

Abstract
Alcoholic liver disease is the result of cascade events, which clinically first lead to alcoholic fatty liver, and then mostly via alcoholic steatohepatitis or alcoholic hepatitis potentially to cirrhosis and hepatocellular carcinoma. Pathogenetic events are linked to the metabolism of ethanol and acetaldehyde as its first oxidation product generated via hepatic alcohol dehydrogenase (ADH) and the microsomal ethanol-oxidizing system (MEOS), which depends on cytochrome P450 2E1 (CYP 2E1), and is inducible by chronic alcohol use. MEOS induction accelerates the metabolism of ethanol to acetaldehyde that facilitates organ injury including the liver, and it produces via CYP 2E1 many reactive oxygen species (ROS) such as ethoxy radical, hydroxyethyl radical, acetyl radical, singlet radical, superoxide radical, hydrogen peroxide, hydroxyl radical, alkoxyl radical, and peroxyl radical. These attack hepatocytes, Kupffer cells, stellate cells, and liver sinusoidal endothelial cells, and their signaling mediators such as interleukins, interferons, and growth factors, help to initiate liver injury including fibrosis and cirrhosis in susceptible individuals with specific risk factors. Through CYP 2E1-dependent ROS, more evidence is emerging that alcohol generates lipid peroxides and modifies the intestinal microbiome, thereby stimulating actions of endotoxins produced by intestinal bacteria; lipid peroxides and endotoxins are potential causes that are involved in alcoholic liver injury. Alcohol modifies SIRT1 (Sirtuin-1; derived from Silent mating type Information Regulation) and SIRT2, and most importantly, the innate and adapted immune systems, which may explain the individual differences of injury susceptibility. Metabolic pathways are also influenced by circadian rhythms, specific conditions known from living organisms including plants. Open for discussion is a 5-hit working hypothesis, attempting to define key elements involved in injury progression. In essence, although abundant biochemical mechanisms are proposed for the initiation and perpetuation of liver injury, patients with an alcohol problem benefit from permanent alcohol abstinence alone.
Keywords: MEOS, microsomal ethanol-oxidizing system, alcohol dehydrogenase, ethanol, acetaldehyde, alcohol metabolism, alcoholic liver disease, alcoholic fatty liver, alcoholic steatohepatitis, alcoholic hepatitis, alcoholic cirrhosis, hepatocellular carcinoma, circadian rhythms, CYP 2E1, endotoxins, intestinal microbiome, SIRT, ROS
1. Introduction
Alcohol is chemically ethyl alcohol, or in condensed form described as ethanol, whereby these terms are often used interchangeably in the clinical context. Ethanol, a short-chain hydrocarbon C2H5OH, has a more non-polar chemical structure, and it is thereby water soluble, but less soluble in lipids with their polar molecules. This ambiguity of polarity facilitates a rapid diffusion of ethanol through biological membranes, allowing at the same time changes in the membrane properties, whereas the mechanism of action remains controversial [1,2]. Ethanol may target the bilayer structures of outer cell membranes or the monolayer membranes of organelles such as mitochondria or the endoplasmic reticulum, considering that both membrane types contain lipids preferentially as phospholipids. There is also evidence for direct alcohol interactions with membrane proteins [3]. This may have an additional impact on functional and injurious changes in organs that are affected by chronic alcohol consumption. Therefore, alcohol related organ injuries and functional alterations could unquestionably be classified as molecular, membrane-targeted diseases, a definition that is well applicable also to alcoholic liver disease (ALD).
Among various liver diseases such as drug-induced liver injury (DILI), herb-induced liver injury (HILI), nonalcoholic fatty liver disease (NAFLD), nonalcoholic steatohepatitis (NASH), or those caused by hepatitis B virus (HBV) and hepatitis C virus (HCV) infections, especially ALD continues to attract much interest from scientists and clinicians worldwide [4,5,6,7,8,9,10,11,12,13,14,15]. Their stimulating and partially controversial discussions focused on the pathogenetic aspects [4,5], clinical features [6], and therapeutic approaches [7,8,9] including liver transplantation [10,11,12,13]. Consensus exists that ALD contributes significantly to the global burden of mortality [14,15]. In 2010, alcoholic cirrhosis (AC) was globally responsible for around 493,300 deaths, corresponding to around 0.9% of all global deaths [15].
This review article provides an update on a few relevant issues of alcohol and acetaldehyde metabolism, molecular and cellular mechanisms leading to liver injury, clinical features, and diagnostic approaches, as well as therapeutic modalities, including the option for liver transplantation. It is well recognized that there exist myriads of alcoholic studies with several hundreds of molecular mechanisms, and many mediators are provided by expert biochemists rather than a satisfactory unifying mechanistic approach that is considered valid for a sound discussion of a yet partially controversial clinical topic.
2. Data Search and Source
The PubMed database was used to identify publications for the following terms: Alcoholic liver disease, alcoholic fatty liver disease, alcoholic steatohepatitis, alcoholic hepatitis, alcoholic cirrhosis, alcohol metabolism, alcohol dehydrogenase, microsomal ethanol-oxidizing system, cytochrome P450 2E1 (CYP 2E1), catalase, and mitochondrial acetaldehyde dehydrogenase. Publications of the first 50 hits from each searched segment were analyzed. The search was completed on 24 September 2018. Prior to the final analysis, the publications were assessed regarding clinical quality and data completeness. The final selection of publications was restricted to those in English language to ensure transparent accessibility.
3. Alcohol Absorption
Alcoholic beverages are commonly nature-based products, initially involving photosynthesis in the plants [16,17]. Glucose is produced, a chemical ingredient of many fruits such as grapes, used as source to produced wine, a process known in many countries since ancient times. During wine production and fermentation, glucose is converted to alcohol. Following ingestion, maximum blood alcohol levels are higher if alcohol is consumed with an empty stomach [18], if highly concentrated alcoholic beverages such as spirits were used within a short time without a meal [19], or if patients had a past medical history (PMH) of gastrectomy surgery causing rapid intestinal alcohol uptake [20].
4. Gastric Alcohol Dehydrogenase and First-Pass Metabolism of Alcohol
Although the liver is the preferred organ of alcohol degradation, it is metabolized in small amounts by the gastric mucosa, which contains alcohol dehydrogenase (ADH), a process called gastric first-pass metabolism (FPM) [20,21,22].
5. Hepatic Alcohol Metabolism
In the liver, three different enzymes are known, which in vitro can metabolize ethanol to acetaldehyde [22,23,24,25]. These are ADH, the microsomal ethanol-oxidizing system (MEOS), and catalase:
| ADH: C2H5OH + NAD+ → CH3CHO + NADH + H+ Ethanol Acetaldehyde |
| MEOS: C2H5OH + NADPH + H+ + O2 → CH3CHO + NADP+ + 2H2O Ethanol Acetaldehyde |
| Catalase: C2H5OH + H2O2 → CH3CHO + 2H2O Ethanol Acetaldehyde |
Within the liver cell, ADH is found in the cytoplasm (cytosol) between the various subcellular structures, MEOS in the endoplasmic reticulum, and catalase in the peroxisomes, also called microbodies. Of relevance for hepatic alcohol metabolism in vivo are ADH and MEOS but not catalase (Figure 1) [22,23,24,25].
Figure 1.

Significant pathways of hepatic alcohol and acetaldehyde metabolism. For alcohol metabolism, presented are cytosolic alcohol dehydrogenase (ADH) and the microsomal ethanol-oxidizing system (MEOS); both pathways metabolize ethanol to acetaldehyde. Reproduced from a previous report [25], with permission of the Publisher Taylor & Francis (Didcot, UK).
6. Hepatic Alcohol Dehydrogenase
Hepatic ADH (EC 1.1.1.1) is a well-studied enzyme of the hepatocyte cytosol. Several aspects of hepatic ADH have been discussed in various reports [22,23,24,25]. The mammalian ADHs represent a group of enzymes that catalyze the oxidation and reduction of a wide variety of alcohols and aldehydes, with individual differences in ADH isozymes.
Human liver ADH consists of five classes, ADH1 through ADH5, characterized by individual subunits. For class I, the subunits α, β, and γ are described [22,24]. Polymorphism is a characteristic feature of ADH2 and ADH3 that encode the β and γ subunits. ADH2 isoenzymes migrate more anodically than ADH1 isoenzymes, and have a relatively high Km for ethanol (34 mM). ADH can easily be differentiated form the other enzymes metabolizing alcohol, MEOS, and catalase (Table 1).
Table 1.
Differentiation between ADH, MEOS, and catalase. Symbols for effects: (+), marginal; + low; ++, moderate; +++, high; ++++, very high. Abbreviations: ADH, alcohol dehydrogenase; Km, Michaelis-Menten constant; MEOS, microsomal ethanol-oxidizing system; N.D., not determined. The original table was published in a previous report [25] and is reproduced with permission of the Publisher Taylor & Francis (Didcot, UK).
| Characteristics | ADH | MEOS | Catalase |
|---|---|---|---|
| Intracellular localization | Cytosol | Endoplasmic reticulum | Peroxisomes |
| Co-factor | NAD+ | NADPH + H+ | N.D. |
| Co-substrate | None | Molecular oxygen | H2O2 |
| Reaction products | Acetaldehyde NADH + H+ |
Acetaldehyde NADP+, H2O |
Acetaldehyde H2O |
| Kinetics | |||
| Km (ethanol) | 0.5–2.0 mM | 7–11 mM | 0.6–10 mM |
| Km (O2) | N.D. | 8.3 μM | 50 μM |
| pH optimum | 11 | 6.9–7.5 | 5.5 |
| Inhibitory effect | |||
| Pyrazole (0.1 mM) | ++++ | 0 | (+) |
| Cyanide (0.1 mM) | N.D. | 0 | ++++ |
| Azide (0.1 mM) | 0 | 0 | ++++ |
| Substrate specificity | |||
| Methanol | ++ | ++ | ++++ |
| Ethanol | +++ | ++++ | ++++ |
| n-Propanol | ++++ | +++ | (+) |
| n-Butanol | ++++ | ++ | 0 |
| n-Pentanol | ++++ | + | 0 |
| i-Propanol | + | + | 0 |
| t-Butanol | 0 | + | 0 |
| Increase in activity following chronic ethanol consumption | 0 | ++++ | 0 |
| Enzyme isolation | + | + | + |
| Isoenzymes | + | + | + |
In animal studies, ADH was characterized and differentiated from MEOS and catalase [22,23,25,26,27,28,29]. ADH is active at a pH optimum of 11, and it has a Km for ethanol of 0.5–2.0 mM, corresponding to 0.025–0.1‰ ethanol. The high pH optimum of ADH in animals is substantially above the physiological pH of 7.4, conditions that may limit the role of hepatic ADH in experimental studies of alcohol metabolism. However, its low Km values for ethanol are theoretically in favor of a major role of ADH in experimental hepatic alcohol metabolism, but only at low and not at higher alcohol concentrations. Since hepatic ADH activity is not inducible after chronic alcohol use, this enzyme cannot account for the increased alcohol metabolism observed after prolonged alcohol intake. Estimates of the role of hepatic ADH in alcohol metabolism of non-alcoholic humans will remain a matter of debate, due to the variability of genetic ADH isoenzymes as confounding variables.
7. Hepatic Microsomal Ethanol-Oxidizing System
The discovery of MEOS by the pioneering scientists Charles S. Lieber and Leonore M. DeCarli was published in 1968 [26], followed by their detailed characterization of MEOS, reported in 1970 [27,28], and its tentative role for ethanol metabolism in vivo, published in 1972 [29]. Subsequent publications focused on this new pathway of alcohol metabolism and related clinical aspects [30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150,151,152,153,154,155,156,157,158,159,160,161,162,163,164,165,166,167,168,169,170,171,172,173,174,175,176,177,178,179,180,181,182,183,184,185,186,187,188,189,190,191,192,193,194,195,196,197,198,199,200,201,202,203,204,205,206,207,208,209,210,211,212,213,214,215,216,217,218,219,220,221,222,223,224,225,226,227,228,229,230,231,232,233,234,235,236,237,238,239,240,241,242,243]. In addition, various review articles discussed new developments of MEOS, including its constituents such as cytochrome P450 (CYP), or more specifically, its isoenzyme, CYP 2E1 [23,25,41,45,52,53,54,55,75,102,234,244,245,246,247,248,249,250,251,252]. The discovery of MEOS [26,27,248,250] goes back to an ultrastructural study, which revealed a striking centrilobular and midzonal proliferation of the smooth endoplasmic reticulum in the livers of rats after consumption of a liquid diet containing ethanol [253]. These ultrastructural changes were associated with a typical alcoholic fatty liver [253], and could be reproduced in volunteers given alcohol under controlled metabolic ward conditions [254]. Such careful observations in experimental and human studies finally facilitated the discovery of MEOS [26,27,28,29,250].
The nature of the newly described MEOS was heavily debated. In particular, there were initial claims of others that this system may be accounted for by enzymes contaminating the liver microsomal fraction during the preparative procedures. The specific critical focus was on ADH, catalase, or both together as possible causative contaminants. However, yet the initially described characteristics of MEOS were clearly different from those of ADH and catalase [26,27,28], with various differentiating features as listed in detail (Table 1). Such possible contaminants had early been considered in the initial publications describing MEOS as a unique enzyme system, but their role in MEOS was clarified using specific inhibitors of ADH and catalase, which showed that MEOS activities remained virtually unchanged under these inhibitory conditions [26,27].
A step forward was the physical separation of MEOS in rat liver from both ADH and catalase activities by DEAE (Diethyl-Amino-Ethyl) cellulose ion exchange column chromatography [30]. MEOS likely consists of cytochrome P450 (CYP), NADPH-cytochrome P450 reductase, and phospholipids (Figure 2).
Figure 2.

Purification of the microsomal ethanol-oxidizing system (MEOS) and its separation from catalase and alcohol dehydrogenase (ADH) activities. Separation was achieved by DEAE (Diethyl-Amino-Ethyl) cellulose ion exchange column chromatography after solubilization of liver microsomes obtained from rats fed an ethanol containing liquid diet for three weeks. In the void volume eluted up to around 220 mL, the highest peak represents the protein curve assessed as E280 nm, and the peak below that is the catalase peak, whereas ADH presents as the lowest peak. Starting with an elution volume of around 330 mL, microsomal components begin to appear. The first peak represents cytochrome P450, the second peak represents E280 nm, followed by a third peak with two shoulders and by a fourth peak representing MEOS. At around 770 mL, the reductase peak emerges, followed by the phospholipid peak at around 790 mL elution volume. Overall, this experimental approach was challenging, putting active MEOS on the top of the column and expecting active MEOS in the effluents. There was a high risk of inactivation of MEOS, not only during the solubilization procedure using ultrasonication and deoxycholate that disintegrated MEOS out of the intact microsomal membranes, but also during the chromatography procedure itself that could lead to the inactive cytochrome P420 from the active P450. The original figure was published in a previous report [30] and is reproduced with permission of the Publisher Elsevier (Amsterdam, The Netherlands).
Based on the observation that MEOS activity was found only in column fractions rich in CYP, this suggested that MEOS is an enzyme system that depends strongly on cytochrome P450 (Figure 3) [30].
Figure 3.

Constituents of MEOS. A key role is attributed to the hepatic microsomal cytochrome P450 2E1, but NADPH-cytochrome P450 reductase plays also an obligatory role; the metabolic reaction of MEOS requires phospholipids, the site of their reaction is unknown. Reproduced from a previous report [25], with permission of the Publisher Taylor & Francis (Didcot, UK).
Of note, the previously described and partially purified ethanol-inducible cyanide binding cytochrome [51,255,256] is presumably identical to the later described ethanol-inducible CYP 2E1 as the most important isoenzyme (Table 2) [192,245,246,247,248,249,250].
Table 2.
Microsomal ethanol-oxidizing system (MEOS) and its cytochrome P450 isoenzymes. To assess the turnover number, MEOS activity (nmoles acetaldehyde/min) is calculated per nmole cytochrome P450, all expressed per mg of microsomal protein as published [192].
| Cytochrome P450 Isoenzyme | MEOS Activity/Cytochrome P450 |
|---|---|
| 1A2 | 10.90 |
| 2A6 | 3.75 |
| 2B6 | 2.89 |
| 2D6 | 0.70 |
| 2E1 | 11.51 |
| 3A4 | 3.38 |
Reconstitution experiments supported the concept that MEOS consists of cytochrome P450, with preference of its isoenzyme CYP 2E1, the reductase, and phospholipids [49,52,55,57]. Other investigators reproduced our initial separation studies [78] published before [30] and thereby confirmed that MEOS is indeed an enzyme system that is independent of ADH and catalase [57,78]. Finally, CYP 2E1 was purified and characterized in human liver as a hemeprotein with a molecular weight of 54 daltons, which oxidizes ethanol at a turnover rate of 12.2 nmole min−1 in a reconstituted system [110]. The capacity of the ethanol-inducible CYP 2E1 to generate reactive oxygen species (ROS) such as superoxide and hydroxyl radicals has been published in several reports [50,56,92,152]. In analogy to many drugs and chemicals, ethanol functions as a substrate for microsomal CYP through an oxidation process, which may partially be incomplete and thereby produce reactive oxygen forms (Figure 4) [257].
Figure 4.

Hepatic microsomal cytochrome P450 and its interaction with substrates. Cytochrome P450 catalyzes the oxidation of substrates such as drugs and ethanol, which bind to the ferric (3+) iron of the cytochrome P450 as the initial metabolic step leading finally to the oxidized substrate. The original figure was published in a recent article [257].
The reactions within the CYP cycle involve the uptake of two electrons and molecular oxygen, yielding the oxidized substrate plus water (Figure 4). The iron in the cytochrome P450 molecule returns to its ferric (3+) state, after having been in a ferrous state (2+), and is again available for binding to the next substrate molecule. AS part of usual life in any healthy organism, radicals are continuously generated through incomplete intracellular oxygen splitting, and are commonly scavenged by antioxidants.
Whereas MEOS activity is induced by chronic alcohol consumption as established by several studies [26,27,28,29,30], even a single dose of ethanol can increase MEOS activity [76]. The induction of MEOS activity was found to be modulated by the composition of the alcohol diets containing various amounts of fat or carbohydrates [40,68,137]. For instance, the induction of MEOS activity was facilitated by a high-fat diet [40] or a low-carbohydrate diet [68]. However, under an alcohol diet with a high amount of carbohydrates the usual alcohol-dependent induction of MEOS activity was offset [68]. Similar to MEOS, hepatic microsomal CYP 2E1 is regulated by dietary lipids and carbohydrates [137]. There is now also sufficient evidence that the induction of the ethanol-specific CYP 2E1 is the result of increased enzyme synthesis [153], found in the course of prolonged ethanol use, and explained via transcription of the CYP 2E1 gene at high alcohol values in the blood [156].
Based on animal studies, it was early recognized that in addition to ADH, a portion of alcohol would be metabolized by a pathway that is independent of ADH, likely by MEOS, which could account for the higher fraction ranging from 20% up to 25% of the alcohol metabolism in vivo [29]. The percentage contribution of MEOS in alcohol metabolism will be increased at higher alcohol levels, commonly achieved for instance, during social drinking, and it will adaptively be enhanced following chronic alcohol use. With respect to MEOS, its high Km value for ethanol (Table 1) [26,27,38,42] favors the role of MEOS at higher alcohol concentrations [22,23,245]. MEOS inducibility by chronic alcohol use (Table 1) [26,27,30] underscores its importance in removing alcohol more quickly under conditions of preexisting and long lasting alcohol consumption, considering that MEOS may contribute >25% of overall hepatic alcohol metabolism [245,246,247]. It has also been suggested that when corrected for microsomal losses during preparation, half to two thirds of the increase in the rate of ethanol oxidation after chronic alcohol use can be accounted for by MEOS [29]. The existence of an ADH-independent pathway such as MEOS was confirmed by a variety of subsequent experimental studies and evaluations in humans [18,48,86,87,245], reinforcing its role at high alcohol concentrations and after chronic alcohol use [22,23,45,48,244,245,246,247,249,250,258,259,260,261,262,263,264].
Assessing the quantitative role of MEOS in alcohol metabolism is a particular issue, due to confounding factors such as the genetic isoenzyme variabilities of hepatic enzymes that are involved in hepatic alcohol metabolism. Other uncertainties relate to the extent of metabolic interactions between ADH and MEOS at the level of reducing equivalents produced by ADH and consumed by MEOS (Table 1). In more detail, MEOS and ADH may promote hepatic alcohol metabolism in joint action, because ethanol oxidation via ADH requires NAD+, and it generates with NADH + H+ reducing equivalents, which are welcome to MEOS, which requires reducing equivalents in form of NADPH + H+ (Figure 5).
Figure 5.

Interconnected action of hepatic alcohol dehydrogenase (ADH) and the microsomal ethanol-oxidizing system (MEOS). ADH produces reducing equivalents that are used by MEOS, showing that both enzymes depend on each other. The original figure was published in an earlier report [25], reproduced with permission of the Publisher Taylor & Francis (Didcot, UK).
Due to the abundancy of publications retrieved from the database of PubMed and restriction of space and references, the selection will inevitably not cover all important aspects, but reports not references are not necessarily of lower quality. For a quick overview the following publications are listed (Table 3) [26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150,151,152,153,154,155,156,157,158,159,160,161,162,163,164,165,166,167,168,169,170,171,172,173,174,175,176,177,178,179,180,181,182,183,184,185,186,187,188,189,190,191,192,193,194,195,196,197,198,199,200,201,202,203,204,205,206,207,208,209,210,211,212,213,214,215,216,217,218,219,220,221,222,223,224,225,226,227,228,229,230,231,232,233,234,235,236,237,238,239,240,241,242,243].
Table 3.
Listing of relevant reports addressing issues of MEOS, CYP 2E1, and related aspects.
| Year | In Short: Selected Details of MEOS, CYP2E1, and Related Aspects | Authors |
|---|---|---|
| 1968 | Discovery of ethanol oxidation by rat liver microsomal enzymes, now called the hepatic microsomal ethanol-oxidizing system (MEOS), which was different from ADH and catalase, using specific inhibitors, and induction by chronic alcohol consumption | Lieber and DeCarli [26] |
| 1970 | Detailed characterization of MEOS | Lieber and DeCarli [27] |
| 1970 | Dissociation of MEOS from NADPH oxidase | Lieber and DeCarli [28] |
| 1972 | Role of MEOS for ethanol metabolism in vivo | Lieber and DeCarli [29] |
| 1972 | Solubilization and purification of MEOS, and its separation from ADH and catalase by DEAE-cellulose ion exchange column chromatography, with the identification of CYP 450, reductase, and phospholipids as components of MEOS | Teschke et al. [30] |
| 1973 | Presence of induced MEOS in hepatic smooth and rough microsomes | Ishii et al. [31] |
| 1973 | Induced NADPH-cytochrome P450 reductase in hepatic smooth and rough microsomes | Joly et al. [32] |
| 1973 | Liver microsomal glycerophosphate acyltransferase activity following prolonged alcohol use | Joly et al. [33] |
| 1973 | Increased activity of glucose-6-phosphatase in liver microsomes due to prolonged alcohol consumption | Ishii et al. [34] |
| 1973 | A component of hepatic microsomes that is rich in CYP oxidizes ethanol | Mezey et al. [35] |
| 1974 | Broad substrate specificity of the microsomal alcohol-oxidizing system (MAOS) for methanol, ethanol, n-propyl alcohol, and n-butyl alcohol, inducible by chronic ethanol consumption | Teschke et al. [36] |
| 1974 | MEOS in acatalasemic mice | Lieber and DeCarli [37] |
| 1974 | Characterization of the solubilized, isolated, and purified MEOS | Teschke et al. [38] |
| 1974 | Enhanced liver injury by carbon tetrachloride after chronic ethanol use: its mechanism | Hasumura et al. [39] |
| 1975 | Role of dietary fat and caloric intake for the induction of MEOS by prolonged ethanol use | Joly and Hétu [40] |
| 1975 | Alteration of acetaldehyde metabolism after prolonged use of ethanol | Lieber et al. [41] |
| 1975 | Detailed description of the microsomal system oxidizing methanol, ethanol, n-propyl alcohol, and n-butyl alcohol as substrates | Teschke et al. [42] |
| 1975 | Isolation of the microsomal alcohol-oxidizing system with methanol, ethanol, n-propyl alcohol, n-butyl alcohol, and n-pentanol in normal and acatalasemic mice | Teschke et al. [43] |
| 1975 | Inhibition of the ethanol-induced cytochrome P450 by tetrahydrofurane | Ullrich et al. [44] |
| 1975 | Ethanol and acetaldehyde metabolism influenced by chronic alcohol use | Lieber et al. [45] |
| 1975 | Chronic alcohol consumption decreases acetaldehyde oxidation in liver mitochondria | Hasumura et al. [46] |
| 1976 | Characteristics of acetaldehyde oxidation in rat liver mitochondria | Hasumura et al. [47] |
| 1976 | Role of MEOS for ethanol metabolism in liver slices, using also n-butyl alcohol as inhibitor | Teschke et al. [48] |
| 1977 | Isolation and reconstitution of MEOS, with substrate specificity of the partially purified ethanol-induced CYP 2E1 for ethanol, n-propyl alcohol, and n-butyl alcohol, and characterization of the reconstituted MEOS | Ohnishi and Lieber [49] |
| 1977 | Involvement of hydroxyl radicals in MEOS | Cederbaum et al. [50] |
| 1977 | Spectral and catalytic properties of an ethanol-induced form of cytochrome P450 | Joly et al. [51] |
| 1977 | Details of MEOS isolation and reconstitution | Teschke et al. [52] |
| 1977 | Current status of MEOS characterization | Teschke et al. [53] |
| 1977 | Biochemical nature and role of MEOS | Teschke et al. [54] |
| 1977 | MEOS described in Methods in Enzymology | Lieber et al. [55] |
| 1978 | Role of superoxide and hydroxyl radicals in MEOS | Ohnishi and Lieber [56] |
| 1978 | Reconstitution of MEOS with highly purified microsomal cytochrome P450, reductase, and phospholipids, free of catalase and ADH | Miwa et al. [57] |
| 1978 | Photochemical action spectrum of MEOS | Fabry and Lieber [58] |
| 1979 | Induction of intestinal MEOS by chronic ethanol administration | Seitz et al. [59] |
| 1979 | Induction of MEOS by thyroid hormones | Moreno et al. [60] |
| 1979 | Prolonged ethanol use augments liver injury due to paracetamol (acetaminophen) | Teschke et al. [61] |
| 1980 | Enhanced chlorpromazine-induced cholestasis following chronic alcohol use | Teschke et al. [62] |
| 1980 | Existence and role of MEOS in deermice genetically lacking ADH | Burnett and Felder [63] |
| 1980 | Prolonged ethanol use ameliorates liver injury due to dimethylnitrosamine (DMN) | Gellert et al. [64] |
| 1980 | Oxidative demethylation of t-butyl alcohol in rat liver microsomes | Cederbaum and Cohen [65] |
| 1980 | Thyroid hormones induce MEOS activity and reduce ADH activity in rat liver | Moreno et al. [66] |
| 1981 | Microsomal system oxidizing isopropyl alcohol | Cederbaum et al. [67] |
| 1981 | Respective role of ethanol and carbohydrates for the induction of MEOS | Teschke et al. [68] |
| 1981 | Induction of pulmonary MEOS by chronic ethanol consumption | Seitz et al. [69] |
| 1981 | Induction of MEOS by propylthiouracil | Moreno et al. [70] |
| 1981 | Prolonged alcohol use potentiates experimental liver injury caused by paracetamol | Sato et al. [71] |
| 1982 | Liver enzymes metabolizing ethanol are altered in male rats treated by sex hormones | Teschke and Heymann [72] |
| 1982 | Purification and characterization of the ethanol-specific CYP 2E1 in rabbits metabolizing ethanol and aniline | Koop et al. [73] |
| 1982 | Substrate specificity of the purified ethanol-induced cytochrome P450 for methanol, ethanol, n-propyl alcohol, n-butyl alcohol, and aniline in rabbits | Morgan et al. [74] |
| 1982 | Induction of the ethanol-specific CYP 2E1 by benzene in rabbits | Ingelman-Sundberg and Hagbjörk [75] |
| 1982 | Increase of MEOS by a single dose of ethanol | Petersen et al. [76] |
| 1982 | Induction of MEOS by testosterone | Teschke and Wiese [77] |
| 1982 | Description of the isolated MEOS by electron microscopy and confirmation by method reproduction of the previous description of MEOS regarding its microsomal constituents and independency of ADH and catalase | Damgaard [78] |
| 1982 | Decreased hepatic vitamin A levels in patients with ALD | Leo and Lieber [79] |
| 1982 | Induction of colonic MEOS by chronic ethanol ingestion | Seitz et al. [80] |
| 1983 | Induction of MEOS by hexachlorobenzene | Teschke et al. [81] |
| 1983 | Interaction of ethanol with vitamin A | Leo and Lieber [82] |
| 1983 | Tumor incidence caused by dimethylnitrosamine is influenced by prolonged alcohol use | Teschke et al. [83] |
| 1983 | Liver injury caused by carbon tetrachloride is modified by ethanol administered acutely | Teschke et al. [84] |
| 1983 | Liver injury due to chlorpromazine, paracetamol, and dimethylnitrosamine is modified by prolonged use of alcohol | Teschke [85] |
| 1983 | MEOS and ethanol metabolism in baboons | Nomura et al. [86] |
| 1983 | The alcohol dehydrogenase (ADH) independent pathway of ethanol metabolism in deermice lacking ADH | Shigeta et al. [87] |
| 1984 | Induction of the ethanol-specific CYP by imidazole in rabbits | Koop et al. [88] |
| 1984 | Induction of the ethanol-specific CYP 2E1 by isoniazid | Gadeholt [89] |
| 1984 | Circadian rhythm of MEOS | Sturtevant and Garber [90] |
| 1984 | Induction of the ethanol-specific CYP 2E1 by pyrazole in rabbits | Ingelman-Sundberg and Jörnvall [91] |
| 1984 | Formation of hydroxyl radical and oxidation of ethanol by CYP 2E1: studies of their mechanisms | Ingelman-Sundberg and Johansson [92] |
| 1984 | Reduced liver levels of vitamin A in humans and rats following drug treatment | Leo et al. [93] |
| 1985 | Induction of the ethanol-specific CYP 2E1 by trichloroethylene, acetone, pyrazole, and isoniazid in rabbit liver microsomes | Koop et al. [94] |
| 1985 | Involvement of the ethanol-specific CYP 2E1 in the microsomal metabolism of dimethylnitrosamine in rats, rabbits, mice, and guinea pigs | Yang et al. [95] |
| 1985 | Involvement of the ethanol-specific CYP in the microsomal metabolism of carbon tetrachloride in rabbits | Johansson and Ingelman-Sundberg [96] |
| 1985 | Ethanol-inducible CYP 2E1 identified as metabolizing acetone and acetol | Koop and Casazza [97] |
| 1985 | Details of liver microsomal CYP induced by isoniazid in the rat | Ryan et al. [98] |
| 1985 | Mixed function oxidation in deermice lacking alcohol dehydrogenase: Modification by acute alcohol administration and prolonged consumption of alcohol | Gellert et al. [99] |
| 1986 | Studies in deermice containing or missing ADH: Metabolic interactions of ethanol oxidation and mixed-function oxidation | Gellert et al. [100] |
| 1986 | Chronic administration of sex hormones and alcohol in female rats and the effect on liver enzymes metabolizing ethanol | Teschke et al. [101] |
| 1986 | Microsomal ethanol-oxidizing system of the liver: Biochemical nature and clinical aspects | Teschke [102] |
| 1986 | Drugs, retinol, and the relevance of their interactions | Leo et al. [103] |
| 1986 | Isoniazid and ethanol: induction of the same microsomal CYP isozyme 3a | Ryan et al. [104] |
| 1986 | Ethanol-inducible human liver demethylase for N-nitrosodimethylamine | Wrighton et al. [105] |
| 1986 | Ethanol-inducible CYP isozyme in rabbit nasal and kidney microsomes | Ding et al. [106] |
| 1986 | Hydroxylation of acetone catalyzed by ethanol- and acetone-inducible CYP in hepatic microsomes and reconstituted membranes | Johansson et al. [107] |
| 1986 | Complementary DNA and protein sequences of ethanol-inducible CYPs. A study in rats and humans | Song et al. [108] |
| 1987 | Induction of cytochrome P-450j: A study in the spontaneously diabetic BB rat, a strain in which about half of the animals develop insulin-dependent diabetes | Bellward et al. [109] |
| 1987 | Purification and characterization of human liver CYP 2E1 | Lasker et al. [110] |
| 1987 | Hepatic microsomal CYP 2E1 inducible by ethanol in rabbits: Details of cDNA and derived amino acid sequence | Khani et al. [111] |
| 1987 | Role of MEOS for interactions with other drugs, carcinogens, and vitamins | Lieber et al. [112] |
| 1987 | Pathways contributing to ethanol metabolism: ethanol-metabolizing pathways in deermice. A study on the estimation of flux calculated from isotope effects | Alderman et al. [113] |
| 1988 | Ethanol-inducible CYP 2E1 expressed in the centrilobular region of the rat liver | Ingelman-Sundberg et al. [114] |
| 1988 | Obesity is considered as a risk factor for drug-induced organ injury: Increased hepatic CYP levels and MEOS activity in the obese overfed rat | Salazar et al. [115] |
| 1988 | Acetaldehyde adducts formed with ethanol-inducible CYP 2E1 in vivo | Behrens et al. [116] |
| 1988 | CYP 2E1 in rabbit olfactory mucosa: its induction by ethanol and acetone | Ding et al. [117] |
| 1988 | Ethanol-inducible CYP 2E1: a study on molecular regulation in hamsters | Kubota et al. [118] |
| 1988 | Metabolism of benzene in microsomes obtained from rat and rabbit liver, and the role of CYP 2E1 induced by ethanol, acetone, and benzene | Johansson and Ingelman-Sundberg [119] |
| 1988 | Ligand-dependent maintenance of ethanol-inducible CYP: an experimental study using primary rat hepatocyte cell cultures | Eliasson et al. [120] |
| 1988 | Hepatic microsomal ethanol-inducible CYP 2E1 and its intralobular distribution of in liver | Tsutsumi et al. [121] |
| 1988 | Prolonged alcohol use enhances oxygen radical dependent inactivation of metabolic enzymes by liver microsomes | Dicker and Cederbaum [122] |
| 1989 | Induction and tissue-specific expression of rabbit CYP 2E1 genes | Porter et al. [123] |
| 1989 | The intralobular distribution of ethanol-inducible CYP 2E1; an experimental study in rat liver and a clinical analysis in human liver | Tsutsumi et al. [124] |
| 1990 | Ethanol-inducible CYP 2E1 and its regional distribution of in the central nervous system: an experimental study in rats | Hansson et al. [125] |
| 1990 | Modification of hepatic CYP 2E1 by pituitary hormones in rats and mice | Hong et al. [126] |
| 1990 | Ethanol-inducible CYP 2E1: multiple mechanisms are involved in its regulation | Koop and Tierney [127] |
| 1990 | Lymphocytes from patients with poorly controlled insulin-dependent diabetes exhibit increased CYP 2E1 | Song et al. [128] |
| 1990 | Induction of rat hepatic CYP 2E1 by pyridine | Kim et al. [129] |
| 1990 | Solvents enhance the translational efficiency in the course of CYP 2E1 induction | Kim et al. [130] |
| 1990 | CYP 2E1 influences the interactions of ethanol with enflurane metabolism and their toxicity | Tsutsumi et al. [131] |
| 1990 | CYP 2E1 is involved in nitrosamine metabolism and regulation | Yang et al. [132] |
| 1990 | Chlorzoxazone hydroxylation is a specific probe for CYP 2E1 in human liver | Peter et al. [133] |
| 1990 | Localization of ethanol-inducible CYP 2E1 assessed by immunohistochemistry in the alimentary tract of rats | Shimizu et al. [134] |
| 1991 | Post-translational reduction of CYP 2E1 by CCl4 | Sohn et al. [135] |
| 1991 | Role of hormones in the phosphorylation and degradation of CYP 2B1 and 2E1: a study in in isolated rat hepatocytes | Johansson et al. [136] |
| 1991 | Dietary lipids and carbohydrates modify the levels of CYP 2E1 in microsomes obtained from rat liver | Yoo et al. [137] |
| 1991 | CYP 2E1 is induced in the in the experimental obese rat model | Raucy et al. [138] |
| 1991 | Acetaldehyde as another substrate for ethanol-inducible CYP 2E1 | Terelius et al. [139] |
| 1991 | Identification and induction of CYP 2E1 in Kupffer cells of an experimental model of rats | Koop et al. [140] |
| 1991 | Genetic polymorphism in the 5′-flanking region change transcriptional regulation of the CYP 2E1 gene: a study in humans | Hayashi et al. [141] |
| 1992 | Interaction of ethanol with β-carotene: Delayed blood clearance and evidence of increased liver injury | Leo et al. [142] |
| 1992 | Intracellular degradation of CYP 2E1 is controlled by hormones and substrates | Eliasson et al. [143] |
| 1992 | Distribution the ethanol-inducible CYP 2E1 in the pancreas of rats fed ethanol combined with high fat or low fat diet | Sohda et al. [144] |
| 1992 | Oxidative and reductive metabolic pathways by CYP 2E1 | Koop [145] |
| 1992 | CYP 2E1 and 2A6 enzymes are the preferred catalysts for metabolic activation of N-nitrosodialkylamines and nitrosamines in the microsomes of human liver | Yamazaki et al. [146] |
| 1993 | Inhibition of chlorzoxazone metabolism by a single dose of disulfiram, and its potential role as a clinical probe for CYP 2E1 | Kharasch et al. [147] |
| 1993 | Enflurane defluorination catalyzed by CYP 2E1 in microsomes of human liver | Thummel et al. [148] |
| 1993 | Human CYP 2E1 stability in HepG2 cells | Day et al. [149] |
| 1993 | DraI and RsaI restriction fragment length polymorphisms analyzed in a study from Finland | Hirvonen et al. [150] |
| 1993 | Pathogenesis of alcoholic liver disease and the role of CYP 2E1 | Morimoto et al. [151] |
| 1993 | NADPH- and NADH-dependent production of superoxide and hydroxyl radical is enhanced in hepatic microsomes obtained following prolonged alcohol use | Rashba-Step et al. [152] |
| 1993 | Increased enzyme synthesis is responsible for the in vivo induction of hepatic CYP 2E1 | Tsutsumi et al. [153] |
| 1993 | Formation of 19(S)-, 19(R)-, and 18(R)-hydroxyeicosatetraenoic acids by alcohol-inducible CYP 2E1 | Laethem et al. [154] |
| 1993 | Induction of CYP 2E1 during prolonged alcohol use is due to the transcription of the CYP 2E1 gene when blood alcohol concentrations are high | Badger et al. [155] |
| 1993 | Contribution of cytochrome P-450s to MEOS: assessed by a specific and sensitive assay of MEOS activity using HPLC with fluorescence labeling | Kunitoh et al. [156] |
| 1993 | Levels of CYP 1A2 and CYP 2E1, and their related monooxygenase activities in human liver obtained as surgical samples | Lucas et al. [157] |
| 1993 | CYP 2E1 induction during chronic ethanol exposure occurs by a two-step mechanism, depending on blood alcohol levels: a study in rats | Ronis et al. [158] |
| 1993 | CYP 2E1 is the preferred enzyme catalyzing the defluorination of sevoflurane, isoflurane, and methoxyflurane in human liver microsomes | Kharasch et al. [159] |
| 1993 | Inhibition of CYP 2E1 by ethanol is caused in the human liver by corresponding increase in encoding messenger RNA | Takahashi et al. [160] |
| 1993 | Use of 4-nitrophenol as an in vitro substrate probe was validated for human liver CYP 2E1 | Tassaneeyakul et al. [161] |
| 1994 | Alcohol-derived radicals and their spin trapping in liver microsomes and reconstituted systems | Albano et al. [162] |
| 1994 | Ethanol augments the content and activity of human CYP 2E1 in a transduced HEPG2 cell line | Carrocio A [163] |
| 1994 | Significance of tissue-specific expression and methylation of the human CYP 2E1 gene | Botto et al. [164] |
| 1994 | Role of genetic CYP 2E1 polymorphism for the development of alcoholic liver disease | Tsutsumi et al. [165] |
| 1994 | Ethnic variation in the CYP 2E1 gene: Polymorphism analysis of 695 African-Americans, European-Americans and Taiwanese | Stephens et al. [166] |
| 1994 | Relationship between CYP 2E1 and acetone catabolism in rats as studied with the inhibitor diallyl sulfide | Chen et al. [167] |
| 1994 | Association between restriction fragment-length polymorphism of the human CYP 2E1 gene and susceptibility to alcoholic liver cirrhosis | Maezawa et al. [168] |
| 1994 | Involvement of CYP 2E1 in the (omega-1)-hydroxylation of lauric acid in rat liver microsomes | Amet et al. [169] |
| 1994 | CYP 2E1 induction by ethanol in a rat hepatoma FGC-4 cell model | McGehee et al. [170] |
| 1994 | Piperine modifies the expression of P4502E1, P4502B, and P4501A in rats | Kang et al. [171] |
| 1994 | Restriction fragment-length polymorphism of the human CYP 2E1 gene and susceptibility to lung cancer: Possible relevance to low smoking exposure | Uematsu et al. [172] |
| 1994 | Differences of regulation and expression of the human CYP 2E1 gene due to the RsaI polymorphism in the 5’ flanking region | Watanabe et al. [173] |
| 1995 | An RsaI polymorphism in the CYP 2E1 gene does not affect lung cancer risk in a Japanese population | Watanabe et al. [174] |
| 1995 | Ethanol induces CYP 2E1 by a mechanism involving protein stabilization | Roberts et al. [175] |
| 1995 | Renal tumorigenicity of 1,1-dichloroethene in mice: the role of male-specific expression of CYP 2E1 in the renal bioactivation of 1,1-dichloroethene | Speerschneider and Dekant [176] |
| 1995 | Intestinal toxicity of acrylonitrile: in vitro metabolism by intestinal CYP 2E1 | Subramanian and Ahmed [177] |
| 1995 | Stable expression of human CYP 2E1 in V79 Chinese hamster cells | Schmalix et al. [178] |
| 1995 | Lacking association of polymorphism at the CYP 2E1 locus with alcoholic liver disease in Caucasian men | Carr et al. [179] |
| 1995 | Modulation of experimental alcoholic liver injury by inhibitors of CYP 2E1 | Morimoto et al. [180] |
| 1995 | Genetic polymorphism of CYP 1A1, 2D6 and 2E1: Regulation and toxicological significance | Rannug et al. [181] |
| 1995 | Genetic polymorphism of CYP 2E1 and risk of alcoholic liver disease in Caucasians | Pirmohamed et al. [182] |
| 1995 | CYP 2E1 genotype and chlorzoxazone metabolism in healthy and alcoholic Caucasians | Lucas et al. [183] |
| 1995 | Decreased CYP 2E1 as assessed by the rate of chlorzoxazone hydroxylation in alcoholics during the withdrawal phase | Lucas et al. [184] |
| 1995 | Respective roles of CYP 2E1 and CYP A2 in chlorzoxazone, and ethanol metabolism in mammalian liver microsomes | Mishin et al. [185] |
| 1995 | CYP 2E1 is not the sole catalyst of chlorzoxazone hydroxylation in rat liver microsomes | Jayyosi et al. [186] |
| 1995 | Selectivity of CYP 2E1 in catalyzing chlorzoxazone 6-hydroxylation | Yamazaki et al. [187] |
| 1995 | Insulin down-regulates CYP 2B and 2E expression at the posttranscriptional level in the rat hepatoma cell line | De Waziers et al. [188] |
| 1995 | Evidence for a tissue-specific induction of cutaneous CYP 2E1 by dexamethasone | Sampo et al. [189] |
| 1995 | Ethanol oxidizing enzymes: Roles in alcohol metabolism and alcoholic liver disease | Crabb [190] |
| 1995 | CYP 2E1 changes in rat liver, kidney and lung microsomes after prolonged alcohol application, either orally or by inhalation | Zerilli et al. [191] |
| 1996 | Microsomal ethanol oxidizing system activity by human hepatic cytochrome P-450s and involvement of CYP 1A2, 2A6, 2B6, 2D6, 2E1, and 3A4 | Asai et al. [192] |
| 1996 | High inducibility of mouse renal CYP 2E1 gene by tobacco smoke and its possible effect on DNA single strand breaks | Seree et al. [193] |
| 1996 | Effects of diet and ethanol on the expression and localization of CYP 2E1 and 2C7 in the colon of male rats | Hakkak et al. [194] |
| 1996 | Induction of CYP 2E1 by ethanol in rat Kupffer cells | Koivisto et al. [195] |
| 1996 | Human CYP 2E1: From genotype to phenotype | Carriere et al. [196] |
| 1996 | Expression, catalytic activity, and inducibility of CYP 2E1 in the rat central nervous system | Tindberg and Ingelman-Sundberg [197] |
| 1997 | Enzymatic degradation of chlorzoxazone by hepatic microsomes from humans and 10 other mammalian species | Court et al. [198] |
| 1997 | Regulation of the hepatic CYP 2E1 gene during prolonged ethanol exposure: Lack of an ethanol response element in the proximal 5′-flanking sequence | McGehee et al. [199] |
| 1997 | Immunohistochemical determination of hepatic CYP 2E1 in formalin-fixed, paraffin-embedded sections | Cohen et al. [200] |
| 1997 | Effect of fatty acids and ketone bodies on CYP 2B, 4A, and 2E1 expression in primary cultured rat hepatocytes | Zangar and Novak [201] |
| 1997 | Ethanol metabolism in the brain | Zimatkin and Deitrich [202] |
| 1997 | Inhibition of CYP 2E1 expression by 2-(allylthio) pyrazine, a potential chemoprotective agent, and considerations on hepatoprotective effects | Kim et al. [203] |
| 1997 | Insulin effects on CYP 2E1, 2B, 3A, and 4A expression in primary cultured rat hepatocytes | Woodcroft and Novak [204] |
| 1997 | Lipid peroxidation, CYP 2E1 and arachidonoid acid metabolism in alcoholic liver disease in rats | French et al. [205] |
| 1997 | Chlormethiazole inhibition of CYP 2E1 as assessed by chlorzoxazone hydroxylation in humans | Gebhardt et al. [206] |
| 1998 | CYP 2E1 activity as assessed by chlorzoxazone hydroxylation: studies in patients with diabetes and obesity | Lucas et al. [207] |
| 1998 | Expression of CYP 2E1 in human liver: Assessment by mRNA, genotype, and phenotype | Powell et al. [208] |
| 1998 | Increased hepatic CYP 2E1 in patients with nonalcoholic steatohepatitis | Weltman et al. [209] |
| 1998 | Respective roles of human CYP 2E1 and 3A4 in the hepatic microsomal ethanol oxidizing system | Salmela et al. [210] |
| 1998 | Microsomal acetaldehyde oxidation is negligible in the presence of ethanol | Wu et al. [211] |
| 1998 | Selective inhibition of CYP 2E1 in vivo and in vitro with trans-1, 2-dichloroethylene | Matthews et al. [212] |
| 1998 | CYP 2E1 and its catalytic activity in rat testis | Jiang et al. [213] |
| 1998 | CYP 2E1 and 1A1 in the rat pancreas | Kessova et al. [214] |
| 1998 | CYP 2E1 is present in the rat pancreas and induced by prolonged alcohol consumption | Norton et al. [215] |
| 1998 | Polyenylphosphatidylcholine opposes the increase of CYP 2E1 by ethanol, and corrects the iron-induced decrease | Aleynik et al. [216] |
| 1998 | Involvement of CYP 2E1 in the (omega-1)-hydroxylation of oleic acid in human and rat liver microsomes | Adas et al. [217] |
| 1998 | Chlorzoxazone pharmacogenetics, a potential marker of hepatic CYP 2E1 in humans | Mishin et al. [218] |
| 1998 | Inhibition of CYP 2E1 by chlormethiazole as measured by chlorzoxazone pharmacokinetics in patients with alcoholism and in healthy volunteers | Eap et al. [219] |
| 1998 | Regulation of rabbit CYP 2E1 expression in HepG2 cells by insulin and thyroid hormones | Peng and Coon [220] |
| 1998 | CYP 2E1 inducibility and hydroxyethyl radical formation among alcoholics | Dupont et al. [221] |
| 1999 | Alcohol, vitamin A, and beta-carotene: adverse interactions, including hepatotoxicity and carcinogenicity | Leo and Lieber [222] |
| 1999 | Expression of CYP 2E1 by human monocyte-derived macrophages | Hutson and Wickramasinghe [223] |
| 1999 | Carbon monoxide, cigarette smoking, and CYP 2E1 activity | Benowitz et al. [224] |
| 1999 | Chlorzoxazone, a selective probe for phenotyping CYP 2E1 in humans | Lucas et al. [225] |
| 2001 | Effects of alcohol and diallylsulphide on CYP 2E1 activity in humans: a phenotyping study using chlorzoxazone | Loizou and Cocker [226] |
| 2001 | Inhibition of CYP 2E1 with natural agents may be a feasible strategy for minimizing liver injury by ethanol | McCarty [227] |
| 2001 | Ethanol and oxidative stress | Sun et al. [228] |
| 2002 | Effect of chronic disulfiram administration on CYP 1A2, CYP 2C19, CYP 2D6, CYP 2E1, and N-acetyltransferase in healthy humans | Frye and Branch [229] |
| 2003 | Rapid determination of enzyme activities of recombinant human CYPs, human liver microsomes, and hepatocytes | Ghosal et al. [230] |
| 2004 | CYP 2E1: biochemistry, toxicology, regulation, and function in alcoholic liver injury | Kessova and Cederbaum [231] |
| 2004 | Robustness of chlorzoxazone as an in vivo measure of CYP 2E1 activity | Ernstgard et al. [232] |
| 2006 | Effect of high-dosed aspirin on CYP 2E1 in healthy humans measured using chlorzoxazone as a probe | Park et al. [233] |
| 2008 | CYP 2E1 and oxidative liver injury caused by alcohol | Choi et al. [234] |
| 2010 | CYP-mediated differential oxidative modification of proteins: Albumin, apolipoprotein E, and CYP 2E1 as targets | Wellman and Siest [235] |
| 2014 | Association studies of CYP, family 2, subfamily E, and polypeptide 1 (CYP 2E1) gene polymorphisms with acute rejection in kidney transplantation recipients | Kim et al. [236] |
| 2014 | Pathogenesis of alcoholic liver disease: Significance of oxidative metabolism | Ceni et al. [237] |
| 2016 | Resveratrol pretreatment affects CYP 2E1 activity in healthy volunteers | Bedada and Neerati [238] |
| 2017 | Effect of piperine on CYP 2E1 enzyme activity in healthy volunteers | Bedada and Boga [239] |
| 2017 | The role of human CYP 2E1 in liver inflammation and fibrosis | Xu et al. [240] |
| 2017 | CYP 2E1 is involved in aging-related kidney damage in mice through increased nitroxidative stress | Abdelmegeed et al. [241] |
| 2018 | Vinyl chloride, CYP 2E1, and liver injury | Fujiwara [242] |
| 2018 | Vinyl chloride, diet, and liver injury | Lang et al. [243] |
Abbreviations: ADH, Alcohol dehydrogenase; MEOS, Microsomal ethanol-oxidizing system.
8. Hepatic Acetaldehyde Dehydrogenase
Acetaldehyde generated via ADH and MEOS is further metabolized in the liver cell by mitochondrial acetaldehyde dehydrogenase (ALDH) to acetate (Table 1, Figure 5) [25], which is released into the bloodstream and oxidized to CO2 in various extrahepatic tissues. Little evidence is provided to show that the classic cytosolic aldehyde dehydrogenase in the liver contributes significantly to the metabolism of acetaldehyde [46,47]. Instead, two other enzymes exist, localized in liver mitochondria: one of these has a low affinity for acetaldehyde and is therefore less important, whereas the other one is described by a high affinity for acetaldehyde, requires NAD+ as cofactor, and is considered as the main pathway for acetaldehyde oxidation in the liver [46,47].
This high-affinity ALDH has been characterized in detail in rat liver mitochondria, which metabolizes acetaldehyde at a rate of 12 nmoles/min/mg of mitochondrial protein [47]. ALDH competes for NAD+ with cytosolic ADH, which also requires NAD+ (Table 1, Figure 5), and both enzymes generate NADH + H+, whereas MEOS requires reducing equivalents in form of NADPH + H+ (Table 1, Figure 5).
This again shows the close interrelation between ALDH, ADH, and MEOS at the level of reducing equivalents. Of more importance, chronic alcohol consumption decreased acetaldehyde oxidation in intact mitochondria and reduced the ALDH activity in liver mitochondria disrupted by deoxycholate [46]. This reduction initiates a vicious circle due to an accumulation of acetaldehyde, which is toxic to the liver (Figure 6) [46].
Figure 6.

Hypothesis of a vicious circle of acetaldehyde in the liver. Acetaldehyde is increasingly generated from ethanol through MEOS, which is adaptively induced in activity following chronic ethanol consumption. Increased acetaldehyde levels in the liver in turn impair mitochondrial functions, including the activity of mitochondrial acetaldehyde dehydrogenase, which again likely enhances hepatic acetaldehyde concentrations at least temporarily, representing a vicious circle. Discussed and presented as a figure in a previous report [46], and reproduced with permission of the Publisher American Association for the Advancement of Science (AAAS, Washington, DC, USA).
Through a condensation process with dopamine or serotonin, acetaldehyde facilitates the development of alcohol dependence [265] and can injury other organs including the brain. Higher blood acetaldehyde levels are found in alcoholic patients, as compared to non-alcoholic individuals [264], assuming a contributory role of the induced MEOS for the increased blood acetaldehyde levels due to an increased metabolism of ethanol to acetaldehyde in the liver. Sufficient circumstantial evidence exists that acetaldehyde is more injurious than ethanol (Figure 7).
Figure 7.

Actions of acetaldehyde. The increasingly generated acetaldehyde in the liver spills over in the blood and reaches many organs, which are injured by direct toxic attacks or through condensation products. Alcohol dependence is considered to be triggered by the condensation of acetaldehyde with dopamine or serotonin. Symbol ↑: Increase.
Acetaldehyde plays a critical role in East Asian individuals, who are genetically deficient in hepatic ALDH2, which upon alcohol drinking causes increased blood acetaldehyde levels and the so-called clinical Asian facial flush [24]. These symptoms are also found in East Asians with a genetically more active form of liver ADH1B*2, converting ethanol to acetaldehyde in excess as compared to individuals lacking this super-active ADH variant [24]. With ADH1B*3, another super-active ADH variant is known, and found primarily in populations of African descent and Native Americans. Ethanol is metabolized through these two super-active ADH variants at a 30–40-fold increased rate to acetaldehyde, compared to normally functioning ADH enzymes encoded by the wild type ADH1B*1 gene. Clearly, individuals suffering from increased acetaldehyde levels due to such genetic variants of ALDH or ADH will abstain from alcohol drinking, preventing flush and the risk of alcohol dependence and alcoholic liver disease.
9. Cascade of Molecular Mechanisms and Cellular Events
9.1. The 5-Hit Working Hypothesis of ALD
ALD in patients with a history of prolonged alcohol abuse includes several stages. Among these are: alcoholic fatty liver (AFL), alcoholic steatohepatitis (ASH), alcoholic hepatitis (AH), alcoholic cirrhosis (AC), and alcoholic hepatocellular carcinoma (AHCC), this warrants discussing the various steps leading to AHCC as the end-stage disease of ALD, and considering a proposed cascade of events promoted along with five injury hits [25]. A multi-hit hypothesis is well applicable in chronic liver injury of ALD caused by prolonged use of alcohol [25,266], and suggests classifying ALD as a multi-hit disease. A hit hypothesis is also useful in idiosyncratic DILI due to use of conventional drugs, considering that one hit will follow the other [267]. The various stages of alcoholic liver disease are presented in connection with their respective hits (Figure 8).
Figure 8.
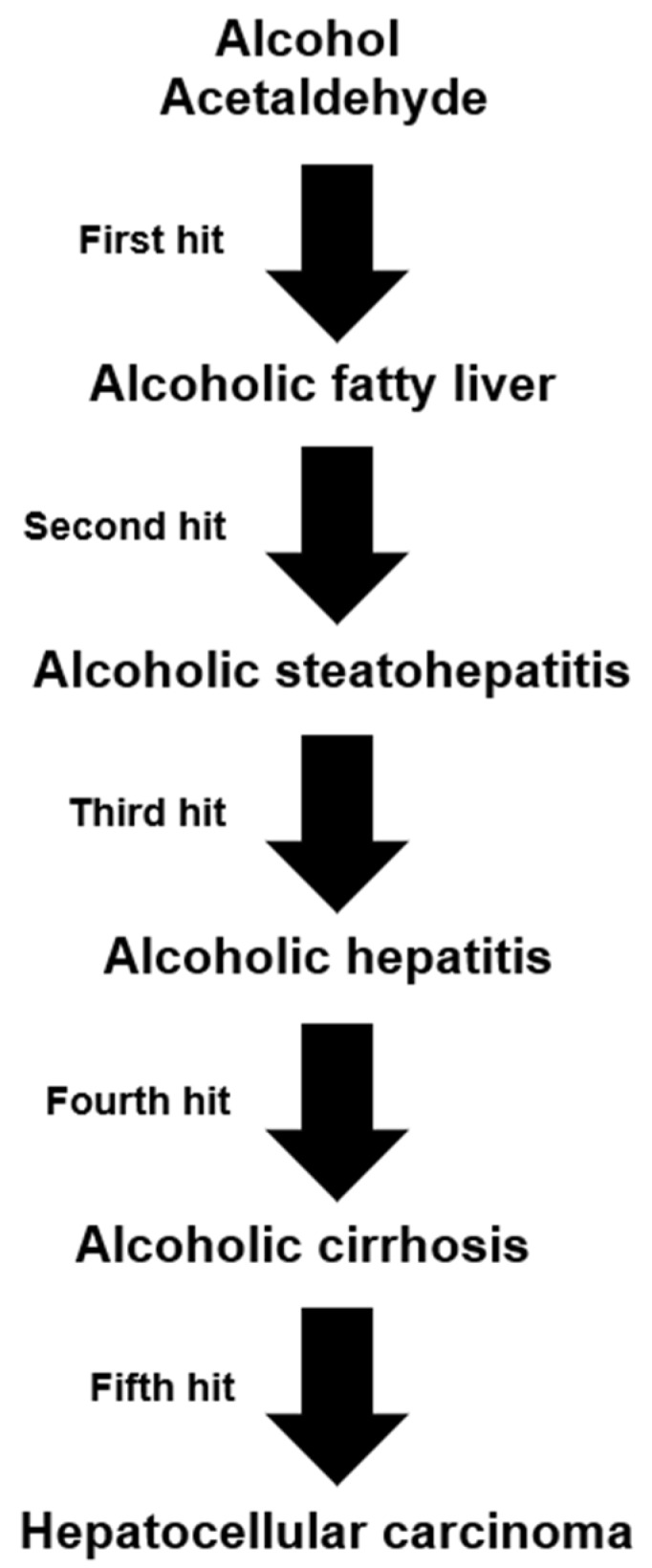
The 5-hit working hypothesis in alcoholic liver disease. The 5-hit hypothesis presents various possible steps leading from alcoholic fatty liver, eventually to hepatocellular carcinoma. In clinical practice, some patients with alcoholic hepatitis do not have steatosis/steatohepatitis as a precursor, with additional details provided in Table 4. The original figure was published in an earlier report [25] and is reproduced with permission of the Publisher Taylor & Francis (Didcot, UK).
The concept of a multi-hit disease as outlined for ALD (Figure 8) is under similar discussion also for a variety of other diseases including cancer, chronic disabling diseases, and more recently, obesity with nonalcoholic fatty liver disease (NAFLD) or nonalcoholic steatohepatitis (NASH) [115,137,138,207,209,268,269,270].
Of limited clinical value is such a multi-hit hypothesis for acute liver injury, caused for instance, by a single ingestion of overdosed acetaminophen that initiates intrinsic rather than idiosyncratic liver injury [271], or by acute intoxication of aliphatic halogenated hydrocarbons such as carbon tetrachloride, leading to acute toxic hepatitis [257,272].
The proposed 5-hit working hypothesis of ALD is based on highly complex and partially disputed conditions, thereby certainly open for critical discussion also regarding its potential clinical impact and whether the patient could benefit therapeutically from these considerations [22,25]. However, it clarifies several key factors leading to the five stages of ALD and outlines mechanistic sequelae as cascade of events (Table 4).
Table 4.
Alcoholic liver disease and the 5-hit working hypothesis with a tentative cascade of events. Hypothetical steps of the five hits leading to end-stage alcoholic liver disease. Adapted from a previous report [25] and reproduced with permission of the PublisherTaylor & Francis (Didcot, UK).
| First Hit. | The first hit is dependent on ADH and occurs at low alcohol levels through the generation not only of NADH + H+ leading to an increased NADH + H+/NAD+ ratio, which stimulates hepatic fatty acid synthesis [22] and increases α-glycerophosphate-trapping fatty acids [22,33], but also of acetaldehyde, which impairs hepatic mitochondrial functions including hepatic mitochondrial fatty acid oxidation [22]. This first hit fully explains at least in part the development of alcoholic fatty liver. |
| Second Hit. | The second hit is classified as a transition from alcoholic fatty liver to alcoholic steatohepatitis, most likely triggered by the increased production of acetaldehyde via MEOS [22,23] and of reactive oxygen species (ROS) with its capacity for irreversible covalently binding to cellular macromolecules, including membrane proteins and phospholipids [45,50,56,92,152,205,231,234,237,244,245,246,247,248,249,250,251,252]. These injurious alterations at the molecular and cellular level cause some necrosis, apoptosis, and inflammatory cells in the fatty liver, justifying the term alcoholic steatohepatitis, as it includes toxic hepatitis in steatosis [25]. Further stages are characterized by perisinusoidal and pericentral fibrosis due to participation of non-hepatocytes such as Kupffer cells, stellate cells, and sinus endothelial cells. Mediators such growth factors, interferons, interleukins, tumor necrosis factor and endotoxins, as well as hepatic iron, are considered as possible active promoters of liver injury, but considering the multiplicity of proposed mediators, it is difficult to predict how they interact with each other and modify the course of liver injury. |
| Third Hit. | The third hit initiates a more severe liver injury stage, whereby alcoholic steatohepatitis is the precursor in most, but certainly not all patients with alcoholic hepatitis. Steatosis is no more a characteristic feature, but is now replaced by necrosis, apoptosis, and inflammation. At this stage, injury becomes more severe and presents with more fibrosis and as a self-perpetuating process, immunity aspects gain additional relevance, because alcohol modifies the innate and adapted immune system, which may explain the individual differences of susceptibility for ALD. With the third hit, the disease may approach a point of no return. |
| Fourth Hit. | The fourth hit is dominated by increased fibrosis, due to increased collagen formation. This allows for a clinically unrecognizable transition from alcoholic hepatitis with fibrosis to irreversible cirrhosis. However, AC can also develop without ASH or AH. |
| Fifth Hit. | In rare cases, a fifth hit initiates the development of a hepatocellular carcinoma (HCC), mostly occurring in patients with cirrhosis. This final hit scenario of carcinogenesis is triggered by acetaldehyde and ROS through the generation of DNA adducts, which promote mutagenesis, and interference with methylation, synthesis, and repair of DNA. Suggested is a possible role of SIRT1. These overall events will enhance AHCC susceptibility, keeping in mind that ethanol itself is not a carcinogenetic chemical. |
9.2. Hepatocytes versus Non-Parenchymal Cells
Hepatocytes are involved in the development of ALD and govern the metabolism of ethanol and acetaldehyde in the liver [22,45,234,244,245,246,247,248,249,250,251,252]. For a quick overview, the complexity of ALD is illustrated as an example for AH, and it includes issues such as the involvement of hepatocytes and several non-parenchymal cells of the liver, the role of mediators, and the generation of multiple ROS (Figure 9).
Figure 9.

Hypothetical steps leading to alcoholic hepatitis. The pathogenesis of alcoholic hepatitis involves various mediators and cell types of the liver, some of the steps need confirmation and are therefore hypothetical. The original figure was published in a recent report [25] and is reproduced with permission of the Publisher Taylor & Francis (Didcot, UK).
In order to maintain the normal biologic and immunologic functions and regulate the homeostasis of the liver, the various cells are closely connected through mediators (Figure 9).
9.2.1. Kupffer Cells
KCs contain CYP 2E1 similar to hepatocytes [195] and contribute to the pathogenesis of ALD through their activation by gut bacteria derived endotoxins and lipopolysaccharides, which are taken up by the liver in increased amounts [273]. As a result and likely due to specific actions of CYP 2E1 possibly upgraded following chronic alcohol consumption, activated KCs produce large amounts of ROS and mediators (Figure 9) and thereby promote ALD in joint actions with hepatocytes, HSCs, and LSECs, but details require further confirmatory studies. Although these events are suggestive as potential targets of pharmacotherapy in ALD [273], such specifically targeted therapy approaches were mostly disappointing with respect to severe AH [25] and these failures are best explained by additional pathogenetic mechanisms.
9.2.2. Hepatic Stellate Cells
HSCs are known as key cells responsible for collagen synthesis, leading to alcoholic liver fibrosis and cirrhosis [274,275]. For this purpose, HSCs proliferate and undergo activation through phenotypic transdifferentiation into collagen-producing myofibroblasts [275]. The mechanism involved in this cell proliferation and activation was considered in response to necrosis and inflammation, partially triggered by mediators derived from KCs. However, the observation that in ALD, fibrosis can develop in the absence of AH, questioned the concept of necrosis an inflammation as obligatory factors of the proliferation and activation process. Instead, activation of HSCs was found to correlate with the severity of steatosis in a study cohort [275], requiring confirmation by other studies.
9.2.3. Liver Sinusoidal Endothelial Cells
LSECs are continuously exposed to the blood of the liver sinusoids, since they form their walls [276]. They are known for their endocytic activity, which is high in well-functioning cells with intact fenestration, but impaired if cell fenestration is reduced [276]. In alcoholic liver injury, capillarization and lack of LSEC fenestration prevail, which is considered permissive for HSC activation and fibrosis under the guidance of various mediators.
9.3. Oxidative Stress and Reactive Oxygen Species
Along with oxidative stress, there is a continuous generation of radicals including ROS by not only hepatocytes but also non-parenchymal cells and leucocytes (Figure 9). Because of incomplete intracellular oxygen splitting, radicals are part of common life in any healthy organism such as plants [277,278,279,280], animals [280], and humans [281,282]. ROS and the other reactive radicals are commonly scavenged by antioxidants and thereby detoxified. Disease occurs if these protective mechanisms are lacking, with obesity as a good example because radicals play an important role for many obesity-related co-morbidities [281]. ROS is also implicated in liver injury [282,283] including alcoholic liver disease [92,231,240] where various forms of ROS and reactive radicals are contributory (Table 5).
Table 5.
Potentially toxic metabolites resulting from the enzymatic degradation of ethanol in the liver Derived from original reports and review articles [25,50,56,92,152,205,231,237,245,246,247,248,249,250,251,252].
| Selected Potentially Toxic Metabolites and Reactive O2-Species due to Hepatic Ethanol Degradation |
|---|
| Acetaldehyde C2H4O |
| Ethoxy radical CH3CH2O |
| Hydroxyethyl radical CH3C(·)HOH |
| Acetyl radical CH3CHO |
| Singlet radical 1O2 |
| Superoxide radical HO2 |
| Hydrogen peroxide H2O2 |
| Hydroxyl radical HO |
| Alkoxyl radical RO |
| Peroxyl radical ROO |
| Lipid peroxides |
Large amounts of ROS are produced in both subcellular areas, the mitochondria where ROS is injurious, and in the endoplasmic reticulum, where MEOS is primarily located. Fairly good evidence exists that ethanol oxidation via MEOS is promoted in some way by hydroxyl radicals [50,56,92,152,162] or superoxide radicals [56,152], leading to intermediates such as hydroxyethyl radicals [221]. Phospholipids are obligatory constituents of MEOS [30,49,57], likely in the form of lipid peroxides [205].
9.4. Signaling Mediators
Abundant signaling mediators are produced by hepatocytes and non-parenchymal cells, with details as examples being referenced in some selected reports [273,274,275,276,277]. Mediators leave the producing cells and modulate other cells; some of these signaling pathways are illustrated, and clearly classified as hypothetical steps, to be on the cautious side (Figure 9). It is outside of the focus of this article to provide additional references and a more thorough discussion on the individual mediators, since many suggested functions are yet debated.
10. Clinical Issues of Alcoholic Liver Disease
10.1. Natural Course
It is estimated that around 90–95% of the patients with a history of alcohol abuse will experience a fatty liver, whereas one third will advance to fibrosis and cirrhosis [6]. However, more robust data can only be achieved in a cohort of AFL with sequential liver biopsies, an approach that is clearly not acceptable, due to ethical reasons. The natural course of ALD is complex and complicated by serious, potentially life-threatening events (Figure 10).
Figure 10.

Stages of alcoholic liver diseases with potential clinical outcomes. The clinical outcome is variable among the different stages. Clinical deterioration is most commonly associated with continuation of alcohol use.
10.2. Questionaires
Prerequisite for the diagnosis is among others that the alcohol abuse is confirmed by specific questionnaires, best to be evaluated by canonical tests [284,285,286,287], as recommended recently [25]. One of these is the CAGE questionnaire [284]. Alternatively, the more complex Michigan Alcoholism Screening Test (MAST) may be applied [285]. Both questionnaires are under discussion, regarding issues of validation, such as sensitivity and specificity [286,287]. These questionnaires facilitate not only the screening for alcoholism itself, but will also help exclude alcohol-unrelated liver diseases as confounding variables, using diagnostic parameters, as outlined in a previous publication [25]. Patients with ALD commonly use drugs and herbal products, making them susceptible for DILI [288,289] and HILI [290]. Their causality should be excluded or verified using the updated version of the RUCAM (Roussel Uclaf Causality Assessment Method) [291], considering that among patients with initially assumed DILI, almost 5% of them had not DILI but ALD [292]. Many other alternative diagnoses are to be excluded in patients with ALD to be sure that ALD is really ALD (Figure 11).
Figure 11.

Differential diagnosis of alcoholic liver disease. Patients with a history of alcohol abuse presenting with increased liver values, require a careful diagnosis to exclude liver diseases that are unrelated to alcohol abuse.
10.3. Laboratory Approaches
The search for individuals with severe alcohol abuse can be assisted by laboratory data with variable percentages of sensitivity: carbohydrate-deficient transferrin (CDT; 63%), gamma-glutamyltransferase (GGT; 58%), mean corpuscular volume of erythrocytes (MCV; 45%), aspartate aminotransferase (AST; 47%), alanine aminotransferase (ALT; 50%), and GGT + CDT (90%) [6]. However, and as expected, the specificity of these parameters is low. Known for a long time in clinical practice, high serum GGT activities in alcoholic patients created interest among hepatologists and led to experimental and clinical studies on its pathogenetic mechanisms.
In experimental studies on GGT activities due to alcohol feeding [293], using the liquid alcohol diet of Lieber and DeCarli [294], it was found that in animals, prolonged alcohol use reproduced the clinical findings of increased serum GGT activities in patients with alcohol abuse. Experimentally, chronic alcohol consumption led to statistically significant increases of GGT activities in the serum by 249% and in the liver by 60% [295]. This led to the conclusion that increased GGT activities in the serum are likely the result of an inducing effect of alcohol on liver GGT, and they cannot be ascribed alone to injurious properties of the alcohol, which would decrease the hepatic GGT activities due to enzyme release out of the liver cell. A similar increase of hepatic GGT activity was found in the liver of patients with ALF, associated with increased values in the serum [295]. Additional experimental studies showed that chronic alcohol use induced and doubled GGT activities in the microsomal fraction obtained after ultracentrifugation of liver homogenates [293]. Prolonged alcohol use increased GGT activities also in the liver plasma membranes, more so in bile canaliculi-enriched ones than in those free of bile canaliculi [296,297]. In other experimental studies, chronic alcohol consumption was found to increase bile flow, biliary GGT excretion, and bile acid output, all parameters that are assessed after cannulation of the bile duct [298]. The relationship between the biliary output of total bile acids and GGT was plotted against each other, and this showed an apparent linear relationship between both parameters in alcohol-fed animals, as well as in their pair-fed controls [298]. It seems that bile acids play a major role as natural chemicals solubilizing the GGT, an enzyme that is firmly bound to or tightly embedded in the liver plasma membranes and microsomal membranes. This view is supported by increased serum bile acid levels after experimental chronic alcohol use [298], in line with similar data obtained in patients with ALD [299].
Moreover, in vitro addition of deoxycholate caused increased GGT activities in liver microsomes of control and alcohol-fed animals, likely because of enzyme solubilization and its release out of the microsomal membrane [293]. Conversely, ethanol added in vitro to the microsomal fraction failed to enhance microsomal GGT activity of control animals, but increased enzyme activity in alcohol-fed animals, substantiating that chronic alcohol use may predispose to membrane GGT solubilization. In other experiments, the acute effect of alcohol on various constituents of the bile was evaluated, whereby rats received intravenously administered 0.9% NaCl solution alone or containing in addition ethanol [300]. Compared to the control group receiving saline alone, ethanol infusion significantly increased biliary gamma-glutamyltransferase excretion by 166% (p < 0.0125); while bile flow and biliary excretion of both total bile acids and alkaline phosphatase remained virtually unchanged. The selective increased biliary excretion of gamma-glutamyltransferase through the action of ethanol is best explained by an augmented solubilization of GGT, an enzyme found in the small bile canaliculi of the liver cells and the epithelial cells of the larger bile ducts. Alcohol infusion failed to change the excretion of total bile acids via the bile, suggesting that the selective solubilization of gamma-glutamyltransferase occurs by a mechanism primarily not involving total bile acids. Instead, it is likely caused by alcohol with its physico-chemical properties that may lead to the increased fluidity of liver plasma membranes [300]. Some aspects of mechanisms leading to alcohol-related GGT increase in the serum have been summarized (Figure 12) [301].
Figure 12.

Hypothesis of events, leading to increased serum gamma-glutamyltransferase (GGT) activities, following chronic alcohol consumption. Mechanisms leading to increased GGT in the serum following alcohol abuse include microsomal GGT induction and enzyme solubilization via ethanol and bile acids. Symbol: ?, process under discussion. Abbreviation: GGT, gamma-glutamyltransferase.
Chronic alcohol consumption causes induction of GGT and various other parameters in the endoplasmic reticulum of the hepatocyte [23], with clinical implications (Figure 13).
Figure 13.

Proliferation of the smooth endoplasmic reticulum associated with microsomal induction of gamma-glutamyltransferase (GGT) due to alcohol abuse. Chronic alcohol consumption induces also various other microsomal functions, which are of potential clinical relevance. In addition, increased GGT activities of the plasma membranes may contribute to increases in the serum [296,297]. Symbol: ?, under discussion. The original figure was published in an earlier report [23] and is reproduced with the permission of the Publisher Wiley (Hoboken, NJ, USA).
In essence, chronic alcohol consumption causes induction of GGT in the endoplasmic reticulum of the hepatocyte (Figure 13) [23], from which GGT is solubilized by alcohol, bile acids, and ROS derived from ethanol-induced CYP 2E1, tightly associated with GGT in the microsomal membrane, is translocated via the Golgi apparatus to and incorporated in the liver plasma membranes (Figure 12). These membranes may be susceptible for partial solubilization of GGT through the action of alcohol and increased bile acid levels, a process that facilitates spilling over of the solubilized GGT into the blood of the liver sinusoids, leading to increased GGT activities in the blood of the systemic circulation (Figure 12 and Figure 13). Here, its activity can then be assayed in the serum of patients.
In a clinical setting of patients with ALD, serum activities of GGT are high, as compared to a control group lacking a history of alcohol abuse and presenting with normal GGT values (Figure 14). The highest serum GGT activities are found in patients with AFL, with a decreasing tendency, along with increasing fibrotic changes and transition of fibrosis to cirrhosis (Figure 15). These results were obtained in a German population, and they differ substantially from a recent study in Beijing that showed no differences of serum GGT activities among the various stages of ALD [302]. Abstinence from alcohol use commonly leads to a decline of serum GGT activity, a useful approach for detecting relapse of alcohol use (Figure 16) [301].
Figure 14.

Serum gamma-glutamyltransferase (GGT) activity in alcoholic liver diseases. Patients with an alcoholic liver disease show increased serum GGT activities as compared to a control group lacking a previous history of alcohol abuse and with normal liver tests.
Figure 15.

Serum gamma-glutamyltransferase (GGT) activities in patients with different stages of alcoholic liver diseases. Highest GGT activities were found in patients with alcoholic fatty liver, with decreasing values along with increasing fibrosis. Relative low values are found in patients with alcoholic cirrhosis, possibly due to reduced GGT enzyme induction because of impaired liver function.
Figure 16.

Decline of serum gamma-glutamyltransferase (GGT) activities due to alcohol abstinence. Alcohol abstinence leads to a reduction of serum activities of GGT in patients with alcoholic liver disease of all stages including alcoholic fatty liver, alcoholic steatohepatitis, alcoholic hepatitis, and alcoholic cirrhosis. This approach is extremely valuable in any clinical setting for checking whether a patient has followed the professional advice to stop alcohol use. The original figure was published in a previous report [301] and is reproduced with the permission of the Publisher Springer (Berlin, Germany).
Patients with alcoholic fatty liver show mostly striking increased GGT activities in the serum, associated with corresponding increases in the liver; these data were reproducible in an animal model of chronic alcohol feeding (Table 6) [295], useful for studying the mechanism of increased serum GGT activities [293,296,297,298,300].
Table 6.
GGT activities in AFL. Studied were 19 patients with AFL and nine patients without AFL. Animals: eight rats with AFL, fed chronically with an alcohol-containing diet were compared with eight controls that were fed control diets. Data are given as means ± SD, adapted from a previous report [295]. Abbreviations: AFL, Alcoholic fatty liver; GGT, gamma-glutamyltransferase.
| Study Cohort | Alcoholic Fatty Liver | Controls | Significance |
|---|---|---|---|
| Patients | |||
| Serum GGT (U/L) | 195.0 ± 93.7 | 13.7 ± 2.0 | p < 0.025 |
| Liver GGT | |||
| (U/g wet weight) | 4.78 ± 0.4 | 1.91 ± 0.2 | p < 0.025 |
| (U/g protein) | 35.9 ± 16.1 | 16.4 ± 6.6 | p < 0.025 |
| Animals | |||
| Serum GGT (U/L) | 4.41 ± 1.64 | 2.19 ± 0.31 | p < 0.025 |
| Liver GGT | |||
| (U/g wet weight) | 0.14 ± 0.06 | 0.07 ± 0.03 | p < 0.001 |
| (U/g protein) | 1.19 ± 0.23 | 0.79 ± 0.19 | p < 0.0.25 |
| (U/100 g body weight) | 0.80 ± 0.28 | 0.34 ± 0.09 | p < 0.001 |
Serum ALT and AST activities are often rather low in ALD [295], including AFL (Table 7).
Table 7.
Serum AST, ALT, and GDH activities in alcoholic fatty liver.
| Patient with AFL | Degree of Steatosis | AST (U/L) | ALT (U/L) | Ratio AST/ALT | GDH (U/L) |
|---|---|---|---|---|---|
| 1 | 50% | 12.4 | 17.7 | 0.70 | 53.9 |
| 2 | 60% | 100.7 | 22.5 | 4.47 | 35.8 |
| 3 | 60% | 50.3 | 19.9 | 2.52 | 16.8 |
| 4 | 80% | 54.7 | 9.2 | 5.94 | 7.7 |
| 5 | 10–15% | 20.6 | 31.7 | 0.65 | 2.0 |
| 6 | 50% | 25.6 | 33.9 | 0.76 | 7.9 |
| 7 | 60–70% | 61.4 | 62.4 | 0.98 | 9.1 |
| 8 | 60–70% | 11.6 | 7.9 | 1.47 | 1.8 |
| 9 | 30–40% | 33.2 | 61.1 | 0.54 | 4.1 |
| 10 | 80% | 53.6 | 19.0 | 2.82 | 1.9 |
| 11 | 80–90% | 34.6 | 46.8 | 0.74 | 9.2 |
| 12 | 30% | 16.0 | 30.6 | 0.52 | 3.3 |
| 13 | 30–40% | 32.9 | 19.9 | 1.65 | 4.8 |
| 14 | 20–30% | 11.3 | 33.8 | 0.33 | 7.3 |
| 15 | 50–60% | 25.7 | 62.2 | 0.41 | 7.2 |
| 16 | 10% | 70.1 | 19.3 | 3.63 | 7.2 |
| 17 | 10–15% | 9.2 | 6.7 | 1.37 | 0.8 |
| 18 | 50% | 26.1 | 68.1 | 0.38 | 7.5 |
| 19 | 50% | 10.9 | 9.2 | 1.19 | 2.1 |
| Means ± SEM | 34.8 ± 5.7 | 30.2 ± 4.6 | 1.53 ± 1.51 | 9.9 ± 3.0 |
Normal range was for AST < 35 U/L, ALT < 45 U/L, and GDH < 7 U/L. Abbreviations: AST, Aspartate aminotransferase; ALT, Alanine aminotransferase; GDH, Glutamate dehydrogenase. Results are derived from 19 patients with alcoholic fatty liver, and with some case details published previously [295].
The serum ratio of AST/ALT provided diagnostic clues in ALD [303]. The ratio is significantly increased in patients with alcoholic hepatitis and cirrhosis (2.85 ± 0.2), and much higher compared to cirrhosis of the so-called postnecrotic type (1.74 ± 0.2), not further specified chronic hepatitis (1.3 ± 0.17), jaundice due to biliary obstruction (0.81 ± 0.06), and not further specified viral hepatitis (0.74 ± 0.07). This led to the proposal that a serum AST/ALT ratio >2.0 is highly suggestive of alcoholic hepatitis and cirrhosis [304], found in as much as 70% of this special cohort, but in only 26% of postnecrotic cirrhosis, 8% of not further specified chronic hepatitis, 4% of acute viral hepatitis without specification, and none of jaundice caused by biliary obstruction. Similar data were published in a study from China, which confirmed a striking increase of the AST/ALT ratio for severe AH (2.08 ± 1.23), but a lesser one for AC (1.44 ± 0.96), whereas the ratio was lower in mild AH (0.66 ± 0.72) and AFL (0.62 ± 0.51) [302]. In Germany, the values of the AST/ALT ratio have been published for patients with AFL (1.64 ± 1.57), as compared with a corresponding control group consisting of individuals with normal liver histology and normal values of AST, ALT, and GGT; this showed a lower AST/ALT ratio (0.72 ± 0.24) that did not achieve a statistically significant level, due to the small control group, and the variability of results within the AFL cohort [295]. Data variability appears indeed to be a major problem in patients with established alcohol abuse and AFL (Table 7). Nevertheless, an AST/ALT ratio of >1.0 can help identify individuals with problematic alcohol use without overt liver alcoholic liver disease, while a ratio <1.0 cannot exclude alcohol abuse with the required d level of certainty. Of interest in the clinical context, serum glutamate dehydrogenase (GDH) activity may be a good marker for alcoholism, with the preference of liver cell necrosis in the alcoholic, showing virtually no overlap between patients with and without AH [305]. Serum GDH activity was also found, with a 5-fold increase in patients with AFL, which was significantly different from controls [304] and showed little variability (Table 7).
With the toxic acetaldehyde as the first oxidation product derived from ethanol, it is not unexpected that antibodies against acetaldehyde proteins are generated and found in the serum of individuals with an alcohol abuse and an obligatory alcohol metabolism. Such serum antibodies can be determined in patients with AH, but their diagnostic validity in a clinical setting is unclear [304]. In this diagnostic context, there are suggestions that blood microRNAs could be of value as diagnostic biomarkers in AH [306], but this issue is yet debated for other liver injuries including DILI or HILI [307,308].
11. Alcoholic Fatty Liver
AFL is a liver injury that is completely reversible upon abstinence from alcohol [22], and it lacks a lethal course during hospitalization [302] except in rare, extremely severe cases [22]. Early recognition is mandatory to advice stopping alcohol use and to prevent more serious stages of ALD. Symptoms are not specific and rarely described [4,5,6,9]. By convention, diagnosis of AFL requires a fat deposition >5% of the liver cells, and presents mostly as macrovesicular steatosis, and rarely as the microvesicular type [309]. Clinical differentiation of AFL is mandatory especially from NAFLD and NASH (Figure 13), commonly associated with obesity and its comorbidities [270,281,310]. Patients with AFL are prone to injurious hits by various drugs and chemicals including carbon tetrachloride [39,257,272], with augmented risk due to increased metabolism related to the induction of CYP 2E1 (Figure 17).
Figure 17.

Risk factors of exogenous substrates for alcoholic fatty liver. At the stage of alcoholic fatty liver, and due to microsomal induction of cytochrome P450, various exogenous substrates are increasingly metabolized, leading to additional liver injury or to decreased blood drug levels.
12. Alcoholic Steatohepatitis and Alcoholic Hepatitis
Reports often do not distinguish between ASH and AH, but a clear differentiation is essential for reasons of clarity to describe their characteristic features of the natural clinical course and treatment options [25]. Unlike ASH, AH commonly presents as a severe disease with a high risk of complications. Similar to AFL, clinical symptoms are rarely observed in patients with ASH, whereas AH is characterized by poor nutritional condition, enlarged liver and spleen, jaundice, ascites, mental alterations, and hepatic as well as renal insufficiency [311].
The liver histology of ASH is described as hepatic injury with steatosis [309,312,313,314,315]; characteristic are ballooned liver cells [313], lytic necrosis is more frequent than apoptosis [313,316]. Not pathognomic are Councilman bodies [313] and Mallory-Denk bodies [309,313]. Lobular inflammation mostly consists of neutrophils, commonly found nearby to ballooned hepatocytes [313]. The extent of histological changes determines the lethality risk of AH [311].
AH is commonly a type of acute-on-chronic liver disease [25]. Liver biopsy is certainly indicated under the assumption that the patient would benefit from the potential results of this invasive procedure, such as if pharmacotherapy options are to be discussed in severe AH [317,318].
Restricted to severe AH, pharmacotherapy options have been thoroughly reviewed [25]. In short, corticosteroids are most promising as the therapy of first choice [25], considering the results of clinical trials [319,320,321,322,323,324,325,326,327,328,329,330]. Pentoxifylline is recommended as a second-line therapy [331,332]. Preferentially indicated in patients with severe AH and concomitant infections or sepsis is pentoxifylline, not allowing steroid treatment [25]. Most importantly, patients not responding to a seven-day corticosteroid therapy should be taken off this treatment and then evaluated for liver transplantation, because other pharmacotherapy options are not available. This also rules out the use of pentoxifylline as rescue therapy because of missing efficacy in this particular cohort subset. A lack of efficiency in patients with severe AH was also provided for drugs like infliximab [333,334,335], propylthiouracil, N-acetylcysteine, silymarin, colchicine, insulin and glucagon, oxandrolone, testosterone, and polyunsaturated lecithin [25].
Despite pharmacotherapy options, the basic approaches of therapy and prevention remain, as are nutritional support and alcohol abstinence [25]. Malnutrition causing nutritional deficiencies due to impaired food and caloric intake is frequently observed [8]. For clinically relevant nutritional deficits that are associated with severe AH, published guidelines should be followed, to correct these conditions [9]. Alcohol abstinence is mandatory in all patients. For those patients with withdrawal risks, the use of Acamprosate (acetylhomotaurine) is advised [9].
Although commonly recommended in various publications [8,9,336], alcohol abstinence does not necessarily prevent short-term hospital lethality in patients that are likely abstinent under hospital surveillance, with lethality rates from 15% [8,311] up to 60% [311]. In such clinical settings of obligatory alcohol abstinence, most patients with AH will likely improve or at least stabilize [336]. Absolute alcohol abstinence leads back to a normal liver (27%), but it eventually progresses to liver cirrhosis (18%), or leads to chronic alcoholic hepatitis (56%); after three years of continued alcohol abstinence, half of these patients reach a normal liver. The conditions are different for those AH patients, who maintain the consumption of alcohol use, because they will develop cirrhosis at a higher rate (38%) or chronic alcoholic hepatitis (62%) (Figure 18) [336].
Figure 18.

Natural course of alcoholic hepatitis under absolute alcohol abstinence or continued alcohol use. Data are compiled from results published in a previous report [336].
Clearly, absolute alcohol abstinence must be the primary goal, not substitutable by reduced alcohol consumption (Figure 18). Many more clinical details can be retrieved from publications referenced recently [25], especially regarding guidelines issued by the AASLD (American Association of the Study of Liver Diseases) [9] and the EASL (European Association of the Study of the Liver) [8]. To evaluate patients with severe AH as potential candidates for liver transplantation, the following diagnostic instruments must be applied [7,8]: Maddrey score (Maddrey discriminant function); GAHS, Glasgow Alcoholic Hepatitis Score; ABIC score (that considers age, serum bilirubin, INR, and serum creatinine, leading to the acronym ABIC); and MELD score (Model-for-End-Stage-Liver-Disease) [25].
Liver transplantation is an option for selected patients with severe AH that are unresponsive to pharmacotherapy [25], but criteria of indications are yet disputed; they vary from country to country, and from continent to continent [8,9,10,11,12,13]. At least some consensus exists that the number of AH patients eligible for liver transplantation will remain low if the strict inclusion criteria for the waiting lists are followed and if donated organs are limited. The AASLD guidelines of 2010 are more cautious, considering recommendations by the American Society of Transplant Physicians and the AASLD, which had been published in 1998, and generally excluded AH patients from liver transplantation [5]. As opposed to AASLD, the EASL guidelines of 2010 mention the proposals of European and North American experts, who previously excluded AH as an indication for liver transplantation, reiterating that this has recently been challenged by a multi-center, case controlled study from France, which provided clear evidence of a higher survival rate in patients who benefitted from early liver transplantation.
Recent developments up to 2017 are summarized in four critical reviews, published on liver transplantation in AH [10,11,12,13]. In the first PRO transplantation article, Toniutto et al. from Italy favored early liver transplantation in a few selected patients with severe AH [10], in line with suggestions published in another PRO transplantation article by Artru et al. from France [11]. The third review was published from the US authored by Luce, who calls for a strong selection procedure, but otherwise titles his paper also as a PRO transplantation view [12]. Finally, the fourth review authored by Fung from Hong Kong was presented as a CON transplantation article [13]. For 2018, no actual guidelines of AASLD or EASL are to be expected, that would clarify the situation by providing a consensus paper on this issue.
13. Alcoholic Cirrhosis
Patients with AC often have no symptoms that would allow for diagnosis and in-time recommendations to abstain from alcohol use. Indeed, the diagnosis of AC would not be established unless until liver functions are heavily disturbed, causing symptoms, which is a common clinical experience. At hospital admission, when the diagnosis of alcoholic cirrhosis was established, in 24% of the patients, complications were not registered, 55% had only ascites, 6% experienced only bleeding from ruptured esophageal varices alone, 4% had variceal bleeding combined with ascites, and 11% had mental problems due to hepatic encephalopathy [337]. Likely due to the initially silent character of the disease, only about one third of the patients with alcoholic cirrhosis had been treated as in-patients before decompensation [8,338,339], a common experience among clinical hepatologists. However, the diseases progresses, and within the first year after admission, these patients are at increased risks of experiencing complications of liver decompensation. For instance, they face a 20% risk of developing ascites, a 6% risk of acute bleeding from ruptured esophageal varices, and a 4% risk of mental disturbances due to hepatic encephalopathy [8,337,339]. Early recognizable under professional health care surveillance, ascites are typically seen as the first complication [8], but other complications such as icterus, acute bleeding from ruptured esophageal varices, and mental disturbances caused by hepatic encephalopathy may also follow within a short period of time [8,337,339].
By liver histology, AC presents with thick collagen strands around the central vein and coursing through the hepatic lobules, also involving the perisinusoidal and pericellular areas [309]. In other areas, focal regeneration predominates, leading to regenerative cirrhotic nodules, mostly of the micronodular type. Regenerating nodules are clearly visible macroscopically on the liver surface with fragmented light reflexes (Figure 19).
Figure 19.

Macroscopic picture of alcoholic cirrhosis. The surface of alcoholic cirrhosis is granular, reflecting the regenerative nodules, which can be seen upon histological evaluation.
The pharmacotherapy of AC is disappointing. Among the specific therapies evaluated in clinical trials of patients with alcoholic cirrhosis, no benefit of outcome was achieved with S-adenosyl-l-methionine (SAMe), propylthiouracil, colchicine, anabolic-androgenic steroids, and silymarin [8]. Current management of alcoholic cirrhosis focuses on correcting nutritional deficiencies and alcohol abstinence, which substantially improves the 5-year survival rate in patients with compensated AC as well as decompensated AC (Figure 20) [338,340,341].
Figure 20.

Prognosis of alcoholic cirrhosis. Shown is the 5-year survival rate, prognosis is better in abstinent patients with compensated cirrhosis as compared to abstinent patients with decompensated cirrhosis. In both cohorts, continued alcohol use deteriorates the survival rate. The original figure was published in a previous report [341] and is reproduced with permission of the Publisher Wiley (Hoboken, NJ, USA).
Liver transplantation (LT) is principally the only chance for some selected patients with decompensated AC to escape the potential fatal outcome, but the number of organs available for LT is limited, and alternatively using split livers donated by family members is not a good approach, due to some mental reservations [8,9,10]. However, LTs are also under general discussion and basic considerations as to whether patients with a history of alcohol abuse should be candidates for a LT [8,9,10]. Although the AC-related lethality rate of 1.36% is low during the hospital stay [302], abstinent patients with decompensated AC have a 5-year survival chance of only around 60%, whereas 40% of this cohort will die (Figure 20) [338,339,341]. Clearly, patients with compensated AC are not candidates for a LT, because of their excellent prognosis with a 90% chance of survival during the next five years, provided that they abstain from alcohol use and adhere to long-term abstinence with professional support; starting and maintaining abstinence is crucial, since non-abstinent patients have a 5-year survival chance of only 45–60% (Figure 20).
For inclusion in the LT waiting list, only a few patients with decompensated AC will be eligible [8,9,10], although at this stage, the 5-year survival rate is low, even in abstinent patients, but more so in non-abstinent ones (Figure 20). Despite the high prevalence of decompensated AC in Western countries, 95% of these patients were never formally assessed for their candidacy for LT [342]. The appropriate patients should be evaluated for LT in a similar way as other patients with decompensated liver disease are assessed, focusing on a careful evaluation of their medical background and psychosocial condition [9]. To achieve a complete assessment, a formal evaluation of the likelihood of long-term abstinence is helpful, considering that a 6-month period of abstinence has been recommended by most centers as a minimal listing criterion [8,9,10]. This period allows issues related to alcohol abuse to be addressed in patients with recent problems of alcohol use, but more importantly, it enables sufficient improvement of the clinical condition prior to the planned liver transplantation, or even better, makes LT unnecessary [9]. Controversial discussions focus on the utility of the 6-month rule as a predictor of long-term sobriety, and additional information on details of other stricter criteria for LT has been published [8,9,10]. Meeting most of these prerequisites, the patient survival rate at five years after LT was 73% [343], and thereby better than the cohort of decompensated AC without LT, although transplanted patients are highly likely to drink after transplantation, but this obviously has no serious effect on the transplanted liver and the overall clinical course [8].
Many risk factors of AC are well established, based on clear evidence, others are based on results that are controversially discussed [9]; some of the risk factors are listed (Figure 21).
Figure 21.

Hypothesis of risk factors of alcoholic cirrhosis.
Abundant factors determine whether cirrhosis develops or not. Most of these cannot be influenced by individuals who drink alcohol, but some factors are under patients’ control, to minimize the risk of upcoming liver injury. Avoidable risk factors include the amount and duration of future alcohol consumption, obesity, and poor nutritional status. It also should be recognized that the development of AC mostly occurs silently. Patients in whom ALF has been diagnosed should abstain from alcohol use, since occasionally fibrosis and cirrhosis starts as central hyaline sclerosis syn. perivenular fibrosis at the stage of AFL, without the clinical warning symptoms of AH (Figure 10) [9,309], an interesting proposal of early diagnostic relevance [344]. Based on several reports [5,6,8,9,345], a percentage sequelae leading to AC is proposed (Figure 22).
Figure 22.

Development of alcoholic cirrhosis Good evidence exists that alcoholic hepatitis is responsible for most cases of alcoholic cirrhosis, but it may emerge also from alcoholic fatty liver with its perivenular and perisinusoidal fibrosis. Symbol: ?, pathway under discussion. The figure was published in a previous report [345], and is reproduced with permission of the Publisher Springer (Berlin, Germany).
Indeed, good clinical evidence is provided that fibrosis can occur in the absence of alcoholic hepatitis. This raises the question as to whether cell necrosis and inflammation are obligatory factors for alcohol-related hepatic stellate cell proliferation and activation and subsequent fibrogenesis and development of cirrhosis.
14. Alcoholic Hepatocellular Carcinoma
Patients with AC are at risk of AHCC, a rare complication of advanced ALD [346,347], but it can also can occur in alcoholic patients without AC [348]. During the development of AHCC, symptoms such as fatigue, weight loss, and anorexia increase in severity. To early recognize AHCC, patients with AC should have regular surveillance with ultrasound and the serum tumor marker α1-fetoprotein. Small localized AHCC can be treated by percutaneous ethanol injections; radiofrequency, microwave ablation, and liver segment resection are the ideal treatments [349], whereas for multifocal AHCC, only systemic chemotherapy or LT remains as a last option, disputed due to the problems of the final outcome [346,348]. The mechanisms leading to AHCC have not clearly been identified, but tumor initiation is likely triggered by ROS or acetaldehyde [346,347]. Experimentally, chronic alcohol consumption leading to AFL actually decreases tumor incidence by dimethylnitrosamine [83] and is protective for liver injury by dimethylnitrosamine [64,85], which is metabolized by CYP 2E1 [95,105]. Whether another carcinogen or procarcinogen may trigger AHCC is not known.
15. Actual Issues and Future Perspectives
A satisfactory unifying mechanism for individual susceptibility, initiation, and progression of alcoholic liver injury is not available [348]. Instead, some dozens of signaling mediators and mechanistic pathways have been presented, including CYP 2E1 and other topics, which are still challenging issues in experimental and clinical alcoholic liver injury, as results are in part contradictory, rarely confirmed, or discussed by other groups [349,350,351,352,353,354,355,356,357,358,359,360,361,362,363,364,365,366,367,368,369,370,371,372,373,374]. Much emphasis has previously been placed on the role of the endoplasmic reticulum as the principal localization of CYP 2E1, and the clinical and pathogenetic importance of this microsomal CYP 2E1, which is considered to play an essential role in endoplasmic reticulum (ER) stress syn. microsomal stress, thereby promoting liver injury [231,232,233,234,235,240,247,249,251,252], likely involving membrane lipid peroxides [348]. However, there is now growing evidence that this isoenzyme is also found in the mitochondrial compartment of the hepatocyte [351,352,353,356], an observation that raises a number of questions regarding the characteristics of this mitochondrial CYP 2E1, as compared to the previously well described microsomal variety [353,355]. In order to study this, special analytical techniques are required and were used, because conventional methods using ultracentrifugation to isolate liver mitochondria from liver homogenates are risky, due to possible contamination of the isolated intact mitochondria by microsomal fragments [353,356]. Among the preferred methods are those with focus on the proteasome complex and knockout and transgenic models [356,360]. More specifically, CYP 2E1 knockout models and transgenic models have been found to be valuable in describing a major functional role of CYP 2E1, as summarized and referenced in detail recently [358]. Using these CYP 2E1 knockout mice, a major role of CYP 2E1 was shown in experimental alcoholic fatty liver. In addition, overexpressed CYP 2E1 was found in a CYP 2E1 transgenic mouse model containing human CYP 2E1, which allowed for studies on the properties of human CYP 2E1, and provided corresponding validation. In these humanized CYP 2E1 knock-in mice, human CYP 2E1 transgene was introduced in the corresponding CYP 2E1 null mouse background [358]. Of some clinical interest are new results published on CYP 2E1 exosomes as possible diagnostic biomarkers in the blood of humans with alcoholism and microsomal stress [360]. They could help assign alcohol as a cause in a liver disease of unknown etiology, but in a clinical setting, other causes commonly have to be excluded, just to be on the safe side [25].
Notably, mitochondrial CYP 2E1 is induced by alcohol, as is microsomal CYP 2E1 [353]. The induction of microsomal CYP 2E1 results from enzyme stabilization by alcohol against degradation, and largely proceeds via a posttranscriptional process [283], while the proteasome complex is involved in regulating CYP 2E1 turnover [353]; induction of mitochondrial CYP 2E1 is caused by a different mechanism [353]. Preliminary studies suggest that mitochondrial CYP 2E1 induction may proceed through the stimulation of adenylate cyclase and N-terminal chimeric signals, activated by cyclic AMP-mediated phosphorylation of Ser-129, and supported by protein kinase A [351,353]. Mitochondrial CYP 2E1 further increases after six weeks of alcohol administration, whereas microsomal CYP 2E1 levels off earlier [353]. Some groups favor a more significant role of mitochondrial CYP 2E1 than microsomal CYP 2E1 promoting ALD [351,353,356], whereas other groups are more cautious and prefer microsomal CYP 2E1 as the primary promoting enzyme [234,355,358]. There is also uncertainty as to whether the CYP 2E1-dependent mitochondrial injury is due to ROS, the acetaldehyde generated from ethanol, or both. CYP 2E1 knockout and transgenic models have been developed, which have proven valuable in describing the contributions of overall CYP 2E1 to ALD, and to the toxicity of other toxins [358]. Despite much experimental data on CYP 2E1 being available, clinical studies have provided contradictory results regarding the question to what extent the polymorphism of CYP 2E1 contributes to human ALD, in line with many other risk factors of alcoholic cirrhosis that are under discussion (Figure 21).
A variety of other clinical aspects of CYP 2E1 merit discussion. In brief, and unrelated to alcohol abuse, an increase of CYP 2E1 is observed under various conditions including obesity [138,207,248,269,270,278,281,348] that renders obese individuals more susceptible to liver injury by acetaminophen that is metabolized via CYP 2E1, which appears to play a critical role [25,138,207,283,362]. Other CYP 2E1-dependent conditions relate to diabetes [109,207,352], fasting and acetone [97,107,119,167], nonalcoholic fatty liver [271], and nonalcoholic steatohepatitis [209,270]. In addition, alcohol abuse predisposes the individual to liver injury by carbon tetrachloride, a substrate for the ethanol-induced CYP 2E1; alcoholics are also be more susceptible to renal injury because renal CYP 2E1 may be induced by alcohol [39,257,272,374].
Sufficient evidence exists that microsomal CYP 2E1 likely plays an important role in ALD, more so than mitochondrial CYP 2E1 [252,358]. Based on present knowledge, a real alternative to microsomal CYP 2E1 promoting ALD is not apparent. Chronic alcohol consumption substantially increases microsomal CYP 2E1 in a process called microsomal or ER stress, which is associated with a high production of ROS as important intermediates initiating and perpetuating liver injury, apoptosis, and cell death [245,246,247,248]. ROS attacks various subcellular organelles by initiation lipid peroxidation, leading to a chain reaction that is hardly stopped unless protective compounds such as hepatic glutathione are available in sufficient amounts.
Nuclear genes of Sirtuin-1 (SIRT1) and Sirtuin-2 (SIRT2) are under broad discussion for their potential involvement in various conditions and diseases including ALD [364,365,366] and experimental lifespan extension and the longevity of Japanese centenarians in Okinawa [270]. In short, NAD-dependent deacetylase SIRT1 is involved in the regulation of mitochondrial energy metabolism [364,365,366]. Prolonged alcohol use inhibited hepatic SIRT1 mRNA by 50%, and impaired the protein expression of SIRT1 that was restored by replacing long-chain triglycerides (LCT) with medium chain triglycerides (MCT) in the experimental alcohol diet, suggesting that that mitochondrial dysfunction due to alcohol abuse can be treated by dietary modifications. Other studies support the view that SIRT1 is involved in experimental alcoholic liver injury and overexpressed in human HCC, studied in human HCC cells that retain ADH and CYP 2E1 [365]. Inhibited by alcohol, SIRT1 deregulates hepatic pathways of lipid metabolism such as lipogenesis, fatty acid β-oxidation, and lipoprotein uptake, and pathways of inflammatory response, including expression of pro-inflammatory cytokines [366], substantiating thereby the important role of SIRT1 signaling in the initiation of AFL.
There is growing evidence that in addition to the known direct toxic effects of alcohol and acetaldehyde, immune reactions may partly be responsible for ALD, but the underlying mechanisms are yet poorly understood, and their individual quantitative impact on disease initiation and perpetuation is, to a large part, not well established and a matter of some debate [22,368,369,370]. A partial pathogenetic role of immune and autoimmune reactions in ALD can be assumed because the majority of patients with AH are successfully treated with corticosteroids [25,317,318,319,320,321,322,323,324,325,326,327,328,329,330,331,332,333,334,335], as opposed to a few others who do not benefit from this treatment, possibly due the fact that their liver disease is caused by other mechanisms that are unrelated to an immunity background [25]. Alcohol can modify both, the innate immune system (IIS) and the adaptive immune system (AIS) [370]. In short, IIS is promoted by macrophages, Kupffer cells, neutrophils, and natural killer cells, whereby macrophages are prepared to attack antigens of bacterial cell walls and respond by providing cytokines. Kupffer cells in alcoholic liver disease are specifically activated by lipopolysaccharide (LPS), also called endotoxins, and derived from the outer membrane of Gram-negative intestinal bacteria [370].
Endotoxins are considered to play a major pathogenetic role in ALD [348,354,357,371]. New experimental studies revealed that binge alcohol use causes gut leakage and endotoxemia through induction of both intestinal and hepatic CYP 2E1 [357], in support of the microbiome hypothesis that endotoxins produced by intestinal bacteria cross the leaky intestinal mucosa and contribute to hepatic apoptosis and steatohepatitis [354,357], which is still a major clinical issue [348].
Additional discussions focus on how alcohol results in the derangement of metabolic pathways. Among these topics are carotenoids [82,93,103,142,222,367], known for their protective properties against liver injury mediated by ROS [367]. Patients with alcoholic cirrhosis had a 25-fold decreased hepatic level of α- and β-carotene, and more than half of these patients had normal blood levels, suggesting that liver disease modulates intestinal resorption, metabolic pathways, and the removal of α- and β-carotene.
Finally, little attention has been paid in the alcohol literature on the circadian rhythms, which are neglected in virtually all publications that are referenced in this article (Table 3), with the exception of one report published in 1984, describing circadian rhythms of ADH and MEOS in an experimental study [90]. The circadian peak in ADH activity fell near the time of maximal blood ethanol clearance rates both in groups of rats injected with a single ethanol dose (acute group), and in rats continuously exposed to ethanol for 22 weeks (chronic group). However, at all time points, investigated ADH activity levels were lower, and they fluctuated less in the chronic group than in either the acute or control (ethanol naive) groups. In contrast, MEOS activity levels revealed a prominent rhythm that was 180 degrees out of phase with the ADH rhythm in the chronic group, while MEOS activity showed very low levels in the acute and control groups and did not vary over the circadian span. Similarly, alcoholic liver injury is under the control of a molecular circadian clock [373], which is also found in plants and it may have negative effects on their quality [278,279]. Evidence from alcohol studies in experimental animals and clinically in humans presented close associations among circadian processes, alcohol use, and liver injury. In particular, alcohol influences the expression of genes that dominate the circadian clock and regulate metabolic pathways [373]. Consequently, alcohol-based disruption in circadian rhythms may contribute to the initiation and perpetuation of alcoholic liver injury.
16. Conclusions
Alcoholic liver disease remains a fascinating topic in clinical and experimental hepatology, and it has gained much input with the discovery of the microsomal ethanol-oxidizing system (MEOS) by Charles S. Lieber and Leonore M. DeCarli, 50 years ago. Some pathogenetic events are linked to acetaldehyde as the first oxidation product that is generated from ethanol, and to cytochrome P450 2E1 as an obligatory component of MEOS. CYP 2E1 produces reactive oxygen species (ROS) such as the ethoxy radical CH3CH2O, the hydroxyethyl radical CH3C(·)HOH, the acetyl radical CH3CHO, the singlet radical 1O2, the superoxide radical HO2, hydrogen peroxide (H2O2), the hydroxyl radical HO, the alkoxyl radical RO, and the peroxyl radical ROO. They attack hepatocytes, Kupffer cells, stellate cells, and liver sinusoidal endothelial cells, and their signaling mediators initiate liver injury, including fibrosis and cirrhosis. Human ALD benefits from appropriate animals, for which experimental models of alcoholic fatty liver are available, allowing for studies on pathogenetic mechanisms. More evidence is now emerging that lipid peroxides, which are generated through ROS and microsomal stress in the membrane of the endoplasmic reticulum, can initiate and perpetuate alcoholic liver injury in joint actions with signaling mediators derived from nonparenchymal cells and hepatocytes. A major pathogenetic role is also assumed for endotoxins that originate from intestinal bacteria and the leaky gut, which results from alcohol-linked modifications of the intestinal microbiome likely as a consequence of hepatic and intestinal CYP 2E1.
Other more recent studies focus on the alcohol-linked inhibition of nuclear genes of Sirtuin-1 (SIRT1). This impairs protein expression and deregulates hepatic pathways, not only of lipid metabolism such as lipogenesis, fatty acid β-oxidation, and lipoprotein uptake, but also of inflammatory responses, including the expression of pro-inflammatory cytokines, thereby substantiating an important role of SIRT1 signaling in the initiation of AFL. The innate immune and adaptive immune systems are modified by alcohol and both could contribute to the individual susceptibility of severe ALD stages, such as AH and AC, which affect only a minority of individuals with alcohol abuse. Finally, and in analogy to the circadian clock in plants, the alcoholic liver is under the control of circadian rhythms that influence many metabolic pathways in the liver, thereby contributing to the natural course of alcoholic liver disease. It is obvious that a unifying mechanism leading to liver injury is not available, but many processes and active mediators are under discussion that may interact among each other. Although these variable pathomechanisms are of interest for biochemists, clinical considerations should focus more on patients with their risk of alcoholic liver injury, and on the early detection of alcoholic fatty liver to prevent disease progression, which is achievable by strict abstinence from alcohol use.
Acknowledgments
The author dedicates this scientific article to Georg Strohmeyer at the occasion of his 90th birthday. As the Head of the Medical Department of Gastroenterology, Hepatology and Infectious Diseases at the Heinrich Heine University in Düsseldorf (Germany), he was not only an outstanding physician and an encouraging scientist, but also a professional clinical mentor for the author from 1975 to 1987.
Abbreviations
| ABIC | Age, bilirubin, INR, and creatinine score |
| AASLD | American Association of the Study of Liver Diseases |
| AC | Alcoholic cirrhosis |
| ADH | Alcohol dehydrogenase |
| AFL | Alcoholic fatty liver |
| ALD | Alcoholic liver disease |
| ALDH | Acetaldehyde dehydrogenase |
| AH | Alcoholic hepatitis |
| AHCC | Alcoholic hepatocellular carcinoma |
| ALT | Alanine transaminase |
| ASH | Alcoholic steatohepatitis |
| AST | Aspartate transaminase |
| CDT | Carbohydrate-deficient transferrin |
| CLD | Chronic liver diseases |
| CYP | Cytochrome P450 |
| CYP2E1 | Cytochrome P450 2E1 |
| DILI | Drug induced liver injury |
| EASL | European Association for the Study of the Liver |
| FPM | First pass metabolism |
| GAHS | Glasgow Alcoholic Hepatitis Score |
| GDH | Glutamate dehydrogenase |
| GGT | Gamma-glutamyltransferase |
| HBV | Hepatitis B virus |
| HCV | Hepatitis C virus |
| HILI | Herb-induced liver injury |
| K m | Michaelis-Menten constant |
| MCV | Mean corpuscular volume of erythrocytes |
| MDB | Mallory-Denk bodies |
| MDF | Maddrey Discriminant Function scale |
| MELD | Model for End-stage Liver Disease score |
| MEOS | Microsomal ethanol-oxidizing system |
| MAST | Michigan Alcoholism Screening Test |
| NAFLD | Nonalcoholic fatty liver disease |
| NASH | Nonalcoholic steatohepatitis |
| PTU | Propylthiouracil |
| ROS | Reactive oxygen species |
| RUCAM | Roussel Uclaf Causality Assessment Method |
Funding
There was no funding of this article.
Conflicts of Interest
The author has no conflict of interest to declare, with respect to this invited review article.
References
- 1.Ingólfsson H.I., Anderson O.S. Alcohol’s effects on lipid bilayer properties. Biophys. J. 2011;101:847–855. doi: 10.1016/j.bpj.2011.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Sergent O., Djoudi-Aliche F., Lagadic-Gossmann D. Up-to date insight about membrane remodeling as a mechanism of action for ethanol-induced liver toxicity. In: Shimizu I., editor. Trends in Alcoholic Liver Disease–Clinical and Scientific Aspects. InTech; London, UK: [(accessed on 26 October 2018)]. Available online: https://cdn.intechopen.com/pdfs-wm/25884.pdf. [Google Scholar]
- 3.Dopico A.M., Lovinger D.M. Acute alcohol action and desensibilization of ligandgated ion channels. Pharmacol. Rev. 2009;61:98–114. doi: 10.1124/pr.108.000430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lieber C.S. Alcoholic fatty liver: Its pathogenesis and mechanism of progression to inflammation and fibrosis. Alcohol. 2004;34:9–19. doi: 10.1016/j.alcohol.2004.07.008. [DOI] [PubMed] [Google Scholar]
- 5.Gao B., Bataller R. Alcoholic liver disease: Pathogenesis and new therapeutic targets. Gastroenterology. 2011;141:1572–1585. doi: 10.1053/j.gastro.2011.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sevastianos V.A., Dourakis S.P. Alcoholic liver disease: A clinical review. J. Nutr. Food Sci. 2016;6:508. doi: 10.4172/2155-9600.1000508. [DOI] [Google Scholar]
- 7.Mathurin P., Lucey M.R. Management of alcoholic hepatitis. J. Hepatol. 2012;56:S39–S45. doi: 10.1016/S0168-8278(12)60005-1. [DOI] [PubMed] [Google Scholar]
- 8.EASL 2010 Guidelines: Management of Alcoholic Liver Disease. [(accessed on 28 April 2018)]; Available online: http://www.easl.eu/research/our-contributions/clinical-practice-guidelines/detail/management-of-alcoholic-liver-disease-easl-clinical-practice-guidelines/report/1.
- 9.O’Shea R.S., Dasarathy S., McCullough A.J. Alcoholic liver disease. [(accessed on 28 April 2018)];Am. J. Gastroenterol. 2010 105:14–32. doi: 10.1038/ajg.2009.593. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19904248. [DOI] [PubMed] [Google Scholar]
- 10.Toniutto P., Zanetto A., Ferrarese A., Burra P. Current challenges and future directions for liver transplantation. Liver Int. 2017;37:317–327. doi: 10.1111/liv.13255. [DOI] [PubMed] [Google Scholar]
- 11.Artru F., Louvet A., Mathurin P. Liver transplantation for patients with alcoholic hepatitis. Liver Int. 2017;37:337–339. doi: 10.1111/liv.13248. [DOI] [PubMed] [Google Scholar]
- 12.Fung J.Y.Y. Liver transplantation for severe alcoholic hepatitis–The CON view. Liver Int. 2017;37:340–342. doi: 10.1111/liv.13286. [DOI] [PubMed] [Google Scholar]
- 13.Lucey M.R. Liver transplantation for severe alcoholic hepatitis—The PRO view. Liver Int. 2017;37:343–344. doi: 10.1111/liv.13343. [DOI] [PubMed] [Google Scholar]
- 14.WHO . Global Status Report on Alcohol and Health. World Health Organization; Geneva, Switzerland: 2011. [(accessed on 20 April 2018)]. Available online: http://www.who.int/substance_abuse/publications/global_alcohol_report/msbgsruprofiles.pdf?ua=1. [Google Scholar]
- 15.Rehm J., Samokhvalov A.V., Shield K.D. Global burden of alcoholic liver diseases. J. Hepatol. 2013;59:160–168. doi: 10.1016/j.jhep.2013.03.007. [DOI] [PubMed] [Google Scholar]
- 16.Lodish H., Berk A., Zipursky S.L., Matsudaira P., Baltimore D., Darnell J. Molecular Cell Biology. 4th ed. W. H. Freeman; New York, NY, USA: 2000. [(accessed on 26 October 2018)]. Molecular analysis of photosystems. Available online: http://www.ncbi.nlm.nih.gov/books/NBK21484/ [Google Scholar]
- 17.Berg J.M., Tymoczko J.L., Stryer L. Biochemistry. 5th ed. W. H. Freeman; New York, NY, USA: 2002. [(accessed on 26 October 2018)]. The Calvin cycle synthesizes hexoses from carbon dioxide and water. Available online: http://www.ncbi.nlm.nih.gov/books/NBK22344/ [Google Scholar]
- 18.Salaspuro M. Epidemiological aspects of alcohol and alcoholic liver disease, ethanol metabolism, and pathogenesis of alcoholic liver injury. In: Bircher J., Benhamou J.P., McIntyre N., Rizzetto N., Rodes J., editors. Oxford Textbook of Clinical Hepatology. Oxford University Press; Oxford, UK: 1999. pp. 791–810. [Google Scholar]
- 19.Mitchell M.C., Teigen E.L., Ramchandani V.A. Absorption and peak blood alcohol concentration after drinking beer, wine, or spirits. Alcohol. Clin. Exp. Res. 2014;38:1200–1204. doi: 10.1111/acer.12355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Caballeria J., Frezza M., Hernández-Muñoz R., DiPadova C., Korsten M.A., Baraona E., Lieber C.S. Gastric origin of the first-pass metabolism of ethanol in humans: Effect of gastrectomy. Gastroenterology. 1989;97:1205–1209. doi: 10.1016/0016-5085(89)91691-0. [DOI] [PubMed] [Google Scholar]
- 21.Frezza M., di Padova C., Pozzato G., Terpin M., Baraona E., Lieber C.S. High blood alcohol levels in women—Role of decreased gastric alcohol dehydrogenase activity and first-pass metabolism. N. Engl. J. Med. 1990;322:95–99. doi: 10.1056/NEJM199001113220205. [DOI] [PubMed] [Google Scholar]
- 22.Lieber C.S. Alcohol and the liver: 1994 update. Gastroenterology. 1994;106:1085–1105. doi: 10.1016/0016-5085(94)90772-2. [DOI] [PubMed] [Google Scholar]
- 23.Teschke R., Gellert J. Hepatic microsomal ethanol-oxidizing system (MEOS): Metabolic aspects and clinical implications. Alcohol. Clin. Exp. Res. 1986;10:20S–32S. doi: 10.1111/j.1530-0277.1986.tb05176.x. [DOI] [PubMed] [Google Scholar]
- 24.Dasgupt A. Genetic polymorphisms of alcohol metabolizing enzymes associated with protection from or increased risk of alcohol. In: Dasgupt A., editor. Alcohol, Drugs, Genes and the Clinical Laboratory. Elsevier; Amsterdam, The Netherlands: 2017. pp. 107–116. [Google Scholar]
- 25.Teschke R. Alcoholic steatohepatitis (ASH) and acute alcoholic hepatitis (AH): Cascade of events, clinical features, and pharmacotherapy options. Exp. Opin. Pharmacother. 2018;19:779–793. doi: 10.1080/14656566.2018.1465929. [DOI] [PubMed] [Google Scholar]
- 26.Lieber C.S., DeCarli L.M. Ethanol oxidation by hepatic microsomes: Adaptive increase after ethanol feeding. Science. 1968;162:917–918. doi: 10.1126/science.162.3856.917. [DOI] [PubMed] [Google Scholar]
- 27.Lieber C.S., DeCarli L.M. Hepatic microsomal ethanol-oxidizing system. In vitro characteristics and adaptive properties in vivo. J. Biol. Chem. 1970;245:2505–2512. [PubMed] [Google Scholar]
- 28.Lieber C.S., DeCarli L.M. Reduced nicotinamide-adenine dinucleotide phosphate oxidase: Enhanced by ethanol consumption. Science. 1970;170:78–80. doi: 10.1126/science.170.3953.78. [DOI] [PubMed] [Google Scholar]
- 29.Lieber C.S., DeCarli L.M. The role of the hepatic microsomal ethanol-oxidzing system (MEOS) for ethanol metabolism in vivo. J. Pharmacol. Exp. Ther. 1972;181:279–287. [PubMed] [Google Scholar]
- 30.Teschke R., Hasumura Y., Joly J.G., Ishii H., Lieber C.S. Microsomal ethanol-oxidizing system (MEOS): Purification and properties of a rat liver system free of catalase and alcohol dehydrogenase. Biochem. Biophys. Res. Commun. 1972;49:1187–1193. doi: 10.1016/0006-291X(72)90594-3. [DOI] [PubMed] [Google Scholar]
- 31.Ishii H., Joly J.G., Lieber C.S. Effect of ethanol on the amount and enzyme activities of hepatic rough and smooth microsomal membranes. Biochim. Biophys. Acta. 1973;291:411–420. doi: 10.1016/0005-2736(73)90493-8. [DOI] [PubMed] [Google Scholar]
- 32.Joly J.G., Ishii H., Teschke R., Hasumura Y., Lieber C.S. Effect of chronic ethanol feeding on the activities of and submicrosomal distribution of reduced nicotinamide adenine dinucleotide phosphate-cytochrome P-450 reductase and the demethylases for aminopyrine and ethylmorphine. Biochem. Pharmacol. 1973;22:1532–1535. doi: 10.1016/0006-2952(73)90334-1. [DOI] [PubMed] [Google Scholar]
- 33.Joly J.G., Feinman L., Ishii H., Lieber C.S. Effect of chronic ethanol feeding on hepatic microsomal glycerophosphate acyltransferase activity. J. Lipid Res. 1973;14:337–343. [PubMed] [Google Scholar]
- 34.Ishii H., Joly J.G., Lieber C.S. Increase of microsomal glucose-6-phosphatase activity after chronic ethanol administration. Metabolism. 1973;22:799–806. doi: 10.1016/0026-0495(73)90050-4. [DOI] [PubMed] [Google Scholar]
- 35.Mezey E., Potter J.J., Reed W.D. Ethanol oxidation by a component of liver microsomes rich in cytochrome P-450. J. Biol. Chem. 1973;248:1183–1187. [PubMed] [Google Scholar]
- 36.Teschke R., Hasumura Y., Lieber C.S. NADPH-dependent oxidation of methanol, ethanol, propanol, and butanol by hepatic microsomes. Biochem. Biophys. Res. Commun. 1974;60:851–857. doi: 10.1016/0006-291X(74)90319-2. [DOI] [PubMed] [Google Scholar]
- 37.Lieber C.S., DeCarli L.M. Oxidation of ethanol by hepatic microsomes of acatalasemic mice. Biochem. Biophys. Res. Commun. 1974;60:1187–1192. doi: 10.1016/0006-291X(74)90324-6. [DOI] [PubMed] [Google Scholar]
- 38.Teschke R., Hasumura Y., Lieber C.S. Hepatic microsomal ethanol oxidizing system: Solubilization, isolation and characterization. Arch. Biochem. Biophys. 1974;163:404–415. doi: 10.1016/0003-9861(74)90492-5. [DOI] [PubMed] [Google Scholar]
- 39.Hasumura Y., Teschke R., Lieber C.S. Increased carbon tetrachloride hepatotoxicity, and its mechanism, after chronic ethanol consumption. Gastroenterology. 1974;66:226–234. [PubMed] [Google Scholar]
- 40.Joly J.G., Hétu C. Effects of chronic ethanol administration in the rat. Relative dependency on dietary lipids—I. Induction of hepatic microsomal drug-metabolizing enzymes in vitro. Biochem. Pharmacol. 1975;24:1475–1480. doi: 10.1016/0006-2952(75)90021-0. [DOI] [PubMed] [Google Scholar]
- 41.Lieber C.S., Hasumura Y., Teschke R., Matsuzaki S., Korsten M. The effect of chronic ethanol consumption on acetaldehyde metabolism. In: Lindros K.O., Eriksson C.J.P., editors. The Role of Acetaldehyde in the Actions of Ethanol. Volume 23. The Finnish Foundation for Alcohol Studies; Helsinki, Finland: 1975. pp. 83–104. [Google Scholar]
- 42.Teschke R., Hasumura Y., Lieber C.S. Hepatic microsomal alcohol-oxidizing system: Affinity for methanol, ethanol, propanol and butanol. J. Biol. Chem. 1975;250:7397–7404. [PubMed] [Google Scholar]
- 43.Teschke R., Hasumura Y., Lieber C.S. Hepatic microsomal alcohol-oxidizng system in normal and acatalasemic mice: Its dissociation from the peroxidatic activity of catalase-H2O2. Mol. Pharmacol. 1975;11:841–849. [PubMed] [Google Scholar]
- 44.Ullrich V., Weber P., Wollenbeg P. Tetrahydrofurane—An inhibitor for ethanol-induced liver micosomal cytochrome P-450. Biochem. Biophys. Res. Commun. 1975;64:808–813. doi: 10.1016/0006-291X(75)90119-9. [DOI] [PubMed] [Google Scholar]
- 45.Lieber C.S., DeCarli L.M., Feinman L., Hasumura Y., Korsten M., Matsuzaki S., Teschke R. Effect of chronic alcohol consumption on ethanol and acetaldehyde metabolism. In: Gross M.M., editor. Alcohol Intoxication and Withdrawal: Experimental Studies II. Volume 59. Plenum Press; New York, NY, USA: 1975. pp. 185–227. Advances in Experimental Medicine and Biology. [DOI] [PubMed] [Google Scholar]
- 46.Hasumura Y., Teschke R., Lieber C.S. Acetaldehyde oxidation by hepatic mitochondria: Its decrease after chronic ethanol consumption. Science. 1975;189:727–729. doi: 10.1126/science.168641. [DOI] [PubMed] [Google Scholar]
- 47.Hasumura Y., Teschke R., Lieber C.S. Characteristics of acetaldehyde oxidation in rat liver mitochondria. J. Biol. Chem. 1976;251:4908–4913. [PubMed] [Google Scholar]
- 48.Teschke R., Hasumura Y., Lieber C.S. Hepatic ethanol metabolism: Respective roles of alcohol dehydrogenase, the microsomal ethanol-oxidizing system, and catalase. Arch. Biochem. Biophys. 1976;175:635–643. doi: 10.1016/0003-9861(76)90554-3. [DOI] [PubMed] [Google Scholar]
- 49.Ohnishi K., Lieber C.S. Reconstitution of the microsomal ethanol-oxidizing system. Qualitative and quantitative changes of cytochrome P-450 after chronic ethanol consumption. J. Biol. Chem. 1977;252:7124–7131. [PubMed] [Google Scholar]
- 50.Cederbaum A.I., Dicker E., Rubin E., Lieber C.S. The effect of dimethylsulfoxide and other radical scavengers on the oxidation of ethanol by rat liver microsomes. Biochem. Biophys. Res. Commun. 1977;78:1254–1562. doi: 10.1016/0006-291X(77)91428-0. [DOI] [PubMed] [Google Scholar]
- 51.Joly J.G., Villeneuve J.P., Mavier P. Chronic ethanol administration induces a form of cytochrome P-450 with specific spectral and catalytic properties. Alcohol. Clin. Exp. Res. 1977;1:17–20. doi: 10.1111/j.1530-0277.1977.tb05760.x. [DOI] [PubMed] [Google Scholar]
- 52.Teschke R., Ohnishi K., Hasumura Y., Lieber C.S. Hepatic microsomal ethanol oxidizing system: Isolation and reconstitution. In: Ullrich V., Roots I., Hildebrandt A., Estabrook R.W., Conney A.H., editors. Microsomes and Drug Oxidations. Pergamon Press; Oxford, UK: 1977. pp. 103–110. [Google Scholar]
- 53.Teschke R., Matsuzaki S., Ohnishi K., DeCarli L.M., Lieber C.S. Microsomal ethanol oxidizing system (MEOS): Current status of its characterization and its role. Alcohol. Clin. Exp. Res. 1977;1:7–15. doi: 10.1111/j.1530-0277.1977.tb05759.x. [DOI] [PubMed] [Google Scholar]
- 54.Teschke R., Matsuzaki S., Ohnishi K., Hasumura Y., Lieber C.S. Metabolism of alcohol at high concentrations: Role and biochemical nature of the hepatic microsomal ethanol oxidizing system. In: Gross M.M., editor. Advances in Experimental Medicine and Biology. Plenum Press; New York, NY, USA: 1977. pp. 257–280. Vol 85A—Alcohol Intoxication and Withdrawal–IIIa. [DOI] [PubMed] [Google Scholar]
- 55.Lieber C.S., DeCarli L.M., Matsuzaki S., Ohnishi K., Teschke R. The microsomal ethanol-oxidizing system. In: Fleischer S., Packer L., editors. Methods in Enzymology. Academic Press; New York, NY, USA: 1978. pp. 355–368. [DOI] [PubMed] [Google Scholar]
- 56.Ohnishi K., Lieber C.S. Respective role of superoxide and hydroxyl radical in the activity of the reconstituted microsomal ethanol-oxidizing system. Arch. Biochem. Biophys. 1978;191:798–803. doi: 10.1016/0003-9861(78)90422-8. [DOI] [PubMed] [Google Scholar]
- 57.Miwa G.T., Lewin W., Thomas P.E., Lu A.Y. The direct oxidation of ethanol by a catalase- free and alcohol dehydrogenase-free reconstituted system containing cytochrome P-450. Arch. Biochem. Biophys. 1978;187:464–475. doi: 10.1016/0003-9861(78)90058-9. [DOI] [PubMed] [Google Scholar]
- 58.Fabry T.L., Lieber C.S. The photochemical action spectrum of the microsomal ethanol oxidizing system. Alcohol. Clin. Exp. Res. 1979;3:219–224. doi: 10.1111/j.1530-0277.1979.tb05303.x. [DOI] [PubMed] [Google Scholar]
- 59.Seitz H.K., Korsten M.A., Lieber C.S. Ethanol oxidation by intestinal microsomes: Increased activity after 303 chronic ethanol administration. Life Sci. 1979;25:1443–1448. doi: 10.1016/0024-3205(79)90422-3. [DOI] [PubMed] [Google Scholar]
- 60.Moreno F., Teschke R., Strohmeyer G. Effect of thyroid hormones on the activities of hepatic alcohol metabolizing enzymes. Biochem. Biophys. Res. Commun. 1979;89:806–812. doi: 10.1016/0006-291X(79)91850-3. [DOI] [PubMed] [Google Scholar]
- 61.Teschke R., Stutz G., Strohmeyer G. Increased paracetamol-induced hepatotoxicity after chronic alcohol consumption. Biochem. Biophys. Res. Commun. 1979;91:368–374. doi: 10.1016/0006-291X(79)90628-4. [DOI] [PubMed] [Google Scholar]
- 62.Teschke R., Stutz G., Moreno F. Cholestasis following chronic alcohol consumption: Enhancement after an acute dose of chlorpromazine. Biochem. Biophys. Res. Commun. 1980;94:1013–1020. doi: 10.1016/0006-291X(80)91335-2. [DOI] [PubMed] [Google Scholar]
- 63.Burnett K.G., Felder M.R. Ethanol metabolism in peromyscus genetically deficient in alcohol dehydrogenase. Biochem. Pharmacol. 1980;29:125–130. doi: 10.1016/0006-2952(80)90318-4. [DOI] [PubMed] [Google Scholar]
- 64.Gellert J., Moreno F., Haydn M., Oldiges H., Frenzel H., Teschke R., Strohmeyer G. Decreased hepatotoxicity of dimethylnitrosamine (DMN) following chronic alcohol consumption. In: Thurman R.G., editor. Alcohol and Aldehyde Metabolizing Systems—IV. Volume 132. Plenum Press; New York, NY, USA: 1980. pp. 237–243. Advances in Experimental Medicine and Biology. [DOI] [PubMed] [Google Scholar]
- 65.Cederbaum A.I., Cohen G. Oxidative demethylation of t-butyl alcohol in rat liver microsomes. Biochem. Biophys. Res. Commun. 1980;97:730–736. doi: 10.1016/0006-291X(80)90325-3. [DOI] [PubMed] [Google Scholar]
- 66.Moreno F., Minzlaff M., Hauptmeier K.H., Teschke R. Alterations of hepatic alcohol metabolizing enzyme activities due to thyroid hormones. Adv. Exp. Med. Biol. 1980;132:109–115. doi: 10.1007/978-1-4757-1419-7_12. [DOI] [PubMed] [Google Scholar]
- 67.Cederbaum A.I., Qureshi A., Messenger P. Oxidation of isopropanol by rat liver micorosomes. Possible role of hydroxyl radicals. Biochem. Pharmacol. 1981;30:825–831. doi: 10.1016/S0006-2952(81)80002-0. [DOI] [PubMed] [Google Scholar]
- 68.Teschke R., Moreno F., Petrides A.S. Hepatic microsomal ethanol-oxidizing system (MEOS): Respective roles of ethanol and carbohydrates for the enhanced activity after chronic alcohol consumption. Biochem. Pharmacol. 1981;30:1745–1751. doi: 10.1016/0006-2952(81)90004-6. [DOI] [PubMed] [Google Scholar]
- 69.Seitz H.K., Garro A.J., Lieber C.S. Enhanced pulmonary and intestinal activation of procarcinogens and mutagens after chronic ethanol consumption in the rat. Eur. J. Clin. Investig. 1981;11:33–38. doi: 10.1111/j.1365-2362.1981.tb01762.x. [DOI] [PubMed] [Google Scholar]
- 70.Moreno F., Petrides A.S., Heinen E., Strohmeyer G., Teschke R. Hepatic microsomal ethanol-oxidizing system (MEOS): Increased activity following propylthiouracil administration. Alcohol. Clin. Exp. Res. 1981;5:85–91. doi: 10.1111/j.1530-0277.1981.tb04870.x. [DOI] [PubMed] [Google Scholar]
- 71.Sato C., Matsuda Y., Lieber C.S. Increased hepatotoxicity of acetaminophen after chronic ethanol consumption in the rat. Gastroenterology. 1981;80:140–148. [PubMed] [Google Scholar]
- 72.Teschke R., Heymann K. Effect of sex hormones on the activities of hepatic alcohol-metabolizing enzymes in male rats. Enzyme. 1982;28:268–277. doi: 10.1159/000459111. [DOI] [PubMed] [Google Scholar]
- 73.Koop D.R., Moran E.T., Tarr G.E., Coon M.J. Purification and characterization of a unique isozyme of cytochrome P-450 from liver microsomes of ethanol-treated rabbits. J. Biol. Chem. 1982;257:8472–8480. [PubMed] [Google Scholar]
- 74.Morgan E.T., Koop D.R., Coon M.J. Catalytic activity of cytochrome P-450 isozyme 3a isolated from liver microsomes of ethanol-treated rabbits. J. Biol. Chem. 1982;257:13951–13957. [PubMed] [Google Scholar]
- 75.Ingelman-Sundberg M., Hagbjörk A.L. On the significance of cytochrome P-450-dependent hydroxyl radical-mediated oxygenation mechanism. Xenobiotica. 1982;12:673–686. doi: 10.3109/00498258209038943. [DOI] [PubMed] [Google Scholar]
- 76.Petersen D.R., Atkinson N., Hjelle J.J. Increase in hepatic microsomal oxidation by a single dose of ethanol. J. Pharmacol. Exp. Ther. 1982;221:275–281. [PubMed] [Google Scholar]
- 77.Teschke R., Wiese B. Sex-dependency of hepatic alcohol-metabolizing enzymes. J. Endocrinol. Investig. 1982;5:243–250. doi: 10.1007/BF03348330. [DOI] [PubMed] [Google Scholar]
- 78.Damgaard S.E. The D(V/K) isotope effect of the cytochrome P-450-mediated oxidation of ethanol and its biological applications. Eur. J. Biochem. 1982;125:593–603. doi: 10.1111/j.1432-1033.1982.tb06724.x. [DOI] [PubMed] [Google Scholar]
- 79.Leo M.A., Lieber C.S. Hepatic vitamin A depletion in alcoholic liver injury. N. Engl. J. Med. 1982;307:597–601. doi: 10.1056/NEJM198209023071006. [DOI] [PubMed] [Google Scholar]
- 80.Seitz H.K., Bösche P., Czygan P., Veith S., Kommerell B. Microsomal ethanol oxidation in the colonic mucosa of the rat: Effect of chronic ethanol ingestion. N-S Arch. Pharmacol. 1982;320:81–84. doi: 10.1007/BF00499078. [DOI] [PubMed] [Google Scholar]
- 81.Teschke R., Bolsen K., Landmann H., Goerz G. Effect of hexachlorobenzene on the activities of hepatic alcohol-metabolizing enzymes. Biochem. Pharmacol. 1983;32:1745–1751. doi: 10.1016/0006-2952(83)90120-X. [DOI] [PubMed] [Google Scholar]
- 82.Leo M.A., Lieber C.S. Interaction of ethanol with vitamin A. Alcohol. Clin. Exp. Res. 1983;7:15–21. doi: 10.1111/j.1530-0277.1983.tb05404.x. [DOI] [PubMed] [Google Scholar]
- 83.Teschke R., Minzlaff M., Oldiges H., Frenzel H. Effect of chronic alcohol consumption on tumor incidence due to dimethylnitrosamine administration. J. Cancer Res. Clin. Oncol. 1983;106:58–64. doi: 10.1007/BF00399898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Teschke R., Hauptmeier K.H., Frenzel H. Effect of an acute dose of ethanol on the hepatotoxicity due to carbon tetrachloride. Liver Int. 1983;3:100–109. doi: 10.1111/j.1600-0676.1983.tb00856.x. [DOI] [PubMed] [Google Scholar]
- 85.Teschke R. Effect of chronic alcohol pretreatment on the hepatotoxicity elicited by chlorpromazine, paracetamol, and dimethylnitrosamine. Biol. Approach Alcohol. Natl. Inst. Alcohol Abuse Alcohol. Res. Monogr. 1983;11:170–189. [Google Scholar]
- 86.Nomura F., Pikkarainen P., Jauhonen M., Arai E.R., Gordon E., Baraona E., Lieber C.S. Effect of ethanol administration on the metabolism of ethanol in baboons. J. Pharmacol. Exp. Ther. 1983;227:78–83. [PubMed] [Google Scholar]
- 87.Shigeta Y., Nomura F., Leo M.A., Iida S., Felder M.R., Lieber C.S. Alcohol dehydrogenase (ADH) independent ethanol metabolism in deermice lacking ADH. Pharmacol. Biochem. Behav. 1983;18(Suppl. 1):195–199. doi: 10.1016/0091-3057(83)90171-5. [DOI] [PubMed] [Google Scholar]
- 88.Koop D.R., Nordblom G.D., Coon M.J. Immunochemical evidence for a role of cytochrome P-450 in liver microsomal ethanol oxidation. Arch. Biochem. Biophys. 1984;235:228–238. doi: 10.1016/0003-9861(84)90272-8. [DOI] [PubMed] [Google Scholar]
- 89.Gadeholt G. Ethanol and isoniazid induce a hepatic microsomal cytochrome P-450-dependent activity with similar properties towards substrate and inhibitors and different properties from those induced by classical inducers. Biochem. Pharmacol. 1984;33:3047–3051. doi: 10.1016/0006-2952(84)90607-5. [DOI] [PubMed] [Google Scholar]
- 90.Sturtevant R.P., Garber S.L. Circadian rhythms of alcohol dehydrogenase and MEOS in the rat. Proc. Soc. Exp. Biol. Med. 1984;175:299–303. doi: 10.3181/00379727-175-3-RC2. [DOI] [PubMed] [Google Scholar]
- 91.Ingelman-Sundberg M., Jörnvall H. Induction of the ethanol-inducible form of rabbit liver microsomal cytochrome P-450 by inhibitors of alcohol dehydrogenase. Biochem. Biophys. Res. Commun. 1984;124:375–382. doi: 10.1016/0006-291X(84)91563-8. [DOI] [PubMed] [Google Scholar]
- 92.Ingelman-Sundberg M., Johansson I. Mechanisms of hydroxyl radical formation and ethanol oxidation by ethanol-inducible and other forms of rabbit liver microsomal cytochrome P-450. J. Biol. Chem. 1984;259:6447–6458. [PubMed] [Google Scholar]
- 93.Leo M.A., Lowe N., Lieber C.S. Decreased hepatic vitamin A after drug administration in humans and rats. Am. J. Clin. Nutr. 1984;40:1131–1136. doi: 10.1093/ajcn/40.6.1131. [DOI] [PubMed] [Google Scholar]
- 94.Koop D.R., Crump B.L., Nordblom G.D., Coon M.J. Immunochemical evidence for induction of the alcohol-oxidizing cytochrome P-450 of rabbit liver microsomes by diverse agents: Ethanol, trichloroethylene, acetone, pyrazole, and isoniazid. Proc. Natl. Acad. Sci. USA. 1985;82:4065–4069. doi: 10.1073/pnas.82.12.4065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Yang C.S., Koop D.R., Wang T., Coon M.J. Immunochemical studies on the metabolism of nitrosamines by ethanol-inducible cytochrome P-450. Biochem. Biophys. Res. Commun. 1985;128:1007–1013. doi: 10.1016/0006-291X(85)90147-0. [DOI] [PubMed] [Google Scholar]
- 96.Johansson I., Ingelman-Sundberg M. Carbon tetrachloride induced lipid peroxidation dependent on an ethanol-inducible form of cytochrome P-450. FEBS Lett. 1985;183:265–269. doi: 10.1016/0014-5793(85)80790-0. [DOI] [PubMed] [Google Scholar]
- 97.Koop D.R., Casazza J.P. Identification of ethanol-inducible P-450 enzyme 3a as the acetone and acetol monooxygenase of rabbit microsomes. J. Biol. Chem. 1985;260:13607–13612. [PubMed] [Google Scholar]
- 98.Ryan D.E., Ramanthan L., Iida S., Thomas P.E., Haniu M., Shively J.E., Lieber C.S., Levin W. Characterization of a major form of rat hepatic microsomal cytochrome P-450 induced by isoniazid. J. Biol. Chem. 1985;260:6385–6393. [PubMed] [Google Scholar]
- 99.Gellert J., Lieber C.S. Effects of acute ethanol administration and chronic ethanol feeding on mixed function oxidation in Deermice lacking ADH. Alcohol. 1985;2:13–15. doi: 10.1016/0741-8329(85)90006-0. [DOI] [PubMed] [Google Scholar]
- 100.Gellert J., Alderman J., Lieber C.S. Interaction between ethanol metabolism and mixed-function oxidation in Alcohol Dehydrogenase positive and negative Deermice. Biochem. Pharmacol. 1986;35:1037–1041. doi: 10.1016/0006-2952(86)90095-X. [DOI] [PubMed] [Google Scholar]
- 101.Teschke R., Wannagat F.J., Löwendorf F., Strohmeyer G. Hepatic alcohol metabolizing enzymes after prolonged administration of sex hormones and alcohol in female rats. Biochem. Pharmacol. 1986;35:521–527. doi: 10.1016/0006-2952(86)90229-7. [DOI] [PubMed] [Google Scholar]
- 102.Teschke R. Hepatic microsomal ethanol-oxidizing system: Biochemical nature and clinical implications. In: Goedde H.W., Agarwal D.P., editors. Genetics and Alcoholism. Alan R. Liss Inc.; New York, NY, USA: 1986. pp. 173–184. [PubMed] [Google Scholar]
- 103.Leo M.A., Lowe N., Lieber C.S. Interaction of drugs and retinol. Biochem. Pharmacol. 1986;35:3949–3953. doi: 10.1016/0006-2952(86)90009-2. [DOI] [PubMed] [Google Scholar]
- 104.Ryan D.E., Koop D.R., Thomas P.E., Coon M.J., Levin W. Evidence that isoniazid and ethanol induce the same microsomal P-450 isozyme 3a. Arch. Biochem. Biophys. 1986;246:633–644. doi: 10.1016/0003-9861(86)90319-X. [DOI] [PubMed] [Google Scholar]
- 105.Wrighton S.A., Thomas P.E., Molowa D.T., Haniu M., Shively J.E., Maines S.L., Watkins P.B., Parker G., Mendez-Picon Levin W., Guzelman P.S. Characterization of ethanol-inducible human liver N-nitrosodimethylamine demethylase. Biochemistry. 1986;25:6731–6735. doi: 10.1021/bi00370a001. [DOI] [PubMed] [Google Scholar]
- 106.Ding X., Koop D.R., Crump B.I., Coon M.J. Immunochemical identification of cytochrome P-450 isozyme 3a [P-450ALC] in rabbit nasal and kidney microsomes and evidence for differential induction by alcohol. Mol. Pharmacol. 1986;30:370–378. [PubMed] [Google Scholar]
- 107.Johansson I.J., Eliasson E., Norsten C., Ingelman-Sundberg M. Hydroxylation of acetone by ethanol- and acetone-inducible cytochrome P-450 in liver microsomes and reconstituted membranes. FEBS Lett. 1986;196:59–64. doi: 10.1016/0014-5793(86)80214-9. [DOI] [PubMed] [Google Scholar]
- 108.Song B.J., Gelboin H.V., Park S.S., Yang C.S., Gonzalez F.J. Complementary DNA and protein sequences of ethanol-inducible rat and human cytochrome P-450s: Transcriptional and posttranscriptional regulation of the rat enzyme. J. Biol. Chem. 1986;261:16689–16697. [PubMed] [Google Scholar]
- 109.Bellward G.D., Chang T., Rodrigues J.H., McNeil J.H., Maines S., Ryan D.E., Levin W., Thomas B.E. Hepatic cytochrome P-450j induction in the spontaneously diabetic BB rat. Mol. Pharmacol. 1987;33:140–143. [PubMed] [Google Scholar]
- 110.Lasker J.M., Raucy J., Kubota S., Bloswick B.P., Black M., Lieber C.S. Purification and characterization of human liver cytochrome P-450-ALC. Biochem. Biophys. Res. Commun. 1987;148:232–238. doi: 10.1016/0006-291X(87)91100-4. [DOI] [PubMed] [Google Scholar]
- 111.Khani S.C., Zaphiropoulos P.G., Fujita V.S., Porter T.D., Koop D.R., Coon M.J. cDNA and derived amino acid sequence of ethanol inducible rabbit liver cytochrome P-450 isozyme 3a (P-450ALC) Proc. Natl. Acad. Sci. USA. 1987;84:638–642. doi: 10.1073/pnas.84.3.638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Lieber C.S., Lasker J.M., Alderman J., Leo M.A. The microsomal ethanol oxidizing system and its interaction with other drugs, carcinogens, and vitamins. Ann. N. Y. Acad. Sci. 1987;492:11–24. doi: 10.1111/j.1749-6632.1987.tb48649.x. [DOI] [PubMed] [Google Scholar]
- 113.Alderman J., Takagi T., Lieber C.S. Ethanol-metabolizing pathways in Deermice: Estimation of flux calculated from isotope effects. J. Biol. Chem. 1987;262:7497–7503. [PubMed] [Google Scholar]
- 114.Ingelman-Sundberg M., Johansson I., Penttila K.E., Glaumann H., Lindros K.O. Centrilobular expression of ethanol-inducible cytochrome P-450 (IIE1) in rat liver. Biochem. Biophys. Res. Commun. 1988;157:55–60. doi: 10.1016/S0006-291X(88)80010-X. [DOI] [PubMed] [Google Scholar]
- 115.Salazar D.E., Sorge C.L., Cocoran G.B. Obesity as a risk factor for drug-induced organ injury. VI. Increased hepatic P450 concentration and microsomal ethanol oxidizing activity in the obese overfed rat. Biochem. Biophys. Res. Commun. 1988;157:315–320. doi: 10.1016/S0006-291X(88)80049-4. [DOI] [PubMed] [Google Scholar]
- 116.Behrens U.H., Hoerner M., Lasker J.M., Lieber C.S. Formation of acetaldehyde adducts with ethanol-inducible P-450IIE1 in vivo. Biochem. Biophys. Res. Commun. 1988;154:584–590. doi: 10.1016/0006-291X(88)90180-5. [DOI] [PubMed] [Google Scholar]
- 117.Ding X., Coon M.J. Induction of cytochrome P-450 isozyme 3a (P-450IIE1) in rabbit olfactory mucosa by ethanol and acetone. Drug Metab. Dispos. 1988;18:742–745. [PubMed] [Google Scholar]
- 118.Kubota S., Lasker J.M., Lieber C.S. Molecular regulation of ethanol inducible cytochrome P-450IIE1 in hamsters. Biochem. Biophys. Res. Commun. 1988;150:304–310. doi: 10.1016/0006-291X(88)90520-7. [DOI] [PubMed] [Google Scholar]
- 119.Johansson I.J., Ingelman-Sundberg M. Benzene metabolism by ethanol-, acetone-, and benzene-inducuble cytochrome P-450 (IIE1) in rat and rabbit liver microsomes. Cancer Res. 1988;48:5387–5390. [PubMed] [Google Scholar]
- 120.Eliasson E., Johansson I., Ingelman-Sundberg M. Ligand-dependent maintenance of ethanol-inducible cytochrome P-450 in primary rat hepatocyte cell cultures. Biochem. Biophys. Res. Commun. 1988;150:436–443. doi: 10.1016/0006-291X(88)90539-6. [DOI] [PubMed] [Google Scholar]
- 121.Tsutsumi M., Shimizu M., Lasker J.M., Lieber C.S. Intralobular distribution of ethanol-inducible cytochrome P-450IIE1 in liver. Hepatology. 1988;8:1237A. doi: 10.1002/hep.1840100407. [DOI] [PubMed] [Google Scholar]
- 122.Dicker E., Cederbaum A.I. Increased oxygen radical dependent inactivation of metabolic enzymes by liver microsomes after chronic ethanol consumption. FASEB J. 1988;2:2901–2906. doi: 10.1096/fasebj.2.13.3169467. [DOI] [PubMed] [Google Scholar]
- 123.Porter T.D., Khani S.C., Coon M.J. Induction and tissue-specific expression of rabbit cytochrome P-450IIE1 and II2 genes. Mol. Pharmacol. 1989;36:61–65. [PubMed] [Google Scholar]
- 124.Tsutsumi M., Lasker J.M., Shimizu M., Rosman A.S., Lieber C.S. The intralobular distribution of ethanol-inducible P-450IIE1 in rat and human liver. Hepatology. 1989;10:437–446. doi: 10.1002/hep.1840100407. [DOI] [PubMed] [Google Scholar]
- 125.Hansson T., Tinberg N., Ingelman-Sundberg M., Köhler C. Regional distribution of ethanol-inducible cytochrome P-450 2E1 in the rat central nervous system. Neuroscience. 1990;34:451–463. doi: 10.1016/0306-4522(90)90154-V. [DOI] [PubMed] [Google Scholar]
- 126.Hong J.Y., Ning S.M., Ma B.L., Lee M.J., Pan J.M., Yang C.S. Roles of pituitary hormones in the regulation of hepatic cytochrome P-450IIE1in rats and mice. Arch. Biochem. Biophys. 1990;281:132–138. doi: 10.1016/0003-9861(90)90422-U. [DOI] [PubMed] [Google Scholar]
- 127.Koop D.R., Tierney D.J. Multiple mechanisms in the regulation of ethanol-inducible cytochrome P-450IIE1. Bioassays. 1990;9:429–435. doi: 10.1002/bies.950120906. [DOI] [PubMed] [Google Scholar]
- 128.Song B.J., Veech R.L., Saenger P. Cytochrome P-450IIE1 is elevated in lymphocytes from poorly controlled insulin-dependent diabetics. J. Clin. Endorinol. Metab. 1990;71:1036–1040. doi: 10.1210/jcem-71-4-1036. [DOI] [PubMed] [Google Scholar]
- 129.Kim S.G., Novak R.F. Induction of rat hepatic cytochrome P-450 IIE1 (CYP 2E1) by pyridine: Evidence for a role of protein synthesis in the absence of transcriptional activation. Biochem. Biophys. Res. Commun. 1990;166:1072–1079. doi: 10.1016/0006-291X(90)90976-T. [DOI] [PubMed] [Google Scholar]
- 130.Kim S.G., Shehin S.E., States J.C., Novak R.F. Evidence for increased translational efficiency in the induction of P-450IIE1 by solvents: Analysis of P-450IIE1 mRNA polyribosomal distribution. Biochem. Biophys. Res. Commun. 1990;172:767–774. doi: 10.1016/0006-291X(90)90740-E. [DOI] [PubMed] [Google Scholar]
- 131.Tsutsumi R., Leo A.M., Kim C., Tsutsumi M., Lasker J.M., Lowe N., Lieber C.S. Interaction of ethanol with enflurane metabolism and toxicity: Role of P-450IIE1. Alcohol. Clin. Exp. Res. 1990;14:174–179. doi: 10.1111/j.1530-0277.1990.tb00466.x. [DOI] [PubMed] [Google Scholar]
- 132.Yang C.S., Yoo J.S., Ishizaki H., Hong J. Cytochrome P-450IIE1: Roles of nitrosamine metabolism and mechanisms of regulation. Drug Metab. Rev. 1990;22:147–159. doi: 10.3109/03602539009041082. [DOI] [PubMed] [Google Scholar]
- 133.Peter R., Bocker R., Beaune P.H., Iwasaki M., Guengerich F.P., Yang C.S. Hydroxylation of chlorzoxazone as a specific probe for human liver cytochrome P-450IIE1. Chem. Res. Toxicol. 1990;3:566–573. doi: 10.1021/tx00018a012. [DOI] [PubMed] [Google Scholar]
- 134.Shimizu M., Lasker J.M., Tsutsumi M., Lieber C.S. Immunohistochemical localization of ethanol-inducible P-450IIE1 in the rat alimentary tract. Gastroenterology. 1990;99:1044–1053. doi: 10.1016/0016-5085(90)90625-B. [DOI] [PubMed] [Google Scholar]
- 135.Sohn D.H., Yun Y.P., Park S.S., Veech R.L., Song B.J. Post-translational reduction of cytochrome P450 2E1 by CCl4, its substrate. Biochem. Biophys. Res. Commun. 1991;179:449–454. doi: 10.1016/0006-291X(91)91391-O. [DOI] [PubMed] [Google Scholar]
- 136.Johansson E., Eliasson E., Ingelman-Sundberg M. Hormone controlled phosphorylation and degradation of CYP2B1 and 2E1 in isolated rat hepatocytes. Biochem. Biophys. Res. Commun. 1991;174:37–42. doi: 10.1016/0006-291X(91)90481-L. [DOI] [PubMed] [Google Scholar]
- 137.Yoo J., Ning S.M., Pantuck E.J., Yang C.S. Regulation of hepatic microsomal cytochrome P-450 2E1 level by dietary lipids and carbohydrates in rats. J. Nutr. 1991;121:959–965. doi: 10.1093/jn/121.7.959. [DOI] [PubMed] [Google Scholar]
- 138.Raucy J.L., Lasker J.M., Kramer J.C., Salazer D.E., Lieber C.S., Corcoran G.B. Induction of P-450IIE1 in the obese rat. Mol. Pharmacol. 1991;39:275–280. [PubMed] [Google Scholar]
- 139.Terelius Y., Norsten-Hoog C., Cronholm T., Ingelman-Sundberg M. Acetaldehyde as a substrate for ethanol-inducible cytochrome P-450 (CYP2E1) Biochem. Biophys. Res. Commun. 1991;179:689–694. doi: 10.1016/0006-291X(91)91427-E. [DOI] [PubMed] [Google Scholar]
- 140.Koop D.R., Chernosky A., Brass E.P. Identification and induction of cytochrome P-450 2E1 in rat Kupffer cells. J. Pharmacol. Exp. Ther. 1991;251:1072–1076. [PubMed] [Google Scholar]
- 141.Hayashi S., Watanabe J., Kaname K. Genetic polymorphism in the 5’-flanking region change transcriptional regulation of the human cytochrome P-450IIE1 gene. J. Biochem. 1991;110:559–565. doi: 10.1093/oxfordjournals.jbchem.a123619. [DOI] [PubMed] [Google Scholar]
- 142.Leo M.A., Kim C.I., Lowe N., Lieber C.S. Interaction of ethanol with β-carotene: Delayed blood clearance and enhanced hepatotoxicity. Hepatology. 1992;15:883–891. doi: 10.1002/hep.1840150522. [DOI] [PubMed] [Google Scholar]
- 143.Eliasson E., Mkrtchian S., Ingelman-Sundberg M. Hormone- and substrate-regulated intracellular degradation of cytochrome P450 (2E1) involving MgATP-activated rapid proteolysis in the endoplasmic reticulum membranes. J. Biol. Chem. 1992;267:15765–15769. [PubMed] [Google Scholar]
- 144.Sohda T., Shimizu M., Okumura M. Distribution in pancreas of ethanol-inducible P-4502E1 in rats fed ethanol plus high fat or low fat diet. Alcohol Metab. Liver. 1992;1:110–113. [Google Scholar]
- 145.Koop D.R. Oxidative and reductive metabolism by cytochrome P-450 2E1. FASEB J. 1992;6:724–730. doi: 10.1096/fasebj.6.2.1537462. [DOI] [PubMed] [Google Scholar]
- 146.Yamazaki H., Inui Y., Yun C.H., Guengrich F.P., Shimada T. Cytochrome P450 2E1 and 2A6 enzymes as major catalysts for metabolic activation of N-nitrosodialkylamines and tobacco-related nitrosamines in human liver microsomes. Carcinogenesis. 1992;13:1789–1794. doi: 10.1093/carcin/13.10.1789. [DOI] [PubMed] [Google Scholar]
- 147.Kharasch E.D., Thummel K.E., Mhyre J., Lillibridge J.H. Single-dose disulfiram inhibition of chlorzoxazone metabolism: A clinical probe for P450 2E1. Clin. Pharmacol. Ther. 1993;53:643–650. doi: 10.1038/clpt.1993.85. [DOI] [PubMed] [Google Scholar]
- 148.Thummel K.E., Kharasch E.D., Podoll T., Kunze K. Human liver microsomal enflurane defluorination catalyzed by cytochrome P-450 2E1. Drug Metab. Dispos. 1993;21:350–357. [PubMed] [Google Scholar]
- 149.Day Y., Rashba-Step J., Cederbaum A.I. Stable expression of human cytochrome P-4502E1 in HepG2 cells. Characterization of catalytic activities and production of reactive intermediates. Biochemistry. 1993;32:6928–6937. doi: 10.1021/bi00078a017. [DOI] [PubMed] [Google Scholar]
- 150.Hirvonen A., Husgafvel-Pursiainen K., Anttilla S., Karjalainen A., Vainio H. The human CYP2E1 gene and lung cancer: Dra I and Rsa I restriction fragment length polymorphisms in a Finish study population. Carcinogensis. 1993;1:85–88. doi: 10.1093/carcin/14.1.85. [DOI] [PubMed] [Google Scholar]
- 151.Morimoto M., Hagbjork A.L., Nanji A.A., Ingelman-Sundberg M., Lindros K.O., Fu P.C., Albano E., French S.W. Role of cytochrome P-450 2E1 in alcoholic liver disease pathogenesis. Alcohol. 1993;10:459–464. doi: 10.1016/0741-8329(93)90065-V. [DOI] [PubMed] [Google Scholar]
- 152.Rashba-Step J., Turro N.J., Cederbaum A.I. Increased NADPH- and NADH-dependent production of superoxide and hydroxyl radical by microsomes after chronic ethanol consumption. Arch. Biochem. Biophys. 1993;300:401–408. doi: 10.1006/abbi.1993.1054. [DOI] [PubMed] [Google Scholar]
- 153.Tsutsumi M., Lasker J.M., Takahashi T., Lieber C.S. The in vivo induction of hepatic P-4502E1: Role of increased enzyme synthesis. Arch. Biochem. Biophys. 1993;304:209–218. doi: 10.1006/abbi.1993.1341. [DOI] [PubMed] [Google Scholar]
- 154.Laethem R.M., Balaxy M., Falck J.R., Laethem C.L., Koop D.R. Formation of 19(S)-, 19(R)-, and 18(R)-hydroxyeicosatetraenoic acids by alcohol-inducible cytochrome P-4502E1. J. Biol. Chem. 1993;268:12912–12918. [PubMed] [Google Scholar]
- 155.Badger T.M., Huang J., Ronis M., Lumpkin C.K. Induction of cytochrome P-450 2E1 during chronic ethanol exposure occurs via transcription of the CYP 2E1 gene when blood alcohol concentrations are high. Biochem. Biophys. Res. Commun. 1993;190:780–785. doi: 10.1006/bbrc.1993.1117. [DOI] [PubMed] [Google Scholar]
- 156.Kunitoh S., Tanaka T., Imaoka S., Funae Y., Monna Y. Contribution of cytochrome P-450s to MEOS (microsomal ethanol-oxidizing system): A specific and sensitive assay of MEOS activity by HPLC with fluorescence labeling. Alcohol Alcohol. 1993;28:63–68. doi: 10.1093/alcalc/28.Supplement_1B.63. [DOI] [PubMed] [Google Scholar]
- 157.Lucas D., Berthou F., Dreano Y., Lozach P., Volant A., Menez J.F. Comparison of levels of cytochrome P-450, CYP1A2, CYP2E1, and their related monooxygenase activities in human surgical samples. Alcohol. Clin. Exp. Res. 1993;17:900–905. doi: 10.1111/j.1530-0277.1993.tb00861.x. [DOI] [PubMed] [Google Scholar]
- 158.Ronis M.J., Huang J., Crouch J., Mercado C., Irby D., Valentine C.R., Lumpkin C.K., Ingelman-Sundberg M., Badger T.M. Cytochrome P-450 CYP2E1 induction during chronic ethanol exposure occurs by a two-step mechanism associated with blood alcohol concentration in rats. J. Pharmacol. Exp. Ther. 1993;264:944–950. [PubMed] [Google Scholar]
- 159.Kharasch E.D., Thummel K.E. Identification of cytochrome P-450 2E1 as the predominant enzyme catalyzing human liver microsomal defluorination of sevoflurane, isoflurane, and methoxyflurane. Anesthesiology. 1993;79:795–807. doi: 10.1097/00000542-199310000-00023. [DOI] [PubMed] [Google Scholar]
- 160.Takahashi T., Lasker J.M., Rosman A.S., Lieber C.S. Inhibition of cytochrome P-4502E1 in the human liver by ethanol is caused by corresponding increase in encoding messenger RNA. Hepatology. 1993;17:236–245. [PubMed] [Google Scholar]
- 161.Tassaneeyakul W., Veronese M.E., Birkett D.J., Gonzalez F.J., Miners O. Validation of 4-nitrophenol as an in vitro substrate probe for human liver CYP2E1 using CDNA expression and microsomal kinetic techniques. Biochem. Pharmacol. 1993;46:1975–1981. doi: 10.1016/0006-2952(93)90639-E. [DOI] [PubMed] [Google Scholar]
- 162.Albano E., Tomasi A., Ingelman-Sundberg M. Spin trapping of alcohol derived radicals in microsomes and reconstituted systems by electron spin resonance. Methods Enzymol. 1994;223:117–127. doi: 10.1016/s0076-6879(94)33014-x. [DOI] [PubMed] [Google Scholar]
- 163.Carrocio A., Wu D., Cederbaum A.I. Ethanol increases content and activity of human cytochrome P-4502E1 in a transduced HEPG2 cell line. Biochem. Biophys. Res. Commun. 1994;203:727–733. doi: 10.1006/bbrc.1994.2242. [DOI] [PubMed] [Google Scholar]
- 164.Botto F., Seree E., Khyari S.E., DeSoussa G., Massacrier A., Placidi M., Cau P., Pellet W., Rahmani R., Barra Y. Tissue-specific expression and methylation of the human CYP2E1 gene. Biochem. Pharmacol. 1994;48:1095–1103. doi: 10.1016/0006-2952(94)90145-7. [DOI] [PubMed] [Google Scholar]
- 165.Tsutsumi M., Takata A., Wang J.S. Genetic polymorphism of cytochrome P-4502E1 related to the development of alcoholic liver disease. Gastroenterology. 1994;107:1430–1435. doi: 10.1016/0016-5085(94)90546-0. [DOI] [PubMed] [Google Scholar]
- 166.Stephens E.A., Taylor J.A., Kaplan N., Yang C.H., Hsieh L.L., Lucier G.W., Bell D.A. Ethnic variation in the CYP2E1 gene: Polymorphism analysis of 695 African-Americans, European-Americans and Taiwanese. Pharmacogenetics. 1994;4:185–192. doi: 10.1097/00008571-199408000-00002. [DOI] [PubMed] [Google Scholar]
- 167.Chen L., Lee M., Hong J.Y., Huang W., Wang E., Yang C.S. Relationship between cytochrome P-450 2E1 and acetone catabolism in rats as studied with diallyl sulfide as an inhibitor. Biochem. Pharmacol. 1994;48:2199–2205. doi: 10.1016/0006-2952(94)00435-8. [DOI] [PubMed] [Google Scholar]
- 168.Maezawa Y., Yamauchi M., Toda G. Association between restriction fragment length polymorphism of the human cytochrome P-450IIE1 gene and susceptibility to alcoholic liver cirrhosis. Am. J. Gastroenterol. 1994;89:561–565. [PubMed] [Google Scholar]
- 169.Amet Y., Berthou F., Goasduff T., Schaun J.P., Le Breton L., Menez J.F. Evidence that cytochrome P-450 2E1 is involved in the (omega-1)-hydroxylation of lauric acid in rat liver microsomes. Biochem. Biophys. Res. Commun. 1994;203:1168–1174. doi: 10.1006/bbrc.1994.2305. [DOI] [PubMed] [Google Scholar]
- 170.McGehee R.E., Ronis M.J.J., Cowherd R.M., Ingelman-Sunderberg M., Badger T.M. Characterization of cytochrome P-450 2E1 induction in a rat hepatoma FGC-4 cell model by ethanol. Biochem. Pharmacol. 1994;48:1823–1833. doi: 10.1016/0006-2952(94)90469-3. [DOI] [PubMed] [Google Scholar]
- 171.Kang M.H., Won S.M., Park S.S., Kim S.G., Novak R.F., Kim N.D. Piperine effects on the expression of P4502E1, P4502B and P4501A in rat. Xenobiotica. 1994;24:1195–1204. doi: 10.3109/00498259409038675. [DOI] [PubMed] [Google Scholar]
- 172.Uematsu F., Ikawa S., Kikuchi H., Sagami I., Kanamara R., Abe T., Satoh K., Motomiya M., Watanabe M. Restriction fragment length polymorphism of the human CYP2E1 (cytochrome P-450IIE1) gene and susceptibility to lung cancer: Possible relevance to low smoking exposure. Pharmacogenetics. 1994;4:58–63. doi: 10.1097/00008571-199404000-00002. [DOI] [PubMed] [Google Scholar]
- 173.Watanabe J., Hayashi S., Kawajiri K. Different regulations and expression of the human CYP2E1 gene due to the Rsal polymorphism in the 5′ flanking region. J. Biochem. 1994;116:321–326. doi: 10.1093/oxfordjournals.jbchem.a124526. [DOI] [PubMed] [Google Scholar]
- 174.Watanabe J., Yang J.P., Eguchi H., Hayashi S., Imai K., Nakachi K., Kawajiri K. An Rsa I polymorphism in the CYP2E1 gene does not affect lung cancer risk in a Japanese population. Jpn. Cancer Res. 1995;86:245–248. doi: 10.1111/j.1349-7006.1995.tb03046.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Roberts B.J., Song B.J., Soh Y., Park S.S., Shoaf S.E. Ethanol induces CYP2E1 by protein stabilization. J. Biol. Chem. 1995;270:29632–29635. doi: 10.1074/jbc.270.50.29632. [DOI] [PubMed] [Google Scholar]
- 176.Speerschneider P., Dekant W. Renal tumorigenicity of 1,1-dichloroethene in mice: The role of male-specific expression of cytochrome P-450 2E1 in the renal bioactivation of 1,1-dichloroethene. Toxicol. Appl. Pharmacol. 1995;130:48–56. doi: 10.1006/taap.1995.1007. [DOI] [PubMed] [Google Scholar]
- 177.Subramanian U., Ahmed A.E. Intestinal toxicity of acrylonitrile: In vitro metabolism by intestinal P-450 2E1. Toxicol. Appl. Pharmacol. 1995;135:1–8. doi: 10.1006/taap.1995.1202. [DOI] [PubMed] [Google Scholar]
- 178.Schmalix W.A., Barrenscheen N., Landsiedel R., Janzowski C., Eisenbrand G., Gonzalez F., Eliasson B., Ingelman-Sundberg M., Perchermeier M., Greim H. Stable expression of human cytochrome P-450 2E1 in V79 Chinese hamster cells. Eur. J. Pharmacol. 1995;293:123–131. doi: 10.1016/0926-6917(95)00008-9. [DOI] [PubMed] [Google Scholar]
- 179.Carr L.G., Hartleroad J.Y., Liang Y.B., Mendenhall C., Moritz T., Thomasson H. Polymorphism at the P-450IIE1 locus is not associated with alcoholic liver disease in Caucasian men. Alcohol. Clin. Exp. Res. 1995;19:182–184. doi: 10.1111/j.1530-0277.1995.tb01489.x. [DOI] [PubMed] [Google Scholar]
- 180.Morimoto M., Hagbjork A.L., Wan Y.J.Y., Fu P.C., Clot P., Albano E., Ingelman-Sundberg M., French S.W. Modulation of experimental alcohol-induced liver disease by cytochrome P-450 2E1 inhibitors. Hepatology. 1995;21:1610–1617. [PubMed] [Google Scholar]
- 181.Rannug A., Alexandrie A.K., Persson I., Ingelman-Sundberg M. Genetic polymorphism of cytochrome P-450 1A1, 2D6 and 2E1: Regulation and toxicological significance. J. Occup. Environ. Med. 1995;37:25–36. doi: 10.1097/00043764-199501000-00005. [DOI] [PubMed] [Google Scholar]
- 182.Pirmohamed M., Kitteringham N.R., Quest L.I., Allott R.L., Green V.J., Gilmore I.T., Park B.K. Genetic polymorphism of cytochrome P-4502E1 and risk of alcoholic liver disease in Caucasians. Pharmacogenetics. 1995;5:351–357. doi: 10.1097/00008571-199512000-00003. [DOI] [PubMed] [Google Scholar]
- 183.Lucas D., Menez C., Girre C., Berthou F., Bodenez P., Joannet I., Hispard E., Bardon L.G., Menez J.F. Cytochrome P-450 2E1 genotype and chlorzoxazone metabolism in healthy and alcoholic Caucasian subjects. Pharmacogenetics. 1995;5:298–304. doi: 10.1097/00008571-199510000-00005. [DOI] [PubMed] [Google Scholar]
- 184.Lucas D., Menez C., Girre C., Bodenez P., Hispard E., Menez J.F. Decrease in cytochrome P-4502E1 as assessed by the rate of chlorzoxazone hydroxylation in alcoholics during the withdrawal phase. Alcohol. Clin. Exp. Res. 1995;19:362–366. doi: 10.1111/j.1530-0277.1995.tb01516.x. [DOI] [PubMed] [Google Scholar]
- 185.Mishin V., Koivisto T., Lieber C.S. Respective roles of CYP2E1 and CYPA2 in chlorzoxazone and ethanol metabolism in mammalian liver microsomes. Alcohol. Clin. Exp. Res. 1995;19:78A. [Google Scholar]
- 186.Jayyosi Z., Knoble D., Muc M., Erick J., Thomas P.E., Kelley N. Cytochrome P-450 2E1 is not the sole catalyst of chlorzoxazone hydroxylation in rat liver microsomes. J. Pharmacol. Exp. Ther. 1995;273:1156–1161. [PubMed] [Google Scholar]
- 187.Yamazaki H., Guo Z., Guengerich F.P. Selectivity of cytochrome P4502E1 in chlorzoxazone 6-hydroxylation. Drug Metab. Dispos. 1995;23:438–440. [PubMed] [Google Scholar]
- 188.De Waziers I., Garlatti M., Bouguet J., Beaune P.H., Barouki R. Insulin down-regulates cytochrome P-450 2B and 2E expression at the posttranscriptional level in the rat hepatoma cell line. Mol. Pharmacol. 1995;47:474–479. [PubMed] [Google Scholar]
- 189.Sampol E., Mirrione A., Villard P.H., Picerelle P., Scoma H., Berbis P., Barra Y., Durand A., Lacarelle B. Evidence for a tissue-specific induction of cutaneous CYP2E1 by dexamethasone. Biochem. Biophys. Res. Commun. 1995;235:557–561. doi: 10.1006/bbrc.1997.6829. [DOI] [PubMed] [Google Scholar]
- 190.Crabb D.W. Ethanol oxidizing enzyme: Roles in alcohol metabolism and alcoholic liver disease. Prog. Liver Dis. 1995;13:151–172. [PubMed] [Google Scholar]
- 191.Zerilli A., Lucas D., Amet Y., Beauge F., Volant A., Floch H.H., Berthou F., Menez J.F. Cytochrome P-450 2E1 in rat liver, kidney and lung microsomes after chronic administration of ethanol either orally or by inhalation. Alcohol Alcohol. 1995;30:357–365. [PubMed] [Google Scholar]
- 192.Asai H., Imaoka S., Kuroki T., Monna T., Funnae Y. Microsomal ethanol oxidizing system activity by 2 DNA single strand breaks. Biochem. Biophys. Res. Commun. 1996;219:429–434. [Google Scholar]
- 193.Seree E.M., Villard P.H., Re J.L., De Meo M., Lacarelle B., Attolini L., Dumenil G., Catalin J., Durand A., Barra Y. High inducibility of mouse renal CYP2E1 gene by tobacco smoke and its possible effect on DNA single strand breaks. Biochem. Biophys. Res. Commun. 1996;219:429–434. doi: 10.1006/bbrc.1996.0250. [DOI] [PubMed] [Google Scholar]
- 194.Hakkak R., Korourian S., Ronis M.J., Ingelman-Sundberg M., Badger T.M. Effects of diet and ethanol on the expression and localization of cytochromes P-450 2E1 and P-450 2C7 in the colon of male rats. Biochem. Pharmacol. 1996;51:61–69. doi: 10.1016/0006-2952(95)02154-X. [DOI] [PubMed] [Google Scholar]
- 195.Koivisto T., Mishin V.M., Mak K.M., Cohen P.A., Lieber C.S. Induction of cytochrome P-4502E1 by ethanol in rat Kuppfer cells. Alcohol. Clin. Res. Exp. Res. 1996;20:207–212. doi: 10.1111/j.1530-0277.1996.tb01631.x. [DOI] [PubMed] [Google Scholar]
- 196.Carriere V., Berthou F., Baird S., Belloc C., Beaune P., De Waziers I. Human cytochrome P-450 2E1 (CYP2E1): From genotype to phenotype. Pharmacogenetics. 1996;6:203–211. doi: 10.1097/00008571-199606000-00002. [DOI] [PubMed] [Google Scholar]
- 197.Tindberg N., Ingelman-Sundberg M. Expression, catalytic activity, and inducibility of cytochrome P-450 2E1 (CYP2E1) in the rat central nervous system. J. Neurochem. 1996;67:2066–2073. doi: 10.1046/j.1471-4159.1996.67052066.x. [DOI] [PubMed] [Google Scholar]
- 198.Court M.H., Von Moltke L.L., Shader R.I., Greenblatt D.J. Biotransformation of chlorzoxazone by hepatic microsomes from humans and ten other mammalian species. Biopharm. Drug Dispos. 1997;18:213–226. doi: 10.1002/(SICI)1099-081X(199704)18:3<213::AID-BDD15>3.0.CO;2-0. [DOI] [PubMed] [Google Scholar]
- 199.McGehee R.E., Ronis M.J.J., Badger T.M. Regulation of the hepatic CYP2E1 gene during chronic alcohol exposure: Lack of an ethanol response element in the proximal 5′-flanking sequence. DNA Cell Biol. 1997;16:725–736. doi: 10.1089/dna.1997.16.725. [DOI] [PubMed] [Google Scholar]
- 200.Cohen P.A., Mak K.M., Rosman A.S., Kessova I., Mishin V.M., Kovisto T., Lieber C.S. Immunohistochemical determination of hepatic cytochrome P-4502E1 in formalin-fixed, paraffin-embedded sections. Alcohol. Clin. Exp. Res. 1997;21:1057–1062. doi: 10.1111/j.1530-0277.1997.tb04254.x. [DOI] [PubMed] [Google Scholar]
- 201.Zangar R.C., Novak R.F. Effect of fatty acids and ketone bodies on cytochromes P-450 2B, 4A, and 2E1 expression in primary cultured rat hepatocytes. Arch. Biochem. Biophys. 1997;337:217–224. doi: 10.1006/abbi.1996.9785. [DOI] [PubMed] [Google Scholar]
- 202.Zimatkin S.M., Deitrich R.A. Ethanol metabolism in the brain. Addict. Biol. 1997;2:387–400. doi: 10.1080/13556219772444. [DOI] [PubMed] [Google Scholar]
- 203.Kim N.D., Kwat M.K., Kim S.G. Inhibition of cytochrome P-450 2E1 expression by 2-(allylthio) pyrazine, a potential chemoprotective agent: Hepatoprotective effects. Biochem. Pharmacol. 1997;53:261–269. doi: 10.1016/S0006-2952(96)00647-8. [DOI] [PubMed] [Google Scholar]
- 204.Woodcroft K.J., Novak R.F. Insulin effects on CYP2E1, 2B, 3A, and 4A expression in primary cultured rat hepatocytes. Chem. Biol. Interact. 1997;107:75–91. doi: 10.1016/S0009-2797(97)00075-6. [DOI] [PubMed] [Google Scholar]
- 205.French S., Morimoto M., Reitz R., Koop D., Klopfenstein B., Estes K., Clot B., Ingelman-Sundberg M., Albano E. Lipidperoxidation, CYP2E1 and arachidonoid acid metabolism in alcoholic liver disease in rats. J. Nutr. 1997;127(Suppl. 5):907S–911S. doi: 10.1093/jn/127.5.907S. [DOI] [PubMed] [Google Scholar]
- 206.Gebhardt A.C., Lucas D., Menez J.F., Seitz H.K. Chlormethiazole inhibition of cytochrome P450 2E1 as assessed by chlorzoxazone hydroxylation in humans. Hepatology. 1997;26:957–961. doi: 10.1002/hep.510260423. [DOI] [PubMed] [Google Scholar]
- 207.Lucas D., Farez C., Bardou L.G., Vaisse J., Attali J.R., Valensi P. Cytochrome P-450 2E1 activity in diabetic and obese patients as assessed by chlorzoxazone hydroxylation. Fundam. Clin. Pharmacol. 1998;12:553–558. doi: 10.1111/j.1472-8206.1998.tb00985.x. [DOI] [PubMed] [Google Scholar]
- 208.Powell H., Kitteringham N.R., Pirmohamed M., Smith D.A., Park B.K. Expression of cytochrome P-4502E1 in human liver: Assessment by MRNA, genotype and phenotype. Pharmacogenetics. 1998;8:411–421. doi: 10.1097/00008571-199810000-00006. [DOI] [PubMed] [Google Scholar]
- 209.Weltman M.D., Farrell G.C., Hall P., Ingelman-Sundberg M., Liddle C. Hepatic cytochrome P4502E1 is increased in patients with nonalcoholic steatohepatitis. Hepatology. 1998;27:128–133. doi: 10.1002/hep.510270121. [DOI] [PubMed] [Google Scholar]
- 210.Salmela K.S., Kessova I.G., Tsyrlov I.B., Lieber C.S. Respective roles of human cytochrome P-4502E1 and 3A4 in the hepatic microsomal ethanol oxidizing system. Alcohol. Clin. Exp. Res. 1998;22:2125–2132. doi: 10.1111/j.1530-0277.1998.tb05926.x. [DOI] [PubMed] [Google Scholar]
- 211.Wu Y., Salmela K., Lieber C.S. Microsomal acetaldehyde oxidation is negible in the presence of ethanol. Alcohol. Clin. Exp. Res. 1998;22:1165–1169. doi: 10.1111/j.1530-0277.1998.tb03717.x. [DOI] [PubMed] [Google Scholar]
- 212.Matthews J.M., Etheridge A.S., Raymer J.H., Black S.R., Pulliam D.W., Bucher J.M. Selective inhibition of cytochrome P-450 2E1 in vivo and in vitro with trans-1, 2-dichloroethylene. Chem. Res. Toxicol. 1998;11:778–785. doi: 10.1021/tx970227g. [DOI] [PubMed] [Google Scholar]
- 213.Jiang Y., Kuo C.L., Pernecky S.J., Piper W.N. The detection of cytochrome P-450 2E1 and its catalytic activity in rat testis. Biochem. Biophys. Res. Commun. 1998;246:578–583. doi: 10.1006/bbrc.1998.8665. [DOI] [PubMed] [Google Scholar]
- 214.Kessova I.G., DeCarli L.M., Lieber C.S. Inducibility of cytochrome P-450 2E1 & P-450 1A1 in rat pancreas. Alcohol. Clin. Exp. Res. 1998;22:501–504. [PubMed] [Google Scholar]
- 215.Norton I.D., Apte M.V., Haber P.S., McCaughan G.W., Pirola R.C., Wilson J.S. Cytochrome P-450 2E1 is present in rat pancreas is induced by chronic ethanol administration. Gut. 1998;42:426–430. doi: 10.1136/gut.42.3.426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Aleynik M.K., Leo M.A., Aleynik S.I., Lieber C.S. Polyenylphosphatidylcholine opposes the increase of cytochrome P-4502E1 by ethanol and corrects the iron-induced decrease. Alcohol. Clin. Exp. Res. 1998;23:96–100. doi: 10.1111/j.1530-0277.1999.tb04028.x. [DOI] [PubMed] [Google Scholar]
- 217.Adas F., Betthou F., Picart D., Lozach P., Beauge F., Amet Y. Involvement of cytochrome P-450 2E1 in the (omega-1)-hydroxylation of oleic acid in human and rat liver microsomes. J. Lipid Res. 1998;39:1210–1219. [PubMed] [Google Scholar]
- 218.Mishin V.M., Rosma A.S., Basu P., Kessova I., Oneta C.M., Lieber C.S. Chlorzoxazone pharmacogenetics as a marker of hepatic cytochrome P-4502E1 in humans. Am. J. Gastroenterol. 1998;93:2154–2161. doi: 10.1111/j.1572-0241.1998.00612.x. [DOI] [PubMed] [Google Scholar]
- 219.Eap C.B., Schnyder C., Besson J., Savary L., Buclin T. Inhibition of CYP2E1 by chlormethiazole as measured by chlorzoxazone pharmacokinetics in patients with alcoholism and in healthy volunteers. Clin. Pharmacol. Ther. 1998;64:52–57. doi: 10.1016/S0009-9236(98)90022-4. [DOI] [PubMed] [Google Scholar]
- 220.Peng H.M., Coon M.J. Regulation of rabbit cytochrome P-450 2E1 expression in HepG2 cells by insulin and thyroid hormones. Mol. Pharmacol. 1998;54:740–747. [PubMed] [Google Scholar]
- 221.Dupont I., Lucas D., Clot P., Menez C., Albano E. Cytochrome P-4502E1 inducibility and hydroxyethyl radical formation among alcoholics. J. Hepatol. 1998;28:564–571. doi: 10.1016/S0168-8278(98)80279-1. [DOI] [PubMed] [Google Scholar]
- 222.Leo M.A., Lieber C.S. Alcohol, vitamin, A.; and beta-carotene: Adverse interactions, including hepatotoxicity and carcinogenicity. Am. J. Clin. Nutr. 1999;69:1071–1085. doi: 10.1093/ajcn/69.6.1071. [DOI] [PubMed] [Google Scholar]
- 223.Hutson J.L., Wickramasinghe S.N. Expression of CYP2E1 by human monocyte-derived macrophages. J. Pathol. 1999;188:197–200. doi: 10.1002/(SICI)1096-9896(199906)188:2<197::AID-PATH295>3.0.CO;2-M. [DOI] [PubMed] [Google Scholar]
- 224.Benowitz N.L., Jacob P., Saunders S., Gourlay S. Carbon monoxide, cigarette smoking and CYP2E1 activity. Clin. Pharmacol. 1999;65:154. doi: 10.1016/S0009-9236(99)80148-9. [DOI] [Google Scholar]
- 225.Lucas D., Ferrara R., Gonzalez E., Bodenez P., Albores A., Manno M., Berthou F. Chlorzoxazone, a selective probe for phenotyping CYP2E1 in humans. Pharmacogenetics. 1999;9:377–388. doi: 10.1097/00008571-199906000-00013. [DOI] [PubMed] [Google Scholar]
- 226.Loizou G.D., Cocker J. The effects of alcohol and diallylsulphide on CYP2E1 activity in humans: A phenotyping study using chlorzoxazone. Hum. Exp. Toxicol. 2001;20:321–327. doi: 10.1191/096032701680350587. [DOI] [PubMed] [Google Scholar]
- 227.McCarty M.F. Inhibition of CYP2E1 with natural agents may be a feasible strategy for minimizing the hepatotoxicity of ethanol. Med. Hypotheses. 2001;56:8–11. doi: 10.1054/mehy.1999.1015. [DOI] [PubMed] [Google Scholar]
- 228.Sun A.Y., Magnus Ingelman-Sundberg M., Neve E., Matsumoto H., Nishitani Y., Minowa Y., Fukui Y., Bailey S.M., Patel V.B., Cunningham C.C., et al. Ethanol and oxidative stress. Alcohol. Clin. Exp. Res. 2001;25:237S–243S. doi: 10.1111/j.1530-0277.2001.tb02402.x. [DOI] [PubMed] [Google Scholar]
- 229.Frye R.F., Branch R.A. Effect of chronic disulfiram administration on the activities of CYP1A2, CYP2C19, CYP2D6, CYP2E1, and N-acetyltransferase in healthy human subjects. Br. J. Clin. Pharmacol. 2002;53:155–162. doi: 10.1046/j.1365-2125.2002.01522.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Ghosal A., Hapangama N., Yuan Y., Lu X., Horne D., Patrick J.E., Zbaida S. Rapid determination of enzyme activities of recombinant human cytochromes P450, human liver microsomes and hepatocytes. Pharm. Drug Dispos. 2003;24:375–384. doi: 10.1002/bdd.374. [DOI] [PubMed] [Google Scholar]
- 231.Kessova I., Cederbaum A.I. CYP2E1: Biochemistry, toxicology, regulation and function in ethanol-induced liver injury. Curr. Mol. Med. 2003;3:509–518. doi: 10.2174/1566524033479609. [DOI] [PubMed] [Google Scholar]
- 232.Ernstgard L., Warholm M., Johanson G. Robustness of chlorzoxazone as an in vivo measure of cytochrome P450 2E1 activity. Br. J. Clin. Pharmacol. 2004;58:190–200. doi: 10.1111/j.1365-2125.2004.02132.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Park J.Y., Kim K.A., Park P.W., Ha J.M. Effect of high-dose aspirin on CYP2E1 activity in healthy subjects measured using chlorzoxazone as a probe. J. Clin. Pharmacol. 2006;46:109–114. doi: 10.1177/0091270005282635. [DOI] [PubMed] [Google Scholar]
- 234.Lu Y., Cederbaum A.I. CYP2E1 and oxidative liver injury by alcohol. Free Radic. Biol. Med. 2008;44:723–738. doi: 10.1016/j.freeradbiomed.2007.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Wellman M., Siest G. Cytochrome P-450-mediated differential oxidative modification of proteins: Albumin, apolipoprotein, E.; and CYP2E1 as targets. J. Toxicol. Environ. Health. 2010;67:2061–2071. doi: 10.1080/15287390490514895. [DOI] [PubMed] [Google Scholar]
- 236.Kim S.K., Park H.J., Seok H., Jeon H.S., Lee T.W., Lee S.H., Moon S.Y., Ihm C.G., Kim T.H., Kim Y.H., et al. Association studies of cytochrome P450, family 2, subfamily, E.; polypeptide 1 (CYP2E1) gene polymorphisms with acute rejection in kidney transplantation recipients. Clin. Transplant. 2014;28:707–712. doi: 10.1111/ctr.12369. [DOI] [PubMed] [Google Scholar]
- 237.Ceni E., Mello T., Galli A. Pathogenesis of alcoholic liver disease: Role of oxidative metabolism. World J. Gastroenterol. 2014;20:17756–17772. doi: 10.3748/wjg.v20.i47.17756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Bedada S.K., Neerati P. Resveratrol pretreatment affects CYP2E1 activity of chlorzoxazone in healthy human. Phytother. Res. 2016;30:463–468. doi: 10.1002/ptr.5549. [DOI] [PubMed] [Google Scholar]
- 239.Bedada S.K., Boga P.K. Effect of piperine on CYP2E1 enzyme activity of chlorzoxazone in healthy volunteers. Xenobiotica. 2017;47:1035–1041. doi: 10.1080/00498254.2016.1241450. [DOI] [PubMed] [Google Scholar]
- 240.Xu J., Ma H.Y., Liang S., Sun M., Karin G., Koyama Y., Hu R., Quehenberger O., Davidson N.O., Dennis E.A., et al. The role of human cytochrome P450 2E1 in liver inflammation and fibrosis. Hepatol. Commun. 2017;1:1043–1057. doi: 10.1002/hep4.1115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Abdelmegeed M.A., Choi Y., Ha S.K., Song B.J. Cytochrome P450-2E1 is involved in aging-related kidney damage in mice through increased nitroxidative stress. Food Chem. Toxicol. 2017;109:48–59. doi: 10.1016/j.fct.2017.08.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Fujiwara R. Exposure to sub-parts per million levels of vinyl chloride can increase the risk of developing liver injury. Hepatol. Commun. 2018;2:227–229. doi: 10.1002/hep4.1169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Lang A.L., Chen L., Poff G.D., Ding W.X., Barnett R.A., Arteel G.E., Beier J.I. Vinylchloride dysregulates metabolic homeostasis and enhances diet-induced liver injury in mice. Hepatol. Commun. 2018;2:270–284. doi: 10.1002/hep4.1151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Matsuzaki S., Teschke R., Ohnishi K., Lieber C.S. Acceleration of ethanol metabolism by high ethanol concentrations and chronic alcohol consumption: Role of the microsomal ethanol oxidizing system (MEOS) In: Fisher M.M., Rankin J.G., editors. Alcohol and the Liver. Volume III. Plenum Press; New York, NY, USA: 1977. pp. 119–143. [Google Scholar]
- 245.Lieber C.S. Mechanism of ethanol induced hepatic injury. Pharmacol. Ther. 1990;46:1–41. doi: 10.1016/0163-7258(90)90032-W. [DOI] [PubMed] [Google Scholar]
- 246.Lieber C.S. Cytochrome P-4502E1: Its physiological and pathological role. Physiol. Rev. 1997;77:517–544. doi: 10.1152/physrev.1997.77.2.517. [DOI] [PubMed] [Google Scholar]
- 247.Lieber C.S. Microsomal ethanol-oxidizing system (MEOS): The first 30 years (1968-1998)—A review. Alcohol. Clin. Exp. Res. 1999;2:991–1007. doi: 10.1097/00000374-199906000-00006. [DOI] [PubMed] [Google Scholar]
- 248.Tanaka E., Terada M., Misawa S. Cytochrome P450 2E1: Its clinical and toxicological role. Clin. Pharm. Ther. 2000;25:165–175. doi: 10.1046/j.1365-2710.2000.00282.x. [DOI] [PubMed] [Google Scholar]
- 249.Lieber C.S. The discovery of the microsomal ethanol oxidizing system and its physiologic and pathologic role. Drug Metab. Rev. 2004;36:511–529. doi: 10.1081/DMR-200033441. [DOI] [PubMed] [Google Scholar]
- 250.Lieber C.S. Milestones in liver disease, a commentary. The unexpected outcomes of medical research: Serendipidy and the microsomal ethanol oxidizing system. J. Hepatol. 2004;40:198–202. doi: 10.1016/j.jhep.2003.12.005. [DOI] [PubMed] [Google Scholar]
- 251.Butura A., Nilsson K., Morgan K., Morgan T.R., French S.W., Johansson I., Schuppe-Koistinen I., Ingelman-Sundberg M. The impact of CYP2E1 on the development of alcoholic liver disease as studied in a transgenic mouse model. J. Hepatol. 2009;50:572–583. doi: 10.1016/j.jhep.2008.10.020. [DOI] [PubMed] [Google Scholar]
- 252.Cederbaum A.I., Lu Y., Wu D. Role of oxidative stress in alcohol-induced liver injury. Arch. Toxicol. 2009;83:519–548. doi: 10.1007/s00204-009-0432-0. [DOI] [PubMed] [Google Scholar]
- 253.Iseri O.A., Lieber C.S., Gottlieb I.S. The ultrastructure of fatty liver induced by prolonged ethanol ingestion. Am. J. Pathol. 1966;48:535–555. [PMC free article] [PubMed] [Google Scholar]
- 254.Lane B.P., Lieber C.S. Ultrastructural alterations in human hepatocytes following ingestion of ethanol with adequate diets. Am. J. Pathol. 1966;49:593–603. [PMC free article] [PubMed] [Google Scholar]
- 255.Joly J.G., Ishii H., Lieber C.S. Microsomal cyanide-binding cytochrome: Its role in hepatic ethanol oxidation. Gastroenterology. 1972;62:174A. [Google Scholar]
- 256.Villeneuve J.P., Mavier P., Joly J.G. Ethanol-induced cytochro 257me P-450: Catalytic activity after prtial purification. Biochem. Biophys. Res. Commun. 1976;70:723–728. doi: 10.1016/0006-291X(76)90652-5. [DOI] [PubMed] [Google Scholar]
- 257.Teschke R. Liver injury by carbon tetrachloride intoxication in 16 patients treated with forced ventilation to accelerate toxin removal via the lungs: A clinical report. Toxics. 2018;6:25. doi: 10.3390/toxics6020025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Salaspuro M.P., Lieber C.S. Nonuniformaty of blood ethanol elimination: Its exaggeration after chronic consumption. Ann. Clin. Res. 1978;10:294–297. [PubMed] [Google Scholar]
- 259.Feinman L., Baraona E., Matsuzaki S., Korsten M., Lieber C.S. Concentration dependence of ethanol metabolism in vivo in rats and man. Alcohol. Clin. Exp. Res. 1978;2:381–385. doi: 10.1111/j.1530-0277.1978.tb04748.x. [DOI] [PubMed] [Google Scholar]
- 260.Cederbaum A.I., Dicker E., Lieber C.S., Rubin E. Ethanol oxidation by isolated hepatocytes from ethanol-treated and control rats: Factors contributing to the metabolic adaptation after chronic ethanol consumption. Biochem. Pharmacol. 1978;27:7–15. doi: 10.1016/0006-2952(78)90250-2. [DOI] [PubMed] [Google Scholar]
- 261.Pikkarainen P.H., Lieber C.S. Concentration dependence of ethanol elimination rates on baboons: Effect of chronic alcohol consumption. Alcohol. Clin. Exp. Res. 1980;4:40–43. doi: 10.1111/j.1530-0277.1980.tb04789.x. [DOI] [PubMed] [Google Scholar]
- 262.Matzsuzaki S., Gordon E., Lieber C.S. Increased alcohol dehydrogenase independent ethanol oxidation at high ethanol concentrations in isolated rat hepatocytes: The effect of chronic ethanol feeding. J. Pharmacol. Exp. Ther. 1981;217:133–137. [PubMed] [Google Scholar]
- 263.Shigeta Y., Nomura F., Iida S., Leo M.A., Felder M.R., Lieber C.S. Ethanol metabolism in vivo by the microsomal ethanol-oxidizing system in Deermice lacking alcohol dehydrogenase (ADH) Biochem. Pharmacol. 1984;33:807–814. doi: 10.1016/0006-2952(84)90466-0. [DOI] [PubMed] [Google Scholar]
- 264.Korsten M.A., Matsuzaki S., Feinman L., Lieber C.S. High blood acetaldehyde levels after ethanol administration—Difference between alcoholic and nonalcoholic subjects. N. Engl. J. Med. 1975;292:386–389. doi: 10.1056/NEJM197502202920802. [DOI] [PubMed] [Google Scholar]
- 265.Davis V.E., Walsh M.J. Alcohol, amines, and alkaloids: A possible biochemical basis for alcohol addition. Science. 1970;167:1005–1007. doi: 10.1126/science.167.3920.1005. [DOI] [PubMed] [Google Scholar]
- 266.Tsukamoto H., Machida K., Dynnyk A., Mkrtchyan H. “Second hit” models for alcoholic liver disease. Semin. Liver Dis. 2009;29:178–187. doi: 10.1055/s-0029-1214373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 267.Iorga A., Dara L., Kaplowitz N. Drug-induced liver injury: Cascade of events leading to cell death, apoptosis or necrosis. Int. J. Mol. Sci. 2017;18:1018. doi: 10.3390/ijms18051018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 268.Chitturi S., Farrell G.C. Etiopathogenesis of nonalcoholic steatohepatitis. Semin. Liver Dis. 2001;21:27–42. doi: 10.1055/s-2001-12927. [DOI] [PubMed] [Google Scholar]
- 269.Emery M.G., Fisher J.M., Chien J.Y., Kharasch E.D., Dellinger E.P., Kowdley K.V., Thummel K.E. CYP2E1 activity before and after weight loss in morbidly obese subjects with nonalcoholic fatty liver disease. Hepatology. 2003;38:428–435. doi: 10.1053/jhep.2003.50342. [DOI] [PubMed] [Google Scholar]
- 270.Teschke R., Xuan T.D. Viewpoint: A contributory role of Shell ginger (Alpinia zerumbet) for human longevity of Okinawa in Japan? Nutrients. 2018;10:166. doi: 10.3390/nu10020166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 271.Yoon E., Babar A., Choudhary M., Kutner M., Pyrsopoulos N. Acetaminophen-induced hepatotoxicity: A comprehensive update. J. Clin. Transl. Hepatol. 2016;4:131–142. doi: 10.14218/JCTH.2015.00052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 272.Teschke R. Intoxications by aliphatic halogenated hydrocarbons: Hepatotoxic risks for patients and clinical issues including role of CO2-induced hyperventilation as therapy option. J. Clin. Exp. Tox. 2018;6:1–21. doi: 10.14218/JCTH.2018.00040. [DOI] [Google Scholar]
- 273.Zeng T., Zhang C.L., Xiao M., Yang R., Xie K.Q. Critical roles of Kupffer cells in the pathogenesis of alcoholic liver disease: From basic science to clinical trials. Front. Immunol. 2016;7:538. doi: 10.3389/fimmu.2016.00538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 274.Suh Y.G., Jeong W.I. Hepatic stellate cells and innate immunity in alcoholic liver disease. World J. Gastroenterol. 2011;17:2543–2551. doi: 10.3748/wjg.v17.i20.2543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 275.Reeves H.L., Burt A.D., Wood S., Day C.P. Hepatic stellate cell activation occurs in the absence of hepatitis in alcoholic liver disease and correlates with the severity of steatosis. J. Hepatol. 1996;25:677–683. doi: 10.1016/S0168-8278(96)80238-8. [DOI] [PubMed] [Google Scholar]
- 276.DeLeve L.D. Liver sinusoidal endothelial cells in hepatic fibrosis. Hepatology. 2015;61:1740–1746. doi: 10.1002/hep.27376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 277.Apel K., Hirt H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Ann. Rev. Plant Biol. 2004;55:373–399. doi: 10.1146/annurev.arplant.55.031903.141701. [DOI] [PubMed] [Google Scholar]
- 278.Teschke R., Eickhoff A., Wolff A., Xuan T.D. Liver injury from herbs and “dietary supplements”: Highlights of a literature review from 2015 to 2017. Curr. Pharmacol. Rep. 2018;4:120–131. doi: 10.1007/s40495-018-0124-7. [DOI] [Google Scholar]
- 279.Teschke R., Melchart D., Xuan T.D. Editorial: Hormesis and dose-responses in herbal traditional Chinese medicine (TCM) alone are insufficient solving real clinical TCM challenges and associated herbal quality issues. Longhua Chin Med. 2018;19:779–793. doi: 10.21037/lcm.2018.03.01. [DOI] [Google Scholar]
- 280.Wang X., Fang H., Huang Z., Shang W., Hou T., Cheng A., Cheng H. Imaging ROS signaling in cells and animals. J. Mol. Med. (Berl) 2013;91:917–927. doi: 10.1007/s00109-013-1067-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 281.Teschke R., Wolff A., Eickhoff A., Danan G. Is obesity rather than dietary supplements used for weight reduction the cause of liver injury? J. Gastroenterol. Hepatol. Open. 2018;2:152–157. doi: 10.1002/jgh3.12057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282.Cichoż-Lach H., Michalak A. Oxidative stress as a crucial factor in liver diseases. World J. Gastroenterol. 2014;20:8082–8091. doi: 10.3748/wjg.v20.i25.8082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 283.Dey A., Cederbaum A.I. Alcohol and oxidative liver injury. Hepatology. 2006;43:S63–S74. doi: 10.1002/hep.20957. [DOI] [PubMed] [Google Scholar]
- 284.Ewing J.A. Detecting alcoholism: The CAGE questionnaire. JAMA. 1984;252:1905–1907. doi: 10.1001/jama.1984.03350140051025. [DOI] [PubMed] [Google Scholar]
- 285.Selzer M.L. The Michigan Alcohol Screening Test: The quest for a new diagnostic instrument. Am. J. Psychiatr. 1971;127:1653–1658. doi: 10.1176/ajp.127.12.1653. [DOI] [PubMed] [Google Scholar]
- 286.Saunders J.B., Aasland O.G., Babor T.F., De La Fuente J.R., Grant M. Development of the Alcohol Use Disorders Identification Test (AUDIT): WHO Collaborative Project on early detection of persons with harmful alcohol consumption—II. Addiction. 1993;88:791–804. doi: 10.1111/j.1360-0443.1993.tb02093.x. [DOI] [PubMed] [Google Scholar]
- 287.Aertgeerts B., Buntinx F., Kester A. Review: Sensitivity of the CAGE questionnaire for the 5DSM diagnosis of alcohol abuse and dependence in general clinical populations was 71% at cut points ≥2. J. Clin. Epidemiol. 2004;57:30–39. doi: 10.1016/S0895-4356(03)00254-3. [DOI] [PubMed] [Google Scholar]
- 288.Teschke R., Danan G. Review article: Diagnosis and management of drug-induced liver injury (DILI) in patients with pre-existing liver disease. Drug Saf. 2016;39:729–744. doi: 10.1007/s40264-016-0423-z. [DOI] [PubMed] [Google Scholar]
- 289.Teschke R., Danan G. Drug-induced liver injury: Is chronic liver disease a risk factor and a clinical issue? Exp. Opin. Drug Metab. Toxicol. 2017;13:425–438. doi: 10.1080/17425255.2017.1252749. [DOI] [PubMed] [Google Scholar]
- 290.Amadi C.N., Orisakwe O.E. Review. Herb-induced liver injuries in developing nations: An update. Toxics. 2018;6:24. doi: 10.3390/toxics6020024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291.Danan G., Teschke R. RUCAM in drug and herb induced liver injury: The update. Int. J. Mol. Sci. 2016;17:14. doi: 10.3390/ijms17010014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Teschke R., Danan G. Systematic review: Drug induced liver injury with analysis of alternative causes as confounding variables. Br. J. Clin. Pharmacol. 2018;84:1467–1477. doi: 10.1111/bcp.13593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 293.Teschke R., Brand A., Strohmeyer G. Induction of hepatic microsomal gamma-glutamyltransferase following chronic alcohol consumption. Biochem. Biophys. Res. Commun. 1977;75:718–724. doi: 10.1016/0006-291X(77)91531-5. [DOI] [PubMed] [Google Scholar]
- 294.Lieber C.S., DeCarli L.M. The feeding of alcohol in liquid diets: Two decades of applications and 1982 update. Alcohol. Clin. Exp. Res. 1982;6:523–531. doi: 10.1111/j.1530-0277.1982.tb05017.x. [DOI] [PubMed] [Google Scholar]
- 295.Teschke R., Neuefeind M., Nishimura N., Strohmeyer G. Hepatic gamma-glutamyltransferase activity in alcoholic fatty liver: Comparison with other liver enzymes in man and rats. Gut. 1983;24:625–630. doi: 10.1136/gut.24.7.625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 296.Nishimura M., Stein H., Berges W., Teschke R. Gamma-glutamyltransferase activity of liver plasma membrane: Induction following chronic alcohol consumption. Biochem. Biophys. Res. Commun. 1981;99:142–148. doi: 10.1016/0006-291X(81)91724-1. [DOI] [PubMed] [Google Scholar]
- 297.Nishimura M., Teschke R. Effect of chronic alcohol consumption on the activities of liver plasma membrane enzymes: Gamma-glutamyltransferase, alkaline phosphatase and 5’-nucleotidase. Biochem. Pharmacol. 1982;31:377–381. doi: 10.1016/0006-2952(82)90185-x. [DOI] [PubMed] [Google Scholar]
- 298.Teschke R., Krukenberg S., Stremmel W., Nishimura M. Enhanced biliary gamma-glutamyltransferase excretion following prolonged alcohol consumption in rats. Eur. J. Clin. Investig. 1987;17:347–353. doi: 10.1111/j.1365-2362.1987.tb02199.x. [DOI] [PubMed] [Google Scholar]
- 299.Milstein H.J., Bloomer J.R., Klatskin G. Serum bile acids in alcoholic liver disease. Am. J. Dig. Dis. 1976;21:281–285. doi: 10.1007/BF01071839. [DOI] [PubMed] [Google Scholar]
- 300.Teschke R., Koch T. Biliary excretion of gamma-glutamyltransferase: Selective enhancement by acute ethanol administration. Biochem. Pharmacol. 1986;35:2521–2525. doi: 10.1016/0006-2952(86)90049-3. [DOI] [PubMed] [Google Scholar]
- 301.Nishimura M., Teschke R. Alcohol and gamma-glutamyltransferase. Klin. Wochenschr. 1983;61:265–275. doi: 10.1007/BF01497775. [DOI] [PubMed] [Google Scholar]
- 302.Huang A., Chang B., Sun Y., Lin H., Li B., Teng G., Zou Z.S. Disease spectrum of alcoholic liver disease in Beijing 302 Hospital from 2002 to 2013. Medicine. 2017;96:e6163. doi: 10.1097/MD.0000000000006163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 303.Cohen J.A., Kaplan M.M. The SGOT/SGPT ratio an indicator of alcoholic liver disease. Dig. Dis. Sci. 1979;24:835–838. doi: 10.1007/BF01324898. [DOI] [PubMed] [Google Scholar]
- 304.Koskinas J., Kenna J.G., Bird G.L., Alexander G.J.M., Williams R. Immunglobulin A antibody to a 200-Kilodalton cytosolic acetaldehyde adduct in alcoholic hepatitis. Gastroenterology. 1992;103:1860–1867. doi: 10.1016/0016-5085(92)91445-A. [DOI] [PubMed] [Google Scholar]
- 305.Van Waes L., Lieber C.S. Glutamate dehydrogenase: A reliable marker of liver cell necrosis in the alcoholic. Br. Med. J. 1977;2:1508–1510. doi: 10.1136/bmj.2.6101.1508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 306.Blaya D., Coll M., Rodrigo-Torres D., Vila-Casadesús M., Altamirano J., Llopis M., Graupera I., Perea L., Aguilar-Bravo B., Díaz A., et al. Integrative microRNA profiling in alcoholic hepatitis reveals a role for microRNA-182 in liver injury and inflammation. Gut. 2016;65:1535–1545. doi: 10.1136/gutjnl-2015-311314. [DOI] [PubMed] [Google Scholar]
- 307.Teschke R., Schulze J., Eickhoff A., Danan G. Drug induced liver injury: Can biomarkers assist RUCAM in causality assessment? Special issue: Molecular Research on Drug Induced Liver Injury. Int. J. Mol. Sci. 2017;18:803. doi: 10.3390/ijms18040803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 308.Teschke R., Larrey D., Melchart D., Danan G. Traditional Chinese Medicine (TCM) and herbal hepatotoxicity: RUCAM and the role of novel diagnostic biomarkers such as microRNAs. Medicines. 2016;3:18. doi: 10.3390/medicines3030018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 309.Celli R., Zhang X. Pathology of alcoholic liver disease. J. Clin. Transl. Hepatol. 2014;2:103–109. doi: 10.14218/JCTH.2014.00010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 310.Mokdad A.H., Ford E.S., Bowman B.A., Dietz W.H., Vinicor F., Bales V.S., Marks J.S. Prevalence of obesity, diabetes, and obesity-related health risk factors, 2001. JAMA. 2003;289:76–79. doi: 10.1001/jama.289.1.76. [DOI] [PubMed] [Google Scholar]
- 311.Basra S., Anand B.S. Definition, epidemiology and magnitude of alcoholic hepatitis. World J. Hepatol. 2011;3:108–113. doi: 10.4254/wjh.v3.i5.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 312.Theise N.D. Histopathology of alcoholic liver disease. Clin. Liver Dis. 2013;2:64–67. doi: 10.1002/cld.172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 313.Sakhuja P. Pathology of alcoholic liver disease, can it be differentiated from nonalcoholic steatohepatitis? World J. Gastroenterol. 2014;20:16474–16479. doi: 10.3748/wjg.v20.i44.16474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 314.Liu Y.C. Histopathology of alcoholic liver disease. Clin. Liver Dis. 1998;2:753–763. doi: 10.1016/S1089-3261(05)70040-4. [DOI] [Google Scholar]
- 315.Toivola D.M., Ku N.O., Resurreccion E.Z., Nelson D.R., Wright T.L., Omary M.B. Keratin 8 and 18 hyperphosphorylation is a marker of progression of human liver disease. Hepatology. 2004;40:459–466. doi: 10.1002/hep.20277. [DOI] [PubMed] [Google Scholar]
- 316.Luedde T., Kaplowitz N., Schwabe R.F. Cell death and cell death responses in liver disease: Mechanisms and clinical relevance. Gastroenterology. 2014;147:765–783. doi: 10.1053/j.gastro.2014.07.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 317.Owens R.E., Snyder H.S., Twilla J.D., Satapathy S.K. Pharmacologic treatment of alcoholic hepatitis: Examining outcomes based on disease severity stratification. J. Clin. Exp. Hepatol. 2016;6:275–281. doi: 10.1016/j.jceh.2016.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 318.Vuittonet C.L., Halse M., Leggio L., Frichione S.B., Brickley M., Haass-Koffler C.L., Tavares T., Swift R.M., Kenna G.A. Pharmacotherapy for alcoholic patients with alcoholic liver disease. Am. J. Health Syst. Pharm. 2014;71:1265–1276. doi: 10.2146/ajhp140028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 319.Porter H.P., Simon F.R., Pope C.E., Volwiler W., Fenster L.F. Corticosteroid therapy in severe alcoholic hepatitis. A double-blind drug trial. N. Engl. J. Med. 1971;284:1350–1355. doi: 10.1056/NEJM197106172842404. [DOI] [PubMed] [Google Scholar]
- 320.Helman R.A., Temko M.H., Nye S.W., Fallon H.J. Alcoholic hepatitis. Natural history and evaluation of prednisolone therapy. Ann. Intern. Med. 1971;74:311–321. doi: 10.7326/0003-4819-74-3-311. [DOI] [PubMed] [Google Scholar]
- 321.Campra J.L., Hamlin E.M., Kirshbaum R.J., Olivier M., Redeker A.G., Reynolds T.B. Prednisone therapy in acute alcoholic hepatitis. Report of a controlled trial. Ann. Intern. Med. 1973;79:625–631. doi: 10.7326/0003-4819-79-5-625. [DOI] [PubMed] [Google Scholar]
- 322.Blitzer B.L., Mutchnick M.G., Joshi P.H., Phillips M.M., Fessel J.M., Conn H.O. Adrenocorticosteroid therapy in alcoholic hepatitis. A prospective, double-blind randomized study. Am. J. Dig. Dis. 1977;22:477–484. doi: 10.1007/BF01072499. [DOI] [PubMed] [Google Scholar]
- 323.Shumaker J.B., Resnick R.H., Galambos J.T., Makopour H., Iber F.L. A controlled trial of 6-methylprednisolone in acute alcoholic hepatitis. With a note on published results in encephalopathic patients. Am. J. Gastroenterol. 1978;69:443–449. [PubMed] [Google Scholar]
- 324.Lesesne H.R., Bozymski E.M., Fallon H.J. Treatment of alcoholic hepatitis with encephalopathy: Comparison with caloric supplements. Gastroenterology. 1978;74:169–173. [PubMed] [Google Scholar]
- 325.Maddrey W.C., Boitnott J.K., Bedine M.S., Weber F.L., Mezey E., White R.I. Corticosteroid therapy of alcoholic hepatitis. Gastroenterology. 1978;75:193–199. [PubMed] [Google Scholar]
- 326.Depew W., Boyer T., Omata M., Redeker A., Reynolds T. Double-blind controlled trial of prednisolone therapy in patients with severe acute alcoholic hepatitis and spontaneous encephalopathy. Gastroenterology. 1980;78:524–529. [PubMed] [Google Scholar]
- 327.Theodossi A., Eddleston A.L., Williams R. Controlled trial of methylprednisolone therapy in severe acute alcoholic hepatitis. Gut. 1982;23:75–79. doi: 10.1136/gut.23.1.75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 328.Mendenhall C.L., Anderson S., Garcia-Pont P., Goldberg S., Kiernan T., Seeff L.B., Sorell M., Tamburro C., Weesner R., Zetterman R., et al. The Veterans Administration Cooperative Study on Alcoholic Hepatitis. Short-term and long-term survival in patients with alcoholic hepatitis treated with oxandrolone and prednisolone. N. Engl. J. Med. 1984;311:1464–1470. doi: 10.1056/NEJM198412063112302. [DOI] [PubMed] [Google Scholar]
- 329.Bories P., Guedj J.Y., Mirouze D., Yousfi A., Michel H. Treatment of acute alcoholic hepatitis with prednisolone. 45 patients. Presse Med. 1987;16:769–772. [PubMed] [Google Scholar]
- 330.Carithers R.L., Herlong H.F., Diehl A.M., Shaw E.W., Combes B., Fallon H.J., Maddrey W.C. Methylprednisolone therapy in patients with severe alcoholic hepatitis. A randomized multicenter trial. Ann. Intern. Med. 1989;110:685–690. doi: 10.7326/0003-4819-110-9-685. [DOI] [PubMed] [Google Scholar]
- 331.Ramond M.J., Poynard T., Rueff B., Mathurin P., Theodore C., Chaput J.C., Benhamou J.P. A randomized trial of prednisolone in patients with severe alcoholic hepatitis. N. Engl. J. Med. 1992;326:507–512. doi: 10.1056/NEJM199202203260802. [DOI] [PubMed] [Google Scholar]
- 332.Mathurin P., OGrady J., Carithers R.L., Phillips M., Louvet A., Mendenhall C.L., Ramond M.J., Naveau S., Maddrey W.C., Morgan T.R. Corticosteroids improve short-term survival in patients with severe alcoholic hepatitis: Meta-analysis of individual patient data. Gut. 2011;60:255–260. doi: 10.1136/gut.2010.224097. [DOI] [PubMed] [Google Scholar]
- 333.Thursz M.R., Richardson P., Allison M., Austin A., Bowers M., Day C.P., Downs N., Gleeson D., MacGilchrist A., Grant A., et al. Prednisolone or Pentoxifylline for alcoholic hepatitis. N. Engl. J. Med. 2015;372:1619–1628. doi: 10.1056/NEJMoa1412278. [DOI] [PubMed] [Google Scholar]
- 334.Akriviadis E., Botla R., Briggs W., Han S., Reynolds T., Shakil O. Pentoxifylline improves short-term survival in severe acute alcoholic hepatitis: A double-blind, placebo-controlled trial. Gastroenterology. 2000;119:16371648. doi: 10.1053/gast.2000.20189. [DOI] [PubMed] [Google Scholar]
- 335.Naveau S., Chollet-Martin S., Dharancy S., Mathurin P., Jouet P., Piquet M.A., Davion T., Oberti F., Broët P., Emilie F. A double blind randomized controlled trial of infliximab associated with prednisolone in acute alcoholic hepatitis. Hepatology. 2004;39:1390–1397. doi: 10.1002/hep.20206. [DOI] [PubMed] [Google Scholar]
- 336.Alexander J.F., Lischner M.W., Galambos J.T. Natural history of alcoholic hepatitis. II. The long-term prognosis. Am. J. Gastroenterol. 1971;56:515–525. [PubMed] [Google Scholar]
- 337.Jepsen P., Ott P., Andersen P.K., Srensen H.T., Vilstrup H. The clinical course of alcoholic liver cirrhosis: A Danish population-based cohort study. Hepatology. 2010;51:1675–1682. doi: 10.1002/hep.23500. [DOI] [PubMed] [Google Scholar]
- 338.Saunders J.B., Walters J.R.F., Davie P., Paton A. A 20-year prospective study of cirrhosis. BMJ. 1981;282:263266. doi: 10.1136/bmj.282.6260.263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 339.Fleming K.M., Aithal G.P., Card T.R., West J. The rate of decompensation and clinical progression of disease in people with cirrhosis: A cohort study. Aliment. Pharmacol. Ther. 2010;32:1343–1350. doi: 10.1111/j.1365-2036.2010.04473.x. [DOI] [PubMed] [Google Scholar]
- 340.Powell W.J., Jr., Klatskin G. Duration of survival in patients with Laennec’s cirrhosis. Influence of alcohol withdrawal, and possible effects of recent changes in general management of the disease. Am. J. Med. 1968;44:406–420. doi: 10.1016/0002-9343(68)90111-3. [DOI] [PubMed] [Google Scholar]
- 341.Martini G.A., Teschke R. Alcohol abstinence in alcoholic liver disease. Acta Med. Scand. 1985;703:185–194. doi: 10.1111/j.0954-6820.1985.tb08914.x. [DOI] [PubMed] [Google Scholar]
- 342.O’Grady J.G. Liver transplantation alcohol related liver disease: (Deliberately) stirring a hornet’s nest! Gut. 2006;55:1529–1531. doi: 10.1136/gut.2005.090506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 343.Poynard T., Naveau S., Doffoel M., Boudjema K., Vanlemmens C., Mantion G., Messner M., Launois B., Samuel D., Cherqui D., et al. Evaluation of efficacy of liver transplantation in alcoholic cirrhosis using matched and simulated controls: 5-year survival. Multi-centre group. J. Hepatol. 1999;30:1130–1137. doi: 10.1016/S0168-8278(99)80269-4. [DOI] [PubMed] [Google Scholar]
- 344.Worner T.M., Lieber C.S. Perivenular fibrosis as precursor lesion of cirrhosis. JAMA. 1985;254:627–630. doi: 10.1001/jama.1985.03360050065027. [DOI] [PubMed] [Google Scholar]
- 345.McCullough A.J., O’Connor J.F.B. Alcoholic liver disease: Proposed recommendations for the American College of Gastroenterology. Am. J. Gastroenterol. 1998;93:2022–2036. doi: 10.1111/j.1572-0241.1998.00587.x. [DOI] [PubMed] [Google Scholar]
- 346.Testino G., Leone S., Borro P. Alcohol and hepatocellular carcinoma: A review and a point of view. World J. Gastroenterol. 2014;20:15943–15954. doi: 10.3748/wjg.v20.i43.15943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 347.Burra P., Zanetto A., Germani G. Liver transplantation for alcoholic liver disease and hepatocellular carcinoma. Cancers. 2018;10:46. doi: 10.3390/cancers10020046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 348.Day C.P., James O.F.W. Steatohepatitis: A tale of two “hits”? Gastroenterology. 1998;114:842–844. doi: 10.1016/S0016-5085(98)70599-2. [DOI] [PubMed] [Google Scholar]
- 349.Robin M.A., Anandatheerthavarada H.K., Biswas G., Sepuri N.B., Gordon D.M., Pain D., Avadhani N.G. Bimodal targeting of microsomal CYP2E1 to mitochondria through activation of an N-terminal chimeric signal by AMP-mediated phosphorylation. J. Biol. Chem. 2002;277:40583–40593. doi: 10.1074/jbc.M203292200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 350.Zhang W., Lu D., Dong W., Zhang L., Zhang X., Quan X., Ma C., Lian H., Zhang L. Expression of CYP2E1 increases oxidative stress and induces apoptosis of cardiomyocytes in transgenic mice. FEBS J. 2011;278:1484–1492. doi: 10.1111/j.1742-4658.2011.08063.x. [DOI] [PubMed] [Google Scholar]
- 351.Bansal S., Liu C.P., Sepuri N.B.V., Anandatheerthavarada H.K., Selvaraj V., Hoek J., Milne G.L., Guengerich F.P., Avadhani N.G. Mitochondria-targeted cytochrome P450 2E1 induced oxidative damage and augments alcohol-mediated stress. [(accessed on 19 September 2018)];J. Biol. Chem. 2010 285:24609–24619. doi: 10.1074/jbc.M110.121822. Available online: http://www.jbc.org/cgi/doi/10.1074/jbc.M110.121822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 352.Raza H., Prabu S.K., Robin M.A., Avadhani N.G. Elevated mitochondrial cytochrome P450 2E1 and glutathione S-transferase A4-4 in streptomycin-induced diabetic rats. Tissue-specific variations and roles in oxidative stress. Diabetes. 2004;53:185–194. doi: 10.2337/diabetes.53.1.185. [DOI] [PubMed] [Google Scholar]
- 353.Robin M.A., Sauvage I., Grandperret T., Descatoire V., Pessayre D., Fromenty B. Ethanol increases mitochondrial cytochrome 2E1 in mouse liver and rat hepatocytes. FEBS Lett. 2005;579:6895–6902. doi: 10.1016/j.febslet.2005.11.029. [DOI] [PubMed] [Google Scholar]
- 354.Nagata K., Suzuki H., Sakaguchi S. Common pathogenic mechanism in development progression of liver injury caused by non-alcoholic or alcoholic steatohepatitis. J. Toxicol. Sci. 2007;32:453–468. doi: 10.2131/jts.32.453. [DOI] [PubMed] [Google Scholar]
- 355.Cederbaum A.I. Role of CYP 2E1 in ethanol-induced oxidant stress, fatty liver and hepatotoxicity. Dig. Dis. 2010;28:802–811. doi: 10.1159/000324289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 356.Yang L., Wu D., Wang X., Cederbaum A.I. Cytochrome P4502E1, oxidative stress, JNK, and autophagy in acute alcohol-induced fatty liver. Free Radic Biol. Med. 2012;53:1170–1180. doi: 10.1016/j.freeradbiomed.2012.06.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 357.Abdelmegeed M.A., Banerjee A., Jang S., Yoo S.H., Yun J.W., Gonzalez F.J., Keshavarzian A., Song B.J. CYP2E1 potentiates binge alcohol-induced gut leakiness, steatohepatitis, and apoptosis. Free Radic Biol. Med. 2013;65:1238–1245. doi: 10.1016/j.freeradbiomed.2013.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 358.Cederbaum A.I. Methodology to assay CYP2E1 mixed function oxidase catalytic activity and its function. Redox Biol. 2014;2:1048–1054. doi: 10.1016/j.redox.2014.09.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 359.Gaviria M.M., Arango G.C., Navas M.C. Alcohol, cirrhosis, and genetic predisposition. Rev. Colomb. Gastroenterol. 2016;31:26–33. doi: 10.22516/25007440.70. [DOI] [Google Scholar]
- 360.Cho Y.E., Mezey E., Hardwick J.P., Salem N., Clemens D.L., Song B.J. Increased ethanol-inducible cytochrome P450-2E1 and cytochrome P450 isoforms in exosomes of alcohol-exposed rodents and patients with alcoholism through oxidative and endoplasmic reticulum stress. Hepatol. Commun. 2017;1:675–690. doi: 10.1002/hep4.1066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 361.Woolbright B.L., Jaeschke H. Alcoholic hepatitis: Lost in translation. J. Clin. Transl. Hepatol. 2018;6:89–96. doi: 10.14218/JCTH.2017.00054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 362.Teschke R., Zhu Y. Paracetamol (acetaminophen), alcohol, and liver injury: Biomarkers, clinical issues, and experimental aspects. SL Pharmacol. Toxicol. 2018;5:11. [Google Scholar]
- 363.Ramachandran A., Jaeschke H. Acetaminophen toxicity: novel insights into mechanisms and future perspectives. Gene Expr. 2018;18:19–30. doi: 10.3727/105221617X15084371374138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 364.Lieber C.S., Leo M.A., Wang X., DeCarli L.M. Effect of chronic alcohol consumption on hepatic SIRT1 and PGC- 1alpha in rats. Biochem. Biophys. Res. Commun. 2008;370:44–48. doi: 10.1016/j.bbrc.2008.03.005. [DOI] [PubMed] [Google Scholar]
- 365.Thompson K.J., Humphries J.R., Niemeyer D.J., Sindram D., McKillop I.H. The effect of alcohol on Sirt1 expression and function in animal and human models of hepatocellular carcinoma (HCC) Adv. Exp. Med. Biol. 2015;815:361–373. doi: 10.1007/978-3-319-09614-8_21. [DOI] [PubMed] [Google Scholar]
- 366.You M., Jogasuria A., Taylor C., Wu J. Sirtuin 1 signaling and alcoholic fatty liver disease. Hepatobiliary Surg. Nutr. 2015;4:88–100. doi: 10.3978/j.issn.2304-3881.2014.12.06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 367.Leo M.A., Rosman A.S., Lieber C.S. Differential depletion of carotenoids and tocopherol in liver disease. Hepatology. 1993;17:977–986. [PubMed] [Google Scholar]
- 368.Albano E., Vidali M. Immune mechanisms in alcoholic liver disease. Genes Nutr. 2010;5:141–147. doi: 10.1007/s12263-009-0151-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 369.Szabo G., Mandrekar P., Petrasek J., Catalano D. The unfolding web of innate immune dysregulation in alcoholic liver injury. Alcohol. Clin. Exp. Res. 2011;35:782–786. doi: 10.1111/j.1530-0277.2010.01398.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 370.Duryee M.J., Klassen L.W., Thiele G.M. Immunological response in alcoholic liver disease. World J. Gastroenterol. 2007;13:4938–4946. doi: 10.3748/wjg.v13.i37.4938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 371.Engen P.A., Green S.J., Voigt R.M., Forsyth C.B., Keshavarzian A. The gastrointestinal microbiome: Alcohol effects on the composition of intestinal microbiota. Alcohol. Res. Curr. Rev. 2015;37:223–236. doi: 10.35946/arcr.v37.2.07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 372.Rahman S., Islam R. Mammalian Sirt1: Insights on its biological functions. Cell Commun. Signal. 2011;9:11. doi: 10.1186/1478-811X-9-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 373.Udoh U.S., Valcin J.A., Gamble K.L., Bailey S.M. The molecular circadian clock and alcohol-induced liver injury. Biomolecules. 2015;5:2504–2537. doi: 10.3390/biom5042504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 374.Teschke R. Aliphatic halogenated hydrocarbons: Liver injury in 60 patients. J. Clin. Transl. Hepatol. 2018;6:1–12. doi: 10.14218/JCTH.2018.00040. [DOI] [PMC free article] [PubMed] [Google Scholar]