

Lipopeptide daptomycin is a last-line cell-membrane-targeting antibiotic to treat multidrug-resistant Staphylococcus aureus. Alarmingly, daptomycin-resistant S. aureus isolates have emerged.
KEYWORDS: Staphylococcus aureus, antibiotic resistance, daptomycin, vancomycin
ABSTRACT
Lipopeptide daptomycin is a last-line cell-membrane-targeting antibiotic to treat multidrug-resistant Staphylococcus aureus. Alarmingly, daptomycin-resistant S. aureus isolates have emerged. The mechanisms underlying daptomycin resistance are diverse and share similarities with resistances to cationic antimicrobial peptides and other lipopeptides, but they remain to be fully elucidated. We selected mutants with increased resistance to daptomycin from a library of transposon insertions in sequent type 8 (ST8) S. aureus HG003. Insertions conferring increased daptomycin resistance were localized to two genes, one coding for a hypothetical lipoprotein (SAOUHSC_00362, Dsp1), and the other for an alkaline shock protein (SAOUHSC_02441, Asp23). Markerless loss-of-function mutants were then generated for comparison. All transposon mutants and knockout strains exhibited increased daptomycin resistance compared to those of wild-type and complemented strains. Null and transposon insertion mutants also exhibited increased resistance to cationic antimicrobial peptides. Interestingly, the Δdsp1 mutant also showed increased resistance to vancomycin, a cell-wall-targeting drug with a different mode of action. Null mutations in both dsp1 and asp23 resulted in increased tolerance as reflected by reduced killing to both daptomycin and vancomycin, as well as an increased tolerance to surfactant (Triton X-100). Neither mutant exhibited increased resistance to lysostaphin, a cell-wall-targeting endopeptidase. These findings identified two genes core to the S. aureus species that make previously uncharacterized contributions to antimicrobial resistance and tolerance in S. aureus.
INTRODUCTION
Staphylococcus aureus is a leading cause of life-threatening infections, both in the hospital and in the community (1, 2). Compounding the problem, effective treatment has become extremely challenging with the emergence of multidrug-resistant lineages, in particular, methicillin-resistant S. aureus (MRSA) (1). Two last-line antibiotics remain generally active against MRSA infections: cell-wall-targeting vancomycin (VAN; glycopeptide family) and daptomycin (DAP; lipopeptide family) (3). Acquired high-level resistance to vancomycin has been described in clinical isolates of S. aureus, but so far, those remain sporadic and rare occurrences. Nevertheless, there is a disconcerting erosion in the efficacy of vancomycin for the treatment of S. aureus associated with MIC creep in MRSA strains (1, 4). In contrast, the cyclic lipopeptide daptomycin shows efficient bactericidal activity against MRSA (5, 6), but the emergence of S. aureus DAP-resistant mutant strains (DAP-R) during the course of protracted treatment is a common finding (7, 8).
Daptomycin shares structural similarities with cationic antimicrobial peptides (CAMPs) and a similar mode of action involving membrane disruption (9). Daptomycin is thought to insert in bacterial membranes of Gram-positive bacteria, in a phosphatidylglycerol-dependent fashion, and induce the formation of pores (10). The formation of such pores, which have high permeability to Na+, K+, and alkali metal ions, leads to the disruption of proton motive force, alterations in cell envelope homeostasis, and ultimately cell death (7). The emergence of resistant isolates in the clinic is generally due to spontaneous mutations in genes and pathways involved in bacterial cell membrane (CM) and cell wall biosynthesis and modification (11). The most common resistance mechanism involves the alteration of the cell surface charge leading to the repulsion of the anionic daptomycin molecules (12). This primarily occurs via gain-of-function nonsynonymous mutations in the phosphatidylglycerol lysyltransferase MprF, producing positively charged lysyl-phosphatidylglycerol, a major component of the bacterial membrane (13, 14). Similarly, overexpression of the dlt operon, encoding proteins required for the incorporation of d-alanine in teichoic acids and resulting in a net increase in positive charge, has been described for daptomycin-resistant S. aureus (15–17). A second major resistance mechanism involves changes in the membrane composition via the alteration of phospholipid membrane metabolism. As such, mutations in phosphatidyltransferase PgsA and in cardiolipin synthase Cls2 provoke a decrease in the amount of phosphatidylglycerol in the membrane, impacting fluidity and ultimately reducing daptomycin binding and efficacy (18, 19). A third known resistance mechanism involves alteration of complex transcriptional regulatory networks governing the cell envelope stress response and membrane homeostasis (20). Decreased or increased transcription of the walKR or vraSR two-component regulatory system, respectively, has been linked to increased daptomycin resistance in S. aureus (12, 21–23). Additionally, polymorphisms in RNA polymerase subunits RpoB and RpoC (19, 24) have been associated with a daptomycin resistance phenotype, but those seem to indirectly affect the expression of the dlt operon or other cell wall biosynthesis pathways, thus explaining the characteristic increased cell wall thickness (25, 26).
To understand better the cell wall and physiologic dynamic in S. aureus that defines susceptibility and resistance to daptomycin, we screened a saturating bank of transposon insertions in S. aureus HG003 for increased resistance likely resulting from a loss-of-function mutation. We isolated, identified, validated, and recapitulated the loss-of-function mutations in two genes that, when mutated, resulted in increased daptomycin resistance. One codes for a hypothetical lipoprotein (SAOUHSC_00362) and the second codes for an alkaline shock protein (SAOUHSC_02441, Asp23). These genes were further characterized for their roles in the resistance and tolerance of S. aureus to daptomycin and other antimicrobials.
RESULTS
Identification of S. aureus transposon mutants with increased MIC to daptomycin.
We first determined the MIC of daptomycin for the S. aureus HG003 transposon mutant library pool (which we described previously to consist of 71,700 unique insertions) (27) and for a control subpool composed of 10 independent transposon mutants selected randomly. The growth of the mutants in the control pool, consisting of insertions not expected to be involved in daptomycin resistance, was inhibited by 0.25 μg/ml of daptomycin (Fig. 1). To determine whether mutants were present in the complete pool that exhibited the increased daptomycin resistance, it was grown in the presence of 0.25 μg/ml of drug (Fig. 1). We further enriched for the desired mutants by subculturing the outgrowth of the MIC assay in fresh medium supplemented with daptomycin (1 μg/ml). After plating and overnight growth on brain heart infusion (BHI) agar plates, 29 S. aureus colonies were confirmed to possess increased daptomycin MICs. The genomes of these strains were sequenced. Most possessed insertions in one of two genes, either SAOUHSC_00362 (n = 20), encoding a hypothetical lipoprotein (named here Dsp1 for daptomycin susceptibility protein 1), or SAOUHSC_02441 (n = 6), encoding an alkaline shock protein, Asp23. Additional transposon insertions were also detected in the gene SAOUHSC_01975 (n = 1), encoding a hypothetical protein, and in an intergenic region (n = 2).
FIG 1.

Transposon library antibiotic treatment and transposon mutant identification. A 10-μl aliquot of an overnight culture of the transposon library (library pool) and a pool generated from 10 independent mutants (control pool), each containing 106 CFU, were inoculated into a final volume of 200 μl Muller-Hinton broth in a 96-well plate broth microdilution format. DAP MIC was determined by the growth of the control pool (MIC-C). The contents of the library pool growing at 0.25 μg/ml DAP (1× MIC-C) were subcultured in 1 μg/ml (2× MIC-L). This culture was diluted and plated on BHI plates to recover isolated resistant mutants. Genomic DNA was harvested from individual colonies and subjected to whole-genome sequencing. Interrupted genes by transposon insertion were identified after alignment with the NCTC8325 whole genome.
Inactivation of either dsp1 or asp23 results in increased resistance to daptomycin and other antimicrobials.
An analysis of all S. aureus genomes available in the GenBank database suggested that dsp1 and asp23 are genes in this species and are present in all genomes. For both genes, we constructed markerless null mutants in S. aureus RN4220 for follow up analysis. In this background, increased daptomycin resistance was also observed, indicating that the resistance initially observed was unlikely to result from polar effects of transposon insertion. The Δdsp1 and Δasp23 null mutants exhibited 2-fold increases in the daptomycin MIC compared to that of the parental RN4220 (Table 1 ), and neither gene deletion was associated with a discernible growth defect in the absence of antibiotic selection (Fig. 2).
TABLE 1.
Antibiotic susceptibilities of the mutant strains
| Strain | MIC (μg/ml)a |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DAP | VAN | BAC | CST | NIS | PMB | CIP | CHL | ERY | GEN | LZD | OXA | TET | |
| RN4220 | 1 | 1 | 32 | 256 | 128 | 32 | 1 | 16 | 0.5 | 1 | 4 | 0.5 | 2 |
| Δdsp1 | 2 | 2 | 64 | 512 | 256 | 64 | 1 | 16 | 0.5 | 1 | 4 | 0.5 | 2 |
| Δasp23 | 2 | 1 | 64 | 512 | 256 | 64 | 1 | 16 | 0.5 | 1 | 4 | 0.5 | 2 |
DAP, daptomycin; VAN, vancomycin; BAC, bacitracin; CST, colistin; NIS, nisin; PMB, polymyxin B; CIP, ciprofloxacin; CHL, chloramphenicol; ERY, erythromycin; GEN, gentamicin; LZD, linezolid; OXA, oxacillin; TET, tetracycline.
FIG 2.

Growth kinetics of Δdsp1 (A to D), Δasp23 (E to H), and complemented strains upon challenge with 1 µg/ml DAP (B and F) or 1 µg/ml VAN (D and H). TSB CaCl2 was used as a control for DAP challenge (A and E), and plain TSB was used as a control for VAN challenge (C and G). The data are median values from three experiments for each condition, and error bars represent the standard deviations.
Interestingly, these mutants also showed increased resistance to various CAMPs (i.e., bacitracin, colistin, nisin, and polymyxin). Unexpectedly, the Δdsp1 mutant (but not the asp23 mutant) also showed 2-fold increased resistance to vancomycin (Table 1, Fig. 2D).
Overexpression of dsp1 or asp23 causes increased susceptibility to daptomycin.
To unambiguously associate Δdsp1 and Δasp23 with daptomycin resistance, we complemented the deletions with wild-type genes cloned into the xylose-inducible expression vector pEPSA5. Growth curves of the Δdsp1 and Δasp23 strains, with an empty pEPSA5, showed that the carriage of this shuttle vector has no effect on the ability of these strains to replicate in the presence of 1 µg/ml of daptomycin, even in the presence of xylose (Fig. 3A and C). When complemented with their respective wild-type alleles cloned into pEPSA5 (Δdsp1_comp and Δasp23_comp), both strains retained the ability to grow in 1 µg/ml of daptomycin in the absence of xylose induction. However, in the presence of 0.25% or greater xylose, both the Δdsp1_comp and Δasp23_comp strains were inhibited by 1 µg/ml of daptomycin.
FIG 3.

Overexpression of genes dsp1 (B) and asp23 (D) in the complemented strains in the presence of DAP (1 µg/ml). Mutant strains with empty vector pEPSA5 were used as a control (A and C). The data are median values from three experiments for each condition, and error bars represent the standard deviations.
Inactivation of dsp1 or asp23 results in reduced killing by vancomycin and daptomycin.
We performed killing assays for both knockout mutants in the presence of 5 µg/ml daptomycin (Fig. 4A). Starting from a 108 CFU/ml inoculum, the survival of the parental RN4200 after 2 h was below our technical limit of detection (1,000 CFU/ml). Strikingly, under the same conditions, >105 CFU/ml was recovered for both mutants. Even after 24 h, the Δdsp1 and Δasp23 mutants both retained readily detectable viable cells (>104 CFU/ml). Interestingly, the increased tolerance of S. aureus Δdsp1 and Δasp23 mutants was found not to be specific to the lipopeptide daptomycin, as a similar phenotype was observed when the cells were challenged with 5 μg/ml of vancomycin (Fig. 4B). For gene dsp1, an increased tolerance to DAP and VAN was observed in a transposon mutant (Tndsp1) obtained from the Nebraska Transposon Mutant Library (NTML) (28) compared to that of wild-type JE2 (Fig. 5), a methicillin-resistant strain derived from the highly characterized community-associated S. aureus USA300 LAC (28). This was independent from their resistance phenotype, as both Tndsp1 and JE2 showed no change in the MIC for DAP (1 μg/ml) or VAN (2 μg/ml). This suggests that gene dsp1, core to S. aureus, is generally involved in vancomycin and daptomycin tolerance in this species. This is likely true for gene asp23 but was not tested experimentally, as no transposon mutants were available in the NTML.
FIG 4.

Increased survival of Δdsp1 and Δasp23 mutants in DAP (A) and VAN (B) exposure assays. Unpaired t tests were used to determine statistical significance after 24-h treatments. For daptomycin: Δdsp1, P = 0.0197 and Δasp23, P = 0.0171; for vancomycin: Δdsp1, P = 0.0243 and Δasp23, P = 0.0193. The experiment was performed in three independent replicates; means and standard deviations are indicated. Detection limit, 1,000 CFU/ml.
FIG 5.

Survival of USA300 wild-type JE2 and mutant Tndsp1 strains in 5 µg/ml and 10 µg/ml DAP (A and C) and 5 µg/ml and 10 µg/ml VAN (B and D) exposure assays. Both JE2 and Tndsp1 strains show similar survival when challenged with 5 µg/ml of antibiotics, while the Tndsp1 strain showed significantly increased tolerance to 10 µg/ml of VAN or DAP. Unpaired t tests were used to determine statistical significance after 24-h treatments. For 10 µg/ml daptomycin: Tndsp1, P = 0.0213; for 10 µg/ml vancomycin: Tndsp1, P = 0.0444. The experiment was performed in three independent replicates; means and standard deviations are indicated. Detection limit, 1,000 CFU/ml.
Mutant strains show reduced autolytic activity, similar susceptibility to lysostaphin, and no alteration in cell surface charge.
Induced autolysis assays were performed to identify the basis for the observed increased resistance to DAP of the mutant strains. The susceptibility to cell lysis was characterized by monitoring the kinetics of Triton X-100-induced autolysis over time. Wild-type RN4220 showed ∼30% cell survival at 3 h in the presence of 0.05% Triton X-100. In contrast, more than 50% of the dsp1 and asp23 mutants survived after 3 h of treatment (Fig. 6A), suggesting enhanced DAP tolerance may result from alterations to the cell envelope. Alternatively, for lysostaphin-induced autolysis, the mutant strains exhibited similar susceptibility to the parental strain, with ∼40% survival after 3 h (Fig. 6B). These results suggest that the Δdsp1 and Δasp23 mutants possess membrane modifications or other cell wall changes that result in DAP resistance and that peptidoglycan itself does not appear to be involved. Therefore, the possibility that the alteration of cell surface charge was be responsible for repelling DAP was evaluated by quantifying the association of the highly cationic molecule, cytochrome c, with the staphylococcal surface (Fig. 6C). However, no difference in cytochrome c binding was observed.
FIG 6.
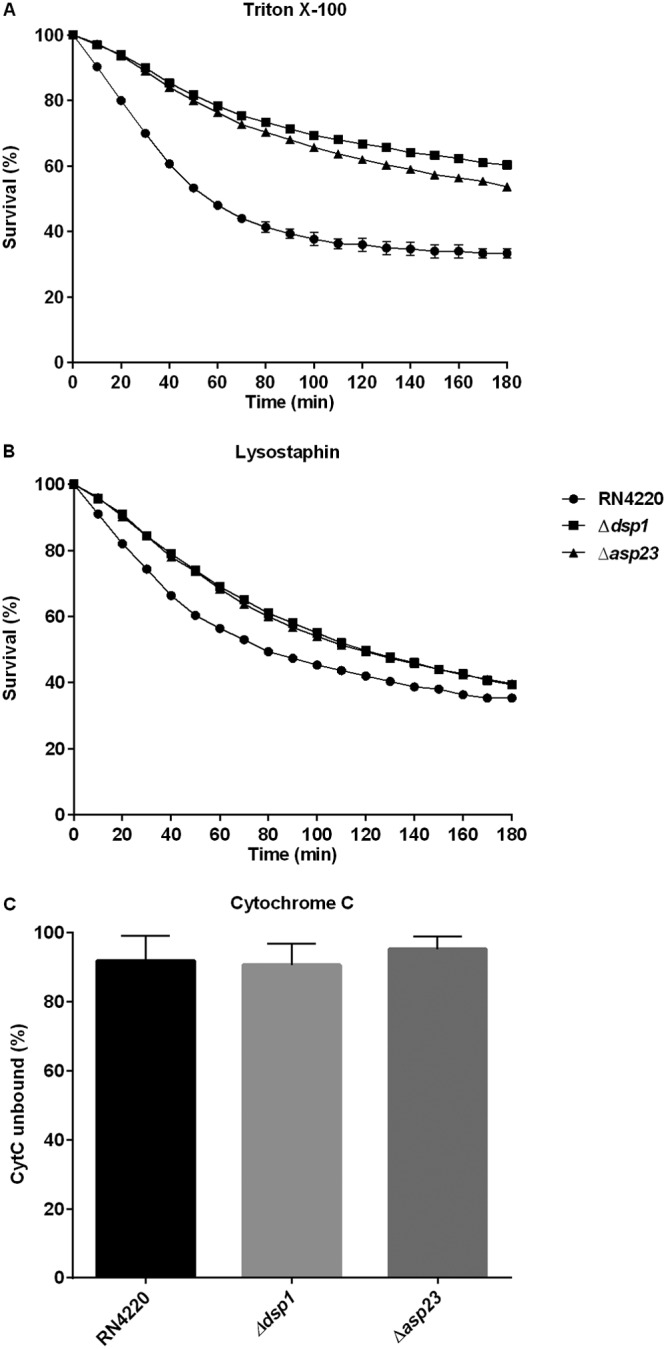
Ability of Δdsp1 and Δasp23 mutants to induced autolysis in the presence of lysostaphin (A) or Triton X-100 (B) and to repulse cationic cytochrome c (C). Unpaired t tests were used to determine statistical significance after 24-h treatment with Triton X-100. Δdsp1, P < 0.0001 and Δasp23, P < 0.0001. The data are mean values from three experiments for each condition, and error bars represent the standard deviations.
DISCUSSION
Prior use of VAN during treatment of VAN-intermediate strains of S. aureus (VISA) has been linked to an increased resistance to DAP (11, 29), suggesting that some alterations of the cell wall have the ability to affect the susceptibility to both drugs. DAP is a drug of last resort for treating multidrug-resistant S. aureus. Therefore, the development of resistance during therapy is a serious threat, as it leaves clinicians without effective options for treatment. Despite numerous studies examining DAP resistance evolution, both in vitro and in vivo, the mechanisms by which bacteria become resistant to DAP are not fully understood (12). Mutations in several genes have been described to trigger DAP resistance, although none of these mutations are sufficient to confer high-level resistance per se (30).
Here, two genes not previously associated with daptomycin resistance were identified in a screen of transposon insertion mutants in S. aureus HG003. A sequence analysis showed that one gene encodes a hypothetical lipoprotein (SAOUHSC_00362) and the other (asp23) an alkaline-shock protein (SAOUHSC_02441). The staphylococcal alkaline shock protein 23 (Asp23) is one of the most abundant proteins expressed by S. aureus. It is a 23-kDa protein that accumulates in the soluble protein fraction of S. aureus cells following alkaline shock (31). The stress response gene asp23 is the last gene in a four-gene operon transcribed from three different σB-dependent promoters (31, 32). The deletion of asp23 has been linked to increased transcription of cell wall stress-associated genes, and most of the upregulated genes were previously shown to also be induced in the presence of VAN (33), including the two-component system VraSR and peptidylprolyl isomerase PrsA (34). Recently, a point mutation in asp23, among other mutations, was reported in a DAP-R strain that resulted from serially passaging S. aureus in increasing DAP concentrations. A C-to-T transition (Glu47Lys) was observed in asp23, but not directly associated with the DAP-R phenotype (30). We show that asp23 loss of function increases DAP resistance and also confers increased tolerance to both DAP and VAN. On the basis of the known function of Asp23 in stress response, we hypothesize that a deletion of asp23 in the context of DAP treatment results in transcriptional changes that indirectly increase DAP resistance.
Beyond DAP and VAN, the Δasp23 and Δdsp1 mutants also exhibited higher tolerance to all CAMPs tested. CAMPs are widely distributed in nature and constitute key effectors of innate immune responses to infection in organisms ranging from mammals to plants. In some respects, DAP resembles CAMPs because of its peptide content, charge, and mode of action targeting membrane function (35). Mishra and coworkers (36) provided evidence that prior exposure to endovascular host cationic peptides could conceivably drive the selection of DAP-R strains. Cross-resistance between DAP and other CAMPs that target the bacterial cell membrane has also been reported, especially in association with mutations that appear to lead to a gain of function of mprF. MprF modifies anionic phospholipids and thereby introduces positive charges into the membrane surface, which reduce the affinity for DAP and CAMPs (14). The overexpression of the dtl operon causes increases in net positive charges in the cell membrane and has also been described in DAP-R strains (15–17).
As most adaptations in DAP-R strains have been associated with cell membrane and cell wall changes, induced autolysis assays were conducted using Triton X-100 or lysostaphin challenge. Bacterial cell autolysis is regulated by various mechanisms, including the modification of the cell wall peptidoglycan and the regulation of the expression of genes associated with cell wall hydrolase activities (37, 38). The ability of the nonionic surfactant Triton X-100 to induce cellular lysis is associated with the disruption of the plasma membrane, triggering the autolysis of the peptidoglycan layer by cellular autolysins (39). Lysostaphin has a direct effect in the cell wall peptidoglycan by cleaving the cross-linking pentaglycine bridges (40). Our results showed that both dsp1 and asp23 deletions are associated with decreased bacterial autolysis induced by the nonionic detergent, Triton X-100, but no effect was observed in controlling lysostaphin-induced lysis. These data support the hypothesis that DAP resistance in the mutant strains is likely caused by changes in the cell membrane, perhaps by modulating the abundance of membrane-associated proteins and thus affecting membrane stability or fluidity. Here, we described two genes that upon inactivation lead to an increased tolerance to both DAP and VAN. Studies are under way to further elucidate the mechanisms by which dsp1 and asp23 contribute to the susceptibility of S. aureus to these antimicrobials.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions.
The bacterial strains and plasmids are listed in Table 2. The transposon library constructed in S. aureus HG003 using the Bursa aurealis system has been previously described (27). S. aureus and Escherichia coli strains were grown in tryptic soy broth (TSB; BD Diagnostics Systems) and Luria broth (LB; BD Diagnostics Systems) medium, respectively, at 37°C under continuous shaking, unless indicated otherwise. E. coli DH5α (Invitrogen) was used as the cloning host for pEPSA5 and pKOR1 plasmids that were subsequently transferred to S. aureus RN4220. Antibiotic markers were selected when appropriate with 50 μg/ml ampicillin (Amp; Sigma-Aldrich) and 10 μg/ml chloramphenicol (Chl; Sigma-Aldrich). Medium broth was supplemented with CaCl2 (50 μg/ml; Sigma-Aldrich) when DAP was tested. Anhydrotetracycline (ATc; Sigma-Aldrich) at a final concentration of 1 μg/ml was used to select mutant strains. Genes cloned into pEPSA5 had their expression induced with xylose (0.5%, Sigma-Aldrich).
TABLE 2.
Strains and plasmids used in this study
| Strain or plasmid | Descriptiona | Source or reference |
|---|---|---|
| Strains | ||
| Escherichia coli DH5α | Cloning host | Invitrogen |
| Staphylococcus aureus | ||
| HG003 | Wild type; NCTC8325 derivative; rsbU and tcaR repaired | 49 |
| RN4220 | Wild type; NCTC8325 derivative; restriction deficient, prophage cured | 50 |
| RN4220 Δdsp1 | Locus SAOUHSC_00362 deletion mutant | This study |
| RN4220 Δasp23 | asp23 deletion mutant | This study |
| RN4220 Δdsp1_Comp | Δdsp1 complemented strain | This study |
| RN4220 Δasp23_Comp | Δasp23 complemented strain | This study |
| JE2 | Wild type; derived from S. aureus USA300 LAC | NTML (28) |
| Tndsp1 | Transposon insertion at position 34 of dsp1 gene in JE2 background | NTML (28) |
| Plasmids | ||
| pKOR1 | E. coli/S. aureus shuttle vector; thermosensitive origin of replication; ATc induced counterselection; Ampr, Chlr | 42 |
| pEPSA5 | E. coli/S. aureus shuttle vector; expression vector; Ampr, Chlr | 45 |
Ampr, ampicillin resistant; Chlr, chloramphenicol resistant.
Transposon mutant library screen.
The mariner-based Bursa aurealis transposon insertion system was previously used to construct a mutagenized library in HG003. This library has been described to consist of 71,700 unique insertions in the HG003 chromosome, with 57,191 insertions occurred within coding regions (27), and it was used in this work as described. Frozen aliquots of the transposon mutant library pool and a “little pool” generated from 10 independent transposon mutants selected at random were thawed and diluted 1:1,000 into 100 ml of Mueller-Hinton (MH; Moltox) broth and grown overnight at 37°C. The culture was diluted 1:10 into 10 ml of MH broth supplemented with CaCl2. The cultures were further diluted (1:20) into 200 μl fresh MH CaCl2 broth with 2-fold dilutions of DAP (16 to 0.008 μg/ml; Sigma-Aldrich) in 96-well plates at 37°C for 24 h. Two rounds of selection in DAP were performed under the same conditions. The MIC of the little pool was determined, and wells above this MIC from the full mutant library were diluted 1:100 and subcultured twice in a higher concentration (2× the MIC) of DAP to increase the sensitivity of the detection of mutants showing decreased susceptibility to DAP. The mutants were serially diluted 1:108 into phosphate-buffered saline (PBS) and then plated on tryptic Soy agar (TSA) plates to recover isolated mutant strains. Two independent experiments were performed.
Molecular cloning and DNA isolation.
The primers used in this study were synthesized by Integrated DNA Technologies (Table 3). Qiagen kits were used for the isolation of bacterial genomic DNA and plasmids. For PCR product cleanup, ExoSAP-IT (Affymetrix; USB) was used. Phusion high-fidelity DNA polymerase, PCR reagents, restriction enzymes, calf-intestinal alkaline phosphatase (CIP), and T4 DNA ligase from New England BioLabs and Gateway BP Clonase II enzyme mix from Invitrogen were used according to the manufacturer’s protocols.
TABLE 3.
Oligonucleotides used in this study
| Primer name | Sequence | Reference or source |
|---|---|---|
| Knockout construction | ||
| 362-51-attB1 | GGGGACAAGTTTGTACAAAAAAGCAGGCTTGAAGAGCAATTGATTAATAAAGG | This study |
| 362-31-OL | AATGATGGCTTTCAGAATGTAATATAACTCCTTAGTTTATCT | This study |
| 362-32-attB2 | GGGGACCACTTTGTACAAGAAAGCTGGGTTAAAGATGAAATCAGTGCGAATCC | This study |
| 362-52-OL | GAGTTATATTACATTCTGAAAGCCATCATTGCGAAA | This study |
| 2441-51-attB1 | GGGGACAAGTTTGTACAAAAAAGCAGGCTAGGTTTTATTTACAAAAACTAGGG | This study |
| 2441-31-OL | TTTTTATCGAATAACAATAGATTCTCCTTTTACTTG | This study |
| 2441-32-attB2 | GGGGACCACTTTGTACAAGAAAGCTGGGTGAACGAAGATAGTACTCAAATTAG | This study |
| 2441-52-OL | AAAGGAGAATCTATTGTTATTCGATAAAAAGGGCTT | This study |
| Complementation construction | ||
| 362-Fw-EcoRI | GCGAGAATTCAGGAGGTACATTATGAAAAAGAGATTACTAC | This study |
| 362-Rev-BamHI | GCGAGGATCCTTATTTATCGATAACATCACTCTTG | This study |
| 2441-Fw-EcoRI | GCGAGAATTCAGGAGGTCTATTATGACTGTAGATAACAATAAAG | This study |
| 2441-Rev-BamHI | GCGAGGATCCTTATTGTAAACCTTGTCTTTCTTGG | This study |
Whole-genome sequencing for identification of mutated genes.
Genomic DNA was isolated with the DNeasy blood and tissue kit (Qiagen) according to the manufacturer’s instructions. Sequencing libraries were prepared using an Illumina Nextera XT Library Prep kit and subjected to 250-bp paired-end sequencing on the MiSeq platform at the Massachusetts Eye and Ear Infirmary Ocular Genomics Institute (Boston, MA, USA). Sequence reads were assembled de novo (CLC Genomics Workbench 8.0.3; Cambridge, MA, USA) and aligned to the transposon sequence using Geneious R8 (41).
Construction of mutant strains and genetic complementation.
Gene deletions from the genome of S. aureus RN4220 were made by homologous recombination with plasmid pKOR1 according Bae and Schneewind (42) with minor modifications. Briefly, two DNA segments of ∼1kb in length up- and downstream of the candidate genes were PCR-amplified from the chromosomal DNA of S. aureus HG003. Overlap regions were introduced in the primer sequence next to the gene to be deleted, and att recombination sites were introduced at the distal ends, as listed in Table 3. The two segments flanking each target gene were recombined by SOEing PCR according to Horton et al. (43). The PCR products were cloned individually into pKOR1 by recombination reactions (Gateway BP Clonase II enzyme mix). The resulting plasmids were transformed into E. coli DH5α chemically competent cells according to the manufacturer’s recommendations and subsequently into S. aureus RN4220 competent cells via electroporation (44). Further allelic replacement was achieved as described previously (42) and verified by Sanger sequence analysis.
To complement the mutant strains, the target genes were PCR amplified using the primers listed in Table 3 and cloned into the pEPSA5 shuttle vector (45) using restriction endonucleases EcoRI and BamHI.
MIC determination.
MICs were determined using the CLSI broth microdilution assay guidelines (46, 47) for different antimicrobials against the parent strain S. aureus RN4220 and isogenic mutants. All MIC assays were performed in triplicates.
Killing curve.
Killing curve assays were performed in the presence of DAP and VAN. Briefly, an overnight culture was diluted 1:100 into MH or MH CaCl2 broth and grown at 37°C to mid-log phase (optical density at 600 nm [OD600] of ∼0.3). Cultures were exposed to 5 µg/ml DAP or VAN, 5× the MIC of parent strain, and incubated at 37°C for 24 h. CFU after 0, 2, 4, 6, and 24 h were quantified by the track dilution method using six 10-fold dilutions and 10 µl aliquot for plating.
Growth curve.
Growth curves were performed with S. aureus RN4220, its mutants, and complemented strains. RN4220 transformed with empty pEPSA5 vector was also included in the analysis. Overnight cultures were diluted and adjusted to an OD600 of 0.08 to 0.1 (∼108 CFU/ml). The cultures were inoculated (1:20) in 200 μl fresh TSB or TSB CaCl2 broth and challenged with 2-fold dilutions of DAP or VAN (4 to 0.125 μg/ml) in 96-well plates at 37°C for 36 h. The OD600 values were monitored every hour using a BioTek plate reader. Chloramphenicol (10 μg/ml) and xylose (0.5%) were included to maintain pEPSA5 plasmid vector and to induce the expression of cloned genes, respectively. Growth curves were generated by plotting the OD600 values over time. The data are reported as mean values from four independent experiments.
Induced autolysis assay.
For Triton X-100-induced autolysis, cells were grown in TSB to an OD600 of 1.0 and chilled on ice before harvesting. After washing the cells with ice-cold water, the bacterial pellets were resuspended in the same volume of 50 mM Tris-HCl (pH 7.5) containing 0.05% Triton X-100 (J. T. Baker Inc.). Induced autolysis was measured spectrophotometrically during incubation at 37°C as a decrease in OD600 over time. For lysostaphin (BioVendor)-induced autolysis assays, strains were also grown to an OD600 of 1.0 and harvested by centrifugation. Cells were washed with water and resuspended in PBS supplemented with 200 ng/ml lysostaphin. Cell autolysis was determined as described above. The results were normalized to the OD600 at time zero (OD0) and the percent survival at indicated times was determined.
Cytochrome c binding assay.
The relative positive surface charge of S. aureus strains was determined by quantifying the association of the positively charged molecule cytochrome c (Sigma) to the staphylococcal surface as previously described (48). Briefly, after overnight growth in TSB, cultures were diluted 1:100 in fresh medium and allowed to regrow to logarithmic phase (OD600 of ∼0.3). After centrifugation, the cells were washed twice with MOPS (morpholinepropanesulfonic acid) buffer (20 mM, pH 7.0, the bacterial suspension was adjusted to an OD600 of 1.0, and 1 ml aliquots were harvested. The cell pellets were resuspended in 200 µl MOPS buffer and 50 µl of cytochrome c solution was added (equine heart, 2.5 mg/ml in MOPS buffer; Sigma). After 10 min at room temperature, the samples were harvested at 12,000 × g, supernatants were recovered, and the OD530 was measured spectrophotometrically. The experiment was repeated in triplicates.
ACKNOWLEDGMENTS
This work was supported by CNPq and has been funded in part by DHHS/NIH/NIAID grants AI072360 and AI083214 (Harvard-wide Program on Antibiotic Resistance) (M.S.G.).
We declare no competing financial interests.
REFERENCES
- 1.Rivera AM, Boucher HW. 2011. Current concepts in antimicrobial therapy against select Gram-positive organisms: methicillin-resistant Staphylococcus aureus, penicillin-resistant pneumococci, and vancomycin-resistant enterococci. Mayo Clin Proc 86:1230–1243. doi: 10.4065/mcp.2011.0514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Sievert DM, Rudrik JT, Patel JB, McDonald LC, Wilkins MJ, Hageman JC. 2008. Vancomycin-resistant Staphylococcus aureus in the United States, 2002–2006. Clin Infect Dis 46:668–674. doi: 10.1086/527392. [DOI] [PubMed] [Google Scholar]
- 3.Sader HS, Mendes RE, Duncan LR, Pfaller MA, Flamm RK. 2018. Antimicrobial activity of dalbavancin against Staphylococcus aureus with decreased susceptibility to glycopeptides, daptomycin, and/or linezolid from U.S. medical centers. Antimicrob Agents Chemother 62: e02397-17. doi: 10.1128/AAC.02397-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Joana S, Pedro P, Elsa G, Filomena M. 2013. Is vancomycin MIC creep a worldwide phenomenon? Assessment of S. aureus vancomycin MIC in a tertiary university hospital. BMC Res Notes 6:65. doi: 10.1186/1756-0500-6-65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lai CC, Sheng WH, Wang JT, Cheng A, Chuang YC, Chen YC, Chang SC. 2013. Safety and efficacy of high-dose daptomycin as salvage therapy for severe gram-positive bacterial sepsis in hospitalized adult patients. BMC Infect Dis 13:66. doi: 10.1186/1471-2334-13-66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Parra-Ruiz J, Hernandez-Quero J. 2012. Safety and efficacy of daptomycin therapy in older adults with pluripathology. Enferm Infecc Microbiol Clin 30 Suppl 1:50–53. (In Spanish.) doi: 10.1016/S0213-005X(12)70072-1. [DOI] [PubMed] [Google Scholar]
- 7.Humphries RM, Pollett S, Sakoulas G. 2013. A current perspective on daptomycin for the clinical microbiologist. Clin Microbiol Rev 26:759–780. doi: 10.1128/CMR.00030-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mulcahy LR, Burns JL, Lory S, Lewis K. 2010. Emergence of Pseudomonas aeruginosa strains producing high levels of persister cells in patients with cystic fibrosis. J Bacteriol 192:6191–6199. doi: 10.1128/JB.01651-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Straus SK, Hancock RE. 2006. Mode of action of the new antibiotic for Gram-positive pathogens daptomycin: comparison with cationic antimicrobial peptides and lipopeptides. Biochim Biophys Acta 1758:1215–1223. doi: 10.1016/j.bbamem.2006.02.009. [DOI] [PubMed] [Google Scholar]
- 10.Zhang T, Muraih JK, Tishbi N, Herskowitz J, Victor RL, Silverman J, Uwumarenogie S, Taylor SD, Palmer M, Mintzer E. 2014. Cardiolipin prevents membrane translocation and permeabilization by daptomycin. J Biol Chem 289:11584–11591. doi: 10.1074/jbc.M114.554444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bayer AS, Schneider T, Sahl HG. 2013. Mechanisms of daptomycin resistance in Staphylococcus aureus: role of the cell membrane and cell wall. Ann N Y Acad Sci 1277:139–158. doi: 10.1111/j.1749-6632.2012.06819.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Tran TT, Munita JM, Arias CA. 2015. Mechanisms of drug resistance: daptomycin resistance. Ann N Y Acad Sci 1354:32–53. doi: 10.1111/nyas.12948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Yang SJ, Mishra NN, Rubio A, Bayer AS. 2013. Causal role of single nucleotide polymorphisms within the mprF gene of Staphylococcus aureus in daptomycin resistance. Antimicrob Agents Chemother 57:5658–5664. doi: 10.1128/AAC.01184-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ernst CM, Peschel A. 2011. Broad-spectrum antimicrobial peptide resistance by MprF-mediated aminoacylation and flipping of phospholipids. Mol Microbiol 80:290–299. doi: 10.1111/j.1365-2958.2011.07576.x. [DOI] [PubMed] [Google Scholar]
- 15.Bertsche U, Yang SJ, Kuehner D, Wanner S, Mishra NN, Roth T, Nega M, Schneider A, Mayer C, Grau T, Bayer AS, Weidenmaier C. 2013. Increased cell wall teichoic acid production and D-alanylation are common phenotypes among daptomycin-resistant methicillin-resistant Staphylococcus aureus (MRSA) clinical isolates. PLoS One 8:e67398. doi: 10.1371/journal.pone.0067398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cafiso V, Bertuccio T, Purrello S, Campanile F, Mammina C, Sartor A, Raglio A, Stefani S. 2014. dltA overexpression: a strain-independent keystone of daptomycin resistance in methicillin-resistant Staphylococcus aureus. Int J Antimicrob Agents 43:26–31. doi: 10.1016/j.ijantimicag.2013.10.001. [DOI] [PubMed] [Google Scholar]
- 17.Yang SJ, Kreiswirth BN, Sakoulas G, Yeaman MR, Xiong YQ, Sawa A, Bayer AS. 2009. Enhanced expression of dltABCD is associated with the development of daptomycin nonsusceptibility in a clinical endocarditis isolate of Staphylococcus aureus. J Infect Dis 200:1916–1920. doi: 10.1086/648473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Khatib TO, Stevenson H, Yeaman MR, Bayer AS, Pokorny A. 2016. Binding of daptomycin to anionic lipid vesicles is reduced in the presence of lysyl-phosphatidylglycerol. Antimicrob Agents Chemother 60:5051–5053. doi: 10.1128/AAC.00744-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Peleg AY, Miyakis S, Ward DV, Earl AM, Rubio A, Cameron DR, Pillai S, Moellering RC Jr, Eliopoulos GM. 2012. Whole genome characterization of the mechanisms of daptomycin resistance in clinical and laboratory derived isolates of Staphylococcus aureus. PLoS One 7:e28316. doi: 10.1371/journal.pone.0028316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Miller WR, Bayer AS, Arias CA. 2016. Mechanism of action and resistance to daptomycin in Staphylococcus aureus and enterococci. Cold Spring Harb Perspect Med 6:a026997. doi: 10.1101/cshperspect.a026997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kuroda M, Kuroda H, Oshima T, Takeuchi F, Mori H, Hiramatsu K. 2003. Two-component system VraSR positively modulates the regulation of cell-wall biosynthesis pathway in Staphylococcus aureus. Mol Microbiol 49:807–821. [DOI] [PubMed] [Google Scholar]
- 22.Mwangi MM, Wu SW, Zhou Y, Sieradzki K, de Lencastre H, Richardson P, Bruce D, Rubin E, Myers E, Siggia ED, Tomasz A. 2007. Tracking the in vivo evolution of multidrug resistance in Staphylococcus aureus by whole-genome sequencing. Proc Natl Acad Sci U S A 104:9451–9456. doi: 10.1073/pnas.0609839104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Camargo IL, Neoh HM, Cui L, Hiramatsu K. 2008. Serial daptomycin selection generates daptomycin-nonsusceptible Staphylococcus aureus strains with a heterogeneous vancomycin-intermediate phenotype. Antimicrob Agents Chemother 52:4289–4299. doi: 10.1128/AAC.00417-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Friedman L, Alder JD, Silverman JA. 2006. Genetic changes that correlate with reduced susceptibility to daptomycin in Staphylococcus aureus. Antimicrob Agents Chemother 50:2137–2145. doi: 10.1128/AAC.00039-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Cui L, Isii T, Fukuda M, Ochiai T, Neoh HM, Camargo IL, Watanabe Y, Shoji M, Hishinuma T, Hiramatsu K. 2010. An RpoB mutation confers dual heteroresistance to daptomycin and vancomycin in Staphylococcus aureus. Antimicrob Agents Chemother 54:5222–5233. doi: 10.1128/AAC.00437-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bæk KT, Thøgersen L, Mogenssen RG, Mellergaard M, Thomsen LE, Petersen A, Skov S, Cameron DR, Peleg AY, Frees D. 2015. Stepwise decrease in daptomycin susceptibility in clinical Staphylococcus aureus isolates associated with an initial mutation in rpoB and a compensatory inactivation of the clpX gene. Antimicrob Agents Chemother 59:6983–6991. doi: 10.1128/AAC.01303-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Valentino MD, Foulston L, Sadaka A, Kos VN, Villet RA, Santa Maria J Jr, Lazinski DW, Camilli A, Walker S, Hooper DC, Gilmore MS. 2014. Genes contributing to Staphylococcus aureus fitness in abscess- and infection-related ecologies. mBio 5:e01729-14. doi: 10.1128/mBio.01729-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Fey PD, Endres JL, Yajjala VK, Widhelm TJ, Boissy RJ, Bose JL, Bayles KW. 2013. A genetic resource for rapid and comprehensive phenotype screening of nonessential Staphylococcus aureus genes. mBio 4:e00537-12. doi: 10.1128/mBio.00537-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.van Hal SJ, Paterson DL, Gosbell IB. 2011. Emergence of daptomycin resistance following vancomycin-unresponsive Staphylococcus aureus bacteraemia in a daptomycin-naive patient–a review of the literature. Eur J Clin Microbiol Infect Dis 30:603–610. doi: 10.1007/s10096-010-1128-3. [DOI] [PubMed] [Google Scholar]
- 30.Muller A, Grein F, Otto A, Gries K, Orlov D, Zarubaev V, Girard M, Sher X, Shamova O, Roemer T, Francois P, Becher D, Schneider T, Sahl HG. 2018. Differential daptomycin resistance development in Staphylococcus aureus strains with active and mutated gra regulatory systems. Int J Med Microbiol 308:335–348. doi: 10.1016/j.ijmm.2017.12.002. [DOI] [PubMed] [Google Scholar]
- 31.Kuroda M, Ohta T, Hayashi H. 1995. Isolation and the gene cloning of an alkaline shock protein in methicillin resistant Staphylococcus aureus. Biochem Biophys Res Commun 207:978–984. doi: 10.1006/bbrc.1995.1281. [DOI] [PubMed] [Google Scholar]
- 32.Gertz S, Engelmann S, Schmid R, Ohlsen K, Hacker J, Hecker M. 1999. Regulation of sigmaB-dependent transcription of sigB and asp23 in two different Staphylococcus aureus strains. Mol Gen Genet 261:558–566. doi: 10.1007/s004380051001. [DOI] [PubMed] [Google Scholar]
- 33.Müller M, Reiß S, Schlüter R, Mäder U, Beyer A, Reiß W, Marles-Wright J, Lewis RJ, Pförtner H, Völker U, Riedel K, Hecker M, Engelmann S, Pané-Farré J. 2014. Deletion of membrane-associated Asp23 leads to upregulation of cell wall stress genes in Staphylococcus aureus. Mol Microbiol 93:1259–1268. doi: 10.1111/mmi.12733. [DOI] [PubMed] [Google Scholar]
- 34.McCallum N, Spehar G, Bischoff M, Berger-Bächi B. 2006. Strain dependence of the cell wall-damage induced stimulon in Staphylococcus aureus. Biochim Biophys Acta 1760:1475–1481. doi: 10.1016/j.bbagen.2006.06.008. [DOI] [PubMed] [Google Scholar]
- 35.Kelley WL, Lew DP, Renzoni A. 2012. Antimicrobial peptide exposure and reduced susceptibility to daptomycin: insights into a complex genetic puzzle. J Infect Dis 206:1153–1156. doi: 10.1093/infdis/jis485. [DOI] [PubMed] [Google Scholar]
- 36.Mishra NN, McKinnell J, Yeaman MR, Rubio A, Nast CC, Chen L, Kreiswirth BN, Bayer AS. 2011. In vitro cross-resistance to daptomycin and host defense cationic antimicrobial peptides in clinical methicillin-resistant Staphylococcus aureus isolates. Antimicrob Agents Chemother 55:4012–4018. doi: 10.1128/AAC.00223-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Rice KC, Firek BA, Nelson JB, Yang SJ, Patton TG, Bayles KW. 2003. The Staphylococcus aureus cidAB operon: evaluation of its role in regulation of murein hydrolase activity and penicillin tolerance. J Bacteriol 185:2635–2643. doi: 10.1128/JB.185.8.2635-2643.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zheng L, Yu C, Bayles K, Lasa I, Ji Y. 2007. Conditional mutation of an essential putative glycoprotease eliminates autolysis in Staphylococcus aureus. J Bacteriol 189:2734–2742. doi: 10.1128/JB.01806-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Cornett JB, Shockman GD. 1978. Cellular lysis of Streptococcus faecalis induced with Triton X-100. J Bacteriol 135:153–160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Schindler CA, Schuhardt VT. 1964. Lysostaphin: a new bacteriolytic agent for the Staphylococcus. Proc Natl Acad Sci U S A 51:414–421. doi: 10.1073/pnas.51.3.414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowitz S, Duran C, Thierer T, Ashton B, Meintjes P, Drummond A. 2012. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28:1647–1649. doi: 10.1093/bioinformatics/bts199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bae T, Schneewind O. 2006. Allelic replacement in Staphylococcus aureus with inducible counter-selection. Plasmid 55:58–63. doi: 10.1016/j.plasmid.2005.05.005. [DOI] [PubMed] [Google Scholar]
- 43.Horton RM, Cai Z, Ho SM, Pease LR. 1990. Gene splicing by overlap extension: tailor-made genes using the polymerase chain reaction. Biotechniques 8:528–535. [PubMed] [Google Scholar]
- 44.Grosser MR, Richardson AR. 2016. Method for preparation and electroporation of S. aureus and S. epidermidis. Methods Mol Biol 1373:51–57. doi: 10.1007/7651_2014_183. [DOI] [PubMed] [Google Scholar]
- 45.Forsyth RA, Haselbeck RJ, Ohlsen KL, Yamamoto RT, Xu H, Trawick JD, Wall D, Wang L, Brown-Driver V, Froelich JM, C KG, King P, McCarthy M, Malone C, Misiner B, Robbins D, Tan Z, Zhu Zy ZY, Carr G, Mosca DA, Zamudio C, Foulkes JG, Zyskind JW. 2002. A genome-wide strategy for the identification of essential genes in Staphylococcus aureus. Mol Microbiol 43:1387–1400. doi: 10.1046/j.1365-2958.2002.02832.x. [DOI] [PubMed] [Google Scholar]
- 46.Clinical and Laboratory Standards Institute. 2015. Performance standards for antimicrobial susceptibility testing; 25th informational supplement. M100-S25. Clinical and Laboratory Standards Institute, Wayne, PA. [Google Scholar]
- 47.Clinical and Laboratory Standards Institute. 2015. Methods for dilution antimicrobial susceptibility tests for bacterial that grow aerobically; approved standard—10th ed M07-A10 Clinical and Laboratory Standards Institute, Wayne, PA. [Google Scholar]
- 48.Berti AD, Baines SL, Howden BP, Sakoulas G, Nizet V, Proctor RA, Rose WE. 2015. Heterogeneity of genetic pathways toward daptomycin nonsusceptibility in Staphylococcus aureus determined by adjunctive antibiotics. Antimicrob Agents Chemother 59:2799–2806. doi: 10.1128/AAC.04990-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Herbert S, Ziebandt AK, Ohlsen K, Schafer T, Hecker M, Albrecht D, Novick R, Gotz F. 2010. Repair of global regulators in Staphylococcus aureus 8325 and comparative analysis with other clinical isolates. Infect Immun 78:2877–2889. doi: 10.1128/IAI.00088-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kreiswirth BN, Lofdahl S, Betley MJ, O'Reilly M, Schlievert PM, Bergdoll MS, Novick RP. 1983. The toxic shock syndrome exotoxin structural gene is not detectably transmitted by a prophage. Nature 305:709–712. doi: 10.1038/305709a0. [DOI] [PubMed] [Google Scholar]