

Abstract
Background
Globally cryptosporidiosis is one of the commonest causes of mortality in children under 24 months old and may be associated with important longterm health effects. Whilst most strains of Cryptosporidium parvum are zoonotic, C. parvum IIc is almost certainly anthroponotic. The global distribution of this potentially important emerging infection is not clear.
Methods
We conducted a systematic review of papers identifying the subtype distribution of C. parvum infections globally. We searched PubMed and Scopus using the following key terms Cryptospor* AND parvum AND (genotyp* OR subtyp* OR gp60). Studies were eligible for inclusion if they had found C. parvum within their human study population and had subtyped some or all of these samples using standard gp60 subtyping. Pooled analyses of the proportion of strains being of the IIc subtype were determined using StatsDirect. Meta-regression analyses were run to determine any association between the relative prevalence of IIc and Gross Domestic Product, proportion of the population with access to improved drinking water and improved sanitation.
Results
From an initial 843 studies, 85 were included in further analysis. Cryptosporidium parvum IIc was found in 43 of these 85 studies. Across all studies the pooled estimate of relative prevalence of IIc was 19.0% (95% CI: 12.9–25.9%), but there was substantial heterogeneity. In a meta-regression analysis, the relative proportion of all C. parvum infections being IIc decreased as the percentage of the population with access to improved sanitation increased and was some 3.4 times higher in those studies focussing on HIV-positive indivduals.
Conclusions
The anthroponotic C. parvum IIc predominates primarily in lower-income countries with poor sanitation and in HIV-positive individuals. Given the apparent enhanced post-infectious virulence of the other main anthroponotic species of Cryptosporidium (C. hominis), it is important to learn about the impact of this subtype on human health.
Electronic supplementary material
The online version of this article (10.1186/s13071-018-3263-0) contains supplementary material, which is available to authorized users.
Keywords: Cryptosporidium parvum, Subtypes, Epidemiology, Sanitation
Background
Cryptosporidium spp. are enteric protozoan parasites found globally throughout the world. They are ubiquitous in the environment and capable of causing infection in both immunocompetent and immunocompromised humans as well as a wide variety of animals. Their ability to cause both sporadic episodes of disease as well as more far-reaching food and waterborne outbreaks is becoming increasingly recognised [1, 2]
Cryptosporidium spp. are now one of the major causes of mortality from an infectious disease in children under 24 months in low income countries and are associated with an increased risk of death in toddlers aged 12–23 months [3]. Even in children who survive, there is growing evidence of a link between cryptosporidiosis, childhood malnutrition and stunting in such countries [4]. It has been estimated that in 2015 1.3 million deaths worldwide were due to diarrhoeal disease, and Cryptosporidium spp. are the second commonest cause of death from diarrhoeal disease in children under five years, after Rotavirus [5]. However, whilst Rotavirus has an internationally available vaccine that is successfully reducing numbers of severe gastro-intestinal disease and death [6], Cryptosporidium spp. have neither a vaccine nor effective treatment to reduce its morbidity and mortality.
Although there have been reported cases of human infection from at least 17 species of Cryptosporidium, C. hominis and C. parvum are the two species that have been most associated with causing human disease [7].
Subtyping of Cryptosporidium species, specifically C. hominis and C. parvum, can provide clarity of mode of transmission in addition to being important epidemiological tools, especially in outbreak situations. At present, the most commonly used and accepted method of subtyping is through sequencing of the gp60 gene, a 60-kDa glycoprotein. The gp60 gene has a number of tandem repeats at the 5' end of the gene consisting of TCA, TCG or TCT [8] and also further variation in the non-repeat 3' end of the gene, enabling classification of C. hominis and C. parvum into subtype families on the basis of the number and type of trinucleotide repeat [9].
Cryptosporidium parvum IIc is of particular interest as it appears to be anthroponotic, i.e. host- restricted to humans, which is in direct contrast to most other subtype families of C. parvum, which cause disease in both humans and animals [8].
Cryptosporidium genotypes and isolates vary in their virulence, with over 25 putative virulence factors identified [10]. Hunter et al. [11] showed that C. hominis was associated with an increased risk of post-infectious sequelae compared with C. parvum [11] and Cama et al. [12] suggested C. hominis Ib is more pathogenic than other subtypes. However, the differences in virulence of C. parvum subtypes have not been systemically studied, and there are very little data available linking subtype to pathology. As existing evidence suggests anthroponotic Cryptosporidium spp. are more virulent than zoonotic Cryptosporidium spp, then as C. parvum IIc is transmitted anthroponotically like C. hominis, it may display enhanced virulence like C. hominis.
The worldwide distribution of C. parvum IIc has not been systematically studied or documented. Thus we aimed to identify all studies that had subtyped C. parvum using gp60 subtyping methods, to characterise the endemic worldwide distribution and proportion of C. parvum IIc and investigate how this differs throughout the world, in particular exploring potential links with gross domestic product (GDP), a measure of economic growth and often used as a proxy for standard of living, and sanitation, in order to provide clarity on proportion and distribution of anthroponotic C. parvum IIc.
Methods
Search strategy
PubMed and Scopus were searched up to and including 1st November 2016 using the following search strategy: Cryptospor* AND parvum AND (genotyp* OR subtyp* OR gp60) without restriction on language or study type. Review articles and book chapters were additionally reviewed for references which would fit the inclusion criteria.
Eligibility criteria
Studies were eligible for inclusion if they had found C. parvum within their study population and had subtyped some or all of these samples using the standard gp60 subtyping classification. The study population of interest was restricted to humans, thus studies which had subtyped C. parvum found in animals were excluded, but for studies that had included both animals and humans the subtyping data for humans only was included. Reviews and editorials were excluded. Studies which reported subtyping of outbreaks were excluded as the aim was to identify the endemic worldwide proportion of C. parvum IIc.
If studies had included data from a previous study, only one study was included to prevent duplication of data.
Data including country of study, population characteristics, subtypes found, number of samples of subtypes found and total number of C. parvum samples subtyped were extracted onto a datasheet by one of the researchers. Independently a second researcher reviewed the studies included in the search and their inclusion or exclusion in the final dataset.
Eligible studies were screened initially by title and abstract, and included if they met the inclusion criteria and full text retrieved. If it was unclear from the title and abstract whether the inclusion criteria were met, then the full paper was reviewed and decision made regarding inclusion or exclusion.
Statistical analysis
Proportion of C. parvum IIc in relation to total C. parvum was calculated using the number of samples of C. parvum IIc and the total C. parvum samples subtyped using gp60 subtyping classification. Forest plots and pooled prevalence estimations were performed using STATS Direct™. When investigating drivers of heterogeneity, negative binomial regression analyses were carried out using STATA™. Given the known association of cryptosporidiosis with drinking water associated outbreaks, the faecal oral transmission pathway and the anthroponotic nature of C. parvum IIc, we hypothesised that much of the heterogeneity between countries could be explained by variation in access to improved drinking water and sanitation. In constructing the regression analyses we took data on Gross Domestic capita per person, % of the population with access to improved sanitation and % of the population with access to an improved water supply. All three variables were taken from the World Bank World Development Indicators Archive [13] and for the year when the study was undertaken or published. Gross domestic product per capita (GDPpc) was expressed in USD for year 2005. The definitions of improved sanitation and improved water supply were as used by the World Bank which are themselves taken from the definitions of the WHO/UNICEF Joint Monitoring Programme [14]. All three country-specific variables were taken to be two years prior to the publication date to adjust for time of collection to publication. Given the marked skewedness of the GDP data we used the log10-transformed data. In addition, publication year and whether or not a focus of the study was Cryptosporidium infections in HIV-positive individuals were also included in the analysis. The regression analysis was run with all predictors individually and then all in a single model removing the least statistically significant until all variables were significant at the P < 0.2 level.
PRISMA guidelines were followed in the preparation of this manuscript (see PRISMA checklist in Additional file 1: Table S1).
Results
The PubMed search gave a total of 750 results, and the Scopus search revealed an additional 90 unique studies. Reviewing of references in review articles and book chapters identified three further studies with subtyping information on C. parvum, resulting in a total of 843 unique studies.
A total of 732 studies were excluded for failing to meet the inclusion criteria, mainly because they were not original studies with subtyping information or had only included animals or environmental samples. A further 23 studies were excluded as they were studies of outbreaks, as the aim was to characterise the endemic proportion of C. parvum IIc. Two further studies were excluded as they contained duplication of material presented in other already included studies. The final number of studies included was 85 (Fig. 1, Table 1).
Fig. 1.
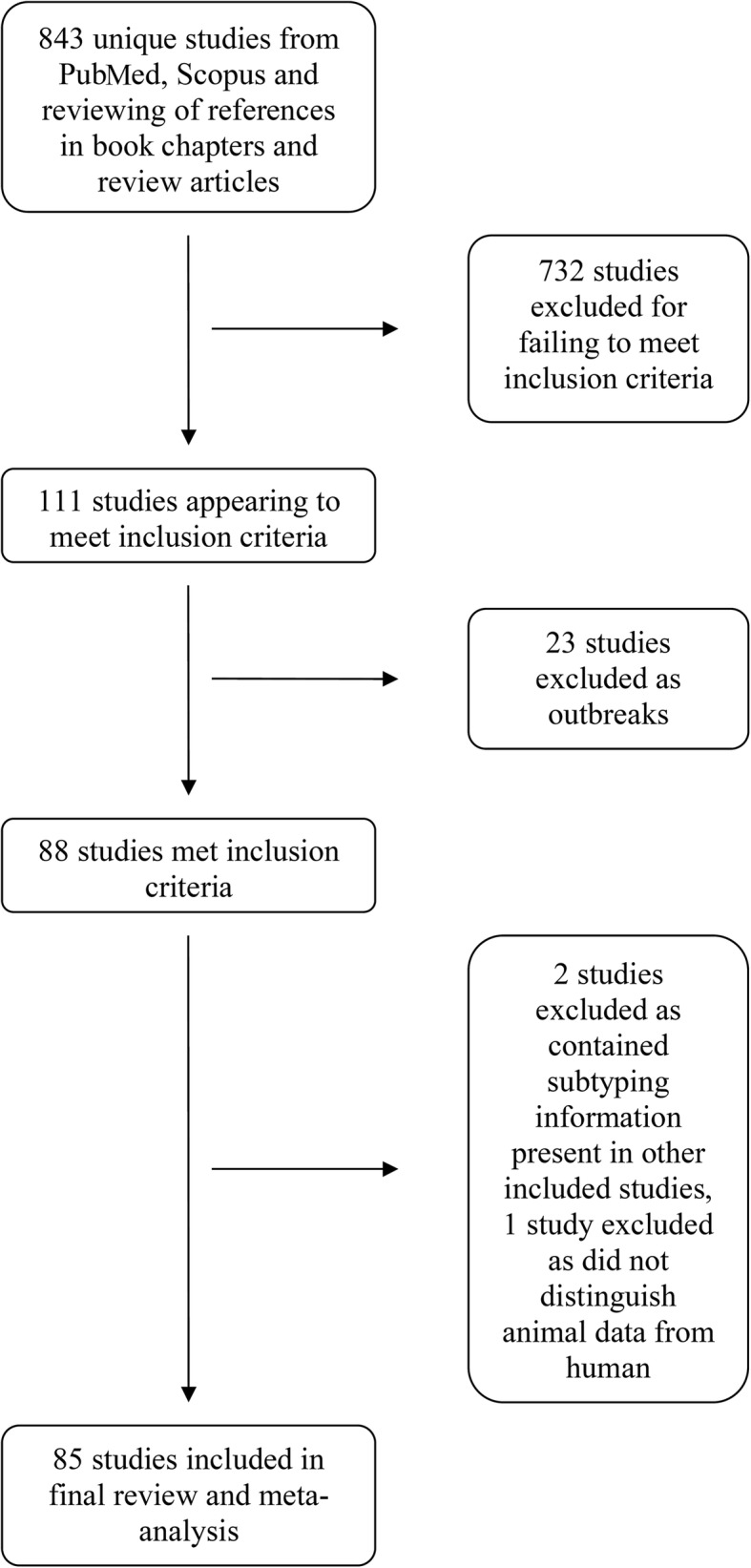
Flow chart depicting inclusion and exclusion of studies with numbers
Table 1.
Cryptosporidium parvum subtype frequency distributions in published literature ranked by continent and country.
| Reference | Country | Population | Total number C. parvum isolates subtyped and numbers of subtypes | Proportion C. parvum IIc of C. parvum subtyped (%) |
|---|---|---|---|---|
| Asia | ||||
| [48] | Bangladesh | Children | (n = 4) (4 × IIm) | 0 |
| [49] | Cambodia | Children | (n = 4) (4 × IIe ) | 0 |
| [50] | China | HIV+ patients | (n = 2) (2 × IId) | 0 |
| [51] | India | Adults and children | (n = 14) (6 × IIc; 5 × IId; 3 × IIe) | 43 |
| [52] | India | Children | (n = 6) (1 × IIc; 1 × IId; 2 × IIm; 2 × IIn) | 17 |
| [21] | India | Children | (n = 7) (7 × IIc) | 100 |
| [53] | India | HIV+ patients | (n = 9) (5 × IIb; 3 × IIc; 2 × IIg) | 33 |
| [54] | Iran | Children | (n = 15) (7 × IIa; 8 × IId) | 0 |
| [55] | Iran | Children | (n = 17) (6 × IIa; 11 × IId) | 0 |
| [56] | Iran | Children | (n = 22) (7 × IIa; 15 × IId) | 0 |
| [57] | Japan | Humans: not specified | (n = 2) (1 × IIa; 1 × IIc) | 50 |
| [58] | Japan | Humans: not specified | (n = 1) (1 × IIa) | 0 |
| [59] | Jordan | Adults and children | (n = 2) (1 × IIa; 1 × IId) | 0 |
| [60] | Jordan | Children | (n = 13) (3 × IIa; 2 × IIc; 8 × IId) | 15 |
| [61] | Kuwait | Children | (n = 61) (29 × IIa; 12 × IIc; 20 × IId) | 20 |
| [9] | Kuwait | Children | (n = 59) (27 × IIa; 2 × IIc; 29 × IId;1 × IIf) | 3 |
| [62] | Lebanon | Adults and children | (n = 5) (5 × IIa) | 0 |
| [63] | Malaysia | HIV+ patients | (n = 13) (12 × IIa; 1 × IId) | 0 |
| [64] | Malaysia | HIV+ patients | (n = 1) (1 × IId) | 0 |
| [65] | Yemen | Adults and children | (n = 7) (7 × IIa) | 0 |
| Africa | ||||
| [66] | Egypt | Adults and children | (n = 5) (5 × IId) | 0 |
| [67] | Egypt | Children | (n = 14) (7 × IIa; 7 × IId) | 0 |
| [17] | Equatorial Guinea | HIV+ patients | (n = 10) (7× IIc; 3 × IIe) | 70 |
| [68] | Ethiopia | HIV+ patients | (n = 82) (71 × IIa; 1 × IIb; 2 × IIc; 5 × IId; 1 × IIe; 2 × If-like 2) | 2 |
| [69] | Ethiopia | Adults and children | (n = 12) (12 × IIa) | 0 |
| [16] | Ghana | Children | (n = 37) (30 × IIc; 7 × IIe) | 81 |
| [15] | Kenya | Children | (n = 19) (19 × IIc) | 100 |
| [70] | Madagascar | Children | (n = 1) (1 × IIc) | 100 |
| [71] | Malawi | Children | (n = 2) (1 × IIc; 1 × IIe) | 50 |
| [72] | Nigeria | HIV+ patients | (n = 1) (1 × IIc) | 100 |
| [73] | Nigeria | HIV+ patients | (n = 1) (1 × IIc) | 100 |
| [74] | Nigeria | Adults and children | (n = 1) (1 × IIe) | 0 |
| [75] | Nigeria | Children | (n = 2) (2 × IIc) | 100 |
| [76] | Nigeria | HIV+ Adults | (n = 8) (4 × IIc; 4 × unspecified) | 50 |
| [18] | Nigeria | Children | (n = 23) (2 × IIa; 17 × IIc; 2 × IIi; 2 × IIm) | 74 |
| [77] | Sao Tome and Principe | Children | (n = 5) (2 × IIa; 3 × IId) | 0 |
| [78] | South Africa | Children | (n = 5) (1 × IIb; 3 × IIc; 1 × IIe) | 60 |
| [79] | South Africa | HIV + children | (n = 5) (5 × IIc) | 100 |
| [80] | Tunisia | Children | (n = 4) (2 × IIa; 2 × IId) | 0 |
| [22] | Uganda | Children | (n = 15) (10 × IIc; 1 × IIg; 1 × IIh; 3 × IIi 3) | 67 |
| Europe | ||||
| [81] | Belgium | Adults and children | (n = 6) (4 × IIa; 1 × IIc; 1 × IId) | 17 |
| [82] | Czech Republic | Adults | (n = 1) (1 × IIa) | 0 |
| [83] | Denmark | Adults and children | (n = 34?) (15 × IIa; Others not stated) | 0 |
| [24] | England & Wales, UK | Adults and children | (n = 66) (56 × IIa; 1 × IIc; 9 × IId) | 2 |
| [84] | Estonia | Human contact with calves | (n = 1) (1 × IIa) | 0 |
| [85] | France | Adults | (n = 1) (1 × IIa) | 0 |
| [86] | Ireland | Adults and children | (n = 249) (249 × IIa) | 0 |
| [87] | Ireland | Adults and children | (n = 79) (78 × IIa; 1 × IId) | 0 |
| [44] | Italy | AIDS patients | (n = 8) (4 × IIa; 4 × IIc) | 50 |
| [88] | Portugal | HIV+ patients | (n = 25) (9 × IIa; 1 × IIb; 7 × IIc; 8 × IId) | 28 |
| [89] | Romania | Children | (n = 4) (4 × IId) | 0 |
| [90] | Scotland | Adults and children | (n = 87) (82 × IIa; 2 × IIc; 2 × IId; 1 × IIg) | 2 |
| [91] | Slovak Republic | Humans: not specified | (n = 1) (1 × IIa) | 0 |
| [92] | Slovenia | Humans: not specified | (n = 31) (29 × IIa; 1 × IIc; 1 × IIl) | 3 |
| [93] | Spain | Adults and children | (n = 12) (7 × IIa; 1 × IIc; 4 × IId) | 8 |
| [94] | Spain | Adults and Children | (n = 7) (6 × IIa; 1 × IId) | 0 |
| [95] | Spain | Children | (n = 3) (3 × IIa) | 0 |
| [96] | Spain | Adults and children | (n = 163) (146 × IIa; 3 × IId; 14 × IIn) | 0 |
| [23] | Sweden | Adults and children | (n = 108) (89 × IIa; 12 × IIc; 24 × IId; 1 × IIe; 2 × IIo) | 11 |
| [25] | The Netherlands | Humans: not specified | (n = 13) (9 × IIa; 1 × IIc; 3 × IId) | 8 |
| [97] | UK | Humans: not specified | (n = 16) (11 × IIa; 1 × IIc; 3 × IId; 1 × IIe) | 6 |
| North America | ||||
| [98] | Canada | Humans: not specified | (n = 7) (7 × IIa) | 0 |
| [99] | Canada | Adults and children | (n = 5) (5 × IIa) | 0 |
| [100] | Canada | Humans: not specified | (n = 4) (4 × IIa) | 0 |
| [19] | Jamaica | HIV+ patients | (n = 7) (7 × IIc) | 100 |
| [101] | Mexico | Children | (n = 2) (2 × IIa) | 0 |
| [102] | USA | Humans: not specified | (n = 5) (5 × IIa) | 0 |
| [103] | USA | Humans: not specified | (n = 30) (30 × IIa) | 0 |
| South America | ||||
| [104] | Brazil, Argentina | Adults and Children | (n = 3) (2 × IIa(mixed with hominis); 1 × IIc) | 33 |
| [12] | Peru | Children | (n = 15) (15 × IIc) | 100 |
| [20] | Peru | HIV+ patients | (n = 22) (22 × IIc) | 100 |
| Australia/Oceania | ||||
| [28] | Australia | Humans | (n = 14) (14 × IIa) | 0 |
| [27] | Australia | Adults and children | (n = 21) (21 × IIa) | 0 |
| [29] | Australia | Farm workers | (n = 7) (5 × IIa; 1 × IId; 1 × IIa/IId) | 0 |
| [30] | Australia | Adults and children | (n = 80) (70 × IIa 79; 1 × IId) | 0 |
| [31] | Australia | Adults and children | (n = 49) (48 × IIa; 1 × IId) | 0 |
| [26] | Australia | Humans: not specified | (n = 32) (30 × IIa; 1 × IIc; 1 × IId) | 3 |
| [105] | Australia | Humans: not specified | (n = 24) (23 × IIa; 1 × IIc) | 4 |
| [32] | Australia | Humans: not specified | (n = 4) (4 × IIa) | 0 |
| [106] | Australia | Humans: not specified | (n = 23) (18 × IIa; 5 × IIc) | 22 |
| [33] | New Zealand | Humans: not specified | (n = 41) (41 × IIa) | 0 |
| Multiple continents | ||||
| [47] | Sub-Saharan Africa and Southeast Asia | Children | (n = 37) (21 × IIc; 13 × IIe; 3 × IId) | 57 |
| [107] | Iran, Malawi, Nigeria, UK, Vietnam | Children | (n = 9) (4 × IIa; 2 × IIc; 3 × IId) | 22 |
| [108] | Australia + Europe | Adults and children | (n = 11) (9 × IIa; 1 × IIc; 1 × IId) | 9 |
| [109] | China, Guatemala, India, Kenya, Portugal, Slovenia | Humans: not specified | (n = 13) (4 × IIa; 5 × IIb; 4 × IIc) | 31 |
Study characteristics
Of the 85 studies included, many (n = 28) were solely in children, 23 studies included both children and adults, and 14 studies focused on HIV positive or AIDS patients. Some studies did not specify the human population they were studying.
Studies had taken place in a wide variety of countries, including high- and low-income countries and both rural and urban settings.
The number of samples subtyped in most studies was small, ranging from 1 to 249 samples. Included studies were published between 2001 and 2016, with the majority of studies published in the later years, reflecting increased access to molecular techniques.
Cryptosporidium parvum IIc distribution
Cryptosporidium parvum IIc was found in 43 studies. In 10 studies, C. parvum IIc was the only subtype of C. parvum to be found. Across all studies the proportion of C. parvum strains typed as IIc using a random effects meta-analysis is 19.0% (95% CI, 12.9–25.9%). However, there was evidence of substantial heterogeneity [Cochran Q = 964.365229, df = 84, P < 0.0001; I2 (inconsistency) = 91.3% (95% CI: 90.1–92.3%)]. The estimates of the pooled proportions for all the GP60 subtypes are shown in Table 2, and Additional file 2: Figure S1 gives the forest plots for the three most common subtypes. It can be seen from Table 2 that the great majority of strains are either IIa, IIc or IId which together account for about 84% of human infections. The remaining 9 subtypes are only detected very rarely, with IIe representing an estimated 2.7% of infections.
Table 2.
Pooled proportion of GP60 subtypes of C. parvum
| Subtype | No. of studiesa (n = 85) |
Pooled proportion | 95% CI |
|---|---|---|---|
| IIa | 58 | 0.53 | 0.43–0.63 |
| IIb | 5 | 0.0097 | 0.0058–0.015 |
| IIc | 43 | 0.19 | 0.13–0.26 |
| IId | 36 | 0.12 | 0.082–0.16 |
| IIe | 11 | 0.027 | 0.016–0.039 |
| IIf | 2 | 0.0087 | 0.0050–0.013 |
| IIg | 3 | 0.0083 | 0.0048–0.013 |
| IIh | 1 | 0.0075 | 0.0042–0.012 |
| IIi | 3 | 0.0087 | 0.0050–0.013 |
| IIm | 3 | 0.0085 | 0.0049–0.013 |
| IIn | 2 | 0.012 | 0.0075–0.017 |
| IIo | 1 | 0.0083 | 0.0048–0.013 |
aNumber of studies with at least one of the subtypes
Abbreviation: CI confidence interval
Even from a simple visual inspection of the data, it was clear that C. parvum IIc was particularly common in middle- and low-income countries.
Mbae et al. [15] investigated the distribution of Cryptosporidium and diversity of subtypes in children in urban Kenya. Of the 19 samples of C. parvum they subtyped, all were the anthroponotic IIc subtype, with no other reported subtypes found. This predominant finding of high numbers of C. parvum IIc was also found in children in rural Ghana, with 81% of the subtyped C. parvum samples found to be the IIc subtype [16]. This finding was also replicated in HIV-positive patients in Equatorial Guinea [17], children in Nigeria [18], HIV-positive patients in Jamaica [19], both HIV-positive patients and children in Peru [12, 20] and children in both India and Uganda [21, 22].
In contrast, high income countries reported much lower numbers of C. parvum IIc as a proportion of total C. parvum subtyped. Often European studies did not find C. parvum IIc amongst their samples, or if they did it tended to be at low levels. Insulander et al. [23] studied adults and children in Sweden and found a C. parvum IIc proportion of just 11%. This finding was replicated by Chalmers et al. [24] studying adults and children in England and Wales who found C. parvum IIc in 2% of their C. parvum-subtyped samples, and Wielinga et al. [25] who reported a C. parvum IIc proportion of 8% from humans in the Netherlands. Studies in Australia and New Zealand tended to report either a very low proportion of C. parvum IIc, e.g. Waldron et al. [26] reported a proportion of 3%, or they did not find any C. parvum IIc within their samples [27–33].
In order to further investigate the possible drivers of the heterogeneity, we undertook regression analysis of all studies reporting data from a single country where we could allocate GDPpc, sanitation and water coverage data. The results of the analyses are shown in Table 3. It can be seen that in the single predictor analysis that increasing GDP, improved access to sanitation and water supply are all strongly associated with a reduced relative proportion of C. parvum IIc. Also, those studies that are focused primarily on people with HIV show a greater relative proportion of C. parvum IIc. In the final model both GDP and % access to improved water was dropped from the model leaving % access to improved sanitation, year of publication and whether the study focus was on people living with HIV. The % access to improved sanitation and focus on HIV was particularly strong. The relative proportion of C. parvum IIc declined by 3.3% (95% CI: 2.8–4.4%) for every 1% increase in national sanitation coverage. Similarly, the relative proportion of C. parvum IIc was 3.4 (95% CI: 1.4–8.2) times greater in studies focusing on people with HIV.
Table 3.
Negative binomial meta-regression analyses of proportion of C parvum that were IIc across 80 analysable studies (studies reporting data from a single country where GDPpc, sanitation and water coverage data could be allocated)
| Predictor | Single predictor analyses | Final model | ||||
|---|---|---|---|---|---|---|
| Proportion ratio | 95% CI | P-value | Proportion ratio | 95% CI | P-value | |
| Log10 GDP per capita US$2005 | 0.303 | 0.167–0.549 | 0.0001 | |||
| % population with access to improved sanitation | 0.969 | 0.955–0.983 | <0.0001 | 0.967 | 0.956–0.972 | <0.0001 |
| % population with access to improved drinking water | 0.943 | 0.914–0.975 | 0.0002 | |||
| Year of publication | 0.907 | 0.815–1.022 | 0.108 | 0.888 | 0.794–0.994 | 0.039 |
| Study focus on HIV-positive individuals | 3.263 | 1.096–9.717 | 0.034 | 3.414 | 1.428–8.162 | 0.006 |
Abbreviation: CI confidence interval
Additional file 2: Figure S1 indicates proportion of C. parvum IIc found in studies ordered in increasing sanitation coverage of country of study. Figure 2 shows the pooled relative prevalences for the three most common subtypes by quartile of proportion of the population with access to improved sanitation. Cryptosporidium parvum IIc is seen in a higher proportion in countries with low % sanitation coverage, and the proportion of C. parvum IIc seen in countries with high % sanitation coverage is much lower, or even none at all. This is in contrast to the subtype IIa, which appears to be seen at a higher proportion in countries with a higher % sanitation coverage, and subtype IId which shows a mixed picture but appears to cluster in Arabic countries. We did not include IIe in this analysis as the numbers were small - IIe was found in small numbers in 11 studies, of which 6 were in the lowest quartile for sanitation provision and one from a mix of countries most of which would have been in the lowest quartile. No IIe strains were reported from countries in the highest quartile.
Fig. 2.

Pooled relative prevalence of three most common C. parvum subtypes grouped by quartile of proportion of population with access to improved sanitation with Q1 representing the quartile with least access to improved sanitation
Discussion
This is the first systematic study, to our knowledge, investigating worldwide prevalence of C. parvum IIc and correlating this with GDP and sanitation data. We have illustrated a striking finding of high proportion of C. parvum IIc in low- and middle-income countries and extremely strong relationship between C. parvum IIc proportion and GDP and inadequate access to improved sanitation. This is especially pertinent when considering the World Health Organisation Millennium Development Goal of improved sanitation as many low-income countries have made little or no progress towards this goal [34].
The subtype C. parvum IIc is interesting as it is different to other C. parvum in that it is considered anthroponotic whereas other subtypes of C. parvum are zoonotic and tend to infect a wide range of animals in addition to humans. Cryptosporidium parvum IIc has never been found in livestock or pet animals, although there are three reports of a particular IIc subtype (IIcA5G3J) being found in hedgehogs [35–37]. However, these may not reflect true infection, rather ingestion from an environment faecally contaminated with C. parvum IIc from human waste. In contrast, C. hominis, the species of Cryptosporidium defined by its predominantly anthroponotic transmission characteristics, has been reported in livestock [7] and also recently in domestic dogs in Spain [38]. This review clearly demonstrates that it is anthroponotic C. parvum that is causing the majority of disease in low- and middle-income countries, rather than zoonotic C. parvum. This is in spite of the often close proximity people may have with animals in low-income settings, and is thus more likely related to the widespread faecal contamination of both food and water sources in these settings. This finding will have implications for public health and should influence measures to prevent infection and risks of ongoing transmission.
The strong association between C. parvum IIc and inadequate access to improved sanitation is worthy of comment. Given the prior association between outbreaks of cryptosporidiosis and waterborne disease [39], one would have expected the association to be strongest with inadequate access to drinking water. The fact that this is not the case needs to be explained. It is accepted that C. hominis is more common in low-income countries and that C. hominis is a human pathogen [40] (although it has rarely been reported in livestock [7] and in dogs [38]). Epidemiological studies that have found inadequate household sanitation is a risk factor for cryptosporidiosis infection in low-income countries including India [41], Venezuela [42] and Peru [43]. Therefore, the higher prevalence of C. hominis compared to C. parvum in low income countries observed suggests that anthroponotic transmission rather than zoonotic transmission is the main pathway in such countries, and this may explain the higher relative proportion of C. parvum IIc in those same countries. However, there may be other reasons for seeing a higher relative proportion of C. parvum IIc, including increased virulence or prolonged shedding for example. Our study focused on C. parvum subtypes, and thus C. hominis subtypes were not included. It is possible that certain C. hominis subtypes would also show increased prevalence in lower income countries, particularly for example IbA10G2 which is thought to be more virulent [12], but this would need to be systematically studied in order to make any conclusions.
Data within this systematic review were not robust enough to draw conclusions about the virulence of C. parvum IIc. However, it is possible that C. parvum IIc is more virulent than other C. parvum subtypes, as we know now C. parvum IIc is causing the majority of C. parvum infections in low-income settings where morbidity and mortality due to Cryptosporidium infections is highest. However, our data cannot provide the evidence for this, and it is plausible that there are host susceptibility factors involved which make C. parvum IIc more prevalent, but not necessarily more virulent, as there may be other subtypes that may cause less cases, but potentially more virulent disease. In addition, as C. parvum IIc is anthroponotic it may act more like C. hominis than zoonotic C. parvum and research has previously suggested that C. hominis is more virulent than C. parvum [11]. One observational study [44] stratified AIDS patients with cryptosporidiosis into three groups (mild, moderate and severe) based on symptom features, and in the severe group the only subtype of Cryptosporidium found was C. parvum IIc. In addition, they suggested that wasting syndrome was strongly linked to the subtype IIc, with wasting seen in four out of four patients with IIc subtype, whereas no wasting syndrome was seen in patients with Ia (1 patient) and IIa (4 patients) subtypes of Cryptosporidium. The study was, however, limited by its small size as it included only nine cases of cryptosporidiosis, and the patient population included AIDS patients only. It does though, raise the question of whether C. parvum IIc causes more severe disease demonstrating the need for further research to address this. Well conducted, adequately powered epidemiological studies investigating differences in clinical symptoms, illness duration and long-term sequelae between different subtypes would be able to provide data to answer this question.
There are several limitations of this review. The first is that it relied on published studies to report the proportion of C. parvum IIc, and these studies are not systematic studies of subtyping. Instead subtyping tends to be done on a subset of samples, often due to expense, which can mean small numbers of subtyped samples are available for analysis. In addition, earlier studies often did not subtype, before gp60 subtyping became more widespread, and thus these studies could not be included in this review. Gp60 subtyping is the commonly used method for subtyping Cryptosporidium, but concerns have been raised as to whether it could be missing some genetic diversity and the role of multi-locus typing has been investigated [45]. However, a consensus is yet to be reached and as such gp60 subtyping has remained the mainstay of Cryptosporidium subtyping, although recently a working group has been set-up to implement and establish a multi-locus genotyping scheme for Cryptosporidium [46]. Regarding the statistical analysis relating to GDP and percentage access to improved sanitation and improved water supply, multiple variate parameters were not undertaken due to high degrees of co-lineality.
With an increased emphasis on Cryptosporidium as a pathogen capable of causing severe disease and now recognised to be the second leading cause of death from diarrhoeal disease in children under the age of five [5], this study emphasises how anthroponotic C. parvum IIc is disproportionately affecting low income countries and demonstrates a clear link with sanitation.
There is an estimated disease burden of 7.6 million diarrhoeal cases due to Cryptosporidium annually [47] and recent understanding has highlighted the importance of anthroponotic Cryptosporidium in causing these infections in sub-Saharan Africa and South Asia, with C. hominis responsible for 77.8% and C. parvum 9.9% of these Cryptospordium infections. Of the C. parvum cases, 91.9% were anthroponotic, of which the far majority were IIc (57%) and then IIe (35%) [47]. Extrapolating this information suggests a C. parvum disease burden of three quarters of a million cases, with more than half of these caused by IIc, resulting in significant morbidity and mortality from the parasites comprising this GP60 subtype. This corresponds to our study finding of high proportion of C. parvum IIc in low-income countries, where the biggest burden of diarrhoeal disease in children is seen, and clearly any intervention to reduce this is desirable.
Given the disease burden associated with cryptosporidiosis in low-income countries and the current lack of an effective treatment or vaccine, there is a need for improved prevention. Our findings and those of the few other studies that have investigated suggest that improving sanitation provision may be the most important intervention to reduce the burden of disease from cryptosporidiosis and its associated increased risk of death in young children. We would support the importance of achieving the Sustainable Development Goal on sanitation provision.
Conclusions
Our systematic study has shown that anthroponotic C. parvum IIc predominates in lower-income countries with poor sanitation and in HIV positive individuals, in contrast to higher-income countries where it is rarely evident. Considering the large disease burden of cryptosporidiosis in low-income countries and the post-infectious virulence of other anthroponotic Cryptosporidium species such as C. hominis, C. parvum IIc plays an increasingly apparent role in this disease process. Given the current lack of effective treatment or vaccine, interventions to improve sanitation provision may be the best option to try and reduce the cryptosporidiosis disease burden and associated childhood deaths in lower income countries.
Additional files
Table S1. PRISMA checklist. (DOC 62 kb)
Figure S1. Forest plots ordered by increasing sanitation coverage in country of study for C. parvum IIc (a) C. parvum IIa (b) and C. parvum IId (c) illustrating the increased proportion of C. parvum IIc found in countries with poor sanitation coverage and low proportion of C. parvum IIc in countries with high % sanitation coverage, in comparison to C. parvum IIa which is frequently seen in a higher proportion in countries with high % sanitation coverage and C. parvum IId which appears to cluster in Arabic countries. Vertical line within the figures equals the pooled relative proportion of all studies. (PDF 1131 kb)
Acknowledgments
Not applicable.
Funding
PRH is supported by the National Institute for Health Research Health Protection Research Unit (NIHR HPRU) in Gastrointestinal Infections at University of Liverpool in partnership with Public Health England (PHE), in collaboration with the University of East Anglia, Norwich Medical School. PK is supported by a National Institute for Health Research Academic Clinical Fellowship.
Availability of data and materials
All data generated or analysed during this study are included in this published article and its additional files.
Authors’ contributions
PRH and KMT proposed the study. PK performed the initial literature search and extracted the data. PRH reviewed the studies included in the search and their inclusion or exclusion in the final dataset and performed the statistical calculations. PK drafted the paper, KMT refined the draft and all authors contributed to, read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Philippa King, Email: philippaking@live.co.uk.
Kevin M. Tyler, Email: k.tyler@uea.ac.uk
Paul R. Hunter, Email: Paul.Hunter@uea.ac.uk
References
- 1.Efstratiou A, Ongerth JE, Karanis P. Waterborne transmission of protozoan parasites: Review of worldwide outbreaks - an update 2011–2016. Water Res. 2017;114:14–22. doi: 10.1016/j.watres.2017.01.036. [DOI] [PubMed] [Google Scholar]
- 2.Ryan U, Hijjawi N, Xiao L. Foodborne cryptosporidiosis. Int J Parasitol. 2018;48:1–12. doi: 10.1016/j.ijpara.2017.09.004. [DOI] [PubMed] [Google Scholar]
- 3.Kotloff KL, Nataro JP, Blackwelder WC, Nasrin D, Farag TH, Panchalingam S, et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): a prospective, case-control study. Lancet. 2013;382:209–222. doi: 10.1016/S0140-6736(13)60844-2. [DOI] [PubMed] [Google Scholar]
- 4.Checkley W, Gilman RH, Epstein LD, Suarez M, Diaz JF, Cabrera L, et al. Asymptomatic and symptomatic cryptosporidiosis: their acute effect on weight gain in Peruvian children. Am J Epidemiol. 1997;145:156–163. doi: 10.1093/oxfordjournals.aje.a009086. [DOI] [PubMed] [Google Scholar]
- 5.Collaborators GMaCoD Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980–2015: a systematic analysis for the Global Burden of Disease Study 2015. Lancet. 2016;388:1459–1544. doi: 10.1016/S0140-6736(16)31012-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.WHO . Rotavirus. 2016. 2016. [Google Scholar]
- 7.Ryan U, Fayer R, Xiao L. Cryptosporidium species in humans and animals: current understanding and research needs. Parasitology. 2014;141:1667–1685. doi: 10.1017/S0031182014001085. [DOI] [PubMed] [Google Scholar]
- 8.Xiao L. Molecular epidemiology of cryptosporidiosis: an update. Exp Parasitol. 2010;124:80–89. doi: 10.1016/j.exppara.2009.03.018. [DOI] [PubMed] [Google Scholar]
- 9.Sulaiman IM, Hira PR, Zhou L, Al-Ali FM, Al-Shelahi FA, Shweiki HM, et al. Unique endemicity of cryptosporidiosis in children in Kuwait. J Clin Microbiol. 2005;43:2805–2809. doi: 10.1128/JCM.43.6.2805-2809.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bouzid M, Hunter PR, Chalmers RM, Tyler KM. Cryptosporidium pathogenicity and virulence. Clin Microbiol Rev. 2013;26:115–134. doi: 10.1128/CMR.00076-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hunter PR, Hughes S, Woodhouse S, Raj N, Syed Q, Chalmers RM, et al. Health sequelae of human cryptosporidiosis in immunocompetent patients. Clin Infect Dis. 2004;39:504–510. doi: 10.1086/422649. [DOI] [PubMed] [Google Scholar]
- 12.Cama VA, Bern C, Roberts J, Cabrera L, Sterling CR, Ortega Y, et al. Cryptosporidium species and subtypes and clinical manifestations in children. Peru. Emerg Infect Dis. 2008;14:1567–1574. doi: 10.3201/eid1410.071273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.World Bank Development Indicators Archive. http://databank.worldbank.org/data/reports.aspx?source=wdi-database-archives-%28beta%29. Accessed 30 Mar 2017.
- 14.WHO/UNICEF Joint Monitoring Programme. https://washdata.org/monitoring/methods. Accessed 4 Nov 2016.
- 15.Mbae C, Mulinge E, Waruru A, Ngugi B, Wainaina J, Kariuki S. Genetic diversity of Cryptosporidium in children in an urban informal settlement of Nairobi, Kenya. PLoS One. 2015;10:e0142055. doi: 10.1371/journal.pone.0142055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Eibach D, Krumkamp R, Al-Emran HM, Sarpong N, Hagen RM, Adu-Sarkodie Y, et al. Molecular characterization of Cryptosporidium spp. among children in rural Ghana. PLoS Negl Trop Dis. 2015;9:e0003551. doi: 10.1371/journal.pntd.0003551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Blanco MA, Montoya A, Iborra A, Fuentes I. Identification of Cryptosporidium subtype isolates from HIV-seropositive patients in Equatorial Guinea. Trans R Soc Trop Med Hyg. 2014;108:594–596. doi: 10.1093/trstmh/tru108. [DOI] [PubMed] [Google Scholar]
- 18.Molloy SF, Smith HV, Kirwan P, Nichols RA, Asaolu SO, Connelly L, et al. Identification of a high diversity of Cryptosporidium species genotypes and subtypes in a pediatric population in Nigeria. Am J Trop Med Hyg. 2010;82:608–613. doi: 10.4269/ajtmh.2010.09-0624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gatei W, Barrett D, Lindo JF, Eldemire-Shearer D, Cama V, Xiao L. Unique Cryptosporidium population in HIV-infected persons, Jamaica. Emerg Infect Dis. 2008;14:841–843. doi: 10.3201/eid1405.071277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Cama VA, Ross JM, Crawford S, Kawai V, Chavez-Valdez R, Vargas D, et al. Differences in clinical manifestations among Cryptosporidium species and subtypes in HIV-infected persons. J Infect Dis. 2007;196:684–691. doi: 10.1086/519842. [DOI] [PubMed] [Google Scholar]
- 21.Ajjampur SS, Gladstone BP, Selvapandian D, Muliyil JP, Ward H, Kang G. Molecular and spatial epidemiology of cryptosporidiosis in children in a semiurban community in South India. J Clin Microbiol. 2007;45:915–920. doi: 10.1128/JCM.01590-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Akiyoshi DE, Tumwine JK, Bakeera-Kitaka S, Tzipori S. Subtype analysis of Cryptosporidium isolates from children in Uganda. J Parasitol. 2006;92:1097–1100. doi: 10.1645/GE-843R.1. [DOI] [PubMed] [Google Scholar]
- 23.Insulander M, Silverlås C, Lebbad M, Karlsson L, Mattsson JG, Svenungsson B. Molecular epidemiology and clinical manifestations of human cryptosporidiosis in Sweden. Epidemiol Infect. 2013;141:1009–1020. doi: 10.1017/S0950268812001665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Chalmers RM, Smith RP, Hadfield SJ, Elwin K, Giles M. Zoonotic linkage and variation in Cryptosporidium parvum from patients in the United Kingdom. Parasitol Res. 2011;108:1321–1325. doi: 10.1007/s00436-010-2199-x. [DOI] [PubMed] [Google Scholar]
- 25.Wielinga PR, de Vries A, van der Goot TH, Mank T, Mars MH, Kortbeek LM, et al. Molecular epidemiology of Cryptosporidium in humans and cattle in The Netherlands. Int J Parasitol. 2008;38:809–817. doi: 10.1016/j.ijpara.2007.10.014. [DOI] [PubMed] [Google Scholar]
- 26.Waldron LS, Ferrari BC, Power ML. Glycoprotein 60 diversity in C. hominis and C. parvum causing human cryptosporidiosis in NSW, Australia. Exp Parasitol. 2009;122:124–127. doi: 10.1016/j.exppara.2009.02.006. [DOI] [PubMed] [Google Scholar]
- 27.Koehler AV, Bradbury RS, Stevens MA, Haydon SR, Jex AR, Gasser RB. Genetic characterization of selected parasites from people with histories of gastrointestinal disorders using a mutation scanning-coupled approach. Electrophoresis. 2013;34:1720–1728. doi: 10.1002/elps.201300100. [DOI] [PubMed] [Google Scholar]
- 28.Koehler AV, Whipp M, Hogg G, Haydon SR, Stevens MA, Jex AR, et al. First genetic analysis of Cryptosporidium from humans from Tasmania, and identification of a new genotype from a traveller to Bali. Electrophoresis. 2014;35:2600–2607. doi: 10.1002/elps.201400225. [DOI] [PubMed] [Google Scholar]
- 29.Ng JS, Eastwood K, Walker B, Durrheim DN, Massey PD, Porigneaux P, et al. Evidence of Cryptosporidium transmission between cattle and humans in northern New South Wales. Exp Parasitol. 2012;130:437–441. doi: 10.1016/j.exppara.2012.01.014. [DOI] [PubMed] [Google Scholar]
- 30.Waldron LS, Dimeski B, Beggs PJ, Ferrari BC, Power ML. Molecular epidemiology, spatiotemporal analysis, and ecology of sporadic human cryptosporidiosis in Australia. Appl Environ Microbiol. 2011;77:7757–7765. doi: 10.1128/AEM.00615-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ng J, MacKenzie B, Ryan U. Longitudinal multi-locus molecular characterisation of sporadic Australian human clinical cases of cryptosporidiosis from 2005 to 2008. Exp Parasitol. 2010;125:348–356. doi: 10.1016/j.exppara.2010.02.017. [DOI] [PubMed] [Google Scholar]
- 32.Ng J, Eastwood K, Durrheim D, Massey P, Walker B, Armson A, et al. Evidence supporting zoonotic transmission of Cryptosporidium in rural New South Wales. Exp Parasitol. 2008;119:192–195. doi: 10.1016/j.exppara.2008.01.010. [DOI] [PubMed] [Google Scholar]
- 33.Grinberg A, Learmonth J, Kwan E, Pomroy W, Lopez Villalobos N, Gibson I, et al. Genetic diversity and zoonotic potential of Cryptosporidium parvum causing foal diarrhea. J Clin Microbiol. 2008;46:2396–2398. doi: 10.1128/JCM.00936-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Unicef, WHO: 25 years progress on sanitation and drinking water, 2015 update and MDG assessment. (2015) www.unicef.org/publications/index_82419.html. Accessed 31 Mar 2017.
- 35.Krawczyk AI, van Leeuwen AD, Jacobs-Reitsma W, Wijnands LM, Bouw E, Jahfari S, et al. Presence of zoonotic agents in engorged ticks and hedgehog faeces from Erinaceus europaeus in (sub) urban areas. Parasit Vectors. 2015;8:210. doi: 10.1186/s13071-015-0814-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Sangster L, Blake DP, Robinson G, Hopkins TC, Sa RC, Cunningham AA, et al. Detection and molecular characterisation of Cryptosporidium parvum in British European hedgehogs (Erinaceus europaeus) Vet Parasitol. 2016;217:39–44. doi: 10.1016/j.vetpar.2015.12.006. [DOI] [PubMed] [Google Scholar]
- 37.Dyachenko V, Kuhnert Y, Schmaeschke R, Etzold M, Pantchev N, Daugschies A. Occurrence and molecular characterization of Cryptosporidium spp. genotypes in European hedgehogs (Erinaceus europaeus L.) in Germany. Parasitology. 2010;137:205–216. doi: 10.1017/S0031182009991089. [DOI] [PubMed] [Google Scholar]
- 38.Gil H, Cano L, de Lucio A, Bailo B, de Mingo MH, Cardona GA, et al. Detection and molecular diversity of Giardia duodenalis and Cryptosporidium spp. in sheltered dogs and cats in northern Spain. Infect Genet Evol. 2017;50:62–69. doi: 10.1016/j.meegid.2017.02.013. [DOI] [PubMed] [Google Scholar]
- 39.Karanis P, Kourenti C, Smith H. Waterborne transmission of protozoan parasites: a worldwide review of outbreaks and lessons learnt. J Water Health. 2007;5:1–38. doi: 10.2166/wh.2006.002. [DOI] [PubMed] [Google Scholar]
- 40.Putignani L, Menichella D. Global distribution, public health and clinical impact of the protozoan pathogen Cryptosporidium. Interdiscip Perspect Infect Dis. 2010;2010:753512. doi: 10.1155/2010/753512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sarkar R, Kattula D, Francis MR, Ajjampur SS, Prabakaran AD, Jayavelu N, et al. Risk factors for cryptosporidiosis among children in a semi urban slum in southern India: a nested case-control study. Am J Trop Med Hyg. 2014;91:1128–1137. doi: 10.4269/ajtmh.14-0304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Chacín-Bonilla L, Barrios F, Sanchez Y. Environmental risk factors for Cryptosporidium infection in an island from western Venezuela. Mem Inst Oswaldo Cruz. 2008;103:45–49. doi: 10.1590/S0074-02762008005000007. [DOI] [PubMed] [Google Scholar]
- 43.Bern C, Ortega Y, Checkley W, Roberts JM, Lescano AG, Cabrera L, et al. Epidemiologic differences between cyclosporiasis and cryptosporidiosis in Peruvian children. Emerg Infect Dis. 2002;8:581–585. doi: 10.3201/eid0806.01-0331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Del Chierico F, Onori M, Di Bella S, Bordi E, Petrosillo N, Menichella D, et al. Cases of cryptosporidiosis co-infections in AIDS patients: a correlation between clinical presentation and GP60 subgenotype lineages from aged formalin-fixed stool samples. Ann Trop Med Parasitol. 2011;105:339–349. doi: 10.1179/1364859411Y.0000000025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Robinson G, Chalmers RM. Assessment of polymorphic genetic markers for multi-locus typing of Cryptosporidium parvum and Cryptosporidium hominis. Exp Parasitol. 2012;132:200–215. doi: 10.1016/j.exppara.2012.06.016. [DOI] [PubMed] [Google Scholar]
- 46.Chalmers RM, Pérez-Cordón G, Cacció SM, Klotz C, Robertson LJ. Participants of the Cryptosporidium genotyping workshop (EURO-FBP). Cryptosporidium genotyping in Europe: the current status and processes for a harmonised multi-locus genotyping scheme. Exp Parasitol. 2018;191:25–30. doi: 10.1016/j.exppara.2018.06.004. [DOI] [PubMed] [Google Scholar]
- 47.Sow SO, Muhsen K, Nasrin D, Blackwelder WC, Wu Y, Farag TH, et al. The burden of Cryptosporidium diarrheal disease among children < 24 months of age in moderate/high mortality regions of sub-Saharan Africa and South Asia, utilizing data from the Global Enteric Multicenter Study (GEMS) PLoS Negl Trop Dis. 2016;10:e0004729. doi: 10.1371/journal.pntd.0004729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hira KG, Mackay MR, Hempstead AD, Ahmed S, Karim MM, O’Connor RM, et al. Genetic diversity of Cryptosporidium spp. from Bangladeshi children. J Clin Microbiol. 2011;49:2307–2310. doi: 10.1128/JCM.00164-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Moore CE, Elwin K, Phot N, Seng C, Mao S, Suy K, et al. Molecular characterization of Cryptosporidium species and Giardia duodenalis from symptomatic Cambodian children. PLoS Negl Trop Dis. 2016;10:e0004822. doi: 10.1371/journal.pntd.0004822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wang L, Zhang H, Zhao X, Zhang L, Zhang G, Guo M, et al. Zoonotic Cryptosporidium species and Enterocytozoon bieneusi genotypes in HIV-positive patients on antiretroviral therapy. J Clin Microbiol. 2013;51:557–563. doi: 10.1128/JCM.02758-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Sharma P, Sharma A, Sehgal R, Malla N, Khurana S. Genetic diversity of Cryptosporidium isolates from patients in North India. Int J Infect Dis. 2013;17:e601–e605. doi: 10.1016/j.ijid.2012.12.003. [DOI] [PubMed] [Google Scholar]
- 52.Ajjampur SS, Liakath FB, Kannan A, Rajendran P, Sarkar R, Moses PD, et al. Multisite study of cryptosporidiosis in children with diarrhea in India. J Clin Microbiol. 2010;48:2075–2081. doi: 10.1128/JCM.02509-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Muthusamy D, Rao SS, Ramani S, Monica B, Banerjee I, Abraham OC, et al. Multilocus genotyping of Cryptosporidium sp. isolates from human immunodeficiency virus-infected individuals in South India. J Clin Microbiol. 2006;44:632–634. doi: 10.1128/JCM.44.2.632-634.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Sharbatkhori M, Nazemalhosseini Mojarad E, Taghipour N, Pagheh AS, Mesgarian F. Prevalence and genetic characterization of Cryptosporidium spp. in diarrheic children from Gonbad Kavoos City, Iran. Iran J Parasitol. 2015;10:441–447. [PMC free article] [PubMed] [Google Scholar]
- 55.Taghipour N, Nazemalhosseini-Mojarad E, Haghighi A, Rostami-Nejad M, Romani S, Keshavarz A, et al. Molecular epidemiology of cryptosporidiosis in Iranian children, Tehran, Iran. Iran J Parasitol. 2011;6:41–5. [PMC free article] [PubMed]
- 56.Nazemalhosseini-Mojarad E, Haghighi A, Taghipour N, Keshavarz A, Mohebi SR, Zali MR, et al. Subtype analysis of Cryptosporidium parvum and Cryptosporidium hominis isolates from humans and cattle in Iran. Vet Parasitol. 2011;179:250–252. doi: 10.1016/j.vetpar.2011.01.051. [DOI] [PubMed] [Google Scholar]
- 57.Abe N, Matsubayashi M, Kimata I, Iseki M. Subgenotype analysis of Cryptosporidium parvum isolates from humans and animals in Japan using the 60-kDa glycoprotein gene sequences. Parasitol Res. 2006;99:303–305. doi: 10.1007/s00436-006-0140-0. [DOI] [PubMed] [Google Scholar]
- 58.Wu Z, Nagano I, Boonmars T, Nakada T, Takahashi Y. Intraspecies polymorphism of Cryptosporidium parvum revealed by PCR-restriction fragment length polymorphism (RFLP) and RFLP-single-strand conformational polymorphism analyses. Appl Environ Microbiol. 2003;69:4720–4726. doi: 10.1128/AEM.69.8.4720-4726.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Hijjawi N, Mukbel R, Yang R, Ryan U. Genetic characterization of Cryptosporidium in animal and human isolates from Jordan. Vet Parasitol. 2016;228:116–120. doi: 10.1016/j.vetpar.2016.08.015. [DOI] [PubMed] [Google Scholar]
- 60.Hijjawi N, Ng J, Yang R, Atoum MF, Ryan U. Identification of rare and novel Cryptosporidium GP60 subtypes in human isolates from Jordan. Exp Parasitol. 2010;125:161–164. doi: 10.1016/j.exppara.2010.01.011. [DOI] [PubMed] [Google Scholar]
- 61.Iqbal J, Khalid N, Hira PR. Cryptosporidiosis in Kuwaiti children: association of clinical characteristics with Cryptosporidium species and subtypes. J Med Microbiol. 2011;60:647–652. doi: 10.1099/jmm.0.028001-0. [DOI] [PubMed] [Google Scholar]
- 62.Osman M, El Safadi D, Benamrouz S, Guyot K, Dei-Cas E, Aliouat el M, et al. Initial data on the molecular epidemiology of cryptosporidiosis in Lebanon. PLoS One. 2015;10:e0125129. doi: 10.1371/journal.pone.0125129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Iqbal A, Lim YA, Surin J, Sim BL. High diversity of Cryptosporidium subgenotypes identified in Malaysian HIV/AIDS individuals targeting gp60 gene. PLoS One. 2012;7:e31139. doi: 10.1371/journal.pone.0031139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Lim YA, Iqbal A, Surin J, Sim BL, Jex AR, Nolan MJ, et al. First genetic classification of Cryptosporidium and Giardia from HIV/AIDS patients in Malaysia. Infect Genet Evol. 2011;11:968–974. doi: 10.1016/j.meegid.2011.03.007. [DOI] [PubMed] [Google Scholar]
- 65.Alyousefi NA, Mahdy MA, Lim YA, Xiao L, Mahmud R. First molecular characterization of Cryptosporidium in Yemen. Parasitology. 2013;140:729–734. doi: 10.1017/S0031182012001953. [DOI] [PubMed] [Google Scholar]
- 66.Ibrahim MA, Abdel-Ghany AE, Abdel-Latef GK, Abdel-Aziz SA, Aboelhadid SM. Epidemiology and public health significance of Cryptosporidium isolated from cattle, buffaloes, and humans in Egypt. Parasitol Res. 2016;115:2439–2448. doi: 10.1007/s00436-016-4996-3. [DOI] [PubMed] [Google Scholar]
- 67.Helmy YA, Krücken J, Nöckler K, von Samson-Himmelstjerna G, Zessin KH. Molecular epidemiology of Cryptosporidium in livestock animals and humans in the Ismailia Province of Egypt. Vet Parasitol. 2013;193:15–24. [DOI] [PubMed]
- 68.Adamu H, Petros B, Zhang G, Kassa H, Amer S, Ye J, et al. Distribution and clinical manifestations of Cryptosporidium species and subtypes in HIV/AIDS patients in Ethiopia. PLoS Negl Trop Dis. 2014;8:e2831. doi: 10.1371/journal.pntd.0002831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Adamu H, Petros B, Hailu A, Petry F. Molecular characterization of Cryptosporidium isolates from humans in Ethiopia. Acta Trop. 2010;115:77–83. doi: 10.1016/j.actatropica.2010.02.003. [DOI] [PubMed] [Google Scholar]
- 70.Areeshi M, Dove W, Papaventsis D, Gatei W, Combe P, Grosjean P, et al. Cryptosporidium species causing acute diarrhoea in children in Antananarivo, Madagascar. Ann Trop Med Parasitol. 2008;102:309–315. doi: 10.1179/136485908X278793. [DOI] [PubMed] [Google Scholar]
- 71.Peng MM, Meshnick SR, Cunliffe NA, Thindwa BD, Hart CA, Broadhead RL, et al. Molecular epidemiology of cryptosporidiosis in children in Malawi. J Eukaryot Microbiol. 2003;50:557–559. doi: 10.1111/j.1550-7408.2003.tb00628.x. [DOI] [PubMed] [Google Scholar]
- 72.Ojuromi OT, Duan L, Izquierdo F, Fenoy SM, Oyibo WA, Del Aguila C, et al. Genotypes of Cryptosporidium spp. and Enterocytozoon bieneusi in human immunodeficiency virus-infected patients in Lagos, Nigeria. J Eukaryot Microbiol. 2016;63:414–418. doi: 10.1111/jeu.12285. [DOI] [PubMed] [Google Scholar]
- 73.Akinbo FO, Okaka CE, Omoregie R, Adamu H, Xiao L. Unusual Enterocytozoon bieneusi genotypes and Cryptosporidium hominis subtypes in HIV-infected patients on highly active antiretroviral therapy. Am J Trop Med Hyg. 2013;89:157–161. doi: 10.4269/ajtmh.12-0635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Maikai BV, Umoh JU, Lawal IA, Kudi AC, Ejembi CL, Xiao L. Molecular characterizations of Cryptosporidium, Giardia, and Enterocytozoon in humans in Kaduna State, Nigeria. Exp Parasitol. 2012;131:452–456. doi: 10.1016/j.exppara.2012.05.011. [DOI] [PubMed] [Google Scholar]
- 75.Ayinmode AB, Fagbemi BO, Xiao L. Molecular characterization of Cryptosporidium in children in Oyo State, Nigeria: implications for infection sources. Parasitol Res. 2012;110:479–481. doi: 10.1007/s00436-011-2531-0. [DOI] [PubMed] [Google Scholar]
- 76.Akinbo FO, Okaka CE, Omoregie R, Dearen T, Leon ET, Xiao L. Molecular characterization of Cryptosporidium spp. in HIV-infected persons in Benin City, Edo State, Nigeria. Fooyin J Health Sci. 2010;2:85–9.
- 77.Lobo ML, Augusto J, Antunes F, Ceita J, Xiao L, Codices V, et al. Cryptosporidium spp., Giardia duodenalis, Enterocytozoon bieneusi and other intestinal parasites in young children in Lobata Province, Democratic Republic of São Tomé and Principe. PLoS One. 2014;9:e97708. doi: 10.1371/journal.pone.0097708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Abu Samra N, Thompson PN, Jori F, Frean J, Poonsamy B, du Plessis D, et al. Genetic characterization of Cryptosporidium spp. in diarrhoeic children from four provinces in South Africa. Zoonoses Public Health. 2013;60:154–159. doi: 10.1111/j.1863-2378.2012.01507.x. [DOI] [PubMed] [Google Scholar]
- 79.Leav BA, Mackay MR, Anyanwu A, O’Connor RM, Cevallos AM, Kindra G, et al. Analysis of sequence diversity at the highly polymorphic Cpgp40/15 locus among Cryptosporidium isolates from human immunodeficiency virus-infected children in South Africa. Infect Immun. 2002;70:3881–3890. doi: 10.1128/IAI.70.7.3881-3890.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Rahmouni I, Essid R, Aoun K, Bouratbine A. Glycoprotein 60 diversity in Cryptosporidium parvum causing human and cattle cryptosporidiosis in the rural region of northern Tunisia. Am J Trop Med Hyg. 2014;90:346–350. doi: 10.4269/ajtmh.13-0522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Geurden T, Levecke B, Cacció SM, Visser A, De Groote G, Casaert S, et al. Multilocus genotyping of Cryptosporidium and Giardia in non-outbreak related cases of diarrhoea in human patients in Belgium. Parasitology. 2009;136:1161–1168. doi: 10.1017/S0031182009990436. [DOI] [PubMed] [Google Scholar]
- 82.Rasková V, Kvetonová D, Sak B, McEvoy J, Edwinson A, Stenger B, et al. Human cryptosporidiosis caused by Cryptosporidium tyzzeri and C. parvum isolates presumably transmitted from wild mice. J Clin Microbiol. 2013;51:360–362. doi: 10.1128/JCM.02346-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Stensvold CR, Ethelberg S, Hansen L, Sahar S, Voldstedlund M, Kemp M, et al. Cryptosporidium infections in Denmark, 2010–2014. Dan Med J. 2015;62:A5086. [PubMed] [Google Scholar]
- 84.Lassen B, Ståhl M, Enemark HL. Cryptosporidiosis - an occupational risk and a disregarded disease in Estonia. Acta Vet Scand. 2014;56:36. doi: 10.1186/1751-0147-56-36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Brunet J, Lemoine JP, Pesson B, Valot S, Sautour M, Dalle F, et al. Ruling out nosocomial transmission of Cryptosporidium in a renal transplantation unit: case report. BMC Infect Dis. 2016;16:363. doi: 10.1186/s12879-016-1661-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Zintl A, Ezzaty-Mirashemi M, Chalmers RM, Elwin K, Mulcahy G, Lucy FE, et al. Longitudinal and spatial distribution of GP60 subtypes in human cryptosporidiosis cases in Ireland. Epidemiol Infect. 2011;139:1945–1955. doi: 10.1017/S0950268810002992. [DOI] [PubMed] [Google Scholar]
- 87.Zintl A, Proctor AF, Read C, Dewaal T, Shanaghy N, Fanning S, et al. The prevalence of Cryptosporidium species and subtypes in human faecal samples in Ireland. Epidemiol Infect. 2009;137:270–277. doi: 10.1017/S0950268808000769. [DOI] [PubMed] [Google Scholar]
- 88.Alves M, Xiao L, Antunes F, Matos O. Distribution of Cryptosporidium subtypes in humans and domestic and wild ruminants in Portugal. Parasitol Res. 2006;99:287–292. doi: 10.1007/s00436-006-0164-5. [DOI] [PubMed] [Google Scholar]
- 89.Vieira PM, Mederle N, Lobo ML, Imre K, Mederle O, Xiao L, et al. Molecular characterisation of Cryptosporidium (Apicomplexa) in children and cattle in Romania. Folia Parasitol (Praha) 2015;62:002. doi: 10.14411/fp.2015.002. [DOI] [PubMed] [Google Scholar]
- 90.Deshpande AP, Jones BL, Connelly L, Pollock KG, Brownlie S, Alexander CL. Molecular characterization of Cryptosporidium parvum isolates from human cryptosporidiosis cases in Scotland. Parasitology. 2015;142:318–325. doi: 10.1017/S0031182014001346. [DOI] [PubMed] [Google Scholar]
- 91.Petrincová A, Valenčáková A, Luptáková L, Ondriska F, Kalinová J, Halánová M, et al. Molecular characterization and first report of Cryptosporidium genotypes in human population in the Slovak Republic. Electrophoresis. 2015;36:2925–2930. doi: 10.1002/elps.201500230. [DOI] [PubMed] [Google Scholar]
- 92.Soba B, Logar J. Genetic classification of Cryptosporidium isolates from humans and calves in Slovenia. Parasitology. 2008;135:1263–1270. doi: 10.1017/S0031182008004800. [DOI] [PubMed] [Google Scholar]
- 93.de Lucio A, Merino FJ, Martínez-Ruiz R, Bailo B, Aguilera M, Fuentes I, et al. Molecular genotyping and sub-genotyping of Cryptosporidium spp. isolates from symptomatic individuals attending two major public hospitals in Madrid, Spain. Infect Genet Evol. 2016;37:49–56. doi: 10.1016/j.meegid.2015.10.026. [DOI] [PubMed] [Google Scholar]
- 94.Segura R, Prim N, Montemayor M, Valls ME, Muñoz C. Predominant virulent IbA10G2 subtype of Cryptosporidium hominis in human isolates in Barcelona: a five-year study. PLoS One. 2015;10:e0121753. doi: 10.1371/journal.pone.0121753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Ramo A, Quílez J, Vergara-Castiblanco C, Monteagudo L, Del Cacho E, Clavel A. Multilocus typing and population structure of Cryptosporidium from children in Zaragoza, Spain. Infect Genet Evol. 2015;31:190–197. doi: 10.1016/j.meegid.2015.01.023. [DOI] [PubMed] [Google Scholar]
- 96.Abal-Fabeiro JL, Maside X, Llovo J, Bello X, Torres M, Treviño M, et al. High-throughput genotyping assay for the large-scale genetic characterization of Cryptosporidium parasites from human and bovine samples. Parasitology. 2014;141:491–500. doi: 10.1017/S0031182013001807. [DOI] [PubMed] [Google Scholar]
- 97.Jex AR, Gasser RB. Analysis of the genetic diversity within Cryptosporidium hominis and Cryptosporidium parvum from imported and autochtonous cases of human cryptosporidiosis by mutation scanning. Electrophoresis. 2008;29:4119–4129. doi: 10.1002/elps.200800422. [DOI] [PubMed] [Google Scholar]
- 98.Iqbal A, Goldfarb DM, Slinger R, Dixon BR. Prevalence and molecular characterization of Cryptosporidium spp. and Giardia duodenalis in diarrhoeic patients in the Qikiqtani Region, Nunavut, Canada. Int J Circumpolar Health. 2015;74:27713. doi: 10.3402/ijch.v74.27713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Budu-Amoako E, Greenwood SJ, Dixon BR, Sweet L, Ang L, Barkema HW, et al. Molecular epidemiology of Cryptosporidium and Giardia in humans on Prince Edward Island, Canada: evidence of zoonotic transmission from cattle. Zoonoses Public Health. 2012;59:424–433. doi: 10.1111/j.1863-2378.2012.01474.x. [DOI] [PubMed] [Google Scholar]
- 100.Trotz-Williams LA, Martin DS, Gatei W, Cama V, Peregrine AS, Martin SW, et al. Genotype and subtype analyses of Cryptosporidium isolates from dairy calves and humans in Ontario. Parasitol Res. 2006;99:346–352. doi: 10.1007/s00436-006-0157-4. [DOI] [PubMed] [Google Scholar]
- 101.Valenzuela O, González-Díaz M, Garibay-Escobar A, Burgara-Estrella A, Cano M, Durazo M, et al. Molecular characterization of Cryptosporidium spp. in children from Mexico. PLoS One. 2014;9:e96128. doi: 10.1371/journal.pone.0096128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Xiao L, Hlavsa MC, Yoder J, Ewers C, Dearen T, Yang W, et al. Subtype analysis of Cryptosporidium specimens from sporadic cases in Colorado, Idaho, New Mexico, and Iowa in 2007: widespread occurrence of one Cryptosporidium hominis subtype and case history of an infection with the Cryptosporidium horse genotype. J Clin Microbiol. 2009;47:3017–3020. doi: 10.1128/JCM.00226-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Feltus DC, Giddings CW, Schneck BL, Monson T, Warshauer D, McEvoy JM. Evidence supporting zoonotic transmission of Cryptosporidium spp. in Wisconsin. J Clin Microbiol. 2006;44:4303–4308. doi: 10.1128/JCM.01067-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Peralta RH, Velásquez JN, FeS C, Pantano ML, Sodré FC, Silva S, et al. Genetic diversity of Cryptosporidium identified in clinical samples from cities in Brazil and Argentina. Mem Inst Oswaldo Cruz. 2016;111:30–36. doi: 10.1590/0074-02760150303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Jex AR, Pangasa A, Campbell BE, Whipp M, Hogg G, Sinclair MI, et al. Classification of Cryptosporidium species from patients with sporadic cryptosporidiosis by use of sequence-based multilocus analysis following mutation scanning. J Clin Microbiol. 2008;46:2252–2262. doi: 10.1128/JCM.00116-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Jex AR, Whipp M, Campbell BE, Cacciò SM, Stevens M, Hogg G, et al. A practical and cost-effective mutation scanning-based approach for investigating genetic variation in Cryptosporidium. Electrophoresis. 2007;28:3875–3883. doi: 10.1002/elps.200700279. [DOI] [PubMed] [Google Scholar]
- 107.Ghaffari S, Kalantari N. A multi-locus study of cryptosporidium parasites isolated from patients living in Iran, Malawi, Nigeria, the United Kingdom, and Vietnam. Iran J Parasitol. 2014;9:79–89. [PMC free article] [PubMed] [Google Scholar]
- 108.O'Brien E, McInnes L, Ryan U. Cryptosporidium GP60 genotypes from humans and domesticated animals in Australia, North America and Europe. Exp Parasitol. 2008;118:118–121. doi: 10.1016/j.exppara.2007.05.012. [DOI] [PubMed] [Google Scholar]
- 109.Peng MM, Matos O, Gatei W, Das P, Stantic-Pavlinic M, Bern C, et al. A comparison of Cryptosporidium subgenotypes from several geographic regions. J Eukaryot Microbiol. 2001;48:28S–31S. [DOI] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. PRISMA checklist. (DOC 62 kb)
Figure S1. Forest plots ordered by increasing sanitation coverage in country of study for C. parvum IIc (a) C. parvum IIa (b) and C. parvum IId (c) illustrating the increased proportion of C. parvum IIc found in countries with poor sanitation coverage and low proportion of C. parvum IIc in countries with high % sanitation coverage, in comparison to C. parvum IIa which is frequently seen in a higher proportion in countries with high % sanitation coverage and C. parvum IId which appears to cluster in Arabic countries. Vertical line within the figures equals the pooled relative proportion of all studies. (PDF 1131 kb)
Data Availability Statement
All data generated or analysed during this study are included in this published article and its additional files.