

Abstract
Diabetes mellitus (DM) is a chronic metabolic disease with high morbimortality rates. DM has two types: type 1, which is often associated with a total destruction of pancreatic beta cells, and non-insulin-dependent or type 2 diabetes mellitus (T2DM), more closely associated with obesity and old age. The main causes of T2DM are insulin resistance and/or inadequate insulin secretion. Protein-tyrosine phosphatase 1B (PTP1B) negatively regulates insulin signaling pathways and plays an important role in T2DM, as its overexpression may induce insulin resistance. Thus, since PTP1B may be a therapeutic target for both T2DM and obesity, the search for novel and promising natural inhibitors has gained much attention. Hence, several marine organisms, including macro and microalgae, sponges, marine invertebrates, sea urchins, seaweeds, soft corals, lichens, and sea grasses, have been recently evaluated as potential drug sources. This review provides an overview of the role of PTP1B in T2DM insulin signaling and treatment, and highlights the recent findings of several compounds and extracts derived from marine organisms and their relevance as upcoming PTP1B inhibitors. In this systematic literature review, more than 60 marine-derived metabolites exhibiting PTP1B inhibitory activity are listed. Their chemical classes, structural features, relative PTP1B inhibitory potency (assessed by IC50 values), and structure–activity relationships (SARs) that could be drawn from the available data are discussed. The upcoming challenge in the field of marine research—metabolomics—is also addressed.
Keywords: protein-tyrosine phosphatase 1B, type 2 diabetes mellitus, insulin signaling pathways, marine metabolites
1. Introduction
Diabetes mellitus (DM) is considered a major health problem worldwide [1]. Obesity and diabetes incidence still continue increasing due to globalization, mechanization, and changes in human lifestyle and daily routines [2]. According to the International Diabetes Federation (IDF), it was estimated that in 2017 there were 451 million (age 18–99 years) people with diabetes worldwide. These figures were expected to increase to 693 million by 2045 [3]. DM is a chronic metabolic disease that results from defects in insulin action, insulin secretion, or both, leading to persistent hyperglycemia [4].
Currently, type 2 diabetes mellitus (T2DM) represents a major threat to health [5]. Characterized by increased blood glucose levels, this is the underlying reason for several complications, including cardiovascular disorders, blindness, kidney failure, and peripheral nerve damages [6]. The development of T2DM and its complications are related, in most cases, to insulin resistance and postprandial hyperglycemic variations [7,8]. Thus, an effective drug for controlling insulin resistance may be beneficial in improving the quality life of T2DM patients. Several pharmacological strategies have been investigated on DM treatment, including insulin release stimulation, gluconeogenesis inhibition, glucose transport activity increase, and intestinal glucose absorption reduction [9]. Insulin supplements and other oral anti-diabetic drugs can be used alone or in combination to improve glycemic regulation [10]. However, some of the available anti-diabetic drugs have either the disadvantage of having low efficacy or serious side effects [11]. Thus, there is a continuous search for more effective and safer anti-hyperglycemic agents, especially from natural origins.
Insulin sensitizers, such as thiazolidinediones (TZDs or glitazones) have been used as effective drugs for T2DM treatment [12]. The identification of the enzyme responsible for the dephosphorylation of insulin receptors, called protein-tyrosine phosphatase 1B (PTP1B), showed that the inhibitors of such an enzyme could be employed as insulin sensitizer agents and, therefore, as promising anti-diabetic drugs [13]. This hypothesis was confirmed in mouse models, where it was found that PTP1B gene disruption can increase insulin sensitivity. Similar results were also obtained when PTP1B antisense nucleotides suppressed PTP1B gene expression [14].
Protein tyrosine phosphatases (PTPs) constitute a huge and structurally variable family of highly regulated enzymes. Most PTPs have been proposed to be targets for advanced drug discovery, and PTP1B is one of the well-established enzymes among PTPs [15,16,17]. It was the first isolated member of the PTP superfamily, and since then, growing evidence has linked it with insulin resistance, obesity, and T2DM. Numerous studies have shown that PTP1B can negatively regulate insulin and leptin signaling pathways. Indeed, PTP1B dephosphorylates both insulin receptor and its substrate IRS-1 in the insulin signaling pathway [18,19], whereas in the leptin pathway, PTP1B binds and dephosphorylates tyrosine kinase downstream of the Janus-Activated Kinase 2 (JAK2) leptin receptor [20]. In cell cultures, PTP1B overexpression causes a decrease in the insulin-stimulated phosphorylation of IR and IRS-1, while PTP1B raises insulin-initiated signaling level reduction [21,22]. The hypothesis that PTP1B expression can contribute to diabetes and obesity is supported by quantitative analysis of trait loci and mutations in the human PTP1B gene [23]. In in vivo studies, PTP1B knockout mice exhibited elevated resistance to high-fat diet-induced obesity and insulin sensitivity [24,25]. In addition, other studies on tissue-specific PTP1B knockout mice have shown that leptin action, adiposity, as well as body weight are controlled by neuronal PTP1B [26]. Generally, many studies suggest that PTP1B inhibitors constitute a highly promising approach for T2DM and obesity amelioration.
Aryl carboxylic acids, such as isoxazole [27], hydroxylpropionic [28], 2-oxalylamino benzoic (OBA) acids [29], and thiophene diacid [30], have been recognized as alternative phosphotyrosine (pTyr) surrogates to overcome the lack of cellular activity of highly charged phosphonates (Figure 1). Furthermore, it was reported that benzyl aryl α-ketoacid derivatives revealed significant PTP1B inhibitory effects in a non-competitive pattern, targeting conserved protein loop (WPD loop) open conformation [31]. It was also noticed that the existence of a benzyl group in these bioactive molecules may enhance PTP1B binding affinity, and being hydrophobic in nature, it also increases their cell membrane permeability. Recent studies also suggested that PTP1B may become an oncogene in breast cancer [16]. Accordingly, multiple studies have been conducted focusing on the development of new PTP1B inhibitors for the treatment of T2DM, obesity, and cancer, but to the authors’ knowledge there are no review articles published on this subject. In this sense, the present review aims to provide an overview of the role of PTP1B in T2DM insulin signaling and treatment, and to highlight the most recent findings on several compounds and extracts discovery from marine organisms and their relevance as upcoming PTP1B inhibitors.
Figure 1.

Structures of phosphotyrosine (pTyr) surrogate acids.
2. Marine Sources as Upcoming Therapeutic Agents
The marine environment is considered a wide and relatively unexploited source of bioactive compounds with high biodiversity, including fatty acids (especially polyunsaturated fatty acids), proteins, polyphenols, sterols, sulfated polysaccharides, and pigments [32,33,34,35,36]. Indeed, marine algae has been increasingly exploited as renowned sources of metabolites with promising biological effects, including antioxidant, hypoglicemic, hypotensive, hypolipidemic, antibacterial, and antiviral activities [37,38]. Specifically, macroalgae are considered healthy foods as they are rich in minerals and dietary fibers. Traditionally, the Far East and Hawaiian Islands, Japan, Korea, and China consume marine algae as a common component of their diets. Macroalgae species can reach 9000 species and can be classified according to their pigment composition into three classes, i.e., Phaeophyta, Rhodophyta, and Chlorophyta (also known as brown, red, and green algae, respectively) [39].
Unique metabolites from diverse classes have been isolated from different marine plants, with in vivo remarkable pharmacological effects [40], such as anticancer, anti-hyperlipidemic, anti-diabetic, anti-hypertensive, antioxidant, anti-inflammatory, anticoagulant, anti-estrogenic, antibacterial, antifungal, antiviral, immunomodulatory, neuroprotective, and tissue healing properties [41]. More recently, as a result of the characterization of a large number of bioactive metabolites from marine macroalgae, there has been a growing interest in the search for potential applications of macroalgae and their metabolites as functional constituents for human and animal health benefits [42]. Functional constituents of macroalgae have been increasingly used as food supplements as well as for anti-diabetic purposes [40]. Hereby, the possible applications of marine macroalgae and/or macroalgae-derived bioactive metabolites for PTP1B inhibitory effects have been greatly expanded.
3. Marine-Derived Molecules with PTP1B Inhibitory Activity
3.1. Ptp1b Inhibitory Activity: In Vitro Findings
Around 300 natural products with PTP1B inhibitory capacity were isolated and characterized from different natural sources, many of them from marine origin [43]. The isolation and identification of sulfircin, a sesterterpene sulfate, from deep-water sponge Ircinia (unknown species), was the first documented marine metabolite possessing PTP1B inhibitory activity [43]. Since then, marine sponges have been considered valuable sources of PTP1B inhibitors with diverse structures [44], such as polybromodiphenyl ether [45], sesquiterpenoids, and sesquiterpene quinones [46]. Nevertheless, the novelty of marine resource screening models has encouraged the development of new studies targeting these resources as upcoming anti-diabetic agents. Marine algae, seaweeds, soft corals, sponges and lichens are considered to be among these models as they were found to exhibit PTP1B inhibitory effects. Table 1, Table 2, Table 3, Table 4, Table 5, Table 6, Table 7 and Table 8 summarize a large number of isolated compounds from marines that have PTP1B inhibitory effects with varying potencies. In the following sections, the PTP1B inhibitory activity of some of these compounds are discussed.
Table 1.
Marine plant-isolated bromophenols with in vitro PTP1B inhibitory effects.
| No. | Compound/Structure | Marine Species | Outcomes/Enzymes | Reference |
|---|---|---|---|---|
| 1 | 2,2′,3,3′-Tetrabromo-4,4′,5,5′-tetra-hydroxydiphenyl methane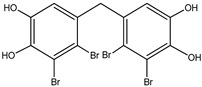
|
Red algae Rhodomela confervoides | PTP1B inhibition (IC50 = 2.4 μM) |
[53] |
| 2 | 3-Bromo-4,5-Bis-(2,3-dibromo-4,5-dihydroxybenzyl)pyrocatechol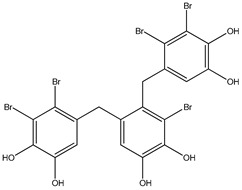
|
Red algae Rhodomela confervoides | PTP1B inhibition (IC50 = 1.7 μM) |
[53] |
| 3 | Bis-(2,3-dibromo-4,5-dihydroxybenzyl) ether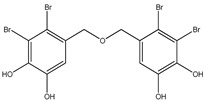
|
Red algae Rhodomela confervoides | PTP1B inhibition (IC50 = 1.5 μM) α-glucosidase inhibition (IC50 = 0.098 μM) |
[53] |
| 4 | 2,2′,3,3′-Tetrabromo-3′,4,4′,5-tetrahydroxy-6′-ethyloxymethyldiphenylmethane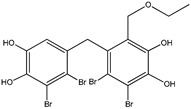
|
Red algae Rhodomela confervoides | PTP1B inhibition (IC50 = 0.8 μM) |
[53] |
| 5 | 3,4-Dibromo-5-(2-bromo-3,4-dihydroxy-6-(ethoxymethyl)benzyl)benzene-1,2-diol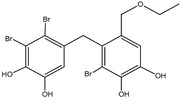
|
Red algae Rhodomela confervoides | PTP1B inhibition (IC50 = 0.8 μM) |
[49,50,51,52] |
| 6 | 3,4-Dibromo-5-(methoxymethyl)benzene-1,2-diol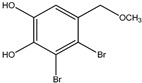
|
Red algae Rhodomela confervoides | PTP1B inhibition (IC50 = 3.4 μM) |
[51,53] |
| 7 | 3-(2,3-Dibromo-4,5-dihydroxyphenyl)-2-methylpropanal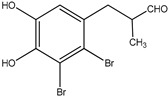
|
Red algae Rhodomela confervoides | PTP1B inhibition (IC50 = 4.5 μM) |
[51,53] |
| 8 | 3,4-Dibromo-5-(2-bromo-3,4-dihydroxy-6-(isobutoxymethyl)benzyl)benzene-1,2-diol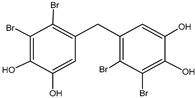
|
Red algae Rhodomela confervoides | PTP1B inhibition (IC50 = 2.4 μM) |
[51,53] |
| 9 | 7-Bromo-1-(2,3-dibromo-4,5-dihydroxy phenyl)-2,3-dihydro-1H-indene-5,6-diol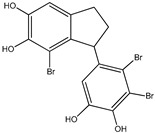
|
Red algae Rhodomela confervoides | PTP1B inhibition (IC50 = 2.8 μM) |
[51,53] |
| 10 | 5,5’-(3-Bromo-4,5-dihydroxy-1,2-phenylene)-Bis-(methylene))Bis-(3,4-dibromobenzene-1,2-diol)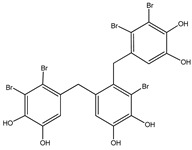
|
Red algae Rhodomela confervoides | PTP1B inhibition (IC50 = 1.7 μM) |
[51,53] |
| 11 | 3,4-Dibromo-5-(2-bromo-3,4-dihydroxy-6-(ethoxymethyl)benzyl)benzene-1,2-diol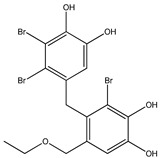
|
Red algae Rhodomela confervoides | PTP1B inhibition (IC50 = 0.84 μM) |
[50,51] |
| 12 | 2-(3′,5′-Dibromo-2′-methoxyphenoxy)-3,5-dibromophenol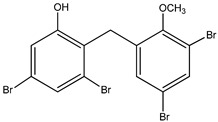
|
Indonesian marine sponge Lamellodysidea herbacea | PTP1B inhibition (IC50 = 0.9 μM) |
[45] |
| 13 | 2-(3′,5′-Dibromo-2′-methoxyphenoxy)-3,5-dibromophenol-methyl ether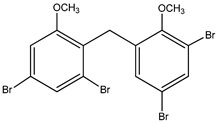
|
Indonesian marine sponge Lamellodysidea herbacea | PTP1B inhibition (IC50 = 1.7 μM) |
[45] |
| 14 | 2,3,6-Tribromo-4,5-dihydroxybenzyl methyl ether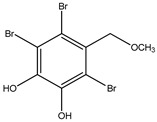
|
Marine red algae Symphyocladia latiuscula | PTP1B inhibition (IC50 = 3.9 µM) |
[55] |
| 15 | Bis-(2,3,6-tribromo-4,5-dihydroxyphenyl) methane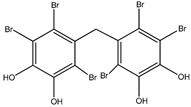
|
Marine red algae Symphyocladia latiuscula | PTP1B inhibition (IC50 = 4.3 µM) |
[55] |
| 16 | 1,2-Bis-(2,3,6-tribromo-4,5-dihydroxyphenyl)-ethane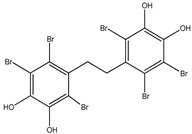
|
Marine red algae Symphyocladia latiuscula | PTP1B inhibition (IC50 = 2.7 µM) |
[55] |
Table 2.
Marine plant-isolated brominated metabolites with in vitro PTP1B inhibitory effects.
| No. | Compound/Structure | Marine Species | Outcomes/Enzymes | Reference |
|---|---|---|---|---|
| 17 | 3’,5’,6’,6-Tetrabromo-2,4-dimethyldiphenyl ether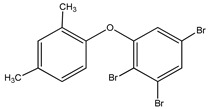
|
Red algae Laurencia similis | PTP1B inhibition (IC50 = 3.0 μM) | [60] |
| 18 | 2’,5’,6’,5,6-Pentabromo-3’,4’,3,4-tetramethoxybenzo-phenone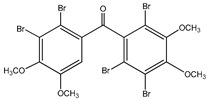
|
Red algae Laurencia similis | PTP1B inhibition (IC50 = 2.7 μM) | [60] |
| 19 | 3’,5’,6’6-Tetrabromo-2,4-dimethyldiphenyl ether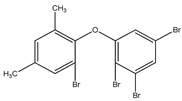
|
Red algae Laurencia similis | PTP1B inhibition (IC50 = 3.0 µg/mL) | [60] |
| 20 | 1,2,5-Tribromo-3-bromoamino-7-bromomethylnaphthalene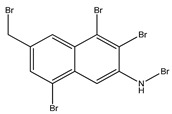
|
Red algae Laurencia similis | PTP1B inhibition (IC50 = 102 µg/mL) | [60] |
| 21 | 2,5,8-Tribromo-3-bromoamino-7-bromomethylnaphthalene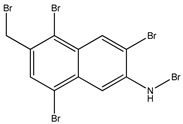
|
Red algae Laurencia similis | PTP1B inhibition (IC50 = 65.3 µg/mL) | [60] |
| 22 | 2,5,6-Tribromo-3-bromoamino-7-bromomethylnaphthalene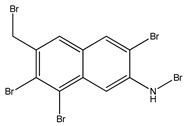
|
Red algae Laurencia similis | PTP1B inhibition (IC50 = 69.8 µg/mL) | [60] |
| 23 | 2’,5’,6’,5,6-Pentabromo-3’,4’,3,4-tetramethoxybenzo-phenone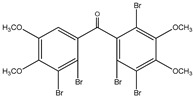
|
Red algae Laurencia similis | PTP1B inhibition (IC50 = 2.7 µg/mL) | [60] |
Table 3.
Marine plant-isolated polybromodiphenyl ether derivatives with in vitro PTP1B inhibitory effects.
| No. | Compound/Structure | Marine Species | Outcomes/Enzymes | Reference |
|---|---|---|---|---|
| 24 | 2-(3’,5’-Dibromo-2’-methoxyphenoxy)-3,5-dibromophenol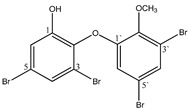
|
Indonesian marine sponge Lamellodysidea herbacea | PTP1B inhibition (IC50 = 0.9 µM) Huh-7 inhibition (IC50 = 32 µM) |
[45] |
| 25 | 3,5-Dibromo-2-(3’,5’-dibromo-2’-methoxyphenoxy)-1-methoxybenzene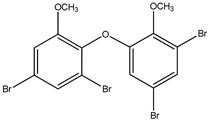
|
Indonesian marine sponge Lamellodysidea herbacea | PTP1B inhibition (IC50 = 1.7 µM) Huh-7 inhibition (IC50 = 48 µM) |
[45] |
| 26 | 3,5-Dibromo-2-(3’,5’-dibromo-2’ -methoxyphenoxy)phenylethanoate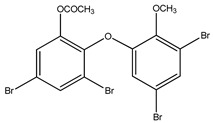
|
Indonesian marine sponge Lamellodysidea herbacea | PTP1B inhibition (IC50 = 0.6 µM) |
[45] |
| 27 | 3,5-Dibromo-2-(3’,5’-dibromo-2’ -methoxyphenoxy)phenylbutanoate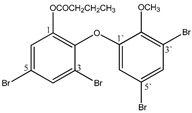
|
Indonesian marine sponge Lamellodysidea herbacea | PTP1B inhibition (IC50 = 0.7 µM) |
[45] |
| 28 | 3,5-Dibromo-2-(3’,5’-dibromo-2’ -methoxyphenoxy)phenylhexanoate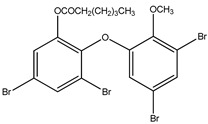
|
Indonesian marine sponge Lamellodysidea herbacea | PTP1B inhibition (IC50 = 0.7 µM) |
[45] |
| 29 | 3,5-Dibromo-2-(3’,5’-dibromo-2’ -methoxyphenoxy)phenyl benzoate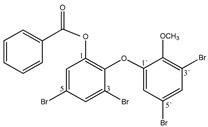
|
Indonesian marine sponge Lamellodysidea herbacea | PTP1B inhibition (IC50 = 1.0 µM) |
[45] |
Table 4.
Marine plant-isolated phlorotannins with in vitro PTP1B inhibitory effects.
| No. | Compound/Structure | Marine Species | Outcomes/Enzymes | Reference |
|---|---|---|---|---|
| 30 | Eckol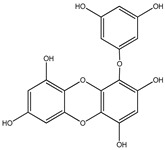
|
Edible brown algae Ecklonia stolonifera and Eisenia bicyclis |
PTP1B inhibition (IC50 = 2.6 µM) α-glucosidase inhibition (IC50 = 22.8 µM) |
[64,65] |
| 31 | Phlorofurofucoeckol-A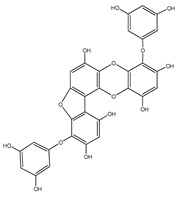
|
Edible brown algae Ecklonia stolonifera and Eisenia bicyclis | PTP1B inhibition (IC50 = 0.6 µM) α-glucosidase inhibition (IC50 = 1.4 µM) |
[64,65] |
| 32 | Dieckol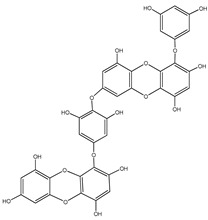
|
Edible brown algae Ecklonia stolonifera and Eisenia bicyclis | PTP1B inhibition (IC50 = 1.2 µM) α-glucosidase inhibition (IC50 = 1.6 µM) |
[64,65] |
| 33 | 7-Phloroeckol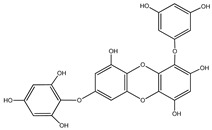
|
Edible brown algae Ecklonia stolonifera and Eisenia bicyclis | PTP1B inhibition (IC50 = 2.1 µM) α-glucosidase inhibition (IC50 = 6.1 µM) |
[65] |
| 34 | Phloroglucinol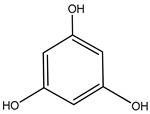
|
Edible brown algae Ecklonia stolonifera and Eisenia bicyclis | PTP1B inhibition (IC50 = 55.5 µM) α-glucosidase inhibition (IC50 = 141.2 µM) |
[65] |
| 35 | Dioxinodehydroeckol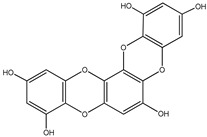
|
Edible brown algae Ecklonia stolonifera and Eisenia bicyclis | PTP1B inhibition (IC50 = 30.0 µM) α-glucosidase inhibition (IC50 = 34.6 µM) |
[65] |
Table 5.
Marine plant-isolated sterols with in vitro PTP1B inhibitory effects.
| No. | Compound/Structure | Marine Species | Outcomes/Enzymes | Reference |
|---|---|---|---|---|
| 36 | 24-Hydroperoxy-24-Vinylcholesterol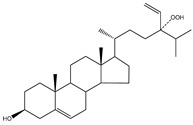
|
Marine invertebrates South China Sea sponge Xestospongia testudinaria Lamarck (Petrosiidae) | - | [68] |
| 37 | 29-Hydroperoxystigmasta-5,24(28)-dien-3-ol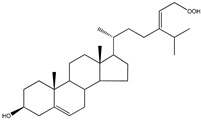
|
Marine invertebrates South China Sea sponge Xestospongia testudinaria Lamarck (Petrosiidae) | PTP1B inhibition (IC50 = 5.8 µg/mL) |
[68] |
| 38 | 5α,8α-Epidioxycholest-6-en-3β-ol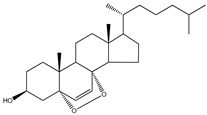
|
Sea urchin Glyptocidaris crenularis A. Agassiz (Glyptocidaridae) | - | [68] |
| 39 | 5α,8α-Epidioxycholest-6,22-dien-3β-ol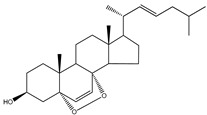
|
Sponge Mycale spp. (Mycalidae) | - | [68] |
| 40 | 5α,8α-Epidioxy-ergosta-6,22-dien-3β-ol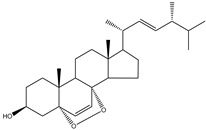
|
Gorgonian Dichotella gemmacea Milne Edwards and Haime (Ellisellidae) | - | [68] |
| 41 | 3β-Hydroxycholest-5-en-25-acetoxy-19-oate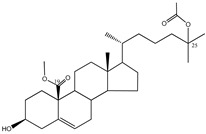
|
Gorgonian Dichotella gemmacea Milne Edwards and Haime (Ellisellidae) | - | [68] |
| 42 | Fucosterol (24-ethylidene cholesterol)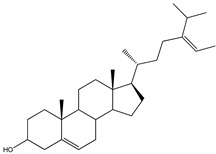
|
Brown algae Eisenia bicyclis and Ecklonia stolonifera |
Non-competitive type inhibitor against PTP1B | [67,68,69] |
Table 6.
Marine plant-isolated terpenes with in vitro PTP1B inhibitory effects.
| No. | Compound/Structure | Marine Species | Outcomes/Enzymes | Reference |
|---|---|---|---|---|
| 43 | Dysidine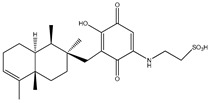
|
Sponge Dysidea villosa | PTP1B inhibition (IC50 = 6.7 μM) | [46,71] |
| 44 | Dysidavarone A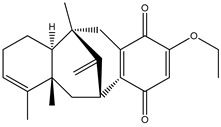
|
South China Sea sponge Dysidea avara | PTP1B inhibition (IC50 = 10.0 μM) | [70] |
| 45 | Dehydroeuryspongin A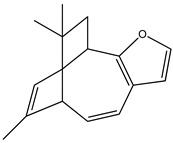
|
Marine Sponge Euryspongia spp. | PTP1B inhibition (IC50 = 3.6 μM) | [72] |
| 46 | Sarsolilide A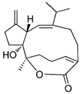
|
Hainan soft coral Sarcophyton trocheliophorum Marenzeller | PTP1B inhibition (IC50 = 6.8 μM) | [73] |
| 47 | Sarsolilide B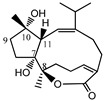
|
Hainan soft coral Sarcophyton trocheliophorum Marenzeller | PTP1B inhibition (IC50 = 27.1 μM) | [73] |
| 48 | Methyl sarcotroates A and B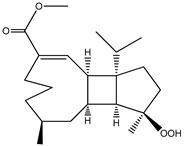
|
Hainan soft coral Sarcophyton trocheliophorum | PTP1B inhibition (IC50 = 7.0 μM) | [73] |
| 49 | 9-Oxa-2-azabicyclo-[3,3,1]-nona-3,7-diene derivative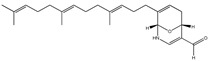
|
Sponge Hippospongia lachne of Yongxing Island | PTP1B inhibition (IC50 = 5.2 μM) | [75] |
| 50 | 2-(Aminomethylene) hepta-3,5-dienedial moiety connected with farnesyl group at C-7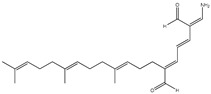
|
Sponge Hippospongia lachne of Yongxing Island | PTP1B inhibition (IC50 = 8.7 μM) | [75] |
| 51 | Hopane-6α,22-diol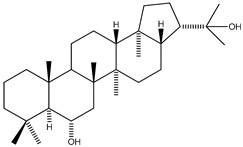
|
Antarctic lichen Lecidella carpathica | PTP1B inhibition (IC50 =3.7 μM) TCPTP inhibition (IC50 = 8.4 μM) SHP-2 inhibition (IC50 > 68 μM) LAR inhibition (IC50 > 68 μM) CD45 inhibition (IC50 > 68 μM) |
[76] |
| 52 | Stellettin G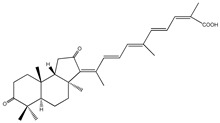
|
Hainan sponge Stelletta spp. | PTP1B inhibition (IC50 = 4.1 μM) | [77] |
TCPTP, T-cell protein tyrosine phosphatase; SHP-2, src homology phosphatase-2; LAR, leukocyte antigen-related phosphatase; CD45, CD45 tyrosine phosphatase.
Table 7.
Marine plant-isolated fungal metabolites with in vitro PTP1B inhibitory effects.
| No. | Compound/Structure | Marine Species | Outcomes/Enzymes | Reference |
|---|---|---|---|---|
| 53 | Fructigenine A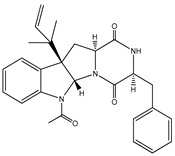
|
Marine-derived fungal strains Penicillium and Eurotium species | PTP1B inhibition (IC50 = 10.7 µM) |
[80] |
| 54 | Cyclopenol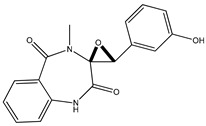
|
Marine-derived fungal strains Penicillium and Eurotium species | PTP1B inhibition (IC50 = 30.0 µM) |
[80] |
| 55 | Echinulin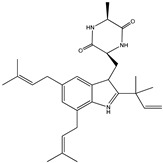
|
Marine-derived fungal strains Penicillium and Eurotium species | PTP1B inhibition (IC50 = 29.4 µM) |
[80] |
| 56 | Flavoglaucin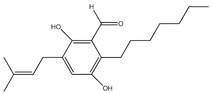
|
Marine-derived fungal strains Penicillium and Eurotium species | PTP1B inhibition (IC50 = 13.4 µM) |
[80] |
| 57 | Viridicatol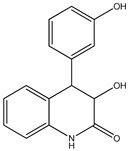
|
Marine-derived fungal strains Penicillium and Eurotium species | PTP1B inhibition (IC50 = 64.0 µM) |
[80] |
| 58 | Penstyrylpyrone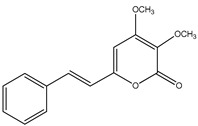
|
Marine-derived fungi Penicillium JF-55 cultures | PTP1B inhibition (IC50 = 5.3 μM) |
[82] |
| 59 | Anhydrofulvic acid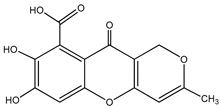
|
Marine-derived fungi Penicillium JF-55 cultures | PTP1B inhibition (IC50 = 1.9 μM) |
[82] |
| 60 | Aquastatin A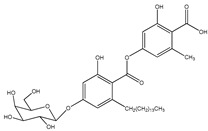
|
Fungus Cosmospora species | PTP1B inhibition (IC50 = 0.2 μM), as well as inhibition of TCPTP, SHP-2, LAR, and CD45 activity | [81,82] |
Table 8.
Marine plant-isolated miscellaneous compounds with in vitro PTP1B inhibitory effects.
| No. | Compound/Structure | Marine Species | Outcomes/Enzymes | Reference |
|---|---|---|---|---|
| 61 | Fucoxanthin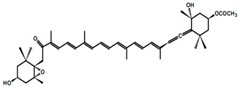
|
Phaeodactylum tricornutum, Eisenia bicyclis (Arame), Undaria pinnatifida (Wakame), and Hi-jikia fusiformis (Hijiki) | PTP1B inhibition (IC50 = 4.8 μM) |
[83,84,85] |
| 62 | Brialmontin 1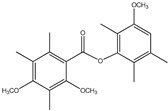
|
Antarctic lichen Lecidella carpathica | PTP1B inhibition (IC50 =14 μM) |
[76] |
| 63 | Atraric acid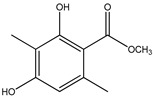
|
Antarctic lichen Lecidella carpathica | PTP1B inhibition (IC50 = 51.5 μM) |
[76] |
| 64 | Saragahydroquinoic acid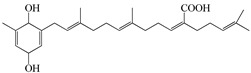
|
Brown algae Sargassum serratifolium C. Agardh | PTP1B inhibition (IC50 = 5.14 μM) |
[89] |
| 65 | Saragaquinoic acid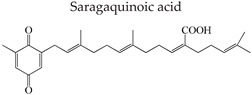
|
Brown algae Sargassum serratifolium C. Agardh | PTP1B inhibition (IC50 = 14.15 μM) |
[89] |
| 66 | Sargachromenol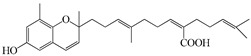
|
Brown algae Sargassum serratifolium C. Agardh | PTP1B inhibition (IC50 = 11.80 μM) |
[89] |
3.1.1. Bromophenols
As the main component of algae, bromophenols may be responsible for the reported anti-diabetic activity of many marine organisms. These compounds arise from the tendency of the phenol moiety to undergo electrophilic bromination to varying degrees (Table 1).
Bromophenols (compounds 1–11) isolated from the red algae Rhodomela confervoides have potent in vitro PTP1B inhibitory effects, with IC50 values fluctuating between 0.8 μM and 4.5 μM [47,48,49,50,51,52,53,54]. This change in potencies could be attributed to the bromine content of these compounds or to their side chains. On the other hand, Yamazaki et al. [45] isolated two bromophenols (compounds 12 and 13) from the Indonesian marine sponge Lamellodysidea herbacea and found positive in vitro PTP1B inhibitory effects, with IC50 values of 0.9 μM and 1.7 μM, respectively. Other brominated phenols (compounds 14–16) isolated from red algae Symphyocladia latiuscula by Liu et al. in 2011 [55] also exerted positive inhibitory activity, with IC50 values of 3.9 μM, 4.3 μM, and 2.7 μM, respectively.
Besides the abovementioned PTP1B inhibitory effects, bromophenols have also been reported to have strong α-glucosidase enzyme inhibitory effects. In fact, α-glucosidase enzyme plays a crucial role in carbohydrates digestion and is a favorite target for anti-diabetic drugs, especially in the case of postprandial hyperglycemia. It has been suggested that Bis-(2,3-dibromo-4,5-di hydroxybenzyl) ether (compound 3) together with bis(2,3,6-tribromo-4,5-dihydroxybenzyl) ether represents a future kind of α-glucosidase inhibitors [47,48,56,57]. Bis-(2,3,6-tribromo-4,5-dihydroxybenzyl) ether exhibited the most potent α-glucosidase inhibition activity compared to the present series of bromophenols, having an IC50 value as low as 0.03 μM [47]. On the other hand, the lowest activity was exhibited by 2,4-dibromophenol (IC50 = 110.4 μM). This may bind α-glucosidase inhibition activity to the bromination degree of these metabolites based on the inversely proportional relationship between their IC50 value and the number of bromines in metabolites. Likewise, its enzymatic inhibition activity increases proportionately with the increase in the number of phenyl units. Thus, Bis-(2,3-dibromo-4,5-dihydroxybenzyl) ether and Bis-(2,3,6-tribromo-4,5-dihydroxybenzyl) ether, with a diphenyl unit, are found to exhibit much more activity than metabolites with a single phenyl unit, such as 3-bromo-4,5-dihydroxybenzyl alcohol. Nevertheless, the underlying reason behind these changes in biological activity needs further clarification.
Interestingly, along with PTP1B and α-glucosidase inhibition activity, some bromophenols have also shown aldose reductase inhibitory effects. Indeed, aldose reductase is considered the basic enzyme of the polyol pathway, which controls sorbitol formation from glucose and plays a significant role in degenerative complications resulting from diabetes development [58]. 2,2′,3,6,6′-pentabromo-3′,4,4′,5-tetrahydroxydibenzyl ether, Bis-(2,3,6-tribromo- 4,5-dihydroxyphenyl) methane, 2′,3,5′,6-pentabromo-3′,4,4′,5-tetrahydroxydiphenyl methane, 2,3,6-tribromo-4,5-dihydro xymethyl benzene, and 2,3,6-tribromo-4,5-dihydroxybenzaldehyde isolated from the red algae S. latiuscula have been reported as aldose reductase inhibitors, and thus could be beneficial in T2DM complications (such as eye and nerve damage management) [59].
3.1.2. Brominated Metabolites
In 2010, Qin et al. [60] assessed the in vitro PTP1B inhibitory action of two brominated metabolites (compounds 17 and 18) isolated from the red algae Laurencia similis. Both compounds showed PTP1B inhibition with IC50 values of 3.0 μM and 2.7 μM, respectively. The authors also studied the effect of highly brominated metabolites (compounds 19–23), but their corresponding IC50 values were much higher than those of the brominated metabolites, ranging from 65.3 µg/mL to 102 µg/mL. This could question the hypothesis that the bromination degree affects the PTP1B inhibitory effect in a directly proportional manner (Table 2).
3.1.3. Polybromodiphenyl Ether Derivatives
Besides the two abovementioned bromophenols (compounds 12 and 13), the Indonesian marine sponge Lamellodysidea herbacea contains six polybromodiphenyl ether derivatives (compounds 24–29). As stated by Yamazaki et al. [45], all of these compounds exhibited in vitro PTP1B inhibitory action with IC50 values ranging from 0.6 μM to 1.7 μM. The authors also determined the activity of compounds 24 and 25 in Huh-7, a well-differentiated hepatocyte-derived cellular carcinoma cell line that has been increasingly investigated, given its ability to secrete mitogen hepatoma-derived growth factor, responsible for promoting cell growth without depending on other growth factors present in serum. The IC50 values obtained for these compounds were, respectively, 32 µM and 48 µM (Table 3).
3.1.4. Phlorotannins
Back in 1977, Glombitza was the first to introduce the term phlorotannins [61]. Briefly, they are a characteristic type of integral tannins found in brown algae, Alariaceae, and are basically classified into six main subclasses (Table 4): eckols, fucols, phlorethols, fucophloretols, fuhalols, and isofuhalos [62,63]. Phlorotannins are thought to modulate cellular signaling, leading to the regulation of different body conditions [49]. Eckol and its derivatives (compounds 30–35) isolated from the edible brown algae Ecklonia stolonifera and Eisenia bicyclis were studied by several authors [64,65,66], who found variable anti-diabetic effects. The in vitro PTP1B inhibitory activity was variable, with IC50 values ranging from 0.6 μM to 55.5 μM, whereas the in vitro α-glucosidase inhibitory action exhibited more potent IC50 values ranging from 1.4 μM to 141.2 μM.
It is worth mentioning that phlorofurofucoeckol-A (compound 31) showed the lowest IC50 values for both enzymes. In contrast, phloroglucinol (compound 34), which is actually the building unit of other polymer phlorotannins, had the highest IC50 values for both enzymes. From this, it is possible to suppose that the activity is due to the product of polymerization, not to the basic monomeric structure.
3.1.5. Sterols
This class of secondary metabolites deserves more attention (Table 5). The few studies performed in this field [67,68] focused on hydroperoxyl sterols, epidioxy sterols, and fucosterol contents in different marine invertebrates, including sea sponge Xestospongia testudinaria Lamarck (Petrosiidae) from South China, sea urchin Glyptocidaris crenularis A. Agassiz (Glyptocidaridae), sea sponge Mycale species (Mycalidae), Gorgonian Dichotella gemmacea Milne Edwards and Haime (Ellisellidae), algae, seaweed, and diatoms. Several sterols (compounds 36–42) were identified in these different marine species, but their PTP1B inhibitory activity is still undetermined, with the exception of compound 37 (29-hydroperoxystigmasta-5,24(28)-dien-3-ol), in which a positive effect was stated (IC50 = 5.8 µg/mL), and compound 42, which showed moderate PTP1B inhibitory activity [69]. These findings encourage further studies on similar compounds.
3.1.6. Terpenes
Sesquiterpenes
Sesquiterpene quinones (compounds 43 and 44) isolated from the sea sponge Dysidea species, available in South China, were evaluated by several research groups [46,70,71] with regards to its in vitro PTP1B inhibitory activity. These sesquiterpenes exhibit positive PTP1B inhibitory activity, with IC50 values of 6.7 μM and 9.98 μM, respectively (Table 6). Dehydroeuryspongin A (compound 45), another sesquiterpene isolated from the marine sponge Euryspongia species, also displayed positive activity with an IC50 value of 3.6 μM [72].
Diterpenes
Diterpenes isolated from Sarcophyton trocheliophorum Marenzeller, a Hainan soft coral, received pivotal attention from Liang et al. over two consecutive years [73,74]. The authors isolated three compounds (compounds 46–48), and assessed their in vitro PTP1B inhibitory potential. These diterpenes showed variable effects, with IC50 values ranging from 6.8 μM to 27.1 μM.
Sesterterpenes
Piao et al. [75] evaluated the activity of two sesterterpenoids (compounds 49 and 50) from the sponge Hippospongia lachne found on Yongxing Island against PTP1B through an in vitro study, and found that they exhibited IC50 values of 5.2 μM and 8.7 μM, respectively.
Triterpenes
Compound 51, a triterpene isolated from the Antarctic lichen Lecidella carpathica, exhibited prominent in vitro anti-diabetic activity through PTP family enzymes inhibition [76]. Specifically, it inhibited PTP1B (IC50 = 3.7 μM) and T-cell protein tyrosine phosphatase (TCPTP) (IC50 = 8.4 μM) enzymes. On the other hand, it showed higher IC50 values (exceeding 68 μM) against the studied phosphatase enzymes, Src homology phosphatase-2 (SHP-2), leukocyte antigen-related phosphatase (LAR), and protein tyrosine phosphatase receptor type C (PTPRC), also known as CD45 antigen tyrosine phosphatase (Table 6).
Stellettin G (compound 52), is an isomalabaricane triterpene that was isolated from the Hainan sponge Stelletta species by Xue et al. [77]. This compound also displayed prominent in vitro PTP1B inhibitory action (IC50 = 4.1 μM). Indeed, isomalabaricanes are currently gaining significant interest as many of them have shown promissory in vitro cytotoxic effects [78].
3.1.7. Fungal Metabolites
In 2013, Sohn et al. and Lee et al. [79,80] investigated the fungal strains derived from marine resources (particularly from Penicillium and Eurotium species) and isolated seven compounds (compounds 53–59). Compounds 53–57 (Table 7) displayed moderate anti-diabetic effects as PTP1B inhibitors, with IC50 values varying from 10.7 µM to 64.0 µM, while compounds 58 and 59 showed much lower IC50 values (5.3 μM and 1.9 μM).
Aquastatin A (compound 60) is also a fungal metabolite isolated from Cosmospora species (Table 7) that has received particular attention from several researchers [81,82]. This attention is attributed not only to its low IC50 value against PTP1B enzyme (0.2 μM), but also due to its selective inhibitory activity against others PTPs, including TCPTP, SHP-2, LAR, and CD45.
3.1.8. Miscellaneous Compounds
Fucoxanthin, compound 61, is a carotenoid that was isolated from Phaeodactylum tricornutum and edible brown seaweeds, such as Eisenia bicyclis (Arame), Undaria pinnatifida (Wakame), and Hi-jikia fusiformis (Hijiki) (Table 8). Fucoxanthin was found to decrease blood glucose and insulin levels as well as water intake in a diabetic/obese KKAy mice model. A significant reduction in mRNA expression levels of monocyte chemoattractant protein-1 and tumor necrosis factor-α (TNF-α) was observed, which is believed to be involved in insulin resistance induction. A prominent PTP1B inhibitory action was also stated, with an IC50 value of 4.8 μM [83,84,85].
Seo et al. [76] isolated brialmontin 1 and atraric acid (compounds 62 and 63) from Lecidella carpathica, an Antarctic lichen. Both compounds were found to have PTP1B inhibitory action with IC50 values of 14 μM and 51.5 μM, respectively (Table 8).
In addition, the crude extracts from other seaweeds, including Derbesia marina, Symphyocladia latiuscula, Codium adhaerens, Attheya longicornis, Chaetoceros socialis, Chaetoceros furcellatus, Skeletonema marinoi, and Porosira glacialis along with the brown algae Hisikia fuziformis have been also evaluated for their in vitro PTP1B inhibitory capacity [86,87,88].
Furthermore, the ethanolic extract of the brown algae S. serratifolium C. Agardh exhibited broad PTP1B and α-glucosidase inhibitory activities (IC50 7.04 and 24.16 µg/mL, respectively) [89]. Among the four subfractions of the ethanol extract, n-hexane exhibited the highest activities (IC50 1.88 and 3.16 µg/mL, respectively), so its major three plastoquinones—sargahydroquinoic acid, sargachromenol, and sargaquinoic acid (compounds 64–66)—were isolated and the three compounds showed potent PTP1B inhibitory activity (Table 8). Sargachromenol also showed the most promising α-glucosidase inhibitory activity with an IC50 value of 42.41 μM, followed by sargaquinoic acid with an IC50 value of 96.17 ± 3.48 μM, while sargahydroquinoic acid was inactive.
3.2. PTP1B Inhibitory Activity: In Vivo Findings
With regards to the in vivo anti-diabetic activity of marine plants, Shi et al. [53] assessed the in vivo PTP1B inhibitory activity of highly brominated derivatives isolated from the red algae R. confervoides, which contains one or two 2,3-dibromo-4,5-dihydroxybenzyl units, in diabetic rats. They found that R. confervoides extracts led to a significant reduction in serum glucose levels. These in vivo results may confirm that the anti-hyperglycemic activity of R. confervoides is partially attributable to the PTP1B inhibition activity of its constituents [53]. Similarly, Nuño et al. [90] investigated the anti-diabetic activities of the microalgae haptophyte Isochrysis galbana and the ochrophyte Nannochloropsis oculata in a diabetic rat model. Different biochemical parameters were investigated, such as glucose level, body weight, lipoproteins, and nitrogenous compounds. In addition, gastrointestinal (GI) histopathology was studied. Both microalgae studied led to an increase in low-density lipoprotein (LDL) and a decrease in high-density lipoprotein (HDL) levels in both control and diabetic rats. More specifically, I. galbana decreased body weight, glucose, triacylglycerides, and cholesterol levels and exhibited just slight signs of inflammation in the gut. The observed activity could be attributed to their high content in docosahexaenoic (DHA) and eicosapentaenoic (EPA) fatty acids. The N. oculata-treated diabetic group did not show any changes in clinical values and had negative effects within the GI tract. Further studies are needed to confirm the effective employment of I. galbana as an anti-diabetic functional food.
4. Current Trends and Upcoming Challenges
4.1. In Vitro and In Vivo Concerns
In overall, T2DM is a metabolic disease characterized by hyperglycemia and hyperinsulinemia, in which the most common risk factor is overweight or obesity [91]. The development of diabetes may occur as a result of insulin secretion and/or signaling deregulation by insulin receptors (IR) [92]. The action of PTPs on IR themselves or their substrates is an important mechanism in insulin signaling regulation [92]. The role of PTPs in insulin signaling pathways and diabetes has previously been studied using vanadium compounds, which are able to reduce serum glucose levels in both type 1 and type 2 diabetic animal models [93,94]. Vanadium compounds show fundamental in vitro and in vivo insulinomimetic effects. Thus, the oral administration of such compounds promotes the normalization of serum glucose levels in T2DM rats, increasing glucose uptake [95]. These rats exhibited increased levels of hepatic cytosolic PTP activity, which decreased following insulin and vanadate treatment, leading to serum glucose levels normalization. These findings can be explained through PTPs inhibition with consequent improvement of cellular tyrosine phosphorylation [96].
Besides IR recognition, structural studies on PTP1B enzymes led to the identification of JAK2 and tyrosine kinase 2 (TYK2) as potential PTP1B substrates. Following interferon stimulation, both kinases were found to be hyperphosphorylated in PTP1B null fibroblasts [97]. This finding was further confirmed by the negative regulation of leptin-stimulated JAK2 phosphorylation produced by PTP1B, which reduced leptin signaling in in vivo models. Thus, null PTP1B mutation was introduced into leptin-deficient obese ob/ob mice, and a significant decrease in weight gain with an increase in resting metabolic rates was found in PTP1B-deficient ob/ob mice. Moreover, fat pads analysis proposed that the weight variations could be attributed to a decrease in adipose tissue. So, in the absence of leptin, PTP1B loss can reduce weight gain without modifying food intake [98,99]. Moreover, PTP1B-deficient mice had an increased response to weight loss caused by leptin and feeding suppression. The hypothalami of these mice evidenced a marked improvement in leptin-induced Transcription Factor STAT3 phosphorylation, indicating that the introduction of exogenous leptin in PTP1B deficiency will also led to leptin sensitivity enhancement [98,99]. Actually, substrate trapping trials using catalytically inactive PTP1B D181A confirmed that leptin-activated JAK2 is considered a PTP1B substrate, and that leptin signaling reduction is an obesity resistance mechanism in PTP1B null mice.
4.2. Human Concerns
In humans, weight loss and improved insulin sensitivity are closely related to decreased PTP activity, along with LAR and PTP1B expression in adipose tissue [100]. It is noticeable that PTP1B activity is not always related to its level of expression.
In obese and diabetic subjects, PTP1B protein levels show a 3- to 5-fold increase in abdominal adipose tissue, while a notable decrease in PTP1B activity was observed per unit of PTP1B protein [101]. It was also observed that total cellular PTP, not increased PTP1B activity, led to a marked raise in adipose tissue in obese individuals. In addition, it has been reported that an increase in PTP activity, but not in PTP1B activity, is accompanied by reduced insulin-stimulated glucose transport, proposing a tissue-specific role in glucose homeostasis for PTP1B [102].
On the other hand, genetic evidence also links PTP1B to diabetes and obesity in humans. Indeed, PTP1B locus maps to chromosome 20 in the region q13.1–q13.2 [103], which is a region recognized as a quantitative trait locus linked to insulin and obesity. A correlation between the role of PTP1B in insulin resistance and various polymorphisms has also been reported. That is, there is a continuing need to identify new PTP inhibitors for diabetes and obesity control.
4.3. Concerns in Culture Conditions
The anti-diabetic activity of several microalgae cultured under stressful conditions was also assessed using PTP1B assay [86,87]. Ingebrigtsen et al. [86] evaluated the non-polar fraction of five diatoms isolated from the North Atlantic (i.e., Attheya longicornis, Chaetoceros socialis, Chaetoceros furcellatus, Skeletonema marinoi, and Porosira glacialis), grown under variable light/temperature conditions. A. longicornis and C. furcellatus extracts displayed anti-PTP1B activity. On the other hand, C. socialis showed activity only when grown under high temperature—low light conditions, whereas P. glacialis showed activity when cultivated only under high temperature—high light conditions. Still, S. marinoi showed no activity in any of the studied conditions. Thus, these findings highlight the importance of culture conditions in activating bioactive metabolites production.
Meanwhile, Lauritano et al. [87] screened a total of 32 crude extracts from microalgal species (four flagellates, seven dinoflagellates, and 21 diatoms) grown under variable culturing conditions. It was found that temperature/light stresses are more important than nutritional stress in microalgal species containing bioactive metabolites that have PTP1B enzyme inhibition.
5. Conclusions
In general, the data collected emphasize the importance of chemical constituents from different marine species, given their PTP1B inhibition activity, as key targets in T2DM and obesity management.
Although distinct anti-diabetic therapeutic strategies are currently available, there is still an urgent need to find more effective and less toxic pharmacological agents. PTP1B has been established as a promising molecular target for the treatment of both T2DM and its major risk factor, obesity. However, PTP1B inhibitors still present significant problems over the closely related enzymes belonging to the PTP family, due to their poor selectivity, which is the main point to overcome their adverse effects.
As summarized herein, marine metabolites have recently gained significant attention from the scientific community, as they are considered to represent a repository of diverse unexploited bioactivities and structural features that could broaden the chemical library and may provide potential targets for the discovery of novel PTP1B inhibitory agents. However, another challenge must be kept in mind, arising from the limited yields of marine metabolites, which hinder their assessment through in vivo studies. In addition, it should be noted that current studies mainly focus on marine metabolites isolation and characterization as PTP1B inhibitors. However, the underlying mechanisms of action and structure–activity relationships need more attention. Thus, intensive efforts should be made through high-throughput screening of marine metabolites, along with structural optimization and synthesis of new PTP1B inhibitors, in order to identify selective, safer, and more effective PTP1B inhibitory agents in the near future.
Acknowledgments
N. Martins would like to thank the Portuguese Foundation for Science and Technology (FCT–Portugal) for the Strategic project, reference numbers UID/BIM/04293/2013 and “NORTE2020-Programa Operacional Regional do Norte” (NORTE-01-0145-FEDER-000012).
Author Contributions
All authors (S.M.E., M.H.E., S.H., B.S., M.S.-R., N. M., and J. S.-R.) contributed equally to this work. All the authors read and approved the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 1.Al-Lawati J.A. Diabetes Mellitus: A Local and Global Public Health Emergency! Om. Med. J. 2017;32:177–179. doi: 10.5001/omj.2017.34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Xiao J.B., Högger P. Dietary polyphenols and type 2 diabetes: Current insights and future perspectives. Curr. Med. Chem. 2015;22:23–38. doi: 10.2174/0929867321666140706130807. [DOI] [PubMed] [Google Scholar]
- 3.Cho N.H., Shaw J.E., Karuranga S., Huang Y., da Rocha Fernandes J.D., Ohlrogge A.W., Malanda B. IDF Diabetes Atlas: Global estimates of diabetes prevalence for 2017 and projections for 2045. Diabetes Res. Clin. Pract. 2018;138:271–281. doi: 10.1016/j.diabres.2018.02.023. [DOI] [PubMed] [Google Scholar]
- 4.American Diabetes Association Classification and diagnosis of diabetes. Diabetes Care. 2015;40:S11–S24. doi: 10.2337/dc17-S005. [DOI] [PubMed] [Google Scholar]
- 5.Dubé N., Tremblay M.L. Involvement of the small protein tyrosine phosphatases TC-PTP and PTP1B in signal transduction and diseases: From diabetes, obesity to cell cycle, and cancer. Biochim. Biophys. Acta. 2005;1754:108–117. doi: 10.1016/j.bbapap.2005.07.030. [DOI] [PubMed] [Google Scholar]
- 6.Brownlee M. Biochemistry and molecular cell biology of diabetic complications. Nature. 2001;414:813–820. doi: 10.1038/414813a. [DOI] [PubMed] [Google Scholar]
- 7.Lee S.H., Min K.H., Han J.S., Lee D.H., Park D.B., Jung W.K., Park P.J., Jeon B.T., Kim S.K., Jeon Y.J. Effects of brown alga, Ecklonia cava on glucose and lipid metabolism in C57BL/KsJ- db/db mice, a model of type 2 diabetes mellitus. Food Chem. Toxicol. 2012;50:575–582. doi: 10.1016/j.fct.2011.12.032. [DOI] [PubMed] [Google Scholar]
- 8.Pontiroli A.E. Type 2 diabetes mellitus is becoming the most common type of diabetes in school children. Acta Diabetol. 2004;41:85–90. doi: 10.1007/s00592-004-0149-8. [DOI] [PubMed] [Google Scholar]
- 9.Thilagam E., Parimaladevi B., Kumarappan C., Mandal S.C. α-Glucosidase and α-amylase inhibitory activity of Senna surattensis. J. Acupunct. Meridian Stud. 2013;6:24–30. doi: 10.1016/j.jams.2012.10.005. [DOI] [PubMed] [Google Scholar]
- 10.Jung M., Park M., Lee H.C., Kang Y.H., Kang E.S., Kim S.K. Antidiabetic agents from medicinal plants. Curr. Med. Chem. 2006;13:1203–1218. doi: 10.2174/092986706776360860. [DOI] [PubMed] [Google Scholar]
- 11.Ray S.D. A Worldwide Yearly Survey of New Data in Adverse Drug Reactions. Elsevier; Waltham, MA, USA: 2017. Side effects of drugs annual. [Google Scholar]
- 12.Scheen A.J., Lefebvre P.J. Troglitazone: Antihyperglycemic activity and potential role in the treatment of type 2 diabetes. Diabetes Care. 1999;22:1568–1577. doi: 10.2337/diacare.22.9.1568. [DOI] [PubMed] [Google Scholar]
- 13.Ahmad F., Li P.M., Meyerovitch J., Goldstein B.J. Osmotic loading of neutralizing antibodies demonstrates a role for protein-tyrosine phosphatase 1B in negative regulation of the insulin action pathway. J. Biol. Chem. 1995;270:20503–20508. doi: 10.1074/jbc.270.35.20503. [DOI] [PubMed] [Google Scholar]
- 14.Zinker B.A., Rondinone C.M., Trevillyan J.M., Gum R.J., Clampit J.E., Waring J.F., Xie N., Wilcox D., Jacobson P., Frost L., et al. PTP1B antisense oligonucleotide lowers PTP1B protein, normalizes blood glucose, and improves insulin sensitivity in diabetic mice. Proc. Nat. Acad. Sci. USA. 2002;99:11357–11362. doi: 10.1073/pnas.142298199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Johnson T.O., Ermolieff J., Jirousek M.R. Protein tyrosine phosphatase 1B inhibitors for diabetes. Nat. Rev. Drug Discov. 2002;1:696–709. doi: 10.1038/nrd895. [DOI] [PubMed] [Google Scholar]
- 16.Zhang S., Zhang Z.Y. PTP1B as a drug target: Recent developments in PTP1B inhibitor discovery. Drug Discov. Today. 2007;12:373–381. doi: 10.1016/j.drudis.2007.03.011. [DOI] [PubMed] [Google Scholar]
- 17.Zhang Z.Y., Lee S.Y. PTP1B inhibitors as potential therapeutics in the treatment of type 2 diabetes and obesity. Expert Opin. Investig. Drugs. 2003;12:223–233. doi: 10.1517/13543784.12.2.223. [DOI] [PubMed] [Google Scholar]
- 18.Bandyopadhyay D., Kusari A., Kenner K.A., Liu F., Chernoff J., Gustafson T.A., Kusari J. Protein-tyrosine phosphatase 1B complexes with the insulin receptor in vivo and is tyrosine-phosphorylated in the presence of insulin. J. Biol. Chem. 1997;272:1639–1645. doi: 10.1074/jbc.272.3.1639. [DOI] [PubMed] [Google Scholar]
- 19.Goldstein B.J., Bittner-Kowalczyk A., White M.F., Harbeck M. Tyrosine dephosphorylation and deactivation of insulin receptor substrate-1 by protein-tyrosine phosphatase 1B. Possible facilitation by the formation of a ternary complex with the Grb2 adaptor protein. J. Biol. Chem. 2000;275:4283–4289. doi: 10.1074/jbc.275.6.4283. [DOI] [PubMed] [Google Scholar]
- 20.Lund I.K., Hansen J.A., Andersen H.S., Moller N.P.H., Billestrup N. Andersen, Mechanism of protein tyrosine phosphatase 1B-mediated inhibition of leptin signalling. J. Mol. Endocrinol. 2005;34:339–351. doi: 10.1677/jme.1.01694. [DOI] [PubMed] [Google Scholar]
- 21.Byon J.C.H., Kusari A.B., Kusari J. Protein-tyrosine phosphatase-1B acts as a negative regulator of insulin signal transduction. Mol. Cell. Biochem. 1998;182:101–108. doi: 10.1023/A:1006868409841. [DOI] [PubMed] [Google Scholar]
- 22.Kenner K.A., Anyanwu E., Olefsky J.M., Kusari J. Protein-tyrosine phosphatase 1B is a negative regulator of insulin- and insulin-like growth factor-I-stimulated signaling. J. Biol. Chem. 1996;271:19810–19816. doi: 10.1074/jbc.271.33.19810. [DOI] [PubMed] [Google Scholar]
- 23.Meshkani R., Taghikhani M., Al-Kateb H., Larijani B., Khatami S., Sidiropoulos G.K., Hegele R.A., Adeli K. Polymorphisms within the protein tyrosine phosphatase 1B (PTPN1) gene promoter: Functional characterization and association with type 2 diabetes and related metabolic traits. Clin. Chem. 2007;53:1585–1592. doi: 10.1373/clinchem.2007.088146. [DOI] [PubMed] [Google Scholar]
- 24.Elchebly M., Payette P., Michaliszy E., Cromlish W., Collins S., Loy A.L., Normandin D., Cheng A., Himms-Hagen J., Chan C., et al. Increased insulin sensitivity and obesity resistance in mice lacking the protein tyrosine phosphatase-1B gene. Science. 1999;283:1544–1548. doi: 10.1126/science.283.5407.1544. [DOI] [PubMed] [Google Scholar]
- 25.Klaman L.D., Boss O., Peroni O.D., Kim J.K., Martino J.L., Zabolotny J.M., Moghal N., Lubkin M., Kim Y.B., Sharpe A.H., et al. Increased energy expenditure, decreased adiposity, and tissue-specific insulin sensitivity in proteintyrosine phosphatase 1B-deficient mice. Mol. Cell. Biol. 2000;20:5479–5489. doi: 10.1128/MCB.20.15.5479-5489.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bence K.K., Delibegovic M., Xue B., Gorgun C.Z., Hotamisligil G.S., Neel B.G., Kahn B.B. Neuronal PTP1B regulates body weight, adiposity and leptin action. Nat. Med. 2006;12:917–924. doi: 10.1038/nm1435. [DOI] [PubMed] [Google Scholar]
- 27.Zhao H., Liu G., Xin Z., Serby M.D., Pei Z., Szczepankiewicz B.G., Hajduk P.J., Abad-Zapatero C., Hutchins C.W., Lubben T.H., et al. Jirousek, Isoxazole carboxylic acids as protein tyrosine phosphatase 1B (PTP1B) inhibitors. Bioorgan. Med. Chem. Lett. 2004;14:5543–5546. doi: 10.1016/j.bmcl.2004.08.063. [DOI] [PubMed] [Google Scholar]
- 28.Leung K.W., Posner B.I., Just G. Periodinates: A new class of protein tyrosine phosphatase inhibitors. Bioorg. Med. Chem. Lett. 1999;9:353–356. doi: 10.1016/S0960-894X(98)00744-6. [DOI] [PubMed] [Google Scholar]
- 29.Andersen H.S., Olsen O.H., Iversen L.F., Sorensen A.L.P., Mortensen S.B., Christensen M., Branner S., Hansen T.K., Lau J.F., Jeppesen L., et al. Discovery and SAR of a novel selective and orally bioavailable nonpeptide classical competitive inhibitor class of protein-tyrosine phosphatase 1B. J. Med. Chem. 2002;45:4443–4459. doi: 10.1021/jm0209026. [DOI] [PubMed] [Google Scholar]
- 30.Wilson D.P., Wan Z.K., Xu W.X., Kirincich S.J., Follows B.C., Joseph-McCarthy D., Foreman K., Moretto A., Wu J., Zhu M., et al. Structure-based optimization of protein tyrosine phosphatase 1B inhibitors: From the active site to the second phosphotyrosine binding site. J. Med. Chem. 2007;50:4681–4698. doi: 10.1021/jm0702478. [DOI] [PubMed] [Google Scholar]
- 31.Liu S., Zeng L.F., Wu L., Yu X., Xue T., Gunawan A.M., Long Y.Q., Zhang Z.Y. Targeting inactive enzyme conformation: Aryl diketoacid derivatives as a new class of PTP1B inhibitors. J. Am. Chem. Soc. 2008;130:17075–17084. doi: 10.1021/ja8068177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lee S.H., Ko S.C., Kang M.C., Lee D.H., Jeon Y.J. Octaphlorethol a, a marine algae product, exhibits antidiabetic effects in type 2 diabetic mice by activating amp-activated protein kinase and upregulating the expression of glucose transporter 4. Food Chem. Toxicol. 2016;91:58–64. doi: 10.1016/j.fct.2016.02.022. [DOI] [PubMed] [Google Scholar]
- 33.Manikkam V., Vasiljevic T., Donkor O.N., Mathai M.L. A review of potential marine-derived hypotensive and anti-obesity peptides. Crit. Rev. Food Sci. Nutr. 2016;56:92–112. doi: 10.1080/10408398.2012.753866. [DOI] [PubMed] [Google Scholar]
- 34.Ruocco N., Costantini S., Guariniello S., Costantini M. Polysaccharides from the marine environment with pharmacological, cosmeceutical and nutraceutical potential. Molecules. 2016;21:551. doi: 10.3390/molecules21050551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Saleh A.S.M., Zhang Q., Shen Q. Recent research in antihypertensive activity of food protein-derived hydrolyzates and peptides. Crit. Rev. Food Sci. Nutr. 2016;56:760–787. doi: 10.1080/10408398.2012.724478. [DOI] [PubMed] [Google Scholar]
- 36.Suleria H.Á.R., Gobe G., Masci P., Osborne S.A. Marine bioactive compounds and health promoting perspectives; innovation pathways for drug discovery. Trends Food Sci. Technol. 2016;50:44–55. doi: 10.1016/j.tifs.2016.01.019. [DOI] [Google Scholar]
- 37.Choochote W., Suklampoo L., Ochaikul D. Evaluation of antioxidant capacities of green microalgae. J. Appl. Phycol. 2014;26:43–48. doi: 10.1007/s10811-013-0084-6. [DOI] [Google Scholar]
- 38.Zhao C., Wu Y.J., Yang C.F., Liu B., Huang Y.F. Hypotensive, hypoglycemic and hypolipidemic effects of bioactive compounds from microalgae and marine microorganisms. Int. J. Food Sci. Technol. 2015;50:1705–1717. doi: 10.1111/ijfs.12860. [DOI] [Google Scholar]
- 39.Khan W., Rayirath U.P., Subramanian S., Jithesh M.N., Rayorath P., Hodges D.M., Critchley A.T., Craigie J.S., Norrie J., Prithiviraj B. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 2009;28:386–399. doi: 10.1007/s00344-009-9103-x. [DOI] [Google Scholar]
- 40.Pangestuti R., Kim S.K. Biological activities and health benefit effects of natural pigments derived from marine algae. J. Funct. Foods. 2011;3:255–266. doi: 10.1016/j.jff.2011.07.001. [DOI] [Google Scholar]
- 41.Mohamed S., Hashim S.N., Rahman H.A. Seaweeds: A sustainable functional food for complementary and alternative therapy. Trends Food Sci. Technol. 2012;23:83–96. doi: 10.1016/j.tifs.2011.09.001. [DOI] [Google Scholar]
- 42.Gupta S., Abu-Ghannam N. Bioactive potential and possible health effects of edible brown seaweeds. Trends Food Sci. Technol. 2011;22:315–326. doi: 10.1016/j.tifs.2011.03.011. [DOI] [Google Scholar]
- 43.Jiang C.S., Liang L.F., Guo Y.W. Natural products possessing protein tyrosine phosphatase 1B (PTP1B) inhibitory activity found in the last decades. Acta Pharmacol. Sin. 2012;33:1217–1245. doi: 10.1038/aps.2012.90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Blunt J.W., Copp B.R., Hu W.P., Munro M.H., Northcote P.T., Prinsep M.R. Marine natural products. Nat. Prod. Rep. 2009;26:170–244. doi: 10.1039/b805113p. [DOI] [PubMed] [Google Scholar]
- 45.Yamazaki H., Sumilat D.A., Kanno S., Ukai K., Rotinsulu H., Wewengkang D.S., Ishikawa M., Mangindaan R.E.P., Namikoshi M. A polybromodiphenyl ether from an Indonesian marine sponge Lamellodysidea herbacea and its chemical derivatives inhibit protein tyrosine phosphatase 1B, an important target for diabetes treatment. J. Nat. Med. 2013;67:730–735. doi: 10.1007/s11418-012-0735-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Li Y., Zhang Y., Shen X., Guo Y.W. A novel sesquiterpene quinone from Hainan sponge Dysidea villosa. Bioorgan. Med. Chem. Lett. 2009;19:390–392. doi: 10.1016/j.bmcl.2008.11.068. [DOI] [PubMed] [Google Scholar]
- 47.Kurihara H., Mitain T., Kawabata J., Takahashi K. Inhibitory potencies of bromophenols from Rhodomelaceae algae against α-glucosidase activity. Fish Sci. 1999;65:300–303. doi: 10.2331/fishsci.65.300. [DOI] [Google Scholar]
- 48.Kurihara H., Mitain T., Kawabata J., Takahashi K. Two new bromophenols from the red alga Odonthalia corymbifera. J. Nat. Prod. 1999;62:882–884. doi: 10.1021/np980324p. [DOI] [PubMed] [Google Scholar]
- 49.Se-Kwon K., Steve T. Marine Medicinal Foods: Implications and Applications, Macro and Microalgae. Academic Press; Waltham, MA, USA: 2011. [Google Scholar]
- 50.Shi D., Xu F., Li J., Guo S.J., Su H., Han L.J. PTP1B inhibitory activities of bromophenol derivatives from algae. Chin. J. Chin. Mater. Med. 2008;33:2238–2240. [PubMed] [Google Scholar]
- 51.Shi D., Guo S., Jiang B., Guo C., Wang T., Zhang L., Li J. HPN, a synthetic analogue of bromophenol from red alga Rhodomela confervoides: Synthesis and anti-diabetic effects in C57BL/KsJ-db/db mice. Mar. Drugs. 2013;11:350–362. doi: 10.3390/md11020350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Shi D., Li J., Jiang B., Guo S., Su H., Wang T. Bromophenols as inhibitors of protein tyrosine phosphatase 1B with antidiabetic properties. Bioorgan. Med. Chem. Lett. 2012;22:2827–2832. doi: 10.1016/j.bmcl.2012.02.074. [DOI] [PubMed] [Google Scholar]
- 53.Shi D., Feng X., He J., Li J., Fan X., Han L. Inhibition of bromophenols against PTP1B and anti-hyperglycemic effect of Rhodomela confervoides extract in diabetic rats. Chin. Sci. Bull. 2008;53:2476–2479. doi: 10.1007/s11434-008-0353-y. [DOI] [Google Scholar]
- 54.Shoeib N.A., Bibby M.C., Blunden G., Linley P.A., Swaine D.J., Wheelhouse R.T., Wright C.W. In-vitro cytotoxic activities of the major bromophenols of the red alga Polysiphonia lanosa and some novel synthetic isomers. J. Nat. Prod. 2004;67:1445–1449. doi: 10.1021/np0305268. [DOI] [PubMed] [Google Scholar]
- 55.Liu X., Li X., Gao L., Cui C., Li C., Li J., Wang B. Extraction and PTP1B inhibitory activity of bromophenols from the marine red alga Symphyocladia latiuscula. Chin. J. Oceanol. Limnol. 2011;29:686–690. doi: 10.1007/s00343-011-0136-1. [DOI] [Google Scholar]
- 56.Kim K.Y., Nam K.A., Kurihara H., Kim S.M. Potent alpha-glucosidase inhibitors purified from the red alga Grateloupia elliptica. Phytochemistry. 2008;69:2820–2825. doi: 10.1016/j.phytochem.2008.09.007. [DOI] [PubMed] [Google Scholar]
- 57.Kim K.Y., Nam K.A., Kurihara H., Kim S.M. Alpha-glucosidase inhibitory activity of bromophenol purified from the red alga Polyopes lancifolia. J. Food Sci. 2010;75:H145–H150. doi: 10.1111/j.1750-3841.2010.01629.x. [DOI] [PubMed] [Google Scholar]
- 58.Suzen S., Buyukbingol E. Recent studies of aldose reductase enzyme inhibition for diabetic complications. Curr. Med. Chem. 2003;10:1329–1352. doi: 10.2174/0929867033457377. [DOI] [PubMed] [Google Scholar]
- 59.Wang W., Okada Y., Shi H., Wang Y., Okuyama T. Structures and aldose reductase inhibitory effects of bromophenols from the red alga Symphyocladia latiuscula. J. Nat. Prod. 2005;68:620–622. doi: 10.1021/np040199j. [DOI] [PubMed] [Google Scholar]
- 60.Qin J., Su H., Zhang Y., Gao J., Zhu L., Wu X., Pan H., Li X. Highly brominated metabolites from marine red alga Laurencia similis inhibit protein tyrosine phosphatase 1B. Bioorgan. Med. Chem. Lett. 2010;20:7152–7154. doi: 10.1016/j.bmcl.2010.08.144. [DOI] [PubMed] [Google Scholar]
- 61.Faulkner D. Marine Natural Product Chemistry. Springer; New York, NY, USA: 1977. [Google Scholar]
- 62.Targett N.M., Arnold T.M. Predicting the effects of brown algal phlorotannins on marine herbivores in tropical and temperate oceans. J. Phycol. 1998;34:195–205. doi: 10.1046/j.1529-8817.1998.340195.x. [DOI] [Google Scholar]
- 63.Riitta K. Brown Algal Phlorotannins: Improving and Applying Chemical Methods. University of Turku; Turku, Finland: 2008. [Google Scholar]
- 64.Jung H.A., Yoon N.Y., Woo M.H., Choi J.S. Inhibitory activities of extracts from several kinds of seaweeds and phlorotannins from the brown alga Ecklonia stolonifera on glucose-mediated protein damage and rat lens aldose reductase. Fish. Sci. 2008;74:1363–1365. doi: 10.1111/j.1444-2906.2008.01670.x. [DOI] [Google Scholar]
- 65.Moon H.E., Islam N., Ahn B.R., Chowdhury S.S., Sohn H.S., Jung H.A., Choi J.S. Protein tyrosine phosphatase 1B and α-glucosidase inhibitory Phlorotannins from edible brown algae, Ecklonia stolonifera and Eisenia bicyclis. Biosci. Biotechnol. Biochem. 2011;75:1472–1480. doi: 10.1271/bbb.110137. [DOI] [PubMed] [Google Scholar]
- 66.Okada Y., Ishimaru A., Suzuki R., Okuyama T. A new phloroglucinol derivative from the brown alga Eisenia bicyclis: Potential for the effective treatment of diabetic complications. J. Nat. Prod. 2004;67:103–105. doi: 10.1021/np030323j. [DOI] [PubMed] [Google Scholar]
- 67.Abdul Q.A., Choi R.J., Jung H.A., Choi J.S. Health benefit of fucosterol from marine algae: A review. J. Sci. Food Agric. 2016;96:1856–1866. doi: 10.1002/jsfa.7489. [DOI] [PubMed] [Google Scholar]
- 68.Zhou X., Sun J., Ma W., Fang W., Chen Z., Yang B., Liu Y. Bioactivities of six sterols isolated from marine invertebrates. Pharm. Biol. 2014;52:187–190. doi: 10.3109/13880209.2013.821664. [DOI] [PubMed] [Google Scholar]
- 69.Jung H.A., Islam M.N., Lee C.M., Oh S.H., Lee S., Jung J.H., Choi J.S. Kinetics and molecular docking studies of an anti-diabetic complication inhibitor fucosterol from edible brown algae Eisenia bicyclis and Ecklonia stolonifera. Chemico-Biol. Interact. 2013;206:55–62. doi: 10.1016/j.cbi.2013.08.013. [DOI] [PubMed] [Google Scholar]
- 70.Jiao W.H., Huang X.J., Yang J.S., Yang F., Piao S.J., Gao H., Li J., Ye W.C., Yao X.S., Chen W.S., et al. Dysidavarones A-D, new sesquiterpene quinones from the marine sponge Dysidea avara. Org. Lett. 2012;14:202–205. doi: 10.1021/ol202994c. [DOI] [PubMed] [Google Scholar]
- 71.Zhang Y.L., Guo Y.W., Jiang H.L., Shen X. A sesquiterpene quinone, dysidine, from the sponge Dysidea villosa, activates the insulin pathway through inhibition of PTPases. Acta Pharm. Sin. 2009;30:333–345. doi: 10.1038/aps.2009.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Yamazaki H., Nakazawa T., Sumilat D.A., Takahashi O., Ukai K., Takahashi S., Namikoshi M. Euryspongins A–C, three new unique sesquiterpenes from a marine sponge Euryspongia sp. Bioorgan. Med. Chem. Lett. 2013;23:2151–2154. doi: 10.1016/j.bmcl.2013.01.102. [DOI] [PubMed] [Google Scholar]
- 73.Liang L., Kurtán T., Mándi A., Gao L.X., Li J., Zhang W., Guo Y.W. Sarsolenane and Capnosane Diterpenes from the Hainan Soft Coral Sarcophyton trocheliophorum Marenzeller as PTP1B Inhibitors. Eur. J. Org. Chem. 2014:1841–1847. doi: 10.1002/ejoc.201301683. [DOI] [Google Scholar]
- 74.Liang L.F., Gao L.X., Li J., Taglialatela-Scafati O., Guo Y.W. Cembrane diterpenoids from the soft coral Sarcophyton trocheliophorum Marenzeller as a new class of PTP1B inhibitors. Bioorg. Med. Chem. Lett. 2013;21:5076–5080. doi: 10.1016/j.bmc.2013.06.043. [DOI] [PubMed] [Google Scholar]
- 75.Piao S.J., Jiao W.H., Yang F., Yi Y.H., Di Y.T., Han B.N., Lin H.W. New Hippolide Derivatives with Protein Tyrosine Phosphatase 1B Inhibitory Activity from the Marine Sponge Hippospongia lachne. Mar. Drugs. 2014;12:4096–4109. doi: 10.3390/md12074096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Seo C., HanYim J., Lee H.K., Oh H. PTP1B inhibitory secondary metabolites from the Antarctic lichen Lecidella carpathica. Mycology. 2011;2:18–23. doi: 10.1080/21501203.2011.554906. [DOI] [Google Scholar]
- 77.Xue D.G., Mao S.C., Yu X.Q., Guo Y.W. Isomalabaricane triterpenes with potent protein-tyrosine phosphatase 1B (PTP1B) inhibition from the Hainan sponge Stelletta sp. Biochem. Syst. Ecol. 2013;49:101–106. doi: 10.1016/j.bse.2013.03.001. [DOI] [Google Scholar]
- 78.Fouad M., Edrada R.A., Ebel R., Wray V., Müller W.E.G., Lin W.H., Proksch P. Cytotoxic Isomalabaricane Triterpenes from the Marine Sponge Rhabdastrella globostellata. J. Nat. Prod. 2006;69:211–218. doi: 10.1021/np050346t. [DOI] [PubMed] [Google Scholar]
- 79.Lee D.S., Jang J.H., Ko W., Kim K.S., Sohn J.H., Kang M.S., Ahn J.S., Kim Y.C., Oh H. PTP1B inhibitory and anti-inflammatory effects of secondary metabolites isolated from the marine-derived fungus Penicillium sp. JF-55. Mar. Drugs. 2013;11:1409–1426. doi: 10.3390/md11041409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Sohn J.H., Lee Y.R., Lee D.S., Kim Y.C., Oh H. PTP1B inhibitory secondary metabolites from marine-derived fungal strains Penicillium spp. and Eurotium sp. J. Microbiol. Biotechnol. 2013;23:1206–1211. doi: 10.4014/jmb.1303.03078. [DOI] [PubMed] [Google Scholar]
- 81.Debbab A., Aly A.H., Lin W.H., Proksch P. Bioactive compounds from marine bacteria and fungi. Microb. Biotechnol. 2010;3:544–563. doi: 10.1111/j.1751-7915.2010.00179.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Seo C., Sohn J.H., Oh H., Kim B.Y., Ahn J.S. Isolation of the protein tyrosine phosphatase 1B inhibitory metabolite from the marine-derived fungus Cosmospora sp. SF-5060. Bioorgan. Med. Chem. Lett. 2009;19:6095–6097. doi: 10.1016/j.bmcl.2009.09.025. [DOI] [PubMed] [Google Scholar]
- 83.Jung H.A., Islam M.N., Lee C.M., Jeong H.O., Chung H.Y., Woo H.C., Choi J.S. Promising antidiabetic potential of fucoxanthin isolated from the edible brown algae Eisenia bicyclis and Undaria pinnatifida. Fish. Sci. 2012;78:1321–1329. doi: 10.1007/s12562-012-0552-y. [DOI] [Google Scholar]
- 84.Maeda H., Hosokawa M., Sashima T., Miyashita K. Dietary combination of fucoxanthin and fish oil attenuates the weight gain of white adipose tissue and decreases blood glucose in obese/diabetic KK-Ay mice. J. Agric. Food Chem. 2007;55:7701–7706. doi: 10.1021/jf071569n. [DOI] [PubMed] [Google Scholar]
- 85.Peng J., Yuan J.P., Wu C.F., Wang J.H. Fucoxanthin, a marine carotenoid present in brown seaweeds and diatoms: Metabolism and bioactivities relevant to human health. Mar. Drugs. 2011;9:1806–1828. doi: 10.3390/md9101806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Ingebrigtsen R.A., Hansen E., Andersen J.H., Eilertsen H.C. Light and temperature effects on bioactivity in diatoms. J. Appl. Phycol. 2015;28:939–950. doi: 10.1007/s10811-015-0631-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Lauritano C., Andersen J.H., Hansen E., Albrigtsen M., Escalera L., Esposito F., Helland K., Hanssen K.Ø., Romano G., Ianora A. Bioactivity screening of microalgae for antioxidant, anti-inflammatory, anticancer, anti-diabetes and antibacterial activities. Front. Mar. Sci. 2016 doi: 10.3389/fmars.2016.00068. [DOI] [Google Scholar]
- 88.Lee H.J., Kim Y.A., Lee J.I., Lee B.J., Seo Y.W. Screening of Korean marine plants extracts for inhibitory activity on protein tyrosine phosphatase 1B. J. Appl. Biol. Chem. 2007;50:74–77. [Google Scholar]
- 89.Ali M.Y., Kim D.H., Seong S.H., Kim H.R., Jung H.A., Choi J.S. α-Glucosidase and Protein Tyrosine Phosphatase 1B Inhibitory Activity of Plastoquinones from Marine Brown Alga Sargassum serratifolium. Mar. Drugs. 2017;15:368. doi: 10.3390/md15120368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Nuño K., Villarruel-Lopez A., Puebla-Pérez A.M., Romero-Velarde E., Puebla-Mora A.G., Ascencio F. Effects of the marine microalgae Isochrysis galbana and Nannochloropsis oculata in diabetic rats. J. Funct. Foods. 2013;5:106–115. doi: 10.1016/j.jff.2012.08.011. [DOI] [Google Scholar]
- 91.American Diabetes Association Diagnosis and classification of diabetes mellitus. Diabetes Care. 2009 doi: 10.2337/dc14-S081. [DOI] [Google Scholar]
- 92.Cheng A., Dube N., Gu F., Tremblay M.L. Coordinated action of protein tyrosine phosphatases in insulin signal transduction. Eur. J. Biochem. 2002;269:1050–1059. doi: 10.1046/j.0014-2956.2002.02756.x. [DOI] [PubMed] [Google Scholar]
- 93.Fantus I.G., Deragon G., Lai R., Tang S. Modulation of insulin action by vanadate: Evidence of a role for phosphotyrosine phosphatase activity to alter cellular signaling. Mol. Cell. Biochem. 1995;153:103–112. doi: 10.1007/BF01075924. [DOI] [PubMed] [Google Scholar]
- 94.Fantus I.G., Tsiani E. Multifunctional actions of vanadium compounds on insulin signaling pathways: Evidence for preferential enhancement of metabolic versus mitogenic effects. Mol. Cell. Biochem. 1998;182:109–119. doi: 10.1023/A:1006853426679. [DOI] [PubMed] [Google Scholar]
- 95.Meyerovitch J., Farfel Z., Sack J., Shechter Y. Characterization and mode of action, Oral administration of vanadate normalizes blood glucose levels in streptozotocin-treated rats. J. Biol. Chem. 1987;262:6658–6662. [PubMed] [Google Scholar]
- 96.Meyerovitch J., Backer J., Kahn C.R. Hepatic phosphotyrosine phosphatase activity and its alterations in diabetic rats. J. Clin. Investig. 1989;84:976–983. doi: 10.1172/JCI114261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Myers M.P., Andersen J.N., Cheng A., Tremblay M.L., Horvath C.M., Parisien J.P., Salmeen A., Barford D., Tonks N.K. TYK2 and JAK2 are substrates of protein-tyrosine phosphatase 1B. J. Biol. Chem. 2001;276:47771–47774. doi: 10.1074/jbc.C100583200. [DOI] [PubMed] [Google Scholar]
- 98.Cheng A., Uetani N., Simoncic P.D., Chaubey V.P., Lee-Loy A., McGlade C.J., Kennedy B.P., Tremblay M.L. Attenuation of leptin action and regulation of obesity by protein tyrosine phosphatase 1B. Dev. Cell. 2002;2:497–503. doi: 10.1016/S1534-5807(02)00149-1. [DOI] [PubMed] [Google Scholar]
- 99.Zabolotny J.M., Bence-Hanulec K.K., Stricker-Krongrad A., Haj F., Wang Y., Minokoshi Y., Kim Y.B., Elmquist J.K., Tartaglia L.A., Kahn B.B., et al. PTP1B regulates leptin signal transduction in vivo. Dev. Cell. 2002;2:489–495. doi: 10.1016/S1534-5807(02)00148-X. [DOI] [PubMed] [Google Scholar]
- 100.Ahmad F., Considine R.V., Bauer T.L., Ohannesian J.P., Marco C.C., Goldstein B.J. Improved sensitivity to insulin in obese subjects following weight loss is accompanied by reduced protein-tyrosine phosphatases in adipose tissue. Metabolism. 1997;46:1140–1145. doi: 10.1016/S0026-0495(97)90206-7. [DOI] [PubMed] [Google Scholar]
- 101.Cheung A., Kusari J., Jansen D., Bandyopadhyay D., Kusari A. Bryer-Ash M., Marked impairment of protein tyrosine phosphatase 1B activity in adipose tissue of obese subjects with and without type 2 diabetes mellitus. J. Lab. Clin. Med. 1999;134:115–123. doi: 10.1016/s0022-2143(99)90115-4. [DOI] [PubMed] [Google Scholar]
- 102.Wu X., Hardy V.E., Joseph J.I., Jabbour S., Mahadev K., Zhu L., Goldstein B.J. Protein-tyrosine phosphatase activity in human adipocytes is strongly correlated with insulin-stimulated glucose uptake and is a target of insulin-induced oxidative inhibition. Metabolism. 2003;52:705–712. doi: 10.1016/S0026-0495(03)00065-9. [DOI] [PubMed] [Google Scholar]
- 103.Brown-Shimer S., Johnson K.A., Lawrence J.B., Johnson C., Bruskin A., Green N.R., Hill D.E. Molecular cloning and chromosome mapping of the human gene encoding protein phosphotyrosyl phosphatase 1B. Proc. Natl. Acad. Sci. USA. 1990;87:5148–5152. doi: 10.1073/pnas.87.13.5148. [DOI] [PMC free article] [PubMed] [Google Scholar]