

Low dose of cannabidiol ameliorates mechanical allodynia and anxious behavior and restores impaired serotonergic transmission in a neuropathic pain model in rats.
Keywords: Cannabidiol, Pain, Dorsal raphe, Electrophysiology, Anxiety
Abstract
Clinical studies indicate that cannabidiol (CBD), the primary nonaddictive component of cannabis that interacts with the serotonin (5-HT)1A receptor, may possess analgesic and anxiolytic effects. However, its effects on 5-HT neuronal activity, as well as its impact on models of neuropathic pain are unknown. First, using in vivo single-unit extracellular recordings in rats, we demonstrated that acute intravenous (i.v.) increasing doses of CBD (0.1-1.0 mg/kg) decreased the firing rate of 5-HT neurons in the dorsal raphe nucleus, which was prevented by administration of the 5-HT1A antagonist WAY 100635 (0.3 mg/kg, i.v.) and the TRPV1 antagonist capsazepine (1 mg/kg, i.v.) but not by the CB1 receptor antagonist AM 251 (1 mg/kg, i.v.). Repeated treatment with CBD (5 mg/kg/day, subcutaneously [s.c.], for 7 days) increased 5-HT firing through desensitization of 5-HT1A receptors. Rats subjected to the spared nerve injury model for 24 days showed decreased 5-HT firing activity, mechanical allodynia, and increased anxiety-like behavior in the elevated plus maze test, open-field test, and novelty-suppressed feeding test. Seven days of treatment with CBD reduced mechanical allodynia, decreased anxiety-like behavior, and normalized 5-HT activity. Antiallodynic effects of CBD were fully prevented by capsazepine (10 mg/kg/day, s.c., for 7 days) and partially prevented by WAY 100635 (2 mg/kg/day, s.c., for 7 days), whereas the anxiolytic effect was blocked only by WAY. Overall, repeated treatment with low-dose CBD induces analgesia predominantly through TRPV1 activation, reduces anxiety through 5-HT1A receptor activation, and rescues impaired 5-HT neurotransmission under neuropathic pain conditions.
1. Introduction
Cannabis triggers a complex set of experiences in humans including euphoria, heightened sensitivity to external experience, and relaxation.91 The primary noneuphorizing and nonaddictive compound of cannabis, cannabidiol (CBD), has recently been shown to possess considerable therapeutic potential for treating a wide range of disorders such as chronic pain,23 nausea,64 epilepsy,28 psychosis, and anxiety.82,88 Cannabidiol in therapeutics is used within a large therapeutic window, ranging from 2.85 to 50 mg/kg/day,28,88 meaning that its therapeutic dose is still unclear. Unlike the main psychoactive ingredient of cannabis, Δ9-tetrahydrocannabinol, CBD lacks addictive properties and euphoric effects, thus representing an interesting pharmacological compound to be further investigated for potential therapeutic utility. Cannabidiol shows low affinity for the cannabinoid G-protein-coupled receptor CB1 (Ki = 2210.5 nM, rat brain)56 and allosteric agonism of the serotonin 5-HT1A receptor.15,16,72,75 Moreover, CBD interacts with the transient receptor potential cation channel subfamily V member 1 (TRPV1) channels10,45,48 and with the mammalian target of rapamycin (mTOR) signaling pathway.78 Intriguingly, several preclinical studies have demonstrated the analgesic properties of CBD.21–23,86,87 In particular, CBD (5-10 mg/kg) prevents the development of cold and mechanical allodynia in mice treated with paclitaxel.87 Moreover, CBD (10 mg/kg) has been shown to abolish carrageenan-induced hyperalgesia to a thermal stimulus in rats.22 Other clinical evidence showed the effectiveness of CBD to treat symptoms of neuropathic pain, alone85 or in combination with tetrahydrocannabinol.52,61
Serotonin (5-HT) is a neurotransmitter implicated in pain,7,89 depression, and anxiety.46,53,71 Indeed, pain is often in comorbidity with mood and anxiety disorders in humans.57,58,84 Preclinical studies have also confirmed the development of anxiety-like behaviors in mice with persistent inflammatory pain.19 Furthermore, neuropathic pain induced by injection of complete Freund adjuvant or by sciatic nerve ligation has been shown to produce a significant anxiogenic effect in the light/dark box test in mice at 4 weeks after the injection or surgery.60 In the context of mood disorders, acute CBD treatment (30 mg/kg) has been shown to exert antidepressant-like effects in the forced swim test (FST),91 whereas 14 days of CBD treatment (30 mg/kg) prevents the anxiogenic phenotype induced by chronic unpredictable stress (CUS) exposure.18 Despite these encouraging findings, few studies have explored the effect of CBD on 5-HT neurotransmission in the dorsal raphe nucleus (DRN), a brain region involved in both mood disorders35,39 and pain.62,70,77,80
Thus, the first aim of our study was to determine whether acute administration of CBD modulates DRN 5-HT neuronal activity in naive animals through 5-HT1A, CB1, or TRPV1 receptor–mediated mechanisms. We subsequently examined the effect of repeated low-dose CBD treatment on mechanical allodynia, anxiety-like behaviors, and DRN 5-HT neuronal activity in the spared nerve injury (SNI) model of neuropathic pain in rats, providing new insights into the therapeutic role of CBD and its mechanism of action.
2. Methods
2.1. Animals
Adult male Wistar rats, 6 weeks old (Charles River, Ste. Constant, Quebec, Canada), weighing 250 to 260 g on arrival were housed in groups of 2 or 3 in standard polycarbonate cages under standard laboratory conditions (12-hour light–dark cycle, lights on at 07:30 and off at 19:30; temperature at 20 ± 2°C; 50%-60% relative humidity). All experimental procedures were conducted in accordance with the guidelines set by the Canadian Institutes of Health Research for animal care and scientific use and the Animal Care Committee of McGill University (protocol number 5766). A total number of 229 animals were used. In particular, we used 114 animals for behavioral experiments (n = 6 or 9 per group) and 115 animals for electrophysiological experiments (n = 4 or 9 per group). All the behavioral experiments were conducted during the light phase between 14:00 and 18:00. In vivo extracellular recordings were mostly performed between 14:00 and 22:00.
2.2. Spared nerve injury
Neuropathic pain was induced using the SNI procedure described by Decosterd and Woolf.26 Under isoflurane anesthesia (4% induction; 1.5% maintenance), the left hind leg sciatic nerve was exposed at the level of trifurcation into the sural, tibial, and common peroneal nerves. The tibial and common peroneal nerves were tightly ligated with 4-0 silk and severed, leaving the sural nerve intact. Sham rats underwent a surgery that exposed the left sciatic nerve without further manipulation. Naive animals did not undergo any surgery. After recovery, rats were housed separately (naive, sham, and SNI) in groups of 3 to 5 individuals.
2.3. In vivo electrophysiology
Naive, sham, and SNI rats were anaesthetized with chloral hydrate (400 mg/kg, intraperitoneally [i.p.]) in their housing room and then transported in light-free boxes to the procedural room. Rats were placed in a stereotaxic apparatus (David Kopf Instruments, Tujunga, CA), and a hole was drilled through the skull according to the coordinates from rat brain atlas of Paxinos and Watson66: 1.2 mm anterior to interaural zero on the midline. Body temperature was measured using a rectal thermometer (Yellow Springs Instrument Co, Yellow Springs, OH) and was maintained at 35 to 36.5°C using an infrared (IR) heating lamp (Philips, Infrared Heat). To maintain a full anesthetic state during the experiments, supplemental doses of chloral hydrate (100 mg/kg, i.p.) were periodically administered. Anesthesia was confirmed by the absence of nociceptive reflex reaction to a tail or paw pinch and of an eye blink response to pressure. Extracellular single-unit recordings were performed using single-barreled glass micropipettes pulled from 2-mm Stoelting (Wood Dale, IL) capillary glass on a Narashige (Tokyo, Japan) PE-21 pipette puller. The micropipettes were preloaded with fiberglass strands to promote capillary filling with 2% pontamine sky blue dye in 2 M NaCl. The micropipette tips were broken down to diameters of 1 to 3 μm to reach an electrode impedance of 2 to 6 MΩ. Single-unit activity was recorded as large-amplitude action potentials captured by a software window discriminator, amplified by an AC Differential MDA-3 amplifier (BAK Electronics, Inc., FL), postamplified and band-pass filtered by a Realistic 10 band frequency equalizer, digitized by a CED 1401 interface system (Cambridge Electronic Design, Cambridge, United Kingdom), processed online, and analyzed off-line using Spike2 software version 5.20 for Windows PC. The first 30 seconds immediately after detecting the neuron was not recorded to eliminate mechanical artifacts due to electrode displacement. The spontaneous single-spike activity of the neuron was then recorded for at least 2 minutes. Once the recordings were terminated, pontamine sky blue dye was injected iontophoretically by passing a constant positive current of 20 μA for 5 minutes through the recording pipette to mark the recording site. Then, rats were decapitated and their brains were extracted and placed in a freezer at −20°C. Subsequent localization of the labeled site was made by cutting 20-μm-thick brain sections using a microtome (Leica CM 3050 S), and the electrode placement was identified with a microscope (Olympus U-TVO.5 × C-3).
2.3.1. Recording of dorsal raphe nucleus 5-HT neurons
In vivo single-unit extracellular recordings of DRN 5-HT neurons were performed as previously described.5,20 The electrode was advanced slowly into the DRN, guided by coordinates from the rat brain atlas of Paxinos and Watson66: 1.2 mm anterior to interaural zero on the midline and 5.0 to 6.5 mm from the dura mater. Under physiological conditions, spontaneously active 5-HT neurons exhibit characteristic electrophysiological properties distinguishable from non–5-HT neurons. These 5-HT neurons exhibit a slow (0.1-4 Hz) and regular firing rate (coefficient of variation [COV], ranges from 0.12 to 0.87) and a broad biphasic (positive–negative) or triphasic waveform (0.8-3.5 ms; 1.4 ms first positive and negative deflections).1,3,6 Although these criteria may vary in response to pharmacological or environmental manipulations,3,5 some spike features (ie, waveform, shape, and spike duration) have been shown to be stable across conditions and are therefore reliable indicators of 5-HT neurons. For experiments in sham and SNI rats, to estimate the cell population spontaneous activity, the electrode was passed within each area in 6 to 9 predetermined tracks separated by 200 μm. The total number of active cells encountered in each area was divided by the number of tracks to give an average number of active neurons per track. A paw pinch was delivered by hand-driven forceps exerting a force between 80 and 100 g/mm2. Firing rate, number of active neurons per descent, percentage of COV, and number of neurons responsive to hind paw pinch were analyzed. These experiments were conducted between 14:00 and 22:00 hours, as in our laboratory it has been demonstrated that the firing activity of serotonergic neurons does not significantly change in control animals according to the phase of the day.20,30
2.4. Mechanical allodynia
Mechanical allodynia was assessed in the SNI model using the up and down method.29 All animals were allowed to acclimate for ∼15 minutes on an elevated mesh platform in an enclosure. Calibrated von Frey filaments for rats (Stoelting, Wood Dale, IL, ranging from 3.61 [0.407 g] to 5.46 [26 g] bending force) were applied to the midplantar surface of the hind paw. For SNI, the sural portion of the plantar surface of the paw was stimulated with a series of ascending force von Frey monofilaments. The threshold was captured as the lowest force (g) that evoked a rapid withdrawal response to 1 of 5 repetitive stimuli.26,83 The mean 50% paw withdrawal threshold (g) was calculated for each group using Dixon's formula.23 Animals were placed in the behavioral room at 13:00 for habituation. Animals were tested at baseline before the SNI/sham procedure and at 15 (D0), 18 (D3), and 23 (D7) days after surgery. These experiments were conducted from 14:00 to 15:00 hours.
2.5. Behavioral assays
At day 23 post-SNI or sham surgery, immediately after the von Frey assessment, sham and SNI rats were tested for depressive- and anxiety-like behavior between 15:00 and 18:00. Because animals tested in the open-field test (OFT) cannot be tested in the novelty-suppressed feeding test (NSFT) (due to unwanted habituation to the novel environment), we randomly divided the animals into 2 groups. This also allowed us to minimize the ability of more invasive behavioral assays (ie, FST) to interfere with performance on subsequent assays. All behavioral experiments were conducted on day 23 after SNI or sham surgery. Sham and SNI rats treated with vehicle or CBD underwent the von Frey assessment from 14:00 to 15:00. The same cohort of rats was subjected to the OFT and FST from 15:00 to 18:00, on the same day. A different cohort of rats underwent the von Frey assessment from 14:00 to 15:00 and were tested in the elevated plus maze test (EPMT) and NSFT from 15:00 to 18:00 on the same day. To examine the pharmacological mechanisms underlying the putative analgesic and anxiolytic effects of CBD administration, separate cohorts of rats were subjected to SNI or sham surgery as described above. In these studies, rats treated with CBD plus the TRPV1 antagonist capsazepine (CPZ) (or CPZ alone) or CBD plus WAY (or WAY alone) underwent the von Frey assessment from 14:00 to 15:00 and were tested in the OFT from 15:00 to 18:00 on the same day. Another cohort of rats underwent the von Frey assessment from 14:00 to 15:00 and was subsequently tested in the EPMT and NSFT from 15:00 to 18:00.
The apparatus was cleaned before each behavioral session using a 70% EtOH solution and paper towel. Behaviors were recorded, stored, and analyzed using an automated behavioral tracking system (Videotrack, View Point Life Science) equipped with IR light-sensitive cameras. Light-phase experiments were conducted using standard room lighting (350 lux) and a white lamp (100 W), and dark-phase experiments using IR light-emitting diodes and a lamp with a red light bulb (8 lux).
2.5.1. Open-field test
Rats were placed in an OFT arena (80 × 80 × 15 cm3), and ambulatory activity (total distance travelled in centimeter), frequency, and total duration of central zone visits were recorded for 20 minutes and analyzed.3
2.5.2. Forced swim test
Passive (immobility) coping behavior was examined in the FST, as previously described.3 Cylindrical glass containers (diameter 35 cm and height 45 cm) filled to 30-cm height with water at a temperature of 24 ± 1°C were used for all swim stress exposures. Consistent with the analysis of stress-coping behaviors in the FST,68 24 hours after a 15-minute pre-exposure, rats were re-exposed for a 5-minute test session in which the duration of immobility was analyzed.
2.5.3. Elevated plus maze test
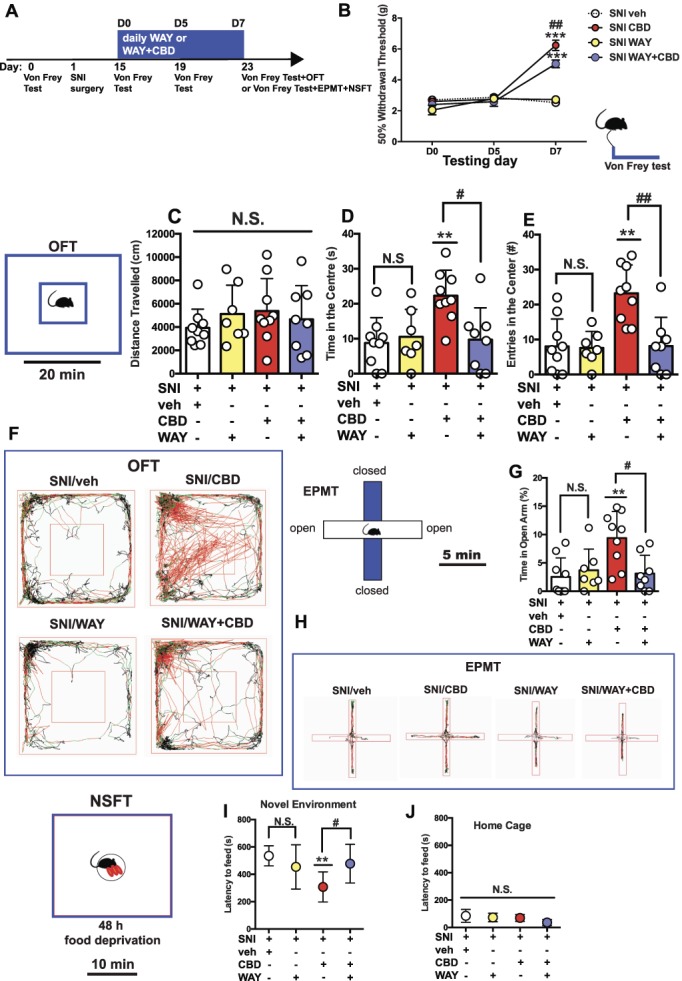
Rats were placed in a cross-shaped, elevated (80 cm) maze consisting of 2 open (50 × 10 cm2) and 2 walled (closed) (50 × 10 × 40 cm3) arms, and behavior was recorded for 5 minutes.5 Rats were singly placed in the central platform facing the open arm, and the following measures were collected: time spent (%) in the open arms and total duration (s) of head dips beyond the borders of the open arms.2 The percentage of time spent in the open arms was calculated using the formula OA% = 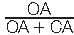 , where OA represents the time (s) spent in the open arms and CA is the time (s) spent in the closed arms.
, where OA represents the time (s) spent in the open arms and CA is the time (s) spent in the closed arms.
2.5.4. Novelty-suppressed feeding test
Latency was measured as the time it takes a rat to consume 3 chow pellets spread across the central area of an unfamiliar arena (80 × 80 × 30 cm3) after 48-hour food deprivation, as previously described.2 Before the test, feeding latency was observed in the familiar home cage. Cutoff time was 10 minutes.
2.6. Drugs
Cannabidiol (Cayman Chemical, Ann Arbor, MI) and the TRPV1 antagonist CPZ (Tocris Bioscience, Ellisville, MO) were prepared in a vehicle of ethanol/Tween 80/0.9% saline (3:1:16). AM 251 (Tocris Bioscience) was prepared in a vehicle of dimethyl sulfoxide/Tween 80/0.9% saline (1:1:18). WAY 100635 (Tocris Bioscience) and d-lysergic diethylamide acid (LSD) (Sigma-Aldrich, London, United Kingdom) were dissolved in 0.9% saline.
2.6.1. Acute treatment
For acute in vivo dose–response electrophysiological experiments, cumulative injections of CBD (0.05-0.25 mg/kg), LSD (10-50 μg/kg), and a single injection of WAY 100635 (300 μg/kg), AM 251 (1 mg/kg), or CPZ (1 mg/kg) were administered intravenously (i.v.) using a 24 G × 3/4″ catheter (Terumo Medical Corporation, Elkton, MD) inserted into the lateral tail vein of naive rats. The maximum volume used for a single injection was 0.1 mL. Once a stable DRN 5-HT neuron was found, its basal firing activity was recorded for at least 5 minutes. Naive rats were injected with vehicle (veh) and then every 5 minutes with sequential doses of CBD or LSD or with a singular injection of WAY (300 μg/kg, i.v.),37 CPZ (1 mg/kg),36 or AM 251 (1 mg/kg).51 Veh, WAY, CPZ, and AM 251 were injected 5 minutes before cumulative injection of CBD. The regimen of cumulative CBD injections took into account the pharmacokinetic properties of CBD: Cmax, Tmax, and T1/2.27,47,59,79
2.6.2. Repeated treatment
In acute electrophysiological experiments, the lowest i.v. dose of CBD able to produce a significant decrease in 5-HT neuronal activity was 0.10 mg/kg (Fig. 1). We have thus calculated the effective subcutaneous (s.c.) dose of CBD for repeated treatment by taking into account the CBD Cmax parameter (14.3 μg/mL) for a dose of 120 mg/kg i.p., and the Tmax (120 minutes).27 Considering that the average blood volume in a rat is 4.59 ± 0.57 cc/100 g body weight, which means 45.9 mL/kg,9 the total drug concentration after i.p. administration of 120 mg/kg corresponds to 0.65 mg/kg. To reach a plasmatic concentration of 0.1 mg/kg, we have approximated to 5 mg/kg/day for a s.c. administration for 7 days, taking into account the high lipophilicity of CBD in s.c. injection and an optimal steady state. Other in vivo studies reported similar low effective doses,63,65,73 in contrast with higher doses ranging from 10 to 100 mg/kg.52,66,84 Furthermore, this regimen mimics that used by patients using CBD to treat chronic neuropathic pain and anxiety.24,88
Figure 1.

Acute and repeated CBD administration on firing activity of DRN 5-HT neurons in naive rats. (A) Representation of coronal sections of the rat brain66 with the photomicrograph of the recording site in the DRN. Central gray dorsal (CGD); central gray lateral ventral (CGLV); aqueduct (Aq). The white arrow indicates the site of the electrode recording labeled with pontamine sky blue dye. (B) The typical spike waveform of 5-HT neuron. (C) Acute intravenous (i.v.) CBD administration decreases firing rate of DRN 5-HT neurons (n = 9), prevented by previous i.v. injection of WAY 100635 (WAY, n = 4) and CPZ (n = 4) but not by AM 251 (n = 4). Each point of the line represents mean ± SEM expressed as percentage of baseline before injections of veh, CBD, or antagonists. White arrow indicates the injection of veh, WAY, CPZ, or AM 251, and black arrow indicates the beginning of cumulative CBD injection. (D) WAY, CPZ, and AM 251 do not affect the basal 5-HT firing rate activity. (E-H) Representative firing rate histograms showing the acute response of 5-HT neurons to CBD alone (E) and with previous injection of WAY (F), CPZ (G), and AM 251 (H). Black arrows indicate sequence of a single injection of antagonists and of increasing doses of CBD. The cumulative doses are indicated on top of each arrow. Two-way ANOVA for RM followed by Bonferroni post hoc comparisons. *P < 0.05 and ***P < 0.001 vs veh;  P < 0.001 vs CBD. (I) Mean firing DRN 5-HT activity of naive rats treated with veh (65 recorded neurons in 4 rats) or with CBD (5 mg/kg/day, subcutaneously [s.c.], for 7 days) (77 neurons recorded in 4 rats). Each bar represents mean ± SEM and each point represents a single neuron recorded in each group. The Student unpaired 2-tailed t test. ***P < 0.001 vs veh. (J) Simple linear regression analysis showing relationship between degree of suppression of 5-HT firing activity in the DRN and the dose of LSD administered i.v. in naive rats treated for 7 days with vehicle or CBD (5.0 mg/kg/day, s.c.). ANOVA, analysis of variance; CBD, cannabidiol; CPZ, capsazepine; DRN, dorsal raphe nucleus; N.S., not significant; RM, repeated measures.
P < 0.001 vs CBD. (I) Mean firing DRN 5-HT activity of naive rats treated with veh (65 recorded neurons in 4 rats) or with CBD (5 mg/kg/day, subcutaneously [s.c.], for 7 days) (77 neurons recorded in 4 rats). Each bar represents mean ± SEM and each point represents a single neuron recorded in each group. The Student unpaired 2-tailed t test. ***P < 0.001 vs veh. (J) Simple linear regression analysis showing relationship between degree of suppression of 5-HT firing activity in the DRN and the dose of LSD administered i.v. in naive rats treated for 7 days with vehicle or CBD (5.0 mg/kg/day, s.c.). ANOVA, analysis of variance; CBD, cannabidiol; CPZ, capsazepine; DRN, dorsal raphe nucleus; N.S., not significant; RM, repeated measures.
Repeated treatment with veh or CBD (5 mg/kg, s.c.) was administered daily for 7 days in naive, sham, and SNI rats, starting from day 15 after SNI or sham surgery. Repeated treatment with WAY (2 mg/kg, s.c.)25 or CPZ (10 mg/kg, s.c.)22 was administered daily for 7 days, alone or 10 minutes before CBD administration, starting from day 15 after SNI or sham surgery. To test the involvement of TRPV1 and 5-HT1A receptors in the effects of CBD, a group of SNI rats was treated with CBD and CPZ (10 mg/kg/day, s.c., for 7 days, 10 minutes before CBD, starting from day 15 after SNI surgery), and another group was tested with CBD and WAY (2 mg/kg/day, s.c., for 7 days, 10 minutes before CBD, starting from day 15 after SNI surgery), respectively.
2.7. Statistical analysis
Data were analyzed using GraphPad Prism version 5.04 (GraphPad Software). Neuronal responses to cumulative administration of drugs were calculated as percentage of change from baseline before drug injections, were reported as mean (% of veh) ± SEM, and were computed using 2-way analysis of variance (ANOVA) for repeated measures (RM) followed by Bonferroni post hoc comparisons using treatment and pretreatment as factors. The ED50 for the suppressant effect of LSD in vehicle-treated and CBD-treated groups was compared using simple linear regression analysis comparing the slopes and elevations of the regression lines for each treatment condition, as reported by Ford et al.34 Data from all behavioral and in vivo electrophysiological experiments involving repeated CBD treatment are expressed as mean ± SEM. The Student t test was used to compare naive rats treated with veh or CBD in in vivo electrophysiological recordings. Three-way ANOVA followed by Bonferroni post hoc tests was used to analyze differences in mechanical allodynia between groups, using surgery (SNI or sham), treatment (CBD or veh), and testing day (D0, D3, and D7) as factors in the analysis. Three-way ANOVA was conducted using SigmaPlot 13 (Systat Software, Inc). Two-way ANOVA followed by Bonferroni post hoc comparisons using surgery and treatment factors was performed for behavioral and repeated treatment in vivo electrophysiological experiments in SNI animals. The chi-square test for electrophysiological population comparisons was also used. For behavioral experiments with CBD and antagonists WAY or CPZ, 2-way ANOVA for RM followed by Bonferroni post hoc comparisons using treatment and testing day factors was performed for von Frey assessments; 1-way ANOVA followed by Bonferroni post hoc comparisons was used to assess differences in performance in the OFT, EPMT, and NSFT.
3. Results
3.1. Acute cannabidiol administration decreases firing activity of dorsal raphe nucleus 5-HT neurons through 5-HT1A and TRPV1 receptors
First, we explored the mechanism of action of CBD on spontaneous firing activity of 5-HT neurons in the DRN. Veh, followed by cumulative doses of CBD, was administered to naive rats, alone or in combination with WAY 100635, AM 251, or CPZ pretreatment. Two-way RM ANOVA revealed a significant main effect of CBD treatment on the firing rate of DRN 5-HT neurons (treatment: F7, 136 = 23.29, P < 0.001) (Fig. 1C). Bonferroni post hoc tests revealed that CBD doses of 0.10 mg/kg significantly decreased the 5-HT firing rate compared with vehicle preinjection (P < 0.05, n = 9) (Figs. 1C and E) and, importantly, the dose of 0.25 mg/kg completely shut down neuronal activity (P < 0.001, n = 9) (Figs. 1C and E).
Separate cohorts of rats were pretreated with WAY 100635 (0.3 mg/kg, i.v.) or CPZ (1 mg/kg, i.v.) before CBD treatment, and single-unit extracellular 5-HT recordings were performed in the DRN. Two-way ANOVA analysis revealed a main effect of antagonist pretreatment on the firing rate of DRN 5-HT neurons (pretreatment: F3, 136 = 81.82, P < 0.001; interaction: F21, 136 = 5.068, P < 0.001). In particular, Bonferroni post hoc comparison revealed that pretreatment with WAY and CPZ each prevented the suppressive effect of CBD at doses of 0.25, 0.5, 1, 1.5, and 2 (P < 0.001, n = 4) (Figs. 1C, F and G). Thus, the ability of CBD to inhibit DRN 5-HT firing was abolished in the presence of either of these antagonists, confirming that CBD decreases 5-HT firing through 5-HT1A and TRPV1 receptor–mediated mechanisms. To assess whether CB1 receptors were also involved in this phenomenon, another cohort of rats (n = 4) received administration of the CB1 receptor antagonist AM 251 before CBD treatment, and single-unit extracellular 5-HT recordings were performed in the DRN. Bonferroni post hoc tests revealed that CBD administered at a dose of 0.25 mg/kg (P < 0.001) led to a complete cessation of 5-HT firing even with AM 251 pretreatment compared to veh (Figs. 1C and H). Thus, AM 251 did not prevent the ability of 0.25 mg/kg dose of CBD to decrease 5-HT firing, ruling out the involvement of CB1 receptors in the suppressive effect of CBD on 5-HT firing. Importantly, 2-way ANOVA revealed that the pretreatment with all these antagonists did not affect the basal activity of 5-HT neurons. Specifically, there was no significant effect of treatment (F2, 18 = 0.2781, P = 0.7604) or time after injection (F1, 18 = 0.1148, P = 0.7387), and there was no significant interaction (F2, 18 = 0.1028, P = 0.9028) (Fig. 1D).
3.2. Repeated cannabidiol treatment increases firing activity of dorsal raphe nucleus 5-HT neurons through desensitization of 5-HT1A receptors
We then tested the extent to which repeated administration of CBD modulates 5-HT neurotransmission. Rats were treated with CBD (5 mg/kg, s.c.) for 7 days. The unpaired 2-tailed t test revealed that after 7 days of treatment, CBD (77 neurons recorded in 4 rats) significantly increased the mean firing rate of DRN 5-HT neurons compared with veh (65 neurons recorded in 4 rats) (t140 = 5.059, P < 0.001) (Fig. 1I). Somatodendritic 5-HT1A autoreceptors play a major role in regulating 5-HT firing activity in the DRN. To determine whether the increase in DRN 5-HT firing after repeated CBD administration was due to an alteration in 5-HT1A autoreceptor sensitivity, cumulative doses of the 5-HT1A receptor agonist LSD42–44 were acutely injected i.v. in anesthetized rats pretreated with CBD or veh for 7 days (Fig. 1J). LSD reduced DRN 5-HT firing activity in both CBD- and veh-treated rats. The median effective dose (ED50) of LSD was 17.07 µg/kg in veh-treated rats and 31.05 µg/kg in CBD-treated rats. Thus, chronic CBD administration shifted the dose–response effect of LSD on 5-HT firing to the right. A linear regression analysis revealed statistically significant differences of slope (F1, 8 = 2.66, P = 0.04) and of regression lines (F1, 9 = 7.74, P = 0.02) (Fig. 1J). Together, these data suggest that 7-day treatment with CBD reduced the sensitivity of 5-HT neurons to the suppressive effect of LSD, likely through desensitization of 5-HT1A autoreceptors in the DRN.
3.3. Repeated cannabidiol treatment increases paw withdrawal threshold after spared nerve injury surgery
Next, we induced neuropathic pain in rats using SNI, and after 15 days from the surgery, a group of rats was treated with CBD for 7 days. Rats were tested at baseline before the SNI/sham procedure and at day 15 (D0), 18 (D5), and 23 (D7) after surgery using von Frey filaments. Three-way ANOVA analysis revealed that mechanical withdrawal threshold measured before surgery in sham-operated rats significantly decreased on the ipsilateral side 15 days after SNI surgery (surgery: F1, 78 = 154.23, P < 0.001; interaction surgery × testing day: F2, 78 = 4.78, P = 0.01; no interaction for factors surgery × treatment) (Fig. 2B). Conversely, the mechanical withdrawal threshold in the contralateral side did not differ from the mechanical withdrawal threshold measured before surgery. Administration of veh did not change the withdrawal threshold in sham (n = 6) and SNI rats (n = 9) (Fig. 2B). Seven days of treatment with CBD (5 mg/kg, s.c.) significantly increased the withdrawal threshold in SNI rats (n = 9) at the 7th day of administration (treatment: F1, 78 = 8.21, P = 0.005; interaction treatment × testing day: F2, 78 = 4.11, P = 0.02) (Fig. 2B). Bonferroni post hoc comparisons revealed a significant difference in withdrawal threshold between SNI rats treated with CBD and SNI rats treated with veh (P < 0.001). Cannabidiol treatment did not modify mechanical threshold in sham rats (n = 6) (Fig. 2B). Moreover, 3-way ANOVA analysis showed a significant interaction for all 3 factors (surgery × treatment × testing day) (F2, 78 = 5.33, P = 0.007). In particular, post hoc comparisons revealed significant differences between SNI/veh and sham/veh rats on testing days D0, D3, and D7 (P < 0.001) (Fig. 2B) and between SNI/veh and SNI/CBD rats on testing day D7 (P < 0.001) (Fig. 2B).
Figure 2.

Repeated CBD treatment increases paw withdrawal threshold and prevents anxiety-like behavior in the OFT, EPMT, and NSFT after SNI surgery. (A) Scheme illustrating the timing of SNI induction, behavioral and electrophysiological experiments. (B) Mechanical hypersensitivity was measured with von Frey filaments at day 15 (D0), 19 (D5), and 23 (D7) after SNI surgery. Sham rats treated with veh (n = 6) or CBD (5 mg/kg/day, subcutaneously [s.c.], for 7 days, n = 6) and SNI rats treated with veh (n = 9) and CBD (5 mg/kg/day, s.c., for 7 days, n = 9) were tested. Each point of the line represents mean ± SEM expressed as mean of 50% paw withdrawal threshold (g). Three-way ANOVA followed by Bonferroni post hoc comparisons. ***P < 0.001 vs sham/veh; ###P < 0.001 vs SNI/veh at D7. (C) Horizontal movement traces in the OFT of sham and SNI rats treated with veh or CBD (5 mg/kg/day, s.c., for 7 days). (D-F) Distance travelled (cm) (D), time spent (s) in the central area (E), and number of entries in the central area (F) of the OFT. Sham rats treated with veh (n = 9) or CBD (5 mg/kg/day, s.c., for 7 days, n = 9) and SNI rats treated with veh (n = 9) and CBD (5 mg/kg/day, s.c., for 7 days, n = 9) were tested. Each bar represents mean ± SEM. In D-F, each point represents data from an individual rat. Two-way ANOVA followed by Bonferroni post hoc comparisons. *P < 0.05 and ***P < 0.001 vs sham/veh; ###P < 0.001 vs SNI/veh. (G) Time of immobility (s) in the FST. (H and I) Percentage of time (%) (H) spent in the open arms in the EPMT and head dipping duration (s) (I). Each bar represents mean ± SEM. (J) Horizontal movement traces in the EPMT of sham and SNI rats treated with veh or CBD (5 mg/kg/day, s.c., for 7 days). (K and L) Latency to feed (s) in the novel (K) and familiar (L) environments in the NSFT. Each symbol represents mean ± SEM. Sham rats treated with veh (n = 8) or CBD (5 mg/kg/day, s.c., for 7 days, n = 9) and SNI rats treated with veh (n = 9) and CBD (5 mg/kg/day, s.c., for 7 days, n = 9) were tested. In G-I, each point represents data from an individual rat. Two-way ANOVA followed by Bonferroni post hoc comparisons. **P < 0.01 vs sham/veh., #P < 0.05, and ##P < 0.01 vs SNI/veh. ANOVA, analysis of variance; CBD, cannabidiol; EPMT, elevated plus maze test; FST, forced swim test; N.S., not significant; NSFT, novelty-suppressed feeding test; OFT, open-field test; SNI, spared nerve injury.
3.4. Repeated cannabidiol treatment reverses spared nerve injury–induced deficits in open-field exploration but has no effect in the forced swim test
Immediately after the von Frey measurements, the same cohort of rats used in the measurement of paw withdrawal threshold was tested in the FST and OFT. Two-way ANOVA analysis performed with surgery and treatment as factors did not reveal any significant difference in locomotion between SNI and sham rats (surgery: F1, 32 = 1.56, P = 0.22; CBD treatment: F1, 32 = 2.03, P = 0.16; and no interaction: F1, 32 = 0.92, P = 0.34) (Fig. 2D). We found that SNI rats (n = 9) showed both decreased time (P < 0.01) (Fig. 2E) and number of entries (P < 0.05) (Fig. 2F) in the center quadrant compared with sham animals (n = 9). Importantly, 7 days of CBD administration increased the time spent in the center (surgery: F1, 32 = 9.83, P = 0.003; treatment: F1, 32 = 5.60, P = 0.02; and interaction: F1, 32 = 8.31, P = 0.007) (Fig. 2E) and increased the number of entries in the center (surgery: F1, 32 = 12.17, P = 0.50; CBD treatment: F1, 32 = 12.54, P = 0.001; and interaction: F1, 32 = 12.17, P = 0.001) (Fig. 2F). Cannabidiol treatment produced no effect in sham animals (n = 9). Immediately after performing the OFT, rats were tested in the FST. Spared nerve injury surgery did not induce changes in immobility time in the FST (F1, 31 = 0.43, P = 0.51, n = 9) (Fig. 2G). There was also no main effect of treatment (F1, 31 = 0.43, P = 0.51) (Fig. 2G) or a significant surgery × treatment interaction (F1, 31 = 0.1065, P = 0.7464).
3.5. Repeated cannabidiol treatment reverses spared nerve injury–induced anxiety-like behavior
A second cohort was subsequently used to assay anxiety-like behavior in the EPMT and NSFT. We found that SNI rats (n = 9) spent less time in the open arms (P < 0.01) in the EPMT compared with sham (n = 8) (surgery: F1, 29 = 13, P = 0.001) (Fig. 2H). Cannabidiol treatment increased the time spent in the open arms in SNI rats (P < 0.05, n = 9) compared with sham rats (n = 9) (treatment: F1, 29 = 4.33, P = 0.04; interaction: F1, 29 = 5.49, P = 0.02). Cannabidiol had no significant effect in sham rats (n = 9) when compared with veh-treated sham rats (n = 8) (Fig. 2H). No significant difference was found in the total duration of head dips between SNI and sham animals treated with either CBD or veh (surgery: F1, 29 = 0.02, P = 0.87; treatment: F1, 29 = 0.08, P = 0.77) (Fig. 2I).
In the NSFT, SNI rats (n = 9) displayed a longer latency to eat in a new environment (P < 0.01) when compared with sham rats (n = 8) (surgery: F1, 29 = 4.49, P = 0.04) (Fig. 2K). Importantly, 7-day CBD treatment reduced the latency to feed in SNI rats (n = 9, P < 0.01) to levels comparable with sham rats (n = 9) (treatment: F1, 29 = 6.15, P = 0.01; interaction: F1, 29 = 7.39, P = 0.01). No differences were observed for the latency to eat in the home cage (Fig. 2L).
3.6. TRPV1 channels are required for the antinociceptive (but not the anxiolytic) effects of repeated cannabidiol administration in spared nerve injury rats
To investigate the possible role of TRPV1 channels in the behavioral effects of CBD, we pretreated SNI rats with CPZ, (10 mg/kg/day, s.c., for 7 days, 10 minutes before CBD). Because CBD did not exert analgesic and anxiolytic effects in sham animals (Fig. 2), this set of experiments was performed only in neuropathic rats to reduce the number of experimental animals. Two-way ANOVA revealed a main effect of treatment (F3, 30 = 33.58, P < 0.001) and testing day (F2, 60 = 18.25, P < 0.001) on mechanical allodynia in rats treated with CBD ± CPZ. Moreover, we observed a significant treatment × testing day interaction (F6, 60 = 28.13, P < 0.001) (Fig. 3B). Bonferroni post hoc analysis indicated that at D7, SNI rats treated with CBD + CPZ were significantly different from SNI animals receiving CBD alone (n = 9) (P < 0.001), but they did not differ from SNI animals treated with veh (n = 9) (Fig. 3B), suggesting that TRPV1 antagonism prevented the antiallodynic effects of CBD. Importantly, treatment with CPZ alone (n = 7) did not alter the mechanical threshold compared with SNI veh-treated rats (n = 9) (Fig. 3B).
Figure 3.
TRPV1 antagonism blocked the antinociceptive, but not the anxiolytic effects, of repeated CBD administration in SNI rats. (A) Scheme illustrating the timing of SNI induction, behavioral and electrophysiological experiments. (B) Mechanical hypersensitivity was measured with von Frey filaments at day 15 (D0), 19 (D5), and 23 (D7) after SNI surgery. Spared nerve injury rats treated with veh (n = 9), CBD alone (5 mg/kg/day, subcutaneously [s.c.], for 7 days, n = 9), or in combination with CPZ (10 mg/kg/day, s.c., for 7 days, 10 minutes before CBD, n = 8) and CPZ alone (10 mg/kg/day, s.c., for 7 days, n = 7) were tested. Each point of the line represents mean ± SEM expressed as mean of 50% paw withdrawal threshold (g). Two-way ANOVA for repeated measures followed by Bonferroni post hoc comparisons. ***P < 0.001 vs SNI/veh; ###P < 0.001 vs SNI/CPZ + CBD at D7. (C) Horizontal movement traces in the OFT of SNI rats treated with veh, CBD alone (5 mg/kg/day, s.c., for 7 days), or in combination with CPZ (10 mg/kg/day, s.c., for 7 days, 10 minutes before CBD) and CPZ alone (10 mg/kg/day, s.c., for 7 days). (D-F) Distance travelled (cm) (D), time spent (s) in the central area (E), and number of entries in the central area (F) of the OFT. Spared nerve injury rats treated with veh (n = 9), CBD alone (5 mg/kg/day, s.c., for 7 days, n = 9), or in combination with CPZ (10 mg/kg/day, s.c., for 7 days, 10 minutes before CBD, n = 8) and CPZ alone (2 mg/kg, s.c., for 7 days, n = 7) were tested. Each bar represents mean ± SEM. In D-F, each point represents data from an individual rat. One-way ANOVA followed by Bonferroni post hoc comparisons. *P < 0.05 and **P < 0.01 vs SNI/veh. (G) Percentage of time (%) spent in the open arms in the EPMT. Each bar represents mean ± SEM. (H) Horizontal movement traces in the EPMT of SNI rats treated with veh (n = 9), CBD alone (5 mg/kg/day, s.c., for 7 days, n = 9), or in combination with CPZ (10 mg/kg/day, s.c., for 7 days, 10 minutes before CBD, n = 8) and CPZ alone (10 mg/kg/day, s.c., n = 7) were tested. (I and J) Latency to feed (s) in the novel (I) and familiar (J) environments in the NSFT. Each symbol represents mean ± SEM. In G, each point represents data from an individual rat. One-way ANOVA followed by Bonferroni post hoc comparisons. *P < 0.05, **P < 0.01, and ***P < 0.001 vs SNI/veh. ANOVA, analysis of variance; CBD, cannabidiol; CPZ, capsazepine; EPMT, elevated plus maze test; N.S., not significant; NSFT, novelty-suppressed feeding test; OFT, open-field test; SNI, spared nerve injury.
Immediately after the von Frey test, animals were tested in the OFT. One-way ANOVA did not reveal any significant difference in the total distance travelled in the novel arena (F3, 29 = 1.653, P = 0.1989) (Fig. 3C). Intriguingly, treatment with CBD + CPZ failed to block the anxiolytic effects of CBD measured as time spent in the central quadrant (F3, 29 = 8.446, P = 0.0003, Fig. 3D) and number of entries in the center of the novel arena (F3, 29 = 9.391, P = 0.0002, Fig. 3E). Indeed, Bonferroni post hoc tests did not reveal statistical differences between SNI rats treated with CBD alone (n = 9) and SNI rats treated with CBD + CPZ (n = 8) in both time in the center (Fig. 3D) and number of entries in the center (Fig. 3E). Notably, treatment with CPZ alone (n = 7) did not induce changes in the time spent in the center (Fig. 3D) or the number of entries in the center (Fig. 3E), compared with SNI rats treated with veh (n = 9). Another cohort of rats that underwent the von Frey assessment was tested in the EPMT. In keeping with the results in the OFT, the injection of CPZ before CBD did not alter the anxiolytic effects of CBD, measured as the percentage of time spent in the open arms (F3, 29 = 5.686, P = 0.003, Fig. 3G). In particular, the Bonferroni post hoc test did not reveal statistical differences between SNI rats treated with CBD alone (5 mg/kg/day, s.c., for 7 days) (n = 9) and SNI rats treated with CBD + CPZ (Fig. 3G). Of note, treatment with CPZ alone (n = 7) did not induce any change in measures of anxiety-like behavior when compared with veh treatment in SNI rats (n = 9). Immediately after the EPMT, rats were tested in the NSFT. Spared nerve injury rats treated with CBD alone (n = 9) did not differ from SNI rats treated with CBD + CPZ (n = 8) (F3, 29 = 10.98, P < 0.001, Fig. 3I), further confirming that the anxiolytic properties of CBD in this model of neuropathic pain are not mediated by TRPV1 receptors. Importantly, treatment with CPZ alone did not affect anxiety-like behavior in SNI rats (Fig. 3I). No treatment differences were observed when the latency to eat in the home cage was analyzed (Fig. 3J). Overall, these results suggest that TRPV1 receptors are necessary for the antinociceptive effects, but not the anxiolytic effects, of CBD in the SNI model of neuropathic pain.
3.7. 5-HT1A receptors are required for the anxiolytic (but not the antinociceptive) effects of repeated cannabidiol administration in spared nerve injury rats
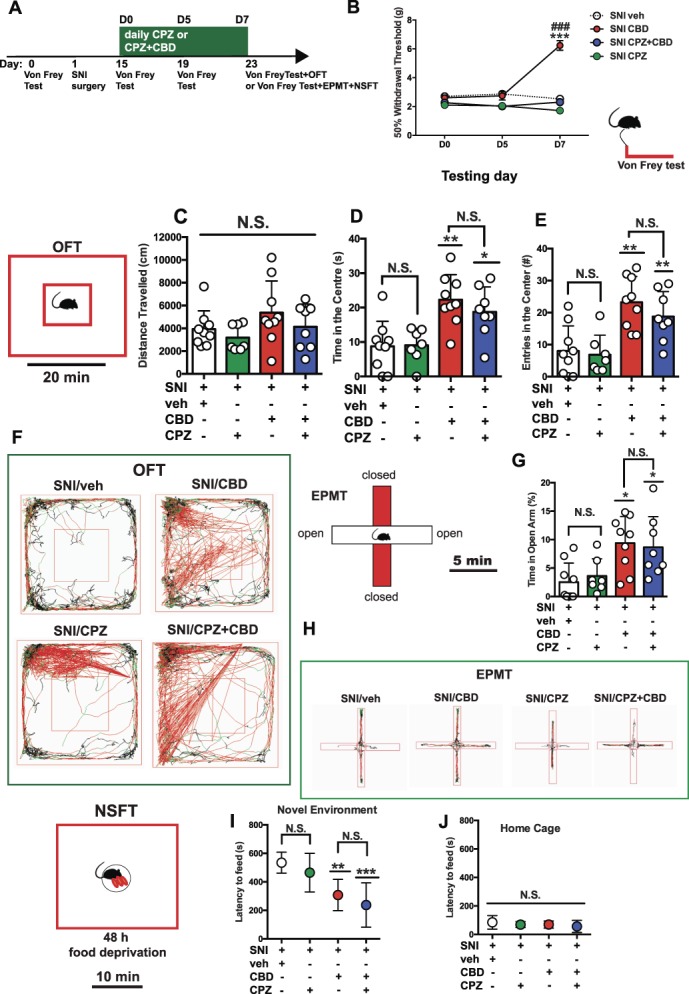
We also tested the possible implication of 5-HT1A receptors in the analgesic and anxiolytic effects of CBD in rats subjected to neuropathic pain by pretreating SNI rats with the 5-HT1A receptor antagonist WAY 100635 (WAY; 2 mg/kg/day, s.c., for 7 days, 10 minutes before CBD). In the von Frey filament test, 2-way ANOVA revealed main effects of treatment (F3, 30 = 15.00, P < 0.001), testing day (F2, 60 = 67.79, P < 0.001), and a significant treatment × testing day interaction (F6, 60 = 24.67, P < 0.001) (Fig. 4B). Bonferroni post hoc analyses revealed that at D7, SNI rats treated with CBD + WAY (n = 8) were significantly different from both SNI rats treated with veh (n = 9) (P < 0.001) and CBD alone (5 mg/kg/day, s.c., for 7 days) (n = 9) (P < 0.01). Interestingly, 2-way ANOVA comparing mechanical threshold of SNI rats treated with CBD alone vs CBD + WAY vs CBD + CPZ revealed a main effect of treatment (F2, 22 = 27.86, P < 0.001), testing day (F2, 44 = 85.02, P < 0.001), and a significant treatment × testing day interaction (F4, 44 = 20.19, P < 0.001) (data not shown in the figure). Importantly, post hoc comparison showed that SNI rats treated with CBD + WAY had a significantly lower mechanical threshold compared with SNI rats receiving CBD + CPZ (P < 0.001) (data not shown in figures). Thus, pretreatment with WAY blocked the antinociceptive effects of CBD to a lesser extent than less CPZ. Treatment with WAY alone did not induce changes in mechanical threshold (n = 7), compared to SNI rats treated with veh (n = 9) (Fig. 4B). These data indicate that the antinociceptive effects of CBD are only partially mediated by 5-HT1A receptors.
Figure 4.
5-HT1A antagonism partially prevented antinociception and completely blocked the anxiolytic properties of repeated CBD administration in SNI rats. (A) Scheme illustrating the timing of SNI induction, behavioral and electrophysiological experiments. (B) Mechanical hypersensitivity was measured with von Frey filaments at day 15 (D0), 19 (D5), and 23 (D7) after SNI surgery. Spared nerve injury rats treated with veh (n = 9), CBD alone (5 mg/kg/day, subcutaneously [s.c.], for 7 days, n = 9), or in combination with WAY (2 mg/kg/day, s.c., for 7 days, 10 minutes before CBD, n = 8) and WAY alone (2 mg/kg/day, s.c., for 7 days, n = 7) were tested. Each point of the line represents mean ± SEM expressed as mean of 50% paw withdrawal threshold (g). Two-way ANOVA for repeated measures followed by Bonferroni post hoc comparisons. ***P < 0.001 vs SNI/veh; ##P < 0.01 vs SNI/WAY + CBD at D7. (C) Horizontal movement traces in the OFT of SNI rats treated with veh, CBD alone (5 mg/kg/day, s.c., for 7 days), or in combination with CPZ (10 mg/kg/day, s.c., for 7 days, 10 minutes before CBD) and WAY alone (2 mg/kg/day, s.c., for 7 days). (D-F) Distance travelled (cm) (D), time spent (s) in the central area (E), and number of entries in the central area (F) of the OFT. Spared nerve injury rats treated with veh (n = 9), CBD alone (5 mg/kg/day, s.c., for 7 days, n = 9), or in combination with WAY (2 mg/kg/day, s.c., for 7 days, 10 minutes before CBD, n = 8) and WAY alone (2 mg/kg, s.c., for 7 days, n = 7) were tested. Each bar represents mean ± SEM. In D-F, each point represents data from an individual rat. One-way ANOVA followed by Bonferroni post hoc comparisons. **P < 0.01 vs SNI/veh; #P < 0.05; and ##P < 0.01 vs SNI/CBD. (G) Percentage of time (%) spent in the open arms in the EPMT. Each bar represents mean ± SEM. (H) Horizontal movement traces in the EPMT of SNI rats treated with veh (n = 9), CBD alone (5 mg/kg/day, s.c., for 7 days, n = 9), or in combination with WAY (2 mg/kg/day, s.c., for 7 days, 10 minutes before CBD, n = 8) and WAY alone (2 mg/kg/day, s.c., n = 7) were tested. (I and J) Latency to feed (s) in the novel (I) and familiar (J) environments in the NSFT. Each symbol represents mean ± SEM. In G, each point represents data from an individual rat. One-way ANOVA followed by Bonferroni post hoc comparisons. **P < 0.01 vs SNI/veh; #P < 0.05 vs SNI/CBD. ANOVA, analysis of variance; CBD, cannabidiol; CPZ, capsazepine; EPMT, elevated plus maze test; N.S., not significant; NSFT, novelty-suppressed feeding test; OFT, open-field test; SNI, spared nerve injury.
Immediately after the von Frey test, animals were tested in the OFT. One-way ANOVA did not reveal any significant difference in the distance travelled (F3, 29 = 0.5803, P = 0.6327 [Fig. 4C]) between veh, CBD, or CBD + WAY treatment groups. Intriguingly, treatment with CBD + WAY completely prevented the anxiolytic effects of CBD (5 mg/kg/day, s.c., for 7 days, 10 minutes after WAY) as measured by the time spent (F3, 29 = 5.746, P = 0.0033, Fig. 4D) and total number of entries (F3, 29 = 9.134, P = 0.0002, Fig. 4E) in the central part of the novel arena. Bonferroni post hoc tests revealed statistically significant differences between SNI rats treated with CBD alone (n = 9) and SNI rats treated with CBD + WAY (n = 8) in both time in the center (P < 0.05) (Fig. 4D) and entries in the center (P < 0.01) (Fig. 4E). Treatment with WAY alone (n = 7) did not alter the amount of time spent in the center (Fig. 4D) or the number of entries in the center (Fig. 4E), compared to SNI rats treated with veh (n = 9). Another cohort of rats was subsequently tested in the EPMT, which revealed significant differences between CBD and CBD + WAY on the percentage of time spent in the open arms (F3, 29 = 6.1116, P = 0.002, Fig. 4G). Similar to the OFT, the anxiolytic effects of CBD were not present when CBD was given in combination with WAY. Bonferroni post hoc tests indicated statistically significant differences between SNI rats treated with CBD alone (n = 9) and SNI rats treated with CBD + WAY (n = 8) (P < 0.05) (Fig. 4G). Moreover, treatment with WAY alone (n = 7) did not induce changes per se compared to SNI rats treated with veh (n = 9). Immediately after the EPMT, rats were tested in the NSFT. Similar results were observed when the latency to feed in the novel environment was measured (F3, 29 = 5.580, P < 0.01, Fig. 4I). Bonferroni post hoc comparisons revealed that the latency to feed in SNI rats treated with CBD alone (n = 9) was significantly shorter than that in SNI rats treated with CBD + WAY (P < 0.05) (n = 8), which was similar to that in animals receiving veh or WAY alone (Fig. 4I). No differences were observed for the latency to eat in the home cage (Fig. 4J). Overall, these results suggest that in the SNI model of neuropathic pain, 5-HT1A receptors modestly contribute to the antinociceptive effects of CBD, but fundamentally contribute to its anxiolytic properties.
3.8. Repeated cannabidiol treatment reverses spared nerve injury–induced deficits in dorsal raphe nucleus 5-HT neuronal activity
Finally, we examined the spontaneous activity of 5-HT DRN neurons in sham and SNI rats treated with vehicle and CBD (5 mg/kg, s.c., for 7 days). The average firing rate of DRN neurons was decreased in SNI rats (n = 86 neurons recorded in 4 rats) when compared with sham rats (n = 167 neurons recorded in 5 rats) (surgery: F1, 446 = 53.33, P < 0.001) (Fig. 5A).
Figure 5.

Repeated CBD treatment prevents SNI-induced alterations in DRN 5-HT neuronal activity. (A) Mean DRN 5-HT firing activity (spikes/s). (B) Representative firing rate histograms of the mean DRN 5-HT firing rate activity recorded in sham and SNI rats treated with veh or CBD (5.0 mg/kg/day, for 7 days, subcutaneously [s.c.]). (C) Mean DRN 5-HT COV (%). Each bar represents mean ± SEM. Two-way ANOVA followed by Bonferroni post hoc comparisons. (D) Representative rate histograms recorded from a DRN neuron excited (top) or not (bottom) by mechanical pinch to the operated hind paw. (E) Contingency interleaved bars showing percentage (%) of DRN 5-HT neurons excited or not by mechanical paw pinch stimulation. (F) Contingency stacked bars showing percentage (%) of bursting and nonbursting DRN 5-HT neurons (the χ2 test). (G) Mean number of DRN 5-HT neuron recorded per electrode descent. Sham rats treated with veh (n = 5) or CBD (5 mg/kg/day, s.c., for 7 days) (n = 5) and SNI rats treated with veh (n = 4) and CBD (5 mg/kg/day, s.c., for 7 days) (n = 7) were tested. Two-way ANOVA followed by Bonferroni post hoc comparisons. In A and C, each point represents a single neuron recorded in each group. In F, each point represents data from an individual rat. **P < 0.01, ***P < 0.001 vs sham/veh., #P < 0.05, and ###P < 0.001 vs SNI/veh. ANOVA, analysis of variance; CBD, cannabidiol; COV, coefficient of variation; DRN, dorsal raphe nucleus; SNI, spared nerve injury.
The analysis of COV of interspike intervals revealed that DRN neurons fired more irregularly in SNI rats when compared with sham rats (surgery: F1, 444 = 119.2, P < 0.001) (Fig. 5C). Seven-day treatment with CBD (5 mg/kg, s.c.) prevented SNI-induced reductions in the mean firing rate of DRN 5-HT neurons (n = 113 neurons recorded in 7 rats) (treatment: F1, 446 = 72.84, P < 0.001, interaction with factor surgery: F1, 446 = 7.45, P = 0.006) and COV (treatment: F1, 446 = 18.54, P < 0.001, interaction with factor surgery: F1, 444 = 18.99, P < 0.001). Similar to the results collected in naive animals (Fig. 1I), post hoc analysis revealed that sham rats treated with CBD (n = 84 neurons recorded in 5 rats) also had an increased firing rate of 5-HT neurons compared with veh (P < 0.001), whereas no difference was found in the COV between sham rats treated with CBD and veh (Figs. 5A and C).
Finally, we investigated the effect of an acute nociceptive stimulus on 5-HT firing in both SNI and sham rats that received repeated injections of CBD or veh. After 2 minutes of stable baseline firing, we applied a 2-second mechanical pinch and found cells that were either responsive (increased their firing rate) or unresponsive (no change in firing rate) during the 5-second interval after the stimulus onset. The chi-square analysis revealed that SNI rats showed an increased percentage of pinch-responsive cells compared with sham rats. Cannabidiol treatment reduced this percentage to that seen in the sham rats (the χ2 test for cell population sham/(veh or CBD) vs SNI/(veh or CBD): 16.33, df: 3, P < 0.001) (Fig. 5E). No significant difference was found in the percentage of neurons discharging burst activity in SNI rats compared with sham or in the numbers of active neurons per descent (Figs. 5F and G).
4. Discussion
We examined the extent to which CBD modulates nociception, anxiety-like behavior, and serotonergic transmission in a rodent model of neuropathic pain. Cumulative dosing of CBD decreased DRN 5-HT firing, which was blocked by pretreatment with the 5-HT1A antagonist WAY or the TRPV1 antagonist CPZ, but not with the CB1 receptor antagonist AM 251. Repeated low-dose CBD treatment increased 5-HT firing activity by desensitizing 5-HT1A autoreceptors. Using the SNI model of neuropathic pain, we found that repeated CBD treatment was able to prevent mechanical allodynia and anxiety-like behavior in rats experiencing neuropathic pain, but through different mechanisms. Specifically, TRPV1 channels were required for the antiallodynic (but not the anxiolytic) effects of CBD, whereas 5-HT1A receptors were required for the anxiolytic (and to a lesser extent the antiallodynic) effects of CBD in rats subjected to the SNI model. In vivo extracellular recordings revealed decreased firing rate, increased irregular firing activity, and increased number of 5-HT neurons responsive to mechanical stimulation in SNI rats compared with sham-operated animals. Importantly, treatment with CBD prevented SNI-induced reductions in 5-HT firing activity.
Cannabidiol can regulate the activity of peroxisome proliferator–activated receptor gamma (PPARγ),15 5-HT1A receptor,75 adenosine transporter, members of the TRP family,48,69 and the G-protein-coupled receptor 5550 to name a few mechanisms. Interestingly, CBD acts similarly to the selective serotonin reuptake inhibitor zimelidine, decreasing the firing of 5-HT neurons after acute injection through stimulation of 5-HT1A autoreceptors and restoring firing after a subchronic treatment, likely due to a 5-HT1A autoreceptor desensitization.12 Indeed, our results show that i.v. doses of CBD inhibit 5-HT firing activity, which were abolished by pretreatment with WAY and CPZ, indicating that the dose-dependent suppression of CBD on 5-HT firing involves both 5-HT1A and TRPV1 receptors. However, after repeated administration of CBD, 5-HT firing is increased compared with the veh-treated group, likely due to desensitization of 5-HT1A autoreceptors, as demonstrated by the attenuated response to LSD. Notably, CBD has been shown to act as an agonist with micromolar affinity at 5-HT1A receptors.75 These findings have been supported by behavioral studies indicating that CBD at the doses used herein suppresses conditioned gaping in rats, which is attenuated by intra-DRN microinfusions of WAY.72
The effectiveness of CPZ to antagonize the inhibitory effect of CBD on 5-HT firing is strongly supported by previous studies. Patch-clamp recordings in transfected HEK293 cells demonstrate that CBD dose dependently activates and rapidly desensitizes TRPV1 channels.48 In addition, CBD activates TRPV1 receptors in HEK293 cells with desensitization and maximal stimulation comparable with the prototypical TRPV1 agonist capsaicin.10 Recently, it has been shown that CBD (10 μM) enhances phagocytosis in cultured microglial cells and that this effect is abolished by TRPV1 antagonists CPZ and AMG 9810.45
Cannabidiol is known to act as an antagonist or inverse agonist at CB1 receptors with micromolar affinity.75 However, CB1 receptor blockade failed to prevent the inhibitory effect of CBD on 5-HT firing in our studies. Thus, actions of CBD at CB1 receptors are not necessary to produce antiallodynic and anxiolytic effects in a model of neuropathic pain.
Clinical76,85 and preclinical studies21–23,67,86 have highlighted the ability of CBD to treat pain conditions. For instance, oral administration of CBD (10, 20, and 40 mg/kg) abolished thermal hyperalgesia in an acute inflammatory pain model.21 Moreover, oral treatment with CBD (20 mg/kg) has antihyperalgesic effects in models of neuropathic and inflammatory pain in rats.23 Our results show that 7-day treatment with CBD significantly increased the withdrawal threshold to mechanical stimulation in neuropathic rats 23 days after SNI surgery, an effect blocked by TRPV1 antagonism. Similarly, Costa et al.22 demonstrated that the antihyperalgesic effect of acute oral administration of CBD (10 mg/kg) is prevented by the TRPV1 antagonist CPZ (10 mg/kg), but not by administration of the nonselective cannabinoid receptor antagonists SR141716 (0.5 mg/kg) or SR144528 (3 and 10 mg/kg), which further supports the hypothesis that the antiallodynic effect observed herein is likely mediated by TRPV1 receptors.
Clinical57,81,84 and preclinical40,41,49,62,74,90 studies have indicated strong comorbidity between chronic pain and anxiety. Furthermore, clinical studies have shown that CBD is effective at reducing anxiety.8,11 In preclinical studies, repeated administration of a high dose of CBD (30 mg/kg) prevents the anxiogenic effect of 14 days of CUS.18 Our data contribute to this growing body of literature by showing that low-dose CBD (5 mg/kg) ameliorates anxiety-like behavior induced by neuropathic pain in the OFT, EPMT, and NSFT through 5-HT1A but not through TRPV1 receptors. No differences were detected in the distance travelled in the OFT or in the latency to feed in the home cage in the NSFT, which confirms that these results are not due to general deficits in locomotor activity or pain-induced hypophagia. In addition, repeated CBD administration failed to alter immobility time in the FST in either sham or SNI rats. This may be due to the low dose used in this study (5 mg/kg) because previous work has shown that a minimum dose of 20 mg/kg is required to exert antidepressant-like effect.17
Studies show that the anxiolytic effect of CBD in naive animals is mediated by 5-HT1A receptors. For instance, CBD injections (30 nmol) into the dorsolateral periaqueductal gray have been shown to decrease anxiety in the EPMT and Vogel conflict test in a 5-HT1A (but not CB1) receptor–dependent manner.16 Moreover, rats receiving injections of CBD (60 nmol) into the bed nucleus of the stria terminalis show increased open-arm exploration in the EPMT, which is similarly blocked by WAY administration (0.37 nmol).38 In addition, pretreatment with WAY prevents the ability of systemic injections of CBD (5 mg/kg/day for 7 days) to prevent the anxiogenic effects of predator exposure in rats.14 These findings contrast with recent work demonstrating that pretreatment with WAY (0.05 mg/kg) does not reverse the anxiolytic properties of CBD (30 mg/kg) in a mouse model of CUS.33 However, this may be due to the different doses of both CBD and WAY used (higher doses of CBD and lower doses of WAY were used in these studies) as well as potential species differences (rats vs mice). In addition, the pathophysiological mechanisms underlying the development of anxiety in the SNI model are likely different from those in a chronic stress model.
A large body of literature supports the involvement of the DRN 5-HT system in nociception.54,55,70,80 Our electrophysiological recordings reveal that neuropathic pain provoked a maladaptation of 5-HT neurotransmission, causing a decrease in the firing activity of spontaneously active DRN 5-HT neurons. These results are in contrast with a previous study indicating that activity of 5-HT neurons was increased at 15 days after SNI.77 This discrepancy could be due to the fact that our in vivo recordings were performed 25 days after SNI surgery, during which time 5-HT neurotransmission may have undergone a shift in firing activity. On the contrary, in accordance with the findings of Sagheddu et al.,77 we found an increased number of neurons responsive to mechanical stimulation in SNI rats compared with sham-operated animals, without alterations in the number of 5-HT neurons recorded, suggesting that 5-HT neurons may be more sensitive to painful stimuli under neuropathological conditions.
A reduction and/or irregularity in the activity of 5-HT neurons has been linked to the emergence of mood and anxiety disorders.2,4 Previous electrophysiological data have shown that, after an initial depression in DRN 5-HT activity as a result of excess 5-HT in the synaptic cleft, long-term treatment with selective 5-HT reuptake inhibitors or 5-HT1A receptor agonists results in the restoration and augmentation of 5-HT firing activity due to the desensitization of 5-HT1A autoreceptors.13,31 Our data indicate that repeated administration of CBD produces a significant increase in the mean firing rate of DRN 5-HT neurons in both sham and SNI rats, which is consistent with results from studies testing more traditional anxiolytics and antidepressants.32 Cannabidiol also reduced the COV and the number of 5-HT neurons that were responsive to mechanical stimulation. Furthermore, the dose–response curve of the 5-HT1A receptor agonist LSD on the 5-HT firing rate was shifted to the right after 7-day CBD treatment, thus supporting the conclusion that a significant desensitization of 5-HT1A autoreceptors occurred as a result of repeated CBD treatment. This study confirms the activity of CBD on 5-HT neurons through 5-HT1A and TRPV1 receptors and rules out a direct involvement of CB1 receptors. Moreover, low-dose CBD treatment is able to (1) prevent mechanical allodynia predominantly through TRPV1 receptor activation (and partially through 5-HT1A receptor activation), (2) relieve pain-induced anxiety-like behavior through 5-HT1A receptor-mediated mechanisms, and (3) prevent aberrations in 5-HT neurotransmission in a SNI model. These results are clinically relevant, as CBD is known to exhibit few side effects14 and supports the initiation of clinical trials testing the efficacy of CBD-based compounds for treating neuropathic pain and comorbid mood disorders.
Conflict of interest statement
The authors declare that they have no competing interests.
Acknowledgements
Author contributions: D. De Gregorio performed electrophysiological and behavioral experiments, analysed data, and wrote the manuscript; R.J. McLaughlin performed electrophysiological experiments, analysed data, and wrote the manuscript; R. Ochoa-Sanchez performed electrophysiological experiments with acute dose–response of CBD; L. Posa performed behavioral experiments; J. Enns performed electrophysiological experiments in neuropathic rats; M. Lopez-Canul assisted in behavioral experiments; M. Aboud performed behavioral experiments with CBD and antagonists; S. Maione and S. Comai read manuscript and gave feedback; and G. Gobbi conceived the hypothesis and experiments, supervised experiments, interpreted results, and wrote the manuscript.
This research was supported by a matching grant (PSR-SIIRI) from the the Ministère de l'Économie Science et Innovation du Québec (MESI, Program PSR-SIIRI) and Aurora Cannabis Inc. D. De Gregorio and R.J. McLaughlin received post-doctoral fellowships from the Fonds de la recherché du Quebec en Santé (FRQS). L. Posa received a PhD fellowship from The Louise and Alan Edwards Foundation and M. Lopez-Canul received the Quebec Merit Scholarship for Foreign Students (PBEEE, FQRNT) for post-doctoral students.
Footnotes
Sponsorships or competing interests that may be relevant to content are disclosed at the end of this article.
D. De Gregorio and R.J. McLaughlin contributed equally to this work.
References
- [1].Allers K, Sharp T. Neurochemical and anatomical identification of fast- and slow-firing neurones in the rat dorsal raphe nucleus using juxtacellular labelling methods in vivo. Neuroscience 2003;122:193–204. [DOI] [PubMed] [Google Scholar]
- [2].Bambico FR, Cassano T, Dominguez-Lopez S, Katz N, Walker CD, Piomelli D, Gobbi G. Genetic deletion of fatty acid amide hydrolase alters emotional behavior and serotonergic transmission in the dorsal raphe, prefrontal cortex, and hippocampus. Neuropsychopharmacol 2010;35:2083–100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Bambico FR, Katz N, Debonnel G, Gobbi G. Cannabinoids elicit antidepressant-like behavior and activate serotonergic neurons through the medial prefrontal cortex. J Neurosci 2007;27:11700–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Bambico FR, Nguyen NT, Gobbi G. Decline in serotonergic firing activity and desensitization of 5-HT1A autoreceptors after chronic unpredictable stress. Eur Neuropsychopharmacol 2009;19:215–28. [DOI] [PubMed] [Google Scholar]
- [5].Bambico FR, Nguyen NT, Katz N, Gobbi G. Chronic exposure to cannabinoids during adolescence but not during adulthood impairs emotional behaviour and monoaminergic neurotransmission. Neurobiol Dis 2010;37:641–55. [DOI] [PubMed] [Google Scholar]
- [6].Baraban J, Aghajanian G. Noradrenergic innervation of serotonergic neurons in the dorsal raphe: demonstration by electron microscopic autoradiography. Brain Res 1981;204:1–11. [DOI] [PubMed] [Google Scholar]
- [7].Bardin L, Lavarenne J, Eschalier A. Serotonin receptor subtypes involved in the spinal antinociceptive effect of 5-HT in rats. PAIN 2000;86:11–18. [DOI] [PubMed] [Google Scholar]
- [8].Bergamaschi MM, Queiroz RHC, Chagas MHN, De Oliveira DCG, De Martinis BS, Kapczinski F, Quevedo J, Roesler R, Schröder N, Nardi AE. Cannabidiol reduces the anxiety induced by simulated public speaking in treatment-naive social phobia patients. Neuropsychopharmacology 2011;36:1219–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Berlin NI, Huff RL, Van Dyke DC, Hennessy TG. The blood volume of the adult rat, as determined by Fe59 and P32 labelled red cells. Proc Soc Exp Biol Med 1949;71:176–8. [DOI] [PubMed] [Google Scholar]
- [10].Bisogno T, Hanuš L, De Petrocellis L, Tchilibon S, Ponde DE, Brandi I, Moriello AS, Davis JB, Mechoulam R, Di Marzo V. Molecular targets for cannabidiol and its synthetic analogues: effect on vanilloid VR1 receptors and on the cellular uptake and enzymatic hydrolysis of anandamide. Br J Pharmacol 2001;134:845–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Blessing EM, Steenkamp MM, Manzanares J, Marmar CR. Cannabidiol as a potential treatment for anxiety disorders. Neurotherapeutics 2015;12:825–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Blier P, De Montigny C. Electrophysiological investigations on the effect of repeated zimelidine administration on serotonergic neurotransmission in the rat. J Neurosci 1983;3:1270–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Blier P, De Montigny C. Modification of 5-HT neuron properties by sustained administration of the 5-HT1A agonist gepirone: electrophysiological studies in the rat brain. Synapse 1987;1:470–80. [DOI] [PubMed] [Google Scholar]
- [14].Campos AC, Ferreira FR, Guimarães FS. Cannabidiol blocks long-lasting behavioral consequences of predator threat stress: possible involvement of 5HT1A receptors. J Psychiatr Res 2012;46:1501–10. [DOI] [PubMed] [Google Scholar]
- [15].Campos AC, Fogaça MV, Sonego AB, Guimarães FS. Cannabidiol, neuroprotection and neuropsychiatric disorders. Pharmacol Res 2016;112:119–27. [DOI] [PubMed] [Google Scholar]
- [16].Campos AC, Guimarães FS. Involvement of 5HT1A receptors in the anxiolytic-like effects of cannabidiol injected into the dorsolateral periaqueductal gray of rats. Psychopharmacology 2008;199:223. [DOI] [PubMed] [Google Scholar]
- [17].Campos AC, Moreira FA, Gomes FV, Del Bel EA, Guimaraes FS. Multiple mechanisms involved in the large-spectrum therapeutic potential of cannabidiol in psychiatric disorders. Philos Trans R Soc Lond B Biol Sci 2012;367:3364–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Campos AC, Ortega Z, Palazuelos J, Fogaca MV, Aguiar DC, Diaz-Alonso J, Ortega-Gutierrez S, Vazquez-Villa H, Moreira FA, Guzman M, Galve-Roperh I, Guimaraes FS. The anxiolytic effect of cannabidiol on chronically stressed mice depends on hippocampal neurogenesis: involvement of the endocannabinoid system. Int J Neuropsychopharmacol 2013;16:1407–19. [DOI] [PubMed] [Google Scholar]
- [19].Chen J, Song Y, Yang J, Zhang Y, Zhao P, Zhu XJ, Su HC. The contribution of TNF-α in the amygdala to anxiety in mice with persistent inflammatory pain. Neurosci Lett 2013;541:275–80. [DOI] [PubMed] [Google Scholar]
- [20].Comai S, Ochoa-Sanchez R, Dominguez-Lopez S, Bambico FR, Gobbi G. Melancholic-like behaviors and circadian neurobiological abnormalities in melatonin MT1 receptor knockout mice. Int J Neuropsychopharmacol 2015;18:pyu075. 10.1093/ijnp/pyu075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Costa B, Colleoni M, Conti S, Parolaro D, Franke C, Trovato AE, Giagnoni G. Oral anti-inflammatory activity of cannabidiol, a non-psychoactive constituent of cannabis, in acute carrageenan-induced inflammation in the rat paw. Naunyn Schmiedebergs Arch Pharmacol 2004;369:294–9. [DOI] [PubMed] [Google Scholar]
- [22].Costa B, Giagnoni G, Franke C, Trovato AE, Colleoni M. Vanilloid TRPV1 receptor mediates the antihyperalgesic effect of the nonpsychoactive cannabinoid, cannabidiol, in a rat model of acute inflammation. Br J Pharmacol 2004;143:247–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Costa B, Trovato AE, Comelli F, Giagnoni G, Colleoni M. The non-psychoactive cannabis constituent cannabidiol is an orally effective therapeutic agent in rat chronic inflammatory and neuropathic pain. Eur J Pharmacol 2007;556:75–83. [DOI] [PubMed] [Google Scholar]
- [24].Crippa JAS, Derenusson GN, Ferrari TB, Wichert-Ana L, Duran FL, Martin-Santos R, Simões MV, Bhattacharyya S, Fusar-Poli P, Atakan Z. Neural basis of anxiolytic effects of cannabidiol (CBD) in generalized social anxiety disorder: a preliminary report. J Psychopharmacol 2011;25:121–30. [DOI] [PubMed] [Google Scholar]
- [25].Dawson L, Nguyen H, Smith D, Schechter L. Effect of chronic fluoxetine and WAY-100635 treatment on serotonergic neurotransmission in the frontal cortex. J Psychopharmacol 2002;16:145–52. [DOI] [PubMed] [Google Scholar]
- [26].Decosterd I, Woolf CJ. Spared nerve injury: an animal model of persistent peripheral neuropathic pain. PAIN 2000;87:149–58. [DOI] [PubMed] [Google Scholar]
- [27].Deiana S, Watanabe A, Yamasaki Y, Amada N, Arthur M, Fleming S, Woodcock H, Dorward P, Pigliacampo B, Close S. Plasma and brain pharmacokinetic profile of cannabidiol (CBD), cannabidivarine (CBDV), Δ 9-tetrahydrocannabivarin (THCV) and cannabigerol (CBG) in rats and mice following oral and intraperitoneal administration and CBD action on obsessive–compulsive behaviour. Psychopharmacology 2012;219:859–73. [DOI] [PubMed] [Google Scholar]
- [28].Devinsky O, Marsh E, Friedman D, Thiele E, Laux L, Sullivan J, Miller I, Flamini R, Wilfong A, Filloux F. Cannabidiol in patients with treatment-resistant epilepsy: an open-label interventional trial. Lancet Neurol 2016;15:270–8. [DOI] [PubMed] [Google Scholar]
- [29].Dixon WJ. Efficient analysis of experimental observations. Annu Rev Pharmacol Toxicol 1980;20:441–62. [DOI] [PubMed] [Google Scholar]
- [30].Domínguez-López S, Mahar I, Bambico FR, Labonté B, Ochoa-Sánchez R, Leyton M, Gobbi G. Short-term effects of melatonin and pinealectomy on serotonergic neuronal activity across the light–dark cycle. J Psychopharmacol 2012;26:830–44. [DOI] [PubMed] [Google Scholar]
- [31].Dong J, De Montigny C, Blier P. Assessment of the serotonin reuptake blocking property of YM992: electrophysiological studies in the rat hippocampus and dorsal raphe. Synapse 1999;34:277–89. [DOI] [PubMed] [Google Scholar]
- [32].El Mansari M, Sánchez C, Chouvet G, Renaud B, Haddjeri N. Effects of acute and long-term administration of escitalopram and citalopram on serotonin neurotransmission: an in vivo electrophysiological study in rat brain. Neuropsychopharmacology 2005;30:1269–77. [DOI] [PubMed] [Google Scholar]
- [33].Fogaça MV, Campos AC, Coelho LD, Duman RS, Guimarães FS. The anxiolytic effects of cannabidiol in chronically stressed mice are mediated by the endocannabinoid system: role of neurogenesis and dendritic remodeling. Neuropharmacology 2018;135:22–33. [DOI] [PubMed] [Google Scholar]
- [34].Ford A, Castonguay A, Cottet M, Little JW, Chen Z, Symons-Liguori AM, Doyle T, Egan TM, Vanderah TW, De Konnick Y. Engagement of the GABA to KCC2 signaling pathway contributes to the analgesic effects of A3AR agonists in neuropathic pain. J Neurosci 2015;35:6057–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Garcia-Garcia AL, Newman-Tancredi A, Leonardo ED. P5-HT1A receptors in mood and anxiety: recent insights into autoreceptor versus heteroreceptor function. Psychopharmacology 2014;231:623–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Gauldie S, McQueen D, Pertwee R, Chessell I. Anandamide activates peripheral nociceptors in normal and arthritic rat knee joints. Br J Pharmacol 2001;132:617–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Gobbi G, Murphy DL, Lesch KP, Blier P. Modifications of the serotonergic system in mice lacking serotonin transporters: an in vivo electrophysiological study. J Pharmacol Exp Ther 2001;296:987–95. [PubMed] [Google Scholar]
- [38].Gomes FV, Resstel LB, Guimarães FS. The anxiolytic-like effects of cannabidiol injected into the bed nucleus of the stria terminalis are mediated by 5-HT1A receptors. Psychopharmacology 2011;213:465–73. [DOI] [PubMed] [Google Scholar]
- [39].Graeff FG, Guimarães FS, De Andrade TG, Deakin JF. Role of 5-HT in stress, anxiety, and depression. Pharmacol Biochem Behav 1996;54:129–41. [DOI] [PubMed] [Google Scholar]
- [40].Guida F, Luongo L, Marmo F, Romano R, Iannotta M, Napolitano F, Belardo C, Marabese I, D'Aniello A, De Gregorio D. Palmitoylethanolamide reduces pain-related behaviors and restores glutamatergic synapses homeostasis in the medial prefrontal cortex of neuropathic mice. Mol Brain 2015;8:47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Guo B, Wang J, Yao H, Ren K, Chen J, Yang J, Cai G, Liu H, Fan Y, Wang W. Chronic inflammatory pain impairs mGluR5-mediated depolarization-induced suppression of excitation in the anterior cingulate cortex. Cereb Cortex 2017;28:2118–30. [DOI] [PubMed] [Google Scholar]
- [42].Haddjeri N, Blier P. Effect of neurokinin-1 receptor antagonists on the function of 5-HT and noradrenaline neurons. Neuroreport 2000;11:1323–7. [DOI] [PubMed] [Google Scholar]
- [43].Haddjeri N, de Montigny C, Blier P. Modulation of the firing activity of rat serotonin and noradrenaline neurons by (±) pindolol. Biol Psychiatry 1999;45:1163–9. [DOI] [PubMed] [Google Scholar]
- [44].Haddjeri N, De Montigny C, Curet O, Blier P. Effect of the reversible monoamine oxidase-A inhibitor befloxatone on the rat 5-hydroxytryptamine neurotransmission. Eur J Pharmacol 1998;343:179–92. [DOI] [PubMed] [Google Scholar]
- [45].Hassan S, Eldeeb K, Millns PJ, Bennett AJ, Alexander SP, Kendall DA. Cannabidiol enhances microglial phagocytosis via transient receptor potential (TRP) channel activation. Br J Pharmacol 2014;171:2426–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Holmes A, Murphy DL, Crawley JN. Abnormal behavioral phenotypes of serotonin transporter knockout mice: parallels with human anxiety and depression. Biol Psychiatry 2003;54:953–9. [DOI] [PubMed] [Google Scholar]
- [47].Huestis M. Pharmacokinetics and metabolism of the plant cannabinoids, Δ 9-tetrahydrocannabinol, cannabidiol and cannabinol. Cannabinoids. New York, NY: Springer, 2005. p. 657–90. [DOI] [PubMed] [Google Scholar]
- [48].Iannotti FA, Hill CL, Leo A, Alhusaini A, Soubrane C, Mazzarella E, Russo E, Whalley BJ, Di Marzo V, Stephens GJ. Nonpsychotropic plant cannabinoids, cannabidivarin (CBDV) and cannabidiol (CBD), activate and desensitize transient receptor potential vanilloid 1 (TRPV1) channels in vitro: potential for the treatment of neuronal hyperexcitability. ACS Chem Neurosci 2014;5:1131–41. [DOI] [PubMed] [Google Scholar]
- [49].Ji G, Fu Y, Ruppert KA, Neugebauer V. Pain-related anxiety-like behavior requires CRF1 receptors in the amygdala. Mol Pain 2007;3:13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Kaplan JS, Stella N, Catterall WA, Westenbroek RE. Cannabidiol attenuates seizures and social deficits in a mouse model of Dravet syndrome. Proc Natl Acad Sci U S A 2017;114:11229–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Komaki A, Hashemi-Firouzi N, Shojaei S, Souri Z, Heidari S, Shahidi S. Study the effect of endocannabinoid system on rat behavior in elevated plus-maze. Basic Clin Neurosci 2015;6:147. [PMC free article] [PubMed] [Google Scholar]
- [52].Kwiatkowska M, Parker LA, Burton P, Mechoulam R. A comparative analysis of the potential of cannabinoids and ondansetron to suppress cisplatin-induced emesis in the Suncus murinus (house musk shrew). Psychopharmacology 2004;174:254–9. [DOI] [PubMed] [Google Scholar]
- [53].Lesch KP, Bengel D, Heils A, Sabol SZ, Greenberg BD, Petri S, Benjamin J, Müller CR, Hamer DH, Murphy DL. Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 1996;274:1527–31. [DOI] [PubMed] [Google Scholar]
- [54].Lopez-Alvarez VM, Puigdomenech M, Navarro X, Cobianchi S. Monoaminergic descending pathways contribute to modulation of neuropathic pain by increasing-intensity treadmill exercise after peripheral nerve injury. Exp Neurol 2018;299:42–55. [DOI] [PubMed] [Google Scholar]
- [55].Mason P. Contributions of the medullary raphe and ventromedial reticular region to pain modulation and other homeostatic functions. Annu Rev Neurosci 2001;24:737–77. [DOI] [PubMed] [Google Scholar]
- [56].McPartland J, Glass M, Pertwee R. Meta-analysis of cannabinoid ligand binding affinity and receptor distribution: interspecies differences. Br J Pharmacol 2007;152:583–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].McWilliams LA, Cox BJ, Enns MW. Mood and anxiety disorders associated with chronic pain: an examination in a nationally representative sample. PAIN 2003;106:127–33. [DOI] [PubMed] [Google Scholar]
- [58].McWilliams LA, Goodwin RD, Cox BJ. Depression and anxiety associated with three pain conditions: results from a nationally representative sample. PAIN 2004;111:77–83. [DOI] [PubMed] [Google Scholar]
- [59].Mechoulam R, Parker LA, Gallily R. Cannabidiol: an overview of some pharmacological aspects. J Clin Pharmacol 2002;42:11S–9S. [DOI] [PubMed] [Google Scholar]
- [60].Narita M, Kaneko C, Miyoshi K, Nagumo Y, Kuzumaki N, Nakajima M, Nanjo K, Matsuzawa K, Yamazaki M, Suzuki T. Chronic pain induces anxiety with concomitant changes in opioidergic function in the amygdala. Neuropsychopharmacology 2006;31:739–50. [DOI] [PubMed] [Google Scholar]
- [61].Nurmikko TJ, Serpell MG, Hoggart B, Toomey PJ, Morlion BJ, Haines D. Sativex successfully treats neuropathic pain characterised by allodynia: a randomised, double-blind, placebo-controlled clinical trial. PAIN 2007;133:210–20. [DOI] [PubMed] [Google Scholar]
- [62].Palazzo E, Luongo L, Guida F, Marabese I, Romano R, Iannotta M, Rossi F, D'Aniello A, Stella L, Marmo F. D-Aspartate drinking solution alleviates pain and cognitive impairment in neuropathic mice. Amino Acids 2016;48:1553–67. [DOI] [PubMed] [Google Scholar]
- [63].Parker LA, Burton P, Sorge RE, Yakiwchuk C, Mechoulam R. Effect of low doses of Δ 9-tetrahydrocannabinol and cannabidiol on the extinction of cocaine-induced and amphetamine-induced conditioned place preference learning in rats. Psychopharmacology 2004;175:360–6. [DOI] [PubMed] [Google Scholar]
- [64].Parker LA, Kwiatkowska M, Mechoulam R. Delta-9-tetrahydrocannabinol and cannabidiol, but not ondansetron, interfere with conditioned retching reactions elicited by a lithium-paired context in Suncus murinus: an animal model of anticipatory nausea and vomiting. Physiol Behav 2006;87:66–71. [DOI] [PubMed] [Google Scholar]
- [65].Parker LA, Mechoulam R, Schlievert C. Cannabidiol, a non-psychoactive component of cannabis and its synthetic dimethylheptyl homolog suppress nausea in an experimental model with rats. Neuroreport 2002;13:567–70. [DOI] [PubMed] [Google Scholar]
- [66].Paxinos GAWC, Watson C. The rat brain atlas in stereotaxic coordinates. San Diego: Academic, 1998. [Google Scholar]
- [67].Philpott HT, O'brien M, McDougall JJ. Attenuation of early phase inflammation by cannabidiol prevents pain and nerve damage in rat osteoarthritis. PAIN 2017;158:2442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Porsolt RD, Brossard G, Hautbois C, Roux S. Rodent models of depression: forced swimming and tail suspension behavioral despair tests in rats and mice. Curr Protoc Neurosci 2001;8:10A. [DOI] [PubMed] [Google Scholar]
- [69].Qin N, Neeper MP, Liu Y, Hutchinson TL, Lubin ML, Flores CM. TRPV2 is activated by cannabidiol and mediates CGRP release in cultured rat dorsal root ganglion neurons. J Neurosci 2008;28:6231–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [70].Qing-Ping W, Nakai Y. The dorsal raphe: an important nucleus in pain modulation. Brain Res Bull 1994;34:575–85. [DOI] [PubMed] [Google Scholar]
- [71].Ressler KJ, Nemeroff CB. Role of serotonergic and noradrenergic systems in the pathophysiology of depression and anxiety disorders. Depress Anxiety 2000;12:2–19. [DOI] [PubMed] [Google Scholar]
- [72].Rock E, Bolognini D, Limebeer C, Cascio M, Anavi-Goffer S, Fletcher P, Mechoulam R, Pertwee R, Parker L. Cannabidiol, a non-psychotropic component of cannabis, attenuates vomiting and nausea-like behaviour via indirect agonism of 5-HT1A somatodendritic autoreceptors in the dorsal raphe nucleus. Br J Pharmacol 2012;165:2620–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [73].Rock EM, Limebeer CL, Petrie GN, Williams LA, Mechoulam R, Parker LA. Effect of prior foot shock stress and Δ9-tetrahydrocannabinol, cannabidiolic acid, and cannabidiol on anxiety-like responding in the light-dark emergence test in rats. Psychopharmacology 2017;234:2207–17. [DOI] [PubMed] [Google Scholar]
- [74].Roeska K, Doods H, Arndt K, Treede RD, Ceci A. Anxiety-like behaviour in rats with mononeuropathy is reduced by the analgesic drugs morphine and gabapentin. PAIN 2008;139:349–57. [DOI] [PubMed] [Google Scholar]
- [75].Russo EB, Burnett A, Hall B, Parker KK. Agonistic properties of cannabidiol at 5-HT1a receptors. Neurochem Res 2005;30:1037–43. [DOI] [PubMed] [Google Scholar]
- [76].Ryz NR, Remillard DJ, Russo EB. Cannabis roots: a traditional therapy with future potential for treating inflammation and pain. Cannabis Cannabinoid Res 2017;2:210–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [77].Sagheddu C, Aroni S, De Felice M, Lecca S, Luchicchi A, Melis M, Muntoni AL, Romano R, Palazzo E, Guida F. Enhanced serotonin and mesolimbic dopamine transmissions in a rat model of neuropathic pain. Neuropharmacology 2015;97:383–93. [DOI] [PubMed] [Google Scholar]
- [78].Sales AJ, Fogaça MV, Sartim AG, Pereira VS, Wegener G, Guimarães FS, Joca SR. Cannabidiol induces rapid and sustained antidepressant-like effects through increased BDNF signaling and synaptogenesis in the prefrontal cortex. Mol Neurobiol 2018. 10.1007/s12035-018-1143-4. [Epub ahead of print]. [DOI] [PubMed] [Google Scholar]
- [79].Samara E, Bialer M, Mechoulam R. Pharmacokinetics of cannabidiol in dogs. Drug Metab disposition 1988;16:469–72. [PubMed] [Google Scholar]
- [80].Schweimer J, Ungless M. Phasic responses in dorsal raphe serotonin neurons to noxious stimuli. Neuroscience 2010;171:1209–15. [DOI] [PubMed] [Google Scholar]
- [81].Scott KM, Bruffaerts R, Tsang A, Ormel J, Alonso J, Angermeyer M, Benjet C, Bromet E, De Girolamo G, De Graaf R. Depression–anxiety relationships with chronic physical conditions: results from the World Mental Health Surveys. J Affect Dis 2007;103:113–20. [DOI] [PubMed] [Google Scholar]
- [82].Scuderi C, Filippis DD, Iuvone T, Blasio A, Steardo A, Esposito G. Cannabidiol in medicine: a review of its therapeutic potential in CNS disorders. Phytother Res 2009;23:597–602. [DOI] [PubMed] [Google Scholar]
- [83].Tal M, Bennett GJ. Neuropathic pain sensations are differentially sensitive to dextrorphan. Neuroreport 1994;5:1438–40. [DOI] [PubMed] [Google Scholar]
- [84].Tsang A, Von Korff M, Lee S, Alonso J, Karam E, Angermeyer MC, Borges GLG, Bromet EJ, De Girolamo G, De Graaf R. Common chronic pain conditions in developed and developing countries: gender and age differences and comorbidity with depression-anxiety disorders. J Pain 2008;9:883–91. [DOI] [PubMed] [Google Scholar]
- [85].Wade DT, Robson P, House H, Makela P, Aram J. A preliminary controlled study to determine whether whole-plant cannabis extracts can improve intractable neurogenic symptoms. Clin Rehabil 2003;17:21–9. [DOI] [PubMed] [Google Scholar]
- [86].Ward SJ, McAllister SD, Kawamura R, Murase R, Neelakantan H, Walker EA. Cannabidiol inhibits paclitaxel-induced neuropathic pain through 5-HT1A receptors without diminishing nervous system function or chemotherapy efficacy. Br J Pharmacol 2014;171:636–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [87].Ward SJ, Ramirez MD, Neelakantan H, Walker EA. Cannabidiol prevents the development of cold and mechanical allodynia in paclitaxel-treated female C57Bl6 mice. Anesth Analg 2011;113:947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [88].Whiting PF, Wolff RF, Deshpande S, Di Nisio M, Duffy S, Hernandez AV, Keurentjes JC, Lang S, Misso K, Ryder S. Cannabinoids for medical use: a systematic review and meta-analysis. JAMA 2015;313:2456–73. [DOI] [PubMed] [Google Scholar]
- [89].Wolfe F, Russell I, Vipraio G, Ross K, Anderson J. Serotonin levels, pain threshold, and fibromyalgia symptoms in the general population. J Rheumatol 1997;24:555–9. [PubMed] [Google Scholar]
- [90].Wu Y, Yao X, Jiang Y, He X, Shao X, Du J, Shen Z, He Q, Fang J. Pain aversion and anxiety-like behavior occur at different times during the course of chronic inflammatory pain in rats. J Pain Res 2017;10:2585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [91].Zanelati TV, Biojone C, Moreira FA, Guimaraes FS, Joca SR. Antidepressant-like effects of cannabidiol in mice: possible involvement of 5-HT1A receptors. Br J Pharmacol 2010;159:122–8. [DOI] [PMC free article] [PubMed] [Google Scholar]