

Abstract
Purpose:
Tumor androgens in castration resistant prostate cancer (CRPC) reflect de novo intra-tumoral synthesis or adrenal androgens. We used C.B.−17 SCID mice in which we observed adrenal CYP17A activity to isolate the impact of adrenal steroids on CRPC tumors in vivo.
Methods:
We evaluated tumor growth and androgens in LuCaP35CR and LuCaP96CR xenografts in response to adrenalectomy. We assessed protein expression of key steroidogenic enzymes in 185 CRPC metastases from 42 patients.
Results:
Adrenal glands of intact and castrated mice expressed CYP17A. Serum DHEA, AED and T in castrated mice became undetectable after adrenalectomy (all p<0.05). Adrenalectomy prolonged median survival (days) in both CRPC models (33 vs 179; 25 vs 301) and suppressed tumor steroids vs castration alone (T 0.64 vs 0.03pg/mg; DHT 2.3 vs 0.23pg/mg; and T 0.81 vs 0.03pg/mg, DHT 1.3 vs 0.04pg/mg; all p=<0.001). A subset of tumors recurred with increased steroid levels, and/or induction of AR, truncated AR variants, and GR. Metastases from 19 of 35 patients with AR positive tumors concurrently expressed enzymes for adrenal androgen utilization and 9 expressed enzymes for de novo steroidogenesis (HSD3B1, CYP17A, AKR1C3 and HSD17B3).
Conclusions:
Mice are appropriate for evaluating adrenal impact of steroidogenesis inhibitors. A subset of adrenalectomy-resistant CRPC tumors demonstrate de novo androgen synthesis. Tumor growth and androgens were suppressed more strongly by surgical adrenalectomy than prior studies using abiraterone, suggesting reduction in adrenally-derived androgens beyond that achieved by abiraterone may have clinical benefit. Proof-of-concept studies with agents capable of achieving true ‘non-surgical ADX’ are warranted.
Keywords: adrenal gland, CYP17A, intratumoral androgens, prostate cancer, castration resistant, xenograft, androgen receptor, splice variant, rodent adrenal androgens, steroidogenesis
INTRODUCTION
Androgen deprivation therapy (ADT) remains front-line treatment for patients with locally recurrent or metastatic prostate cancer (PCa), but patients uniformly progress to castration resistant prostate cancer (CRPC). Residual intratumoral androgen are believed to play a critical role in maintaining ligand-dependent mechanisms of androgen receptor (AR) activation.(1) The source of residual tissue androgens in castrated patients is believed to reflect the tumoral uptake and intracellular conversion of the adrenal androgens such as DHEA-sulfate (DHEAS) to testosterone (T) and dihydrotestosterone (DHT), and/or the de novo intratumoral synthesis of androgens from cholesterol or progesterone precursors.(2, 3) CYP17A, expressed in the human adrenal gland, testis and ovary, is a single enzyme with one active site that catalyzes sequential but independent hydroxylase and lyase reactions both of which are required for converting pregnenolone or progesterone to the adrenal androgens DHEA or androstenedione (AED), respectively. In particular, circulating levels of DHEA-S (the primary circulating form of DHEA) in eugonadal men are extremely high and are not reduced by standard castration (CX) therapy.(4–6)
The contribution of adrenal steroids to CRPC progression is directly supported by historical case reports documenting clinical responses to adrenalectomy (ADX) in men with CRPC,(7) and indirectly supported in the modern era by the efficacy of the CYP17A inhibitor abiraterone (ABI) in decreasing circulating adrenal androgens and improving overall survival for men with metastatic PCa in the androgen sensitive and CRPC settings.(8, 9) A direct impact of adrenal CYP17A inhibition on suppressing prostate tissue androgens can be inferred from the decrease in prostate androgen levels achieved by addition of ABI to standard ADT in the neoadjuvant setting, wherein tumoral CYP17A expression and de novo intratumoral androgen synthesis has not yet been upregulated.(4, 10, 11) However, the anti-tumor activity of ABI in the setting of CRPC may also occur at the level of the tumor via inhibition of intratumoral CYP17A activity or via the direct anti-AR activity of ABI.(2, 3, 12) Thus, the anti-tumor activity of ABI cannot necessarily be ascribed to inhibition of adrenal androgen synthesis alone, and the specific contribution of adrenally-derived steroids to intratumoral androgens and CRPC progression remains unproven.
The role of adrenal steroids in xenograft models of CRPC grown in castrated mice has not been previously investigated due to early literature suggesting a lack of functional CYP17A in rodent adrenal glands.(13–17) Accordingly, the source of tumor androgens detected in patient derived xenograft (PDX) models of CRPC grown in castrated male mice has been attributed solely to de novo intratumoral androgen synthesis.(18) However, both historical studies using radioimmunoassays (RIA) and more recent studies using mass spectrometry (MS) have suggested functional adrenal CYP17A activity in rats and mice, including residual prostate androgens in castrated rats that were eliminated by ADX.(19–23) While the physiologic contribution of these steroid levels to normal prostatic biology was considered negligible,(22, 23) this does not rule out a potential pathologic role in stimulating prostate tumor cells which are hypersensitive to androgen stimulation. We sought to experimentally demonstrate the contribution of adrenal steroids to intratumoral androgens and tumor progression in PDX models of CRPC grown in castrated male C.B-17 SCID mice in which we demonstrate adrenal CYP17A expression and activity.
MATERIALS AND METHODS
Generation of Murine Samples from Intact, Castrated and Adrenalectomized Mice
All experiments involving animals were performed in accordance with protocols approved by the Fred Hutchinson Center Institutional Animal Care Use Committee (file 1775) and recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. All surgery was carried out under isofluorane anesthesia. All studies utilized male C.B-17 SCID mice (Taconic, Rensselaer, NY). Serum samples were collected by retro-orbital bleed from non-tumor bearing 18–20 week old intact (eugonadal) male mice, an age-matched cohort of mice at 12 weeks after CX (CX performed at 6–8 weeks), and an age-matched cohort at 12 weeks after both CX and ADX. Serum was obtained from a separate cohort of intact and castrated male mice for independent measurement of serum steroids in the laboratory of Dr. Penning. Adrenal glands, kidney, liver and quadriceps muscle were resected for tissue steroid measurements Drinking water was replaced with normal saline supplemented with corticosterone (Sigma, St. Louis, MO; 25 μg/mL in 0.2% ethanol-0.5% NaCl) at time of ADX. (24) Adrenal glands for assessment of CYP17A methylation and transcript profiling were resected from intact and castrated male mice ranging in age from 12–36 weeks (3–8 months) to mirror the ages spanning the course of a typical xenograft study.
LuCaP Human Prostate Cancer PDX models and CRPC Tissues
All studies utilizing human tissue sources were conducted in accordance with recognized ethical guidelines (Belmont Report and U.S. Common Rule). The LuCaP35CR and LuCaP96CR lines are CX resistant (CR) PCa PDX models expressing wild type AR established as part of the University of Washington Medical Center (UWMC) Genitourinary Biorepository as previously described. (21) Cell line authentication is regularly performed via STR profiling. Castrated male C.B-17 SCID mice aged 8–10 weeks were subcutaneously implanted in bilateral flanks with 30mm3 tumor pieces. Tumor volume was measured three times per week and calculated as (length x (width2)/ 2. When tumors reached 200–250mm3, drinking water was replaced with normal saline supplemented with corticosterone as described above, and mice were randomized to no further treatment (CX alone) or to undergo ADX (CX plus ADX). Tumors from n=4 mice in each PDX model were harvested at 21–30 days following ADX. Mice were monitored until tumors reached approximately 1000mm3 (end of study, EOS) or animals became compromised, at which point animals were euthanized according to institutional protocol and tumors harvested for flash freezing and formalin fixation. Due to their prolonged time to tumor regrowth, a subset of mice in the ADX plus CX arms were euthanized and the data censored prior to reaching the size endpoint due to concerns for animal health.
In the LuCaP96CR study, n=8 mice were bilaterally implanted to yield approximately 9 evaluable tumors per arm for long term follow-up (~60% take rate). Based on 25% variation in re-growth rates, 9 tumors per arm is predicted to provide 80% power with two-tailed alpha of 0.05 to detect a 35% difference in mean time to regrowth (defined as 750 mm3). Due to the unexpectedly prolonged time to regrowth following CX plus ADX in this model (with interval attrition of mice due to health reasons) animal number in the combined treatment arm of the LuCaP35CR study was increased to n=12.
Metastatic CRPC tissues for creation of the CRPC tissue microarrays (TMA) were obtained as part of the UWMC Prostate Cancer Rapid Autopsy Program, and comprised metastases from 42 patients (including 65 soft tissue metastases and 120 bone metastases, with up to 4 metastatic sites from each patient) as previously described.(25, 26) The UWMC Institutional Review Board approved all procedures involving human subjects, and all subjects signed written informed consent.
Pyrosequencing, Western Blotting and Quantitative RT-PCR
Detailed methods for assessment of adrenal CYP17A methylation status by pyrosequencing, adrenal CYP17A protein expression by western blotting, and quantitative RT-PCR in murine adrenal glands and xenograft tissues are given in the Supplementary Methods and Supplementary Table 1.
Steroid Measurements
Detailed methods for determination of steroids in serum and tissue samples by mass spectrometry are given in the Supplementary Methods. The lower limits of quantitation (LLOQ) for steroids in serum were 0.01 ng/ml for pregnenolone, progesterone, AED, T and androsterone; 0.02 ng/ml for DHEA and cortisol, 0.04 ng/ml for DHT and corticosterone, and 0.002 ng/ml for deoxycorticosterone. The LLOQ for steroids in tissue was 0.02 pg/mg for progesterone, AED, DHT, and T; 0.04 pg/mg for DHEA; and 0.08 pg/mg for pregnenolone. Methods for determination of steroids in serum by the Penning Laboratory are as previously published.(27)
Immunohistochemistry and Image Analysis
Details for creation of the PDX TMA are given in the Supplementary Methods. The CRPC TMA was created as previously published.(26) Detailed methods for immunostaining and specific antibody clones are given in the Supplementary Methods and Supplementary Table 2. Quantitative image analysis of PDX TMA slides stained for AR and ARV7 was performed using metrics we have previously described and further detailed in the Supplementary Methods.(28) Scoring for all other stains carried out in a blinded fashion by an experienced pathologist (L.T. or M.T.). A quasi-continuous immunohistochemical (IHC) score was calculated by multiplying each intensity level (0 for no stain, 1 for faint stain, and 2 for intense stain) by the corresponding percentage of cells (0–100%) at the corresponding intensity and totaling the results. For all stains data from the two TMA spots per sample was averaged.
Statistical Analyses.
The non-parametric Mann Whitney Rank test was used for all two sample comparisons. P values < 0.05 were considered significant and those < 0.10 as trending toward significance. Progression free survival in CX vs. CX plus ADX treated mice (defined as tumor size <750mm3) was determined via Kaplan Meier analysis with comparison of curves using the Mantel-Haenszel logrank test. Statistical analyses were carried out using GraphPad Prism Software (LaJolla, CA).
RESULTS
Impact of Adrenalectomy on Circulating Steroid Levels in Castrated Mice
To determine the contribution of adrenal androgens to circulating serum levels in castrated mice we evaluated steroid levels in serum from intact (INT) male C.B-17SCID mice, and from age-matched mice 12 weeks after CX or after CX plus ADX (Fig 1). As expected, median levels of pregnenolone and progesterone in serum were similar in castrated vs intact mice (0.52 and 0.48 ng/ml pregnenolone, and 1.9 vs 3.2 ng/ml progesterone, p=not significant (ns) for both) and were significantly decreased following ADX (to 0.12 ng/ml, p=0.002 pregnenolone, and to below detectable limits (bdl) for progesterone, p<0.001; Fig 1A-B), consistent with the adrenal origin of these steroids. Notably, median levels of DHEA in castrate mice were also similar to those in intact mice (0.03 vs 0.04 ng/ml, p=ns) and decreased to the limit of detection following ADX (to 0.02 ng/ml p=0.001, Fig 1C). Following CX median levels of AED and T observed in intact mice were decreased but detectable (from 0.03 to 0.01ng/ml, p=0.08 AED; from 1.9 to 0.02 ng/ml, p<0.001 T), only becoming undetectable following ADX (p=0.05 and p=0.004, respectively; Fig 1D-E), while levels of DHT in intact mice (0.15 ng/ml) became undetectable following CX alone (p=0.05, Fig 1F). Serum levels of DHEA, AED, T and DHT from a separate cohort of intact and 6 week castrated male C.B-17 mice, were independently measured in the laboratory of Dr. Penning, and were consistent with our measurements, with the higher castrate T levels in the latter likely reflecting the shorter duration of castration (Supp Table 3).
Figure 1. Serum steroid levels and adrenal CYP17A expression in male C.B-17 SCID mice.
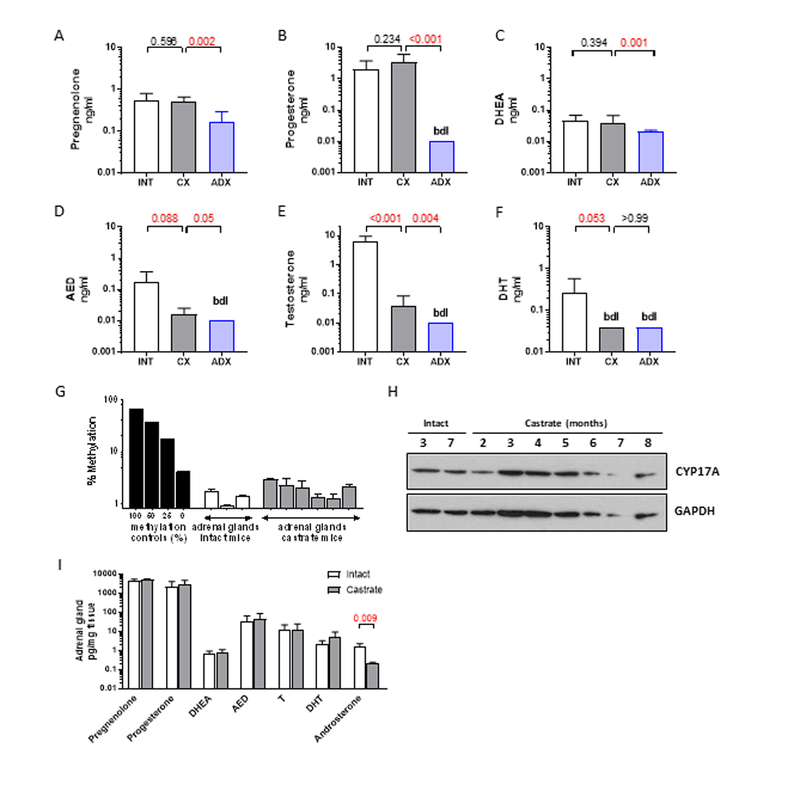
A-G) Levels of the indicated steroids were measured by mass spectrometry in eugonadal male mice (intact, white bars, n=8) and age-matched mice at 12 weeks after castration alone (CX, gray bars, n=7) or castration plus adrenalectomy (ADX, blue bars, n=10). G) Methylation status of CYP17A in adrenal glands resected from intact (age 3 months) and castrated male mice (age 1–3 months), compared to a methylation control ranging from 100% methylated to un-methylated (0%). H) Protein levels of CYP17A by western blot in adrenal glands resected from intact and castrated male mice at the indicated age (in months). I) Levels of the indicated steroids in adrenal glands resected from (intact, white bars) and age-matched mice at 12 weeks after castration alone (CX, gray bars). Data in A-F and I are shown as mean and standard deviation and represent a minimum of 6 animals per group. P values calculated using the Mann-Whitney rank test between the indicated groups (P values <0.05 and <0.10 were considered as significant and trending towards significance, respectively).
Consistent with literature documenting corticosterone as the primary circulating mineralocorticoid in mice,(13, 14) circulating levels of cortisol were undetectable (not shown), while levels of 11-deoxycorticosterone were similar in intact and castrated mice (2.6 ng/dl vs 4.8 ng/ml, p=ns) and became significantly decreased following ADX (0.005 ng/dl, p<0.001, Supp Fig 1A). Due to the requirement for mineralocorticoid replacement (administered as corticosterone in drinking water), levels of corticosterone in the adrenalectomized mice were similar to those in the intact and castrated mice, Supp Fig 1B).
Expression of CYP17A and Steroid Levels in Adrenal Glands of Castrated Mice
Previously published data in rats and in the mouse adrenocortical Y1 cell line suggested epigenetic regulation of CYP17A expression in adrenal cells via methylation, possibly via activity of cAMP responsive element modulator (CREM) isoforms. (29–31) However, we show that CpG islands in the CYP17A promoter DNA were unmethylated (Fig 1G) and protein expression was detectable by western blot (Fig 1H) in adrenal glands of intact and castrated mice ranging in age from 12–36 months (3–8 months; to mirror the ages spanning the course of a typical xenograft study). Transcripts of steroidogenic genes required for de novo and adrenal androgen synthesis (STAR, CYP11A, CYP17A, HSD3B1) and of SRD5A (required for production of DHT) were detectable in adrenal glands of intact mice and were significantly increased after castration (Supp Fig 1C), as observed previously in some rodent studies.(19, 32, 33) Notably, adrenal expression of melanocortin 2 receptor (MC2R; also known as adrenocorticotropic hormone receptor; ACTH) was also increased following castration, and is consistent with response to ACTH as a driver of adrenal steroidogenesis in the castrate setting (sufficient serum for ACTH measurements was not available).
To confirm the functional significance of adrenal steroidogenic enzyme expression we evaluated steroid levels in adrenal glands resected from eugonadal mice and age-matched castrated mice (12 weeks after CX; Fig 1I). As expected, median levels of pregnenolone (4238 pg/mg) and progesterone (1876 mg/pg) were very high in adrenal glands of intact mice and remained unchanged by CX. Notably, median levels of AED (24.6 pg/mg) and T (10 pg/mg) in adrenal glands from intact mice were substantial, while levels of DHEA (0.7 pg/mg), DHT (1.7 pg/mg) and androsterone (1.4 pg/ml) were easily measurable. Similar to pregnenolone and progesterone, adrenal levels of AED, T and DHT were also unchanged by CX, suggesting that testicular steroids do not serve as a source of these androgens detected within the adrenal gland. In contrast, median levels of androsterone in the adrenal gland were substantially decreased after CX, suggesting a testicular contribution to this steroid level (p=0.095).
As serum androgen levels in castrated mice may be influenced by steroid production in other organs with steroidogenic potential, we assessed steroid levels in liver, kidney and muscle (Sup Fig 2).(34, 35) Notably, levels of pregnenolone, progesterone, AED, T, and DHT in the adrenal gland of castrated mice were 2–3 orders of magnitude higher than levels in liver, kidney and muscle (Supp Fig 2A-F), suggesting active production specific to the adrenal gland. Moreover, while CX did not affect steroid levels in the adrenal gland (other than androsterone), it significantly decreased levels of AED in liver (p=0.10; Supp Fig 2D), of T in liver, kidney and muscle (p=0.013; p=0.05; p=0.028; Supp Fig 2E), of DHT in kidney (p=0.028; Supp Fig 2F), and of androsterone in liver (p=0.039; Supp Fig 2G). These observations demonstrate the testicular contribution to steroid levels detected in liver, kidney and muscle, and support the hypothesis that circulating steroid levels observed in castrated mice are primarily derived from the adrenal gland rather than other organs.
Impact of Adrenalectomy on Tumor Growth and Steroid Levels in Castration Resistant PDX Models
To evaluate the functional relevance of adrenally-derived steroids in CRPC tumor models in vivo, we determined the impact of ADX on suppressing tumor growth and intra-tumoral androgens in the LuCaP96CR and LuCaP35CR PDX models in castrated C.B-17 SCID mice. In the LuCaP96CR PDX model (Fig 2A), tumor regrowth was markedly delayed by CX plus ADX (CX + ADX, blue curves) compared to CX alone (CX, black curves), with an increase in median survival (to tumor endpoint of 750mm3) from 25 days (in the CX group) to 300 days (in the CX + ADX grou; (p=0.0002; Fig 2B). In the LuCaP35CR model (Fig 2C), tumor growth was markedly delayed by CX + ADX in a subset of tumors (Slow, blue curves), while a subset (Rapid, red curves) regrew with kinetics similar to the CX only group (black curves). Median survival in the CX only vs the entire CX + ADX cohort was delayed from 33 to 179 days, (p=0.005; Fig 2D), with the growth prolongation primarily contributed by the Slow subset of tumors (median survival not reached vs 66 days in the Rapid group, p=0.008; Fig 2E).
Figure 2. Impact of adrenalectomy on growth of CRPC tumor models in vivo.
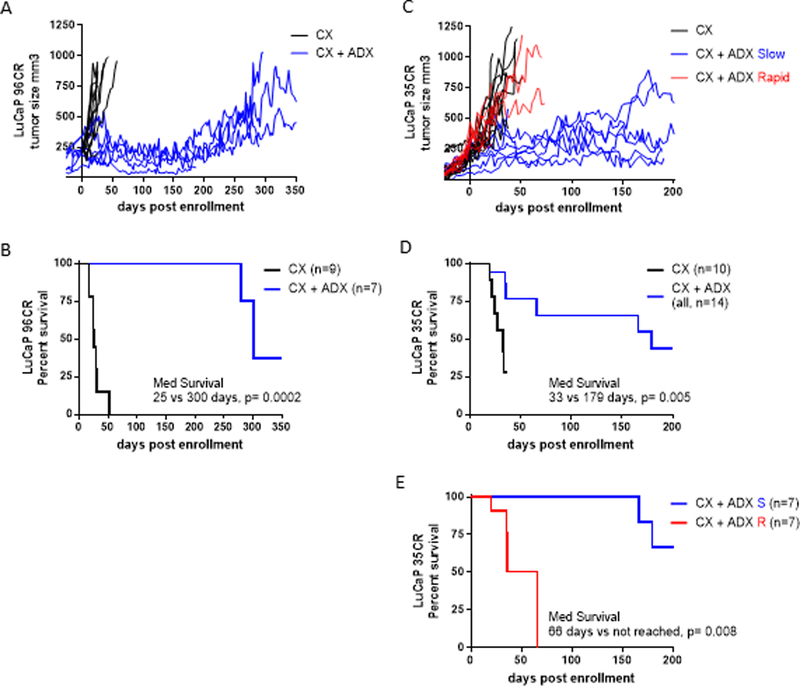
A) Individual tumor volume curves for the LuCaP96CR patient derived xenograft (PDX) model in mice treated with castration alone (CX, black curves) or castration plus adrenalectomy (CX + ADX, blue curves). B) Kaplan Meier analysis of progression free survival in CX vs. CX + ADX treated LuCaP96CR tumors (defined as tumor size <750mm3) with comparison of curves using the Mantel-Haenszel logrank test. C) Individual tumor growth curves for the LuCaP35CR patient derived xenograft (PDX) model in mice treated with CX (black curves) or CX + ADX, among which a subset of tumors regrew slowly (Slow, blue curves), while a subset (Rapid, red curves) regrew with kinetics similar to the CX only group (black curves). D) Kaplan Meier analysis of progression free survival in CX vs. CX + ADX treated LuCaP35CR tumors. E) Kaplan Meier analysis of progression free survival in LuCaP35CR tumors treated with CX + ADX, comparing median survival in the tumor subsets with Slow (blue) vs Rapid (red) re-growth.
In LuCaP96CR median intratumor levels of pregnenolone, progesterone, T and DHT were decreased by an order of magnitude at 30 days after ADX + CX (ADX d30) compared to CX alone (pregnenolone 24.3 vs 2.7 pg/mg, p=0.008 (not shown); progesterone 1.4 vs 0.03 pg/mg, p=0.001; T 0.9 vs 0.03 pg/mg, p=0.001; DHT 1.2 vs 0.03 pg/mg, p=0.001; Fig 3A). Tumor levels of DHEA and AED were lower at ADX d30 vs CX alone, although the decreases did not reach statistical significance (p=ns for both). While levels of DHEA, AED, T and DHT remained suppressed near the limit of detection in the end of study (EOS) tumors that regrew after CX + ADX (ADX EOS), progesterone levels in these EOS tumors were increased compared to tumors resected at day 30 after ADX (0.03 vs 0.125 pg/mg, p=0.0317), although still lower than tumors in the CX only group.
Figure 3. Impact of adrenal inhibition on tumor steroid levels in CRPC tumor models in vivo.
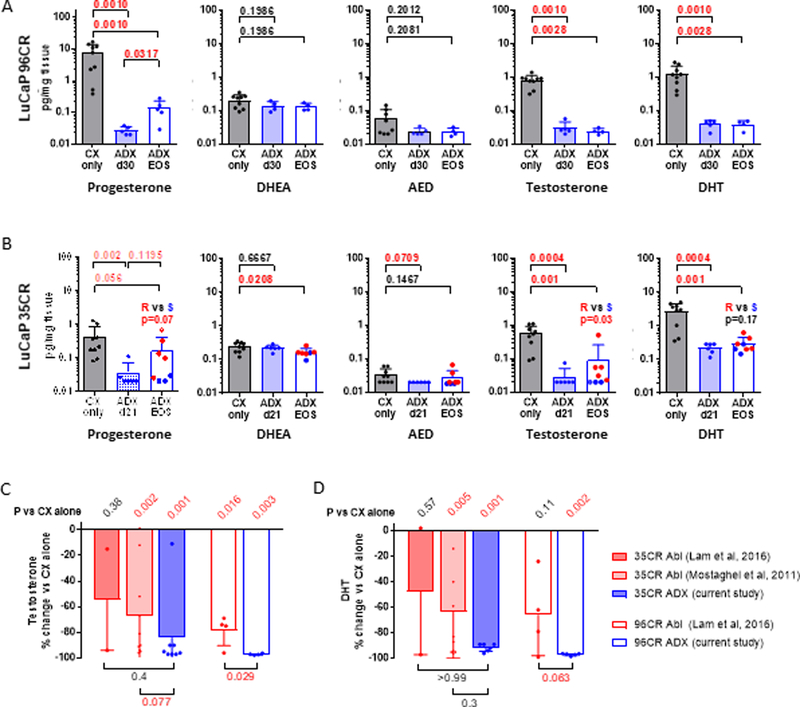
Levels of the indicated steroids were measured by mass spectrometry in tumors resected from the A) LuCaP96CR and B) LuCaP35CR PDX studies shown in Figure 2, in mice treated with castration alone (CX, gray bars) or castration plus adrenalectomy (ADX, blue bars), at short time points (shaded blue bars) or at end of study (EOS, open blue bars). For LuCaP35CR the subset of EOS tumors that regrew slowly (S, blue dots) vs rapidly (R, red dots) are indicated. Data are shown as mean and standard deviation. P values calculated using the Mann-Whitney rank test between the indicated groups (P values <0.05 and <0.10 were considered as significant and trending towards significance, respectively). The impact of ABI (red bars, prior studies) vs ADX (blue bars, current study) on the percent change in C) Testosterone and D) DHT compared to castration alone is shown for LuCaP35CR (closed bars) and LuCaP96CR (open bars). P values for the difference in % change between ABI and ADX treated tumors are given under the horizontal bars at the bottom of each graph. P values for the difference in tumor steroids in each treatment group vs Cx alone are given in diagonal font at the top of each graph.
In LuCaP35CR median intratumor levels of progesterone, T and DHT were also decreased by an order of magnitude by 21 days after ADX (ADX d21) compared to CX alone (progesterone 0.21 vs 0.02 pg/mg, p=0.002; T 0.6 vs 0.02 pg/mg, p=0.0004; DHT 3.2 vs 0.25 pg/mg, p=0.0004; Fig 3B), while decreases in AED trended toward significance (0.03 vs 0.02 pg/mg, p=0.07, respectively). Steroid levels in tumors that regrew after CX + ADX (ADX EOS) were significantly below those in the CX alone group for progesterone, DHEA, T and DHT (p=0.056, p=0.02, p=0.001, and p=0.001, respectively); however, median levels in tumors that regrew rapidly (red dots in Fig 3B) trended toward being higher than levels in tumors that regrew slowly (blue dots) for progesterone (p=0.07), T (p=0.03), and DHT (p=0.17), suggesting the more rapid regrowth in these tumors was in part stimulated by the elevation in tumor androgens. In both models serum androgens levels following ADX were largely below detectable limits (not shown), precluding assessment of correlation with tumor androgens
Notably, surgical ADX showed a significantly more profound impact on suppressing tumor androgens and CRPC tumor growth than did previously published studies from our group using ABI, in which LuCaP96CR was classified as an intermediate responder, and LuCaP35CR showed no response in one study and moderate response in another.(18, 36) Compared to these studies, ADX more consistently suppressed tumor levels of pregnenolone, progesterone, DHEA and AED (blue bars, Supp Fig 3A-D) than did ABI (red bars), whereas in at least in one model (LuCaP35CR) ABI was associated with increases in pregnenolone and progesterone (immediately upstream of the first enzymatic function of CYP17A inhibited by ABI), and DHEA (immediately upstream of HSD3B1, which is also inhibited by ABI).(37) Similarly, ABI clearly suppressed EOS levels of T and DHT (red bars, Fig 3C-D); however, ADX (blue bars) appeared to do so more consistently.
Induction of AR, ARV7 and GR in Adrenalectomy-Resistant CRPC
We determined the impact of ADX on key mechanisms of resistance identified in treatment refractory CRPC including expression of AR, the prevalent AR splice variant – ARV7, GR, and steroidogenic enzymes, as potential mechanisms driving tumor re-growth under the stringent sup
ression of ligand levels observed in the setting of CX + ADX.(3, 18, 36)
In LuCaP96CR we observed no change in transcript levels of AR, but a significant induction of ARV7 (4.9 fold, p=0.005), decrease in PSA (−2.6 fold, p=0.009), and increase in expression of GR (3.6 fold, p=0.028) in the EOS tumors (Table 1). IHC staining of a TMA created from this PDX model (Fig 4A) similarly showed no change in AR following ADX, an increase in nuclear ARV7 staining (p=0.08 at EOS, p=0.05 for EOS vs ADX d30), and a decrease in cytoplasmic PSA (p=0.016 at ADX d30, p=0.035 at EOS). However, an increase in nuclear GR staining was not observed, which instead showed an apparent decrease (p=0.025 at ADX d30). These data suggest induction of ARV7 as a significant mechanism of resistance in this PDX model, potentially accounting for the observation that transcript levels of some androgen regulated genes (e.g. TMPRSS2 and FKBP5) were not necessarily decreased in the EOS tumors.
Table 1.
Change in transcript levels of AR, AR regulated genes, GR, and steroidogenic enzymes in CRPC tumors after castration vs castration plus adrenalectomy.
| Gene Transcript | LuCaP96CR | LuCaP35CR | ||
|---|---|---|---|---|
| Fold | P value* | Fold | P value* | |
| AR | 1.6 | ns | 3.5 | <0.001 |
| V7 | 4.9 | 0.005 | 3.6 | 0.004 |
| GR (NR3C1) | 3.6 | 0.028 | 1.6 | 0.017 |
| PSA | −2.6 | 0.009 | −1.0 | ns |
| TMPRSS2 | −1.3 | ns | −1.4 | ns |
| FKBP5 | 2.9 | 0.068 | 1.3 | ns |
| CYP17A1 | −1.2 | ns | −2.1 | ns |
| CYB5A | 2.1 | 0.018 | 3.8 | <0.001 |
| NR5A1 | 2.1 | 0.026 | 3.1 | 0.001 |
| HSD3B1 | 1.0 | ns | −1.1 | ns |
| AKR1C3 | 1.3 | ns | 8.6 | <0.001 |
| HSD17B3 | −2.1 | ns | 1.9 | 0.06 |
| SRD5A1 | 3.1 | 0.018 | 1.8 | <0.001 |
| UGT2B15 | 4.9 | 0.014 | 1.4 | ns |
| UGT2B17 | −1.4 | 0.010 | deleted | |
P values from non-parametric Mann Whitney test comparing end of study tumors treated with castration alone vs castration plus adrenalectomy.
Figure 4. Impact of adrenalectomy on immunohistochemical (IHC) staining for AR, ARV7, PSA and GR in CRPC tumor models in vivo.
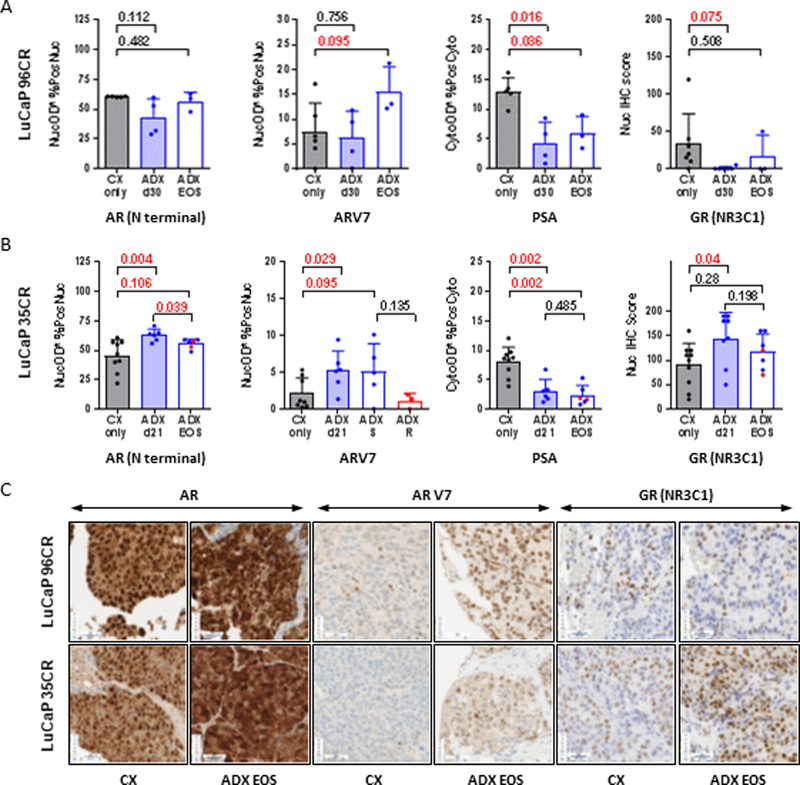
IHC staining for the indicated protein in tumors resected from the A) LuCaP96CR and B) LuCaP35CR PDX studies shown in Figure 3 in mice treated with castration alone (CX, gray bars) or castration plus adrenalectomy (ADX, blue bars), at short time points (shaded blue bars) or at end of study (EOS, open blue bars). For LuCaP35CR the subset of EOS tumors that regrew slowly (S, blue dots) vs rapidly (R, red dots) are indicated. Staining for AR, ARV7 and PSA quantified as the average nuclear (AR, ARV7) or cytoplasmic (PSA) staining intensity within tumor epithelium multiplied by the percentage of positive nuclei (AR, ARV7) or cells with positive cytoplasm (PSA) in tumor epithelium (denoted as AvgNuclearOD*%PosNuclei or AvgCytoOD*%PosCyto). Semi-quantitative scoring for nuclear GR expression calculated by multiplying the intensity level (0 for no stain, 1 for faint stain, and 2 for intense stain) by the percentage of cells (0–100%) at each intensity level and totaling the results, ranging from 0 (no staining in any cell) to 200 (intense staining in 100% of the cells). Data are shown as mean and standard deviation. P values calculated using the Mann-Whitney rank test between the indicated groups (P values <0.05 and <0.10 were considered as significant and trending towards significance, respectively). C) Representative examples of the indicated stains for LuCaP96CR (top) and LuCaP35CR (bottom) PDX models, showing each stain in an EOS tumor from a CX only or CX plus ADX treatment arm.
In LuCaP35CR we observed a significant induction of AR (3.5 fold, p=<0.001), ARV7 (3.6 fold, p=0.004) and GR message (1.6 fold, p=0.017) in the EOS tumors (Table 1). There was no change in the androgen-regulated genes PSA, TMPRSS2 or FKBP5 at the transcript level in this PDX model, although IHC staining (Fig 4B) did show a decrease in cytoplasmic PSA (at ADX d21 and at EOS, p=0.0016 for both). Consistent with the transcript results, we observed an increase in nuclear AR, ARV7 and GR staining in this model (Fig 4B) that was moderately more significant at ADX d21 (p=0.004, p=0.029and p=0.025, respectively) than in the EOS tumors (p=0.106, p=0.095 and p=0.15, respectively). We did not observe significant differences in AR, PSA, or GR staining in the EOS tumors that re-grew rapidly (red dots, Fig 4B) vs those that regrew slowly (blue dots). However, the increase in ARV7 seen at ADX d21 i appeared to be sustained in the EOS tumors that recurred with low androgen levels (blue dots, here and in EOS T and DHT levels, Fig 3B), but not in the EOS tumors that recurred with an increase in tumor androgens (red dots, here and in EOS T and DHT levels, Fig3B). These data suggest induction of AR, ARV7 and GR as potentially significant mechanism of resistance in this model, with ARV7 playing a more important role in tumors that regrew without an increase in intratumoral androgen levels, consistent with the known inverse relationship between androgen levels and ARV7 expression.(38) Of note, corticosterone supplementation itself had no impact on tumor growth (Supp Fig 4A) or IHC expression of GR (Supp Fig4B) or of AR and ARV7 (not shown). This suggests the induction of GR expression in the ADX-treated tumors in this model reflects the impact of ADX and not the replacement corticosterone dosing.
Induction of Steroidogenic Enzyme Expression in Adrenalectomy Resistant CRPC
Consistent with the increased intratumoral levels of T and DHT observed in a subset of the ADX recurrent LuCaP35CR tumors (red dots, Fig 3B) we observed increased expression of several genes involved in mediating steroidogenic enzyme activity (Table 1; see Supp Fig5 for schema of androgen biosynthesis enzymes), including CYB5A (enhances activity of CYP17A and HSD3B, 3.8 fold p<0.001), NR5A1 (transcriptional regulator of multiple steroidogenic genes, 3.1 fold p=0.001), AKR1C3 (8.3 fold p<0.001), HSD17B3 (1.9 fold p=0.06), and SRD5A1 (1.8 fold p<0.001). (39, 40) In contrast, while ADX EOS tumors in the LuCaP96 model also showed increases in expression of CYB5A1 and NR5A1 (2 fold for both, p=0.018 and 0.026, respectively) and SRD5A1 (3.1 fold p=0.018), they did not show induction of the steroid producing genes AKR1C3 or HS17B3 but rather induction of UGT2B15 (4.9 fold p=0.014) which conjugates T and DHT for excretion, potentially explaining the differences in androgen levels observed in the two models.
Expression of Steroidogenic Enzyme in CRPC Metastases
To further assess the translational relevance of the steroidogenic enzyme transcripts observed in the ADX-treated tumors, we evaluated AR axis proteins (AR, PSA) and key enzymes required for de novo steroid synthesis and/or conversion of adrenal androgens (HSD3B1, CYP17A1, AKR1C3 and HSD17B3; Supp Fig 5) in a TMA of bone and soft tissue CRPC metastases from 43 patients (~2–6 metastases per patient). Despite some degree of intra-patient heterogeneity, grouping of tumors by patient of origin revealed several distinct staining profiles, including subsets of patients with i. AR- and PSA- tumors with low expression of steroidogenic enzymes (Fig 5A), ii. AR+ tumors with negative (Fig5B top) or positive (Fig 5B bottom) PSA expression and low steroidogenic potential (based on generally low expression of HSD3B1, which is required for both de novo synthesis and conversion of adrenal androgen), and iii. AR+ and PSA+ tumors with high steroidogenic potential (Fig5C, based on moderate to high expression of HSD3B1). Tumors in the latter group can be further sub-divided based on potential for adrenal androgen conversion (e.g. coordinate expression of HSD3B1, HSD17B3/AKR1C3 (which catalyze the same reaction) but low CYP17A1, Fig 5C top), or potential for de novo steroidogenesis (e.g. coordinate expression HSD3B1, HSD17B3/AKR1C3 and high CYP17A, Fig 5C bottom). Representative images of tumors showing coordinate expression of HSD3B1, HSD17B3/AKR1C3 and CYP17A are shown in Supp Fig6. As we have previously published, a subset of these tumors show loss of C terminal AR staining, consistent with the presence of truncated AR variant species.(41)
Figure 5. Expression of steroidogenic enzymes in CRPC metastases.
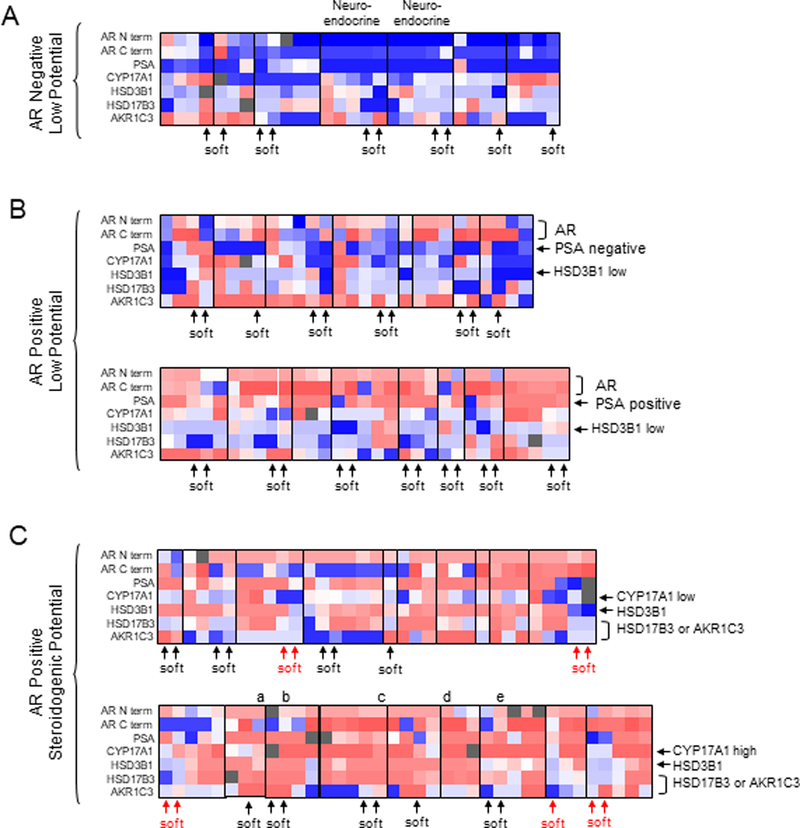
The heatmap summarizes staining observed on a TMA containing multiple bone and soft tissue CRPC metastases from 43 men collected via rapid autopsy. Each column represents serial staining of the same metastasis for the indicated proteins, ranging from negative expression (dark blue) to high expression (dark red). Vertical lines demarcate the set of metastases from each patient. Tumors are grouped based on negative (A) or positive (B, C) AR expression (in the majority of tumors in an individual patient), and then by steroidogenic potential. A) AR negative tumors with negative PSA staining and low expression of steroidogenic enzymes. Tumors from the two patients indicated had neuroendocrine histology. B) AR positive tumors with negative (top) or positive (bottom) PSA expression and low steroidogenic potential (based on low HSD3B1 expression). C) AR positive tumors with high steroidogenic potential. Tumors are grouped based on potential for adrenal androgen conversion (coordinate expression of HSD3B1and HSD17B3/AKR1C3 with low CYP17A1, top), or potential for de novo steroidogenesis (coordinate expression of CYP17A1, HSD3B1 and HSD17B3/AKR1C3, bottom). Among the group with high steroidogenic potential, patients in whom the soft tissue metastases appear distinctly different than the bone metastases are indicated in red. Representative IHC stains of the metastases labeled a-e in panel C are shown in Supplementary Figure 6.
DISCUSSION
Clinical studies of ABI in men with metastatic PCa are consistent with an important role for CYP17A-mediated adrenal androgen production in CRPC progression, particularly as higher pretreatment levels of circulating adrenal androgens associate with better response to CYP17A inhibition.(42–44) However, inhibition of intratumoral CYP17A activity is not excluded, and ABI and its metabolites can directly target the AR.(12) Thus these studies do not actually isolate the specific contribution of adrenal steroids in CRPC progression. It has been widely held that rodent adrenal glands do not produce adrenal androgens and that rodent models cannot, therefore, be used to test this facet of PCa biology. We now show that adrenal CYP17A is present in C.B-17 SCID mice, that steroids downstream of CYP17A are detectable in the adrenal glands and serum of castrated mice, and that surgical ADX markedly suppresses serum and tumor steroid levels and delays tumor re-growth beyond that observed with CX alone in two PDX models of CRPC. This is the first study to demonstrate the specific impact of adrenal steroids on intratumoral androgens and CRPC tumor growth in vivo.
Functional CYP17A is transiently present in rodent adrenal glands during the fetal/early post-natal period (45, 46) and then decreases, likely via epigenetic methylation and repression.(29) This phenomenon is not universal however, as we show that adrenal CYP17A DNA is unmethylated and detectable in C.B-17 SCID mice, consistent with the spiny mouse (Acomys cahirinus) that is known to express functional CYP17A.(46) Gonadectomy induces adrenal gland expression of steroidogenic genes and increases circulating levels of AED and T in certain inbred mouse strains.(33) We did not observe differences in CYP17A methylation status or in transcript and protein levels between intact and castrated mice, although we did observe statistically significant increases in transcript expression of STAR, CYP11A, HSD3B1 and SRD5A2 at 12 weeks following CX.
Importantly, we show levels of adrenal androgens (DHEA, AED) and downstream metabolites (T, DHT and androsterone) are easily detectable in adrenal glands of intact mice and do not decrease following CX. The adrenal levels of DHEA (0.7 pg/mg, range 0.44 – 1.2 pg/mg) and AED (44 pg/mg, range 4 – 97 pg/mg) in our study are consistent with the findings of Hu et al who reported levels of DHEA (~4–7 pg/mg) and AED (~2–4 pg/mg) in the adrenal glands of rats (Wistar, via GC/MS) 8 weeks after CX, equivalent to levels in intact rats.(23) Moreover, we show adrenal levels of AED, T, and DHT in castrated mice to be nearly 2 orders of magnitude higher than in kidney, liver or muscle (other organs shown to have steroidogenic capacity in mice)(34, 35), suggesting active production specific to the adrenal gland.
An existing body of data, published primarily in the endocrine literature and not well-recognized in the PCa field, supports functional adrenal CYP17A activity in adult rodents and may reflect strain specific differences. While some rodent adrenal suspension studies failed to detect production of 17-hydroxyprogesterone or AED (products of CYP17A hydroxylase and lyase activity, respectively) via RIA,(13) others did measure AED or DHEA (via RIA(46, 47)) or showed metabolism of [3H]DHEA or [3H]AED to T and DHT in vitro.(48, 49). Similarly, serum levels of T, DHT and AED were undetectable in some strains of castrated rats (Holtzmann, via RIA(50); Wistar, via LC/MS(51)), but detectable in others (Sprague–Dawley, via LC/MS(19)(52)). Kyprianou and Isaacs detected circulating (T 0.10 ng/ml and DHT 0.16 ng/ml) and prostatic levels of T and DHT (via RIA) in castrated Copenhagen rats that were eliminated by ADX,(22) while a study in Wistar rats showed a decrease in prostate levels of T and DHT after castration but no sustained decrease in AED or DHEA (via GC/MS), consistent with an adrenal origin.(23) In context of the present study, the relevance of this literature becomes more apparent and provides significant support for our findings.
Strain specific differences in adrenal androgen production are suggested by the observation that certain inbred strains of mice (DBA/2J, CE/J, C3H, NU/J, BALB/c and B6D2F1) are highly sensitive to gonadectomy induced sex-steroid producing adrenocortical neoplasia, while others (C57BL/6 and FVB/N) are not.(53) The mouse strains associated with adrenal androgen production in the literature are consistent with those susceptible to adrenal neoplasia, with androgens observed in BALB/c mice (via RIA(13)) but not in C57BL/6 mice (via LC/MS(54). In this respect it is notable that the C.B-17 SCID mice used in our study are derived from the BALB/C strain.
Notably, surgical ADX showed a significantly more profound impact on suppressing tumor androgens and CRPC tumor growth than did previously published studies from our group using ABI.(18, 36) While ABI clearly suppressed EOS levels of T and DHT in the prior studies (red bars, Fig 3C-D), ADX (blue bars) appeared to do so more consistently. Differences in growth suppression might also reflect the AR agonist activity attributed to a five-alpha reduced metabolite of ABI (55), an effect that would be absent in the ADX treated tumors.
Given the clear impact of both ADX and ABI on tumor androgen levels we sought to determine whether mechanisms of resistance to ADX mirrored those observed with ABI. Consistent with our previous findings in LuCaP96CR tumors treated with ABI,(36) we observed increased transcript levels of ARV7 and GR, and no increase in transcript or nuclear staining for full length AR in the ADX-treated tumors (although we did not observe increased nuclear expression of GR as noted in that study). In LuCaP35CR tumors we observed increased transcript expression and nuclear staining of AR and ARV7 and an increase in nuclear GR staining after ADX, consistent with prior data (although increased AR expression was only observed in one of the prior studies likely due to PDX heterogeneity).(18, 36) These data suggest induction of ARV7 as a potentially significant mechanism of resistance following ADX in LuCaP96CR, and induction of AR, ARV7 and GR as potentially significant mechanisms of resistance in LuCaP35CR. While AR and ARV7 share an overlapping transcriptome, transcripts of AR target genes are not consistently increased in the ADX-treated tumors, consistent with prior observations thatARV7 is not necessarily as potent as full length AR in inducing the expression of canonical AR genes.(56, 57)
Our data suggests that the adrenal gland contributes substantially to levels of T and DHT in CRPC tumors grown in castrated mice, and is consistent with our data demonstrating expression of enzymes required for adrenal androgen utilization in a majority of AR+ CRPC metastases examined (Fig 5C). However, a subset of ADX-resistant LuCaP35CR tumors demonstrate induction of steroidogenic enzymes and moderate reconstitution of intratumoral progesterone, T and DHT (similar to our prior data in ABI-recurrent LuCaP35CR tumors.(18) This suggests that at least some tumor models are also capable of de novo steroid production and is consistent with the concurrent expression of enzymes required for de novo steroidogenesis in a subset of the CRPC tumors we evaluated (Fig 5C bottom). As increased progesterone levels were present in a subset of the ADX EOS tumors vs earlier time points in both PDX models we determined whether these EOS tumors showed induction of the progesterone receptor (PR) as possible mechanism of progression.(58) However, staining for PR was low to undetectable in the CX only tumors, without any difference in the ADX-treated tumors (data not shown). A direct effect of progesterone on stimulation of growth via wild-type (WT) AR cannot necessarily be excluded, as Kumagai et al did not detect downstream conversion of progesterone to T or DHT in VCaP cells (which has WT AR), yet still observed modest activation of growth.(59) Activation of WT AR by a metabolite directly downstream of progesterone (20beta-hydroxy-5alpha-dihydroprogesterone) has also been recently demonstrated.(60) We did not measure levels of this particular steroid in our assay, but as its production from progesterone is not mediated by CYP17A it suggests another reason why surgical ADX appears to have better anti-tumor efficacy than CYP17A inhibition with ABI.
We and others have previously demonstrated transcript expression of steroidogenic enzymes in subsets of CRPC metastases, but data on concordant protein expression of key enzymes required for androgen synthesis from adrenal androgens or earlier precursors are limited.(2, 3, 61–66) Here, we demonstrate clear subsets of tumors with coordinate expression of the critical enzymes required for de novo steroid synthesis and/or conversion of adrenal androgens. Steroidogenic enzyme expression is generally low in AR negative tumors (Fig 5A), and in AR positive tumors with low PSA expression (Fig5B, top), indicative of poor response to agents targeting the AR or ligand synthesis. Low expression of steroidogenic enzymes is also observed in a subset of AR and PSA positive tumors (Fig5B, bottom), suggesting AR axis signaling in response to exogenous ligand is maintained, but without intratumoral androgen production. In contrast, tumors from nearly half the patients (Fig 5C) appear potentially capable of androgen production from adrenal androgens (based on concordant expression of HSD3B1 with AKR1C3 or HSD17B3,), and of these, half also show concordant expression of CYP17A (Fig 5C, bottom), suggesting capacity for de novo steroidogenesis.
Although we did not observe consistent differences in steroidogenic capacity between bone and soft tissue tumors overall, there was more pronounced staining in bone vs soft tissue metastases in several patients (red arrows in Fig 5C), potentially consistent with the observation that osteoblasts may induce steroidogenic enzymes in CRPC cells.(67) Unexpectedly, some tumors with loss of C terminal AR staining (consistent with the presence of ligand-independent AR variants) also demonstrated steroidogenic enzyme expression, suggesting that in certain cases the response to castration may include both induction of AR variants and upregulated steroid production. Alternatively, ARV7 is a strong inducer of the glucuronidating enzyme UGT2B17 which conjugates active androgens for excretion, and could therefore continue to promote AR variant production in the presence of concomitant steroidogenesis.(41)
In summary, we show that adrenally-derived steroids are produced in C.B-17 SCID mice and contribute to tumor androgen levels and growth in two PDX models of CRPC, demonstrating that C.B-17 SCID mice are an appropriate model for evaluating the impact of steroidogenesis inhibitors in CRPC xenograft studies. Although the potential impact/availability of low circulating T levels may be magnified in mice compared to humans due to the lack of circulating sex hormone binding globulin (SHBG) in mice,(68) castrate levels of DHEA, AED, T and DHT in this study were similar to those in ABI-treated CRPC patients (as previously reported for T(68)). This suggests a reduction in adrenally-derived androgens beyond those achieved by ABI may have clinical benefit in men with CRPC, and is consistent with our observation that tissue androgens and tumor growth were suppressed more strongly by surgical ADX in this study than in previously reported studies using ABI. This suggests that proof-of-concept studies testing agents capable of achieving true ‘non-surgical ADX’ are warranted (such as the recently reported CYP11A inhibitor ODM-208),(69) and that optimal clinical efficacy may be obtained by a combination of CYP11A and CYP17A inhibition. While beyond the scope of the current work, PDX studies testing CYP11A inhibition are planned. We find that mechanisms of resistance after surgical ADX in CRPC models are similar to those observed following ABI, including the induction of AR, truncated AR splice variants, GR, and steroidogenesis. This is consistent with the fact that both approaches act via suppression of tissue androgen levels, and suggests optimal clinical activity will be achieved by combination strategies co-targeting AR or GR with agents capable of fully abrogating adrenal and intratumoral ligand synthesis.
Supplementary Material
STATEMENT OF TRANSLATIONAL RELEVANCE (150).
Response to the adrenal CYP17A inhibitor abiraterone in men with castration resistant prostate cancer (CRPC) does not isolate the contribution of adrenally-derived steroids, as inhibition of intratumoral CYP17A and/or direct anti-AR effects cannot be excluded. We show C.B-17 SCID mice have adrenal CYP17A and are appropriate for testing inhibitors of adrenal steroidogenesis. Adrenalectomy suppressed tumor growth and androgens in two xenograft models of CRPC, and did so more strongly than prior studies using abiraterone. This suggests that reduction in adrenally-derived androgens beyond that achieved by abiraterone may have clinical benefit, and that proof of concept studies with agents that block ligand synthesis upstream of CYP17A, such as novel CYP11A inhibitors, are warranted. A subset of adrenalectomy-resistant tumors demonstrate de novo androgen synthesis and/or induction of AR, truncated AR variants, and GR, suggesting co-targeting AR or GR with abrogation of ligand synthesis will still be required for optimal clinical activity.
ACKNOWLEDGEMENTS
The authors wish to acknowledge reagents and technical assistance in pyrosequencing provided by Dr. William Grady, and expert technical assistance with image analysis provided by Jonathan Henriksen and NWBioSpecimen (a core service for procurement and annotation of research biospecimens) which is supported by National Cancer Institute grant P30 CA015704 (G. Gilliland, principal investigator [PI]), Institute of Translational Health Sciences grant UL1 TR000423 (M. Disis, PI), the University of Washington School of Medicine and Department of Pathology, and Fred Hutchinson Cancer Research Center.
Support: NIH Pacific Northwest Prostate Cancer SPORE P50 CA97186 (EAM, EC, LDT, PSN), NIH P01 CA163227 (EAM, TMP, SPB, EC, LDT, PSN), DOD W81XWH-12–1-0208 (EAM), NIH P30 CA015704 (JH), NIH UL1 TR000423 (JH), Department of Veterans Affairs Puget Sound Health Care System (EAM, AMM)
Footnotes
The authors declare no financial conflicts of interest.
REFERENCES
- 1.Scher HI. Current management of hormone-refractory prostate cancer. Clin Adv Hematol Oncol 2004;2(11):724–6. [PubMed] [Google Scholar]
- 2.Stanbrough M, Bubley GJ, Ross K, Golub TR, Rubin MA, Penning TM, et al. Increased expression of genes converting adrenal androgens to testosterone in androgen-independent prostate cancer. Cancer Res 2006;66(5):2815–25. [DOI] [PubMed] [Google Scholar]
- 3.Montgomery RB, Mostaghel EA, Vessella R, Hess DL, Kalhorn TF, Higano CS, et al. Maintenance of intratumoral androgens in metastatic prostate cancer: a mechanism for castration-resistant tumor growth. Cancer Res 2008;68(11):4447–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Taplin ME, Montgomery B, Logothetis CJ, Bubley GJ, Richie JP, Dalkin BL, et al. Intense androgen-deprivation therapy with abiraterone acetate plus leuprolide acetate in patients with localized high-risk prostate cancer: results of a randomized phase II neoadjuvant study. J Clin Oncol 2014;32(33):3705–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Arai S, Miyashiro Y, Shibata Y, Tomaru Y, Kobayashi M, Honma S, et al. Effect of castration monotherapy on the levels of adrenal androgens in cancerous prostatic tissues. Steroids. 2011;76(3):301–8. [DOI] [PubMed] [Google Scholar]
- 6.Tamae D, Mostaghel E, Montgomery B, Nelson PS, Balk SP, Kantoff PW, et al. The DHEA-sulfate depot following P450c17 inhibition supports the case for AKR1C3 inhibition in high risk localized and advanced castration resistant prostate cancer. Chemico-biological interactions. 2015;234:332–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bhanalaph T, Varkarakis MJ, Murphy GP. Current status of bilateral adrenalectomy or advanced prostatic carcinoma. Annals of surgery. 1974;179(1):17–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Fizazi K, Tran N, Fein L, Matsubara N, Rodriguez-Antolin A, Alekseev BY, et al. Abiraterone plus Prednisone in Metastatic, Castration-Sensitive Prostate Cancer. N Engl J Med 2017;377(4):352–60. [DOI] [PubMed] [Google Scholar]
- 9.Mostaghel EA. Abiraterone in the treatment of metastatic castration-resistant prostate cancer. Cancer Manag Res 2014;6:39–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cho E, Mostaghel EA, Russell KJ, Liao JJ, Konodi MA, Kurland BF, et al. External beam radiation therapy and abiraterone in men with localized prostate cancer: safety and effect on tissue androgens. Int J Radiat Oncol Biol Phys 2015;92(2):236–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mostaghel EA, Cho E, Zhang A, Alyamani M, Kaipainen A, Green S, et al. Association of Tissue Abiraterone Levels and SLCO Genotype with Intraprostatic Steroids and Pathologic Response in Men with High-Risk Localized Prostate Cancer. Clin Cancer Res 2017;23(16):4592–601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Li Z, Bishop AC, Alyamani M, Garcia JA, Dreicer R, Bunch D, et al. Conversion of abiraterone to D4A drives anti-tumour activity in prostate cancer. Nature. 2015;523(7560):347–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.van Weerden WM, Bierings HG, van Steenbrugge GJ, de Jong FH, Schroder FH. Adrenal glands of mouse and rat do not synthesize androgens. Life Sci 1992;50(12):857–61. [DOI] [PubMed] [Google Scholar]
- 14.Belanger B, Belanger A, Labrie F, Dupont A, Cusan L, Monfette G. Comparison of residual C-19 steroids in plasma and prostatic tissue of human, rat and guinea pig after castration: unique importance of extratesticular androgens in men. J Steroid Biochem 1989;32(5):695–8. [DOI] [PubMed] [Google Scholar]
- 15.Perkins LM, Payne AH. Quantification of P450scc, P450(17) alpha, and iron sulfur protein reductase in Leydig cells and adrenals of inbred strains of mice. Endocrinology. 1988;123(6):2675–82. [DOI] [PubMed] [Google Scholar]
- 16.Le Goascogne C, Sananes N, Gouezou M, Takemori S, Kominami S, Baulieu EE, et al. Immunoreactive cytochrome P-450(17 alpha) in rat and guinea-pig gonads, adrenal glands and brain. J Reprod Fertil 1991;93(2):609–22. [DOI] [PubMed] [Google Scholar]
- 17.Pelletier G, Li S, Luu-The V, Tremblay Y, Belanger A, Labrie F. Immunoelectron microscopic localization of three key steroidogenic enzymes (cytochrome P450(scc), 3 beta-hydroxysteroid dehydrogenase and cytochrome P450(c17)) in rat adrenal cortex and gonads. The Journal of endocrinology. 2001;171(2):373–83. [DOI] [PubMed] [Google Scholar]
- 18.Mostaghel EA, Marck BT, Plymate SR, Vessella RL, Balk S, Matsumoto AM, et al. Resistance to CYP17A1 inhibition with abiraterone in castration-resistant prostate cancer: induction of steroidogenesis and androgen receptor splice variants. Clin Cancer Res 2011;17(18):5913–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Chen J, Liang Q, Hua H, Wang Y, Luo G, Hu M, et al. Simultaneous determination of 15 steroids in rat blood via gas chromatography-mass spectrometry to evaluate the impact of emasculation on adrenal. Talanta. 2009;80(2):826–32. [DOI] [PubMed] [Google Scholar]
- 20.Ando S, Canonaco M, Beraldi E, Valenti A, Maggiolini M, Piro A, et al. The evaluation of androgen circulating levels following castration in adult male rats. Experimental and clinical endocrinology. 1988;91(3):311–8. [DOI] [PubMed] [Google Scholar]
- 21.Nguyen HM, Vessella RL, Morrissey C, Brown LG, Coleman IM, Higano CS, et al. LuCaP Prostate Cancer Patient-Derived Xenografts Reflect the Molecular Heterogeneity of Advanced Disease an--d Serve as Models for Evaluating Cancer Therapeutics. The Prostate. 2017;77(6):654–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kyprianou N, Isaacs JT. Biological significance of measurable androgen levels in the rat ventral prostate following castration. The Prostate. 1987;10(4):313–24. [DOI] [PubMed] [Google Scholar]
- 23.Hu M, Xin D, Chen J, Sun G, Wang Y, Na Y. Changes in the androgen levels in the ventral prostate of spontaneously hypertensive rats after castration. BJU Int. 2009;104(3):406–11. [DOI] [PubMed] [Google Scholar]
- 24.Mongrain V, Hernandez SA, Pradervand S, Dorsaz S, Curie T, Hagiwara G, et al. Separating the contribution of glucocorticoids and wakefulness to the molecular and electrophysiological correlates of sleep homeostasis. Sleep. 2010;33(9):1147–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Roudier MP, True LD, Higano CS, Vesselle H, Ellis W, Lange P, et al. Phenotypic heterogeneity of end-stage prostate carcinoma metastatic to bone. Hum Pathol 2003;34(7):646–53. [DOI] [PubMed] [Google Scholar]
- 26.Roudier MP, Winters BR, Coleman I, Lam HM, Zhang X, Coleman R, et al. Characterizing the molecular features of ERG-positive tumors in primary and castration resistant prostate cancer. The Prostate. 2016;76(9):810–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Tamae D, Byrns M, Marck B, Mostaghel EA, Nelson PS, Lange P, et al. Development, validation and application of a stable isotope dilution liquid chromatography electrospray ionization/selected reaction monitoring/mass spectrometry (SID-LC/ESI/SRM/MS) method for quantification of keto-androgens in human serum. J Steroid Biochem Mol Biol 2013;138:281–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Rizzardi AE, Johnson AT, Vogel RI, Pambuccian SE, Henriksen J, Skubitz AP, et al. Quantitative comparison of immunohistochemical staining measured by digital image analysis versus pathologist visual scoring. Diagnostic pathology. 2012;7:42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Missaghian E, Kempna P, Dick B, Hirsch A, Alikhani-Koupaei R, Jegou B, et al. Role of DNA methylation in the tissue-specific expression of the CYP17A1 gene for steroidogenesis in rodents. The Journal of endocrinology. 2009;202(1):99–109. [DOI] [PubMed] [Google Scholar]
- 30.Hornsby PJ, Cheng CY, Lala DS, Maghsoudlou SS, Raju SG, Yang L. Changes in gene expression during senescence of adrenocortical cells in culture. J Steroid Biochem Mol Biol 1992;43(8):951–60. [DOI] [PubMed] [Google Scholar]
- 31.Kosir R, Zmrzljak UP, Bele T, Acimovic J, Perse M, Majdic G, et al. Circadian expression of steroidogenic cytochromes P450 in the mouse adrenal gland--involvement of cAMP-responsive element modulator in epigenetic regulation of Cyp17a1. The FEBS journal. 2012;279(9):1584–93. [DOI] [PubMed] [Google Scholar]
- 32.Parker L, Lai M, Wolk F, Lifrak E, Kim S, Epstein L, et al. Orchiectomy does not selectively increase adrenal androgen concentrations. J Clin Endocrinol Metab 1984;59(3):547–50. [DOI] [PubMed] [Google Scholar]
- 33.Johnsen IK, Slawik M, Shapiro I, Hartmann MF, Wudy SA, Looyenga BD, et al. Gonadectomy in mice of the inbred strain CE/J induces proliferation of sub-capsular adrenal cells expressing gonadal marker genes. The Journal of endocrinology. 2006;190(1):47–57. [DOI] [PubMed] [Google Scholar]
- 34.Vianello S, Waterman MR, Dalla Valle L, Colombo L. Developmentally regulated expression and activity of 17alpha-hydroxylase/C-17,20-lyase cytochrome P450 in rat liver. Endocrinology. 1997;138(8):3166–74. [DOI] [PubMed] [Google Scholar]
- 35.Dalla Valle L, Vianello S, Belvedere P, Colombo L. Rat cytochrome P450c17 gene transcription is initiated at different start sites in extraglandular and glandular tissues. J Steroid Biochem Mol Biol. 2002;82(4–5):377–84. [DOI] [PubMed] [Google Scholar]
- 36.Lam HM, McMullin R, Nguyen HM, Coleman I, Gormley M, Gulati R, et al. Characterization of an Abiraterone Ultraresponsive Phenotype in Castration-Resistant Prostate Cancer Patient-Derived Xenografts. Clin Cancer Res. 2017;23(9):2301–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Li R, Evaul K, Sharma KK, Chang KH, Yoshimoto J, Liu J, et al. Abiraterone inhibits 3beta-hydroxysteroid dehydrogenase: a rationale for increasing drug exposure in castration-resistant prostate cancer. Clin Cancer Res. 2012;18(13):3571–9. [DOI] [PubMed] [Google Scholar]
- 38.Luo J, Attard G, Balk SP, Bevan C, Burnstein K, Cato L, et al. Role of Androgen Receptor Variants in Prostate Cancer: Report from the 2017 Mission Androgen Receptor Variants Meeting. Eur Urol. 2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Storbeck KH, Swart AC, Fox CL, Swart P. Cytochrome b5 modulates multiple reactions in steroidogenesis by diverse mechanisms. J Steroid Biochem Mol Biol. 2015;151:66–73. [DOI] [PubMed] [Google Scholar]
- 40.Mostaghel EA. Steroid hormone synthetic pathways in prostate cancer. Translational andrology and urology. 2013;2(3):212–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Zhang X, Morrissey C, Sun S, Ketchandji M, Nelson PS, True LD, et al. Androgen receptor variants occur frequently in castration resistant prostate cancer metastases. PLoS ONE. 2011;6(11):e27970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kim W, Zhang L, Wilton JH, Fetterly G, Mohler JL, Weinberg V, et al. Sequential use of the androgen synthesis inhibitors ketoconazole and abiraterone acetate in castration-resistant prostate cancer and the predictive value of circulating androgens. Clin Cancer Res. 2014;20(24):6269–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ryan CJ, Molina A, Li J, Kheoh T, Small EJ, Haqq CM, et al. Serum androgens as prognostic biomarkers in castration-resistant prostate cancer: results from an analysis of a randomized phase III trial. J Clin Oncol. 2013;31(22):2791–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ryan CJ, Peng W, Kheoh T, Welkowsky E, Haqq CM, Chandler DW, et al. Androgen dynamics and serum PSA in patients treated with abiraterone acetate. Prostate cancer and prostatic diseases. 2014;17(2):192–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Pignatelli D, Xiao F, Gouveia AM, Ferreira JG, Vinson GP. Adrenarche in the rat. The Journal of endocrinology. 2006;191(1):301–8. [DOI] [PubMed] [Google Scholar]
- 46.Quinn TA, Ratnayake U, Dickinson H, Nguyen TH, McIntosh M, Castillo-Melendez M, et al. Ontogeny of the adrenal gland in the spiny mouse, with particular reference to production of the steroids cortisol and dehydroepiandrosterone. Endocrinology. 2013;154(3):1190–201. [DOI] [PubMed] [Google Scholar]
- 47.Bell JB, Gould RP, Hyatt PJ, Tait JF, Tait SA. Properties of rat adrenal zona reticularis cells: production and stimulation of certain steroids. The Journal of endocrinology. 1979;83(3):435–47. [DOI] [PubMed] [Google Scholar]
- 48.Canonaco M, Ando S, Valenti A, Tavolaro R, Panno ML, Maggiolini M, et al. The in-vitro transformation of [3H]dehydroepiandrosterone into its principal metabolites in the adrenal cortex of adult castrated male rats and following steroid treatment. The Journal of endocrinology. 1989;121(3):419–24. [DOI] [PubMed] [Google Scholar]
- 49.Ando S, Canonaco M, Valenti A, Aquila S, Tavolaro R, Maggiolini M, et al. The in vitro conversion of 3H androstenedione to testosterone and dihydrotestosterone in the adrenal gland of castrated male rat: influence of gonadal steroid administration. Experimental and clinical endocrinology. 1989;93(1):83–9. [DOI] [PubMed] [Google Scholar]
- 50.Coyotupa J, Parlow AF, Kovacic N. Serum testosterone and dihydrotestosterone levels following orchiectomy in the adult rat. Endocrinology. 1973;92(6):1579–81. [DOI] [PubMed] [Google Scholar]
- 51.Kashiwagi B, Shibata Y, Ono Y, Suzuki R, Honma S, Suzuki K. Changes in testosterone and dihydrotestosterone levels in male rat accessory sex organs, serum, and seminal fluid after castration: establishment of a new highly sensitive simultaneous androgen measurement method. Journal of andrology. 2005;26(5):586–91. [DOI] [PubMed] [Google Scholar]
- 52.Ando S, Aquila S, Beraldi E, Canonaco M, Panno ML, Valenti A, et al. Physiological changes in androgen plasma levels with elapsing of time from castration in adult male rats. Hormone and metabolic research = Hormon- und Stoffwechselforschung = Hormones et metabolisme. 1988;20(2):96–9. [DOI] [PubMed] [Google Scholar]
- 53.Basham KJ, Hung HA, Lerario AM, Hammer GD. Mouse models of adrenocortical tumors. Mol Cell Endocrinol. 2016;421:82–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Nilsson ME, Vandenput L, Tivesten A, Norlen AK, Lagerquist MK, Windahl SH, et al. Measurement of a Comprehensive Sex Steroid Profile in Rodent Serum by High-Sensitive Gas Chromatography-Tandem Mass Spectrometry. Endocrinology. 2015;156(7):2492–502. [DOI] [PubMed] [Google Scholar]
- 55.Li Z, Alyamani M, Li J, Rogacki K, Abazeed M, Upadhyay SK, et al. Redirecting abiraterone metabolism to fine-tune prostate cancer anti-androgen therapy. Nature. 2016;533(7604):547–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hu R, Lu C, Mostaghel EA, Yegnasubramanian S, Gurel M, Tannahill C, et al. Distinct transcriptional programs mediated by the ligand-dependent full-length androgen receptor and its splice variants in castration-resistant prostate cancer. Cancer Res 2012;72(14):3457–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Hu R, Dunn TA, Wei S, Isharwal S, Veltri RW, Humphreys E, et al. Ligand-independent androgen receptor variants derived from splicing of cryptic exons signify hormone-refractory prostate cancer. Cancer Res 2009;69(1):16–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Chen EJ, Sowalsky AG, Gao S, Cai C, Voznesensky O, Schaefer R, et al. Abiraterone treatment in castration-resistant prostate cancer selects for progesterone responsive mutant androgen receptors. Clin Cancer Res 2015;21(6):1273–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kumagai J, Hofland J, Erkens-Schulze S, Dits NF, Steenbergen J, Jenster G, et al. Intratumoral conversion of adrenal androgen precursors drives androgen receptor-activated cell growth in prostate cancer more potently than de novo steroidogenesis. The Prostate. 2013. [DOI] [PubMed] [Google Scholar]
- 60.Ando T, Nishiyama T, Takizawa I, Miyashiro Y, Hara N, Tomita Y. A carbon 21 steroidal metabolite from progestin, 20beta-hydroxy-5alpha-dihydroprogesterone, stimulates the androgen receptor in prostate cancer cells. The Prostate. 2018;78(3):222–32. [DOI] [PubMed] [Google Scholar]
- 61.Holzbeierlein J, Lal P, LaTulippe E, Smith A, Satagopan J, Zhang L, et al. Gene expression analysis of human prostate carcinoma during hormonal therapy identifies androgen-responsive genes and mechanisms of therapy resistance. Am J Pathol 2004;164(1):217–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Mitsiades N, Sung CC, Schultz N, Danila DC, He B, Eedunuri VK, et al. Distinct patterns of dysregulated expression of enzymes involved in androgen synthesis and metabolism in metastatic prostate cancer tumors. Cancer Res 2012;72(23):6142–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Hofland J, van Weerden WM, Dits NF, Steenbergen J, van Leenders GJ, Jenster G, et al. Evidence of limited contributions for intratumoral steroidogenesis in prostate cancer. Cancer Res 2010;70(3):1256–64. [DOI] [PubMed] [Google Scholar]
- 64.Jernberg E, Thysell E, Bovinder Ylitalo E, Rudolfsson S, Crnalic S, Widmark A, et al. Characterization of prostate cancer bone metastases according to expression levels of steroidogenic enzymes and androgen receptor splice variants. PLoS ONE. 2013;8(11):e77407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Sakai M, Martinez-Arguelles DB, Aprikian AG, Magliocco AM, Papadopoulos V. De novo steroid biosynthesis in human prostate cell lines and biopsies. The Prostate. 2016;76(6):575–87. [DOI] [PubMed] [Google Scholar]
- 66.Efstathiou E, Titus M, Tsavachidou D, Tzelepi V, Wen S, Hoang A, et al. Effects of abiraterone acetate on androgen signaling in castrate-resistant prostate cancer in bone. J Clin Oncol 2012;30(6):637–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Hagberg Thulin M, Nilsson ME, Thulin P, Ceraline J, Ohlsson C, Damber JE, et al. Osteoblasts promote castration-resistant prostate cancer by altering intratumoral steroidogenesis. Mol Cell Endocrinol 2016;422:182–91. [DOI] [PubMed] [Google Scholar]
- 68.Michiel Sedelaar JP, Dalrymple SS, Isaacs JT. Of mice and men--warning: intact versus castrated adult male mice as xenograft hosts are equivalent to hypogonadal versus abiraterone treated aging human males, respectively. The Prostate. 2013;73(12):1316–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Oksala R CYP11A1 inhibition as a therapeutic approach for the treatment of castration resistant prostate cancer. Journal of clinical oncology. 2018;36(6_suppl):340-. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.