiently expressed in N. benthamiana by agroinfiltration and pulled-down with amylose resin. MBP-AKIN10 [37] is an unrelated protein, used as negative control with a similar size to the MBP-NPRs. The panel shows GFP-NPR2 detected by immunoblot before treatment with resin. Ponceau-S staining of the nitrocellulose membrane is shown as a loading control. (b) Expression of MBP-NPRs detected by immunoblot analysis before treatment with resin. (c) The panel shows the eluted fraction from the resin detecting GFP-NPR2 by immunoblot analysis. (d) The same eluted fraction detecting MBP-NPRs.
Protein stability and function of the NPRs
NPR3 and NPR4 regulate the stability of NPR1 [19]. We wondered if the same regulation affects NPR2, so we coexpressed NPR1 with the paralogs (Fig 8a) and NPR2 with the paralogs (Fig 8b). With NPR1, we did not see any evidence of this regulation, in fact there is less GFP-NPR1 in the presence of MBP-NPR1 than in the presence of MBP alone (Fig 8a). It is true that in the presence of NPR3, there is less NPR1 than with other paralogs, but the levels of protein are still above the control with MBP alone. In the case of NPR2, the levels of protein are quite constant, except in the presence of MBP-NPR1 (Fig 8b).
Fig 8. Stability of NPR1 and NPR2 in the presence of the NPR1 paralogs.

(a) Stability of GFP-NPR1 when co-expressed with the NPR1 paralogs fused to MBP. The constructs were agroinfiltrated with 1mM SA treatment one day before sampling. (b) Stability of GFP-NPR2 when co-expressed with the NPR1 paralogs, as in (a). Ponceau-S staining is shown below as a loading control.
NPR1, NPR3, and NPR4 bind SA [15];[19]. Perhaps this characteristic is common to all the NPRs, or perhaps is only found in the NPRs that have a direct function in SA perception. We analyzed the SA binding of all the NPRs together [41], and Fig 9a shows that all the NPRs have some SA binding with this particular technique, while Fig 9b shows the amount of protein used in the experiment. In this assay, BOP1 and BOP2 had a very weak binding, NPR3 and NPR4 had strong activity, and as we would expect given the presented phenotypes, NPR2 bound SA stronger than NPR1.
Fig 9. SA binding of the NPR1 paralogs.

(a) Purified recombinant proteins were incubated with 4-AzSA, then treated with UV light. 4-AzSA-cross-linked proteins were detected by immunoblot analysis with anti-SA antibody. The first line corresponds to a negative control, where the UV light was omitted. (b) Loading control, the same amount of protein used in A was detected with an anti-MBP. The image was cut to show the MBP empty vector, as indicated by the vertical line.
Discussion
NPR2 is part of the SA perception
NPR1 is the master regulator of SA perception, a position reached by genetic (e.g. [7]), biochemical (e.g. [15]), and molecular approaches (e.g. [50]). However, there are several lines of evidence that suggest other genes participate in SA signalling. First, NPR3 and NPR4 bind SA and negatively regulate the levels of NPR1 [19]. Secondly, plants with npr1 null mutation retain a part of the SA perception [9]. Third, there are genes induced by SA independently of NPR1 [28].
NPR2, in spite of being more similar to NPR1 than the other NPRs, has not been studied in detail until now. We considered the hypothesis that NPR2 could act with NPR3 and NPR4, or alternatively, with NPR1. NPR2 complemented an npr1-1 mutation (Fig 1), while the other NPRs did not. NPR2 and NPR1 were not interchangeable, since NPR2 was not as efficient as NPR1 in complementing the different phenotypes. Moreover, in our experiments NPR2 was more stable than NPR1, measured as protein accumulation (Fig 8, S1, S2 and S3 Figs).
The phenotypes produced in the 35S:NPR2 plants could be attributed to the ectopic and unregulated overexpression, although none of the other NPRs have these phenotypes. However, while the mutant npr2 did not have a phenotype in our conditions, the presence of npr2 did produce a phenotype when combined with npr1-1 or other nprs (Fig 2). Therefore, NPR2 plays a role in SA perception.
If NPR2 plays a role in SA perception, it has to be in the right place, meet with the right proteins, and with the right chemical. NPR2 localized in the nucleus and did not change its localization upon SA treatment (S2 and S3 Figs). This behaviour is different from NPR1, which concentrates in the nucleus in the presence of SA [30]. But NPR1 is the exception, since none of the others NPRs were affected by the SA (S2 and S3 Figs). In fact, a detailed study of two NPR1 orthologs from tobacco showed a continuous nuclear localization [26].
Regarding the interactions, NPR2 clearly interacted with NPR1. Note that in our yeast system (see Methods for details), the interaction between NPR2 and NPR1 was the strongest one (Fig 5), with other interactions present with less stringent conditions, including the interaction between NPR4 and NPR2 in the presence of SA (Fig 5). This result partially disagrees with some of the published interactions in yeast [19], although no interactions were found between NPR1 and NPR3/4 in yeast in a recent work [20]. Our data demonstrated that NPR1 interacts with NPR4 in the absence of SA and this interaction is disrupted by SA as published (Fig 5 [19]). We attribute this discrepancy to the use of a different system, which in our case is known to be more restrictive [51]. In fact, the yeast system reproduced the interaction of NPR1 with itself, which can be detected in planta [52], only in the less stringent conditions (Fig 5). In the BiFC system all the NPRs interacted with the others (Fig 6), but a more detailed experiment of co-sedimentation showed that NPR2 interacted in planta with NPR3 and NPR4 to some degree, as reported for NPR1. Moreover, NPR2 also interacted with NPR1 and with itself (Fig 7). NPR2 also interacted with part of the NPR1 interactors like three TGAs (Fig 4), so the connection with the gene activation upon SA perception is direct.
NPR3/4 have been proposed to be relevant in SA perception by regulating the amount of NPR1 [19], and perhaps NPR2 could regulate the amount of NPR1 in opposition to NPR3/4. The results in transient assays in N. benthamiana showed that, in the presence of NPR2, there was indeed an increase of NPR1 with respect to the empty vector control (Fig 8). It was not a symmetric interaction, since NPR1 reduced the levels of NPR2, with no effect on the rest of NPRs in these conditions (Fig 8). We did not detect a decrease of NPR1 in the presence of NPR3 or NPR4 as reported [19], perhaps due to the transient nature of our experiment.
NPR2 binds SA at least as strongly as NPR1 (Fig 9). That NPR1 binds SA has been described before [15];[53], and the same is true for NPR3/4 [19]. There was even a small amount of SA binding in BOP1/2 in this assay, requiring further confirmation by other methods. Although there was no apparent phenotype with SA in the bop1 bop2 mutant [29] (Fig 2), these weak interactions could explain the phenotypes in some combinations of mutants, described below.
To sum up, our results show that NPR2 can complement npr1, the lack of NPR2 produces a measurable phenotype, NPR2 interacts with some of the proteins with which NPR1 interacts, and binds SA. Therefore NPR2 acts as a SA receptor in the same way as NPR1.
What is the role of NPR2 in planta? NPR2 reaches peak expression in senescent leaves and later stages of fruit formation (including dry seeds [54]). In terms of global expression, NPR2 is expressed five times lower than NPR1 ([55]). The data of NPR2 expression could explain why its role in SA perception is only detected when ectopically expressed, or in an npr1 background. In normal conditions, we speculate that NPR2 would play a role in SA perception in the mentioned peaks of expression, and also as an evolutive reservoir for NPR1.
SA dependent, NPRs independent gene expression
Since npr2 has a phenotype with SA in combination with npr1-1, we reasoned that perhaps the genes induced by SA in an npr1-1 mutant (SdNi) would alter their expression in an npr1-1 npr2 background. When we performed the experiment, the results were not as clear-cut as expected (Fig 3). In four cases, the sextuple mutant is more extreme than npr1-1 (either in induction or repression), while in two the sextuple is closer to wt than to npr1-1. In the four cases where npr1-1 npr2 was tested alongside the sextuple, there was no significant difference between npr1-1 and npr1-1 npr2. Thus, instead of some genes being NPR1 and NPR2 dependent or independent, it would be more accurate to describe them as NPRs dependent or independent. It is clear that part of the SdNi genes are NPRs dependent. In the case of the gene GRX480, which we could call NPRs independent in the short term, it is more difficult to categorize in the long term (Fig 3).
The expression of SdNi genes did not have an impact on the defence or growth of the plant with SA that we could measure, since npr1-1 plants with SA are as susceptible as with a mock treatment (Fig 2c). Perhaps this gene activation is unrelated to the defence response and is a consequence of the chemical characteristics of SA. In the four genes tested, HBA did not have the same effect as SA, so it seems that this gene activation is SA specific. It is possible that there are additional receptors for SA besides the NPRs, and perhaps these putative receptors are relevant for the set of responses to SA in development [56].
The rest of the paralogs regulate SA perception
BOP1/2 are two paralogs of NPR1 with a defined role in development [29]. We found that the rest of paralogs did not have a function in the same phenotypes affected by BOP1/2 (S5 Fig), so their role in development is not shared among the NPRs. Since the sextuple is viable, we can say that there is no other obvious function in development shared by the NPRs. The defensive role of BOP1/2 was considered to affect only part of the resistance triggered by MeJA [23], but the work here presented shows that they also affect SA response (Fig 2a and 2b). Since npr3 npr4 bop1 bop2 plants respond more strongly to BTH than their parentals npr3 npr4, and bop1 bop2, perhaps BOP1/2 have the same function as that attributed to NPR3/4, namely to regulate the levels of NPR1 [19]. The effect of the bops is only perceived when they are in an npr3 npr4 background, so there is a hierarchy, where NPR3/4 are more relevant, and BOP1/2 are less relevant for this phenotype. The effect of the BOPs could be through their interaction with NPR2 and NPR4 (Fig 5), or perhaps due to competition for shared factors between BOP1/2 and the rest of the paralogs. It has been said that BOP1/2 have a different C-terminal domain than NPR1 and NPR3/4, a domain involved in SA-binding in these paralogs [15]; [20]. The response of these genotypes in the growth altered by BTH could be considered unique, since it was not observable in the growth of the pathogen (Fig 2c). However, Fig 3 shows that in three out of four genes tested, there was a stronger induction by SA in npr3 npr4 bop1 bop2 with respect to the wt. Therefore, this is not a unique phenotype of BTH, or related to growth, but related to SA perception, and NPR3/4 and BOP1/2 negatively regulate SA perception.
At the same time, the sextuple mutant pointed in the opposite direction, where not only NPR2, but all the NPRs positively participate in SA perception. The best example was PR1 (Fig 3c). While in an npr1-1 background the levels of PR1 were quite low (giving NPR1 its name), in the sextuple mutant the levels were even lower. In the rest of the genes and conditions, the sextuple mutant behaved differently than npr1-1 in all but in GRX480 at short times. Note that in some cases this difference took the sextuple closer to wt than to npr1-1 (Fig 3a and 3b). Thus, besides NPR1, some of the SA signalling goes through the NPRs, and yet part of the SA signalling is going through unknown mechanism(s).
This contradiction between a positive and a negative role for NPR3/4 and BOP1/2 could be explained by the difference in their background. The evidence for a positive role is presented in an npr1-1 background, while the negative role is presented in an NPR1 background. Consequently the difference in behaviour is due to the relation of NPR3/4 and BOP1/2 with NPR1. Since these four proteins bind SA to some degree, we suggest that although these proteins perceive SA, they are less efficient in producing the signal. When NPR1 is present, these four proteins bind SA and act as competitors to NPR1. When NPR1 and NPR2 are not functional, NPR3/4 and BOP1/2 (in small amounts) are able to trigger SA signalling.
Supporting information
(PDF)
(PDF)
(PDF)
(PDF)
(PDF)
(PDF)
(PDF)
Acknowledgments
We appreciate the opinions and generous help of Drs. Pablo Vera and Jeff Dangl with the manuscript.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by the Spanish Ministry of Economy and Competitiveness (www.idi.mineco.gob.es/portal/site/MICINN, Grant BIO2013-45444-P to PT) and the Valencia Regional Government (www.ceice.gva.es/web/ciencia, Grant ACOMP/2013/052 to PT). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Goossens J, Fernandez-Calvo P, Schweizer F, Goossens A. Jasmonates: signal transduction components and their roles in environmental stress responses. Plant Mol Biol. 2016;91(6):673–89. 10.1007/s11103-016-0480-9 . [DOI] [PubMed] [Google Scholar]
- 2.Yan S, Dong X. Perception of the plant immune signal salicylic acid. Curr Opin Plant Biol. 2014;20:64–8. 10.1016/j.pbi.2014.04.006 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Robert-Seilaniantz A, Grant M, Jones JD. Hormone crosstalk in plant disease and defense: more than just jasmonate-salicylate antagonism. Annu Rev Phytopathol. 2011;49:317–43. Epub 2011/06/15. 10.1146/annurev-phyto-073009-114447 . [DOI] [PubMed] [Google Scholar]
- 4.Canet JV, Dobon A, Tornero P. Non-recognition-of-BTH4, an Arabidopsis mediator subunit homolog, is necessary for development and response to salicylic acid. Plant Cell. 2012;24(10):4220–35. 10.1105/tpc.112.103028 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Delaney TP, Friedrich L, Ryals JA. Arabidopsis signal transduction mutant defective in chemically and biologically induced disease resistance. Proc Natl Acad Sci U S A. 1995;92(14):6602–6. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Glazebrook J, Rogers EE, Ausubel FM. Isolation of Arabidopsis mutants with enhanced disease susceptibility by direct screening. Genetics. 1996;143:973–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cao H, Glazebrook J, Clarke JD, Volko S, Dong X. The Arabidopsis NPR1 gene that controls systemic acquired resistance encodes a novel protein containing ankyrin repeats. Cell. 1997;88(1):57–63. . [DOI] [PubMed] [Google Scholar]
- 8.Shah J, Tsui F, Klessig DF. Characterization of a salicylic acid-insensitive mutant (sai1) of Arabidopsis thaliana identified in a selective screen utilizing the SA-inducible expression of the tms2 gene. Mol Plant Microbe Interact. 1997;10:69–78. 10.1094/MPMI.1997.10.1.69 [DOI] [PubMed] [Google Scholar]
- 9.Canet JV, Dobón A, Roig A, Tornero P. Structure-Function Analysis of npr1 Alleles in Arabidopsis Reveals a Role for its Paralogs in the Perception of Salicylic Acid. Plant, Cell & Environ. 2010;33:1911–22. Epub Jun 18. 10.1111/j.1365-3040.2010.02194.x [DOI] [PubMed] [Google Scholar]
- 10.Dong X. NPR1, all things considered. Curr Opin Plant Biol. 2004;7(5):547–52. 10.1016/j.pbi.2004.07.005 [DOI] [PubMed] [Google Scholar]
- 11.Rochon A, Boyle P, Wignes T, Fobert PR, Despres C. The coactivator function of Arabidopsis NPR1 requires the core of its BTB/POZ domain and the oxidation of C-terminal cysteines. Plant Cell. 2006;18(12):3670–85. Epub 2006 Dec 15. 10.1105/tpc.106.046953 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Weigel RR, Pfitzner UM, Gatz C. Interaction of NIMIN1 with NPR1 modulates PR gene expression in Arabidopsis. Plant Cell. 2005;17(4):1279–91. 10.1105/tpc.104.027441 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Medina-Puche L, Castello MJ, Canet JV, Lamilla J, Colombo ML, Tornero P. beta-carbonic anhydrases play a role in salicylic acid perception in Arabidopsis. PLoS One. 2017;12(7):e0181820 10.1371/journal.pone.0181820 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhou J-M, Trifa Y, Silva H, Pontier D, Lam E, Shah J, et al. NPR1 differentially interacts with members of the TGA/OBF family of transcription factors that bind an element of the PR-1 gene required for induction by salicylic acid. Mol Plant Microbe Interact. 2000;15:191–202. [DOI] [PubMed] [Google Scholar]
- 15.Wu Y, Zhang D, Chu Jee Y, Boyle P, Wang Y, Brindle Ian D, et al. The Arabidopsis NPR1 Protein Is a Receptor for the Plant Defense Hormone Salicylic Acid. Cell Reports. 2012;1(6). [DOI] [PubMed] [Google Scholar]
- 16.Tada Y, Spoel SH, Pajerowska-Mukhtar K, Mou Z, Song J, Wang C, et al. Plant immunity requires conformational charges of NPR1 via S-nitrosylation and thioredoxins. Science. 2008;321(5891):952–6. 10.1126/science.1156970 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Seyfferth C, Tsuda K. Salicylic acid signal transduction: the initiation of biosynthesis, perception and transcriptional reprogramming. Frontiers in plant science. 2014;5:697 10.3389/fpls.2014.00697 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Zhang Y, Cheng YT, Qu N, Zhao Q, Bi D, Li X. Negative regulation of defense responses in Arabidopsis by two NPR1 paralogs. Plant J. 2006;48(5):647–56. 10.1111/j.1365-313X.2006.02903.x . [DOI] [PubMed] [Google Scholar]
- 19.Fu ZQ, Yan S, Saleh A, Wang W, Ruble J, Oka N, et al. NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature. 2012;486(7402):228–32. 10.1038/nature11162 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ding Y, Sun T, Ao K, Peng Y, Zhang Y, Li X, et al. Opposite Roles of Salicylic Acid Receptors NPR1 and NPR3/NPR4 in Transcriptional Regulation of Plant Immunity. Cell. 2018. 10.1016/j.cell.2018.03.044 . [DOI] [PubMed] [Google Scholar]
- 21.Ha CM, Kim GT, Kim BC, Jun JH, Soh MS, Ueno Y, et al. The BLADE-ON-PETIOLE 1 gene controls leaf pattern formation through the modulation of meristematic activity in Arabidopsis. Development. 2003;130(1):161–72. Epub 2002/11/21. . [DOI] [PubMed] [Google Scholar]
- 22.Canet JV, Dobón A, Ibáñez F, Perales L, Tornero P. Resistance and biomass in Arabidopsis: a new model for Salicylic Acid perception. Plant Biotech J 2010; 8:126–41. [DOI] [PubMed] [Google Scholar]
- 23.Canet JV, Dobon A, Fajmonova J, Tornero P. The BLADE-ON-PETIOLE genes of Arabidopsis are essential for resistance induced by methyl jasmonate. BMC Plant Biol. 2012;12(1):199 Epub 2012/11/03. 10.1186/1471-2229-12-199 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bai W, Chern M, Ruan D, Canlas PE, Sze-To WH, Ronald PC. Enhanced disease resistance and hypersensitivity to BTH by introduction of an NH1/OsNPR1 paralog. Plant biotechnology journal. 2011;9(2):205–15. 10.1111/j.1467-7652.2010.00544.x . [DOI] [PubMed] [Google Scholar]
- 25.Malnoy M, Jin Q, Borejsza-Wysocka EE, He SY, Aldwinckle HS. Overexpression of the apple MpNPR1 gene confers increased disease resistance in Malus x domestica. Mol Plant Microbe Interact. 2007;20(12):1568–80. 10.1094/MPMI-20-12-1568 . [DOI] [PubMed] [Google Scholar]
- 26.Maier F, Zwicker S, Huckelhoven A, Meissner M, Funk J, Pfitzner AJ, et al. NONEXPRESSOR OF PATHOGENESIS-RELATED PROTEINS1 (NPR1) and some NPR1-related proteins are sensitive to salicylic acid. Mol Plant Pathol. 2011;12(1):73–91. Epub 2010/12/02. 10.1111/j.1364-3703.2010.00653.x . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lawton KA, Friedrich L, Hunt M, Weymann K, Delaney T, Kessmann H, et al. Benzothiadiazole induces disease resistance in Arabidopsis by activation of the systemic acquired resistance signal transduction pathway. Plant J. 1996;10(1):71–82. . [DOI] [PubMed] [Google Scholar]
- 28.Blanco F, Garretón V, Frey N, Dominguez C, Pérez-Acle T, Van der Straeten D, et al. Identification of NPR1-Dependent and Independent Genes Early Induced by Salicylic Acid Treatment in Arabidopsis. Plant Molecular Biology. 2005;59(6):927–44. 10.1007/s11103-005-2227-x [DOI] [PubMed] [Google Scholar]
- 29.Hepworth SR, Zhang Y, McKim S, Li X, Haughn GW. BLADE-ON-PETIOLE-dependent signaling controls leaf and floral patterning in Arabidopsis. Plant Cell. 2005;17(5):1434–48. 10.1105/tpc.104.030536 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kinkema M, Fan W, Dong X. Nuclear localization of NPR1 is required for activation of PR gene expression. Plant Cell. 2000;12(12):2339–50. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Cao H, Li X, Dong X. Generation of broad-spectrum disease resistance by overexpression of an essential regulatory gene in systemic acquired resistance. Proc Natl Acad Sci U S A. 1998;95(11):6531–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Tornero P, Dangl JL. A high throughput method for quantifying growth of phytopathogenic bacteria in Arabidopsis thaliana. Plant J. 2001;28(4):475–81. [DOI] [PubMed] [Google Scholar]
- 33.Ihaka R, Gentleman R. R: A Language for Data Analysis and Graphics. Journal of Computational and Graphical Statistics. 1996;5(3):299–314. [Google Scholar]
- 34.Ciannamea S, Kaufmann K, Frau M, Tonaco IA, Petersen K, Nielsen KK, et al. Protein interactions of MADS box transcription factors involved in flowering in Lolium perenne. J Exp Bot. 2006;57(13):3419–31. Epub 2006/09/29. 10.1093/jxb/erl144 . [DOI] [PubMed] [Google Scholar]
- 35.Vidal M, Legrain P. Yeast forward and reverse ‘n’-hybrid systems. Nucleic Acids Res. 1999;27(4):919–29. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Curtis MD, Grossniklaus U. A gateway cloning vector set for high-throughput functional analysis of genes in planta. Plant Physiol. 2003;133(2):462–9. Epub 2003/10/14. 10.1104/pp.103.027979 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Belda-Palazon B, Ruiz L, Marti E, Tarraga S, Tiburcio AF, Culianez F, et al. Aminopropyltransferases involved in polyamine biosynthesis localize preferentially in the nucleus of plant cells. PLoS One. 2012;7(10):e46907 10.1371/journal.pone.0046907 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002;3(7):RESEARCH0034 Epub 2002/08/20. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kauffmann S, Legrand M, Fritig B. Isolation and characterization of six pathogenesis-related (PR) proteins of Samsun NN tobacco. Plant Mol Biol. 1990;14(3):381–90. . [DOI] [PubMed] [Google Scholar]
- 40.Busso D, Delagoutte-Busso B, Moras D. Construction of a set Gateway-based destination vectors for high-throughput cloning and expression screening in Escherichia coli. Anal Biochem. 2005;343(2):313–21. 10.1016/j.ab.2005.05.015 . [DOI] [PubMed] [Google Scholar]
- 41.Tian M, von Dahl CC, Liu PP, Friso G, van Wijk KJ, Klessig DF. The combined use of photoaffinity labeling and surface plasmon resonance-based technology identifies multiple salicylic acid-binding proteins. Plant J. 2012;72(6):1027–38. 10.1111/tpj.12016 . [DOI] [PubMed] [Google Scholar]
- 42.Breen S, Williams SJ, Outram M, Kobe B, Solomon PS. Emerging Insights into the Functions of Pathogenesis-Related Protein 1. Trends Plant Sci. 2017;22(10):871–9. 10.1016/j.tplants.2017.06.013 . [DOI] [PubMed] [Google Scholar]
- 43.Uquillas C, Letelier I, Blanco F, Jordana X, Holuigue L. NPR1-independent activation of immediate early salicylic acid-responsive genes in Arabidopsis. Mol Plant Microbe Interact. 2004;17(1):34–42. 10.1094/MPMI.2004.17.1.34 . [DOI] [PubMed] [Google Scholar]
- 44.Blanco F, Salinas P, Cecchini NM, Jordana X, Van Hummelen P, Alvarez ME, et al. Early genomic responses to salicylic acid in Arabidopsis. Plant Mol Biol. 2009;70(1–2):79–102. 10.1007/s11103-009-9458-1 . [DOI] [PubMed] [Google Scholar]
- 45.De Leon IP, Sanz A, Hamberg M, Castresana C. Involvement of the Arabidopsis alpha-DOX1 fatty acid dioxygenase in protection against oxidative stress and cell death. Plant J. 2002;29(1):61–2. . [DOI] [PubMed] [Google Scholar]
- 46.Ndamukong I, Abdallat AA, Thurow C, Fode B, Zander M, Weigel R, et al. SA-inducible Arabidopsis glutaredoxin interacts with TGA factors and suppresses JA-responsive PDF1.2 transcription. Plant J. 2007;50(1):128–39. 10.1111/j.1365-313X.2007.03039.x . [DOI] [PubMed] [Google Scholar]
- 47.He Y, Gan S. Identical promoter elements are involved in regulation of the OPR1 gene by senescence and jasmonic acid in Arabidopsis. Plant Mol Biol. 2001;47(5):595–605. . [DOI] [PubMed] [Google Scholar]
- 48.Hong Z, Zhang Z, Olson JM, Verma DP. A novel UDP-glucose transferase is part of the callose synthase complex and interacts with phragmoplastin at the forming cell plate. Plant Cell. 2001;13(4):769–79. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kerppola TK. Visualization of molecular interactions using bimolecular fluorescence complementation analysis: characteristics of protein fragment complementation. Chemical Society reviews. 2009;38(10):2876–86. 10.1039/b909638h . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Zhang Y, Fan W, Kinkema M, Li X, Dong X. Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene. Proc Natl Acad Sci U S A. 1999;96(11):6523–8. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Caufield JH, Sakhawalkar N, Uetz P. A comparison and optimization of yeast two-hybrid systems. Methods (San Diego, Calif. 2012;58(4):317–24. 10.1016/j.ymeth.2012.12.001 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Mou Z, Fan W, Dong X. Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes. Cell. 2003;113(7):935–44. . [DOI] [PubMed] [Google Scholar]
- 53.Manohar M, Tian M, Moreau M, Park SW, Choi HW, Fei Z, et al. Identification of multiple salicylic acid-binding proteins using two high throughput screens. Frontiers in plant science. 2015;5:777 10.3389/fpls.2014.00777 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Winter D, Vinegar B, Nahal H, Ammar R, Wilson GV, Provart NJ. An "Electronic Fluorescent Pictograph" browser for exploring and analyzing large-scale biological data sets. PLoS One. 2007;2(8):e718 10.1371/journal.pone.0000718 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Swarbreck D, Wilks C, Lamesch P, Berardini TZ, Garcia-Hernandez M, Foerster H, et al. The Arabidopsis Information Resource (TAIR): gene structure and function annotation. Nucl Acids Res. 2008;36(suppl_1):D1009–14. 10.1093/nar/gkm965 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Rivas-San Vicente M, Plasencia J. Salicylic acid beyond defence: its role in plant growth and development. J Exp Bot. 2011;62(10):3321–38. Epub 2011/03/02. 10.1093/jxb/err031 . [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(PDF)
(PDF)
(PDF)
(PDF)
(PDF)
(PDF)
(PDF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.