

Abstract
Mosquitoes, like other insects, produce a diversity of peptide hormones that are processed from different precursor proteins and have a range of activities. Early studies relied on purification of bioactive peptides for hormone identification, but more recently genomic data have provided the information needed to more comprehensively identify peptide hormone genes and associated receptors. The first part of this chapter summarizes the known or predicted peptide hormones that are produced by mosquitoes. The second part of this chapter discusses the sources of these molecules and their functions.
1. INTRODUCTION
All multicellular organisms produce peptide hormones that are classified on the basis of shared structural features and/or function (Fricker, 2012). Most derive from precursors called prepropeptides that are produced and enzymatically processed in different types of endocrine or neurosecretory cells. Endocrine cells usually store and directly release peptide hormones into circulation, whereas neurosecretory cells can either release hormones directly or transport them through axons to neurohemal organs or other locations where they are stored and released (Näassel and Winther, 2010; Szapiro and Barbour, 2009). Peptide hormones mediate activities by binding to membrane receptors that activate intracellular signalling and effector functions in different types of cells. A majority of peptide hormones bind receptors in the large G protein-coupled receptor (GPCR) family, but some bind protein kinase receptors (PKRs) or receptor guanylyl cyclases (RGCs) (Biarc et al., 2011; Lemmon and Schlessinger, 2010; Potter, 2011). The focus of this chapter is the peptide hormones of mosquitoes. We first discuss the known or predicted peptide hormones mosquitoes produce and the receptors they bind. We then examine the sources of these hormones and their functions. Insects, including mosquitoes, use some peptide hormones as neurotransmitters, while also producing other peptides that have important roles in development, locomotion, circadian rhythms, olfaction, and mating. We largely do not discuss these topics because of space constraints and/or the limited mosquito literature that is available.
2. PEPTIDE HORMONES AND RECEPTOR DISCOVERY IN MOSQUITOES
The first endocrine studies in mosquitoes began in the mid-20th century with experiments showing that a humoral factor released from the head of female mosquitoes stimulated development of eggs after blood feeding (Clements, 1956; Detinova, 1945; Gillett, 1956). This factor was initially named egg development neurosecretory hormone (EDNH) because experiments showed it was produced by brain medial secretory cells (Lea, 1967, 1972). Studies later established that EDNH was a protein or small peptide that stimulated ovaries of Aedes aegypti to produce the steroid hormone ecdysone, which was shown to be essential for yolk biosynthesis by the fat body, yolk uptake by primary oocytes and egg maturation (Hagedorn et al., 1975, 1979; Lea, 1972) (see chapter “Regulation of Reproductive Processes in Female Mosquitoes” by Roy et al.). This finding resulted in the renaming of EDNH as ovary ecdysteroidogenic hormone (OEH) (Matsumoto et al., 1989a). However, identification of OEH as a 140 amino acid peptide required many years of effort because large quantities of starting material from mosquito heads were needed to ultimately purify the bioactive peptide and determine its sequence (Brown et al., 1998).
Other studies during this period provided evidence that mosquitoes produce peptide hormones with functions in nutrient storage (Lea and Van Handel, 1970), water balance and diuresis (Maddrell and Phillips, 1978; Nijhout and Carrow, 1978), host seeking (Klowden and Lea, 1979), and blood meal digestion (Graf et al., 1998). Yet the small size of mosquitoes presented similar difficulties to those confronted with OEH. As a result, few of these factors were identified. Instead, most peptide hormones that were successfully purified and identified came from larger insects like certain moths, locusts, and cockroaches (Gäade et al., 1997; Hauser et al., 1997, 2006a,b; Raabe, 1989). In some cases, these sequence data were then used to identify homologous peptide sequences from mosquitoes (Veenstra, 1994, 1999; Veenstra et al., 1997a,b, 1999).
Approaches to studying peptide hormones and their receptors dramatically changed with the sequencing and annotation of the first insect genome from Drosophila melanogaster (Adams et al., 2000). This was soon followed by the sequencing of the mosquito, Anopheles gambiae (Holt et al., 2002), and several other mosquito species including Ae. aegypti and Culex quinquefasciatus (Arensburger et al., 2010; Jiang et al., 2014; Neafsey et al., 2015; Nene et al., 2007; Waterhouse, 2015). These data together with the genomes for several other insects, invertebrates, and vertebrates provided the foundation needed to identify candidate peptide hormone and receptor genes through homology-based searches (Hauser et al., 2010; Hewes and Taghert, 2001; Hummon et al., 2006; Li et al., 2008; Predel et al., 2010; Riehle et al., 2002; Roller et al., 2008; Southey et al., 2008; Veenstra et al., 2012).
2.1. Peptide Hormone Genes
Annotation of the Ae. aegypti and An. gambiae genomes originally identified 43 and 35 peptide hormone genes, respectively (Predel et al., 2010; Riehle et al., 2002). Additions to the literature since these studies were published together with annotation improvements increase this number to 50 for Ae. aegypti, 47 for An. gambiae, and 45 C. quinquefasciatus. We list these in alphabetical order in Table 1 along with their identifiers, which will be useful to readers because some of these genes are still difficult to find or correctly recognize in databases. We also list the 45 genes that have been identified from D. melanogaster, which is of value to this discussion from two perspectives. First, more peptide hormones have been functionally studied in D. melanogaster than in any mosquito species. Second, mosquitoes and D. melanogaster reside in families (Culicidae and Drosophilidae) that arose at different times in the evolutionary history of the order Diptera. The Culicidae is an early lineage that arose 225 million years ago (mya) while the Drosophilidae evolved much more recently (40–65 mya) in concert with other cyclorrhaphan flies (Wiegmann et al., 2003). Thus, the peptide hormone genes from mosquitoes and drosophilids together provide information across the breadth of the order.
Table 1.
Peptide Hormone Genes for Three Mosquito Species and D. melanogastera
| Hormone | Ae. aegypti | An. gambiae | C. quinquefasciatus | D. melanogaster | Mosquito Species with Reported Localization Datab |
|---|---|---|---|---|---|
| Adipokinetic hormone (AKH) | AAEL011996 | AGAP008834 | CPIJ000869 | CG1171 | Aa, Ag |
| Adipokinetic/corazonin peptide (ACP) | AAEL010950 | AGAP002430 | CPIJ001379 | ND | Aa, Ag |
| Agatoxin-like neuropeptide | PA | AGAP007821 | CPIJ000956 | ND | |
| Allatostatin A (ASTA) | AAEL015251 | AGAP003712 | CPIJ008017 | CG13633 | Aa |
| Allatostatin C (ASTC) | AAEL005747 | AGAP010157 | partial unan. | CG14919 | Aa |
| Allatotropin | AAEL009541 | AGAP012130 | CPIJ007896 | ND | Aa |
| Apis-ITG-like | AAEL006369 | AGAP008993 | CPIJ005482 | CG8216 | |
| Bursicon | PA | AGAP002537 | CPIJ009600 | CG13419 | Ag |
| Partner of bursicon | AAEL013722 | AGAP004506 | CPIJ012985 | CG15284 | Ag |
| CCHamide 1 | AAGE02019353 | BM585352 | AAWU01008744 | CG14358 | |
| CCHamide 2 | AAEL004890 | AGAP004553 | AAWU01038417 | CG14375 | |
| Corazonin | AAEL005252 | AGAP012665 | ND | CG3302 | Aa |
| Crustacean cardioactive peptide (CCAP) | AAEL000630 | AGAP009729 | CPIJ005842 | CG4910 | Ag |
| Diuretic hormone 31 (DH31) | AAEL008070 | AGAP001382 | ND | CG13094 | Aa |
| Diuretic hormone 44 (DH44) | AAEL008292 | AGAP003269 | CPIJ008822 | CG8348 | Aa |
| Ecdysis triggering hormone (ETH) | AAEL001762 | AGAP007062 | CPIJ004945 | CG18105 | Aa |
| Eclosion hormone | AAEL011229 | AGAP010437 | CPIJ011911 | CG5400 | |
| FMRFamide | AAEL013645 | AGAP005518 | CPIJ000101 | CG2346 | Aa |
| Glycoprotein A2 | BN001241 | AGAP008301 | CPIJ013236 | CG17878 | |
| Glycoprotein B5 | AAEL001474 | AGAP008302 | CPIJ013235 | CG40041 | |
| Insulin-like peptide (ILP) 1 | AAEL000937 | AGAP010605 | CPIJ018051 | CG14173 | |
| ILP2 | AAEL000960 | AY324308 | CPIJ018049 | CG8167 | |
| ILP3 | DQ845751 | AY324309 | CPIJ018050 | CG14167 | Aa, Ag, As |
| ILP4 | AAEL000932 | AY324310 | ND | CG6736 | |
| ILP5 | AAEL003000 | AGAP003927 | CPIJ001698 | CG33273 | |
| ILP6 | DQ845755 | AY324313 | CPIJ003329 | CG14049 | |
| ILP7 | DQ845757 | AY324314 | ND | CG13317 | |
| ILP8 | DQ845754 | ND | ND | CG14059 | |
| Ion transport peptide | AAEL015332 | AGAP005055 | CPIJ003972 | CG13586 | |
| Limostatin 1 | AAEL008355 | AGAP013197 | ND | CG8317 | |
| Limostatin 2 | AAEL008359 | ND | ND | ND | |
| Kinin (leucokinin) | AAEL010172 | AGAP013518 | CPIJ010343 | CG13480 | Aa |
| Myoinhibitory peptide | AAEL012139 | AGAP000833 | CPIJ802231 | CG6456 | Aa |
| Myosuppressin | AAEL007294 | AGAP001474 | CPIJ012769 | CG6440 | |
| Natalisin | AAEL003260 | AGAP005277 | CPIJ001072 | CG34388 | |
| Neuropeptide F | AAEL002733 | AGAP004642 | PA | CG10342 | Aa |
| Neuropeptide-like peptides (NPLPs) | AAEL014708 | AGAP010366 | CPIJ014175 | CG3441 | Aa |
| Orcokinin | AAEL010172 | AGAP012220 | CPIJ010343 | CG13565 | |
| Ovary ecdysteroidogenic hormone (OEH) | AAEL004155 | AGAP000108 | CPIJ010626 | ND | Aa, Ag |
| Pigment dispersing hormone | AAEL001754 | AGAP005776 | CPIJ004895 | CG6496 | |
| Proctolin | ND | ND | ND | CG7105 | |
| Prothoracicotropic hormone (PTTH) | PA | AGAP000859 | CPIJ003196 | CG13687 | |
| Pyrokinin 1 (PK1) | AAEL012060 | AGAP002292 | CPIJ005970 | CG6371 | Aa, As, Cp |
| Pyrokinin 2 (PK2) | AAEL005444 | AGAP000347 | partial unan. | CG15520 | Aa, As, Cp |
| RYamide | AAEL011702 | AGAP006765 | CPIJ008988 | CG40733 | |
| Short neuropeptide F 1 (sNPF1) | AAEL012542 | DQ437578 | CPIJ009049 | CG13968 | Aa |
| Short neuropeptide F 2 (sNPF2) | AF155738.1 | ND | ND | ND | Aa |
| SIFamide | AAEL009858 | AGAP007056 | CPIJ004953 | CG33527 | Aa |
| Sulfakinin | PA | AGAP009275 | CPIJ004208 | CG18090 | Aa |
| Tachykinin | AAEL006644 | AGAP010014 | Partial unan. | CG14734 | Aa, Cs |
| Trissin | AAEL008756 | AGAP012496 | CPIJ016124 | CG14871 |
Gene entries for each species are by their VectorBase, GenBank, or FlyBase identifier. PA: identification of a partial exon in the genome that is unannotated. ND: gene not detected in databases.
Aa, Aedes aegypti; Ag, Anopheles gambiae; As, An stephensi; Cp, Culex pipiens; and Cs, C. salinarius.
2.1.1. Comparative Genomics
Most peptide hormone genes are single copy and produce mature hormones that are monomeric. However, differential processing can yield multiple bioactive forms for some peptide hormones with the most extreme example in mosquitoes being prepro-FMRFamide, which is processed into nine functional FMRFamide isoforms (Predel et al., 2010). Some peptide hormone genes along with their receptors are duplicated in mosquitoes and select other insects. These include adipokinetic hormone (AKH), corazonin, and AKH/corazonin peptide (ACP) (Table 1). Some of these duplications, however, have been lost in drosophilids including D. melanogaster (Hauser and Grimmelikhuijzen, 2014; Vogel et al., 2013). The only multimember peptide hormone genes in mosquitoes, drosophilids, and other insects are the insulin-like peptides (ILPs), but it is unclear all family members derive from a common ancestral gene (Grönke et al., 2010). Lastly, bursicon and glycoprotein A2/B5 are dimeric peptide hormones that are produced from cleavage products of different prepropeptides (bursicon/partner of bursicon and glyprotein hormone A2/A5), whose corresponding genes are also expressed in different neurosecretory cells (Table 1). Trypsin modulating oostatic factor (TMOF) is a proline rich, 10 amino acid peptide identified in Ae. aegypti that is often discussed as a peptidyl regulator of blood meal digestion and egg formation in mosquitoes (Borovsky et al., 1990; Verlinden et al., 2014). However, TMOF is not included in Table 1 or further discussed here because it matches a 10 amino acid sequence of the vitelline envelope protein (Lin et al., 1993), which clearly is not a peptide hormone precursor protein. In addition, no data compellingly supports that TMOF binds a specific receptor including any GPCR, PRK, or RGC, or activates signalling in any target cell that is consistent with it functioning as a peptide hormone.
Overall, the data in Table 1 show Ae. aegypti, An. gambiae, and C. quinquefasciatus encode the same peptide hormone genes with two exceptions. First the short neuropeptide (sNPF) gene in Ae. aegypti has duplicated, which we refer to here as sNPF1 and sNPF2 (see Section 4.1.16), whereas An. gambiae and C. quinquefasciatus have only one sNPF gene. Second, Ae. aegypti has eight ILP genes while An. gambiae has seven and C. quinquefasciatus has five. Table 1 notes a few other instances of genes not being detected in C. quinquefasciatus but this is likely due to annotation issues. D. melanogaster lacks genes for ACP, allatotropin, and OEH but orthologs of all other peptide hormone genes in mosquitoes are present. Orthologs of most peptide hormone genes in mosquitoes and drosophilids are also present in insects in other orders. The proctolin gene is present in D. melanogaster and other insects but is absent in mosquitoes. A few other peptide hormone genes present in one or more insects from other orders are absent from both mosquitoes and drosophilids (Table 1). The latter could indicate these peptide hormones are absent from all of the Diptera.
2.1.2. Transcription and Peptidomics
Transcriptional profiling of individual genes has been reported for several peptide hormones in mosquitoes, but only a few studies report expression data in specific tissues or life stages. In An. gambiae, tissue and/or life stage expression data are available for AKH and ACP (Kaufmann and Brown, 2006, 2008), bursicon (Robertson et al., 2007), ILPs, (Arsic and Guerin, 2008; Krieger et al., 2004), neuropeptide F (NPF) (Garczynski et al., 2005), and sNPF (Garczynski et al., 2007). In Ae. aegypti, such data are available for AKH and ACP (Kaufmann et al., 2009), allatostatin A and C (ASTA and C) (Hernández-Martínez et al., 2005; Li et al., 2006; Veenstra et al., 1997a), allatotropin (Hernández-Martínez et al., 2005; Veenstra and Costes, 1999), ILPs (Brown et al., 2008; Riehle et al., 2006), kinin (Veenstra, 1994), NPF (Stanek et al., 2002), OEH (Brown et al., 1998), and sNPF2 (Stracker et al., 2002). Data are also available for what we name in this paper pyrokinin 2 (Choi et al., 2013) in Ae. aegypti (see Section 4.1.15) and prothoracicotropic hormone (PTTH) in Culex pipiens (Zhang and Denlinger, 2011). Profiling of multiple An. gambiae or Ae. aegypti genes through array based or RNAseq studies provide additional data on peptide hormone gene expression (Akbari et al., 2013; Baker et al., 2011; Matthews et al., 2016). The only expression data available for other mosquito species are for ILP genes in C. pipiens (Sim and Denlinger, 2009) and Anopheles stephensi (Marquez et al., 2011; Pietri et al., 2015), and tachykinins in Culex salinaris (Meola et al., 1998). Insights about prepropeptide processing are currently only available for Ae. aegypti through peptidomic analysis of the central nervous system and select other tissues (Predel et al., 2010; Siju et al., 2013). It is also important to note that the accuracy of the above data depends on the quality of the reference genomes that were available at the time the above studies were conducted.
2.2. Peptide Hormone Receptors
Insights into the known or predicted receptors for mosquito peptide hormones derive from two lines of investigation. The first is experimental data in one or more mosquito species showing that a particular receptor is activated or required for the biological activity of a given peptide hormone. The second is homology-based studies of mosquito receptor genes relative to D. melanogaster, other insects, or other animals where experimental data on orthologs are available. While several studies discuss GPCRs that bind peptide hormones (see later), Vogel et al. (2013) provides the only comprehensive summary of the receptors in mosquitoes and drosophilids that are known or predicted to bind peptide hormones. Key features of these receptors are summarized below.
2.2.1. GPCRs
GPCRs form a very large protein family that is present in all multicellular animals. However, the first studies showing that peptide hormones are often ligands for GPCRs derive from experiments first conducted in mammals. GPCRs range from 40 to 60 kDa in size and reside as monomers in the plasma membrane of cells. All exhibit a standard topology which consists of three regions: a variable extracellular N-terminal domain, a conserved transmembrane domain with seven hydrophobic α-helices that functions as a ligand binding pocket, and an intracellular C-terminal domain that mediates signalling through interactions with G proteins (Bockhaert and Pin, 1999). GPCRs are further subdivided into six classes (A–F) of which only Class A (rhodopsin-like) and Class B (secretin-like) contain members that bind peptide hormones (Attwood and Findlay, 1994; Hauser et al., 2006a,b).
The conserved transmembrane domain shared by GPCRs was used during annotation to interrogate the D. melanogaster genome. This led to identification of more than 200 predicted GPCR genes of which 44 were Class A and B members (Brody and Cravchik, 2000; Hewes and Taghert, 2001). A few functional studies of GPCRs preceded sequencing of the D. melanogaster genome but most of these predicted GPCRs were ‘orphans’ that were subsequently targeted for study by many groups using different experimental approaches. This resulted in ‘deorphanization’ and identification of GPCRs that bind several of the peptide hormones listed in Table 1 (see Johnson et al., 2003; Näassel and Winther, 2010). The same approach was used to annotate the GPCR genes of An. gambiae, Ae. aegypti, and C. quinquefasciatus (Arensburger et al., 2010; Hill et al., 2002; Nene et al., 2007). However, experimental studies identifying mosquito GPCRs that bind peptide hormones are currently restricted in Ae. aegypti to ASTC (Mayoral et al., 2010), allatotropin (Hayes et al., 1997; Nouzova et al., 2012), kinins (Kersch and Pietrantonio, 2011; Kwon et al., 2012; Pietrantonio et al., 2005; Taneja-Bageshwar et al., 2009), NPF (Liesch et al., 2013), pyrokinin 2 (Choi et al., 2013), and sNPFs (Liesch et al., 2013). In anopheline mosquitoes, GPCRs that bind AKH, crustacean cardioactive peptide (CCAP), and corazonin (Belmont et al., 2006), ACP (Hansen et al., 2010), myosuppressin (Scholler et al., 2005), NPF (Garczynski et al., 2005), and sNPFs (Garczynski et al., 2007) have been identified in An. gambiae while the GPCR receptor for a kinin has been identified from A. stephensi (Radford et al., 2004).
The likely GPCRs for other peptide hormones in mosquitoes were comparatively assessed by Vogel et al. (2013) using all of the Class A (245) and B (82) genes from Ae. aegypti, An. gambiae, C. quinquefasciatus, D. melanogaster, and D. mojavensis. This produced phylogenies comprised of well-supported monophyletic clades that were subdivided into assemblages and subassemblages. A majority of clades contained receptors whose peptide hormone ligands are known from experimental studies in D. melanogaster or other species, which in turn provided evidence for the likely ligand of corresponding orthologs in mosquitoes. These ‘characterized’ GPCR clades and their known or predicted peptide hormone ligands are listed in Table 2 using the nomenclature of Vogel et al. (2013). Data on where different receptors are expressed in mosquitoes and drosophilids are difficult to summarize because of inconsistencies between studies in regard to the life stages and tissues that were examined. We therefore discuss such expression data, if available in mosquitoes, in Section 4.1.
Table 2.
Peptide Hormone GPCR Genes for Three Mosquito Species and D. melanogastera
| GPCR Class | Hormone | Ae. aegypti | An. gambiae | C. quinquefasciatus | D. melanogaster |
|---|---|---|---|---|---|
| A, Cluster 1 | |||||
| 1a | Bursicon | AAEL004777 | AGAP008347 | CPIJ011619 | CG8930 |
| 1a | Glycoprotein A2/B5 | AAEL004399 | AGAP004035 | CPIJ007712 | CG7665 |
| 1a | Drosophila ILP8 | CG31096 | |||
| 1b | CCAP R1 | AAEL008652 | AGAP001961 | CPIJ006268 | |
| 1b | CCAP R2 | AAEL008655 | AGAP001962 | CPIJ006269 | CG33344 |
| 1b | Corazonin | AAEL011475 | AGAP001558 | CPIJ016679 CPIJ016678 |
CG10698 |
| 1b | ACP | AAEL009673 | AGAP001532 | CPIJ001199 | |
| 1b | AKH | AAEL011325 | AGAP002156 | CG11325 | |
| 1b | Allatotropin | AAEL005310 AAEL011680 |
CPIJ014752 | ||
| 2a | Sulfakinin R1 | AAEL010207 | AGAP001022 | CPIJ005574 | CG32540 |
| 2a | Sulfakinin R2 | CPIJ016281 CPIJ016282 |
CG42301 | ||
| 2b | RYamide R1 | AAEL008296 | AGAP000351 | CPIJ019394 | CG5811 |
| 2b | RYamide R2 | AAEL015418 | AGAP000115 | CPIJ018504 | |
| 2b | Tachykinin | AGAP002824 | CPIJ014103 CPIJ007277 |
CG6515 | |
| 2b | Natalisin | AAEL006947 | AGAP001592 | CG7887 | |
| 2b | Kinin | AAEL011026 AAEL006636 |
AGAP010851 | CPIJ012071 | CG10626 |
| 2b | NPF | AAEL010626 NPYR2 |
AGAP004122-3 | CPIJ006984 CPIJ018265 |
CG1147 |
| 2b | sNPF | AAEL013505 AAEL007924 |
AGAP012378 | CPIJ13069 | CG7395 |
| 2b | SIFamide | AAEL009698 AAEL005994 |
AGAP003335 | CPIJ017622 CPIJ016970 |
CG10823 |
| 2d | CCH amide R1 | AAEL016997 | AGAP011452 | CPIJ017639 | CG30106 |
| 2d | CCH amide R2 | AAEL017410 | AGAP003631 | CG14593 | |
| 2d | ASTC R1 | AAEL012920 | AGAP010486 | CPIJ011191 | CG7285 |
| 2d | ASTC R2 | AAEL012356 | AGAP012268 | CPIJ010469 | CG13702 |
| 2d | ASTA R1 | AAEL007169 | AGAP003658 | CPIJ016163 | CG2872 |
| 2d | ASTA R2 | AAEL006076-7 | AGAP001773-4 | CPIJ013095 CPIJ011118 |
CG10001 |
| 2e | Trissin | AAEL004732 | AGAP008702 | CPIJ001962-3 | CG34381 |
| 2f | PK R1 | AAEL017335 AAEL003747 |
AGAP003076 | CPIJ011105-07 | CG8784 CG8795D |
| 2f | PK R2 | AAEL012796 KC155994 |
AGAP000658 | CPIJ014065 CPIJ019566 CPIJ015545 |
CG9918 |
| 2f | ETH R1 | AAEL005803 | AGAP002881 | CPIJ003421 | CG5911 |
| 2f | ETH R2 | AAEL005804 | CPIJ003422 | ||
| A, Cluster 2 | |||||
| FMRFamide | AAEL015604 AAEL003378 |
AGAP001862 | CPIJ007187 | CG2114 | |
| MIP | AAEL010313 | AGAP012427 | CPIJ007973 | CG16752 | |
| Myosuppressin | AAEL006283 | AGAP005229 | CPIJ000143 | CG43745 CG8985 |
|
| B | |||||
| Pigment dispersing hormone | AAEL009024 | AGAP003654 | CPIJ009749 | CG13758 | |
| DH44 R1 | AAEL008292 | AGAP005464 | CPIJ008822 | CG12370 | |
| DH44 R2 | AAEL005894 | AGAP005465 | CPIJ008820 | CG8422 | |
| DH33 | AAEL010043 | AGAP009770 | CPIJ014419 | CG32843 | |
Gene entries for each species as in Table 1. Receptor classification per Vogel et al. (2013). The Class A GPCR phylogeny forms two clusters (1 and 2). Cluster 1 is further divided into subassemblages 1a, b, and 2a—f.
Vogel et al. (2013) also identified several GPCR ‘orphan’ clades. In some cases placement in the phylogeny provides insights into predicted ligands. For example, subassemblage 1a of the Class A GPCRs contains two orphan clades (OA1 and OA2) that are sister to the bursicon and glycoprotein A2/B5 receptors. This makes them strong candidates for binding relaxin/ILPs such as ILP6 in Ae. aegypti and ILP8 in D. melanogaster (Brown et al., 2008). Notably, this prediction was recently supported experimentally for D. melanogaster ILP8, which deorphanizes OA1 (Garelli et al., 2015; Vallejo et al., 2015). Other orphan clades provide no insights about their ligands but their relationship to other GPCRs in the phylogeny strongly suggest they bind peptide hormones. If correct, this suggests that mosquitoes and drosophilids produce several peptide hormones that have not been identified. Lastly, these data show that dipteran Class A and B GPCRs have experienced several instances of duplication or loss in particular species, which results in some characterized clades containing ‘in group’ orphans (Vogel et al., 2013). The functional significance of this is unclear although duplicated receptors may preferentially bind different isoforms of a given peptide hormone since differential processing can produce variants of the same ligand (Predel et al., 2010). Recent studies also provide some support for this suggestion (Liesch et al., 2013).
2.2.2. PKRs
Most PKRs are single subunit receptors that have an extracellular ligand binding domain, a single hydrophobic transmembrane-spanning domain, and an intracellular kinase domain that upon ligand binding autophosphorylates either tyrosine or serine/threonine residues (Hubbard and Miller, 2007; Lemmon and Schlessinger, 2010). Most PKRs form noncovalent dimers upon entry into the plasma membrane or ligand binding, although some, like the insulin receptor (IR) (see later), form a covalent heterotetramer through intra- and intersubunit disulphide bonds. PKRs with tyrosine phosphorylation activity are referred to as receptor tyrosine kinases (RTKs) which can activate one or more signalling pathways. PKRs with serine/threonine kinase domains are also capable of activating one or more signalling pathways.
PKR ligands in mammals include insulin, several growth factors, and certain cell surface molecules (Lemmon and Schlessinger, 2010). A homologue of the vertebrate IR, was cloned from D. melanogaster prior to sequencing of its genome, which was named the Drosophila IR (Fernandez-Almonacid and Rosen, 1987). Biochemical studies showed this receptor binds vertebrate insulin (Fernandez-Almonacid and Rosen, 1987; Sajid et al., 2011) while genetic studies provided evidence that several endogenous ILPs activate the insulin signalling pathway (Brogiolo et al., 2001). Genetic studies in D. melanogaster also identified an RTK as the receptor for PTTH (Rewitz et al., 2009). In mosquitoes, experiments conducted in Ae. aegypti show that its IR binds endogenous ILP3 with high affinity which activates the canonical insulin signalling pathway (Brown et al., 2008; Dhara et al., 2013; Wen et al., 2010). In contrast, no functional studies have been conducted in mosquitoes on the PTTH receptor (see Section 4.1).
Results from Vogel et al. (2013) assemble the mosquito and drosophilid PKR genes into two branches: 12 well-supported clades consisting of RTKs and 5 clades containing PKRs with conserved serine–threonine kinase domains that are related to mammalian transforming growth factor beta (TGF-β) receptors. Unlike GPCRs, this analysis also shows that dipteran PKRs have undergone few lineage-specific gains or losses, which suggest the same genes are likely present in most or all Diptera. Mosquito and drosophilid IRs form a single clade that is closely related to a clade that contains homologues of human anaplastic lymphoma RTK (ALK) and leucocyte RTK plus a second orphan clade (OR1) lost from D. melanogaster. The ALK-like receptor has been shown to have functional roles in development, ethanol sensitivity, and learning in D. melaogaster but its ligand is unknown (Gouzi et al., 2011; Lasek et al., 2011; Lorén et al., 2001). The OR1 receptor from Ae. aegypti was recently deorphanized by showing that it binds OEH (Vogel et al., 2015; see Section 4.1). Many of the remaining clades whose ligands are growth factors have been studied in D. melanogaster but have not been examined in mosquitoes. The RTK genes with known or predicted peptide hormone ligands in mosquitoes and D. melanogaster are summarized in Table 3.
Table 3.
Peptide Hormone RTK and RGC Genes for Three Mosquito Species and D. melanogastera
| Receptor | Hormone | Ae. aegypti | An. gambiae | C. quinquefasciatus | D. melanogaster |
|---|---|---|---|---|---|
| RTK | ILPs | AAEL002317 | AGAP012424 | CPIJ006878 | CG18402 |
| RTK | ILPs | AAEL002317 | AGAP012424 | CPIJ006878 | CG18402 |
| RTK | PTTH | AAEL002404 AAEL010923 |
AGAP005763 | CPIJ001844 CPIJ000058 |
CG1389 |
| RGC | NPLPs | AAEL008390 | AGAP012163 | CPIJ010083 | CG42636 CG42637 |
| RGC | EH | AAEL008387 | AGAP012161 | CPIJ010082 | CG10738 |
Gene entries for each species as in Table 1.
2.2.3. RGCs
RGCs are conserved homodimeric membrane proteins which have an extracellular ligand binding domain, a single pass transmembrane domain, and an intracellular catalytic domain with adenylyl or guanylyl cyclase activities that are sensitive to intracellular calcium levels (Potter, 2011). RGCs also usually form dimers prior to or after ligand binding. Studies in mammals were the first to show that RGCs bind peptide hormones called brain and atrial natriuretic peptides (Potter, 2011). In insects, experiments with the oriental fruitfly, Bactrocera dorsalis, identified an RGC as the receptor for eclosion hormone (EH) (Chang et al., 2009), while a second RGC was identified as the receptor for neuropeptide-like peptide 1 (NPLP1) in D. melanogaster (Overend et al., 2012). Phylogenetic analysis of mosquito and drosophilid RGCs generated one clade containing predicted EH receptors, a second containing NPLP receptors, and four orphan clades (OGC1–4) (Vogel et al., 2013) (Table 3). One of these orphan clades appears orthologous to the vertebrate RGCs that bind natriuretic peptides, while a second appears orthologous to vertebrate retinal RGCs. The other two orphan clades have no vertebrate ortholog.
3. SOURCES, PROCESSING, AND RELEASE OF PEPTIDE HORMONES IN MOSQUITOES
In mosquitoes, most insights into where different peptide hormones are produced derive from studies of adult females. This bias mirrors the literature at-large where most of what we know about mosquito genetics, biochemistry, physiology, and behaviour also come from studies of adults, which is the life stage of interest from the perspective of vector biology and disease transmission. A different situation, however, exists in a number of other insects including D. melanogaster where the source of many peptide hormones have been carefully mapped in larvae but distribution in adults is less detailed (Näassel and Winther, 2010). Studies on peptide hormone production and function in Lepidoptera also overall focus on larvae because of the historic interest in these insects as models for the study of moulting (Nijhout, 1998; Smith and Rybczynski, 2012; Truman and Riddiford, 2002). Thus, while most peptide hormone genes in mosquitoes are conserved with other insects, differences in functional priorities prevent some comparisons from being made in regard to where they are produced. The emphasis on adults has also affected which peptide hormones have been most studied in mosquitoes.
3.1. Peptide Hormone Producing Cells
Immunocytochemistry and peptidomics provide information on cells in mosquitoes where different peptide hormones have been detected. We list the species for which such data are available in Table 1 and illustrate these sites in Fig. 1. For the central nervous system several hormones are detected in the brain, specific neurosecretory cells, and ganglia of the ventral nerve chord (Brown and Cao, 2001; Brown and Lea, 1988; Brown et al., 2008; Cao and Brown, 2001; Estévez-Lao et al., 2013; Hellmich et al., 2014; Hernández-Martínez et al., 2005; Kaufmann and Brown, 2006; Kaufmann et al., 2009; Krieger et al., 2004; Marquez et al., 2011; Meola and Lea, 1972; Meola et al., 1998; Moffett and Moffett, 2005; Predel et al., 2010; Riehle et al., 2006; Siju et al., 2013; Stracker et al., 2002). The corpus cardiacum (CC) and perivisceral organs iteratively present in abdominal segments are neurohemal organs that store and release several different peptide hormones in mosquitoes (Brown and Cao, 2001; Brown and Lea, 1988; Cao and Brown, 2001; Estévez-Lao et al., 2013; Hernández-Martínez et al., 2005; Honegger et al., 2011; Kaufmann and Brown, 2006; Kaufmann et al., 2009; Marquez et al., 2011; Predel et al., 2010; Riehle et al., 2006; Veenstra and Costes, 1999). Studies conducted primarily in D. melanogaster show that the midgut epithelium contains intestinal stem cells, which self-renew and differentiate into two other cell types: enterocytes, that produce digestive enzymes and mediate nutrient uptake, and endocrine cells (Jiang and Edgar, 2012). In Ae. aegypti, midgut endocrine cells are known to produce and/or release several peptide hormones (Brown et al., 1986; Hernández-Martínez et al., 2005; Marquez et al., 2011; Moffett and Moffett, 2005; Predel et al., 2010; Stracker et al., 2002; Veenstra et al., 1995) (Table 1, Fig. 1).
Fig. 1.
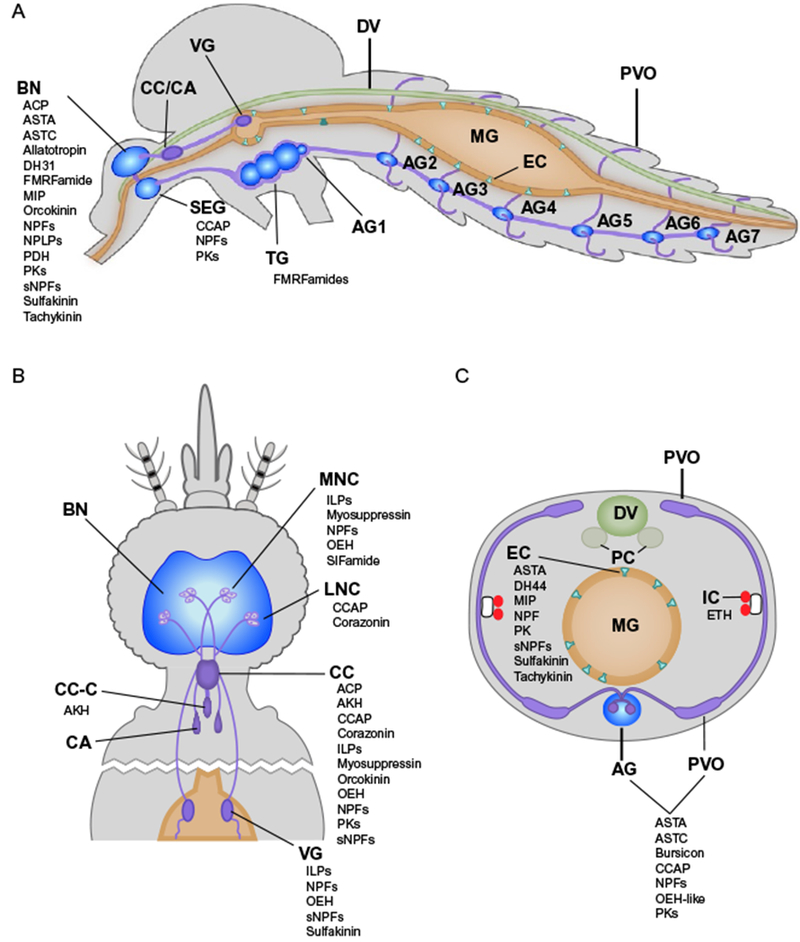
Schematic illustrating the sources of different peptide hormones in adult female mosquitoes as determined by immunocytochemistry or peptidomics. Sagittal view of the whole body (A), dorsal view of the head and thoracic regions (B), and a cross section through the third abdominal segment (C). Select regions of the nervous system, organs and cells are labelled in bold. Peptide hormones in Table 1 are listed alphabetically below detected sites. Abbreviations: AG1–7, abdominal ganglia 1 through 7; BN, brain; CA, corpora allata; CC, corpus cardiacum; CC-C, corpus cardiacum intrinsic cells; DV, dorsal vessel; EC, gut endocrine cells; IC, Inka cells; LNC, lateral neurosecretory cells; MG, midgut; MNC, medial neurosecretory cells; PC, pericardia; PVO, perivisceral organs; SEG, subesophegial ganglion; VG, ventricular ganglion. Peptide hormone abbreviations are listed in Table 1. Most of these data derive from studies of adults with the exception being for ETH, which is shown in (C) although data derive from studies of larvae (see text).
3.2. Processing Enzymes and Hormone Secretion
Microscopy studies show that peptide hormones accumulate in the body of endocrine and neurosecretory cells where these are packaged into secretory vesicles as propeptides along with processing enzymes (Brown and Lea, 1988; Raabe, 1989). Studies of mammalian peptidergic cells indicate these include endopeptidases, which are primarily convertases that process propeptides, and other modifying enzymes that can alter the amino- and carboxyl termini of processed peptides in ways that are important for function (Hook et al., 2008; Isaac et al., 2009). Amidation of carboxyl termini also confers resistance to peptidases, which potentially extends half-life after release into circulation. Functional studies of convertases and the enzymes responsible for carboxy-terminal amidation have been conducted in D. melanogaster (Jiang et al., 2000; Reiher et al., 2011; Taghert and Veenstra, 2003; Wegener et al., 2011). Genes encoding homologous processing enzymes have been identified in mosquitoes but no functional studies have been conducted (Akbari et al., 2013; Matthews et al., 2016; Riehle et al., 2002).
Peptide hormones are released from endocrine and neurosecretory cells in response to external and internal stimuli. For neurosecretory cells in the brain, this release occurs primarily from axons extending to the CC and along the gut (Brown and Cao, 2001; Cao and Brown, 2001; Raabe, 1989). In contrast, neurosecretory cells located in other ganglia or tissues release peptide hormones in several other locations throughout the body (Raabe, 1989). Midgut endocrine cells release peptides into the space between epithelial cells for diffusion to neighbouring cells and the hemolymph (Brown et al., 1986, 1988). Several enzymes that degrade peptide hormones in circulation or after binding to target cells have been identified in mammals (Isaac et al., 2009). While homologues of these enzymes are known in both mosquitoes and D. melanogaster (Riehle et al., 2002; Taghert and Veenstra, 2003), little information is available regarding their expression or function. The only exception to this is in the context of data showing that some kinin agonists are resistant to proteolysis in mosquitoes, which suggests they could be useful as control agents (Taneja-Bageshwar et al., 2009).
Some peptide hormones are also likely degradated by endosomal enzymes after receptor binding and internalization. One example of this is insulin degrading enzymes (IDE, insulinases) that are conserved between mammals and insects including An. gambiae and D. melanogaster (Duckworth et al., 1989; Riehle et al., 2002). A recent study showed that an IDE is globally expressed in D. melanogaster throughout development, is present in both the membrane and cytoplasm of cells, and affects ILP levels and signalling (Galagovsky et al., 2014). Studies from mammals also suggest IDEs may have important roles in intracellular degradation of other peptides (Fernández-Gamba et al., 2009).
4. PEPTIDE HORMONE FUNCTIONS IN MOSQUITOES
Many studies have been conducted on the functions of peptide hormones in insects, but this literature is also large and difficult to summarize across species that have been studied. As a result, most reviews focus on either: (1) processes that peptide hormones have roles in regulating like metabolism (Baker and Thummel, 2007; Broughton and Partridge, 2009), moulting and metamorphosis (Smith and Rybczynski, 2012; White and Ewer, 2014), and water balance (Dow and Davies, 2006) or (2) particular types of peptide hormones such as FMRFamides (Nichols, 2003), ILPs (Antonova et al., 2012), or PRXamides (Jurenka, 2015). A few recent summaries discuss peptide hormone functions more broadly but they primarily emphasize nonmosquito species, which results in a number of studies from the mosquito literature being uncited (Hauser et al., 2008; Näassel and Winther, 2010; Verlinden et al., 2014). Our own assessment identifies functional data in one or more mosquito species for about a third of the entries in Table 1. Below, we first summarize these findings in alphabetical order with the absence of a given peptide from Table 1 indicating that no functional studies to our knowledge have been conducted in mosquitoes. We also provide a succinct summary of comparative information, so that the reader can put into firmer context findings from mosquitoes. We then follow this information with some concluding remarks.
4.1. Peptide Hormones with Reported Activities
4.1.1. Adipokinetic Hormone (AKH) and AKH/Corazonin Peptide (ACP)
Most insects including D. melanogaster have one AKH gene that is processed into an N-terminally pyroglutamate blocked peptide of 8–10 amino acids (Gäade, 2009). Functional studies outside of mosquitoes report that AKHs mobilize stored lipid and/or carbohydrate from the fat body into the hemolymph; making this hormone a functional analogue of mammalian glucagon (Van der Host, 2003). Studies from D. melanogaster focus primarily on AKH function in concert with ILPs in regulating metabolism (Baker and Thummel, 2007; Näassel and Winther, 2010) although one study also reports that AKH is cardiostimulatory (Noyes et al., 1995). As noted earlier, mosquitoes have AKH and ACP genes, which give rise to two peptides that bind class A GPCRs in subassemblage 1b. These receptors are also related to the receptors for CCAP and allatotropin (Belmont et al., 2006; Caers et al., 2012; Mugumbate et al., 2011, 2013; Vogel et al., 2013). Functional experiments indicate that AKH mobilizes stored carbohydrate but not lipid in An. gambiae while ACP has no effect on either lipid or carbohydrate mobilization (Kaufmann and Brown, 2008).
4.1.2. Allatostatin A and C (ASTA and C)
ASTs are structurally variable peptides from distinctly different genes (Veenstra, 2008). The first AST was identified from the cockroach Diploptera punctata as an allatostatin (Woodhead et al., 1989). However, studies report activities for ASTs in other insects that do not include the inhibition of juvenile hormone (JH). Thus, the name AST for these peptides is largely historical. We thus include allatostatin B (ASTB) with the myoinhibitory peptides (MIP) on the basis of sequence similarity (Table 1). In mosquitoes and drosophilids, ASTA and C bind related class A GPCRs that cluster together into subassemblage 2c along with the receptor for CCHamide (Mayoral et al., 2010; Vogel et al., 2013). ASTA and C have both been shown to regulate JH biosynthesis in Ae. aegypti (Li et al., 2004, 2006), while one report suggests a role for ASTs in gut motility (Onken et al., 2004). The role of ASTs in regulation of JH is comprehensively discussed in chapter “The Role of Juvenile Hormone in Mosquito Development and Reproduction” by Zhu and Noriega.
4.1.3. Allatotropin
Allatotropins are 14 amino acid peptides that stimulate JH biosynthesis in insects from several orders. As previously noted, however, drosophilids lack orthologous genes for both allatotropin and its receptor (Vogel et al., 2013). Functional studies identify the Class A, subassemblage 1a GPCR that binds allatotropin in Ae. aegypti while also demonstrating that allatotropin stimulates JH biosynthesis (Hernández-Martínez et al., 2007; Li et al., 2003; Nouzova et al., 2012). These results are further discussed in chapter “The Role of Juvenile Hormone in Mosquito Development and Reproduction” by Zhu and Noriega. Vogel et al. (2013) identified a closely related paralogue of the allatotropin receptor in Ae. aegypti but its ligand is unknown.
4.1.4. Bursicon and Partner of Bursicon
Bursicon has long been known in the literature as a factor required for tanning of insect cuticle after moulting (Nijhout, 1998). However, it was only recently identified in D. melanogaster as a 30 kDa heterodimer that is produced from products of the bursicon and partner of bursicon genes (Luo et al., 2005; Mendive et al., 2005). The Class A GPCR that binds bursicon in D. melanogaster clusters in subassemblage 1a with single orthologs in all mosquitoes (Vogel et al., 2013). This subassemblage also contains receptors that bind biogenic amines, steroid hormones, and certain large peptide ligands. As such, subassemblage 1a receptors are referred to as derived-peptide GPCRs (Vogel et al., 2013). The adult bias of the literature largely underlies the paucity of data on bursicon function in mosquitoes although one study does show that bursicon-producing neurosecretory cells undergo apoptosis in An. gambiae after adult ecdysis (Honegger et al., 2011).
4.1.5. Corazonin
An 11 amino acid peptide structurally similar to AKHs was isolated and named corazonin based on its activation of heart contractions in the cockroach Periplaneta americana (Veenstra, 1989). Characterization of the corazonin gene in mosquitoes and other insects revealed this similarity extended to their prepropeptides and led to the identification of a gene encoding another peptide named ACP (Hansen et al., 2010). These prepropeptides show structural similarity to the gonadotropin-releasing hormones (GnRH) in mammals. AKH, corazonin, and ACP bind different but closely related Class A GPCRs in An. gambiae that belong to subassemblage 1b (Belmont et al., 2006; Hansen et al., 2010; Vogel et al., 2013). These GPCRs also share homology with the GnRH GPCRs (Hauser and Grimmelikhuijzen, 2014). The function and expression of ACP and its receptor are unknown in insects, whereas corazonin has a range of reported activities including being cardioactive (Hauser and Grimmelikhuijzen, 2014). However, studies in An. gambiae indicate no significant effect on dorsal vessel contraction after knockdown of this factor and its receptor by RNA interference (RNAi) (Hillyer et al., 2012).
4.1.6. Crustacean Cardioactive Peptide (CCAP)
CCAPs are cyclic, amidated nine amino acid peptides that were first identified from the crab Carcinus maenas but are widely conserved regulators of dorsal vessel contraction in insects and other arthropods (Stangier et al., 1999; White and Ewer, 2014). CCAP has also been implicated in regulating ecdysis (White and Ewer, 2014). The Class A, subassemblage 1b GPCR that binds CCAP was first identified in D. melanogaster and later in An. gambiae (Belmont et al., 2006). Expression studies in adult An. gambiae establish that CCAP localizes to brain neurosecretory cells with axons to the CC, and cells in abdominal ganglia with axons to the heart (Estévez-Lao et al., 2013) (Fig. 1). CCAP injected into adult female An. gambiae stimulates heart contractions, whereas RNAi knockdown of CCAP had the opposite effect (Chen and Hillyer, 2013; Estévez-Lao et al., 2013). Both water and sucrose deprivation reduce heart contraction rate but these effects occur independently of CCAP activity (Ellison et al., 2015).
4.1.7. Diuretic Hormone 31 (DH31, Calcitonin-Like)
This hormone was first identified from the cockroach Di. punctata as a 31 amino acid peptide with a C-terminal GXPamide that shares structural features and diuretic activity with vertebrate calcitonins (Furuya et al., 2000). DH31 homologues are now known from several other insects (Zandawala, 2012). Early studies showing that a peptide hormone regulates diuresis in mosquitoes after blood feeding (see earlier) was identified as a DH31 homologue in An. gambiae (Coast et al., 2005). Identification of the DH31 receptor in D. melanogaster as a Class B GPCR was followed by identification of its ortholog in Ae. aegypti, which is expressed as a gradient along the Malpighian tubules (Kwon et al., 2012). Expression in the hindgut further led to the discovery that DH31 exhibits myotropic activity (Kwon and Pietrantonio, 2013). These data are further elaborated upon in chapter “Renal Excretory Processes in Mosquitoes” by Piermarini.
4.1.8. Diuretic Hormone 44 (Corticotropin-Releasing Factor-Like)
A 41 amino acid peptide with diuretic activity that was initially identified from the moth Manduca sexta with its receptor (Kataoka et al., 1989; Reagan, 1994). DH44 sequences exhibit similarities to peptide hormones in the vertebrate corticotropin-releasing factor (CRF) family that bind Class B GPCRs (Balment and Lovejoy, 1999; Cardoso et al., 2014). A DH44 homologue was isolated from whole body extracts of Culex salinarius that increased cAMP levels in M. sexta Malpighian tubules and stimulated diuresis in Ae. aegypti tubules (Clark et al., 1998). Subsequently, two Class B GPCRs that bind DH44 were identified from D. melanogaster while one was identified from Ae. aegypti (Hector et al., 2009; Jagge and Pietrantonio, 2008). More information on DH44 is provided in chapter “Renal Excretory Processes in Mosquitoes” by Piermarini.
4.1.9. Ecdysis Triggering Hormone (ETH)
Studies conducted in M. sexta identified ETH as a 26 amino acid peptide that ended at its C-terminus with the sequence PRMamide (Zitnan et al., 1996). ETH genes encoding usually two ETH peptides have since been identified from insects in most orders including Diptera (Jurenka, 2015; Zitnan et al., 2003). As noted earlier, ETH peptides are conservatively produced in Inka (=epitracheal) cells (Fig. 1). ETH genes and the mature peptides they encode exhibit distinct features. However, the C-terminal PRXamide motif of ETHs is shared with another group of peptide hormones we refer to below as pyrokinins (see Section 4.1.15). ETHs also bind Class A GPCRs that are closely related to the pyrokinin receptors (Vogel et al., 2013). In D. melanogaster the ETH receptor gene is alternatively spliced to produce receptors that preferentially bind the two mature ETHs that are processed from prepro-ETH (Iversen et al., 2002; Park et al., 2002). Mosquito ETH genes also produce two ETH isoforms but no studies have examined receptor binding. Functional data support a conserved role for ETH in triggering ecdysis in Ae. aegypti during larval and pupal moulting (Dai and Adams, 2009). A recent study also implicates ETH in the timing of JH synthesis (Areiza et al., 2014).
4.1.10. FMRFamides
These neuropeptides are 7–10 amino acids in length, share C-terminal RFamide sequences, and have broadly conserved myotropic functions. The literature initially treated peptides ending in RF/Pamide including insect FMRFamides, sulfakinins, myosuppressins, NPF, and sNPF as related ‘FMRFamide’ or ‘FaRPamide’ peptides. These peptides also all bind Class A GPCRs. Subsequent studies, however, showed that FMRFamides, sulfakinins, myosuppressins, NPFs, and sNPFs derive from distinctly different genes, which suggests their similar C-termini reflects evolutionary convergence driven by currently unknown factors. The receptors for mosquito and drosophilid FMRFamides all reside in a single cluster 2 clade (Vogel et al., 2013). The myotropic action of FMRFamides on the heart and gut of D. melanogaster as well as their expression in the nervous system are well studied (Nichols, 2003). The first functional studies in mosquitoes used an FMRFamide antiserum to identify FMRFamide cells that were immunopositive in the nervous system and midgut of Ae. aegypti (Brown and Lea, 1988; Brown et al., 1986; Moffett and Moffett, 2005). These data though provide unclear results regarding which of these cells actually produce and/or store FMRFamides because the antisera used recognizes only the RFamide epitope. Studies in An. gambiae show that a single dose of FMRFamide stimulates larval heart contractions (Duttlinger et al., 2003). Hillyer et al. (2014) profiled FMRFamide gene expression in An. gambiae and predicted prepro-FMRFamide produces eight FMRFamide peptides, which is consistent with previously discussed peptidomic data showing that prepro-FMRFamide from Ae. aegypti produces nine peptides (Predel et al., 2010). Functional experiments indicate that two FMRFamide peptides increase heart contractions in An. gambiae at low doses but reduce contractions at high doses (Hillyer et al., 2014). A direction for study that would aid in interpreting the physiological importance of FMRFamides in mosquito physiology would be to clearly identify the cell sources for these peptides and release dynamics.
4.1.11. Insulin-Like Peptides (ILPs)
All ILPs are 6–8 kDa, share a common structural motif called the insulin fold, and are processed from precursors with similar domain structure (Pre, B, C, A) (De Meyts et al., 2009). ILPs are also broadly conserved among metazoans and are the most studied peptide hormones because of their important regulatory roles in metabolism, growth, and development. In mammals, insulin is the only ILP that binds with high affinity to the IR; an RTK, which activates the insulin signalling pathway (De Meyts et al., 2009). Mammals also produce two other types of ILPs: (1) insulin-like growth factors (IGFs) that preferentially bind another RTK, the IGF receptor (IGFR), and activate MAPK signalling and (2) relaxins that bind GPCRs and activate several signalling pathways (Bani, 1997; McDonald et al., 1989). Insulin primarily regulates metabolic responses, IGFs primarily have functions in growth, while relaxins are implicated in several activities.
This background is important because mosquitoes and other insects produce multiple ILPs but usually have only one IR/IGFR homologue which as previously noted is named the IR. Several but not all ILP family members are functionally redundant in D. melanogaster but it is currently unknown whether all ILP family members are capable of binding and activating the IR (Grönke et al., 2010; Zhang et al., 2009). That ILP8 requires a GPCR for function suggests it is relaxin-like and may not interact with the IR (Garelli et al., 2015; see earlier). Factors released from the fat body plus the peptide hormone CCHamide2 from gut endocrine cells are both implicated in regulating ILP release from brain medial neurosecretory cells in D. melanogaster (Sano et al., 2015).
Studies in mosquitoes have mostly been conducted in Ae. aegypti where five ILP genes (1, 3, 4, 7, and 8) are expressed in brain medial neurosecretory cells while three (2, 5, and 6) are detected in other tissues including the midgut and fat body (Riehle et al., 2006). Each prepro-ILP exhibits features consistent with processing like vertebrate insulin/relaxins except ILP6, which has features that suggest it is processed like a vertebrate IGF (Brown et al., 2008; Riehle et al., 2006). Most functional data focus on ILP3 because it is most similar to vertebrate insulin, binds with high affinity to the Ae. aegypti IR, and activates the insulin signalling pathway (Brown et al., 2008; Dhara et al., 2013; Wen et al., 2010).
ILP3 exhibits vertebrate insulin-like activity by reducing blood sugar (trehalose) in adult female Ae. aegypti while elevating carbohydrate and lipid stores in the fat body (Brown et al., 2008). ILP3 also has several functions in female reproduction. As noted earlier, the mosquito endocrinology literature began with characterization of EDNH, which was renamed OEH. In the process of identifying OEH, data suggested factors smaller than OEH were released from the brain after blood feeding that had similar activity (Brown et al., 1998; Matsumoto et al., 1989a). Subsequent experiments demonstrated this factor was ILP3 which is released with OEH from medial neurosecretory cells (Fig. 1). The factors that regulate ILP and OEH release after blood feeding are unknown but data strongly support that ILP3 directly stimulates ovary follicle cells to produce ecdysone by binding the IR (Brown et al., 2008; Riehle et al., 2002; Wen et al., 2010). ILP3 also stimulates late-phase trypsin expression by the midgut, which is required for blood meal digestion, and proliferation of hemocytes (Castillo et al., 2011; Gulia-Nuss et al., 2011). Other factors involved in regulation of egg formation are discussed in chapter “Regulation of Reproductive Processes in Female Mosquitoes” by Roy et al.
Functional studies of ILPs in other mosquitoes are restricted two lines of investigation. First, data support a role for insulin signalling in arrest of ovary development and lipid storage in overwintering diapause by C. pipiens (Sim and Denlinger, 2009). Second, ingestion of vertebrate insulin in a blood meal activates insulin signalling and synthesis of reactive oxygen species in the midgut of anopheline mosquitoes (Kang et al., 2008).
4.1.12. Kinin
The first kinin was identified from the cockroach Rhyparobia (formerly Leucophaea) maderae (Holman et al., 1986), which was followed by identification of structurally similar peptides from other insects (Hayes et al., 1997; Hewes and Taghert, 2001). The single kinin gene in Ae. aegypti encodes a prepropeptide that is processed into three isoforms but the kinin gene from D. melanogaster is processed into only one (Hewes and Taghert, 2001; Veenstra et al., 1997b). In turn, Ae. aegypti encodes two class A GPCRs that bind kinins while D. melanogaster encodes one (Hewes and Taghert, 2001; Pietrantonio et al., 2005; Taneja-Bageshwar et al., 2009; Vogel et al., 2013). Immunostaining localizes the kinin receptor to the membrane of endocrine cells in the posterior midgut, Malpighian tubules, and the rectal pads of female Ae. aegypti (Kersch and Pietrantonio, 2011). Vogel et al. (2013) assigns these receptors to subassemblage 2b along with the receptors that bind RYamide, tachykinin, NPF, sNPF, and SIFamide. Functional studies in Ae. aegypti provide strong support for the role of kinins in diuresis (Beyenbach and Piermarini, 2010; Kersch and Pietrantonio, 2011), which is further discussed in chapter “Renal Excretory Processes in Mosquitoes” by Piermarini.
4.1.13. Neuropeptide F (NPF)
NPFs are 28–40 amino acid peptides that share a consensus C-terminal RxRFamide motif (Näassel and Wegener, 2011). They are broadly conserved among invertebrates and also share homology with vertebrate neuropeptide Y (NPY) and pancreatic polypeptides that regulate appetite, digestion, reproduction, stress, and other activities (Holzer et al., 2012). The first insect NPF was identified from D. melanogaster on the basis of its RFamide immunoreactivity (Brown et al., 1999), which was followed by cloning of an NPF gene from Ae. aegypti (Stanek et al., 2002). Drosophilids and mosquitoes both encode a single NPF gene while mature NPFs bind Class A GPCRs in subassemblage 2b that are most closely related to the receptors that bind sNPFs (Näassel and Wegener, 2011; Vogel et al., 2013). This finding is interesting because NPFs and sNPFs are structurally distinct and derive from unrelated genes (see below). Functional studies in D. melanogaster indicate NPF has similar activities to NPY in mammals (Näassel and Wegener, 2011). In contrast, little is known about NPF function in mosquitoes. Expression studies detect NPF mRNA in the brain and midgut of Ae. aegypti while binding studies show that mature NPF from Ae. aegypti and An. gambiae both bind the An. gambiae NPF receptor (Garczynski et al., 2005; Stanek et al., 2002). More recently, studies of five Class A, subassemblage 2 GPCRs from Ae. aegypti identified two that bind Ae. aegypti NPF (Liesch et al., 2013). The only reported function for NPF in mosquitoes is the inhibition of anterior midgut peristalsis in larval stage Ae. aegypti (Onken et al., 2004), but it is highly likely these peptide hormones have other, more functionally significant activities, given the greater literature.
4.1.14. Ovary Ecdysteroidogenic Hormone (OEH)
OEH is a relatively large 9–13 kDa hormone that as previously noted was identified from Ae. aegypti (Brown et al., 1998). It is structurally related to neuroparsins which are peptides of unclear function that were first isolated from locusts (Badisco et al, 2007). OEH is released with ILPs from medial neurosecretory cells with both stimulating ovary follicle cells to produce ecdysone (Fig. 1). Functional redundancy with ILPs together with the absence of an identified receptor obscured the mode of action of OEH for many years. This changed with identification of the OEH receptor as an RTK that is closely related to the IR (Vogel et al., 2015). Results also show that OEH directly induces ecdysone production by activating PI3K/Akt signalling (Dhara et al., 2013; Vogel et al., 2015). Thus, redundancy is due to OEH and ILPs binding different receptors on follicle cells that appear to both activate insulin pathway signalling. In contrast, OEH does not activate late-phase trypsin expression or haemocyte proliferation because OEH receptor expression is restricted to ovary follicle cells (Vogel et al., 2015). Some mosquito species have evolved from blood feeding relatives to produce eggs without blood feeding (autogeny). Recent studies show that facultatively autogenous Georgecraigius (=Aedes) atropalpus releases OEH and ILP from medial neurosecretory cells shortly after adult emergence but OEH is the primary regulator of ecdysone synthesis by the ovaries (Gulia-Nuss et al., 2012). OEH alone also stimulates limited egg development in nonblood fed Ae. aegypti (Gulia-Nuss et al., 2015). These findings suggest the evolution of autogeny is due in part to the shift from blood meal dependent to blood meal-independent release of OEH and ILPs from the nervous system.
4.1.15. Pyrokinins (PKs)
Peptide hormones we name PKs are relatively small and usually end with the C-terminal motif PRL/Vamide. This motif is similar to that of ETHs discussed earlier, which is why these peptides are often referred to collectively as PRXamides (Jurenka, 2015). The PRXamide literature is very difficult to follow for most individuals. One reason is because of the range of activities PRXamide peptides exhibit. The other is confusing nomenclature. PRXamides are named for either the biological activity or structural features exhibited by the first peptides that were identified. Thus, cardioacceleratory peptide (CAPA) was the name given for a PRXamide purified from M. sexta, leucopyrokinin (PK), and perviscerokinin (PVK) for myotropic PRXamides from cockroaches (R. maderae and P. americana), diapause hormone for PRXamides from Bombyx mori and Helicoverpa zea (DH-2), ETHs for PRXamides from M. sexta, pheromone biosynthesis activating peptide (PBAN) for a PRXamide from H. zea, and diuretic hormone (DH-1) for a PRXamide from P. americana (see Jurenka, 2015 for primary citations).
Peptide hormones are often named this way. It also works well for ETHs, which derive from a single gene and exhibit conservation in sequence and function. In contrast, the other PRXamides often exhibit cross-reactivity or activities that differ from given names when studied in different species. They also derive from two distinct genes. The first of these was named pban in H. zea because it encoded the corresponding peptide. However, pban from H. zea also contains four other PRXamides: DH-2 and three others referred to as PKs (Jurenka, 2015). Orthologs of this gene are known in many insects including D. melanogaster where it is named hugin. Hugin though encodes only two PRXamides named PK and hugin λ (see Näassel and Winther, 2010). The second gene is capability (capa), which was first identified in D. melanogaster with orthologs also now known from other insects. This gene usually contains three PRXamides: two named CAPA or CAPA-PVK peptides and a third named DH-1 or PK depending on publication (see Jurenka, 2015; Näassel and Winther, 2010). These names derive from sequence similarities with the originally identified CAPA, PVK, or PKs but again functional data often differ when studied in different species.
Given this background, we discuss ETHs as distinct peptide hormones (see earlier) because their structure and function support this. In contrast, the functional literature does not currently support giving different names to the peptides encoded by the pban/hugin or capa genes. We therefore name these genes in mosquitoes pk1 and pk2. In Ae. aegypti pk1 encodes four peptides we name PK1a–d while pk2 encodes three we name PK2a–c. The Class A GPCRs in mosquitoes and drosophilids that bind peptides encoded by the above genes form one clade in subassemblage 2F while the receptors that bind ETHs form a sister group (Choi et al., 2013; Dai and Adams, 2009; Olsen et al., 2007; Vogel et al., 2013). Peptidomic and immunocytochemistry methods detect PKs in the CNS, periviseral organs, CC, and midgut of adult mosquitoes (Hellmich et al., 2014; Predel et al., 2010). The only functional data in mosquitoes show that PK2a from Ae. aegypti inhibits fluid secretion in larval Malpighian tubules at femtomolar concentration but stimulates diuresis at higher concentrations (Ionescu and Donini, 2012). By previous nomenclature PK2a would be referred to as CAPA1 (Jurenka, 2015), which is inconsistent with its diuretic activity.
4.1.16. Short Neuropeptide F (sNPF)
sNPFs are peptides of variable length (6–19 amino acids) that end with the consensus C-terminal motif PxLRLRFamide in insects (Näassel and Wegener, 2011). All sequenced insect and other invertebrate genomes contain one or more sNPF genes but orthologs are absent from vertebrates. Among holometabolous insects like mosquitoes, prepropeptides often contain multiple sNPF sequences, whereas hemimetabolous insects and other invertebrates tend to produce prepropeptides that contain only one sNPF. The naming of NPFs and sNPFs derives from early studies in the literature where multiple peptides with related C-termini were identified. Later studies showed that NPFs and sNPFs derive from different genes and bind distinct but related class A, GPCRs that for Diptera reside in subassemblage 2b (Näassel and Wegener, 2011; Vogel et al., 2013). We earlier noted that the sNPF gene has duplicated in Ae. aegypti but not in An. gambiae or C. quinquefasciatus. Ae. aegypti sNPF1 encodes what was historically named sNPF while sNPF2 contains three copies of what was originally named head peptide (Matsumoto et al., 1989b; Stracker et al., 2002).
A diversity of neurons in the CNS and chemosensory structures like the antennae express sNPF in D. melanogaster while functional data implicate sNPFs in regulating feeding, growth, and ILP production by medial neurosecretory cells (Näassel and Winther, 2010). The literature likewise suggests sNPFs are broadly expressed in the adult mosquito CNS (Garczynski et al., 2007; Matsumoto et al., 1989b; Predel et al., 2010; Siju et al., 2013; Stracker et al., 2002; Veenstra, 1999) (Fig. 1), while a recent study reports male accessory glands in Ae. aegypti produce and transfer sNPF2 to females during mating (Naccarati et al., 2012). Early functional experiments in Ae. aegypti showed that sNPF2 titer increases in the hemolymph of adult females after consuming a blood meal, which coincides with females exhibiting no host-seeking behaviour (Brown et al., 1994). Nonblood fed females in contrast are highly responsive to vertebrate hosts but injection of sNPF2 inhibited host seeking for up to 5 h which suggested a role for sNPF2 in regulating host-seeking behaviour (Brown et al., 1994). More recent studies confirm that Ae. aegypti NPF and sNPFs bind different receptors but also detected overlap in receptor binding between sNPF1 and sNPF2 (Liesch et al., 2013). Bioassays identified roles for both NPF and sNPFs in host-seeking behaviour by nonblood females, yet curiously mutagenesis of the GPCR that binds sNPFs has no effect on host seeking, sugar, and blood-feeding behaviour, egg laying, or locomotion (Liesch et al., 2013). Naccarati et al. (2012) tested whether sNPF2 regulates female mating receptivity in Ae. aegypti because a structurally unrelated factor named sex peptide in male accessory gland secretions of D. melanogaster exhibits this activity. However, sNPF2 had no effect on female mating receptivity. Lastly, one study reports that sNPF1 and two both inhibit peristalsis of the Ae. aegypti larval anterior midgut (Onken et al., 2004).
4.2. Concluding Remarks
Significant progress has been made in identifying peptide hormone genes and receptors in mosquitoes. The functional literature is also strong in a few areas of study that focus on adult females. These include the roles of peptide hormones in regulating egg formation, JH biosynthesis, and diuresis, which are also subjects of separate chapters in this volume. In contrast, mapping the cell sources of different peptides hormones across life stages remains weak as is the literature on functional activities during immature development. For example, the endocrine regulation of moulting and metamorphosis in D. melanogaster implicates PTTH in regulating ecdysone production by prothoracic glands (PGs) and insulin signalling in nutrition-dependent growth (Nijhout et al., 2014; Rewitz et al., 2009). No data, however, support that PTTH or ILPs directly activate ecdysone production (Niwa and Niwa, 2014). Given the literature on many other insects, it is somewhat ironic that surprisingly little is known about the regulation of moulting in mosquitoes. An early study showed that larval thoracic and abdominal wall tissues produce ecdysone yet also reported that PGs do not (Jenkins et al., 1992). These findings together with the Drosophila literature indicate our understanding of peptide hormone functions in moulting are far from complete for Diptera. The orphan GPCR, RTK, and RGCs in mosquitoes also suggests some peptide hormones remain to be identified and characterized.
ACKNOWLEDGEMENTS
Research from the authors that is cited here was supported by the National Institutes of Health (R01AI1033108, R01AI106892, and F32GM109750) and Georgia Agricultural Experiment Station. We thank J.A. Johnson for her assistance with Fig. 1.
REFERENCES
- Adams MD, Celniker SE, Holt RA, Evans CA, Gocayne JD, Amanatides PG, Scherer SE, Li PW, Hoskins RA, Galle RF, George RA, et al. , 2000. The genome sequence of Drosophila melanogaster. Science 287, 2185–2195. [DOI] [PubMed] [Google Scholar]
- Akbari OS, Antoshechkin I, Amrhein H, Williams B, Diloreto R, Sandler J, Hay BA, 2013. The developmental transcriptome of the mosquito Aedes aegypti, an invasive species and major arbovirus vector. G3 (Bethesda) 3, 1493–1509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Antonova Y, Arik AJ, Moore W, Riehle MR, Brown MR, 2012. Insulin-like peptides: structure, signaling, and function In: Gilbert LI (Ed.), Insect Endocrinology. Elsevier, New York, pp. 63–92. [Google Scholar]
- Areiza M, Nouzova M, Rivera-Perez C, Noriega FG, 2014. Ecdysis triggering hormone ensures proper timing of juvenile hormone biosynthesis in pharate adult mosquitoes. Insect Biochem. Mol. Biol 54, 98–105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arensburger P, Megy K, Waterhouse RM, Abrudan J, Amedeo P, Antelo B, Bartholomay L, Bidwell S, Caler E, Camara F, Campbell CL, Campbell KS, et al. , 2010. Sequencing of Culex quinquefasciatus establishes a platform for mosquito comparative genomics. Science 330, 86–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arsic D, Guerin PM, 2008. Nutrient content of diet affects the signaling activity of the insulin/target of rapamycin/p70 S6 kinase pathway in the African malaria mosquito Anopheles gambiae. J. Insect Physiol 54, 1226–1235. [DOI] [PubMed] [Google Scholar]
- Attwood TK, Findlay JB, 1994. Fingerprinting G-protein-coupled receptors. Protein Eng 7, 195–203. [DOI] [PubMed] [Google Scholar]
- Badisco L, Claeys I, van Loy T, van Hiel M, Franssens CV, Simonet G, Vanden Broeck J, 2007. Neuroparsins, a family of conserved arthropod neuropeptides. Gen. Comp. Endocrinol 153, 64–71. [DOI] [PubMed] [Google Scholar]
- Baker KD, Thummel CS, 2007. Diabetic larvae and obese flies-emerging studies of metabolism in Drosophila. Cell Metab 6, 257–266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baker DA, Nolan T, Fishcer B, Pinder A, Crisanti A, Russell S, 2011. A comprehensive gene expression atlas of sex- and tissue-specificity in the malaria vector, Anopheles gambiae. BMC Genomics 12, 296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balment RJ, Lovejoy DA, 1999. Evolution and physiology of corticotropin-releasing factor (CRF) family of neuropeptides in vertebrates. Gen. Comp. Endocrinol 115, 1–22. [DOI] [PubMed] [Google Scholar]
- Bani D, 1997. Relaxin: a pleiotrophic hormone. Gen. Pharmacol 28, 13–22. [DOI] [PubMed] [Google Scholar]
- Belmont M, Cazzamali G, Williamson M, Hauser F, Grimmelikhuijzen CJ, 2006. Identification of four evolutionarily related G protein-coupled receptors from the malaria mosquito Anopheles gambiae. Biochem. Biophys. Res. Commun 344, 160–165. [DOI] [PubMed] [Google Scholar]
- Beyenbach KW, Piermarini PM, 2010. Transcellular and paracellular pathways of transepithelial fluid secretion in Malphighian (renal) tubules of the yellow fever mosquito Aedes aegypti. Acta Physiol 202, 387–407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biarc J, Chalkley RJ, Burlingame AL, Bradshaw RA, 2011. Receptor tyrosine kinase signaling—a proteomic perspective. Adv. Enzym. Regul 51, 293–305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bockhaert J, Pin JP, 1999. Molecular tinkering of G protein-coupled receptors: an evolutionary success. EMBO J 18, 1723–1729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borovsky D, Carlson DA, Griffin PR, Shabanowitz J, Hunt DF, 1990. Mosquito oostatic factor: a novel decapeptide modulating trypsin-like enzyme biosynthesis in the midgut. FASEB J 4, 3015–3020. [DOI] [PubMed] [Google Scholar]
- Brody T, Cravchik A, 2000. Drosophila melanogaster G protein-coupled receptors. J. Cell Biol 150, F83–F88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brogiolo W, Stocker H, Ikeya T, Rintelen F, Fernandez R, Hafen E, 2001. An evolutionarily conserved function of the Drosophila insulin receptor and insulin-like peptides in growth control. Curr. Biol 11, 213–221. [DOI] [PubMed] [Google Scholar]
- Broughton S, Partridge L, 2009. Insulin/IGF-like signaling, the central nervous system, and aging. Biochem. J 15, 1–12. [DOI] [PubMed] [Google Scholar]
- Brown MR, Cao C, 2001. Distribution of ovary ecdysteroidogenic hormone I in the nervous system and gut of mosquitoes. J. Insect Sci 1, 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown MR, Lea AO, 1988. FMRFamide- and adipokinetic hormone-like immunoreactivity in the nervous system of the mosquito, Aedes aegypti. J. Comp. Neurol 270, 606–614. [DOI] [PubMed] [Google Scholar]
- Brown MR, Crim JW, Lea AO, 1986. FMRFamide- and pancreatic polypeptide-like immunoreactivity of endocrine cells in the midgut of a mosquito. Tissue Cell 18, 419–428. [DOI] [PubMed] [Google Scholar]
- Brown MR, Raikhel AS, Lea AO, 1988. Ultrastructure of midgut endocrine cells in the adult mosquito, Aedes aegypti. Tissue Cell 17, 709–721. [DOI] [PubMed] [Google Scholar]
- Brown MR, Klowden MJ, Crim JW, Young L, Shrouder LA, Lea AO, 1994. Endogenous regulation of mosquito host-seeking behavior by a neuropeptide. J. Insect Physiol 40, 399–406. [Google Scholar]
- Brown MR, Graf R, Swiderek KM, Fendley D, Stracker TM, Champagne DE, Lea AO, 1998. Identification of a steroidogenic neurohormone in female mosquitoes. J. Biol. Chem 273, 3967–3971. [DOI] [PubMed] [Google Scholar]
- Brown MR, Crim JW, Arata RC, Cai HN, Chun C, Shen P, 1999. Identification of a Drosophila brain-gut peptide related to the neuropeptide Y family. Peptides 20, 1035–1042. [DOI] [PubMed] [Google Scholar]
- Brown MR, Clark KD, Gulia M, Zhao Z, Garczynski SF, Crim JW, Sulderman RJ, Strand MR, 2008. An insulin-like peptide regulates egg maturation and metabolism in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. U. S. A 105, 5716–5721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caers J, Peeters L, Janssen T, De Haes W, Gade G, Schoofs L, 2012. Structure-activity studies of Drosophila adipokinetic (AKH) by a cellular expression system of dipteran AKH receptors. Gen. Comp. Endocrinol 77, 332–337. [DOI] [PubMed] [Google Scholar]
- Cao C, Brown MR, 2001. Localization of an insulin-like peptide in brains of two flies. Cell Tissue Res 304, 317–321. [DOI] [PubMed] [Google Scholar]
- Cardoso JC, Félix RC, Bergqvist CA, Larhammar D, 2014. New insights into the evolution of vertebrate CRH (corticotropin-releasing hormone) and invertebrate DH44 (diuretic hormone 44) receptors in metazoans. Gen. Comp. Endocrinol 209, 162–170. [DOI] [PubMed] [Google Scholar]
- Castillo J, Brown MR, Strand MR, 2011. Blood feeding and insulin-like peptide 3 stimulate proliferation of hemocytes in the mosquito Aedes aegypti. PLoS Pathog 7, e1002274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang JC, Yang RB, Adams ME, Lu KH, 2009. Receptor guanylyl cyclases in Inka cells targeted by eclosion hormone. Proc. Natl. Acad. Sci. U. S. A 106, 13371–13376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen W, Hillyer JF, 2013. FlyNap (triethylamine) increases the heart rate of mosquitoes and eliminates the cardioacceleratory effect of the neuropeptide CCAP. PLoS One 16, e70414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi MY, Estep A, Sanscrainte N, Becnel J, Vander Meer RK, 2013. Identification and expression of PBAN/diapause hormone and GPCRs from Aedes aegypti. Mol. Cell. Endocrinol 375, 113–120. [DOI] [PubMed] [Google Scholar]
- Clark TM, Hayes TK, Beyenbach KW, 1998. Dose-dependent effects of CRF-like diuretic peptide on transcellular and paracellular transport pathways. Am. J. Physiol. Renal Physiol 274, F834–F840. [DOI] [PubMed] [Google Scholar]
- Clements AN, 1956. Hormonal control of ovary development in mosquitoes. J. Exp. Biol 33, 211–223. [Google Scholar]
- Coast GM, Garside CS, Webster SG, Schegg KM, Schooley DA, 2005. Mosquito natriuretic peptide identified as a calcitonin-like diuretic hormone in Anopheles gambiae (Giles). J. Exp. Biol 208, 3281–3291. [DOI] [PubMed] [Google Scholar]
- Dai L, Adams ME, 2009. Ecdysis triggering hormone signaling in the yellow fever mosquito Aedes aegypti. Gen. Comp. Endocrinol 162, 43–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Meyts P, Gauguin L, Svendsen AM, Sarhan M, Knudsen L, Nøhr J, Kiselyov VV, 2009. Structural basis of allosteric ligand–receptor interactions in the insulin/relaxin peptide family: implications for other receptor tyrosine kinases and G-protein-coupled receptors. Ann. N. Y. Acad. Sci 1160, 45–53. [DOI] [PubMed] [Google Scholar]
- Detinova TS, 1945. On the influence of glands of internal secretion upon the ripening of the gonads and the imaginal diapause of Anopheles maculipennis. Zool. Zhurnal 34, 291–298. [Google Scholar]
- Dhara A, Eum JH, Robertson A, Gulia-Nuss M, Vogel KJ, Clark KD, Graf R, Brown MR, Strand MR, 2013. Ovary ecdysteroidogenic hormone functions independently of the insulin receptor in the yellow fever mosquito, Aedes aegypti. Insect Biochem. Mol. Biol 43, 1100–1108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dow JA, Davies SA, 2006. The Malphighian tubule: rapid insights from post-genomic biology. J. Insect Physiol 52, 365–378. [DOI] [PubMed] [Google Scholar]
- Duckworth WC, Garcia JV, Liepnieks JJ, Hamel FG, Hermodson MA, Frank BH, Rosner MR, 1989. Drosophila insulin degrading enzyme and rat skeletal muscle insulin protease cleave insulin at similar sites. Biochemistry 28, 2471–2477. [DOI] [PubMed] [Google Scholar]
- Duttlinger A, Mispelon M, Nichols R, 2003. The structure of the FMRFamide receptor and activity of the cardioexcitatory neuropeptide are conserved in mosquito. Neuropeptides 37, 120–126. [DOI] [PubMed] [Google Scholar]
- Ellison HE, Estévez-Lao TY, Murphree CS, Hillyer JF, 2015. Deprivation of both sucrose and water reduces the mosquito heart contraction rate while increasing the expression of nitric oxide synthase. J. Insect Physiol 74, 1–9. [DOI] [PubMed] [Google Scholar]
- Estévez-Lao TY, Boyce DS, Honegger HW, Hillyer JF, 2013. Cardioacceleratory function of the neurohormone CCAP in the mosquito Anopheles gambiae. J. Exp. Biol 216, 601–613. [DOI] [PubMed] [Google Scholar]
- Fernandez-Almonacid R, Rosen OM, 1987. Structure and ligand specificity of the Drosophila melanogaster insulin receptor. Mol. Cell. Biol 7, 2718–2727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernández-Gamba A, Leal MC, Morelli L, Castaño EM, 2009. Insulin-degrading enzyme: structure–function relationship and its possible roles in health and disease. Curr. Pharm. Des 15, 3644–3655. [DOI] [PubMed] [Google Scholar]
- Fricker LD, 2012. Neuropeptides and Other Bioactive Peptides: From Discovery to Function. Morgan & Claypool Life Sciences, San Rafael, CA, 107 pp. [Google Scholar]
- Furuya K, Milchak RJ, Schegg KM, Zhang J, Tobe SS, Coast GM, Schooley DA, 2000. Cockroach diuretic hormone: characterization of a calcitonin-like peptide in insects. Proc. Natl. Acad. Sci. U. S. A 97, 6469–6474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gäade G, 2009. Peptides of the adipokinetic hormone/red pigment-concentrating hormone family: a new take on biodiversity. Ann. N. Y. Acad. Sci 1163, 125–136. [DOI] [PubMed] [Google Scholar]
- Gäade G, Hoffmann KH, Spring JH, 1997. Hormonal regulation in insects: facts, gaps, and future directions. Physiol. Rev 77, 963–1032. [DOI] [PubMed] [Google Scholar]
- Galagovsky D, Katz MJ, Acevedo JM, Sorianello E, Glavic A, Wappner P, 2014. The Drosophila insulin-degrading enzyme restricts growth by modulating the PI3K pathway in a cell-autonomous manner. Mol. Biol. Cell 25, 916–924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garczynski SF, Crim JW, Brown MR, 2005. Characterization of neuropeptide F and its receptor from the African malaria mosquito, Anopheles gambiae. Peptides 26, 99–107. [DOI] [PubMed] [Google Scholar]
- Garczynski SF, Crim JW, Brown MR, 2007. Characterization and expression of the short neuropeptide F receptor in the African malaria mosquito, Anopheles gambiae. Peptides 28, 109–118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garelli A, Heredia F, Casimiro AP, Macedo A, Nunes C, Garcez M, Dias AR, Volonte YA, Uhlmann T, Caparros E, Koyama T, Gontijo AM, 2015. Dilp8 requires the neuronal relaxin receptor Lgr3 to couple growth to developmental timing. Nat. Commun 6, 8732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gillett JD, 1956. Initiation and promotion of ovarian development in the mosquito Aedes (Stegomyia) aegypti (Linnaeus). Ann. Trop. Med. Parasitol 50, 375–380. [DOI] [PubMed] [Google Scholar]
- Gouzi JY, Moressis A, Walker JA, Apostolopoulou AA, Palmer RH, Bernards A, Skoulakis EM, 2011. The receptor tyrosine kinase Alk controls neurofibromin functions in Drosophila growth and learning. PLoS Genet 7, e1002281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graf R, Lea AO, Briegel H, 1998. A temporal profile of the endocrine control of trypsin synthesis in the yellow fever mosquito, Aedes aegypti. J. Insect Physiol 44, 451–454. [DOI] [PubMed] [Google Scholar]
- Grönke S, Clarke D-F, Broughton S, Andrews TD, Partridge L, 2010. Molecular evolution and functional characterization of Drosophila insulin-like peptides. PLoS Genet 6, e1000857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gulia-Nuss M, Robertson AE, Brown MR, Strand MR, 2011. Insulin-like peptides and the target of rapamycin pathway coordinately regulate blood digestion and egg maturation in the mosquito Aedes aegypti. PLoS One 6, e20401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gulia-Nuss M, Eum JH, Strand MR, Brown MR, 2012. Ovary ecdysteroidogenic hormone activates egg maturation in the mosquito Georgecraigius atropalpus after adult eclosion or a blood meal. J. Exp. Biol 215, 3758–3767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gulia-Nuss M, Elliot A, Brown MR, Strand MR, 2015. Multiple factors contribute to anautogenous reproduction by the mosquito Aedes aegypti. J. Insect Physiol 82, 8–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagedorn HH, O’Connor J, Fuchs MS, Sage B, Schlaeger DA, Bohm M, 1975. The ovary as a source of alpha-ecdysone in an adult mosquito. Proc. Natl. Acad. Sci. U. S. A 72, 3255–3259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagedorn HH, Shapiro JP, Hanaoka K, 1979. Ovarian ecdysone secretion is controled by a brain hormone in the adult mosquito. Nature 282, 92–94. [DOI] [PubMed] [Google Scholar]
- Hansen KK, Stafflinger E, Schneider M, Hauser F, Cazzamali G, Williamson M, Kollmann M, Schachtner J, Grimmelikhuijzen CJ, 2010. Discovery of a novel insect neuropeptide signaling system closely related to the insect adipokinetic hormone and corazonin hormonal systems. J. Biol. Chem 285, 10736–10747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauser F, Grimmelikhuijzen CJ, 2014. Evolution of the AKH/corazonin/ACP/GnRH receptor superfamily and their ligands in the Protostomia. Gen. Comp. Endocrinol 209, 35–49. [DOI] [PubMed] [Google Scholar]
- Hauser F, Nothacker HP, Grimmelikhuijzen CJ, 1997. Molecular cloning, genomic organization, and developmental regulation of a novel receptor from Drosophila melanogaster structurally related to members of the thyroid-stimulating hormone, follicle-stimulating hormone, luteinizing hormone/choriogonadotropin receptor family from mammals. J. Biol. Chem 272, 1002–1010. [DOI] [PubMed] [Google Scholar]
- Hauser F, Cazzamali G, Williamson M, Blenau W, Grimmelikhuijzen CJP, 2006a. A review of neurohormone GPCRs present in the fruitfly Drosophila melanogaster and the honey bee Apis mellifera. Prog. Neurobiol 80, 1–19. [DOI] [PubMed] [Google Scholar]
- Hauser F, Williamson M, Cazzamali G, Grimmelikhuijzen CJ, 2006b. Identifying neuropeptide and protein hormone receptors in Drosophila melanogaster by exploiting genomic data. Brief. Funct. Genomic. Proteomic 4, 321–330. [DOI] [PubMed] [Google Scholar]
- Hauser F, Cazzamali G, Williamson M, Park Y, Li B, Tanaka Y, Predel R, Neupert S, Schachtner J, Verleyen P, Grimmelikhuijzen CJP, 2008. A genome-wide inventory of neurohormone GPCRs in the red flour beetle Tribolium castaneum. Front. Neuroendocrinol 29, 142–165. [DOI] [PubMed] [Google Scholar]
- Hauser F, Neupert S, Williamson M, Predel R, Tanaka Y, Grimmelikhuijzen CJP, 2010. Genomics and peptidomics of neuropeptides and protein hormones present in the parasitic wasp Nasonia vitripennis. J. Proteome Res 9, 5296–5310. [DOI] [PubMed] [Google Scholar]
- Hayes TK, Strey A, Belk S, Holman GM, Nachman RJ, Petzel D, Readio J, Meola R, Pannabecker T, Beyenbach K, 1997. Biochemical characterization of mosquito kinin and related receptors. Ann. N. Y. Acad. Sci 814, 342–345. [DOI] [PubMed] [Google Scholar]
- Hector CE, Bretz CA, Zhao Y, Johnson EC, 2009. Functional differences between two CRF-related diuretic hormone receptors in Drosophila. J. Exp. Biol 212, 3142–3147. [DOI] [PubMed] [Google Scholar]
- Hellmich E, Nusawardani T, Bartholomay L, Jurenka R, 2014. Pyrokinin/PBAN-like peptides in the central nervous system of mosquitoes. Cell Tissue Res 356, 39–47. [DOI] [PubMed] [Google Scholar]
- Hernández-Martínez S, Li Y, Lanz-Mendoza H, Rodríguez MH, Noriega FG, 2005. Immunostaining for allatotropin and allatostatin-A and -C in the mosquitoes Aedes aegypti and Anopheles albimanus. Cell Tissue Res 321, 105–113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hernández-Martínez S, Mayoral JG, Li Y, Noriega FG, 2007. Role of juvenile hormone and allatotropin on nutrient allocation, ovarian development and survivorship in mosquitoes. J. Insect Physiol 53, 230–234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hewes RS, Taghert PH, 2001. Neuropeptides and neuropeptide receptors in the Drosophila melanogaster genome. Genome Res 11, 1126–1142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill CA, Fox AN, Pitts RJ, Kent LB, Tan PL, Chrystal MA, Cravchik A, Collins FH, Robertson HM, Zwiebel LJ, 2002. G protein coupled receptors in Anopheles gambiae. Science 298, 176–178. [DOI] [PubMed] [Google Scholar]
- Hillyer JF, Estevez-Lao TY, Funkhouser LJ, Aluoch VA, 2012. Anopheles gambiae corazonin: gene structure, expression and effect on mosquito heart physiology. Insect Mol. Biol 21, 343–355. [DOI] [PubMed] [Google Scholar]
- Hillyer JF, Estevez-Lao TY, de la Parte LE, 2014. Myotropic effects of FMRFamide containing peptides on the heart of the mosquito Anopheles gambiae. Dev. Comp. Immunol 202, 15–25. [DOI] [PubMed] [Google Scholar]
- Holman GM, Cook BJ, Nachman RJ, 1986. Primary structure and synthesis of a blocked myotropic neuropeptide isolated from the cockroach, Leucophaea maderae. Comp. Biochem. Physiol. C 85, 219–224. [DOI] [PubMed] [Google Scholar]
- Holt RA, Subramanian GM, Halpern A, Sutton GG, Charlab R, Nusskern DR, Wincker P, Clark AG, Ribeiro JMC, Wides R, Salzberg SL, et al. , 2002. The genome sequence of the malaria mosquito Anopheles gambiae. Science 298, 129–135. [DOI] [PubMed] [Google Scholar]
- Holzer P, Reichmann F, Farzi A, 2012. Neuropeptide Y, peptide YY and pancreatic polypeptide in the gut-brain axis. Neuropeptides 46, 261–274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honegger HW, Estevez-Lao TY, Hillyer JF, 2011. Bursicon-expressing neurons undergo apoptosis after adult ecdysis in the mosquito Anopheles gambiae. J. Insect Physiol 57, 1017–1022. [DOI] [PubMed] [Google Scholar]
- Hook V, Funkelstein L, Lu D, Bark S, Wegrzyn J, Hwang SR, 2008. Proteases for processing proneuropeptides into peptide neurotransmitters and hormones. Annu. Rev. Pharmacol. Toxicol 48, 393–423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hubbard SR, Miller WT, 2007. Receptor tyrosine kinases: mechanisms of activation and signaling. Curr. Opin. Cell Biol 19, 117–123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hummon AB, Richmond TA, Verleyen P, Baggerman G, Huybrechts J, Ewing MA, Vierstraete E, Rodriguez-Zas SL, Schoofs L, Robinson GE, Sweedler JV, 2006. From the genome to the proteome: Uncovering peptides in the Apis brain. Science 314, 647–649. [DOI] [PubMed] [Google Scholar]
- Ionescu A, Donini A, 2012. AedesCAPA-PVK-1 displays diuretic and dose dependent antidiuretic potential in the larval mosquito Aedes aegypti (Liverpool). J. Insect Physiol 58, 1299–1306. [DOI] [PubMed] [Google Scholar]
- Isaac RE, Bland ND, Shirras AD, 2009. Neuropeptidases and the metabolic inactivation of insect neuropeptides. Gen. Comp. Endocrinol 162, 8–17. [DOI] [PubMed] [Google Scholar]
- Iversen A, Cazzamali G, Williamson M, Hauser F, Grimmelikhuijzen CJP, 2002. Molecular identification of the first insect ecdysis triggering hormone receptors. Biochem. Biophys. Res. Commun 299, 924–931. [DOI] [PubMed] [Google Scholar]
- Jagge CL, Pietrantonio PV, 2008. Diuretic hormone 44 receptor in Malpighian tubules of the mosquito Aedes aegypti: evidence for transcriptional regulation paralleling urination. Insect Mol. Biol 17, 413–426. [DOI] [PubMed] [Google Scholar]
- Jenkins SP, Brown MR, Lea AO, 1992. Inactive prothoracic glands in larvae and pupae of Aedes aegypti: Ecdysteroid release by tissues in the thorax and abdomen. Insect Biochem. Mol. Biol 22, 553–559. [Google Scholar]
- Jiang H, Edgar BA, 2012. Intestinal stem cell function in Drosophila and mice. Curr. Opin. Genet. Dev 22, 354–360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang N, Kolhekarm AS, Jacobs PS, Mains RE, Eipper BA, Taghert PH, 2000. PHM is required for normal developmental transitions and for biosynthesis of secretory peptides in Drosophila. Dev. Biol 226, 118–136. [DOI] [PubMed] [Google Scholar]
- Jiang X, Peery A, Hall AB, Sharma A, Chen XG, Waterhouse RM, Komissarov A, Riehle MM, Shouche Y, Sharakhova MV, Lawson D, et al. , 2014. Genome analysis of a major urban malaria vector mosquito, Anopheles stephensi. Genome Biol 15, 459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson EC, Bohn LM, Barak LS, Birse RT, Näassel DR, Caron MG, Taghert PH, 2003. Identification of Drosophila neuropeptide receptors by G protein-coupled receptors-beta-arrestin2 interactions. J. Biol. Chem 278, 52172–52178. [DOI] [PubMed] [Google Scholar]
- Jurenka R, 2015. The PRXamide neuropeptide signalling system: conserved in animals. Adv. Insect Physiol 49, 123–169. [Google Scholar]
- Kang MA, Mott TM, Tapley EC, Lewis EE, Luckhart S, 2008. Insulin regulates aging and oxidative stress in Anopheles stephensi. J. Exp. Biol 211, 741–748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kataoka H, Troetschler RG, Li JP, Kramer SJ, Carney RL, Schooley DA, 1989. Isolation and identification of a diuretic hormone from the tobacco hornworm, Manduca sexta. Proc. Natl. Acad. Sci. U.S.A 86, 2976–2980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaufmann C, Brown MR, 2006. Adipokinetic hormones in the African malaria mosquito, Anopheles gambiae: identification and expression of genes for two peptides and a putative receptor. Insect Biochem. Mol. Biol 36, 466–481. [DOI] [PubMed] [Google Scholar]
- Kaufmann C, Brown MR, 2008. Regulation of carbohydrate metabolism and flight performance by a hypertrehalosaemic hormone in the mosquito Anopheles gambiae. J. Insect Physiol 54, 367–377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaufmann C, Merzendorfer H, Gade G, 2009. The adipokinetic hormone system in Culicinae (Diptera: Culicidae): Molecular identification and characterization of two adipokinetic hormone (AKH) precursors from Aedes aegypti and Culex pipiens and two putative AKH receptor variants from Ae. aegypti. Insect Biochem. Mol. Biol 39, 770–781. [DOI] [PubMed] [Google Scholar]
- Kersch CN, Pietrantonio PV, 2011. Mosquito Aedes aegypti (L.) leucokinin receptor is critical for in vivo fluid excretion post blood feeding. FEBS Lett 585, 3507–3512. [DOI] [PubMed] [Google Scholar]
- Klowden MJ, Lea AO, 1979. Humoral inhibition of host-seeking in Aedes aegypti during oocyte maturation. J. Insect Physiol 25, 231–235. [DOI] [PubMed] [Google Scholar]
- Krieger MJB, Jahan N, Riehle MA, Cao C, Brown MR, 2004. Molecular characterization of insulin-like peptide genes and their expression in the African malaria mosquito, Anopheles gambiae. Insect Mol. Biol 13, 305–315. [DOI] [PubMed] [Google Scholar]
- Kwon H, Pietrantonio PV, 2013. Calcitonin receptor 1 (AedaeGPCRCAL1) hindgut expression and direct role in myotropic action in females of the mosquito Aedes aegypti (L.). Insect Biochem.Mol. Biol 43, 588–593. [DOI] [PubMed] [Google Scholar]
- Kwon H, Lu HL, Longnecker MT, Pietrantonio PV, 2012. Role in diuresis of a calcitonin receptor (GPRCAL1) expressed in a distal-proximal gradient in renal organs of the mosquito Aedes aegypti (L.). PLos One 7, e50374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lasek AW, Lim J, Kliethermes CL, Berger KH, Joslyn G, Brush G, Xue L, Robertson M, Moore MS, Vranizan K, Morris SW, et al. , 2011. An evolutionary conserved role for anaplastic lymphoma kinase in behavioral responses to ethanol. PLoS One 6, e22636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lea AO, 1967. The medial neurosecretory cells and egg maturation in mosquitoes. J. Insect Physiol 13, 419–429. [DOI] [PubMed] [Google Scholar]
- Lea AO, 1972. Regulation of egg maturation in the mosquito by the neurosecretory system: the role of the corpus cardiacum. Gen. Comp. Endocrinol 3 (Suppl.), 602–608. [Google Scholar]
- Lea AO, Van Handel E, 1970. Suppression of glycogen synthesis in the mosquito by a hormone from the medial neurosecretory cells. J. Insect Physiol 16, 319–321. [DOI] [PubMed] [Google Scholar]
- Lemmon MA, Schlessinger J, 2010. Cell signaling by receptor tyrosine kinases. Cell 141, 1117–1134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Y, Unnithan GC, Veenstra JA, Feyereisen R, Noriega FG, 2003. Stimulation of JH biosynthesis by the corpora allata of adult female Aedes aegypti in vitro: effect of farnesoic acid and Aedes allatotropin. J. Exp. Biol 206, 1825–1832. [DOI] [PubMed] [Google Scholar]
- Li YP, Hernandez-Martinez S, Noriega FG, 2004. Inhibition of juvenile hormone biosynthesis in mosquitoes: effect of allatostatic head factors, PISCF- and YXFGL-amide-allatostatins. Reg. Peptides 118, 175–182. [DOI] [PubMed] [Google Scholar]
- Li Y, Hernandez-Martinez S, Fernandez F, Mayoral JG, Topalis P, Priestap H, Perez M, Navare A, Noriega FG, 2006. Biochemical, molecular, and functional characterization of PISCF-allatostatin, a regulator of juvenile hormone biosynthesis in the mosquito Aedes aegypti. J. Biol. Chem 281, 34048–34055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li B, Predel R, Neupert S, Hauser F, Tanaka Y, Cazzamali G, Williamson M, Arakane Y, Verleyen P, Schoofs L, Schachtner J, Grimmelikhuijzen CJP, Park Y, 2008. Genomics, transcriptomics, and peptidomics of neuropeptides and protein hormones in the red flour beetle Tribolium castaneum. Genome Res 18, 113–122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liesch J, Bellani LL, Vosshall LB, 2013. Functional and genetic characterization of neuropeptide Y-like receptors in Aedes aegypti. PLoS Negl. Trop. Dis 7, e2486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin Y, Hamblin MT, Edwards MJ, Barillas-Mury C, Kanost MR, Knipple DC, Wolfner MF, Hagedorn HH, 1993. Structure, expression, and hormonal control of genes from the mosquito, Aedes aegypti, which encode proteins similar to the vitelline membrane proteins of Drosophila melanogaster. Dev. Biol 155, 558–568. [DOI] [PubMed] [Google Scholar]
- Lorén CE, Scully A, Grabbe C, Edeen PT, Thomas J, McKeown M, Hunter T, Palmer RH, 2001. Identification and characterization of DAlk: a novel Drosophila melanogaster RTK which drives ERK activation in vivo. Genes Cells 6, 531–544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo CW, Dewey EM, Sudo S, Ewer J, Hsu SY, Honegger HW, Hsueh AJW, 2005. Bursicon, the insect cuticle-hardening hormone, is a heterodimeric cystine knot protein that activates G protein-coupled receptor LGR2. Proc. Natl. Acad. Sci. U. S. A 102, 2820–2825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maddrell SHP, Phillips JE, 1978. Induction of sulphate transport and hormonal control of fluid secretion by Malpighian tubules of larvae of the mosquito Aedes taeniorhynchus. J. Exp. Biol 72, 181–202. [Google Scholar]
- Marquez AG, Pietri JE, Smithers HM, Nuss A, Antonova Y, Drexler AL, Riehle MA, Brown MR, Luckhart S, 2011. Insulin-like peptides in the mosquito Anopheles stephensi: identification and expression in response to diet and infection with Plasmodium falciparum. Gen. Comp. Endocrinol 173, 303–312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsumoto S, Brown MR, Suzuki A, Lea AO, 1989a. Isolation and characterization of ovarian ecdysteroidogenic hormones from the mosquito, Aedes aegypti. Insect Biochem. Mol. Biol 19, 651–656. [Google Scholar]
- Matsumoto S, Brown MR, Crim JW, Vigna SR, Lea AO, 1989b. Isolation and primary structure of neuropeptides from the mosquito, Aedes aegypti, immunoreactive to FMRFamide antiserum. Insect Biochem 19, 277–283. [Google Scholar]
- Matthews BJ, McBride CS, DeGennaro M, Despo O, Vosshall LB, 2016. The neurotransciptome of the Aedes aegypti mosquito. BMC Genomics 17, 32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayoral JG, Nouzova M, Brockhoff A, Goodwin M, Hernandez-Martinez S, Richter D, Meyerhof W, Noriega FG, 2010. Allatostatin-C receptors in mosquitoes. Peptides 31, 442–450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDonald N, Murray-Rust J, Blundell T, 1989. Structure–function relationships of growth factors and their receptors. Br. Med. Bull 45, 554–569. [DOI] [PubMed] [Google Scholar]
- Mendive FM, Van Loy T, Claeysen S, Poels J, Williamson M, Hauser F, Grimmelikhuijzen CJP, Vassart G, Broeck JV, 2005. Drosophila molting neurohormone bursicon is a heterodimer and the natural agonist of the orphan receptor DLGR2. FEBS Lett 579, 2171–2176. [DOI] [PubMed] [Google Scholar]
- Meola SM, Lea AO, 1972. The ultrastructure of the corpus cardiacum of Aedes sollicitans and the histology of the cerebral neurosecretory system of mosquitoes. Gen. Comp. Endocrinol 18, 210–234. [DOI] [PubMed] [Google Scholar]
- Meola SM, Clottens FL, Holman GM, Nachman RJ, Nichols R, Schoofs L, Wright MS, Olson JK, Hayes TK, Pendleton MW, 1998. Isolation and immunocytochemical characterization of three tachykinin-related peptides from the mosquito, Culex salinarius. Neurochem. Res 23, 189–202. [DOI] [PubMed] [Google Scholar]
- Moffett SB, Moffett DF, 2005. Comparison of immunoreactivity to serotonin, FMRFamide and SCPb in the gut and visceral nervous system of larvae, pupae and adults of the yellow fever mosquito Aedes aegypti. J. Insect Sci 5, 20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mugumbate G, Jackson GE, Van Der Spoel D, 2011. Open conformation of adipokinetic hormone receptor from the malaria mosquito facilitates hormone binding. Peptides 32, 553–559. [DOI] [PubMed] [Google Scholar]
- Mugumbate G, Jackson GE, van der Spoel D, Kövér KE, Szilágyi L, 2013. Anopheles gambiae, Anoga-HrTH hormone, free and bound structure—a nuclear magnetic resonance experiment. Peptides 41, 94–100. [DOI] [PubMed] [Google Scholar]
- Naccarati C, Audsley N, Keen JN, Kim JH, Howell GJ, Kim YJ, Isaac RE, 2012. The host-seeking inhibitory peptide, Aea-HP-1, is made in the male accessory gland and transferred to the female during copulation. Peptides 34, 150–157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Näassel DR, Wegener C, 2011. A comparative review of short and long neuropeptide F signaling in invertebrates: any similarities to vertebrate neuropeptide Y signaling? Peptides 32, 1335–1355. [DOI] [PubMed] [Google Scholar]
- Näassel DR, Winther AME, 2010. Drosophila neuropeptides in regulation of physiology and behavior. Prog. Neurobiol 92, 42–104. [DOI] [PubMed] [Google Scholar]
- Neafsey DE, Waterhouse RM, Abai MR, Aganezov SS, Alekseyev MA, Allen JE, Amon J, Arcà B, Arensburger P, Artemov G, Assour LA, et al. , 2015. Mosquito genomics. Highly evolvable malaria vectors: the genomes of 16 Anopheles mosquitoes. Science 347, 1258522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nene V, Wortman JR, Lawson D, Haas B, Kodira C, Tu ZJ, Loftus B, Xi ZY, Megy K, Grabherr M, Ren QH, et al. , 2007. Genome sequence of Aedes aegypti, a major arbovirus vector. Science 316, 1718–1723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nichols R, 2003. Signaling pathways and physiological functions of Drosophila melanogaster FMRFamide-related peptides. Annu. Rev. Entomol 48, 485–503. [DOI] [PubMed] [Google Scholar]
- Nijhout HF, 1998. Insect Hormones. Princeton University Press, Princeton, NJ, 260 pp. [Google Scholar]
- Nijhout HF, Carrow GM, 1978. Diuresis after a bloodmeal in female Anopheles freeborni. J. Insect Physiol 24, 293–298. [Google Scholar]
- Nijhout HF, Riddiford LM, Mirth C, Shingleton AW, Suzuki Y, Callier V, 2014. The developmental control of size in insects. Wiley Interdiscip. Rev. Dev. Biol 3, 113–134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niwa R, Niwa YS, 2014. Enzymes for ecdysteroid biosynthesis: their biological functions in insects and beyond. Biosci. Biotechnol. Biochem 78, 1283–1292. [DOI] [PubMed] [Google Scholar]
- Nouzova M, Brockhoff A, Mayoral JG, Goodwin M, Meyerhof W, Noriega FG, 2012. Functional characterization of an allatotropin receptor expressed in the corpora allata of mosquitoes. Peptides 34, 201–208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noyes BE, Katz FN, Schaffer MH, 1995. Identification and expression of the Drosophila adipokinetic hormone gene. Mol. Cell. Endocrinol 109, 133–141. [DOI] [PubMed] [Google Scholar]
- Olsen SS, Cazzamali G, Williamson M, Grimmelikhuijzen CJP, Hauser F, 2007. Identification of one capa and two pyrokinin receptors from the malaria mosquito Anopheles gambiae. Biochem. Biophys. Res. Commun 362, 245–251. [DOI] [PubMed] [Google Scholar]
- Onken H, Moffett SB, Moffett DF, 2004. The anterior stomach of larval mosquitoes (Aedes aegypti): effects of neuropeptides on transepithelial ion transport and muscular motility. J. Exp. Biol 207, 3731–3739. [DOI] [PubMed] [Google Scholar]
- Overend G, Cabrero P, Guo AX, Sebastian S, Cundall M, Armstrong H, Mertens I, Schoofs L, Dow JAT, Davies SA, 2012. The receptor guanylate cyclase Gyc76C and a peptide ligand, NPLP1-VQQ, modulate the innate immune IMD pathway in response to salt stress. Peptides 34, 209–218. [DOI] [PubMed] [Google Scholar]
- Park Y, Filippov V, Gill SS, Adams ME, 2002. Deletion of the ecdysis-triggering hormone gene leads to lethal ecdysis deficiency. Development 129, 493–503. [DOI] [PubMed] [Google Scholar]
- Pietrantonio PV, Jagge C, Taneja-Bageshwar S, Nachman RJ, Barhoumi R, 2005. The mosquito Aedes aegypti (L.) leucokinin receptor is a multiligand receptor for the three Aedes kinins. Insect Mol. Biol 14, 55–67. [DOI] [PubMed] [Google Scholar]
- Pietri JE, Pietri EJ, Potts R, Riehle MA, Luckhart S, 2015. Plasmodium falciparum suppresses the host immune response by inducing the synthesis of insulin-like peptides (ILPs) in the mosquito Anopheles stephensi. Dev. Comp. Immunol 53, 134–144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Potter LR, 2011. Regulation and therapeutic targeting of peptide-activated receptor guanylyl cyclases. Pharmacol. Ther 130, 71–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Predel R, Neupert S, Garczynski SF, Crim JW, Brown MR, Russell WK, Kahnt J, Russell DH, Nachman RJ, 2010. Neuropeptidomics of the mosquito Aedes aegypti. J. Proteome Res 9, 2006–2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raabe M, 1989. Recent Developments in Insect Neurohormones. Plenum Press, New York: 503 pp. [Google Scholar]
- Radford JC, Terhzaz S, Cabrero P, Davies SA, Dow JAT, 2004. Functional characterisation of the Anopheles leucokinins and their cognate G-protein coupled receptor. J. Exp. Biol 207, 4573–4586. [DOI] [PubMed] [Google Scholar]
- Reagan JD, 1994. Expression cloning of an insect diuretic hormone receptor. A member of the calcitonin/secretin receptor family. J. Biol. Chem 269, 9–12. [PubMed] [Google Scholar]
- Reiher W, Shirras C, Kahnt J, Baumeister S, Isaac RE, Wegener C, 2011. Peptidomics and peptide hormone processing in the Drosophila midgut. J. Proteome Res 10, 1881–1892. [DOI] [PubMed] [Google Scholar]
- Rewitz KF, Yamanaka N, Gilbert LI, O’Connor MB, 2009. The insect neuropeptide PTTH activates Receptor Tyrosine Kinase Torso to initiate metamorphosis. Science 326, 1403–1405. [DOI] [PubMed] [Google Scholar]
- Riehle MA, Garczynski SF, Crim JW, Hill CA, Brown MR, 2002. Neuropeptides and peptide hormones in Anopheles gambiae. Science 298, 172–175. [DOI] [PubMed] [Google Scholar]
- Riehle MA, Fan Y, Cao C, Brown MR, 2006. Molecular characterization of insulin-like peptides in the yellow fever mosquito, Aedes aegypti: expression, cellular localization, and phylogeny. Peptides 27, 2547–2560. [DOI] [PubMed] [Google Scholar]
- Robertson HM, Navik JA, Walden KK, Honegger HW, 2007. The bursicon gene in mosquitoes: an unusual example of mRNA trans-splicing. Genetics 176, 1351–1353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roller L, Yamanaka N, Watanabe K, Daubnerova I, Zitnan D, Kataoka H, Tanaka Y, 2008. The unique evolution of neuropeptide genes in the silkworm Bombyx mori. Insect Biochem. Mol. Biol 38, 1147–1157. [DOI] [PubMed] [Google Scholar]
- Sajid W, Kulahin N, Schluckebier G, Ribel U, Henderson HR, Tatar M, Hansen BF, Svendsen AM, Kiselyov VV, Norgaard P, Wahlund PO, Brandt J, Kohanski RA, Andersen AS, De Meyts P, 2011. Structural and biological properties of the Drosophila insulin-like peptide 5 show evolutionary conservation. J. Biol. Chem 286, 661–673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sano H, Nakamura A, Texada MJ, Truman JW, Ishimoto H, Kamilkouchi A, Nibu Y, Kume K, Ida T, Kojima M, 2015. The nutrient-responsive hormone CCHamide-2 controls growth by regulating insulin-like peptides in the brain of Drosophila melanogaster. PLoS Genet 11, e1005209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scholler S, Belmont M, Cazzamali G, Hauser F, Williamson M, Grimmelikhuijzen CJP, 2005. Molecular identification of a myosuppressin receptor from the malaria mosquito Anopheles gambiae. Biochem. Biophys. Res. Commun 327, 29–34. [DOI] [PubMed] [Google Scholar]
- Siju KP, Reifenrath A, Scheiblich H, Neupert S, Predel R, Hansson BS, Schachtner J, Ignell R, 2013. Neuropeptides in the antennal lobe of the yellow fever mosquito, Aedes aegypti. J. Comp. Neurol 522, 592–608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sim C, Denlinger DL, 2009. A shut-down in expression of an insulin-like peptide, ILP-1, halts ovarian maturation during the overwintering diapause of the mosquito Culex pipiens. Insect Mol. Biol 18, 325–332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith W, Rybczynski R, 2012. Prothoracicotropic hormone In: Gilbert LI (Ed.), Insect Endocrinology. Elsevier, New York, pp. 1–62. [Google Scholar]
- Southey BR, Sweedler JV, Rodriguez-Zas SL, 2008. Prediction of neuropeptide cleavage sites in insects. Bioinformatics 24, 815–825. [DOI] [PubMed] [Google Scholar]
- Stanek DM, Pohl J, Crim JW, Brown MR, 2002. Neuropeptide F and its expression in the yellow fever mosquito, Aedes aegypti. Peptides 23, 1367–1378. [DOI] [PubMed] [Google Scholar]
- Stangier J, Hilbich C, Beyreuther K, Keller R, 1999. Unusual cardioactive peptide (CCAP) from pericardial organs of the shore crab Carcinus maenas. Proc. Natl. Acad. Sci. U. S. A 84, 575–579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stracker TH, Thompson S, Grossman GL, Riehle MA, Brown MR, 2002. Characterization of the AeaHP gene and its expression in the mosquito Aedes aegypti (Diptera: Culicidae). J. Med. Entomol 39, 331–342. [DOI] [PubMed] [Google Scholar]
- Szapiro G, Barbour B, 2009. Parasynaptic signalling by fast neurotransmitters: the cerebellar cortex. Neuroscience 162, 644–655. [DOI] [PubMed] [Google Scholar]
- Taghert PH, Veenstra JA, 2003. Drosophila neuropeptide signaling. Adv. Genet 49, 1–65. [DOI] [PubMed] [Google Scholar]
- Taneja-Bageshwar S, Strey A, Isaac RE, Coast GM, Zubrzak P, Pietrantonio PV, Nachman RJ, 2009. Biostable agonists that match or exceed activity of native insect kinins on recombinant arthropod GPCRs. Gen. Comp. Endocrinol 162, 122–128. [DOI] [PubMed] [Google Scholar]
- Truman JW, Riddiford LM, 2002. Endocrine insights into the evolution of metamorphosis in insects. Annu. Rev. Entomol 47, 467–500. [DOI] [PubMed] [Google Scholar]
- Vallejo DM, Juarez-Carrewno S, Bolivar J, Morante J, Dominguez M, 2015. A brain circuit that syntchronizes growth and maturation revealed through Dilp8 binding to Lgr3. Science 350, aac6767. [DOI] [PubMed] [Google Scholar]
- Van der Host DJ, 2003. Insect adipokinetic hormones: release and integration of flight energy metabolism. Comp. Biochem. Physiol. B Biochem. Mol. Biol 136, 217–226. [DOI] [PubMed] [Google Scholar]
- Veenstra JA, 1989. Isolation and structure of corazonin, a cardioactive peptide in the American cockroach. FEBS Lett 250, 231–234. [DOI] [PubMed] [Google Scholar]
- Veenstra JA, 1994. Isolation and identification of three leucokinins from the mosquito Aedes aegypti. Biochem. Biophys. Res. Commun 202, 715–719. [DOI] [PubMed] [Google Scholar]
- Veenstra JA, 1999. Isolation and identification of three RFamide-immunoreactive peptides from the mosquito, Aedes aegypti. Peptides 20, 31–38. [DOI] [PubMed] [Google Scholar]
- Veenstra JA, 2008. Allatostatin C and its paralog allatostatin double C: the arthropod somatostatins. Insect Biochem. Mol. Biol 39, 161–170. [DOI] [PubMed] [Google Scholar]
- Veenstra JA, Costes L, 1999. Isolation and identification of a peptide and its cDNA from the mosquito Aedes aegypti related to Manduca sexta allatotropin. Peptides 20, 1145–1151. [DOI] [PubMed] [Google Scholar]
- Veenstra JA, Lau GW, Agricola HJ, Petzel DH, 1995. Immunohistological localization of regulatory peptides in the midgut of the female mosquito Aedes aegypti. Histochem. Cell Biol 104, 337–347. [DOI] [PubMed] [Google Scholar]
- Veenstra JA, Noriega FG, Graf R, Feyereisen R, 1997a. Identification of three allatostatins and their cDNA from the mosquito Aedes aegypti. Peptides 18, 937–942. [DOI] [PubMed] [Google Scholar]
- Veenstra JA, Pattillo JM, Petzel DH, 1997b. A single cDNA encodes all three Aedes leucokinins, which stimulate both fluid secretion by the Malpighian tubules and hindgut contractions. J. Biol. Chem 272, 10402–10407. [DOI] [PubMed] [Google Scholar]
- Veenstra JA, Rombauts S, Grbic M, 2012. In silico cloning of genes encoding neuropeptides, neurohormones and their putative G-protein coupled receptors in a spider mite. Insect Biochem. Mol. Biol 42, 277–295. [DOI] [PubMed] [Google Scholar]
- Verlinden H, Vleugels R, Zels S, Dillen S, Lenaerts C, Crabbe K, Spit J, Broeck JV, 2014. Receptors for neuronal or endocrine signalling molecules as potential targets for the control of insect pests. Adv. Insect Physiol 46, 167–303. [Google Scholar]
- Vogel KJ, Brown MR, Strand MR, 2013. Phylogenetic investigation of peptide hormone and growth factor receptors in five dipteran genomes. Front. Endocrinol 4, 193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vogel KJ, Brown MR, Strand MR, 2015. Ovary ecdysteroidogenic hormone requires a receptor tyrosine kinase to activate egg formation in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. U. S. A 112, 5057–5062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waterhouse RM, 2015. A maturing understanding of the composition of the insect gene repertoire. Curr. Opin. Insect Sci 7, 15–23. [DOI] [PubMed] [Google Scholar]
- Wegener C, Herbert H, Kahnt J, Bender M, Rhea JM, 2011. Deficiency of prohormone convertase dPC2 (AMONTILLADO) results in impaired production of bioactive neuropeptide hormones in Drosophila. J. Neurochem 118, 581–595. [DOI] [PubMed] [Google Scholar]
- Wen Z, Gulia M, Clark KD, Dhara A, Crim JW, Strand MR, Brown MR, 2010. Two insulin-like peptide family members from the mosquito Aedes aegypti exhibit differential biological and receptor binding activities. Mol. Cell. Endocrinol 328, 47–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- White BH, Ewer J, 2014. Neural and hormonal control of postecdysial behaviors in insects. Annu. Rev. Entomol 59, 363–381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiegmann BM, Yeates DK, Thorne JL, Kishino H, 2003. Time flies, a new molecular time-scale for brachyceran fly evolution without a clock. Syst. Biol 52, 745–756. [PubMed] [Google Scholar]
- Woodhead AP, Stay B, Seidel SL, Khan MA, Tobe SS, 1989. Primary structure of four allatostains: neuropeptide inhibitors of juvenile hormone synthesis. Proc. Natl. Acad. Sci. U. S. A 86, 5997–6001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zandawala M, 2012. Calcitonin-like diuretic hormones in insects. Insect Biochem. Mol. Biol 42, 816–825. [DOI] [PubMed] [Google Scholar]
- Zhang Q, Denlinger DL, 2011. Molecular structure of the prothoracicotropic hormone gene in the northern house mosquito, Culex pipiens, and its expression analysis in association with diapause and blood feeding. Insect Mol. Biol 20, 201–213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang H, Liu J, Li CR, Momen B, Kohanski RA, Pick L, 2009. Deletion of Drosophila insulin-like peptides causes growth defects and metabolic abnormalities. Proc. Natl. Acad. Sci. U. S. A 106, 19617–19622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zitnan D, Kingan TG, Hermesman JL, Adams ME, 1996. Identification of ecdysistriggering hormone from an epitracheal endocrine system. Science 271, 88–91. [DOI] [PubMed] [Google Scholar]
- Zitnan D, Spalovska ZI, Takac P, Park Y, Adams ME, 2003. Conservation of ecdysis-triggering hormone signalling in insects. J. Exp. Biol 206, 1275–1289. [DOI] [PubMed] [Google Scholar]