

Abstract
Distancing is a type of emotion regulation that involves simulating a new perspective to alter the psychological distance and emotional impact of a stimulus. The effectiveness and versatility of distancing relative to other types of emotion regulation make it a promising tool for clinical applications. However, the neurocognitive mechanisms of this tactic are unclear, and inconsistencies in terminology and methods across studies make it difficult to synthesize the literature. To promote more effective research, we propose a taxonomy of distancing within the broader context of emotion regulation strategies; review the effects of this tactic; and offer a preliminary neurocognitive model describing key cognitive processes and their neural bases. Our model emphasizes three components—self-projection, affective self-reflection, and cognitive control. Additionally, we present results from a supporting meta-analysis of neuroimaging studies of distancing. These efforts are presented within the overarching goals of supporting effective applications of distancing in laboratory, clinical, and other real-world contexts, and advancing understanding of the relevant high-level cognitive functions in the brain.
Keywords: distancing, reappraisal, emotion regulation, functional magnetic resonance imaging, meta-analysis
Affective dysregulation is a transdiagnostic feature of mental health disorders including mood and anxiety disorders, trauma-related disorders, borderline personality disorder, addiction, and more (Kring & Sloan, 2010). These conditions have devastating personal and societal impacts (Insel, 2011), but effective training in emotion regulation can help alleviate affective disturbances and allow individuals to optimize their wellbeing (Berking, Ebert, Cuijpers, & Hofmann, 2013). Across the myriad methods available to regulate emotion, distancing is particularly versatile and effective (Webb, Miles, & Sheeran, 2012). These features make it a promising candidate for systematic investigation and clinical application.
Distancing refers to one of the two main reappraisal tactics for emotion regulation, the other being reinterpretation (Ochsner, Silvers, & Buhle, 2012; refer to “A Taxonomy of Distancing and Emotion Regulation” below for discussion on the distinction between tactics and strategies in emotion regulation). Distancing involves simulating a new perspective to alter the psychological distance and emotional impact of a stimulus. This distance can take several forms such as spatial distance, temporal distance, or objectivity. For instance, upon feeling hurt by criticism from a co-worker, you might try to minimize the impact of the incident by imagining how a neutral, objective observer would perceive the situation. In contrast, reinterpretation refers to deriving an alternative outcome or meaning for some stimulus to alter its emotional effect. In this case, you might change your emotional response by imagining that your co-worker is not simply criticizing you, but is trying to help you. The difference between these tactics is that reinterpretation is more focused on transforming the content or meaning of the stimulus, while distancing is more focused on transforming the viewpoint from which the stimulus is considered. Another way of thinking about this difference is that reappraisals involving reinterpretation are stimulus-dependent, whereas those involving distancing are not. In the example above, a neutral viewpoint could be considered for any situation, but reimagining the co-worker’s intentions is dependent on the circumstances of this situation.
Reappraisal, more broadly, tends to produce some of the largest effect sizes in the emotion regulation literature (Webb et al., 2012), and its versatility makes it an appealing alternative to other strategies in many situations. For instance, strategies characterized by avoiding or changing the external aspects of a situation are not always possible. Likewise, shifting attention away from aversive stimuli can be a powerful approach to regulating emotion (Webb et al., 2012), but it might not be feasible for aversive situations that require focused attention (e.g. treating an injury). Strategies focused on modifying the outcomes of emotional responses (e.g. relaxation techniques) can be effective in some contexts, but they can also interfere with ongoing goal-directed behavior. Reappraisal tactics are advantageous because they can be implemented in a wide range of situations with less attentional and behavioral disruption. These tactics still occupy cognitive resources, but, unlike distraction, the relevant event is still being attended and processed. Thus, reappraisal tactics are favorable over other emotion regulation strategies in some contexts.
Although the reappraisal tactics of distancing and reinterpretation are both effective, some differences between them suggest that distancing may be particularly promising. Denny and Ochsner (2014) found that longitudinal training in distancing was associated with decreased stress in daily life and more neutral evaluations of aversive content when no regulation was instructed. These benefits were not associated with longitudinal training in reinterpretation. Additionally, applying distancing across diverse situations may require less effort than reinterpretation because distancing solutions are not bound by the specific features of a stimulus. Although reinterpretation has received greater attention in emotion regulation research (Koenigsberg et al., 2010; Ochsner et al., 2012), the benefits of distancing motivate further investigation of this tactic.
Obstacles in Distancing Research
Below we identify some of the major obstacles currently hindering the research and application of distancing. In this article, we address each of these challenges and offer guidance for future research.
In the distancing literature, the specific emotion regulation technique used in a study is sometimes referred to as broadly as reappraisal (Goldin, McRae, Ramel, & Gross, 2008; Gruber, Hay, & Gross, 2014; Opialla et al., 2015). This practice can foster the misleading assumption that the results of these studies are directly comparable to any other reappraisal findings. As noted above, though, reappraisal encompasses a variety of specific techniques, and differences have been found between variants of reappraisal (Denny & Ochsner, 2014; Webb et al., 2012). To complicate matters further, consistent techniques have been labeled with numerous different terms across studies. For example, researchers using distancing focused on objective perspective taking have referred to these techniques as reappraisal (Goldin et al., 2008; Gruber et al., 2014), inhibition (Beauregard, Lévesque, & Bourgouin, 2001), cognitive regulation (Erk, von Kalckreuth, & Walter, 2010; Schardt et al., 2010), self-focused regulation (Ochsner, Ray, et al., 2004), detachment (Erk, Mikschl, et al., 2010), detached reappraisal (Shiota & Levenson, 2009) and suppression (Lévesque et al., 2003). These weaknesses in terminology have likely contributed to inconsistent methods including participant instructions, training criteria, and measures of regulation success.
Because distinctions within reappraisal are not consistently recognized, the mechanisms that support distancing are not clear. Understanding these cognitive mechanisms and their brain bases could lead to improved measures of distancing (e.g. functional neuroimaging indices of neural circuit activation) which, in turn, could inform better interventions (e.g. targeted neurostimulation). In addition to allowing for better applications of distancing in clinical contexts, these efforts could also better integrate distancing research with related domains of inquiry, such as perspective taking. For example, distancing extends traditional social and visual perspective-taking research into self-generated, imagined perspectives that challenge a simple conceptualization of the assignment of self and other (for example, see D’Argembeau & Van der Linden, 2004). Investigating the distinctive applications of perspective taking in distancing may reveal more nuance into how the brain accomplishes these complex cognitve processes.
Goals of this Article
To address these obstacles, we pursue three aims in this article. First, we offer a taxonomy in which we define distinct subtypes of distancing using construal-level theory of psychological distance (Trope & Liberman, 2010), and place them within the broader framework of the process model of emotion regulation (Gross, 1998b). By integrating existing terminology from areas related to distancing, this taxonomy provides a foundation for better communication in emotion regulation research. Second, we leverage the definition of distancing developed in our taxonomy to review literature regarding the effectiveness of distancing, including how it compares to other methods of regulation. In doing so, we also identify areas for future work that could promote more effective applications of distancing. Third, we perform a quantitative meta-analysis of functional neuroimaging findings and integrate these results with additional literature to propose a preliminary neurocognitive model of distancing. This model presents a mechanistic outline of distancing, which can inform hypothesis-driven research on these mechanisms and targets for clinical interventions.
Defining Distancing Within Emotion Regulation
In this section, we use a cognitive theory of psychological distance to define distancing in detail. We then discuss distancing within the broader context of emotion regulation and propose a system of terminology for unifying experimental approaches. We conclude with brief remarks about the theoretical implications of this taxonomy and its flexibility to evolve with emotion regulation research.
Defining Distancing Using Construal-Level Theory
Construal-level theory (CLT) is the dominant theory on psychological distance. It posits four dimensions of psychological distance: spatial, temporal, social, and hypothetical (Trope & Liberman, 2010). It further states that distance is measured egocentrically. In other words, the reference point is the self in the here and now, and distance increases as representations become more removed from the self, the present, etc. While CLT identifies four types of psychological distance, it also proposes that they share a common underlying dimension of abstraction that ranges from close and concrete to far and abstract. In the terminology of CLT, this underlying dimension reflects construal level. With increasing distance, regardless of the type, the mental construal of an object becomes more abstract.
For instance, if a person imagines a dog in the same room as herself, then her mental representation of the dog will likely be grounded in concrete details, such as the color and texture of the fur. On the other hand, if she is instructed to imagine a dog on the far side of a large field, her representation of the dog will likely be more abstract, consisting of more general dog features, such as having four legs, having a tail, or the general shape of a dog. D’Argembeau and Van der Linden (2004) demonstrated this relationship between psychological distance and abstraction by having participants recall past events or imagine future events. As the temporal distance of the events increased in either direction, participants reported less sensorial and contextual detail as well as weaker feelings of re-experiencing or pre-experiencing.
Because the various dimensions of psychological distance are all linked through construal level, CLT also suggests a predictive relationship among these dimensions. For example, when the person imagines the spatially distant dog versus the nearby dog, she is more likely to report that the dog feels less real (more hypothetical), less familiar (more socially distant), and less present in time (more temporally distant). These features of CLT have been further supported by studies of how psychological distance is expressed in language (Stephan, Liberman, & Trope, 2010), how it affects behavior (Bar-Anan, Liberman, Trope, & Algom, 2007), and how it is represented in the brain (Parkinson, Liu, & Wheatley, 2014).
CLT was developed independently from the emotion regulation tactic of distancing, but it corresponds well with the ways distancing has been implemented in emotion regulation studies. Four forms of distancing have emerged in the emotion regulation literature, which we refer to as spatial, temporal, objective, and hypothetical distancing. Each form of distancing requires the user to generate a new, imagined perspective of emotional content that increases or decreases separation between the person and content in some way. Most commonly, the imagined perspective involves increased distance of some form to attenuate an emotional response (i.e. downregulation). For example, if someone were upset after losing a credit card, he could imagine his situation from the perspective of a removed, objective observer. This perspective-taking process may then enable more affectively neutral reflection on the event, such as, “The situation is inconvenient, but the problem will not likely take long to resolve or result in any major harm. Getting upset would just be a waste of energy.” While taking a more or less distant perspective of the stimulus is the defining component of distancing, the new appraisals of the stimulus generated under this perspective help to shape a new affective response. Here, the use of objective distancing interrupts and replaces the current response with a more neutral response based on the alternative perspective. Thus, new affective associations are formed with the stimulus. These new associations then allow for the more moderate emotional response to persist beyond the period of active engagement with the alternative perspective (Kross & Ayduk, 2008). Descriptions and examples of general techniques for each form of distancing in the context of affective downregulation are included in Table 1. Note that in some contexts distancing can also be applied for upregulating affective responses. In addition to using these forms separately, regulation techniques can also combine forms of distancing or blend distancing with other methods of regulation (e.g. performing deep breathing while distancing).
Table 1.
Forms of Distancing as Applied to Emotion Downregulation
| Form | Description | Example Technique |
|---|---|---|
| Spatial | The user downregulates an emotional response by taking a perspective that is more spatially distant from the stimulus. | Imagine how you would feel about the content if you were very far away from it. |
| Temporal | The user downregulates an emotional response by taking a perspective in which the stimulus is more distant in time. | Imagine how you would feelabout the content after manyyears have passed. |
| Objective | The user downregulates an emotional response by taking an objective perspective. This perspective is often linked to an imagined observer, who may be a generic neutral party or a contextually appropriate professional, such as a clinician, investigator, or reporter. | Imagine you are observing the content from the perspective of a neutral, objective observer. |
| Hypothetical | The user downregulates an emotional response by taking a perspective in which the stimulus represents a hypothetical scenario as opposed to reality. | Imagine how you would feel about the content if it were not real, but instead, a contrived or hypothetical scenario. |
Conceptually, the CLT dimensions of spatial, temporal, and hypothetical distance clearly map onto the spatial, temporal, and hypothetical forms of distancing, respectively. This begs the question of whether the remaining CLT dimension of social distance is similarly matched to objective distancing, the remaining form of the emotion regulation tactic. Objective distancing techniques often include an explicit social component by instructing the user to take the perspective of an observer (Schardt et al., 2010; Walter et al., 2009; Winecoff, LaBar, Madden, Cabeza, & Huettel, 2011). Curiously, while all distancing techniques involve simulating a new perspective, the user is only instructed to assign the new perspective to another agent in this way with certain objective distancing techniques. This shift in perspective from self to other entails an increase in social distance, but taking an observer’s perspective is only one of the features of these techniques. The other main feature relates to the quality of that observer, namely the observer’s neutrality, or objectivity.
What about techniques in which an individual is instructed to think about a stimulus in a neutral, objective way, but without mention of an observer perspective? We assert that neutrality is inherently related to social distance, and, therefore, objective distancing is associated with social distance even for techniques that do not explicitly involve another agent. When one perceives a personally relevant, emotionally salient stimulus, the self naturally experiences an emotional response. This event is also relevant to a socially close other, such as a family member or close friend, because her social connection with the original person has relevance to her. Thus, a socially close agent may experience an empathic emotional response (Zaki, 2014). On the other hand, the original person and event would have less personal relevance for a socially distant other, who would likely have a more objective response than the close other. A practical example of this association can be seen in the principles of juror selection. In its protections against selecting impartial jurors, the American Bar Association specifically mandates the exclusion of any person with a familial relation to a participant in the trial (American Bar Association, American Jury Project, 2005). In other words, we assume that low social distance may compromise objectivity.
Manipulations designed to decrease objectivity in distancing studies also support this relationship. In these studies, decreased objectivity and greater emotional engagement is accomplished by instructing participants to imagine themselves or a loved one involved with the eliciting stimulus, often a picture or film clip (Domes et al., 2010; Eippert et al., 2007). These instructions amount to decreasing social distance. Therefore, we can conclude that objectivity is closely related to social distance, although it may be mediated by other factors as well. Further research on distancing could help to better characterize this relationship and inform theories of social distance like CLT.
Finally, hypothetical distancing is distinct from other forms of distancing in a way that warrants special consideration. In studies using hypothetical distancing, the stimuli are generally pictures or film clips, and participants are instructed to focus on how the stimuli do not constitute the events they depict (e.g. “It is only a picture.” or “It is not real.”; Morris, Sparks, Mitchell, Weickert, & Green, 2012; Mulej Bratec et al., 2015; Pitskel, Bolling, Kaiser, Crowley, & Pelphrey, 2011). These laboratory tasks obscure the challenge of applying hypothetical distancing to most real-world stimuli because these experimental stimuli are only pictures and film clips, and many of them are contrived (e.g. portraying actors). In CLT, hypotheticality is framed as the probability of a future event or the imagined versus real nature of some content; the more hypothetically distant a situation is, the less likely it is to occur or to be real. Emotion regulation, however, is most often prompted by real circumstances.
All forms of distancing involve a change in perspective, but hypothetical distancing does not allow the stimulus to be reconstrued in the way that spatial, temporal, and objective distancing do. For these three forms, the person considers how her response would be impacted if her perspective of the event were not A, but were instead B. For instance, how would she feel about an argument with her friend if it were not only 20 minutes in the past, but several months? Such a mental exercise may help her to perceive the event as a fleeting disagreement of little consequence in the larger scope of their friendship, and she might achieve a more moderate emotional response. With hypothetical distancing, though, the person stops at considering how her response would be impacted if she did not have perspective A of the event. An alternative perspective of the event does not need to be simulated because, in this case, the event did not happen. In the above example, the person would imagine that the argument with her friend was just a situation that could have happened but really did not. This simple denial of reality, without any resulting reconstrual, seems unlikely to be productive. No new associations are formed with the event’s content to promote a persistent change in emotional response after active regulation has ended. Again, this problem is not present when using hypothetical distancing with pictures and film clips, as is often the case in experimental studies, because these stimuli are already representations of scenarios instead of the scenarios themselves. Thus, distancing can be applied by imagining that the stimulus is depicting a hypothetical scenario without having to imagine that the stimulus itself does not exist. Recognition of the hypothetical form of distancing supports a clear scheme for categorizing techniques in the distancing literature and emphasizes consistency with CLT, but, for the above reasons, this form of distancing may have less practical utility and therapeutic value than the others.
A Taxonomy of Distancing and Emotion Regulation
Precise and consistent use of concepts and terminology is crucial to advancing research on distancing and emotion regulation. At present, substantial variation in the use of terms makes it difficult to synthesize existing findings in the literature and integrate new data. We propose a system that integrates extant conventions in the literature of emotion regulation and psychological distance (Figure 1).
Figure 1.
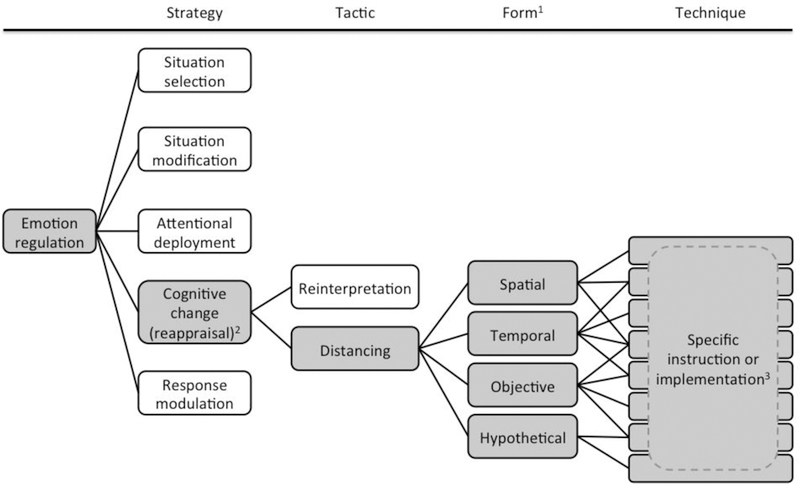
Taxonomy of distancing as an emotion regulation tactic. Gray boxes denote items at each level that relate to distancing. 1We refer to categories of distancing as forms; however, including a level between tactic and technique is not necessarily informative for all methods of emotion regulation. 2While distancing is regarded as a reappraisal tactic, there is discrepancy in the literature over whether reappraisal is synonymous with cognitive change or a subcategory of it. 3A distancing technique refers to any specific way that one or more forms of distancing is implemented (e.g. spatially distancing by imagining a stimulus receding away or imagining the stimulus being in another part of the world; spatially and temporally distancing by imagining that a stressful situation happened long ago and far away).
The first of these conventions is the process model of emotion regulation (Gross, 1998b). This widely referenced framework considers emotional responses in stages at which they can be modified. The process model states that regulation can occur at any of five principal stages during the course of an emotional response through situation selection, situation modification, attentional deployment, cognitive change, or response modulation. Situation selection refers to avoiding or seeking out situations based on the emotional responses they are likely to elicit, while situation modification refers to altering the circumstances of a situation to promote a different emotional outcome. Attentional deployment encompasses methods of directing attention toward or away from content to alter an emotional response; cognitive change involves thinking about a situation in a different way to modify its emotional impact; and response modulation describes efforts to regulate the behavioral or physiological outcomes of an emotional response. This model provides an overarching structure for categorizing more specific types of regulation.
The second convention involves the relationship between distancing and reappraisal. The term reappraisal is often used synonymously with the process model concept of cognitive change, although some researchers view reappraisal more narrowly as a subcategory of cognitive change (Gross, 2015). Despite disagreement over the scope of reappraisal, several researchers endorse a scheme of categorization in which the general approach of reappraisal refers to a strategy while the more specific approaches within it constitute tactics for implementing that strategy (McRae, Ciesielski, & Gross, 2012; Ochsner et al., 2012). Ochsner et al. (2012) further name distancing and reinterpretation as the two main tactics studied under reappraisal.
Under tactic, we propose the term technique be used to refer to the specific way that a tactic is instructed or implemented. As such, technique refers to the most detailed level of distinction among methods of emotion regulation. The purpose of this level is not so much to further categorize emotion regulation, but simply to provide a terminology for acknowledging differences in instruction and implementation. For example, one study might instruct participants to use distancing by thinking about stimuli in an objective, neutral way while another might instruct participants to consider stimuli from the factual perspective of a reporter. These studies would both be employing objective distancing, but with somewhat different approaches. The former defines the goal of being neutral and objective without a specific method for achieving it, while the latter specifically directs users to achieve this goal by activating their concept of a reporter and considering how that person would relate to material as part of their profession. In research aimed at defining or manipulating the neurocognitive mechanisms of distancing, it is important to acknowledge that these two instructions may recruit partially distinct mechanisms. Labeling these approaches as different objective distancing techniques provides a common language for addressing finer distinctions between different implementations of a tactic when these mechanistic differences may be relevant. This tripartite structure–strategy, tactic, and technique–constitutes the primary framework of our proposed taxonomy for emotion regulation. These levels can be applied to other categories in the process model in addition to cognitive change. For example, attentional deployment would refer to a general strategy, while distraction would represent a tactic within this strategy. One specific technique for implementing distraction could be intentionally shifting focus to an unrelated positive memory while another might be rehearsing a lengthy series of digits.
Further distinctions can be incorporated as needed within this structure. Distinctions within distancing have rarely been recognized in the literature, but distancing techniques can be further classified by the type(s) of psychological distance involved. Drawing on the conventions of CLT, distancing techniques can be classified as spatial, temporal, objective, hypothetical, or some combination of these types. In this review, we use form to refer to these types of distance (Figure 1). The number of useful levels of distinction might vary across methods of emotion regulation, though, so this additional level between tactic and technique might not be applicable to all regulation tactics. For distancing, however, this level of distinction could have implications for the mechanisms of these techniques, matching the optimal technique to an emotional elicitor, and developing brain-targeted methods to enhance distancing performance.
Conclusions and Future Directions
This system of terminology should be expanded and revised as research in distancing and emotion regulation progresses, but presently it captures meaningful distinctions within distancing and related concepts. Research under this framework may help to evaluate and develop its foundational models and theories, such as CLT. Specifically, comparing the mechanisms of different forms of distancing may inform the validity of these categories in CLT. For example, we might find evidence of a mechanistic distinction between hypothetical distancing and the other forms that would call into question the inclusion of hypotheticality as a dimension of psychological distance. Similarly, comparing the mechanisms of distancing and reinterpretation may help to refine conceptual distinctions within reappraisal. More broadly, the proposed taxonomy establishes a conceptual structure to promote hypothesis-driven research and effective interaction among emotion regulation researchers. Thus, this system facilitates a transition from individual studies of distancing to more cumulative progress as a research community.
The Effectiveness of Distancing
Below, we review the distancing literature as it concerns the effectiveness of this emotion regulation tactic. Specifically, we highlight the effects of distancing measured through self-report, neuroimaging, and psychophysiology; we review work comparing distancing with other methods of emotion regulation; and we discuss some future directions for this work. By far, the most commonly studied application of distancing is to decrease unpleasant or unwanted emotional responses by increasing the psychological distance between the research subject and the eliciting stimulus. This prevalence is driven by the fact that decreasing aversive emotions is the most common regulatory goal. As such, we focus on distancing techniques for downregulating emotion in this review. It is worth noting, however, that techniques for decreasing psychological distance to intensify emotions have also been studied (Ayduk & Kross, 2008; Davis, Gross, & Ochsner, 2011; Eippert et al., 2007; Schulze et al., 2011).
No gold standard exists for measuring the success of an emotion regulation manipulation, so studies have employed a wide variety of measures. We summarize the effects observed with these measures in healthy individuals as the effects of distancing in psychopathological contexts are beyond the scope of this review. Nonetheless, studies have demonstrated positive effects of distancing across a wide range of clinical populations (Denny, Fan, et al., 2015; Gaebler et al., 2014; Hermann et al., 2009; Lang et al., 2012), although in some cases, effects were attenuated relative to control groups (Erk, Mikschl, et al., 2010; Morris et al., 2012).
Self-Report Measures
Some studies have used self-reported valence to measure distancing success. In these studies, participants rated the degree of positivity or negativity of their experienced affect in response to an emotion-eliciting stimulus on a numbered (Koenigsberg et al., 2009) or pictorial scale (Vrtička, Bondolfi, Sander, & Vuilleumier, 2012; Winecoff et al., 2011; Winecoff et al., 2013). Valence ratings were then compared between distancing trials and natural response trials, in which participants were instructed to permit their natural emotional responses while attending to aversive stimuli. Natural response is a common control condition in studies of emotion regulation, but, importantly, the influence of implicit regulation on natural responses is unknown. Regardless, these studies indicated significant positive shifts in valence with distancing relative to natural response in healthy adults. Other studies have reported similar effects for spatial and objective distancing using arousal ratings (Davis et al., 2011; Grecucci et al., 2015).
In other work, self-report measures have been created that blend valence and arousal into a general construct of affective intensity (Ochsner, Ray, et al., 2004; Wang, Lin, Huang, & Yeh, 2012). Participants rated the strength of their negative or positive affect, or they rated individual emotions that were later collapsed into negative and positive affective intensity scores. More complex composite measures of emotional reactivity have been used as well. For instance, Ayduk and Kross (2008) instructed participants to recall anger-evoking autobiographical experiences and rate the extent to which they re-experienced their original feelings and the intensity of these feelings. These ratings were then averaged into a metric of emotional reactivity. For each of these measures, distancing resulted in decreased emotional responses.
Self-reported regulation success is another measure in common use. In these studies, participants were asked to provide numeric ratings of their self-appraised success at reducing emotion. Schardt et al. (2010) used a success rating scale of one (not at all successful) to nine (very successful) and found a mean participant rating of approximately six, while Leiberg, Eippert, Veit, and Anders (2012) used a similar rating scale of one to five and found a mean rating of approximately 3.7. Lévesque et al. (2003) asked participants to estimate their objective distancing performance, which was reported as 84-percent successful on average and was corroborated by decreased online ratings of sadness during distancing relative to natural response. In summary, the above studies in healthy adults indicate that distancing has been reliably associated with decreases in subjective emotional experience and successful strategy implementation.
Researchers have also compared distancing with other methods of regulation using self-report measures. Gross (1998a) found that objective distancing reduced disgust ratings to film clips relative to natural response, whereas expressive suppression, or intentionally inhibiting outward expressions of emotion, did not. Ochsner, Ray, et al. (2004) found that groups instructed to use either reinterpretation or objective distancing were comparably effective at decreasing negative affect using a rating of affective intensity. Denny and Ochsner (2014) compared the effects of distancing and reinterpretation over a period of approximately two weeks in a longitudinal training study. In this study, distancing could include spatial, temporal, or objective distancing at the participants’ discretion. Relative to natural response to negative pictures, both reappraisal tactics were associated with decreases in ratings of negative affect that were strengthened over the course of the study; however, only the group using distancing showed a longitudinal decrease in negative affect when responding naturally on negative trials and decreased perceived stress in daily life. These results suggest that distancing may have greater potential to generalize beyond its instructed use. They also suggest that within-subject comparisons of regulation and natural response may underestimate the effect size of distancing in well-practiced participants. Kross and Ayduk (2008) compared a technique combining spatial and objective distancing with a technique of passive neutral distraction while participants recalled a depressive experience. The distraction technique involved presenting neutral content to draw participants’ attention, but they were not given an explicit emotion regulation goal. These techniques comparably reduced depressive affect ratings immediately after regulation, but only distancing had sustained regulatory effects when the same experience was recalled again on a later day. These studies suggest that distancing is at least as effective as some other common methods of emotion regulation, if not more so, and distancing may specifically promote auxiliary regulatory effects.
Webb et al. (2012) performed a meta-analysis on the effectiveness of various regulation methods in nonclinical samples. This study distinguished several tactics within the strategies of attentional deployment, cognitive change, and response modulation outlined in the process model of emotion regulation. Out of seven attentional categories, four cognitive categories, and four response modulation categories, distancing had the third largest pooled mean effect size (d+ = 0.45) after active positive distraction (d+ = 0.47) and non-specific reappraisal (d+ = 0.89). Active positive distraction is a tactic in which participants are explicitly instructed to think about positive content unrelated to the stimulus to distract themselves, while in non-specific reappraisal, participants freely choose among reappraisal methods. In the non-specific reappraisal studies, distancing techniques were sometimes indicated through instructions or post-task feedback (Harenski, Kim, & Hamann, 2009; Kim & Hamann, 2007). In other words, distancing also contributed to the regulation category with the largest effect size. Overall, these findings demonstrate the relative effectiveness of distancing within the broader landscape of emotion regulation methods.
Neuroimaging Markers of Distancing Success
Owing to its central role in emotional processing, amygdala activation has been the most common neuroimaging measure of emotion regulation success, as well as distancing success more specifically. Amygdala activity is often presumed to correspond to affective intensity (Phan, Wager, Taylor, & Liberzon, 2002; Satpute, Shu, Weber, Roy, & Ochsner, 2013), such that reduced amygdala activation is interpreted as a marker of emotion downregulation. The processes underlying distancing may have downstream regulatory effects on this region, allowing its activation to serve as an outcome measure. Thus, we focus on effects in the amygdala below, but for a discussion of the processes directly involved in distancing and their neural mechanisms, see “A Preliminary Neurocognitive Model of Distancing” later in this article.
Eippert et al. (2007) associated mixed objective and hypothetical distancing with reduced activity in the left amygdala, which was replicated by Domes et al. (2010) in the bilateral amygdala using a similar distancing technique. Walter et al. (2009) examined objective distancing using a two-part neuroimaging task to evaluate sustained regulatory effects. In the first part, participants viewed aversive and neutral images and were instructed to distance for a subset of the stimuli. In the second part, the images were presented again without explicit regulation instructions. Distancing was associated with decreased bilateral amygdala activity in response to aversive images in both the first and second parts of the task, indicating a sustained decrease in this marker for post-regulation stimulus presentation. A subsequent study showed sustained effects on right amygdala activity one week after regulation using a mixed distancing technique, but only for stimuli that had been reappraised repeatedly during the initial session (Denny, Inhoff, Zerubavel, Davachi, & Ochsner, 2015). These latter findings are suggestive of a dose-dependent response of distancing on amygdala activity.
Another group of studies have implicated the serotonergic system in the relationship between distancing (specifically objective distancing) and amygdala activity. Outhred et al. (2015) examined left amygdala activity and found that decreases related to distancing were enhanced by acute administration of escitalopram, a selective serotonin reuptake inhibitor used to treat anxiety or depression. Another study examined the relationship between distancing and a common polymorphism in the promoter region of the serotonin transporter gene SLC6A4, the short allele of which has been associated with hyperreactivity in the amygdala to negative emotional stimuli (Hariri et al., 2005; Schardt et al., 2010). Right amygdala activity was higher for short-allele carriers when responding naturally to fear-related images, although this relationship did not hold for disgust-related images. When using distancing, though, amygdala activity decreased to comparable levels between groups. These results suggest that distancing may be capable of compensating for some biological susceptibilities to high emotional reactivity. This polymorphism has further been shown to moderate the relationship between escitalopram administration and amygdala effects during distancing (Outhred et al., 2016). Given the systemic nature of drug administration in these studies, mechanistic interpretations of these findings should be made with caution. Nevertheless, these results collectively suggest that the serotonergic system may have a moderating effect on distancing.
Regarding distancing in comparison with other methods of regulation, an analysis collapsing across objective distancing and reinterpretation yielded a positive correlation between negative affect scores and bilateral amygdala activity (Ochsner, Ray, et al., 2004); however, a subsequent study that directly compared these tactics found that objective distancing resulted in greater attenuation of left amygdala activity than reinterpretation (Dörfel et al., 2014). Additionally, Vrtička, Sander, and Vuilleumier (2011) compared hypothetical distancing to expressive suppression and found greater modulation of left amygdala activity with distancing and right amygdala activity with suppression. The reasons for these particular patterns, however, are unclear.
These and other results suggest that amygdala activity may serve as a neural marker for emotion regulation success in certain contexts, but further investigation is needed to explain inconsistencies in laterality and differences related to stimulus type (e.g. digust-versus fear-related images in Schardt et al., 2010). zThe amygdala is also a particularly difficult region to image with fMRI due to its small signal intensity and proneness to susceptibility artifacts because of its anatomic location (LaBar, Gitelman, Mesulam, & Parrish, 2001). Such limitations should always be considered when interpreting findings for this region, and they may have contributed to some of the inconsistencies noted here, particularly with regard to laterality effects and the potential for null findings. Finally, emotional processing is known to be distributed across numerous brain areas (e.g., Kragel & LaBar, 2016), which calls for an expansion of neural metrics of emotion downregulation beyond the amygdala.
Psychophysiological Effects
Data on the psychophysiological effects of distancing are limited. Early work by Gross (1998a) compared objective distancing and expressive suppression in response to disgusting stimuli while measuring finger pulse amplitude, skin conductance, and finger temperature as measures of sympathetic activity as well as general body movement and heart rate. As discussed above (see “Self-Report Measures”), self-reported affect was reduced for distancing relative to both suppression and natural response. Nonetheless, all physiological responses were comparable between the distancing and natural response groups, while sympathetic measures were elevated in the suppression group. Similarly, Eippert et al. (2007) found no effect of distancing on skin conductance or startle eyeblink amplitude, another measure that has been associated with autonomic arousal (Dillon & LaBar, 2005), in response to aversive images using a combined objective and hypothetical technique.
However, more recent work has yielded positive findings. Ayduk and Kross (2008) reported a decrease in blood pressure in participants recalling anger-evoking experiences using mixed spatial and objective distancing relative to first-person perspective recall. In a subsequent study, Ayduk and Kross (2010) evaluated spontaneous regulation in the form of taking an observer perspective and found that spontaneous distancing negatively correlated with total peripheral resistance, an autonomic measure of vasoconstriction. Paret et al. (2011) associated mixed spatial and objective distancing with decreased skin conductance during threat of shock, while Leiberg et al. (2012) evaluated the effect of mixed objective and hypothetical distancing on responses to aversive scenes using startle eyeblink amplitude and skin conductance (following methods from Eippert et al. [2007]). Startle and skin conductance responses were lowest when participants were instructed to increase psychological distance, although distancing was not directly compared to natural response. Instead, these effects were demonstrated through linear contrasts of increased distance, natural response, and decreased distance.
It is too early to draw conclusions about the psychophysiological effects of distancing from this handful of studies. Differences among distancing techniques and dependent measures may have contributed to the apparent discrepancy in the findings to date. Future studies may require more comprehensive psychophysiological assessment to identify which measures or combinations of measures show the most sensitivity to distancing, paying careful attention to their relationship to sympathetic and parasympathetic activation. Finally, the field may benefit from applying sensitive multivariate analytic tools that can better account for covariance among multiple physiological measures (Kragel & LaBar, 2014).
Future Directions
From a translational perspective, a primary goal in emotion regulation research should be to maximize the effectiveness of regulation. The above literature review attests to the effectiveness and versatility of distancing within emotion regulation studies, making this tactic an ideal target for further optimization. Some research directions in this pursuit include directly comparing distancing techniques, exploring the influence of individual and contextual factors on distancing, and investigating compound regulation techniques, which are discussed below. In addition, more research should incorporate long-term outcomes.
While determining how to maximize regulatory effects, some practical concerns bear consideration: namely, determining whether it is adaptive or maladaptive to regulate a given emotional response, and with which techniques. Wang et al. (2012) proposed that individual differences in attachment style, reflecting differences in default psychological distance, might be important in determining the effectiveness of distancing techniques for individuals. Their results indicated that individuals with a general tendency for more self-immersive processing may benefit from techniques based on increasing psychological distance whereas those with a tendency for processing from a distance may benefit from techniques for decreasing psychological distance. Additionally, successful emotion regulation can have negative consequences, such as poor social outcomes (Ben-Naim, Hirschberger, Ein-Dor, & Mikulincer, 2013; Butler et al., 2003), under certain circumstances. Thus, increasing psychological distance to blunt an emotional response is not necessarily optimal for a given scenario, and further research into these questions is warranted. Nevertheless, effective tools are required for when regulation is needed, and distancing may be well suited in many cases.
Comparing distancing techniques.
Different distancing techniques are likely supported by partially distinct neurocognitive mechanisms, and therefore, these techniques might differ in their effectiveness. We are not aware of any studies that have directly compared the effects of different distancing techniques, although some work has begun to investigate the mechanisms of specific forms of distancing (Ahmed, Somerville, & Sebastian, 2017; Bruehlman-Senecal & Ayduk, 2015). Direct comparisons could aid clinicians in selecting the most beneficial techniques to include in emotion regulation skills training. Furthermore, the effects of different techniques may differ by population. These questions are clearly relevant for optimizing distancing applications in treatment contexts and merit further exploration.
Individual factors.
Individual differences in cognition likely contribute to individual differences in regulatory success. Therefore, identifying the cognitive resources that support distancing and developing ways to assess individual differences in these components could yield predictors of distancing success. Some of these factors may relate to distancing more generally, while others may be more specific to particular forms or techniques. These predictors could then be used to target individuals for whom distancing techniques are most likely to be effective, an especially important goal in the age of personalized medicine.
Little work has been done to characterize individual differences in distancing, but studies relating distancing performance to development and cognitive capacity provide a general demonstration of this approach. Studies of children and adolescents have found that the effectiveness of these techniques increases over early-life development (Silvers, Insel, et al., 2016; Silvers, Shu, Hubbard, Weber, & Ochsner, 2015; White, Kross, & Duckworth, 2015). Conversely, research in healthy adults has demonstrated reduced distancing success in association with older age (Shiota & Levenson, 2009; Winecoff et al., 2011). General cognitive capacity may be an underlying factor contributing to this effect. Schmeichel, Volokhov, and Demaree (2008) found that participants with higher working memory capacity used both objective distancing and expressive suppression more effectively than participants with lower working memory capacity in response to emotional film clips. These results indicate that working memory capacity may predict regulation success more generally for techniques involving executive functioning. Further supporting this conclusion, Winecoff et al. (2011) found a relationship between higher cognitive capacity and decreased amygdala activity across young and older adults during objective distancing. These findings implicate cognitive capacity as a potential mediator of age-related differences in distancing success. Differences in visuospatial skills, mentalizing, empathy, fantasizing, and other processes related to perspective taking may be more specific predictors of distancing success among other cognitive methods of emotion regulation, but these relationships have yet to be tested.
Contextual factors.
Distancing is presumably better suited for some situations than others. Distancing effects may be influenced by contextual factors, such as emotional category (e.g. fear, sadness, disgust); the presence of competing, non-regulatory goals; the regulatory target (personal memories, movies, photos); the time frame of the stimulus (remembered, present, or anticipated); and the intensity of emotional response. Research determining the contexts for which distancing is best suited could be used to help individuals select distancing for circumstances in which it is most likely to be effective.
While few studies have focused on contextual factors related to distancing, emotional intensity has received some attention. Some evidence suggests that distancing might not be best suited for regulating high-intensity emotional responses, although it is likely to be effective for responses of low-to-moderate intensity (Sheppes, Scheibe, Suri, & Gross, 2011; Wisco et al., 2015). Certain forms of distraction may be more effective at reducing high-intensity emotions (Sheppes et al., 2011; Smoski, LaBar, & Steffens, 2014), but other contextual factors may also impact technique selection. Diverting attention from upsetting stimuli and sacrificing accurate encoding may be maladaptive in some real-world situations, such as responding to an emergency. Objective distancing, on the other hand, has been shown to have no impact (Erk, von Kalckreuth, & Walter, 2010; Richards & Gross, 2000) or even positive effects on memory performance relative to natural response (Dillon, Ritchey, Johnson, & LaBar, 2007). Thus, for situations in which effective encoding is still needed despite high emotional intensity, distancing may be preferable to distraction. Further exploration of these and other contextual factors will help promote more optimal use of distancing and other emotion regulation methods.
Compound regulation techniques.
In this article, we assert that dividing distancing into distinct forms may be informative for determining mechanisms; for maximizing regulation, however, combining emotion regulation techniques or giving individuals a range of options may have a powerful impact. In a meta-analysis of the effectiveness of emotion regulation tactics, Webb et al. (2012) found that the pooled mean effect size associated with non-specific reappraisal (d+ = 0.89) was much higher than those related specifically to distancing (d+ = 0.45) and reinterpretation (d+ = 0.36). In non-specific reappraisal conditions, participants had a choice in their selection of reappraisal methods. The studies in this category tended to use vague reappraisal instructions (Ochsner, Bunge, Gross, & Gabrieli, 2002) or include several forms of reappraisal (Lu & Stanton, 2010). Similarly, many of the studies we discuss in this article included or combined multiple forms of distancing in their instructed techniques (e.g. Denny & Ochsner, 2014; Kim & Hamann, 2007). We refer to these methods, in which individuals freely choose among multiple regulation techniques or employ multiple techniques simultaneously, as compound regulation techniques. More work is needed to clarify the striking difference in effect sizes observed by Webb and colleagues. Nevertheless, their results suggest that compound regulation techniques could be a valuable approach for optimizing distancing in interventions for emotion dysregulation.
Conclusions
Distancing has been shown to regulate emotion across a wide range of healthy and clinical populations. These effects have been most commonly and consistently demonstrated through measures of self-report with minimal training and short-term outcome evaluation. Analyses of amygdala activity have generally indicated decreases in emotional reactivity in association with distancing, but some questions remain concerning the scope of this measure and inconsistencies in the laterality of these findings. Several studies have also evaluated distancing using psychophysiology, but these early results do not yet provide clear support for any conclusions and still await replication. Only a small number of studies have directly compared the effects of distancing to other methods of emotion regulation, but initial findings generally support a favorable profile of distancing relative to other tactics within moderate levels of emotional intensity. Future research controlling for differences in distancing techniques will allow for better characterization of these comparisons. Directions for future research include comparing different distancing techniques, examining relevant individual and contextual factors, and pursuing compound regulation techniques, perhaps in combination with other means of engaging plasticity in relevant brain networks (e.g. neuromodulation).
A Preliminary Neurocognitive Model of Distancing
While the neurocognitive mechanisms of reappraisal have been discussed elsewhere (Ochsner & Gross, 2007; Ochsner et al., 2012), little attention has been given to the mechanisms that specifically support distancing. We propose that understanding these mechanisms may lead to greater insight into the cognitive processes involved in distancing and the development of brain-based techniques to enhance distancing performance. In this section, we aim to identify cognitive processes supporting this tactic and leverage relevant neuroscientific literature to establish an initial neurocognitive account of distancing. We begin with a discussion of putative component processes and their neural bases, emphasizing processing most specific to distancing. Unlike the above discussion on neuroimaging measures of distancing outcomes, this section considers neuroimaging evidence of its mechanisms. We then review comparisons between this tactic and other methods of emotion regulation. Based on this evidence, we propose a preliminary model of distancing and consider some future directions in relation to this model in both basic and clinical research.
Cognitive Processes and Neural Substrates
The core processes that contribute to distancing should be relatively consistent across instances despite some contextual variation in how it may be initiated. One approach to defining these processes is to consider how emotional responses involving distancing differ from natural responses. This approach is mirrored in experimental studies that contrast a distancing condition with natural response to control for lower-level stimulus processing. We will reference a common experimental paradigm for distancing as a framework to discuss its constituent processes. This laboratory paradigm involves emotional elicitors such as affective pictures, film clips, or memory cues and an early condition cue preceding each stimulus to instruct the method of response (refer to Ochsner et al., 2012 for discussion of early vs. late cues).
Using this paradigmatic case, we propose that the following cognitive processes would characterize a typical instance of distancing. In a given experimental trial, an individual is first presented with a cue that references instructions to distance or respond naturally to the upcoming stimulus, which initiates a corresponding goal state to either regulate the upcoming response or abstain from regulation. Next, an emotionally salient stimulus is presented, such as an evocative picture. The stimulus is perceived and interpreted, triggering an initial emotional response, which is recognized by the individual through a process of affective self-reflection. This self-reflective processing would likely occur in real-world contexts as well, although it is particularly emphasized in experimental tasks that include affective self-report. In the natural response condition, the individual cycles through these processes of stimulus perception, interpretation, and affective self-reflection. In the distancing condition, the individual constructs and adopts an alternative perspective of the stimulus through a process of self-projection. This change in perspective prompts a new cycle of stimulus perception, interpretation, and affective self-reflection based on the alternative perspective of the stimulus. Meanwhile, cognitive control processes are required to monitor pursuit of the regulatory goal and modify self-projection processes as needed. Notably, distancing diverges from other reappraisal tactics and other types of emotion regulation in the self-projection process. Below we discuss this cognitive process and its basis in the brain in greater detail as well as the affective self-reflection and cognitive control processes more briefly.
Self-projection.
Buckner and Carroll (2007) defined self-projection as the ability to shift perspective from the here and now to a simulated time, place, or person. The concept of self-projection effectively describes the mental transformations involved in distancing. Buckner and Carroll proposed that self-projection provided a unifying description of the various abilities that have been linked to a common neural network. These abilities include remembering the past, prospection (imagining future scenarios), theory of mind, and navigation. They described a neural network largely overlapping with the default mode network (DMN) that includes the medial prefrontal cortex (PFC), medial temporal lobe (MTL), medial parietal cortex, and lateral parietal cortex. A key component of their proposal is the idea that these various forms of self-projection all rely on memory. Past associations are referenced to construct simulated perspectives, regardless of their temporal orientation (i.e. simulating a past, present, or future scenario; Barrett & Satpute, 2013). Buckner, Andrews-Hanna, and Schacter (2008) went on to hypothesize that the primary function of the DMN is to facilitate an internal processing system focused on prospective simulation.
Hassabis and Maguire (2007) also noted the striking overlap in the brain areas associated with these abilities, and offered an expanded functional account that includes the imagination of fictitious scenarios that are not tied to the self (e.g., imagining a squirrel digging in the ground). Although the perception of such imagined scenarios would still be tied to an egocentric perspective (e.g., I am seeing the squirrel in my imagination), they suggested that these simulations do not rely on self-concept in a comparable way to true instances of self-projection. Therefore, they proposed that scene construction was the more basic and probable function of this neural network. Later, Spreng, Mar, and Kim (2009) followed up on this discussion with a series of meta-analyses testing the degree of overlap in the networks for autobiographical memory, prospection, theory of mind, navigation, and the default mode. They concluded that a common core network does underlie these processing states, although some states were characterized by more overlap than others. The core network they found included the medial and lateral PFC, MTL, posterior cingulate cortex and surrounding parietal regions (PCC+), temporoparietal junction (TPJ), and portions of the lateral temporal cortex (LTC) in the vicinity of the superior temporal sulcus and middle temporal gyrus.
These studies and others have also discussed the more specific roles that some of these network components might play in relation to self-projection. The medial PFC has been consistently associated with self-referential processing (Buckner et al., 2008; Northoff et al., 2006; Spreng et al., 2009) and may manage attributions to self versus other during self-projection (Buckner & Carroll, 2007; Kurczek et al., 2015; Satpute et al., 2013) or attributions of personal relevance (Ochsner & Gross, 2007). The MTL’s more general role in associative memory could provide the building blocks for constructing simulated scenes (Buckner et al., 2008; Kurczek et al., 2015). Due to its widespread connectivity, the PCC+ has been hypothesized to serve as a processing hub integrating medial PFC- and MTL-driven systems in the network (Buckner et al., 2008). As a result, it may integrate self-reflective processing and scene construction to support the egocentric experience of mental simulations (Cavanna & Trimble, 2006). The TPJ has been associated with a wide range of processes related to perspective taking including mentalizing (Frith & Frith, 2006; Saxe & Kanwisher, 2003), representing alternative visual perspectives (Aichhorn, Perner, Kronbichler, Staffen, & Ladurner, 2006; Schurz, Aichhorn, Martin, & Perner, 2013), egocentric spatial representation (Schindler & Bartels, 2013), and representing the position of the body in space (Blanke et al., 2005; Ionta, Perruchoud, Draganski, & Blanke, 2012). One hypothesis regarding the role of the TPJ in self-projection is that it manages the separation of true and simulated mental states (Mitchell, 2009). In other words, it functions to distinguish one’s personal mental state from a simulated alternative perspective. Finally, the LTC has been related to semantic processing (Ochsner et al., 2012; Spreng et al., 2009), which could explain why it has been consistently associated with the DMN, or self-projection network, but often as a less robust component (Buckner et al., 2008). It may be that LTC recruitment only becomes prominent when constructing perspectives more dissimilar (i.e. distant) to the self (Denny, Kober, Wager, & Ochsner, 2012), which may require greater semantic elaboration. Despite these hypothesized roles for specific regions, their interdependence in a functional network is strongly implicated in the literature.
The neural representation of psychological distance may be closely related to the self-projection network described above. CLT does not directly address the brain, but the theory implies that psychological distance of any type might have a common neural signature (Trope & Liberman, 2010). A few neuroscientific studies testing this idea have been reported, but results have been inconsistent to date. Parkinson et al. (2014) presented participants with stimuli that varied in spatial, temporal, or social distance while collecting fMRI data. Using a multivariate classification technique, they found the right TPJ to be involved in coding all three forms of distance. In contrast, Tamir and Mitchell (2011) reported that activity in the medial PFC was linked to changes in perspective involving spatial, temporal, social, and hypothetical distance. Greater MPFC activity was associated with more proximal perspectives across distance dimensions. Finally, another study demonstrated robust activity in association with spatial, temporal, and social distance in the PCC+ and TPJ, with considerable extension into the LTC for temporal distance and the medial PFC for social distance (Peer, Salomon, Goldberg, Blanke, & Arzy, 2015). Considering the abilities that have been associated with self-projection, such as mentalizing and prospection, it is not surprising that the neural substrates of psychological distance seem to reside in the same network. Thus, the neural underpinnings of self-projection and psychological distance predict a distribution of activity roughly consistent with the DMN when individuals engage in distancing to regulate their emotions.
Affective self-reflection.
Appraising one’s own affective state is a crucial process in cognitive emotion regulation tactics such as distancing. This information is critical for control processes assessing regulatory needs and the effectiveness of regulatory attempts. It is important to note that affective self-reflection is also likely to occur during natural emotional response, particularly in experimental settings that require affective self-report. Therefore, neural activity related to this process may not emerge from a standard contrast of distancing with natural response (Buhle et al., 2014). Nevertheless, given the key role of affective self-reflection in distancing, we briefly discuss the proposed neural substrates of this process.
Affective self-reflection has been most consistently linked with the dorsomedial PFC. Ochsner, Knierim, and colleagues (2004) compared conditions of judging one’s own emotional state, judging another’s emotional state, and non-emotional judgments about scenes. They identified regions in the medial PFC including the dorsomedial PFC to be specifically associated with emotional judgments of self, while activation in the PCC+, superior LTC, and medial PFC was related to emotional judgments in general. This study expanded on similar findings by Lane, Fink, Chau, and Dolan (1997) also linking the medial PFC with evaluating subjective emotional responses. Ochsner and Gross (2007) broadly implicated the dorsal PFC and adjacent cingulate regions in generating descriptions of one’s own emotional state. In a later review, Ochsner and colleagues (2012) more specifically highlighted the dorsomedial PFC and its putative function of attributing mental states, including attending to and interpreting one’s own emotional state. They also addressed the role of the ventromedial PFC, a region that has been implicated in various affective processes, but especially fear extinction (Milad & Quirk, 2012; Schiller & Delgado, 2010). This region seems to be involved in representing the subjective value of a stimulus in a context- and goal-dependent manner (Delgado et al., 2016; Ochsner & Gross, 2014; Ochsner et al., 2012). As such, this region may support affective appraisals more generally rather than directly managing affective self-reflection.
More recently, Satpute et al. (2013) investigated the neural basis of affective self-report, and found the dorsomedial PFC to be particularly tied to directing attention to one’s affective state, while generating verbal descriptions of these states was more specifically associated with the ventrolateral PFC. Similarly, Lieberman et al. (2007) have related the ventrolateral aspect of the PFC to selecting verbal labels of emotion for face stimuli. This evidence suggests that ventrolateral PFC may be more associated with the linguistic component of affect labeling. However, Ochsner, Knierim, et al. (2004) identified this region in a contrast of rating general valence for an emotional picture’s subject versus rating the viewer’s own valence, indicating it may be involved in evaluating the affective states of others more generally. It is worth noting that activation in this contrast was only significant in the left PFC, while Lieberman and colleagues have specifically identified right PFC in their work. Thus, it is possible that some laterality of function exists in this region with regard to evaluating affective states. Nevertheless, the ventrolateral PFC seems to be more related to evaluating affect in others than the self.
In sum, the dorsomedial PFC likely supports affective self-reflection during distancing with various other regions contributing to affective appraisal and labeling processes more generally. This role for dorsomedial PFC converges with the accounts of its function presented above in the discussion on self-projection. These sources point to a general function of evaluating internal states with regard to the self.
Cognitive control.
Distancing, like other active methods of emotion regulation, involves cognitive control processes to support goal-directed cognition. These processes include maintaining a regulatory goal, maintaining the distanced perspective of the stimulus, monitoring regulation (i.e. monitoring changes in output from affective self-reflection), and adjusting cognitive efforts to more effectively meet the regulatory goal. One study comparing the neural correlates of reappraisal and distraction found common activity in the dorsolateral PFC, pre-supplementary motor area (pre-SMA), PCC+, and inferior parietal lobe (Kanske, Heissler, Schonfelder, Bongers, & Wessa, 2011; see also McRae et al., 2010). The authors identified this activity as reflecting a cognitive control network supporting both regulation strategies, and they note its consistency with the activity elicited by executive control tasks such as Stroop or flanker paradigms. Ochsner et al. (2012) proposed a similar network subserving working memory in their model of cognitive regulation, which involved the dorsolateral PFC, pre-SMA, and inferior parietal regions. A recent neuroimaging meta-analysis provided further support for this cognitive control network in reappraisal (Buhle et al., 2014).
In particular, the lateral PFC has been reliably associated with the maintenance and manipulation of information: core functions of working memory and cognitive control more broadly (Braver, 2012; D’Esposito, Postle, Ballard, & Lease, 1999). In a meta-analysis, Wager and Smith (2003) found that activity in dorsolateral PFC was especially recruited by tasks involving continuous updating of working memory contents. Distancing would presumably recruit these working memory functions for maintaining regulatory goals and tracking goal pursuit over time. In addition, in a qualitative comparison of neuroimaging associations with distancing and reinterpretation, distancing was more strongly associated with right dorsolateral PFC activity (Ochsner & Gross, 2008). Ochsner and Gross suggest that this result may reflect more attentional control processes in association with distancing relative to reinterpretation. Right inferior parietal cortex has also been implicated in selective attention as part of a frontoparietal network recruited during reorienting of visuospatial attention (Thiel, Zilles, & Fink, 2004; Yantis et al., 2002). Thus, dorsolateral PFC and inferior parietal cortex may support the maintenance of reappraisals and reorientation of attention to alternative perspectives during distancing (Buhle et al., 2014). The ventrolateral PFC has been implicated in working memory functions along with the dorsolateral aspect in some previous work (D’Esposito et al., 1999); however, the emotion regulation literature collectively relates the dorsolateral PFC more specifically to cognitive control functions in this context.
Monitoring distancing outcomes would likely recruit the anterior cingulate cortex (ACC) or adjacent cortex in the pre-SMA. This reasoning is consistent with Botvinick’s (2007) integrated account of cognitive control in the ACC, descriptions of ACC function within emotional processing (Etkin, Egner, & Kalisch, 2011), and the model of cognitive regulation offered by Ochsner et al. (2012; see also Ochsner & Gross, 2007). Some have argued, though, that the conflict monitoring activity attributed to the ACC may be more accurately localized to nearby pre-SMA cortex (Nachev, Kennard, & Husain, 2008). Regardless of this anatomical debate, distancing likely involves monitoring processes attributed to this general area of cortex, along with other cognitive control processes supported by the dorsolateral PFC and inferior parietal regions.
Neuroimaging Comparisons with Other Emotion Regulation Methods
We are only aware of a handful of studies that have compared the neural bases of distancing to other methods of emotion regulation. One study revealed greater activity in the left inferior parietal lobe and right ventromedial PFC when downregulating with distancing relative to reinterpretation (Ochsner, Ray, et al., 2004). In a review of neuroimaging studies on cognitive regulation, Ochsner et al. (2012) compared the distribution of regions that have been associated with distancing and reinterpretation. The authors noted that parietal regions linked to spatial attention, perspective taking, and agency have been more frequently associated with distancing, whereas the ventrolateral PFC has more often been associated with reinterpretation. They also noted that the distribution of distancing results in the PFC was somewhat right lateralized relative to reinterpretation, possibly reflecting less semantic process and greater spatial and attentional processing in distancing. Dörfel et al. (2014) found further evidence for selective involvement of the right angular gyrus in distancing in a comparison with reinterpretation, distraction, and expressive suppression. Although there is little direct evidence to date, preliminary findings suggest that recruitment of the parietal cortex around the TPJ and perhaps right lateralization within PFC might distinguish distancing from other forms of regulation including reinterpretation.
Neurocognitive Model
The previous sections provide a foundation for conceptualizing distancing in terms of its neurocognitive mechanisms. Here, we integrate this evidence to propose a preliminary model of how various cognitive processes and their underlying neural regions may interact dynamically to support distancing. The architecture of this model is illustrated in Figure 2.
Figure 2.
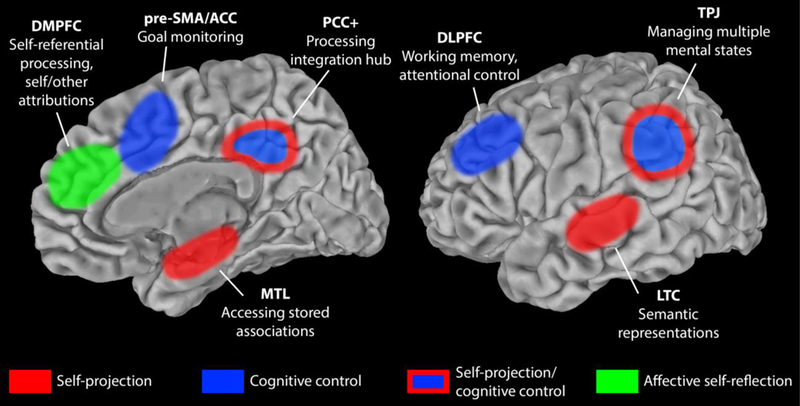
Neurocognitive model of distancing. Regions of interest are highlighted on a single hemisphere for ease of illustration. No laterality claims are intended for medial regions. Distancing may recruit lateral regions bilaterally, but some evidence suggests an emphasis on the right DLPFC and left LTC. ACC = anterior cingulate cortex; DLPFC = dorsolateral prefrontal cortex; DMPFC = dorsomedial prefrontal cortex; LTC = lateral temporal cortex including middle temporal gyrus and superior temporal sulcus; MTL = medial temporal lobe; PCC+ = posterior cingulate cortex and surrounding parietal cortex; pre-SMA = pre-supplementary motor area; TPJ = temporoparietal junction.
Distancing begins with a regulatory goal. In this review, we have focused on the general goal of downregulating a negative emotional response. This goal and the planned distancing technique for achieving it are likely maintained through working memory processes recruiting the dorsolateral PFC. This region is part of a larger network supporting the cognitive control processes involved in distancing. The regulatory goal may be established before or after exposure to an emotionally salient stimulus, depending on whether the event was anticipated or not. Regardless, perception and interpretation of the salient event triggers an initial emotional response. An individual must then recognize this emotional response through self-reflective processes (and deem it undesirable) to motivate explicit regulation through distancing. This affective self-reflection is dependent on activity in the dorsomedial PFC. The current mental state, involving both interpretations of the event and self-evaluations, is likely maintained through activity in control regions including the dorsolateral PFC, TPJ, and PCC+, and can be referenced for comparison during regulation.
Next, the individual engages in constructive processes to simulate an alternative perspective of the event with a different psychological distance. A network of several regions supports this self-projection. The simulated perspective is constructed from mental building blocks that include stored associations and semantic knowledge. Similar to memory-related processing, the MTL likely supports the construction of a mental scene from these stored associations, and the LTC, particularly in the left hemisphere, likely contains semantic representations that are referenced in simulation. The PCC+ serves as a hub to integrate the constructed mental scene with self-referential processing in the dorsomedial PFC. This integration is critical for allowing the mental scene to be experienced as a perspective referenced to the self. Meanwhile, the TPJ may play an important role in maintaining the separation of this projection from other mental states, including the initial, reality-based perspective, and, along with the dorsolateral PFC (Kim, Cilles, Johnson, & Gold, 2012), the TPJ may also enable switching focus between these states, or perspectives.
The subjective experience of the alternative, distanced perspective may then prompt a new emotional response to the event, which would be detected through recapitulation of an affective self-reflection process. In an explicit effort to regulate emotion, changes in affect must be monitored to assess and manage this goal-directed pursuit. This monitoring likely involves activity in the area of the anterior cingulate cortex and pre-SMA. Signals from this area can then be used to update regulation goals and techniques being maintained in the dorsolateral PFC. These processes cycle to support adaptive changes in self-projection and regulation.
Change in emotional response is likely a product of natural emotion generative processes responding to the alternative perspective of the stimulus in place of the original perspective. As such, the effects of distancing on neural mechanisms of emotion generation would likely be indirect. This mechanism of action stands in contrast to the more direct neural mechanism that has been proposed for emotion modulation in fear extinction, for example. In fear conditioning paradigms, activity in ventromedial PFC has been associated with successful retrieval of the safe, or non-fearful, association with the stimulus during extinction (Milad & Quirk, 2012). This activity in the ventromedial PFC is thought to directly attenuate activity in the amygdala, and as a result, attenuate the fear response to the stimulus (but see Fullana et al., 2018). Based on studies of this circuit, two general neural mechanisms for linking emotion regulatory and generative processes in reappraisal have been discussed (Buhle et al., 2014). The first candidate mechanism is that control regions involved in reappraisal upregulate activity in the ventromedial PFC, which results in attenuation of amygdala activity via the circuit described in fear extinction (Schiller & Delgado, 2010; Silvers, Insel, et al., 2016). The second candidate mechanism is that regions involved in reappraisal alter semantic and perceptual representations in LTC in ways that alter the emotional significance of the stimulus, which manifests through natural emotion generative processes. Little work has been done to directly test potential mechanisms linking emotion regulatory and generative systems in the brain in reappraisal (Ochsner et al., 2012), but in their meta-analysis of neuroimaging studies of reappraisal, Buhle et al. found consistent activity in left LTC but not ventromedial PFC, supporting the latter mechanism. This mechanism, based on altering semantic and perceptual representations in the brain, is consistent with the description of reappraisal as mentally transforming the content of the stimulus (reinterpretation) or the viewpoint from which it is observed (distancing). Therefore, distancing likely acts indirectly on emotion generative neural processes and not through one specific circuit.
As for which regions support these emotion generative processes, the relevant regions likely vary based on the specific emotional response. Ochsner and colleagues have previously described a model of cognitive emotion regulation that highlights the ventral and subcortical regions that have been commonly associated with emotion generation in general (see Ochsner et al., 2012), although recent work has suggested that more distributed patterns of activity that include cortical areas likely support various emotional responses (Kragel & LaBar, 2015).
In summary, this model identifies a network of cognitive control regions that manages the alternative perspectives of the stimulus, maintains the goal of regulation, and adapts regulatory processes to achieve this goal. A partially overlapping network of regions supports the construction and experience of an alternative perspective of the relevant event. This new perspective, characterized by a shift in psychological distance, can lead to new emotional responses, which are recognized through self-reflective processing mediated by the dorsomedial PFC.
Future Directions
Theory.
Systematic investigations of the different forms of distancing may support a more nuanced account of the neurocognitive mechanisms discussed in this model, specifically for mechanisms related to self-projection and self-referential processing. As such, the model proposed herein should be considered an initial description. CLT and some preliminary neural evidence suggests that there could be a unified neural substrate for different forms psychological distance in the brain (Parkinson et al., 2014; Tamir & Mitchell, 2011); however, these neuroimaging studies also suggest that some brain areas may be more specialized for specific forms (Peer et al., 2015). Investigations of the differences in neural substrates among these forms may reveal more specific functional subdivisions within the proposed model or additional brain areas related to forms that have not been well represented in the current literature. For instance, the medial PFC is recognized as a key area in social cognition (Amodio & Frith, 2006). In the proposed model, we attribute self-referential processes to the dorsomedial PFC that are general to distancing. Given its putative role in attributions to self versus other, though (Mitchell, Macrae, & Banaji, 2006; Ochsner, Knierim, et al., 2004), it is possible that some processing in this area may relate more specifically to objective distancing, which is most connected to social distance.
More generally, the medial PFC encompasses a large area of cortex, and defining the functional organization of this area has been an ongoing challenge. Some researchers have proposed functional gradients or subdivisions for this area. Mitchell et al. (2006) found a distinction between activity in the ventral and dorsomedial PFC corresponding to judgments more closely related to the self versus dissimilar others, respectively. CLT conceptualizes psychological distance as egocentric, such that representations become more abstract as social distance from the self increases. In light of this theory, these results suggest that dorsal regions of the medial PFC process more abstract representations relative to ventral regions. In contrast, Amodio and Frith (2006) have proposed that representations in the medial PFC become more abstract moving from posterior to more anterior regions. Taren, Venkatraman, and Huettel (2011) have proposed a similar posterior-to-anterior axis of abstraction specifically within the dorsomedial PFC. Thus, the relationship between psychological distance and abstraction would indicate some discrepancy in these descriptions of medial PFC function. Distancing research could aid in clarifying this functional organization. While previous distancing studies have tended to focus simply on increasing psychological distance, more nuanced studies using multiple levels of distance could help to better characterize the functional arrangement of the medial PFC, discriminate activity in this area supporting different aspects of distancing, and evaluate the tenets of CLT.
Similarly, the function of the TPJ has been difficult to define given the wide variety of experimental manipulations that elicit activity in this area. In an effort to make sense of these findings, Carter and Huettel (2013) proposed the Nexus Model for TPJ function, in which different functional regions overlap to define loci with more specific functions. Through this model, the authors attempted to differentiate portions of the TPJ that relate to social cognition from those that do not. Associations with distancing have been widely distributed throughout this area of cortex, and it is possible that different regions in this area contribute distinct functions to distancing. Again, given the particular relationship of objective distancing with social distance, it would be interesting to examine whether this form of distancing specifically recruits some regions of the TPJ related to social cognition that other forms do not. These types of comparisons could potentially validate and inform the functional subdivision proposed in the Nexus Model and characterize the mechanisms of distancing in greater detail.
Application.
Understanding the neural processes underlying distancing creates the potential for brain-targeted interventions. These interventions could include techniques for enhancing emotion regulation through distancing, but they could also include techniques for enhancing specific processes outlined in the proposed model. For example, it may be possible to improve an individual’s ability to manage multiple mental states, even outside of the context of distancing, by modulating activity in specific regions of the TPJ. Targeted forms of brain stimulation are already being employed in the treatment of major depressive disorder (Carpenter et al., 2012), and more brain stimulation treatments are being developed as these technologies and our understanding of neurocognitive mechanisms advance. These methods are particularly suited for modulating activity in superficial cortical areas, making several regions within the proposed model of distancing potential targets (see Figure 2). More research is needed to fully optimize the stimulation parameters of these methods, but they are capable of both inhibiting and facilitating cognitive function (Parkin, Ekhtiari, & Walsh, 2015).
An effective intervention for enhancing distancing or related processing using brain stimulation would require that facilitative effects can persist beyond the period of stimulation. Some research has demonstrated that such sustained benefits to cognitive function are possible (Luber & Lisanby, 2014). Luber et al. (2013) achieved a sustained benefit to working memory performance using a course of stimulation that involved multiple sessions over multiple days during which participants engaged in a working memory task. Improvements in distancing performance may be possible using similar methods. Further basic research is needed to more precisely characterize the mechanisms of distancing and identity specific targets for interventions in clinical disorders. Nevertheless, the proposed model could provide a guide for enhancing interventions for emotion dysregulation using brain stimulation. Preliminary efforts to enhance distancing with brain stimulation are already underway (Feeser, Prehn, Kazzer, Mungee, & Bajbouj, 2014), and the success of these interventions will likely improve as understanding of the relevant brain mechanisms increases.
Meta-Analysis of Brain Areas Associated with Distancing
To provide an initial evaluation of the neurocognitive model proposed here, we performed a meta-analysis of fMRI studies of distancing. We hypothesized the meta-analysis would reveal convergent activation related to distancing in the brain areas included in our model (Figure 2), possibly excepting the dorsomedial PFC. As noted above, affective self-reflection has been consistently associated with the dorsomedial PFC. While we believe this process is a critical component of intentional emotion regulation, it is likely also recruited in the control conditions of distancing experiments, for which participants are generally also required to report on their affective state. Thus, activation in this region may not differ significantly between distancing and control conditions. This analysis expands on previous meta-analyses on the neural mechanisms of reappraisal (Buhle et al., 2014; Diekhof, Geier, Falkai, & Gruber, 2011; Messina, Bianco, Sambin, & Viviani, 2015) by evaluating distancing specifically. Synthesizing these results from the literature provides an assessment of the neural architecture of the model, and we discuss these implications below.
Methods
Because researchers have used inconsistent terminology to refer to distancing techniques (refer to “Obstacles in Distancing Research” above), we employed multiple methods to identify relevant experimental studies. These included research database searches, examining reviews and meta-analytic reports related to reappraisal, and backward search. Figure 3 illustrates the results of these search procedures.
Figure 3.

A flow diagram of the literature search.
We began by searching the PubMed (www.ncbi.nlm.nih.gov/pubmed/) and PsycINFO (http://www.apa.org/pubs/databases/psycinfo/) databases. Initial screening criteria were as follows.
The title or abstract must include the term “distancing,” “reappraisal,” or “detachment” in combination with “functional magnetic resonance imaging,” “functional MRI,” or “fMRI” (nine searches in total for each database).
The year of publication must be 1992 or later (excluding studies predating fMRI).
The report must contain an analysis of original fMRI data (excluding reports consisting solely of review or meta-analyses).
The report must be a journal article (excluding non-peer-reviewed dissertations, etc.)
The report must be in English.
The fourth criterion was implemented using the “journal article” filter in the PubMed search engine and the “Peer Reviewed Journal” filter in the PsycINFO search engine. The remaining criteria were also implemented through search engine filters, with the exception of the third criterion, which was implemented by manually screening the search results after eliminating duplicate reports. Duplicate reports were first eliminated from search results from within databases and then between databases. Of note, one study was excluded after initial screening for failing to meet the third criterion (Koenigsberg et al., 2010). The relevant results from this study were already reported in a previous study that was also included (Koenigsberg et al., 2009), so it was excluded as a duplicate report.
The full texts of the remaining reports were then screened using the following eligibility criteria.
Participants used an emotion regulation technique that exclusively employed one or more forms of distancing.
Distancing was used for the purpose of downregulating negative emotional responses.
Results were available for a group comprised solely of healthy participants (i.e. no known inclusion of participants with neurological or psychiatric conditions).
Results were available from a whole-brain contrast of distancing > natural response.
Coordinates were provided in standard Montreal Neurological Institute (MNI) or Talairach spaces.
Of note, three studies did not meet the fourth criterion because their control conditions were deemed qualitatively different from a non-regulation, natural response control condition (Bruneau, Jacoby, & Saxe, 2015; Silvers, Insel, et al., 2016; Silvers et al., 2015). In each of these studies, the downregulation condition of increasing distance was compared to a condition of decreasing distance as opposed to natural response. Also of note, several studies refer to hypothetical distancing techniques as reinterpretation. As a result, it is possible that some studies of hypothetical distancing were not found in our searches or were excluded based on the first criterion if they did not include sufficient description to distinguish their regulation technique from reinterpretation as we have defined it here. This issue did not likely result in many erroneous exclusions since most studies did provide enough detail about their regulation techniques to support clear decisions.
We then examined reviews and meta-analyses that contained lists of reports that distinguished distancing from other forms of reappraisal to identify additional reports not found in our database searches. The reviews and meta-analytic reports we examined included the following: Buhle et al., 2014; Diekhof, Geier, Falkai, & Gruber, 2011; Frank et al., 2014; Messina, Bianco, Sambin, & Viviani, 2015; Ochsner et al., 2012; Webb et al., 2012; and Zilverstand, Parvaz, & Goldstein, 2016.
Finally, we performed a backward search using all included studies. We identified reports for further screening based on citations within the introduction or methods sections as well as report titles in the reference section. Ultimately, backward search revealed no new, qualifying experimental studies.
In total, 22 studies were included in the meta-analysis (refer to Table A1 for a description of these studies). We also explored the possibility of performing a meta-analysis of the reverse contrast (natural response > distancing) using our selected studies, but the number of studies reporting this contrast did not meet the minimum recommendation for our meta-analytic method (Eickhoff, Laird, P. M. Fox, Lancaster, & P. T. Fox, 2017). For the included studies, only results from whole-brain analyses were included in the meta-analysis, while results from region-of-interest analyses and small-volume correction were excluded. This decision was made to promote maximal consistency in our data set, as the original researchers’ decisions regarding these additional analyses would have varied across studies. Similarly, reported cluster sub-peaks were excluded from analysis, as the original decisions regarding reporting these coordinates and the criteria used would also have added uncontrolled variance into our data set. An exception was made for one study (Hayes et al., 2010), which reported only local maxima with no indication of the clustering.
Meta-analysis was performed using the Activation Likelihood Estimation approach (ALE; Eickhoff, Bzdok, Laird, Kurth, & Fox, 2012; Eickhoff et al., 2017) with the BrainMap GingerALE software (version 2.3.6; brainmap.org/ale/). This meta-analytic technique identifies areas where the included experimental results overlap greater than would be expected by chance. Using the software, all results reported in Talairach space were transformed into the MNI coordinate system, which served as the standard space for our analysis. Based on guidelines recommended by the developers of this method in the software’s manual and peer-reviewed publication (Eickhoff et al., 2012), we ran our analysis using the following settings and parameters: larger mask, Non-Additive Modeled Activation algorithm, cluster-forming threshold of p < .001 uncorrected, and cluster-level inference threshold of .05 with 10,000 permutations.
Results and Discussion
Analysis revealed convergent regions of activation related to distancing in the left and right TPJ, right dorsolateral PFC, left LTC, PCC+, and pre-SMA (Figure 4). Descriptions of significant clusters are provided in Table 2. These regions are consistent with our proposed neurocognitive model of distancing.
Figure 4.
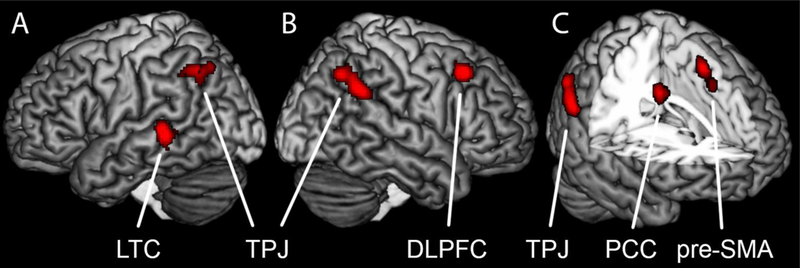
Rendering of convergent activity associated with distancing. Panels A (left hemisphere) and B (right hemisphere) illustrate lateral cortical results. Panel C illustrates medial cortical results using a cutout from a right-anterior view of the brain. DLPFC = dorsolateral prefrontal cortex; LTC = lateral temporal cortex; PCC = posterior cingulate cortex; pre-SMA = pre-supplementary motor area; TPJ = temporoparietal junction.
Table 2.
Cluster Report of Convergent Activity Related to Distancing
| Anatomical locus (Brodmann area) | Volume (mm3) | Peak ALE statistic | MNI coordinates |
||
|---|---|---|---|---|---|
| X | Y | Z | |||
| L middle temporal gyrus (22) | 2072 | 0.028 | −60 | −36 | −2 |
| R middle frontal gyrus (8) | 1296 | 0.028 | 40 | 22 | 42 |
| R supramarginal gyrus (40) | 2560 | 0.023 | 58 | −48 | 32 |
| L superior frontal gyrus (6) | 1840 | 0.021 | −6 | 14 | 58 |
| Posterior cingulate gyrus (23) | 864 | 0.021 | 0 | −22 | 30 |
| L inferior parietal lobule (40) | 1312 | 0.018 | −54 | −58 | 42 |
Notes. ALE = activation likelihood estimation; L = left; MNI = Montreal Neurological Institute; R = right. The ALE statistic is a voxel-wise measure of the convergence of significant results across the experiments included in the meta-analysis. Computationally, it is defined as the estimated probability that a true peak of activation of at least one included experiment lies within the volume of a given voxel (Eickhoff et al., 2009).
In particular, the TPJ, LTC, and PCC+ are all part of the network defined by Spreng et al. (2009) through meta-analyses of abilities that Buckner and Carroll (2007) have related to self-projection. The MTL component of this network was notably absent in our meta-analytic results. Although null results are challenging to interpret, this omission may be due to difficulties in reliably imaging this region using whole-brain approaches and standard pre-processing methods. Offering support for this possibility, several of the studies incorporated in the current meta-analysis reported reduced activity in amygdala in association with distancing, but these results were only obtained through region-of-interest analysis or small volume correction rather than whole-brain contrasts (e.g. Denny, Inhoff, et al., 2015; Walter et al., 2009). Alternatively, it is possible that the simulation required for distancing does not rely on scene construction in a way comparable to recalling events or prospective thinking, and is therefore not dependent on processing in the MTL. Also, these constructive processes may be particularly minimized in laboratory tasks in which the target stimuli of distancing may be freely perceived during the regulation period, while for real-world applications involving autobiographical experiences, they may play a larger role. Spreng et al. also identified left dorsolateral PFC as part of this core network, but we observed convergent activity only in right dorsolateral PFC, consistent with the characterization of distancing by Ochsner et al. (2012). Our results were consistent with other predicted cognitive control regions, particularly the pre-SMA, but also the PCC+ and inferior parietal lobe.
Finally, we might have expected results in medial PFC, in line with the networks involved in self-projection and affective self-reflection; however, as discussed above (see “Affective self-reflection”), engagement of this region is likely common across both distancing and natural response given the explicit instructions to reflect on one’s affective state in both conditions. Recent work also suggests that increasing social distance (as with many of the objective distancing techniques included here) can cause a shift in affective attributions from self to other, which results in less medial PFC activation (Moser et al., 2017; note, participants were not given the goal of downregulating emotion through this manipulation); although, this area is likely still engaged to some degree in the context of explicit emotion regulation so that the individual may track regulation success. Thus, the particular contrasts included in this analysis may have precluded the observation of results in this region.
The neuroimaging literature of distancing is dominated by the objective form of distancing (see Table A1). As such, the literature cannot currently support meta-analytic approaches to differentiating the various forms of distancing, and the present meta-analysis collapsed across all forms of distancing. Additionally, most studies investigated downregulation of negative affect, which led to our exclusion of less prevalent applications, such as decreasing reward anticipation (Staudinger, Erk, Abler, & Walter, 2009; Staudinger, Erk, & Walter, 2011). Characterizing distinctions in the mechanisms supporting different forms of distancing and different regulatory goals are important areas for future research; nevertheless, this meta-analysis fills a critical gap in the literature by synthesizing the neuroimaging findings specific to this tactic of reappraisal.
While the inclusion criteria of the meta-analysis were indeed specific to distancing, it is important to note that the specificity of the results are limited by the neuroimaging contrast. The results suggest the observed regions are sensitive to distancing manipulations relative to natural response, but we would also expect other methods of emotion regulation that share component processes with distancing to have commensurate overlap in brain activation. Specifically, we would expect some degree of activation related to affective self-reflection in other explicit methods of emotion regulation as well as activation related to cognitive control with other cognitively focused techniques. Despite this limitation, the results of this meta-analysis are largely consistent with the neural mechanisms of distancing proposed in the model and thus offer initial support for this account.
Given these results and their agreement with our model, there are many ways in which future research could expand our understanding of how these areas dynamically interact to support distancing. One potential approach would be to search for time-varying co-activation maps during distancing. We would predict such an analysis to reveal networks comparable to those described in the model corresponding to the various processes underlying distancing. Transitions between these brain states during distancing may be beyond the temporal resolution of standard fMRI protocols, but general trends may emerge such as an early bias toward a self-projection network and a late bias toward an affective self-reflection network. Another approach could be to describe the effective connectivity of these brain areas using dynamic causal modelling. Using careful manipulations to distancing procedures, this approach could evaluate neural communications described in the model, such as the PCC+ serving as a hub to integrate the self-projection network with self-referential processing in the dorsomedial PFC. These future directions would further clarify the brain dynamics of distancing and related processes as well as network interactions more generally.
Conclusions
Distancing is an effective and versatile method for regulating emotion. As such, individuals with various forms of emotion dysregulation may benefit from interventions incorporating distancing techniques, but many questions remain about the mechanisms underlying this tactic and how it can best be utilized. Several obstacles have prevented more efficient progress in distancing research. In this review, we have begun to address these challenges and discussed several strategies and directions for further progress.
Currently, it is difficult to synthesize findings from distancing literature due to inconsistency in the terminology used to discuss distancing and emotion regulation. To promote more effective communication in this area, we have integrated extant systems of terminology from the literatures of emotion regulation and psychological distance into a unified taxonomic framework for discussing distancing and emotion regulation more broadly. This framework draws on the process model of emotion regulation, conventions in reappraisal research, and CLT, and it establishes basic categorical levels of strategy, tactic, and technique for descriptions of emotion regulation. Additionally, by applying CLT to distancing, we delineate different forms of this tactic. Distinctions between these forms have received little attention in previous work, but we propose that these forms may be supported by partially distinct mechanisms. Therefore, these forms might differ in their effects. Future research should aim to address these potential differences.
Other key areas for future work involve the theoretical foundations of objective and hypothetical distancing. Objective distancing is the most prevalent form of distancing in emotion regulation research. Despite this fact, its relationship to the dimensions of CLT is the least straightforward. More research is needed to characterize the relationship between objective distancing and social distance and determine what other factors may mediate objectivity. Additionally, hypothetical distancing might not have comparable utility to the other forms of distancing. Future work could help to clarify whether the corresponding CLT dimension of hypotheticality may be fundamentally distinct from the other dimensions in this theory.
Using the conceptual framework of distancing that we have proposed, we were able to consolidate and review studies of distancing to evaluate the effectiveness of this tactic. Overall, this review indicated positive effects of distancing with subjective and physiological measures in a variety of contexts. It has also performed favorably in comparisons with other methods of emotion regulation. Questions remain, though, concerning how to best measure distancing performance, and emotion regulation performance more generally. Self-report of subjective experience has been most common, but the specific measures have varied widely. Amygdala activity has shown some utility as a physiological metric for distancing success, although its utility may be limited to certain emotional contexts and affective representations are known to be more distributed in the brain. The evidence for psychophysiological effects is inconclusive at this point, given the small number of studies and the inconsistent choice of measures across these studies. Differences in distancing techniques may have contributed to some of the inconsistency in neural and psychophysiological effects between studies. We hope that the information provided in this article will help to clarify the communication of distancing methods and establish more consistency in future studies.
Despite the favorable profile of distancing demonstrated through this review, the effects associated with distancing often indicate only moderate levels of regulation, similar to other effective methods in emotion regulation research. We have highlighted several approaches to potentially improve the effectiveness of distancing applications. These include determining whether certain distancing techniques are more effective than others, identifying individual and contextual factors influencing distancing success, and exploring compound regulation techniques that may have greater effect sizes than individual techniques. Little work has been done in these areas, but these approaches could help to optimize distancing, which already demonstrates promising regulatory effects.
Another approach to increasing the effectiveness of distancing involves enhancing the neurocognitive mechanisms that support it. A mechanistic account of distancing that would support this approach was lacking in previous literature. Therefore, we aimed to examine the cognitive and neural processes involved in distancing and offer an initial account of these mechanisms. We have proposed that distancing recruits processes of affective self-reflection and cognitive control, similar to many other methods of emotion regulation, but it is distinguished by self-projection based in the DMN. By reviewing literature on the neural bases of each of these processes, we established predictions regarding the neural systems supporting distancing. A meta-analysis of neuroimaging findings related to distancing and a review of the work comparing distancing to other methods of emotion regulation both provided initial support for these predictions. Based on this evidence, we have proposed a preliminary model of distancing that describes how relevant cognitive and neural processes might unfold during regulation and identifies key functional areas in this tactic in the TPJ, PCC+, PFC, MTL, and LTC.
This model provides a basis for developing brain-targeted interventions for enhancing distancing. Brain stimulation techniques offer a promising approach to selectively modulating cortical activity and cognitive function. Although these techniques are relatively early in development, attempts to apply them to improve distancing performance have already begun. These approaches would benefit from an expansion of our initial model to include more detailed accounts of the functions of specific brain regions in distancing. Some of this detail may be added by future work comparing different forms of distancing. Investigations of these differences may reveal functional subdivisions within some of the cortical areas in our model, particularly the dorsomedial PFC and TPJ.
A substantial body of literature has demonstrated that distancing is an effective tactic for emotion regulation. This fact makes it an appealing target for further research and development, as there are several potential routes for further optimization. At the same time, distancing provides a means for investigating several high-level cognitive processes and their neural bases. We hope that the taxonomy, literature review, and preliminary model offered here will enable researchers to further advance applications and theoretical studies related to distancing.
Highlights.
Distancing is an effective and versatile method of emotion regulation
We relate distancing to construal-level theory of psychological distance
We propose a taxonomy of distancing and emotion regulation
We offer a preliminary neurocognitive model of distancing
We present a meta-analysis of fMRI studies of distancing in support of this model
Acknowledgments
The authors would like to thank Tobias Egner and Daniel Stjepanović for their valuable feedback on previous drafts of this manuscript. This work was supported by the National Science Foundation Graduate Research Fellowship Program [DGE-1644868 to J. P. P.] and NIH [R01 MH113238 to K. S. L.].
Appendix
Table A1.
Description of Studies Included in Meta-Analysis
| Study | N (F/M) | Mean age (SD) | Distancing form(s) |
|---|---|---|---|
| Denny, Fan, et al., 2015 | 11/10 | 29.0 (6.7) | obj |
| Denny, Inhoff, et al., 2015 | 12/5 | 24.1 | obj, spat, temp |
| Domes et al., 2010 | 17/16 | male: 25.2 (1.9); female: 24.6 (1.6) |
obj |
| Dörfel et al., 2014 | 17/0 | within 18–39 | obj |
| Eippert et al., 2007 | 24/0 | 23.3 | obj |
| Erk, Mikschl, et al., 2010 | 8/9 | 43.9 (10.1) | obj |
| Goldin et al., 2008 | 17/0 | 22.7 (3.5) | obj |
| Goldin et al., 2009 | 9/8 | 32.1 (9.3) | obj |
| Hayes et al., 2010 | 11/14 | 21.6 (2.5) | obj |
| Kim & Hamann, 2007 | 10/0 | 20.7 | hyp, obj, spat |
| Koenigsberg et al., 2009 | 9/7 | 31.8 (7.7) | obj |
| Lang et al., 2012 | 15/0 | 24.7 (5.6) | obj |
| Leiberg et al., 2012 | 24/0 | 24.1 | hyp, obj |
| Morris et al., 2012 | 9/6 | 35 (12) | hyp, obj |
| Mulej Bratec et al., 2015 | 20/0 | 24.8 (2.3)* | hyp, obj |
| Paret et al., 2011 | 0/21 | 28 (4) | obj, spat |
| Paschke et al., 2016 | 55/53 | 26.1 (3.7) | obj |
| Pitskel et al., 2011 | 6/9 | 13.0 (2.2) | hyp |
| Walter et al., 2009 | 18/0 | 24 (3)* | obj |
| Winecoff et al., 2011 | 42 (N/A); 20 older adults, 22 young adults |
older adults: 69.0 (3.9); young adults: 23.1 (4.0) |
obj |
| Winecoff et al., 2013 | 21/10 | 25 | obj |
| Xie et al., 2016 | 19/0 | 25.0 (2.3)* | hyp, obj, spat |
Notes. F = female; hyp = hypothetical; M = male; N/A = not reported; obj = objective; spat = spatial; temp = temporal.
reported prior to exclusions
Footnotes
Declarations of interest: none
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Ahmed SP, Somerville LH, & Sebastian CL (2017). Using temporal distancing to regulate emotion in adolescence: Modulation by reactive aggression. Cognition and Emotion, 1–15. 10.1080/02699931.2017.1358698 [DOI] [PubMed]
- Aichhorn M, Perner J, Kronbichler M, Staffen W, & Ladurner G (2006). Do visual perspective tasks need theory of mind? NeuroImage, 30(3), 1059–1068. 10.1016/j.neuroimage.2005.10.026 [DOI] [PubMed] [Google Scholar]
- American Bar Association, American Jury Project. (2005). Principles for juries and jury trials Retrieved from https://www.americanbar.org/content/dam/aba/administrative/american_jury/principles.authcheckdam.pdf
- Amodio DM, & Frith CD (2006). Meeting of minds: The medial frontal cortex and social cognition. Nature Reviews Neuroscience, 7(4), 268–277. 10.1038/nrn1884 [DOI] [PubMed] [Google Scholar]
- Ayduk Ö, & Kross E (2008). Enhancing the pace of recovery: Self-distanced analysis of negative experiences reduces blood pressure reactivity. Psychological Science, 19(3), 229–231. 10.1111/j.1467-9280.2008.02073.x [DOI] [PubMed] [Google Scholar]
- Ayduk Ö, & Kross E (2010). From a distance: Implications of spontaneous self-distancing for adaptive self-reflection. Journal of Personality and Social Psychology, 98(5), 809–829. 10.1037/a0019205 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bar-Anan Y, Liberman N, Trope Y, & Algom D (2007). Automatic processing of psychological distance: Evidence from a Stroop task. Journal of Experimental Psychology: General, 136(4), 610–622. 10.1037/0096-3445.136.4.610 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barrett LF, & Satpute AB (2013). Large-scale brain networks in affective and social neuroscience: Towards an integrative functional architecture of the brain. Current Opinion in Neurobiology, 23(3), 361–372. 10.1016/j.conb.2012.12.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beauregard M, Lévesque J, & Bourgouin P (2001). Neural correlates of conscious self-regulation of emotion. The Journal of Neuroscience, 21(18), RC165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ben-Naim S, Hirschberger G, Ein-Dor T, & Mikulincer M (2013). An experimental study of emotion regulation during relationship conflict interactions: The moderating role of attachment orientations. Emotion, 13(3), 506–519. 10.1037/a0031473 [DOI] [PubMed] [Google Scholar]
- Berking M, Ebert D, Cuijpers P, & Hofmann SG (2013). Emotion regulation skills training enhances the efficacy of inpatient cognitive behavioral therapy for major depressive disorder: A randomized controlled trial. Psychotherapy and Psychosomatics, 82(4), 234–245. 10.1159/000348448 [DOI] [PubMed] [Google Scholar]
- Blanke O, Mohr C, Michel CM, Pascual-Leone A, Brugger P, Seeck M, … Thut G (2005). Linking out-of-body experience and self processing to mental own-body imagery at the temporoparietal junction. Journal of Neuroscience, 25(3), 550–557. 10.1523/JNEUROSCI.2612-04.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Botvinick MM (2007). Conflict monitoring and decision making: Reconciling two perspectives on anterior cingulate function. Cognitive, Affective, & Behavioral Neuroscience, 7(4), 356–366. [DOI] [PubMed] [Google Scholar]
- Braver TS (2012). The variable nature of cognitive control: A dual mechanisms framework. Trends in Cognitive Sciences, 16(2), 106–113. 10.1016/j.tics.2011.12.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruehlman-Senecal E, & Ayduk Ö (2015). This too shall pass: Temporal distance and the regulation of emotional distress. Journal of Personality and Social Psychology, 108(2), 356–375. [DOI] [PubMed] [Google Scholar]
- Bruneau EG, Jacoby N, & Saxe R (2015). Empathic control through coordinated interaction of amygdala, theory of mind and extended pain matrix brain regions. NeuroImage, 114, 105–119. 10.1016/j.neuroimage.2015.04.034 [DOI] [PubMed] [Google Scholar]
- Buckner RL, Andrews-Hanna JR, & Schacter DL (2008). The brain’s default network: Anatomy, function, and relevance to disease. Annals of the New York Academy of Sciences, 1124(1), 1–38. 10.1196/annals.1440.011 [DOI] [PubMed] [Google Scholar]
- Buckner RL, & Carroll DC (2007). Self-projection and the brain. Trends in Cognitive Sciences, 11(2), 49–57. 10.1016/j.tics.2006.11.004 [DOI] [PubMed] [Google Scholar]
- Buhle JT, Silvers JA, Wager TD, Lopez R, Onyemekwu C, Kober H, … Ochsner KN (2014). Cognitive reappraisal of emotion: A meta-analysis of human neuroimaging studies. Cerebral Cortex, 24(11), 2981–2990. 10.1093/cercor/bht154 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Butler EA, Egloff B, Wilhelm FH, Smith NC, Erickson EA, & Gross JJ (2003) The social consequences of expressive suppression. Emotion, 3(1), 48–67. [DOI] [PubMed] [Google Scholar]
- Carpenter LL, Janicak PG, Aaronson ST, Boyadjis T, Brock DG, Cook IA, … Demitrack MA (2012). Transcranial magnetic stimulation (TMS) for major depression: A multisite, naturalistic, observational study of acute treatment outcomes in clinical practice. Depression and Anxiety, 29(7), 587–596. 10.1002/da.21969 [DOI] [PubMed] [Google Scholar]
- Carter RM, & Huettel SA (2013). A nexus model of the temporal–parietal junction. Trends in Cognitive Sciences, 17(7), 328–336. 10.1016/j.tics.2013.05.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavanna AE, & Trimble MR (2006). The precuneus: A review of its functional anatomy and behavioural correlates. Brain, 129(3), 564–583. 10.1093/brain/awl004 [DOI] [PubMed] [Google Scholar]
- D’Argembeau A, & Van der Linden M (2004). Phenomenal characteristics associated with projecting oneself back into the past and forward into the future: Influence of valence and temporal distance. Consciousness and Cognition, 13(4), 844–858. 10.1016/j.concog.2004.07.007 [DOI] [PubMed] [Google Scholar]
- Davis JI, Gross JJ, & Ochsner KN (2011). Psychological distance and emotional experience: What you see is what you get. Emotion, 11(2), 438–444. 10.1037/a0021783 [DOI] [PubMed] [Google Scholar]
- Delgado MR, Beer JS, Fellows LK, Huettel SA, Platt ML, Quirk GJ, & Schiller D (2016). Viewpoints: dialogues on the functional role of the ventromedial prefrontal cortex. Nature Neuroscience, 19(12), 1545–1552. [DOI] [PubMed] [Google Scholar]
- Denny BT, Fan J, Liu X, Ochsner KN, Guerreri S, Mayson SJ, … Koenigsberg HW (2015). Elevated amygdala activity during reappraisal anticipation predicts anxiety in avoidant personality disorder. Journal of Affective Disorders, 172, 1–7. 10.1016/j.jad.2014.09.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denny BT, Inhoff MC, Zerubavel N, Davachi L, & Ochsner KN (2015). Getting over it: Long-lasting effects of emotion regulation on amygdala response. Psychological Science, 26(9), 1377–1388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denny BT, Kober H, Wager TD, & Ochsner KN (2012). A meta-analysis of functional neuroimaging studies of self- and other judgments reveals a spatial gradient for mentalizing in medial prefrontal cortex. Journal of Cognitive Neuroscience, 24(8), 1742–1752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denny BT, & Ochsner KN (2014). Behavioral effects of longitudinal training in cognitive reappraisal. Emotion, 14(2), 425–433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D’Esposito M, Postle BR, Ballard D, & Lease J (1999). Maintenance versus manipulation of information held in working memory: An event-related fMRI study. Brain and Cognition, 41, 66–86. [DOI] [PubMed] [Google Scholar]
- Diekhof EK, Geier K, Falkai P, & Gruber O (2011). Fear is only as deep as the mind allows: A coordinate-based meta-analysis of neuroimaging studies on the regulation of negative affect. NeuroImage, 58(1), 275–285. 10.1016/j.neuroimage.2011.05.073 [DOI] [PubMed] [Google Scholar]
- Dillon DG, & LaBar KS (2005). Startle modulation during conscious emotion regulation is arousal-dependent. Behavioral Neuroscience, 119(4), 1118–1124. [DOI] [PubMed] [Google Scholar]
- Dillon DG, Ritchey M, Johnson BD, & LaBar KS (2007). Dissociable effects of conscious emotion regulation strategies on explicit and implicit memory. Emotion, 7(2), 354–365. 10.1037/1528-3542.7.2.354 [DOI] [PubMed] [Google Scholar]
- Domes G, Schulze L, Böttger M, Grossmann A, Hauenstein K, Wirtz PH, … Herpertz SC (2010). The neural correlates of sex differences in emotional reactivity and emotion regulation. Human Brain Mapping, 31(5), 758–769. 10.1002/hbm.20903 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dörfel D, Lamke J-P, Hummel F, Wagner U, Erk S, & Walter H (2014). Common and differential neural networks of emotion regulation by detachment, reinterpretation, distraction, and expressive suppression: A comparative fMRI investigation. NeuroImage, 101, 298–309. [DOI] [PubMed] [Google Scholar]
- Eickhoff SB, Bzdok D, Laird AR, Kurth F, & Fox PT (2012). Activation likelihood estimation meta-analysis revisited. NeuroImage, 59(3), 2349–2361. 10.1016/j.neuroimage.2011.09.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eickhoff SB, Laird AR, Fox PM, Lancaster JL, & Fox PT (2017). Implementation errors in the GingerALE software: Description and recommendations. Human Brain Mapping, 38(1), 7–11. 10.1002/hbm.23342 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eickhoff SB, Laird AR, Grefkes C, Wang LE, Zilles K, & Fox PT (2009). Coordinate-based activation likelihood estimation meta-analysis of neuroimaging data: A random-effects approach based on empirical estimates of spatial uncertainty. Human Brain Mapping, 30, 2907–2926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eippert F, Veit R, Weiskopf N, Erb M, Birbaumer N, & Anders S (2007). Regulation of emotional responses elicited by threat-related stimuli. Human Brain Mapping, 28(5), 409–423. 10.1002/hbm.20291 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erk S, Mikschl A, Stier S, Ciaramidaro A, Gapp V, Weber B, & Walter H (2010). Acute and sustained effects of cognitive emotion regulation in major depression. Journal of Neuroscience, 30(47), 15726–15734. 10.1523/JNEUROSCI.1856-10.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erk S, von Kalckreuth A, & Walter H (2010). Neural long-term effects of emotion regulation on episodic memory processes. Neuropsychologia, 48(4), 989–996. 10.1016/j.neuropsychologia.2009.11.022 [DOI] [PubMed] [Google Scholar]
- Etkin A, Egner T, & Kalisch R (2011). Emotional processing in anterior cingulate and medial prefrontal cortex. Trends in Cognitive Sciences, 15(2), 85–93. 10.1016/j.tics.2010.11.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feeser M, Prehn K, Kazzer P, Mungee A, & Bajbouj M (2014). Transcranial direct current stimulation enhances cognitive control during emotion regulation. Brain Stimulation, 7, 105–112. [DOI] [PubMed] [Google Scholar]
- Frank DW, Dewitt M, Hudgens-Haney M, Schaeffer DJ, Ball BH, Schwarz NF, … Sabatinelli D (2014). Emotion regulation: Quantitative meta-analysis of functional activation and deactivation. Neuroscience & Biobehavioral Reviews, 45, 202–211. 10.1016/j.neubiorev.2014.06.010 [DOI] [PubMed] [Google Scholar]
- Frith CD, & Frith U (2006). The neural basis of mentalizing. Neuron, 50(4), 531–534. 10.1016/j.neuron.2006.05.001 [DOI] [PubMed] [Google Scholar]
- Fullana MA, Albajes-Eizagirre A, Soriano-Mas C, Vervliet B, Cardoner N, Benet O, … Harrison BJ (2018). Fear extinction in the human brain: A meta-analysis of fMRI studies in healthy participants. Neuroscience & Biobehavioral Reviews, 88, 16–25. [DOI] [PubMed] [Google Scholar]
- Gaebler M, Daniels J, Lamke J-P, Fydrich T, & Walter H (2014). Behavioural and neural correlates of self-focused emotion regulation in social anxiety disorder. Journal of Psychiatry & Neuroscience, 39(4), 249–258. 10.1503/jpn.130080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldin PR, Manber T, Hakimi S, Canli T, & Gross JJ (2009). Neural bases of social anxiety disorder: Emotional reactivity and cognitive regulation during social and physical threat. Archives of General Psychiatry, 66(2), 170–180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldin PR, McRae K, Ramel W, & Gross JJ (2008). The neural bases of emotion regulation: Reappraisal and suppression of negative emotion. Biological Psychiatry, 63(6), 577–586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grecucci A, De Pisapia N, Thero DK, Paladino MP, Venuti P, & Job R (2015). Baseline and strategic effects behind mindful emotion regulation: Behavioral and physiological investigation. PloS One, 10(1), e0116541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gross JJ (1998a). Antecedent-and response-focused emotion regulation: Divergent consequences for experience, expression, and physiology. Journal of Personality and Social Psychology, 74(1), 224. [DOI] [PubMed] [Google Scholar]
- Gross JJ (1998b). The emerging field of emotion regulation: An integrative review. Review of General Psychology, 2(3), 271–299. [Google Scholar]
- Gross JJ (2015). Emotion regulation: Current status and future prospects. Psychological Inquiry, 26(1), 1–26. 10.1080/1047840X.2014.940781 [DOI] [Google Scholar]
- Gruber J, Hay AC, & Gross JJ (2014). Rethinking emotion: Cognitive reappraisal is an effective positive and negative emotion regulation strategy in bipolar disorder. Emotion, 14(2), 388–396. 10.1037/a0035249 [DOI] [PubMed] [Google Scholar]
- Hariri AR, Drabant EM, Munoz KE, Kolachana BS, Mattay VS, Egan MF, & Weinberger DR (2005). A susceptibility gene for affective disorders and the response of the human amygdala. Archives of General Psychiatry, 62(2), 146–152. [DOI] [PubMed] [Google Scholar]
- Harenski CL, Kim SH, & Hamann S (2009). Neuroticism and psychopathy predict brain activation during moral and nonmoral emotion regulation. Cognitive Affective & Behavioral Neuroscience, 9, 1–15. 10.3758/CABN.9.1.1 [DOI] [PubMed] [Google Scholar]
- Hassabis D, & Maguire EA (2007). Deconstructing episodic memory with construction. Trends in Cognitive Sciences, 11(7), 299–306. 10.1016/j.tics.2007.05.001 [DOI] [PubMed] [Google Scholar]
- Hayes JP, Morey RA, Petty CM, Seth S, Smoski MJ, McCarthy G, & LaBar KS (2010). Staying cool when things get hot: Emotion regulation modulates neural mechanisms of memory encoding. Frontiers in Human Neuroscience, 4 10.3389/fnhum.2010.00230 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hermann A, Schäfer A, Walter B, Stark R, Vaitl D, & Schienle A (2009). Emotion regulation in spider phobia: Role of the medial prefrontal cortex. Social Cognitive and Affective Neuroscience, 4(3), 257–267. 10.1093/scan/nsp013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Insel T (2011, September 28). The global cost of mental Illness Retrieved from http://www.nimh.nih.gov/about/director/2011/the-global-cost-of-mental-illness.shtml
- Ionta S, Perruchoud D, Draganski B, & Blanke O (2012). Body context and posture affect mental imagery of hands. PloS One, 7(3), e34382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanske P, Heissler J, Schonfelder S, Bongers A, & Wessa M (2011). How to regulate emotion? Neural networks for reappraisal and distraction. Cerebral Cortex, 21(6), 1379–1388. 10.1093/cercor/bhq216 [DOI] [PubMed] [Google Scholar]
- Kim C, Cilles SE, Johnson NF, & Gold BT (2012). Domain general and domain preferential brain regions associated with different types of task switching: A meta-analysis. Human Brain Mapping, 33(1), 130–142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim S, & Hamann SB (2007). Neural correlates of positive and negative emotion regulation. Journal of Cognitive Neuroscience, 19(5), 776–798. [DOI] [PubMed] [Google Scholar]
- Koenigsberg HW, Fan J, Ochsner KN, Liu X, Guise KG, Pizzarello S, … Siever LJ (2009). Neural correlates of the use of psychological distancing to regulate responses to negative social cues: A study of patients with borderline personality disorder. Biological Psychiatry, 66(9), 854–863. 10.1016/j.biopsych.2009.06.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koenigsberg HW, Fan J, Ochsner KN, Liu X, Guise K, Pizzarello S, … Siever LJ (2010). Neural correlates of using distancing to regulate emotional responses to social situations. Neuropsychologia, 48(6), 1813–1822. 10.1016/j.neuropsychologia.2010.03.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kragel PA, & LaBar KS (2014). Advancing Emotion Theory with Multivariate Pattern Classification. Emotion Review, 6(2), 160–174. 10.1177/1754073913512519 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kragel PA, & LaBar KS (2015). Multivariate neural biomarkers of emotional states are categorically distinct. Social Cognitive and Affective Neuroscience, 10(11), 1437–1448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kragel PA, & LaBar KS (2016). Decoding the nature of emotion in the brain. Trends in Cognitive Sciences, 20, 444–455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kring AM, & Sloan DM (2010). Emotion regulation and psychopathology: A transdiagnostic approach to etiology and treatment New York, NY: Guilford Press. [Google Scholar]
- Kross E, & Ayduk Ö (2008). Facilitating adaptive emotional analysis: Distinguishing distanced-analysis of depressive experiences from immersed-analysis and distraction. Personality and Social Psychology Bulletin, 34(7), 924–938. 10.1177/0146167208315938 [DOI] [PubMed] [Google Scholar]
- Kurczek J, Wechsler E, Ahuja S, Jensen U, Cohen NJ, Tranel D, & Duff M (2015). Differential contributions of hippocampus and medial prefrontal cortex to self-projection and self-referential processing. Neuropsychologia, 73, 116–126. 10.1016/j.neuropsychologia.2015.05.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- LaBar KS, Gitelman DR, Mesulam MM, & Parrish TB (2001). Impact of signal-to-noise on functional MRI of the human amygdala. NeuroReport, 12(16), 3461–3464. [DOI] [PubMed] [Google Scholar]
- Lane RD, Fink GR, Chau PM-L, & Dolan RJ (1997). Neural activation during selective attention to subjective emotional responses. NeuroReport, 8, 3969–3972. [DOI] [PubMed] [Google Scholar]
- Lang S, Kotchoubey B, Frick C, Spitzer C, Grabe HJ, & Barnow S (2012). Cognitive reappraisal in trauma-exposed women with borderline personality disorder. NeuroImage, 59(2), 1727–1734. 10.1016/j.neuroimage.2011.08.061 [DOI] [PubMed] [Google Scholar]
- Leiberg S, Eippert F, Veit R, & Anders S (2012). Intentional social distance regulation alters affective responses towards victims of violence: An FMRI study. Human Brain Mapping, 33(10), 2464–2476. 10.1002/hbm.21376 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lévesque J, Eugene F, Joanette Y, Paquette V, Mensour B, Beaudoin G, … Beauregard M (2003). Neural circuitry underlying voluntary suppression of sadness. Biological Psychiatry, 53(6), 502–510. [DOI] [PubMed] [Google Scholar]
- Lieberman MD, Eisenberger NI, Crockett MJ, Tom SM, Pfeifer JH, & Way BM (2007). Putting feelings into words: Affect labeling disrupts amygdala activity in response to affective stimuli. Psychological Science, 18(5), 421–428. 10.1111/j.1467-9280.2007.01916.x [DOI] [PubMed] [Google Scholar]
- Lu Q, & Stanton AL (2010). How benefits of expressive writing vary as a function of writing instructions, ethnicity and ambivalence over emotional expression. Psychology & Health, 25(6), 669–684. 10.1080/08870440902883196 [DOI] [PubMed] [Google Scholar]
- Luber B, & Lisanby SH (2014). Enhancement of human cognitive performance using transcranial magnetic stimulation (TMS). NeuroImage, 85, 961–970. 10.1016/j.neuroimage.2013.06.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luber B, Steffener J, Tucker A, Habeck C, Peterchev AV, Deng Z-D, … Lisanby SH (2013). Extended remediation of sleep deprived-induced working memory deficits using fMRI-guided transcranial magnetic stimulation. Sleep, 36(6), 857–871. 10.5665/sleep.2712 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McRae K, Ciesielski B, & Gross JJ (2012). Unpacking cognitive reappraisal: Goals, tactics, and outcomes. Emotion, 12(2), 250–255. 10.1037/a0026351 [DOI] [PubMed] [Google Scholar]
- McRae K, Hughes B, Chopra S, Gabrieli JDE, Gross JJ, & Ochsner KN (2010). The neural bases of distraction and reappraisal. Journal of Cognitive Neuroscience, 22(2), 248–262. 10.1162/jocn.2009.21243 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Messina I, Bianco S, Sambin M, & Viviani R (2015). Executive and semantic processes in reappraisal of negative stimuli: Insights from a meta-analysis of neuroimaging studies. Frontiers in Psychology, 6 10.3389/fpsyg.2015.00956 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milad MR, & Quirk GJ (2012). Fear extinction as a model for translational neuroscience: Ten years of progress. Annual Review of Psychology, 63(1), 129–151. 10.1146/annurev.psych.121208.131631 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitchell JP (2009). Inferences about mental states. Philosophical transactions of the Royal Society of London. Series B, Biological sciences, 364(1521), 1309–1316. 10.1098/rstb.2008.0318 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitchell JP, Macrae CN, & Banaji MR (2006). Dissociable medial prefrontal contributions to judgments of similar and dissimilar others. Neuron, 50(4), 655–663. 10.1016/j.neuron.2006.03.040 [DOI] [PubMed] [Google Scholar]
- Morris RW, Sparks A, Mitchell PB, Weickert CS, & Green MJ (2012). Lack of cortico-limbic coupling in bipolar disorder and schizophrenia during emotion regulation. Translational Psychiatry, 2(3), e90 10.1038/tp.2012.16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moser JS, Dougherty A, Mattson WI, Katz B, Moran TP, Guevarra D, … Kross E (2017). Third-person self-talk facilitates emotion regulation without engaging cognitive control: Converging evidence from ERP and fMRI. Scientific Reports, 7(1), 4519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mulej Bratec S, Xie X, Schmid G, Doll A, Schilbach L, Zimmer C, … Sorg C (2015). Cognitive emotion regulation enhances aversive prediction error activity while reducing emotional responses. NeuroImage, 123, 138–148. 10.1016/j.neuroimage.2015.08.038 [DOI] [PubMed] [Google Scholar]
- Nachev P, Kennard C, & Husain M (2008). Functional role of the supplementary and pre-supplementary motor areas. Nature Reviews Neuroscience, 9(11), 856–869. 10.1038/nrn2478 [DOI] [PubMed] [Google Scholar]
- Northoff G, Heinzel A, de Greck M, Bermpohl F, Dobrowolny H, & Panksepp J (2006). Self-referential processing in our brain—A meta-analysis of imaging studies on the self. NeuroImage, 31(1), 440–457. 10.1016/j.neuroimage.2005.12.002 [DOI] [PubMed] [Google Scholar]
- Ochsner KN, Bunge SA, Gross JJ, & Gabrieli JDE (2002). Rethinking feelings: An fMRI study of the cognitive regulation of emotion. Journal of Cognitive Neuroscience, 14(8), 1214–1229. [DOI] [PubMed] [Google Scholar]
- Ochsner KN, & Gross JJ (2007). The neural architecture of emotion regulation. In Gross JJ (Ed.), Handbook of Emotion Regulation (pp. 87–109). New York, NY: Guilford Press. [Google Scholar]
- Ochsner KN, & Gross JJ (2008). Cognitive emotion regulation insights from social cognitive and affective neuroscience. Current Directions in Psychological Science, 17(2), 153–158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ochsner KN, & Gross JJ (2014). The neural bases of emotion and emotion regulation: A valuation perspective. In Handbook of emotion regulation (2nd ed.; pp. 23–42). New York, NY: Guilford Press. [Google Scholar]
- Ochsner KN, Knierim K, Ludlow DH, Hanelin J, Ramachandran T, Glover G, & Mackey SC (2004). Reflecting upon feelings: An fMRI study of neural systems supporting the attribution of emotion to self and other. Journal of Cognitive Neuroscience, 16(10), 1746–1772. [DOI] [PubMed] [Google Scholar]
- Ochsner KN, Ray RD, Cooper JC, Robertson ER, Chopra S, Gabrieli JDE, & Gross JJ (2004). For better or for worse: Neural systems supporting the cognitive down-and up-regulation of negative emotion. NeuroImage, 23(2), 483–499. 10.1016/j.neuroimage.2004.06.030 [DOI] [PubMed] [Google Scholar]
- Ochsner KN, Silvers JA, & Buhle JT (2012). Functional imaging studies of emotion regulation: A synthetic review and evolving model of the cognitive control of emotion. Annals of the New York Academy of Sciences, 1251(1), E1–E24. 10.1111/j.1749-6632.2012.06751.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Opialla S, Lutz J, Scherpiet S, Hittmeyer A, Jäncke L, Rufer M, … Brühl AB (2015). Neural circuits of emotion regulation: A comparison of mindfulness-based and cognitive reappraisal strategies. European Archives of Psychiatry and Clinical Neuroscience, 265(1), 45–55. 10.1007/s00406-014-0510-z [DOI] [PubMed] [Google Scholar]
- Outhred T, Das P, Dobson-Stone C, Felmingham KL, Bryant RA, Nathan PJ, … Kemp AH (2016). Impact of 5-HTTLPR on SSRI serotonin transporter blockade during emotion regulation: A preliminary fMRI study. Journal of Affective Disorders, 196, 11–19. 10.1016/j.jad.2016.02.019 [DOI] [PubMed] [Google Scholar]
- Outhred T, Das P, Felmingham KL, Bryant RA, Nathan PJ, Malhi GS, & Kemp AH (2015). Facilitation of emotion regulation with a single dose of escitalopram: A randomized fMRI study. Psychiatry Research: Neuroimaging, 233(3), 451–457. 10.1016/j.pscychresns.2015.07.018 [DOI] [PubMed] [Google Scholar]
- Paret C, Brenninkmeyer J, Meyer B, Yuen KSL, Gartmann N, Mechias M-L, & Kalisch R (2011). A test for the implementation-maintenance model of reappraisal. Frontiers in Psychology, 2 10.3389/fpsyg.2011.00216 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parkin BL, Ekhtiari H, & Walsh VF (2015). Non-invasive human brain stimulation in cognitive neuroscience: A primer. Neuron, 87(5), 932–945. 10.1016/j.neuron.2015.07.032 [DOI] [PubMed] [Google Scholar]
- Parkinson C, Liu S, & Wheatley T (2014). A common cortical metric for spatial, temporal, and social distance. Journal of Neuroscience, 34(5), 1979–1987. 10.1523/JNEUROSCI.2159-13.2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paschke LM, Dörfel D, Steimke R, Trempler I, Magrabi A, Ludwig VU, … Walter H (2016). Individual differences in self-reported self-control predict successful emotion regulation. Social Cognitive and Affective Neuroscience, 11(8), 1193–1204. 10.1093/scan/nsw036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peer M, Salomon R, Goldberg I, Blanke O, & Arzy S (2015). Brain system for mental orientation in space, time, and person. Proceedings of the National Academy of Sciences, 112(35), 11072–11077. 10.1073/pnas.1504242112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phan KL, Wager T, Taylor SF, & Liberzon I (2002). Functional neuroanatomy of emotion: A meta-analysis of emotion activation studies in PET and fMRI. NeuroImage, 16(2), 331–348. 10.1006/nimg.2002.1087 [DOI] [PubMed] [Google Scholar]
- Pitskel NB, Bolling DZ, Kaiser MD, Crowley MJ, & Pelphrey KA (2011). How grossed out are you? The neural bases of emotion regulation from childhood to adolescence. Developmental Cognitive Neuroscience, 1(3), 324–337. 10.1016/j.dcn.2011.03.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richards JM, & Gross JJ (2000). Emotion regulation and memory: The cognitive costs of keeping one’s cool. Journal of Personality and Social Psychology, 79(3), 410. [DOI] [PubMed] [Google Scholar]
- Satpute AB, Shu J, Weber J, Roy M, & Ochsner KN (2013). The functional neural architecture of self-reports of affective experience. Biological Psychiatry, 73(7), 631–638. 10.1016/j.biopsych.2012.10.001 [DOI] [PubMed] [Google Scholar]
- Saxe R, & Kanwisher N (2003). People thinking about thinking people: The role of the temporo-parietal junction in “theory of mind.” NeuroImage, 19(4), 1835–1842. 10.1016/S1053-8119(03)00230-1 [DOI] [PubMed] [Google Scholar]
- Schardt DM, Erk S, Nüsser C, Nöthen MM, Cichon S, Rietschel M, … Walter H (2010). Volition diminishes genetically mediated amygdala hyperreactivity. NeuroImage, 53(3), 943–951. 10.1016/j.neuroimage.2009.11.078 [DOI] [PubMed] [Google Scholar]
- Schiller D, & Delgado MR (2010). Overlapping neural systems mediating extinction, reversal and regulation of fear. Trends in Cognitive Sciences, 14(6), 268–276. 10.1016/j.tics.2010.04.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schindler A, & Bartels A (2013). Parietal cortex codes for egocentric space beyond the field of view. Current Biology, 23(2), 177–182. 10.1016/j.cub.2012.11.060 [DOI] [PubMed] [Google Scholar]
- Schmeichel BJ, Volokhov RN, & Demaree HA (2008). Working memory capacity and the self-regulation of emotional expression and experience. Journal of Personality and Social Psychology, 95(6), 1526–1540. 10.1037/a0013345 [DOI] [PubMed] [Google Scholar]
- Schulze L, Domes G, Krüger A, Berger C, Fleischer M, Prehn K, … Herpertz SC (2011). Neuronal correlates of cognitive reappraisal in borderline patients with affective instability. Biological Psychiatry, 69(6), 564–573. 10.1016/j.biopsych.2010.10.025 [DOI] [PubMed] [Google Scholar]
- Schurz M, Aichhorn M, Martin A, & Perner J (2013). Common brain areas engaged in false belief reasoning and visual perspective taking: A meta-analysis of functional brain imaging studies. Frontiers in Human Neuroscience, 7(712). 10.3389/fnhum.2013.00712 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheppes G, Scheibe S, Suri G, & Gross JJ (2011). Emotion-regulation choice. Psychological Science, 22(11), 1391–1396. 10.1177/0956797611418350 [DOI] [PubMed] [Google Scholar]
- Shiota MN, & Levenson RW (2009). Effects of aging on experimentally instructed detached reappraisal, positive reappraisal, and emotional behavior suppression. Psychology and Aging, 24(4), 890–900. 10.1037/a0017896 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silvers JA, Insel C, Powers A, Franz P, Helion C, Martin RE, … Ochsner KN (2016). vlPFC–vmPFC–amygdala interactions underlie age-related differences in cognitive regulation of emotion. Cerebral Cortex 10.1093/cercor/bhw073 [DOI] [PMC free article] [PubMed]
- Silvers JA, Shu J, Hubbard AD, Weber J, & Ochsner KN (2015). Concurrent and lasting effects of emotion regulation on amygdala response in adolescence and young adulthood. Developmental Science, 18(5), 771–784. 10.1111/desc.12260 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smoski MJ, LaBar KS, & Steffens DC (2014). Relative effectiveness of reappraisal and distraction in regulating emotion in late-life depression. The American Journal of Geriatric Psychiatry, 22(9), 898–907. 10.1016/j.jagp.2013.01.070 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spreng RN, Mar RA, & Kim AS (2009). The common neural basis of autobiographical memory, prospection, navigation, theory of mind, and the default mode: A quantitative meta-analysis. Journal of Cognitive Neuroscience, 21(3), 489–510. [DOI] [PubMed] [Google Scholar]
- Staudinger MR, Erk S, Abler B, & Walter H (2009). Cognitive reappraisal modulates expected value and prediction error encoding in the ventral striatum. NeuroImage, 47(2), 713–721. 10.1016/j.neuroimage.2009.04.095 [DOI] [PubMed] [Google Scholar]
- Staudinger MR, Erk S, & Walter H (2011). Dorsolateral prefrontal cortex modulates striatal reward encoding during reappraisal of reward anticipation. Cerebral Cortex, 21(11), 2578–2588. 10.1093/cercor/bhr041 [DOI] [PubMed] [Google Scholar]
- Stephan E, Liberman N, & Trope Y (2010). Politeness and psychological distance: A construal level perspective. Journal of Personality and Social Psychology, 98(2), 268–280. 10.1037/a0016960 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tamir DI, & Mitchell JP (2011). The default network distinguishes construals of proximal versus distal events. Journal of Cognitive Neuroscience, 23(10), 2945–2955. [DOI] [PubMed] [Google Scholar]
- Taren AA, Venkatraman V, & Huettel SA (2011). A parallel functional topography between medial and lateral prefrontal cortex: Evidence and implications for cognitive control. Journal of Neuroscience, 31(13), 5026–5031. 10.1523/JNEUROSCI.5762-10.2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thiel CM, Zilles K, & Fink GR (2004). Cerebral correlates of alerting, orienting and reorienting of visuospatial attention: An event-related fMRI study. NeuroImage, 21(1), 318–328. 10.1016/j.neuroimage.2003.08.044 [DOI] [PubMed] [Google Scholar]
- Trope Y, & Liberman N (2010). Construal-level theory of psychological distance. Psychological Review, 117(2), 440–463. 10.1037/a0018963 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vrtička P, Bondolfi G, Sander D, & Vuilleumier P (2012). The neural substrates of social emotion perception and regulation are modulated by adult attachment style. Social Neuroscience, 7(5), 473–493. 10.1080/17470919.2011.647410 [DOI] [PubMed] [Google Scholar]
- Vrtička P, Sander D, & Vuilleumier P (2011). Effects of emotion regulation strategy on brain responses to the valence and social content of visual scenes. Neuropsychologia, 49(5), 1067–1082. 10.1016/j.neuropsychologia.2011.02.020 [DOI] [PubMed] [Google Scholar]
- Wager TD, & Smith EE (2003). Neuroimaging studies of working memory: A meta-analysis. Cognitive, Affective, & Behavioral Neuroscience, 3(4), 255–274. [DOI] [PubMed] [Google Scholar]
- Walter H, von Kalckreuth A, Schardt D, Stephan A, Goschke T, & Erk S (2009). The temporal dynamics of voluntary emotion regulation. PloS One, 4(8), e6726 10.1371/journal.pone.0006726 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y-L, Lin Y-C, Huang C-L, & Yeh K-H (2012). Benefitting from a different perspective: The effect of a complementary matching of psychological distance and habitual perspective on emotion regulation. Asian Journal of Social Psychology, 15(3), 198–207. 10.1111/j.1467-839X.2012.01372.x [DOI] [Google Scholar]
- Webb TL, Miles E, & Sheeran P (2012). Dealing with feeling: A meta-analysis of the effectiveness of strategies derived from the process model of emotion regulation. Psychological Bulletin, 138(4), 775–808. 10.1037/a0027600 [DOI] [PubMed] [Google Scholar]
- White RE, Kross E, & Duckworth AL (2015). Spontaneous self-distancing and adaptive self-reflection across adolescence. Child Development, 86(4), 1272–1281. 10.1111/cdev.12370 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winecoff A, Clithero JA, Carter RM, Bergman SR, Wang L, & Huettel SA (2013). Ventromedial prefrontal cortex encodes emotional value. Journal of Neuroscience, 33(27), 11032–11039. 10.1523/JNEUROSCI.4317-12.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winecoff A, LaBar KS, Madden DJ, Cabeza R, & Huettel SA (2011). Cognitive and neural contributors to emotion regulation in aging. Social Cognitive and Affective Neuroscience, 6(2), 165–176. 10.1093/scan/nsq030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wisco BE, Marx BP, Sloan DM, Gorman KR, Kulish AL, & Pineles SL (2015). Self-distancing from trauma memories reduces physiological but not subjective emotional reactivity among veterans with posttraumatic stress disorder. Clinical Psychological Science, 3(6), 956–963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie X, Mulej Bratec S, Schmid G, Meng C, Doll A, Wohlschläger A, … Sorg C (2016). How do you make me feel better? Social cognitive emotion regulation and the default mode network. NeuroImage, 134, 270–280. 10.1016/j.neuroimage.2016.04.015 [DOI] [PubMed] [Google Scholar]
- Yantis S, Schwarzbach J, Serences JT, Carlson RL, Steinmetz MA, Pekar JJ, & Courtney SM (2002). Transient neural activity in human parietal cortex during spatial attention shifts. Nature Neuroscience, 5(10), 995–1002. 10.1038/nn921 [DOI] [PubMed] [Google Scholar]
- Zaki J (2014). Empathy: A motivated account. Psychological Bulletin, 140(6), 1608–1647. 10.1037/a0037679 [DOI] [PubMed] [Google Scholar]
- Zilverstand A, Parvaz MA, & Goldstein RZ (2016). Neuroimaging cognitive reappraisal in clinical populations to define neural targets for enhancing emotion regulation: A systematic review. NeuroImage 10.1016/j.neuroimage.2016.06.009 [DOI] [PMC free article] [PubMed]