

Abstract
The bloodstream form of the parasite Trypanosoma brucei obtains iron from its mammalian host by receptor-mediated endocytosis of host transferrin through its own unique transferrin receptor (TbTfR). Expression of TbTfR rapidly increases upon iron starvation by post-transcriptional regulation through a currently undefined mechanism that is distinct from the mammalian iron response system. We have created reporter cell lines by fusing the TbTfR 3’UTR or a control Aldolase 3’UTR to reporter genes encoding GFP or firefly Luciferase, and inserted the fusions into a bloodstream form cell line at a tagged ribosomal RNA locus. Fusion of the TbTfR 3’UTR is sufficient to significantly repress the expression of the reporter proteins under normal growth conditions. Under iron starvation conditions we observed upregulation of the mRNA and protein level of the TbTfR 3’UTR fusions only, with a magnitude and timing consistent with that reported for upregulation of the TbTfR. We conclude that the dynamic regulation of the T. brucei transferrin receptor in response to iron starvation is mediated via its 3’UTR, and that the effect is independent of genomic location.
Introduction
The obligate extracellular parasite Trypanosoma brucei has a complex digenetic lifecycle between a tsetse fly vector and a range of mammalian hosts. Mammals limit the availability of essential element iron to prevent growth of invasive pathogens by sequestering iron within the major serum glycoprotein transferrin (Tf). The bloodstream form of T. brucei has evolved a unique transferrin receptor (TbTfR) that allows it to obtain iron through receptor mediated endocytosis of host transferrin (Tf) [1, 2]. After binding of host Tf to the TbTfR in the flagella pocket, endocytosis occurs and subsequent acidification releases free iron for further processing and transportation, whilst the apo-Tf is degraded and the TbTfR is recycled to the flagella pocket [3]. Different TbTfR genes encode proteins with varying affinities for Tf from different mammals, and the occurrence of multiple TbTfR genes has been suggested to allow the parasite to adapt to a wide host range [4, 5].
A subset of T. brucei genes are transcribed by RNA polymerase I (RNA pol I), including the essential Variant Surface Glycoprotein (VSG) which forms a dense surface coat that enables the parasite to evade the host’s innate and adaptive immune responses, and which undergoes antigenic variation from a repertoire of ~1500 VSG genes [6]. Of the 15 subtelomeric VSG bloodstream expression sites (BES), only one is active at a time so that a single VSG is transcribed from a discrete location within the nucleus [7]. Antigenic variation requires replacement of the VSG within the active BES or a switch to a different BES, with the latter also causing a change in the identity of the expression site associated genes (ESAGs) that are expressed, as they are located on the VSG polycistronic transcriptional unit. The VSG promoter proximal genes ESAG6 & ESAG7 form the heterodimeric TbTfR [1], which is evolutionarily distinct from mammalian TfR and structurally resembles a truncated VSG homodimer [8]. Both monomers of TbTfR are extensively N-glycosylated and are membrane associated via the GPI anchor present on ESAG6 [9].
Under basal conditions only 3 × 103 TbTfR heterodimers [1] are expressed despite ESAG6 & 7 being located on the same polycistronic transcriptional unit as the highly abundant VSG (5 × 106 homodimers), demonstrating that transcript abundance and/or processing are differentially regulated. Under iron starvation conditions expression of the TbTfR rapidly increases equally at the mRNA and protein level, with lack of increase in VSG mRNA suggesting that regulation occurs through a currently undefined post-transcriptional mechanism [10, 11]. Reducing the uptake of iron using the iron chelator deferoxamine, culturing with different mammalian serum, incubation with anti-TfR antibodies, or competition with apo-Tf all result in a rapid 2.5–5-fold upregulation of the TbTfR and a corresponding increase in Tf uptake [5, 10–12]. Interestingly, TbTfR upregulation occurs before intracellular iron stores are depleted and cells continue to divide for 48 h, suggesting that cells are responding to changes in iron flux. The mechanism is distinct from the post-transcriptional Iron Response Element (IRE) / Iron Response Protein (IRP) system found in mammals, as knockout of the T. brucei IRP-1 homologue aconitase has no effect on TbTfR regulation [10]. As TbTfR is a multi-gene family that occurs in an atypically regulated locus, and available antibodies cross-react with different ESAG6 & 7 glycoproteins, the direct study of TbTfR regulation is challenging.
The BES that is usually active in T. brucei bloodstream form culture-adapted cell lines is BES1, which expresses ESAG6 & 7 genes with nanomolar affinity for bovine Tf, but that bind to canine Tf only poorly [4, 5]. Prolonged iron starvation (>7 days) induced by changing from growth in media supplemented with bovine serum to canine serum selects for cells that have altered the identity of the expressed ESAG6 & 7 and VSG, either by switching to another BES or replacing the genes in the active BES1 [5, 12]. Switching events can be prevented by supplementing the canine serum with bovine Tf, demonstrating that the adaption is driven by iron starvation [4]. Under normal physiological conditions the concentration of available host Tf is unlikely to limit trypanosome growth, but uptake may become limiting in later stages of an infection when competition with anti-TbTfR antibodies and/or host anaemia come into consideration [12, 13].
There is mounting evidence of the importance of the 3’UTR in the post-transcriptional regulation of developmentally regulated genes in T. brucei. The stage-specific regulation of both RNA pol I transcribed VSG [14] and procyclin [15], and the RNA pol II transcribed COX genes [16] has been demonstrated to occur, at least in part, due to recognition of motifs within their respective 3’UTRs. The developmental regulation of ESAG9 depends on a 34-nucleotide bifunctional element in the 3’UTR that confers both positive and negative regulation [17], and an RNA binding protein that negatively regulates ESAG9 has recently been identified through a genome-wide RNA interference screen [18]. Here, we investigate the importance of the TbTfR 3’UTR in the dynamic regulation of TbTfR in response to iron starvation using a simplified reporter system. By fusing the ESAG6-3’UTR to reporter genes encoding GFP or firefly Luciferase (fLUC), we demonstrate that the 3’UTR alone is sufficient to confer dynamic regulation of gene expression in response to iron starvation.
Material and methods
Cell lines
The culture adapted monomorphic T. brucei brucei Lister 427 bloodstream form 2T1 cell line [19], containing a tagged RRNA locus, were cultured in HMI-11T [20] containing 0.2 μg/mL Puromycin and 0.5 μg/mL Phleomycin at 37 0C in a 5% CO2 incubator. Transfected 2T1 cell lines were selected and maintained with 2.5 μg/mL Hygromycin in place of Puromycin.
Cloning of ESAG6 3’UTR
RNA was extracted from ~1 × 107 logarithmic phase cells using the RNeasy plus kit (Qiagen) according to the manufacturer’s instructions. A two-step RT-PCR reaction was performed by first transcribing 0.25 μg of RNA using an Omniscript Reverse Transcriptase (Promega) with a Oligo-dT adapter primer (5’-CGCGTCGACTAGTACTTTTTTTTTTTTTTTT-3’) and then using 5 μl of the resulting cDNA as a template for a PCR amplification using Hot-start RED-Taq (Sigma) with primers specific for BES1 ESAG6 ORF [7] (5’-GCAGTACATTTGAGTCTT T-3’) and the adapter sequence (5’-CGCGTCGACTAGTAC-3’). The resulting amplicon was ligated into pGEM-T-easy (Promega) prior to DNA sequencing.
Generation of reporter cell lines
The pRPaΔGFP X vector, a version of the pRPaGFP X vector [19] with tetracycline operator removed, was a kind gift from Sam Alsford, LSTHM. The firefly Luciferase (fLUC) ORF was PCR amplified from the pCRm-LUC-HYG vector [21] (a kind gift from Phillip Yates, Oregon Health & Science University, USA) using a mutagenic forward primer incorporating a HindIII site (underlined) to removes an internal ApaI site (mismatch in bold) (5’-ATTATAAGCTTATGGAAGATGCCAAAAACATTAAGAAAGGCCCAGGG-3’) and a reverse primer incorporating a BamHI site (underlined) (5’-TTCGCGGATCCTCACACGGCGATCTTGC-3’). The PCR product was ligated into pGEM-T-easy (Promega) to allow DNA sequencing prior to subsequent subcloning into pRPaΔGFP X using the HindIII and BamHI sites to replace the GFP-stuffer protein fusion, resulting in pRPaΔfLUC that contains the fLUC gene fused to the aldolase 3’UTR.
The 335bp ESAG6 3’UTR was PCR amplified from T. brucei gDNA using a forward primer incorporating XbaI (italics) and BamHI (underlined) sites (5’-AATGATCTAGATAGGGATCCGGGAAGGATGCGAC-3’) and reverse primer incorporating an ApaI (underlined) site (5’-AATAGGGCCCAGTAGAATTAGTCTAGTTT-3’). Digestion with XbaI and ApaI allowed subcloning into pRPaΔGFP X, replacing the stuffer protein X and creating pRPaΔGFP-ESAG6-3’UTR where the GFP gene is fused to ESAG6-3’UTR. Digestion of the same PCR product with BamHI and ApaI allowed subcloning into pRPaΔ-fLUC creating pRPaΔfLUC-ESAG6-3’UTR where the fLUC gene is fused to ESAG6-3’UTR. Finally, the ESAG6-3’UTR of pRPaΔGFP-ESAG6-3’UTR was replaced with the aldolase 3’UTR of pRPaΔGFP X using the BamHI and ApaI sites creating pRPaΔ-GFP that contains the GFP gene fused to the aldolase 3’UTR.
The reporter constructs were digested with AscI, ethanol precipitated and resuspended in water prior to transfection. 3 μg DNA was mixed with 1 × 107 2T1 cells suspended in transfection buffer [22] in a 2 mm cuvette (BioRad) and electroporated in an Amaxa Nucleofector (Lonza Biosciences) using program X-001. Cells were allowed to recover for 6h at 37 0C, 5% CO2 in HMI-11T antibiotics before selection with 2.5 μg/mL Hygromycin and 0.5 μg/mL Phleomycin.
Induction of iron starvation
The iron chelator deferoxamine (Sigma Aldrich) was added to T. brucei cell cultures at 25 μM to induce iron starvation. For serum switching starvation experiments, cells were harvested at 800 × g, washed once in HMI-11T without serum, and resuspended in HMI-11T containing either 10% fetal calf serum (Labtech) or 10% donor dog serum (Labtech) with and without the addition of 200 μg/ml holo bovine transferrin (Sigma).
Western blots
Cells were counted, harvested at 800 × g, the pellet resuspended at 1 × 106 cells/μl in 1 × SDS loading buffer (Melford), and the cells lysed for 5 min at 95 0C. The lysate was separated on a 10% Tris-acetate SDS-PAGE gel, transferred to a PVDF membrane using a Trans blot Turbo semi-dry blotter (Biorad) and blocked overnight in 5% w/w ECL blocking agent (GE Healthcare) in TTBS (0.05% Tween 20 in Tris buffered saline). The expression of GFP was detected with a rabbit anti-GFP primary (1:1000, Roche) and an anti-rabbit-HRP conjugated secondary (1:20,000, GE Healthcare) with expression level quantified with Clarity ECL chemiluminescent detection (Biorad) in a ChemiDoc XPS+ system (Biorad). The detection of tubulin using mouse anti-tubulin KMX-1 primary (1:100; Gift from Keith Gull, Oxford) and anti-mouse-HRP secondary (1:20,000, GE Healthcare) was used to verify equal sample loading, and to normalise the GFP signal.
Luciferase assays
The expression of the fLUC reporter gene was quantified by incubating 100 μl of a 5 × 105 cells/ml T. brucei culture in white 96 well plates with an equal volume the OneGlo luciferase reagent (Promega) for 5 min, with the resulting luminescence detected using a Fluoroskan ascent FL plate reader (Thermo Scientific) with a 10 sec acquisition time per well.
qRT-PCR
Cells were lysed and total RNA extracted using the EZNA total RNA kit (Omega Bio-TEK), treated with DNase I (Thermo Scientific), and reverse transcribed using the iScript cDNA Synthesis Kit (Bio-Rad Laboratories) according to manufacturer’s instructions. qRT-PCR amplification was carried out using FastStart Universal SYBR Green master mix (Roche) on a LightCycler 96 realtime PCR instrument (Roche) using primers for the fLUC (fLUC-F 5’-GACACCGCTATCCTCAGCGT-3’ and fLUC-R 5’- CATGAGCACGACCCGAAAGC -3’) and an Actin control (Actin-F 5’- GTACCACTGGCATTGTTCTCG -3’ and Actin-R 5’-CTTCATGAGATATTCCGTCAGGTC-3’). The cycling parameters were as follows: initial denaturation at 95 0C for 600 secs; 40 cycles at 95 0C for 10 secs and at 60 0C for 30 secs, followed by a melt curve step. Quantification cycle for the mRNA’s of interest were first normalised to an Actin reference mRNA, and then converted to fold change relative to the untreated fLUC-3’ALD signal.
Immunofluorescence microscopy
Approximately 1 × 106 cells were harvested by centrifugation at 800 × g, washed once with 1 × PBS and allowed to adhere to glass slides for 5 min. Slides were incubated with 4% PFA for 5 min at room temperature, washed twice in 1 × PBS for 5 minute. A drop of Fluoroshield with DAPI (Sigma) was added to the slide, a coverslip was applied and the slide examined on a Leica DMRXA2 fluorescent microscope.
Results
Identification of the ESAG6-3’UTR
In order to investigate whether the 3’UTR of the TbTfR is involved in its dynamic regulation it was necessary to first identify the size of the 3’UTR, which due to its telomeric location is not present in the previous polyadenylation site mappings [23, 24]. As expression of ESAG7 may not be essential [7], we focussed our efforts on ESAG6. To identify the size of the ESAG6 3’UTR expressed in the active BES1, a two-step RT-PCR was performed with an oligo-dT adapter primer to transcribe cDNA and subsequent amplification of the ESAG6-3’UTR with primers specific for BES1 ESAG6 open reading frame and the adapter sequence. Sequencing of the resulting amplicons identified that polyadenylation occurred at closely spaced sites 317, 330 or 335 bases downstream of the ESAG6 stop codon. The observed heterogeneity and 3’UTR length are consistent with previous observations in T. brucei [23, 24]. Sequence alignment revealed a high level of sequence conservation between the 13 copies of the ESAG6-3’UTR found in the 15 different BES [7], with pairwise comparison of the nucleotide sequences revealing 91.0–99.7% identity [25] (Fig 1 & Figure A in S1 File). Interestingly the 3’UTR of the putative non-telomeric copy of ESAG6 (Tb927.9.15680) was less conserved, with only 38.9–42.2% sequence identity with the telomeric sequences [25] (Figure B in S1 File).
Fig 1. Identification of the ESAG6-3’UTR.

The observed polyadenylation sites are indicated with triangles above the sequence, and sequence identity with the other 13 BES ESAG6-3’UTR sequences indicated by stars below the sequence. Sequences immediately downstream of the stop codon were aligned with T-Coffee [26].
Expression of reporters with the ESAG6-3’UTR under normal growth conditions
To investigate the potential involvement of the ESAG6-3’UTR in regulation of the TbTfR, constructs were created with the reporter genes GFP or firefly Luciferase (fLUC) ORF followed by either the aldolase (ALD) 3’UTR or the 335 bp ESAG6-3’UTR (Fig 2). The reporter genes were located downstream of a RRNA promoter and integrated into the tagged RRNA locus in the 2T1 cell line to ensure high levels of constitutive transcription by RNA pol I whilst avoiding positional effects [19].
Fig 2. Reporter constructs used in this study.

RNA processing regions and promoters: HygΔStop—portion of the hygromycin resistance targets the Hyg-tagged RRNA locus; PRRNA—RRNA promoter; PPRO—procyclin promoter; PRO—procyclin 5’UTR; ALD—Aldolase 3’UTR; ESAG6—ESAG6-3’UTR; GFP–green fluorescent protein; fLUC–firefly luciferase.
Observation of the GFP reporter cell lines by immunofluorescence microscopy confirmed cytosolic expression of GFP, with the ESAG6-3’UTR reporter cells producing noticeably diminished GFP signal compared to the ALD-3’UTR reporter cells (Fig 3A). The steady state expression level of GFP during exponential growth was quantified by western blotting with anti-GFP antibodies for three independent clones, revealing that expression of GFP in the ESAG6-3’UTR reporter cells was repressed by ~80% compared to the ALD-3’UTR reporter cells (Fig 3B). Similarly, measurement of luciferase activity in the fLUC expressing cell lines revealed a significant difference between the ESAG6-3’UTR and ALD-3’UTR for each of three independent clones, with the fusion of the ESAG6-3’UTR repressing the expression level by ~90% (Fig 3C). The level of expression of TbTfR has previously been reported to increase up to 5-fold at high cell density [27], and we observed similar cell density dependent changes in luciferase activity in the ESAG6-3’UTR fusion (Fig 3D).
Fig 3. Reporter protein expression under normal culture conditions.

A. Immunofluorescence of GFP reporter constructs. B. Quantification of GFP expression by western blotting, using anti-GFP and KMX1 as a loading control. C. Luciferase activity assay, D. Luciferase activity of the ESAG6-3’UTR cell line varies with cell density. Data in panel B-D represents three independent clones of each cell line. Luciferase signal is normalised to the aldolase-3’UTR signal; data point for three biological replicates shown. ALD–aldolase-3’UTR, ESAG6 –ESAG6-3’UTR. * p < 0.05, ** p < 0.001.
The ESAG6 3’UTR mediates an iron starvation response
The addition of the iron chelator deferoxamine to T. brucei cells reduces the availability of Tf-bound iron, and results in a 2.4–3.7-fold increase in TbTfR protein and mRNA levels after 5 hours [10]. Treatment of the two fLUC reporter cell lines with deferoxamine for 5 hours reduced the luciferase activity of the ALD-3’UTR reporter cells by 0.8-fold, but significantly increased the luciferase activity of the ESAG6-3’UTR reporter cells by ~10-fold, so that the luciferase activity was 73 ± 3% of that measured in the untreated ALD-3’UTR reporter cells (Fig 4A). Analysis of the fLUC reporter mRNA by qRT-PCR confirmed that the effect was also evident at the level of mRNA abundance, with the ALD-3’UTR reporter mRNA level reduced (0.9-fold) and that of the ESAG6-3’UTR reporter increased by 3.2-fold so that the later was 83 ± 8% of untreated ALD-3’UTR level (Fig 4B). Although both the protein and mRNA levels of the luciferase in deferoxamine treated ESAG6-3’UTR reporter cells are increased to ~80% of the level in the untreated ALD-3’UTR reporter cells, the discrepancy in the fold-change at mRNA and protein level suggests that the iron starvation response may affect translation efficiency via the ESAG6-3’UTR.
Fig 4. Luciferase expression in the ESAG6-3’UTR reporter increases under iron starvation conditions.
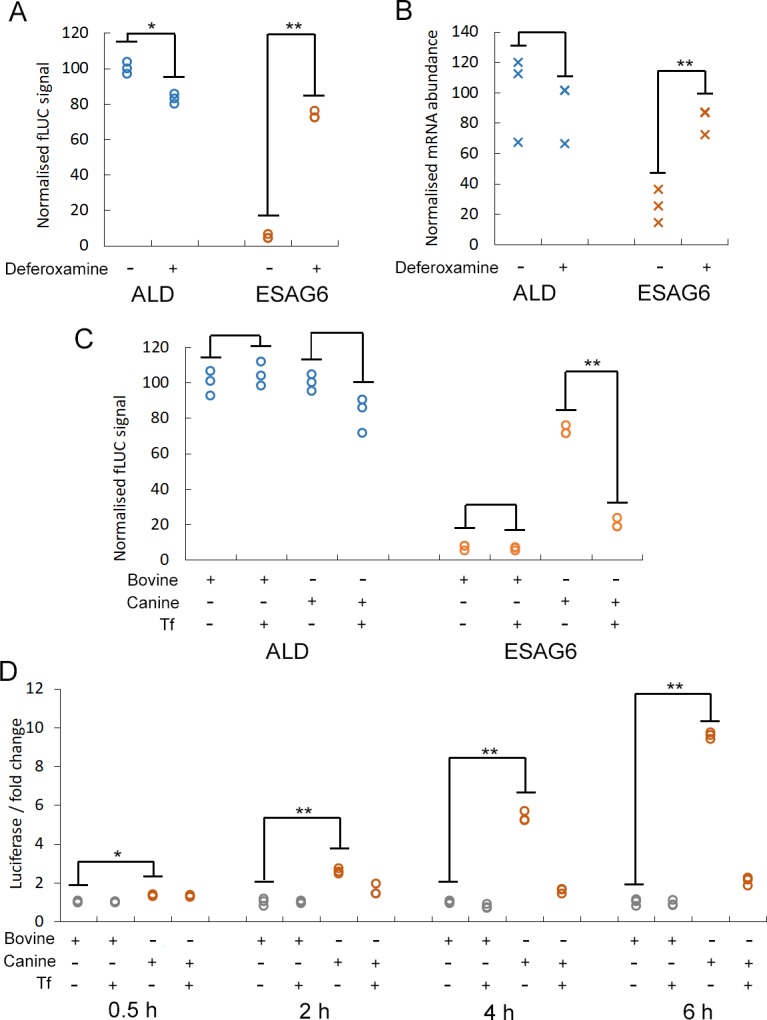
A. Luciferase activity of fLUC reporter cells treated with 25 μM deferoxamine for 5 h, normalised to untreated ALD signal. B. fLUC mRNA levels in reporter cells treated with 25 μM deferoxamine for 5 h, normalised to untreated ALD mRNA level. C. Luciferase activity of fLUC reporter cells switched to HMI11-T + 10% serum (bovine or canine as indicated) for 5 h with or without the addition of 200 μg/ml bovine transferrin (Tf), normalised to untreated ALD signal. D. Time course of luciferase activity of fLUC ESAG6-3’UTR reporter cells switched to HMI11-T + 10% serum (bovine or canine as indicated) with or without the addition of 200 μg/ml bovine Tf, normalised to signal in HMI11-T + 10% FCS. ALD–aldolase 3’UTR, ESAG6 –ESAG6 3’UTR. Data point for three biological replicates shown, * p < 0.05, ** p < 0.001.
Switching T. brucei cells with an active BES1 from growth in media supplemented with 10% bovine serum to growth in media supplemented with 10% canine serum results in rapid 2.5–5-fold upregulation of ESAG6 and ESAG7 at the mRNA and protein level, as the TbTfR expressed from BES1 has a particularly low affinity for canine Tf [4, 10, 11, 27]. The addition of excess bovine Tf to media supplemented with 10% canine serum prevents the upregulation of TbTfR as cells are no longer starved of iron, validating the specificity of the response [4, 5]. Switching the fLUC reporter cell lines to canine-serum-supplemented media for 5 hours increased the luciferase activity of the ESAG6-3’UTR reporter cells by ~3.5-fold compared to growth in canine-serum-supplemented media with excess bovine Tf (Fig 4C). In contrast, the addition of excess bovine Tf to bovine-serum-supplemented media for 5 hours had no significant effect on the luciferase activity of either reporter cell line compared to growth in bovine-serum-supplemented media alone (Fig 4C).
The fLUC ESAG6-3’UTR reporter cell line was used to investigate the temporal profile of the response to serum switching. In the absence of additional bovine Tf, a time-dependent increase in luciferase activity was observed in cells grown in canine-serum-supplemented media compared to cells grown in bovine-serum-supplemented media (Fig 4D), with a significant 1.4-fold increase observed as early as 30 minutes after serum switching and increasing to ~10-fold after 6 hours. Addition of bovine Tf to the canine-serum-supplemented media reduced the magnitude of the increase to ~2-fold after 6 hours but did not completely negate the effect, suggesting that the amount of additional bovine Tf used here is not sufficient to completely restore iron intake.
These data demonstrate that fusion of the ESAG6-3’UTR to a reporter gene is sufficient to confer dynamic regulation of reporter at both the mRNA and protein level in response to iron starvation conditions, and that up-regulation is more rapid than previously thought. The changes in the luciferase activity observed here are broadly consistent with the magnitude of increase in TfR expression previously observed for the iron starvation response, with levels as high as 10-fold reported [27, 28]. The slightly larger changes observed with the fLUC reporter compared TbTfR might be due to the superior quantitation and dynamic range of the bioluminescence assay compared to western blotting with polyclonal anti-TbTfR antibodies.
Discussion
Our data demonstrate that fusion of the ESAG6-3’UTR to reporter genes is sufficient to confer a specific response to iron starvation conditions that increases the mRNA and protein level of the reporter with a magnitude and temporal profile consistent with that previously observed for TbTfR upregulation. Since the reporter gene is located in a RRNA locus, this provides evidence that the dynamic regulation of TbTfR is independent of transcription within the BES site or location at the telomere, and decouples the rapid dynamic regulation of TbTfR from subsequent VSG switching events [4, 5, 12]. To our knowledge, this is the first time that a T. brucei 3’UTR has been demonstrated to be involved in dynamic post-transcriptional regulation of a gene in response to a specific nutritional stimulus, rather than mediated through irreversible developmental regulation in response to lifecycle changes.
We have demonstrated that fusion of the ESAG6-3’-UTR causes the expression of the gene product to be repressed under basal conditions, which is consistent with the low level of ESAG6 mRNA present despite being located on the highly transcribed VSG polycistronic transcriptional unit. Under iron starvation conditions the repressive effect of the ESAG6-3’UTR is reduced, resulting in an increase of expression that is driven at least in part by an increase in transcript abundance, just as reported for TbTfR [10, 11]. Whether the effect is driven by an increase in mRNA stability and/or increased translation efficiency remain to be investigated. The involvement of the 3’UTR in iron starvation response suggest that it contains secondary structural elements or sequence motifs that are recognised by RNA binding protein(s) (RBPs). We performed simultaneous folding and alignment of the 13 ESAG6-3’UTRs to predicted a minimum free energy structure of the consensus sequence [29, 30], which revealed a high level of structure (Figure C in S1 File) suggesting that such features are more extensive than the 16-mer regulatory sequence identified in VSG mRNAs [31]. Further experiments will be required to identify any regulatory features present in the ESAG6-3’UTR, and whether they have a positive or negative regulatory role.
TbTfR is a multi-gene family that occurs in a promoter proximal position in the 15 BES, and Ansorge et al. have previously used RT-PCR analysis to propose that ~20% of ESAG6 mRNA is transcribed from the 14 silent BES due promoter-proximal de-repression of silencing [32]. Recent analysis of the surface proteome in culture adapted monomorphic cell line by Ghadella et al. [33] detected VSGs and ESAGs from multiple BES in addition to the major active BES, which the authors suggest is due to the occurrence of low abundance of a subpopulation of cells that have switched their active BES. Analysis of our own global proteomic data sets [20, 34, 35] supports their conclusion, as although the most abundant ESAGs detected correlate with the BES of the most abundant VSG, several other VSGs from ‘silent’ BES are detected at lower abundance along with their corresponding ESAGs. Therefore we propose that the transcription of TbTfR from supposedly ‘silent’ BES can at least in part be explained by the occurrence of a low abundance of subpopulation of cells that have switched their active BES, and that any contribution of de-repression of silent BES to the rapid dynamic regulation of TbTfR expression is likely to be minimal.
Conclusions
Taken together, the data presented here demonstrate that fusion of the ESAG6-3’UTR to a reporter gene is sufficient to confer a specific response to iron starvation conditions that increases the mRNA and protein level of the reporter with a magnitude and temporal profile consistent with that previously observed for TbTfR upregulation. The effect is independent from transcription within the expression site or location at the telomere, and is therefore decoupled from VSG switching events. The fLUC-ESAG6-3’UTR reporter system is experimentally tractable and provides a simple and reproducible signal-and-response system in cultured monomorphic cells with which to elucidate a signalling pathway essential for the clinically-relevant bloodstream form parasite.
Supporting information
Supporting information containing Figures A–C. Figure A—Sequence alignment of the BES ESAG6 3’UTRs. Figure B—Sequence alignment of the BES and putative genomic ESAG6 3’UTRs. Figure C—Extensive secondary structure is predicted in the ESAG6-3’UTR.
(PDF)
Acknowledgments
We thank Dr Sam Alsford (London School of Hygiene and Tropical Medicine, UK) for providing the pRPaΔGFP X vector, Prof Phillip A. Yates (Oregon Health & Science University, USA) for providing the pCRm-LUC-HYG vector, and Prof Keith Gull (School of Pathology, Oxford, UK) for providing the KMX-1 antibody.
Data Availability
All relevant data are within the manuscript and its Supporting Information file.
Funding Statement
CB and MDU are funded by a New Investigator Research Grant (BB/M009556/1) from the Biotechnology and Biological Sciences Research Council (https://bbsrc.ukri.org/). The funder had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Salmon D, Geuskens M, Hanocq F, Hanocq-Quertier J, Nolan D, Ruben L, et al. A novel heterodimeric transferrin receptor encoded by a pair of VSG expression site-associated genes in Trypanosoma brucei. Cell. 1994;78:75–86. PubMed Central PMCID: PMC8033214. [DOI] [PubMed] [Google Scholar]
- 2.Steverding D, Stierhof YD, Fuchs H, Tauber R, Overath P. Transferrin-binding protein complex is the receptor for transferrin uptake in Trypanosoma brucei. J Cell Biol. 1995;131:1173–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Taylor MC, Kelly JM. Iron metabolism in trypanosomatids, and its crucial role in infection. Parasitology. 2010;137(6):899–917. 10.1017/S0031182009991880 . [DOI] [PubMed] [Google Scholar]
- 4.Bitter W, Gerrits H, Kieft R, Borst P. The role of transferrin-receptor variation in the host range of Trypanosoma brucei. Nature. 1998;391:499–501. 10.1038/35166 [DOI] [PubMed] [Google Scholar]
- 5.Gerrits H, Mußmann R, Bitter W, Kieft R, Borst P. The physiological significance of transferrin receptor variations in Trypanosoma brucei. Mol Biochem Parasitol. 2002;119:237–47. [DOI] [PubMed] [Google Scholar]
- 6.Cross GAM. Antigenic variation in trypanosomes: secrets surface slowly. Bioessays. 1996;18:283–91. 10.1002/bies.950180406 [DOI] [PubMed] [Google Scholar]
- 7.Hertz-Fowler C, Figueiredo LM, Quail MA, Becker M, Jackson A, Bason N, et al. Telomeric expression sites are highly conserved in Trypanosoma brucei. PloS one. 2008;3(10):e3527 10.1371/journal.pone.0003527 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Salmon D, Hanocq-Quertier J, Paturiaux-Hanocq F, Pays A, Tebabi P, Nolan DP, et al. Characterization of the ligand-binding site of the transferrin receptor in Trypanosoma brucei demonstrates a structural relationship with the N-terminal domain of the varient surface glycoprotein. EMBO J. 1997;16:7272–8. 10.1093/emboj/16.24.7272 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mehlert A, Wormald MR, Ferguson MA. Modeling of the N-glycosylated transferrin receptor suggests how transferrin binding can occur within the surface coat of Trypanosoma brucei. PLoS Pathog. 2012;8(4):e1002618 10.1371/journal.ppat.1002618 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fast B, Kremp K, Boshart M, Steverding D. Iron-dependent regulation of transferrin receptor expression in Trypanosoma brucei. Biochem J. 1999;342:691–6. [PMC free article] [PubMed] [Google Scholar]
- 11.Mußmann R, Janssen H, Calafat J, Engstler M, Ansorge I, Clayton C, et al. The expression level determines the surface distribution of the transferrin receptor in Trypanosoma brucei. Mol Microbiol. 2003;47(1):23–35. [DOI] [PubMed] [Google Scholar]
- 12.van Luenen HG, Kieft R, Mussmann R, Engstler M, ter Riet B, Borst P. Trypanosomes change their transferrin receptor expression to allow effective uptake of host transferrin. Mol Microbiol. 2005;58(1):151–65. 10.1111/j.1365-2958.2005.04831.x . [DOI] [PubMed] [Google Scholar]
- 13.Steverding D. The significance of transferrin receptor variation in Trypanosoma brucei. Trends Parasitol. 2003;19(3):125–7. 10.1016/s1471-4922(03)00006-0 [DOI] [PubMed] [Google Scholar]
- 14.Jefferies D, Tebabi P, Pays E. Transient activity assays of the Trypanosoma brucei variant surface glycoprotein gene promoter: control of gene expression at the posttranscriptional level. Mol Cell Biol. 1991;11(1):338–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Roditi I, Furger A, Ruepp S, Schurch N, Butikofer P. Unravelling the procyclin coat of Trypanosoma brucei. Mol Biochem Parasitol. 1998;91:117–30. [DOI] [PubMed] [Google Scholar]
- 16.Mayho M, Fenn K, Craddy P, Crosthwaite S, Matthews K. Post-transcriptional control of nuclear-encoded cytochrome oxidase subunits in Trypanosoma brucei: evidence for genome-wide conservation of life-cycle stage-specific regulatory elements. Nucleic Acids Res. 2006;34(18):5312–24. 10.1093/nar/gkl598 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Monk SL, Simmonds P, Matthews KR. A short bifunctional element operates to positively or negatively regulate ESAG9 expression in different developmental forms of Trypanosoma brucei. J Cell Sci. 2013;126(Pt 10):2294–304. 10.1242/jcs.126011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Rico E, Ivens A, Glover L, Horn D, Matthews KR. Genome-wide RNAi selection identifies a regulator of transmission stage-enriched gene families and cell-type differentiation in Trypanosoma brucei. PLoS Pathog. 2017;13(3):e1006279 10.1371/journal.ppat.1006279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Alsford S, Kawahara T, Glover L, Horn D. Tagging a Trypansoma brucei RRNA locus improves stable transfection efficiency and circumvents inducible expression position effects. Mol Biochem Parasitol. 2005;144(2):142–8. 10.1016/j.molbiopara.2005.08.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Urbaniak MD, Martin DM, Ferguson MA. Global quantitative SILAC phosphoproteomics reveals differential phosphorylation is widespread between the procyclic and bloodstream form lifecycle stages of Trypanosoma brucei. Journal of proteome research. 2013;12(5):2233–44. Epub 2013/03/15. 10.1021/pr400086y ; PubMed Central PMCID: PMC3646404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Soysa R, Carter NS, Yates PA. A dual luciferase system for analysis of post-transcriptional regulation of gene expression in Leishmania. Mol Biochem Parasitol. 2014;195(1):1–5. 10.1016/j.molbiopara.2014.05.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Schumann Burkard G, Jutzi P, Roditi I. Genome-wide RNAi screens in bloodstream form trypanosomes identify drug transporters. Mol Biochem Parasitol. 2011;175(1):91–4. Epub 2010/09/21. 10.1016/j.molbiopara.2010.09.002 . [DOI] [PubMed] [Google Scholar]
- 23.Siegel TN, Hekstra DR, Wang X, Dewell S, Cross GA. Genome-wide analysis of mRNA abundance in two life-cycle stages of Trypanosoma brucei and identification of splicing and polyadenylation sites. Nucleic Acids Res. 2010;38(15):4946–57. 10.1093/nar/gkq237 ; PubMed Central PMCID: PMC2926603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kolev NG, Franklin JB, Carmi S, Shi H, Michaeli S, Tschudi C. The transcriptome of the human pathogen Trypanosoma brucei at single-nucleotide resolution. PLoS Pathog. 2010;6(9):e1001090 Epub 2010/09/15. 10.1371/journal.ppat.1001090 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Stothard P. The Sequence Manipulation Suite: JavaScript programs for analyzing and formatting protein and DNA sequences. BioTechniques. 2010;28:1102–4. PubMed Central PMCID: PMCPMID: 10868275. [DOI] [PubMed] [Google Scholar]
- 26.Notredame C, Higgins D, Heringa J. T-Coffee: A novel method for multiple sequence alignment. J Mol Biol. 2000;302:205–17. 10.1006/jmbi.2000.4042 [DOI] [PubMed] [Google Scholar]
- 27.Mussmann R, Engstler M, Gerrits H, Kieft R, Toaldo CB, Onderwater J, et al. Factors affecting the level and localization of the transferrin receptor in Trypanosoma brucei. J Biol Chem. 2004;279(39):40690–8. 10.1074/jbc.M404697200 . [DOI] [PubMed] [Google Scholar]
- 28.Schwartz KJ, Peck RF, Tazeh NN, Bangs JD. GPI valence and the fate of secretory membrane proteins in African trypanosomes. J Cell Sci. 2005;118(Pt 23):5499–511. Epub 2005/11/18. 10.1242/jcs.02667 . [DOI] [PubMed] [Google Scholar]
- 29.Will S, Joshi T, Hofacker IL, Stadler PF, Backofen R. LocARNA-P: accurate boundary prediction and improved detection of structural RNAs. RNA. 2012;18(5):900–14. Epub 2012/03/28. 10.1261/rna.029041.111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lai D, Proctor JR, Zhu JY, Meyer IM. R-CHIE: a web server and R package for visualizing RNA secondary structures. Nucleic Acids Res. 2012;40(12):e95 Epub 2012/03/22. 10.1093/nar/gks241 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ridewood S, Ooi CP, Hall B, Trenaman A, Wand NV, Sioutas G, et al. The role of genomic location and flanking 3'UTR in the generation of functional levels of variant surface glycoprotein in Trypanosoma brucei. Mol Microbiol. 2017;106(4):614–34. Epub 2017/09/15. 10.1111/mmi.13838 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ansorge I, Steverding D, Melville S, Hartmann C, Clayton C. Transcription of ‘inactive’ expression sites in African trypanosomes leads to expression of multiple transferrin receptor RNAs in bloodstream forms. Mol Biochem Parasitol. 1999;101:81–94. [DOI] [PubMed] [Google Scholar]
- 33.Gadelha C, Zhang W, Chamberlain JW, Chait BT, Wickstead B, Field MC. Architecture of a Host-Parasite Interface: Complex Targeting Mechanisms Revealed Through Proteomics. Mol Cell Proteomics. 2015;14(7):1911–26. Epub 2015/05/02. 10.1074/mcp.M114.047647 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Urbaniak MD, Guther MLS, Ferguson MAJ. Comparative SILAC Proteomic Analysis of Trypanosoma brucei Bloodstream and Procyclic Lifecycle Stages. PloS one. 2012;7(5):e36619 10.1371/journal.pone.0036619 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Domingo-Sananes MR, Szoor B, Ferguson MA, Urbaniak MD, Matthews KR. Molecular control of irreversible bistability during trypanosome developmental commitment. J Cell Biol. 2015;211(2):455–68. 10.1083/jcb.201506114 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supporting information containing Figures A–C. Figure A—Sequence alignment of the BES ESAG6 3’UTRs. Figure B—Sequence alignment of the BES and putative genomic ESAG6 3’UTRs. Figure C—Extensive secondary structure is predicted in the ESAG6-3’UTR.
(PDF)
Data Availability Statement
All relevant data are within the manuscript and its Supporting Information file.