

Abstract
Background:
The capsulo-osseous layer (COL), short lateral ligament, mid–third lateral capsular ligament, lateral capsular ligament, and anterolateral ligament (ALL) are terms that have been used interchangeably to describe what is probably the same structure. This has resulted in confusion regarding the anatomy and function of the anterolateral complex of the knee and its relation to the distal iliotibial band (ITB).
Purpose:
To characterize the macroscopic anatomy of the anterolateral complex of the knee, in particular the femoral condylar attachment of the distal ITB. We identified a specific and consistent anatomic structure that has not been accurately described previously; it connects the deep surface of the ITB to the condylar area and is distinct from the ALL, COL, and Kaplan fibers.
Study Design:
Descriptive laboratory study.
Methods:
Sixteen fresh-frozen human cadaveric knees were used to study the anterolateral complex of the knee. Standardized dissections were performed that included qualitative and quantitative assessments of the anatomy through both anterior (n = 5) and posterior (n = 11) approaches.
Results:
The femoral condylar attachment of the distal ITB was not reliably identified by anterior dissection but was in all posterior dissections. A distinct anatomic structure, hereafter termed the “condylar strap” (CS), was identified between the femur and the lateral gastrocnemius on one side and the deep surface of the ITB on the other, in all posteriorly dissected specimens. The structure had a mean thickness of 0.88 mm, and its femoral insertion was located between the distal Kaplan fibers and the epicondyle. The proximal femoral attachment of the structure had a mean width of 15.82 mm, and the width of the distal insertion of the structure on the ITB was 13.27 mm. The mean length of the structure was 26.33 mm on its distal border and 21.88 mm on its proximal border. The qualitative evaluation of behavior in internal rotation revealed that this anatomic structure became tensioned and created a tenodesis effect on the ITB.
Conclusion:
There is a consistent structure that attaches to the deep ITB and the femoral epicondylar area. The orientation of fibers suggests that it may have a role in anterolateral knee stability.
Clinical Relevance:
This new anatomic description may help surgeons to optimize technical aspects of lateral extra-articular procedures in cases of anterolateral knee laxity.
Keywords: distal iliotibial band, distal iliotibial tract, anterolateral ligament, capsulo-osseous layer, Lemaire procedure
During the past 5 decades, the Lemaire procedure has been performed in conjunction with intra-articular anterior cruciate ligament (ACL) reconstruction to control anterolateral laxity of the knee.14 In the original technique, an 18 cm–long by 1 cm–wide strip of the iliotibial band (ITB) that remained attached to the Gerdy tubercle was harvested. The graft was passed deep to the fibular collateral ligament (FCL), through a periosteal bridge, and then through a femoral tunnel located proximal to the lateral femoral epicondyle. Then, the graft came back again under the FCL to be sutured to itself or fixed in a tibial tunnel. The graft was fixed in external rotation between 30° and 45° of flexion. This technique as well as other types of lateral extra-articular procedures, including anterolateral ligament (ALL) reconstruction, have shown good results with respect to abolishing the pivot shift and reducing ACL graft rupture rates.1,5,10,16,20 Despite these clinical results, the literature demonstrates that there is still a lack of consensus concerning the precise anatomy and biomechanics of the anterolateral structures of the knee.19
Vincent et al24 reported that several terms, including capsulo-osseous layer (COL), short lateral ligament, mid–third lateral capsular ligament, lateral capsular ligament, lateral femorotibial ligament,17 ligamentum tractotibiale,7 and ALL,2,8 have been used interchangeably in the literature to describe what is probably the same structure. Porrino et al18 highlighted that previous reports are also inconsistent with respect to the relationship of this structure with the ITB. This lack of consistency in nomenclature has led to confusion in anatomic and biomechanical studies, in which the use of nonstandardized dissection protocols has resulted in different interpretations about anatomy and function.2,4,12 Furthermore, one of the limitations of previous anatomic studies is that they have typically used a single anterior surgical approach.9,13 This restriction to 1 type of approach limits the overall perspective that can be gained and therefore risks interpretation bias and also the inadvertent creation of artificial tissue planes around tightly confluent structures.
The history of the description of the ITB relies on some landmark papers. Early anatomists described the fascia lata as an important muscle of the lower limb, but an accurate description of the distal insertion of the fascia lata was only reported during the past century, and some inaccuracies and controversies are still present. Vesalius22 described the fascia lata. He grouped it with other muscles and called it the sixth muscle of the tibia, but probably the most thorough and extensive work on the anatomy and functions of the ITB was written in 1843 by Jacques Maissiat.11 The publication of his monograph was responsible for the widespread adoption of the eponymous name “band of Maissiat.”11 In the modern literature, the terms “iliotibial band” and “iliotibial tract” are commonly used.
In 1958, Kaplan11 described the iliotibial tract as intimately connected with the intermuscular septum and the linea aspera from the greater trochanter to the supracondylar tubercle of the lateral condyle of the femur. He presented the iliotibial tract as free of bony attachments/connections except at the level of the upper portions of the femoral condyle and also at the Gerdy tubercle. In this landmark article, it was already clear that the ITB made a connection between the area of the lateral epicondyle and the Gerdy tubercle. However, it is interesting to mention that the descriptions of what would later be called the Kaplan fibers,21 as originally reported by Kaplan11 himself, were quite vague. One of the illustrations in his paper showed a connection between the ITB and the supracondylar area of the femur, oriented from proximal-lateral to distal-medial (Figure 1).
Figure 1.

Original drawing from Kaplan11 showing the connection between the iliotibial band and the supracondylar area of the femur, oriented from proximal-lateral to distal-medial (reprinted with permission).
The aim of this study was to characterize the macroscopic anatomy of the anterolateral complex of the knee, in particular the femoral condylar attachment of the distal ITB, by using 2 separate standardized dissection protocols. In preliminary dissections, we had found that a specific and consistent anatomic structure, which has not been accurately described previously, connects the deep surface of the ITB to the condylar area and is distinct from the ALL, the COL, and the Kaplan fibers. We hypothesized that this structure, which we called the “condylar strap” (CS), would be consistently found in human knee specimens.
Methods
The methodology of the current study comprised 2 main parts. For both parts, nonpaired, fresh-frozen human cadaveric knees without evidence of previous injuries or surgery were used to perform a qualitative and quantitative study. Three authors (P.L., R.S., A.C.) conducted the dissections together. These were performed in a standardized manner, commencing with removal of the skin on the lateral side of the knee to create a large rectangular window.
In the first part of the study, 5 specimens were studied (mean age, 79 years [range, 65-87 years]). The technique was performed in accordance with previous descriptions in the literature.3,6,11,15 A transverse incision was made in the ITB approximately 10 cm proximal to the knee joint. An anterior longitudinal incision of the ITB was made to progressively reflect distally and posteriorly the ITB. As we flipped distally and posteriorly the ITB, we exposed progressively the vastus lateralis muscle and the deeper attachments of the ITB.
In the second part of the study, 11 specimens were studied (mean age, 82 years [range, 71-87 years]). In contrast to the previous dissection, the ITB was approached from posterior (Figure 2A). To do so, the space between the biceps and the ITB was approached. The biceps muscle was retracted posteriorly, allowing excellent visualization of the intermuscular septum. A Kelly forceps was passed between the distal part of the ITB and the capsular structures. The correct plane was confirmed by the absence of resistance to instrument insertion. Then, the Kelly forceps was moved from inferior (just proximal to the Gerdy tubercle attachment of the ITB) to superior, where the instrument was stopped by a robust structure, hereafter referred to as the CS of the ITB, connecting the deep part of the ITB and the femoral epicondylar area (Figure 2B). At this stage of the dissection, an internal rotational force was applied between 30° and 60° of knee flexion, and a qualitative assessment of tension in the structure was conducted by pulling on it with the forceps (Figure 2C).
Figure 2.

Right knee specimen. (A) Posterior approach: the space between the biceps and the iliotibial band (ITB) was approached. The biceps muscle was retracted posteriorly, allowing excellent visualization of the intermuscular septum. (B) The Kelly forceps was moved from inferior (just proximal to the Gerdy tubercle attachment of the ITB) to superior, where the instrument was stopped by a robust structure (asterisk) connecting the deep part of the ITB and the femoral epicondylar area. (C) An internal rotational force was applied between 30° and 60° of knee flexion, and a qualitative assessment of tension in the structure was conducted by pulling on it with the forceps. (D) Progressive and careful dissection was performed to identify the septum and the Kaplan fibers. Proximal and distal Kaplan fibers (PKF and DKF, respectively) were always found proximal to the identified anatomic structure (asterisk).
Once the structure attaching to the deep part of the ITB and the femoral epicondylar area was clearly identified, the next steps were to describe its precise relation to the anatomic structures already described in the literature. Fatty tissue was always encountered distal to the septum. This was excised cautiously. Progressive and careful dissection was performed to identify the septum and the proximal and distal Kaplan fibers (Figure 2D); the terminology of proximal and distal Kaplan fibers was used according to the study of Godin et al.6
Only after clear identification of these previously described anatomic structures was the distal attachment of the ITB detached and redirected proximally, allowing excellent visualization of the CS and a thorough qualitative assessment. Throughout the dissection, a Vernier caliper with an accuracy of 0.1 mm was used to define and record the anatomy of this structure and its relationships to known anatomic structures.
The caliper was used to measure the following aspects of the CS: width of the proximal attachment, thickness (measured 1 cm from the femoral attachment), length of the proximal and distal borders, and width of the distal attachment to the ITB. Several distances were measured to precisely define the relationship with known anatomic structures of the lateral part of the knee. This included distances between the following: distal Kaplan fibers (proximal, middle, and distal) and the proximal attachment of the structure, the lateral epicondyle and the proximal and distal attachments of the structure on the femur, the middle of the Gerdy tubercle attachment of the ITB and the proximal and distal attachments of the structure on the femur, and the distal attachment of the structure on the ITB (anterior border and posterior border) and the ITB attachment to the Gerdy tubercle. Finally, to investigate a potential correlation between the size of the structure and the size of the joint, the following distances were calculated: width of the femoral attachment of distal Kaplan fibers, distance between the lateral epicondyle and the middle of the Gerdy tubercle, and the surface of the distal attachment of the ITB on the Gerdy tubercle (calculated using the size of anterior, posterior, and proximal borders of the attachment to the Gerdy tubercle). The measurements were performed with the knee in extension.
Statistical Analysis
All data were assessed for normality using the Shapiro-Wilk test before analysis. Pearson correlation coefficients (r) were used to analyze the association between the width of distal Kaplan fibers, the distance between the lateral epicondyle and the middle of the Gerdy tubercle, and the Gerdy tubercle surface as independent variables and the different anatomic characteristics of the structure. Statistical significance was set at P < .05. Statistical analyses were carried out using SPSS 22.0 for Mac OS X (IBM).
Results
In all 5 anterior dissections, it was not possible to accurately identify the structure that attached to the distal ITB and the femoral epicondylar region. In contrast, this structure was easily and reliably identified in all posterior dissections.
Qualitative Anatomy
The distinct flat bundle of fibers of the CS was identified in all cadaveric specimens. These fibers coursed from proximal and femoral to distal and deep into the ITB. The femoral attachment was located on an area sited proximal to the epicondyle, just proximal and anterior to the femoral insertion of the lateral gastrocnemius tendon (Figure 3A). At this location, some expansions were noted in all knees. These expansions were extended to the lateral gastrocnemius muscle and the plantaris tendons (Figure 3B).
Figure 3.

Schematic representation of the distal femur of a right knee demonstrating (A) the insertion sites of the condylar strap (CS) related to the known anatomic structures. (B) The CS (asterisk) connecting the deep portion of the iliotibial band (ITB) and the lateral epicondylar area. ALL, anterolateral ligament; DKF, distal Kaplan fibers; FCL, fibular collateral ligament; LE, lateral epicondyle; LGT, lateral gastrocnemius tendon; PKF, proximal Kaplan fibers; PLT, popliteus tendon.
The distal attachment of the CS was located on the deep surface of the ITB. No direct connection between this structure and the Gerdy tubercle was identified. The superior genicular artery was identified as previously described. This was consistently located proximal to the CS (Figure 4).
Figure 4.
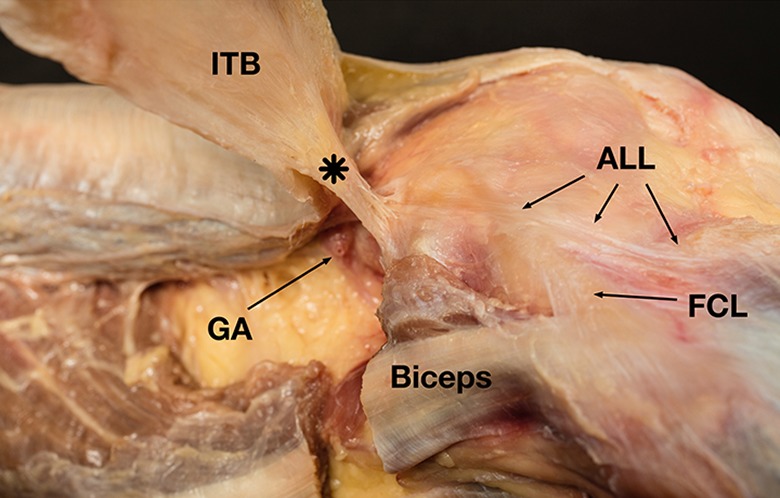
Right knee. The superior genicular artery (GA) was consistently located proximal to the condylar strap (asterisk). The iliotibial band (ITB) was detached from the Gerdy tubercle and redirected proximally. ALL, anterolateral ligament; FCL, fibular collateral ligament.
The proximal and distal Kaplan fibers were consistently found in our study, according to the study of Godin et al.6 The orientation of the Kaplan fibers was from lateral-proximal (on the deep surface of the ITB) to medial-distal (on the femur). In all specimens, the qualitative evaluation of behavior in tibial internal rotation revealed that the CS became tensioned and created a “tenodesis effect” on the ITB (see the online Video Supplement for this qualitative evaluation).
In every specimen, the ALL could be identified.2,8 As the purpose of our study was not focused on the ALL, only a qualitative evaluation of this ligament was performed. In all specimens, the femoral insertion of the CS was found proximal to the femoral attachment of the ALL. In some cases, the proximal fibers of the ALL were inserted closely to the femoral insertion of the CS.
Quantitative Anatomy
In all 11 specimens in which the ITB was approached posteriorly, a clear anatomic structure, the CS, was identified between the femur and the lateral gastrocnemius on one side and the deep surface of the ITB on the other side. Systematic measurements were performed. Descriptive data are presented as mean ± SD (Table 1).
TABLE 1.
Measurements of the Identified CS, Its Distance to Other Known Structures, and Some Anatomic Variablesa
| Specimen Profile | CS Measurements, mm | CS Attachments, mm | Anatomic Independent Variables, mm | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Specimen | Side | Sex | Donor Height, in | Thickness | Length of Distal Border | Length of Proximal Border | Width of Distal Insertion on ITB | Width of Proximal Attachment | Middle of DKF to Proximal Attachment | Proximal End of DKF to Proximal Attachment | Distal End of DKF to Proximal Attachment | Proximal Attachment to Lateral Epicondyle | Distal Attachment to Lateral Epicondyle | Proximal Attachment to Middle of Gerdy Tubercle | Distal Attachment to Middle of Gerdy Tubercle | Distance Between Posterior Distal Attachment on ITB and Gerdy Tubercle | Distance Between Anterior Distal Attachment on ITB and Gerdy Tubercle | Width of DKF | Distance of Lateral Epicondyle to Middle of Gerdy Tubercle | Anterior Border of Gerdy Tubercle | Posterior Border of Gerdy Tubercle | Proximal Border of Gerdy Tubercle |
| 1 | R | M | 72.00 | 0.70 | 31.20 | 21.00 | 13.00 | 13.60 | 6.00 | 11.90 | –8.50 | 21.90 | 20.20 | 69.40 | 64.20 | 50.50 | 65.50 | 14.10 | 57.20 | 17.30 | 22.20 | 18.50 |
| 2 | R | M | 73.00 | 0.80 | 30.70 | 23.90 | 9.60 | 13.50 | –2.90 | 4.90 | –10.40 | 26.20 | 18.60 | 68.70 | 59.70 | 59.30 | 62.60 | 11.30 | 45.30 | 16.30 | 15.40 | 14.10 |
| 3 | R | F | 61.00 | 1.00 | 21.60 | 12.30 | 7.60 | 15.60 | 10.40 | 14.80 | 5.10 | 17.20 | 13.70 | 56.40 | 45.20 | 42.20 | 53.90 | 10.50 | 41.50 | 12.10 | 14.30 | 13.50 |
| 4 | L | M | 72.00 | 0.70 | 32.00 | 33.70 | 11.10 | 15.40 | 8.10 | 13.80 | 5.20 | 17.40 | 6.70 | 68.80 | 57.30 | 31.90 | 56.90 | 12.30 | 57.30 | 16.30 | 17.20 | 16.80 |
| 5 | L | F | 64.00 | 0.60 | 23.40 | 17.50 | 13.80 | 14.10 | 3.20 | 5.50 | –5.60 | 18.60 | 11.40 | 55.70 | 43.30 | 22.50 | 55.20 | 12.90 | 40.40 | 14.70 | 17.40 | 16.10 |
| 6 | R | F | 68.00 | 0.80 | 29.40 | 16.10 | 10.80 | 13.20 | 12.50 | 15.40 | 10.60 | 18.70 | 19.10 | 60.30 | 52.40 | 46.30 | 63.40 | 11.40 | 48.60 | 15.10 | 18.20 | 16.70 |
| 7 | L | M | 67.00 | 1.90 | 20.10 | 26.80 | 13.20 | 20.70 | 17.50 | 22.80 | 15.00 | 22.70 | 13.80 | 68.00 | 59.20 | 45.70 | 56.20 | 9.00 | 50.20 | 15.10 | 18.90 | 17.20 |
| 8 | R | M | 71.00 | 1.30 | 33.50 | 24.00 | 19.50 | 26.50 | 18.90 | 24.70 | 14.10 | 30.30 | 9.50 | 74.20 | 53.20 | 36.00 | 58.30 | 12.20 | 51.50 | 14.40 | 16.10 | 17.80 |
| 9 | R | M | 71.00 | 0.90 | 19.30 | 21.50 | 19.60 | 20.20 | 5.10 | 9.70 | –8.20 | 13.60 | 18.40 | 64.50 | 50.60 | 40.60 | 62.60 | 12.60 | 55.30 | 13.80 | 16.40 | 16.00 |
| 10 | L | F | 60.00 | 0.40 | 27.40 | 19.90 | 13.50 | 13.20 | 1.50 | 3.70 | 0.00 | 20.40 | 13.40 | 56.60 | 46.50 | 42.20 | 54.70 | 5.30 | 37.70 | 13.40 | 17.50 | 15.80 |
| 11 | R | F | 60.00 | 0.60 | 21.00 | 24.00 | 14.30 | 8.00 | 8.60 | 13.30 | 4.10 | 15.90 | 12.10 | 52.90 | 47.20 | 30.50 | 52.50 | 12.00 | 40.80 | 10.70 | 12.60 | 12.60 |
| Mean ± SD | 67.18 ± 5.12 | 0.88 ± 0.41 | 26.33 ± 5.33 | 21.88 ± 5.71 | 13.27 ± 3.70 | 15.82 ± 4.95 | 8.08 ± 6.57 | 12.77 ± 6.80 | 1.95 ± 9.18 | 20.26 ± 4.80 | 14.26 ± 4.34 | 63.23 ± 7.12 | 52.62 ± 6.80 | 40.70 ± 10.19 | 58.35 ± 4.44 | 11.24 ± 2.37 | 47.80 ± 7.12 | 14.47 ± 1.92 | 16.93 ± 2.51 | 15.92 ± 1.83 | ||
| Range | 60.00 to 73.00 | 0.40 to 1.90 | 19.30 to 33.50 | 12.30 to 33.70 | 7.60 to 19.60 | 8.00 to 26.50 | –2.90 to 18.90 | 3.70 to 24.70 | –10.40 to 15.00 | 13.60 to 30.30 | 6.70 to 20.20 | 52.90 to 74.20 | 43.30 to 64.20 | 22.50 to 59.30 | 52.50 to 65.50 | 5.30 to 14.10 | 37.70 to 57.30 | 10.70 to 17.30 | 12.60 to 22.20 | 12.60 to 18.50 | ||
aCS, condylar strap; DKF, distal Kaplan fibers; F, female; ITB, iliotibial band; L, left; M, male; R, right.
The mean thickness of the CS was 0.88 mm. The proximal attachment of the CS on the femur had a mean width of 15.82 mm, and the width of the distal insertion of the CS on the ITB was 13.27 mm. The mean length of the CS was 26.33 mm on its distal border and 21.88 mm on its proximal border.
The femoral insertion of the CS was located between the distal Kaplan fibers (the proximal insertion was located at a mean distance of 8.08 mm from the center of the distal Kaplan fibers) and the epicondyle (the distal end of the CS was a mean of 14.26 mm from the epicondyle). In these specimens, the epicondyle (with the knee in extension) was located a mean of 47.80 mm from the center of the Gerdy tubercle. In comparison, the distance between the femoral attachment of the CS and the center of the Gerdy tubercle was 63.23 mm and 52.62 mm on the proximal and distal ends of the CS, respectively. The distance between the distal attachment of the CS to the ITB and the center of the Gerdy tubercle was 40.70 mm on the posterior border and 58.35 mm on the anterior border.
No significant correlations were detected between the width of the distal Kaplan fibers and any of the different anatomic characteristics of the CS. However, significant correlations were found between the distance from the lateral epicondyle to the middle of the Gerdy tubercle and the Gerdy tubercle surface and 3 anatomic characteristics of the CS (Table 2).
TABLE 2.
Correlation Coefficients and P Values of the Association Between Independent Variables and Different Anatomic Characteristics of the CSa
| Gerdy Tubercle Surface | Proximal Attachment of CS in Middle of Gerdy Tubercle | Distal Attachment of CS in Middle of Gerdy Tubercle | Distal Anterior ITB | |
|---|---|---|---|---|
| Distance of lateral epicondyle to middle of Gerdy tubercle | ||||
| Pearson correlation coefficient (r) | 0.666b | 0.779b | 0.731b | 0.642b |
| P value | .025 | .005 | .011 | .033 |
| Gerdy tubercle surface | ||||
| Pearson correlation coefficient (r) | 0.627b | 0.700b | 0.646b | |
| P value | .039 | .017 | .032 |
aCS, condylar strap; ITB, iliotibial band.
bSignificant correlation (P < .05).
Discussion
The main findings of this study are that the anterior9,13 and posterior approaches provide very different perspectives of the anatomy of the distal ITB and that posterior dissection reliably demonstrated that there is a distinct anatomic structure, termed the CS, that attaches the deep surface of the ITB to the lateral part of the distal femur and the lateral gastrocnemius tendon. This CS is not well visualized from anterior, as it is disconnected with the anterior approach. The qualitative “biomechanical” evaluation revealed that the CS became tensioned and created a “tenodesis effect” on the ITB. To our knowledge, this structure has not previously been described in this way, and this finding is likely attributable to the use of a posterior approach.
In contrast to the findings of the current study, Terry et al,21 in 1986, described a COL, localized under the deep layer of the ITB. The authors stated that “posteriorly, the capsulo-osseous layer forms a superficial arcuate whose proximal origin is continuous with fascia covering the plantaris and lateral gastrocnemius and whose distal tibial insertion is just posterior to Gerdy’s tubercle on the lateral tibial tuberosity.” In the current study, the CS that was identified was attached to the lateral condyle and had some expansions on the lateral gastrocnemius. Therefore, it is probably the case that the same anatomic structure is described in both studies, at least with respect to the proximal attachment. However, in the current study, a direct attachment to the tibia was not identified. Instead, the fibers were attached to the deep surface of the ITB. Furthermore, in all specimens, it was possible to measure the distance between the distal attachment of the fibers on the deep surface of the ITB and the Gerdy tubercle. The mean distance between the distal attachment of the CS on the ITB and the Gerdy tubercle was 40.70 mm on the posterior border and 58.35 mm on the anterior border. The current study demonstrates that these fibers blend with the deep surface of the ITB with an orientation that suggests that they contribute to the distal part of the ITB along with contributions from the superficial structures approaching from more proximally.
At the level of the CS, it was possible to separate the 2 main layers of the ITB, confirming the previous description of ITB layers. However, it was not possible to separate these layers in the last centimeters of the ITB, close to the Gerdy tubercle. Therefore, the distal part of the ITB should be considered as a 1-layer entity, created by the unity of different layers and more proximal structures, including the CS with a condylar attachment identified in the current study. This is something that can be easily confirmed while performing a modified Lemaire procedure or any lateral tenodesis with the ITB.14 The layers are often visible during ITB harvesting on the proximal part of the graft but never in the last few centimeters, close to the tibial attachment.
Anatomically, it is hard to reconcile the reason why Terry et al21 and other authors have described this COL attached to the Gerdy tubercle. In their article, Terry et al21 did not describe any structure like the one described later, the ALL,2,8,24 but they mentioned that the COL “functions as a medial retaining wall for the deep layer and allows the deep layer to extend its more anterior and proximal bony origin down onto the lateral capsule. It thus acts as an anterolateral ligament of the knee.” In the current study, the structure of interest did not have any capsular attachment, as its distal attachment was to the deep aspect of the ITB. In our dissections, we noticed that the fibers blended in with the ITB, but we were not able to follow them into the Gerdy tubercle. It is possible that a microscopic assessment would yield more information on their terminal insertion. Furthermore, in every case, the ALL was identified as a distinct and separate entity, as previously described by Claes et al.2 Specifically, there was no connection between the ALL and the deep surface of the ITB. This allows clarification that the ALL and the CS identified are clearly separated. However, it is our opinion that the COL defined by Terry et al21 corresponds to a description combining these 2 structures. In fact, it is interesting that Terry et al21 mentioned that the COL “acts as an anterolateral ligament of the knee.” Nonetheless, these 2 anatomic structures are distinct and should not be described as a unique bundle. Therefore, it is proposed that the term “capsulo-osseous layer” should be abandoned, principally because it causes confusion by suggesting an anatomic association (capsular attachment) that has not been found to be present and because the terms “anterolateral ligament” and “condylar attachment of the distal ITB” clarify that these are anatomically discrete structures.
In the Terry et al21 study, dissection was performed using an anterior approach. Upon initiating our anatomic study, 5 knee specimens were anteriorly dissected, assuming that a deep structure connects the ITB and the epicondylar area of the femur. Furthermore, our first approach was anterior, in keeping with the majority of the previously published literature on this topic.9,13 However, it was always difficult to correctly dissect the structure that we could easily feel by finger, in the epicondylar region, through the ITB, when the tibia was brought into internal rotation.
In our experience, the anterior and posterior approaches allowed a very different view of the deep anatomic structures. The posterior approach was associated with significant advantages in terms of visualization because the proximal attachment of the CS is spread over the condyle and the posterior surface of the lateral gastrocnemius. The anterior approach allowed only a partial view of the bony attachment and no view of the gastrocnemius expansions.9,13
Moreover, during an anterior approach, posterior retraction of the ITB artificially creates a fold that forms continuity between the condyle and the Gerdy tubercle, through the ITB, with an arch-like appearance. Even if we agree on some points with the previous descriptions, this aspect of an arch is a result of the anterior dissection that was performed.
Vieira et al,23 in 2007, conducted an anatomic study with methodology similar to Terry et al21 and confirmed the same conclusions. The photographs of Vieira et al’s23 anatomic dissection mention the “capsular-osseous layer of the ITB with its origin at the supra-epicondylar region of the femur, the arched direction of its fibers, and the insertion lateral to Gerdy’s tubercle.” On the basis of the current study, it is clear that the distal attachment of the COL on the Gerdy tubercle is actually the attachment of the ITB. Vieira et al23 concluded that the COL can be considered as a true ALL of the knee and that it forms, “in conjunction with the ACL, the figure of an inverted ‘U’ or a horseshoe shape, being the ACL the medium portion of the ‘U’ and the capsulo-osseous layer of the ITB its lateral portion. Although it is clear that some lateral structures can play a role, along with the ACL, to control anterolateral rotatory laxity, the anatomic dissections performed in the current study did not demonstrate any connection between the proximal attachment of the CS that we identified and the ACL or even any posterior structure that could connect them together. Therefore, if the 2 structures are “working” together, it is in a separate anatomic manner. It is important to clarify this point to avoid any overinterpretation for surgical application. More specifically, it should be highlighted that the Macintosh concept for surgical reconstruction in cases of anterolateral laxity should not be considered as strict anatomic reconstruction of an existing inverted U-shaped structure, even if it has demonstrated some clinical efficiency.
Lobenhoffer et al,15 in 1987, presented an anatomic study on distal femoral fixation of the iliotibial tract; 100 knee joints were dissected. They concluded that there is a fiber bundle system with 3 main parts: (1) the supracondylar bundle oriented from proximal-lateral to distal-medial and fixed to the supracondylar area of the femur, (2) the fibers near to the septum with a transverse course between the superficial tract and the dorsolateral femur, and (3) the retrograde tracts that connect the Gerdy tubercle with the dorsolateral femur and form an arc, bridging the knee joint. All the dissections were performed from an anterior approach. What the authors described as the retrograde fiber tract appears to correspond directly to the COL previously described by Terry et al.21 The description by Lobenhoffer et al15 is more detailed concerning the proximal insertion, with 2 types according to their position beside the septum. It is remarkable that they described this structure as either “septum-like” (78% of specimens) or “ligament-like” (22% of specimens).15 It confirms the findings observed in the current study with a significant structure connecting the deep surface of the ITB and the lateral femur. Interestingly, the authors did not report an attachment to the lateral gastrocnemius. In contrast, Godin et al6 reported that the proximal aspect of the COL was associated with the fascia surrounding the lateral gastrocnemius tendon, further supporting the view that all authors are describing the same anatomic structure.
It is important to highlight that the connection between the CS and the proximal insertion of the lateral gastrocnemius tendon is difficult to interpret from this anatomic study. Through the connection with this muscle, a dynamic control of this CS and a proprioceptive role can be hypothesized, however this anatomic study cannot present any conclusion on this topic, and further study should be performed to evaluate this concept further.
Lobenhoffer et al15 reported that the retrograde fibers connect the Gerdy tubercle with the dorsolateral femur and form an arc, bridging the knee joint. Although we agree with this statement from a functional perspective, because of the connection between the femur and the Gerdy tubercle, it is our opinion that this occurs through the distal ITB because the fibers are attached directly to the ITB. Interestingly, in their published illustrations, Lobenhoffer et al15 show the distal attachment of these retrograde fibers on the deep ITB and not on the Gerdy tubercle. We believe that their interpretation and the lack of information concerning the posterior attachment of this structure, especially on the lateral gastrocnemius, are the result of the anterior approach of their dissection. However, it should be noted that Godin et al6 reported 2 distinct deep layers of the distal ITB, which they called the proximal and distal Kaplan fibers. Their findings are consistent with those of the current study. The distal Kaplan fibers were always proximal to the CS that we identified. Godin et al6 suggested that there may be a potential action of the proximal and distal Kaplan fibers in internal rotation control of the tibia, but it should be noted that their assessment was performed only after dissecting the ITB off the tibia. Furthermore, in the current study, we identified that the Kaplan fibers are oriented from proximal-lateral (at the deep surface of the ITB) to medial-distal (on the femur) in all specimens. This seemingly precludes an important role in the control of tibial internal rotation. In contrast, we found that the fiber orientation of the identified CS was from proximal-medial to distal-lateral, making this structure more likely to be important in the control of anterolateral laxity. This concept was also supported by the fact that it was tensioned by application of an internal rotation force in all degrees of flexion evaluated, although clearly a more sophisticated biomechanical study would be required to validate these findings.
Claes et al,2 in their anatomic description of the ALL, mentioned that both the “deep layer” and the COL should not be confused with the ALL. We are in complete agreement with this statement. In all of the specimens, the ALL could be clearly identified as a bundle superficial to the lateral collateral ligament, with a proximal insertion quite close to the bony insertion of the CS identified. Vincent et al24 reported their observation of the lateral anatomy during total knee arthroplasty procedures. They described “a relatively consistent structure in the lateral knee, linking the lateral femoral condyle, the lateral meniscus, and the lateral tibial plateau.” This structure seems to correspond to the ALL and not the structure that we have dissected because in the current study, no attachment of the CS to the capsule or the lateral meniscus was identified.
We believe that the control of anterolateral knee laxity is probably the result of several anatomic structures in addition to the ACL. This study showed that, besides the ALL, there is another anatomic structure with a proximal condylar insertion and a distal insertion on the deep surface of the ITB. This is the reason why we termed this anatomic structure the “condylar strap of the iliotibial band.” Its orientation from proximal-medial to distal-lateral suggests that this structure may have a role in the control of internal rotation and anterior translation of the lateral tibial plateau. A potentially important biomechanical role was further suggested by the qualitative assessment of the CS. However, it was not possible to assess the strength of internal rotation control, especially in comparison with other structures such as the ALL.
The limitations of this study include the small number of specimens evaluated. Despite this, the CS was identified in all 11 specimens that were evaluated with a posterior approach. An anatomic study with a higher number of specimens and microscopic dissection could help to confirm our results and better define the terminal insertion of the CS fibers. The age of the specimens was relatively high (mean age, 82 years [range, 71-87 years]), suggesting that the quality of the tissue is lower than in a younger population. However, we assume that if it was possible to accurately identify the CS in all of the included specimens evaluated with a posterior approach, its identification in younger specimens would be even easier. Our main limitation was the absence of a quantitative biomechanical evaluation. We consider our anatomic study as a first step to identify a structure that was either underestimated or understudied previously in comparison with other anatomic structures. We acknowledge that the next step is to perform a biomechanical study assessing the exact role of the CS.
Conclusion
The distal ITB attaches to the Gerdy tubercle. Proximal to this, there is a consistent structure that attaches to the deep ITB and the femoral epicondylar area, which we have named the condylar strap of the ITB. This structure is clearly distinct from the ALL and the Kaplan fibers. The COL as has been previously described probably includes both the CS and the ALL. The orientation of fibers and qualitative assessment of behavior in internal rotation suggest that the CS may have a role in anterolateral knee rotatory stability, but further study is required to evaluate a potentially important biomechanical role.
A Video Supplement for this article is available at http://journals.sagepub.com/doi/suppl/10.1177/2325967118818064.
Acknowledgment
The authors thank Montassar Taben for his contribution to statistical analyses and Khloud Sebak for her contribution during cadaveric dissections.
Footnotes
One or more of the authors has declared the following potential conflict of interest or source of funding: P.L. receives royalties from Springer-Verlag and Medacta, is a consultant for Smith & Nephew, and receives institutional support from Smith & Nephew. A.S. is a consultant for Arthrex and Smith & Nephew. B.S.-C. is a consultant for, receives research support from, and receives royalties from Arthrex. AOSSM checks author disclosures against the Open Payments Database (OPD). AOSSM has not conducted an independent investigation on the OPD and disclaims any liability or responsibility relating thereto.
Ethical approval was not sought for the present study.
References
- 1. Cerciello S, Batailler C, Darwich N, Neyret P. Extra-articular tenodesis in combination with anterior cruciate ligament reconstruction: an overview. Clin Sports Med. 2018;37(1):87–100. [DOI] [PubMed] [Google Scholar]
- 2. Claes S, Vereecke E, Maes M, et al. Anatomy of the anterolateral ligament of the knee. J Anat. 2013;223(4):321–328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Daggett M, Busch K, Sonnery-Cottet B. Surgical dissection of the anterolateral ligament. Arthrosc Tech. 2016;5(1):e185–e188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Daggett M, Claes S, Helito CP, et al. The role of the anterolateral structures and the ACL in controlling laxity of the intact and ACL-deficient knee: letter to the editor. Am J Sports Med. 2016;44(4):NP14–NP15. [DOI] [PubMed] [Google Scholar]
- 5. Ferretti A, Monaco E, Ponzo A, et al. Combined intra-articular and extra-articular reconstruction in anterior cruciate ligament deficient knee: 25 years later. Arthroscopy. 2016;32(10):2039–2047. [DOI] [PubMed] [Google Scholar]
- 6. Godin JA, Chahla J, Moatshe G, et al. A comprehensive reanalysis of the distal iliotibial band: quantitative anatomy, radiographic markers, and biomechanical properties. Am J Sports Med. 2017;45(11):2595–2603. [DOI] [PubMed] [Google Scholar]
- 7. Hassler H, Jakob RP. [On the cause of the anterolateral instability of the knee joint: a study on 20 cadaver knee joints with special regard to the tractus iliotibialis (authors’ transl)]. Arch Orthop Trauma Surg. 1981;98(1):45–50. [DOI] [PubMed] [Google Scholar]
- 8. Helito CP, Demange MK, Bonadio MB, et al. Anatomy and histology of the knee anterolateral ligament. Orthop J Sports Med. 2013;1(7):2325967113513546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Herbst E, Albers M, Burnham JM, et al. The anterolateral complex of the knee: a pictorial essay. Knee Surg Sports Traumatol Arthrosc. 2017;25(4):1009–1014. [DOI] [PubMed] [Google Scholar]
- 10. Hewison CE, Tran MN, Kaniki N, et al. Lateral extra-articular tenodesis reduces rotational laxity when combined with anterior cruciate ligament reconstruction: a systematic review of the literature. Arthroscopy. 2015;31(10):2022–2034. [DOI] [PubMed] [Google Scholar]
- 11. Kaplan EB. The iliotibial tract: clinical and morphological significance. J Bone Joint Surg Am. 1958;40(4):817–832. [PubMed] [Google Scholar]
- 12. Kittl C, El-Daou H, Athwal KK, et al. The role of the anterolateral structures and the ACL in controlling laxity of the intact and ACL-deficient knee. Am J Sports Med. 2016;44(2):345–354. [DOI] [PubMed] [Google Scholar]
- 13. Kowalczuk M, Herbst E, Burnham JM, Albers M, Musahl V, Fu FH. A layered anatomic description of the anterolateral complex of the knee. Clin Sports Med. 2018;37(1):1–8. [DOI] [PubMed] [Google Scholar]
- 14. Lemaire M. Ruptures anciennes du ligament croisé antérieur du genou. J Chir. 1967;93:311–320. [Google Scholar]
- 15. Lobenhoffer P, Posel P, Witt S, Piehler J, Wirth CJ. Distal femoral fixation of the iliotibial tract. Arch Orthop Trauma Surg. 1987;106(5):285–290. [DOI] [PubMed] [Google Scholar]
- 16. Marcacci M, Zaffagnini S, Giordano G, et al. Anterior cruciate ligament reconstruction associated with extra-articular tenodesis: a prospective clinical and radiographic evaluation with 10- to 13-year follow-up. Am J Sports Med. 2009;37(4):707–714. [DOI] [PubMed] [Google Scholar]
- 17. Müller W. [Functional anatomy and clinical findings of the knee joint]. Helv Chir Acta. 1984;51(5):505–514. [PubMed] [Google Scholar]
- 18. Porrino J, Jr, Maloney E, Richardson M, Mulcahy H, Ha A, Chew FS. The anterolateral ligament of the knee: MRI appearance, association with the Segond fracture, and historical perspective. AJR Am J Roentgenol. 2015;204(2):367–373. [DOI] [PubMed] [Google Scholar]
- 19. Segond P. Recherches cliniques et expérimentales sur les épanchements sanguins du genou par entorse. Progrès Méd. 1879;7:297–341. [Google Scholar]
- 20. Sonnery-Cottet B, Saithna A, Cavalier M, et al. Anterolateral ligament reconstruction is associated with significantly reduced ACL graft rupture rates at a minimum follow-up of 2 years: a prospective comparative study of 502 patients from the SANTI study group. Am J Sports Med. 2017;45(7):1547–1557. [DOI] [PubMed] [Google Scholar]
- 21. Terry GC, Hughston JC, Norwood LA. The anatomy of the iliopatellar band and iliotibial tract. Am J Sports Med. 1986;14(1):39–45. [DOI] [PubMed] [Google Scholar]
- 22. Vesalius A. De Humani Corporis Fabrica. Lib II, Cap 53 Padua: University of Padua; 1543. [Google Scholar]
- 23. Vieira EL, Vieira EA, da Silva RT, Berlfein PA, Abdalla RJ, Cohen M. An anatomical study of the iliotibial tract. Arthroscopy. 2007;23(3):269–274. [DOI] [PubMed] [Google Scholar]
- 24. Vincent JP, Magnussen RA, Gezmez F, et al. The anterolateral ligament of the human knee: an anatomical and histologic study. Knee Surg Sports Traumatol Arthrosc. 2012;20(1):147–152. [DOI] [PubMed] [Google Scholar]