

Abstract
Telomeres, the protective structures at the ends of chromosomes, can be shortened when individuals are exposed to stress. In some species, the enzyme telomerase is expressed in adult somatic tissues, and potentially protects or lengthens telomeres. Telomeres can be damaged by ionizing radiation and oxidative stress, although the effect of chronic exposure to elevated levels of radiation on telomere maintenance is unknown for natural populations. We quantified telomerase expression and telomere length (TL) in different tissues of the bank vole Myodes glareolus, collected from the Chernobyl Exclusion Zone, an environment heterogeneously contaminated with radionuclides, and from uncontaminated control sites elsewhere in Ukraine. Inhabiting the Chernobyl Exclusion Zone was associated with reduced TL in the liver and testis, and upregulation of telomerase in brain and liver. Thus upregulation of telomerase does not appear to associate with longer telomeres but may reflect protective functions other than telomere maintenance or an attempt to maintain shorter telomeres in a stressful environment. Tissue specific differences in the rate of telomere attrition and apparent radiosensitivity weaken the intra-individual correlation in telomere length among tissues in voles exposed to radionuclides. Our data show that ionizing radiation alters telomere homeostasis in wild animal populations in tissue specific ways.
Introduction
Telomeres are nucleo-protein complexes that protect the ends of linear (i.e. eukaryotic) chromosomes, principally by preventing chromosome ends from fusing or being recognized as double strand DNA breaks1. Almost all vertebrate telomere sequences are composed of the motif TTAGGG that is repeated as a long, tandem array. Telomere length (TL) is reduced with cell division due to incomplete end replication1, with cell senescence triggered at a critically short TL2. As TL is widely correlated with longevity, health and fitness in humans and other animals, there is much interest in understanding the processes that impact TL dynamics. Stressors, such as psychological stress3,4, oxidative stress5,6 and environmental pollution7, correlate with accelerated rate of telomere shortening in humans and animals.
Telomere length homeostasis is more complex than the rate of shortening per se. A key mechanism by which telomeres may be repaired or extended is by the action of the enzyme telomerase8. Telomerase expression varies among tissues and among species9,10. Notably, small (~2 kg or less) rodents9,11,12 and some birds13 express telomerase in many somatic tissues as adults; conversely, many large long-lived mammals repress telomerase expression in somatic tissues as adults14,15, albeit with some exceptions (e.g. pig16). Moreover, the level of telomerase expression varies among tissues9. In adult mice (Mus musculus and M. spretus), telomerase expression tends to be higher in tissues with longer telomeres (such as testis and liver), and weak or absent in tissues with shorter telomeres (such as spleen, kidney and brain)17. Similar pattern is seen in adult rats where telomerase is highly expressed in the liver, but has low expression in the brain tissue18. Stress also affects telomerase expression, but in an unpredictable way, with exposure to stress capable of stimulating and repressing telomerase activity3,19–22. Typically, inadequate expression of telomerase is associated with accelerated telomere shortening and aging1,2. However, an increase in telomerase expression is not always beneficial, for example with cancerous cells becoming immortalized by overexpression of telomerase14. Indeed, the level of telomerase expression for appropriate telomere maintenance likely interacts with species’ life-history, particularly whether animals tend to repress or express telomerase when mature. As the levels of expression of telomerase in wildlife exposed to stress and its impact on TL dynamics is unknown, quantification of TL dynamics and telomerase expression in different tissues and in animals under stress is needed.
Contaminated environments present complex stressors and health impacts on wildlife23. Numerous human actions have led to the accidental release of radionuclides into the environment, leaving many contaminated areas worldwide24,25. The wildlife inhabiting the area surrounding the former Nuclear Power Plant (NPP) at Chernobyl, Ukraine, provide the best-studied model of the biological impacts of exposure to environmental radionuclides26. On April 26, 1986, an explosion and subsequent fire at the Chernobyl NPP Unit 4 led to the release of more than 9 × 103 PBq of radionuclides over much (>200,000 km2) of Europe and eastern Russia26. Subsequently, the Chernobyl Exclusion Zone (CEZ) was established at a 30 km radius around the site to limit human exposure to contamination. The organisms that inhabit the areas surrounding the Chernobyl NPP are exposed to elevated levels of radionuclides with long half-lives, notably strontium-90 (90Sr), cesium-137 (137Cs), and plutonium-239 (239Pu) that have half-lives of 29, 30 and 24,100 years respectively26,27. Diverse genomic impacts have been reported in wildlife inhabiting the areas surrounding Chernobyl25, including elevated levels of DNA damage28 and mutations27. However, these studies failed to identify specific genomic regions that are sensitive to damaging effects of inhabiting an area with elevated radioactivity.
We hypothesized that exposure to environmental radionuclides within the CEZ is likely to impact telomere homeostasis as telomeres present a specific target for damaging effects of ionizing radiation (IR) because G-rich regions of the genome are expected to be radiosensitive. Additionally, IR induced increase in cellular reactive oxygen species (ROS) and oxidative stress can disrupt telomere maintenance, as telomeres are sensitive to oxidative damage5,19,29. Indeed, both TL and telomerase expression are linked with radiosensitivity in vitro: cell lines with short telomeres show increased radiosensitivity when they lack a functioning telomerase19 (also in vivo in mice30). Other cell lines show a reduction in telomere length and an increase in telomerase expression after exposure to ionizing radiation31, and radiosensitive cells have accelerated telomere shortening and telomere dysfunction32,33. The extent to which wildlife experiencing a chronic, low dose exposure of ionizing radiation parallel these studies of cell lines exposed to acute, high doses of radiation (e.g. for clinical context) is unknown.
Radiosensitivity defines the relative susceptibility of cells, tissues, organs, organisms, or other substances to the injurious action of radiation. Radiosensitive cells are typically those with a high rate of division, high metabolic rate and/or are non-specialized34. Thus, TL in mice testis and skin is particularly sensitive to chronic oxidative stress due to the rapid rate of cell division in these cell types35. Liver, heart and brain tissues have high metabolic demands, but differ in their proliferative capabilities and production of ROS36. While tissues invariably cope with their intrinsic oxidative stress, the additional stress caused by exposure to radionuclides (i.e. inhabiting the CEZ) could pose a challenge for telomere maintenance.
In summary, different tissues possess an intrinsically different balance between telomere erosion and telomere repair and their radiosensitivity. The aim of this study was to quantify the effect that chronic exposure to ionizing radiation, associated with inhabiting the CEZ, has upon telomere homeostasis in wild populations by quantifying telomerase expression and TL in different tissues of the bank vole Myodes glareolus.
Methods
Model species
The bank vole Myodes glareolus is a muroid rodent that inhabits deciduous and coniferous woodland throughout much of northern Europe and Asia37. This species presents an ideal model to quantify biological effects of exposure to environmental radiation as it was one of the first mammals to recolonize areas contaminated by radionuclides following the Chernobyl accident38. Bank voles inhabiting the CEZ can experience considerable (i.e. >10 mGy/d, but average around 3.7 mGy/d) absorbed doses of radiation38. Evidence for genetic damage in bank voles inhabiting the CEZ is equivocal, with reports of increased levels of chromosomal aberrations contrasting with no evidence of increased micronuclei formation (reviewed by25) or accumulations of heteroplasmic mutations in mitochondrial DNA39. Nonetheless, bank voles inhabiting the CEZ upregulate some DNA repair enzymes40.
Sample collection
During 2015, bank voles were captured from 13 trapping locations within the CEZ and from control locations outside the CEZ: Brody, Lubny and Korostyshev (Fig. 1). The CEZ presents a mosaic of radionuclide contamination with areas of elevated levels of contamination and relatively uncontaminated areas across a broad landscape. Bank voles can move distance of 1 km during the breeding season41 and thus animals captured within the CEZ have the opportunity to be exposed to radionuclides prior to capture. In contrast, the control sites are located some 170 to 450 km from the CEZ and individuals from these locations would not have been exposed to environmental radionuclides. Levels of ambient ground-level soil radiation at trapping locations within the CEZ varied from 0.12 to 12.34 µSv/h (locations with elevated contamination: mean = 5.21 µSv/h, SD = 2.71 and clean areas with lower levels: mean = 0.19 µSv/h, SD = 0.11). The external radiation exposure of bank voles at Chernobyl strongly correlates (r = 0.86, Pearson) with the ambient background dose rates of the trapping sites (Lavrinienko et al. unpublished). The means for the uncontaminated control areas were 0.13 μSv/h, 0.14 μSv/h and 0.13 μSv/h for Brody, Lubny and Korostyshev, respectively. Thus, the level of soil background radiation was significantly different between the CEZ and control areas (t-test: t = 4.95, df = 58, P < 0.001).
Figure 1.

Myodes glareolus sampling sites. Location of uncontaminated control sampling sites (a) and the Chernobyl Exclusion Zone (CEZ, border marked with dotted line) in Ukraine (b). For the sampling locations within the CEZ (b), sites with ambient radiation > 1 µSv/h are marked red circles and sites with radiation level of <1 µSv/h are marked with green circles.
All procedures complied with the legal requirements and international guidelines for the use of animals in research. All necessary permissions for the experiments were obtained from the Animal Experimentation Committee (permission no. ESAVI/7256/04.10.07/2014).
Relative telomere length (RTL) and telomerase expression
We quantified relative TL in bank voles (N = 41) using the quantitative polymerase chain reaction (qPCR) method described by Cawthon, in which relative telomere length (RTL) is presented as the T/S ratio, where T is the amount of telomere in a sample and S reflects the amount of standard single copy gene (S). Additionally, the experimental samples are compared to a reference ‘golden standard’ DNA sample run on each qPCR plate42. TL was quantified in liver, brain, testis, ovary and heart tissue. Telomerase expression was quantified by qPCR in liver, brain, ovary and heart tissue. Telomerase expression is presented as relative quantities (fold change) to a reference cDNA sample that was present on all plates (see Supplementary Information for full details of both protocols).
Statistical analyses
Differences in TL and telomerase expression between samples collected from within the CEZ and the control areas were analyzed using generalized linear models (GLM) in R v.3.4.343, using sample area (CEZ, control) and sex as fixed factors. Intra-individual correlations for telomere length were examined for CEZ and control individuals separately, and pairwise comparison for all tissues were performed (Pearson’s correlation in R). REST 2.0.13 software44 was used for comparing general levels of telomerase expression among tissues.
Results
Telomere length
Relative telomere lengths were estimated in liver, brain, heart, testis and ovary tissue from bank voles within the CEZ and control animals from Ukraine which were not exposed to increased levels of ambient background radiation in their environment. Samples from the two groups within the CEZ were combined since animals from elevated radiation areas and relatively clean areas did not differ in TL. Animals from the CEZ had shorter telomeres compared to control animals in liver and testis. Brain, heart and ovary samples showed no differences between the two groups (Table 1, Fig. 2). Males had longer telomeres in all studied somatic tissues compared to females (Supplementary Information, Fig. S1). In general, telomeres were longer in liver and testis compared to brain, heart and ovary (Table 2, Fig. 2).
Table 1.
Summary of GLM results for relative telomere length (RTL) and telomerase expression for bank voles within the CEZ and control sites outside the CEZ, and the effect of sex.
| Telomere length (RTL) | Telomerase expression | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| B | SE | t | P | B | SE | t | P | ||
| Brain | intercept | 1.027 | 0.039 | 26.61 | 0.000 | 0.493 | 0.147 | 3.36 | 0.001 |
| Control | 0.045 | 0.044 | 1.03 | 0.311 | −0.481 | 0.167 | −2.89 | 0.006 | |
| Female | −0.281 | 0.044 | −6.37 | 0.000 | −0.025 | 0.167 | −0.15 | 0.883 | |
| Heart | intercept | 1.243 | 0.068 | 18.35 | 0.000 | −1.952 | 0.251 | −7.79 | 0.000 |
| Control | 0.064 | 0.076 | 0.84 | 0.407 | −0.382 | 0.358 | −1.066 | 0.292 | |
| Female | −0.375 | 0.076 | −4.96 | 0.000 | 0.372 | 0.358 | 1.038 | 0.305 | |
| Liver | intercept | 1.784 | 0.162 | 11.03 | 0.000 | 1.033 | 0.290 | 3.56 | 0.001 |
| Control | 0.435 | 0.187 | 2.33 | 0.026 | −0.878 | 0.320 | −2.75 | 0.008 | |
| Female | −0.508 | 0.187 | −2.72 | 0.009 | −0.311 | 0.320 | −0.97 | 0.337 | |
| Ovary | intercept | 1.026 | 0.104 | 9.86 | 0.000 | 0.105 | 0.118 | 0.896 | 0.380 |
| Control | −0.018 | 0.154 | −0.12 | 0.908 | −0.105 | 0.163 | −0.646 | 0.525 | |
| Testis | intercept | 1.594 | 0.258 | 6.17 | 0.000 | ||||
| Control | 1.020 | 0.357 | 2.86 | 0.010 | |||||
Values in bold are statistically significant at the 0.05 level.
Figure 2.
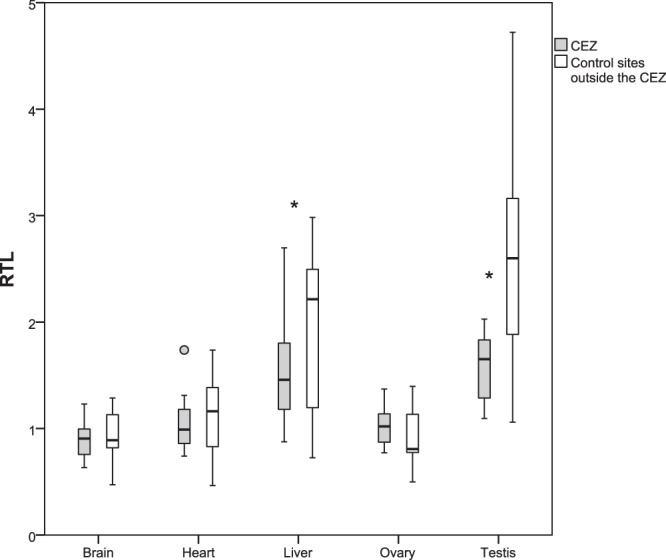
Telomere length. Relative telomere length (RTL) in the different tissues from bank voles from the Chernobyl Exclusion Zone (CEZ) and uncontaminated control sites. Box plots show medians, quartiles, 5- and 95-percentiles and extreme values. Statistically significant comparisons are marked with * (significant at the 0.05 level).
Table 2.
Mean relative telomere length (RTL) and relative telomerase expression in bank vole tissues from animals from the Chernobyl Exclusion Zone (CEZ) and control sites outside the CEZ.
| Tissue (N CEZ/control) | CEZ RTL (SD) | control RTL (SD) | combined |
|---|---|---|---|
| RTL | |||
| Brain (20/21) | 0.89 (0.17) | 0.94 (0.22) | 0.92 (0.20) |
| Heart (20/21) | 1.06 (0.24) | 1.13 (0.35) | 1.10 (0.30) |
| Liver (19/21) | 1.55 (0.46) | 1.98 (0.76) | 1.78 (0.66) |
| Ovary (12/10) | 1.03 (0.18) | 1.01 (0.49) | 1.02 (0.35) |
| Testis (10/11) | 1.59 (0.30) | 2.61 (1.07) | 2.13 (0.95) |
| Tissue (N CEZ/control) | CEZ telomerase (SD) | control telomerase (SD) | combined |
| Telomerase | |||
| Brain (30/29) | 1.69 (0.92) | 1.17 (0.62) | 1.44 (0.82) |
| Heart (25/24) | 0.37 (0.28) | 0.24 (0.17) | 0.31 (0.25) |
| Liver (28/28) | 0.80 (0.43) | 0.54 (0.34) | 0.67 (0.40) |
| Ovary (12/13) | 1.25 (0.31) | 1.16 (0.33) | 1.20 (0.32) |
N = sample size, SD = standard deviation.
Telomerase
Expression of telomerase was higher in the brain and ovary tissues compared to liver and heart (Fig. 3, Table 2). There was a 5.5 fold difference between the tissues with highest and lowest telomerase expression, brain and heart, respectively. Two tissues showed significant increase in telomerase expression in animals from the CEZ (Table 1). Telomerase was upregulated (1.4 fold change) in brain tissue of Chernobyl animals compared to individuals from control sites outside the CEZ, with no difference between males and females. No difference was seen between CEZ animals from elevated radiation or clean areas when three groups were analyzed. Increase in telomerase expression was also observed in liver tissue of Chernobyl voles compared to control individuals (1.7 fold change, no difference between sexes). In ovaries and heart tissue, there were no differences in telomerase expression between animals from Chernobyl and control areas (or between sexes for heart).
Figure 3.
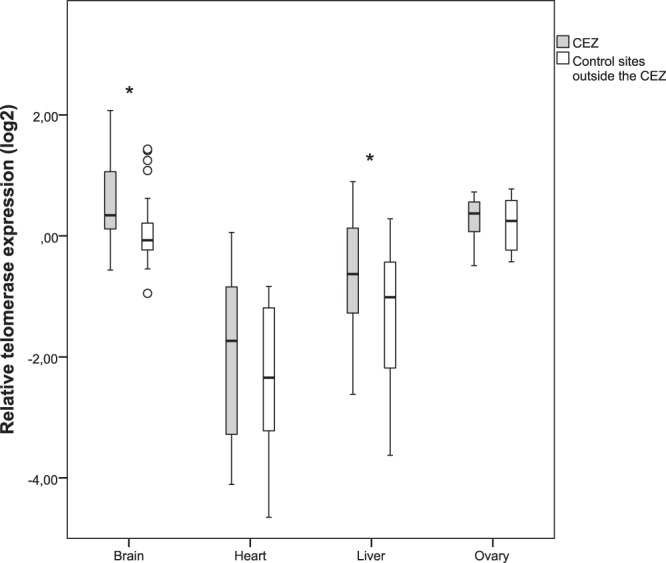
Telomerase expression. Relative telomerase expression in the different tissues from bank voles from the Chernobyl Exclusion Zone (CEZ) and uncontaminated control sites. Box plots show medians, quartiles, 5- and 95-percentiles and extreme values. Statistically significant comparisons are marked with * (significant at the 0.05 level).
Correlation in telomere length among tissues
In animals from uncontaminated control locations outside the CEZ, within individual correlations between telomere lengths in different tissues were positive and significant (pairwise correlations ranging from r = 0.50–0.82, excluding testis, Table 3), with the highest correlation seen between the post-mitotic heart and brain tissues (r = 0.82). The exception was testis, where patterns were weak and non-significant. However, in animals from Chernobyl, the intra-individual correlations in TL between tissues were weaker, and for example, correlations involving liver tissue were degraded (r = 0.14 in TL of liver and the somatic tissues brain and heart) (Table 3).
Table 3.
Intra-individual Pearson correlations for telomere length in the different tissues.
| Brain | Heart | Liver | Ovary | Testis | |
|---|---|---|---|---|---|
| Brain | 0.40* | 0.14 | 0.58* | −0.61* | |
| Heart | 0.82*** | 0.14 | 0.01 | −0.21* | |
| Liver | 0.50** | 0.55*** | −0.13 | 0.61 | |
| Ovary | 0.64** | 0.80*** | 0.73** | — | |
| Testis | 0.27 | 0.14 | 0.40 | — |
Values above the diagonal are within animals from the Chernobyl Exclusion Zone (CEZ) and values below the diagonal are from animals from uncontaminated control sites.
Significance level: ***< 0.001, **0.01, *0.05.
Discussion
Genetic damage is a characteristic biomarker for organisms inhabiting areas contaminated by environmental radioactivity25, and previous studies have shown that mammalian cell lines with short telomeres and/or malfunctioning telomerase are radiosensitive19,45. In bank voles inhabiting the CEZ, exposure to environmental radioactivity is associated with upregulation of telomerase (principally in brain and liver) and a general reduction in telomere length (TL) (in liver and testis). Additionally, tissue-specific differences in the sensitivity to exposure to radionuclides disrupts the within-individual correlation in TL of different tissues. To the best of our knowledge this is the first evidence that exposure to environmental radionuclides impacts telomere homeostasis in wild animals.
Sex specific differences in TL are common in mammals, with adult females having longer telomeres than males due to a slower rate of telomere attrition throughout life10,11. Why female bank voles have shorter telomeres than male voles is unclear, but may reflect greater growth (cell division) in the larger females or higher costs of reproduction46. Additionally, the intracellular oxidative stress environment may differ among bank vole sexes; for example, female rats have greater protection against oxidative stress and longer telomeres47. Nonetheless, we found no interaction between sex and the effects of exposure to radionuclides on telomere homeostasis.
As exposure to stressful events impacts telomere dynamics48,49, and telomere length (TL) is equivalent among tissues of newborn animals17,50, any differences in TL among adult tissues reflects tissue-specific rates of erosion (e.g. cell division, oxidative stress) and repair (e.g. expression of telomerase). In bank vole, the apparently high radiosensitivity of the testes34 (i.e. the reduction in mean and variance in TL, Fig. 2) is consistent with the high rate of cell division in this tissue35. Nonetheless, telomeres of bank vole testes are longer than telomeres of somatic tissues, consistent with tissue specific variation in TL among mouse17, implying that it is critical to maintain long telomeres in testes. Maintenance of TL in bank vole testes is presumably achieved by expression of telomerase, as the testes are generally characterized by high telomerase expression, even in species that repress telomerase in somatic tissues as adults9,14,51. Our qPCRs confirmed expression of telomerase in bank vole testes in all sample locations, although we were unable to determine relative telomerase expression associated with exposure to radionuclides (see Supplementary Information). Repression of telomerase associated with inhabiting the CEZ is unlikely, given the general trend in the somatic bank vole tissues (Fig. 3), and that a lack of telomerase expression in testes has critical fitness implications (such as depletion of sperm, decrease in testis size and increased apoptosis associated with excessive telomere shortening)52. The shorter TL of testes in the contaminated areas is important as it implies that radionuclide contamination can impact the multigenerational dynamics of TL in wild mammal populations. In humans and other mammals there is evidence that TL is determined by the paternal TL at the age of conception, with offspring inheriting longer telomeres from older fathers with longer telomeres in the sperm cells53. Short telomeres in bank voles inhabiting the CEZ may reflect reproduction by younger males, for example due to accelerated mortality in the contaminated environment, and/or accelerated telomere erosion due to the stressful environment (and potentially inadequate repair by telomerase). Irrespective of the mechanism(s) associated with telomere attrition, young bank voles likely begin their lives with shorter telomeres than typical of bank voles born in uncontaminated environments.
While ovary tissue is considered to be sensitive to radiation induced damage34, neither TL nor telomerase expression in ovaries were notably impacted by exposure to radionuclides. Low telomerase expression is common in mature ovary tissue and oocytes in mammals (e.g. macaque Macaca fascicularis ovaries54, Mus spretus oocytes55) as in the bank vole. Some studies have implicated a role for maternal TL inheritance in mammals56. While the mode of telomere heritance is not known for bank voles, the short TL in ovaries is perhaps consistent with the putative role of paternal, not maternal, mode of TL inheritance.
The contrasting effects of exposure to radionuclides on TL of liver, brain and heart tissues appears consistent with their comparative radiosensitivity, with no differences in TL among the CEZ and control samples in brain and heart (Fig. 2), which are less sensitive to radiation damage than liver tissue34. However, the comparatively low TL in bank vole brain and heart could indicate that these tissues naturally experience telomere attrition during development, and the short telomeres in adults reduce statistical power to detect an effect of radiation. Samples of newborn and adolescent bank voles are required to determine whether exposure to radionuclides accelerate telomere erosion during development. Given the short telomeres in bank vole brain and heart, it is notable that telomerase expression is low or absent in mouse and rat brain and heart tissues17,18,35, and similar pattern of expression occurs in other rodents and birds as well9,13. However, while telomerase expression was also low in bank vole heart, there was relatively high expression of telomerase in brain tissue, and upregulation of this enzyme in voles inhabiting the CEZ. This potentially indicates an attempt to prevent telomeres of the brains from becoming critically short in adults. It is possible that during adulthood, bank vole brain tissue is somewhat resistant to telomere attrition, as brains do not show age-related telomere shortening in rats10 and there is low mitochondrial ROS production in rodent brains36. Alternatively, telomerase may have wider functions than telomere maintenance, such as protection of cellular and mitochondrial function during oxidative stress57, inhibition of apoptosis58 and/or activation of DNA repair pathways59. Interestingly, small brain size is a characteristic malformation in birds affected by radioactive fallout from the CEZ60 and in adult bank voles inhabiting contaminated areas of the CEZ (Kivisaari et al. unpublished). Small brain size can imply fewer cell divisions during development, and this may be a further mechanism that limits the extent that exposure to radionuclides erodes TL. Whether small brain size could represent an ‘adaptation’ by bank voles inhabiting the CEZ, is an intriguing topic for further study.
Many (but not all) small rodents express telomerase in their livers as adults9,18 and have inherently long (compared to brain and kidney in mouse17) telomeres as adults. Long telomeres (relative to heart and brain) and expression of telomerase indicate a need to maintain telomere length in adult bank vole liver. Nonetheless, exposure to environmental stress impacts TL dynamics in the liver tissue, as inhabiting CEZ was associated with a reduction in TL and upregulation of telomerase. To highlight the complex relationship between telomerase expression and TL, the increased expression of telomerase in the liver appears insufficient to elongate/maintain TL in bank voles inhabiting the CEZ, but as noted above (for brain tissue) telomerase may have pleiotropic roles associated with the molecular stress response beyond safeguarding telomeres. Interestingly, expression of Mre11, which is involved with telomere maintenance, is upregulated in CEZ bank vole livers40. In any case, we find the liver tissue of adult bank voles exhibits some radiosensitivity as expected34.
In addition to the impact of telomerase, telomere length homeostasis can be regulated via multiple other mechanisms, for example recombination-mediated alternative lengthening of telomeres (ALT61) or stochastic events such as telomere rapid deletions (TDR62) can lengthen or shorten telomeres63,64. However the relevance of these is unclear especially in species and tissues where telomerase in continuously expressed, such as in the bank vole.
The similar patterns of telomere dynamics in animals inhabiting the CEZ, irrespective of the background level of contamination, is interesting in light of a recent debate about the possible role of historic dose influencing contemporary biological response65. As bank voles can disperse among the mosaic of radionuclide contamination within the CEZ, animals inhabiting apparently uncontaminated areas may have experienced radionuclide contamination at some point in their life. This prior experience of radionuclide exposure may trigger a persistent biological response, and epigenetic effects may even persist across generations66. Alternatively, changes in telomere homeostasis might reflect persistent adaptation(s) to radionuclide contamination that was driven by selection in the population of bank voles that recolonized the CEZ soon after the accident.
Despite the intrinsic tissue-specific variation in the rate of telomere attrition during an individual’s life, a positive correlation in telomere lengths among tissues within individuals is typical in humans54,67 and other animals54,68,69. This intra-individual correlation in TL exists in natural populations of bank voles, except for comparisons involving the testis where telomere lengths are long and variable. A crucial outcome of our analysis is that exposure to environmental radioactivity (and potentially other stressors) weakens or removes the intra-individual correlation in TL, presumably due to tissue-specific differences in radiosensitivity that impact telomere homeostasis (i.e. the rate of telomere attrition and effects of telomerase) in some tissues more than others.
Exposure to stress can induce telomerase expression in animals19,21,22. Upregulation of telomerase and short TL characterizes bank voles exposed to environmental radionuclides in the CEZ. The impact on telomere dynamics in humans is unclear, as contrasting results of either slight lengthening70, or shortening71 of leukocyte telomeres have been reported in studies of the late effects of exposure to radioactive fallout in Chernobyl clean-up workers. Humans can increase telomerase expression in response to acute stress20, but telomerase expression associated with exposure to environmental radiation in humans is unknown. Of course, it is hard to compare these human data with our study as bank voles are continually exposed to radionuclides in soil and internally via their diet; moreover, small and large animals have different telomere homeostasis, notably in the pattern of telomerase expression in somatic tissues of adults9,12. However, our data on telomerase are comparable with studies on cell lines; upregulation of telomerase is a common response by cell lines to acute exposure to ionizing radiation (>1 Gy)19,31. Given the tissue specific differences in telomerase expression and telomere dynamics in bank voles, further work on other mechanisms possibly affecting telomere dynamics in response to stress is required. For example, methylation levels at subtelomeric regions can impact telomere dynamics by regulating the frequency of telomeric recombination events or the expression of genes involved in telomere maintenance72, and may be relevant for telomere maintenance in species inhabiting the CEZ, as radiation exposure has been shown to alter genome wide methylation levels in Pinus silvestris from the CEZ73.
Currently, telomerase expression is a neglected aspect of our understanding of telomere dynamics in wild animal populations (but see13), however it is important to study both TL and telomerase expression to understand telomere homeostasis, especially in species expressing telomerase in somatic tissues as adults. Thus, while TL and rate of telomere loss are widely associated with lifespan74, expression of telomerase is another mechanism associated with (cellular) longevity75,76. We also caution against an interpretation that the generally short telomere length associated with the CEZ is evidence that telomerase does not maintain telomeres or that telomerase is directed towards other molecular functions. As telomeres are sensitive to damage by factors that increase oxidative stress (such as radiation), it is not clear whether long telomeres per se would be better adapted to a stressful environment. For example, murine cell lines or mice with short telomeres and which lack telomerase expression are the most sensitive to acute exposure to ionizing radiation19,30,77, indicating that telomerase could protect short telomeres against radiation-induced damage78. Longer telomeres present a greater target (physical length) for damage by radiation, and excessively long telomeres can confer genomic instability79. Therefore, a combination of short telomeres that are better protected/repaired (i.e. via telomerase expression), rather than attempting to maintain long telomeres, may be a more effective method of maintaining cell function and genomic stability in areas contaminated by radionuclides. Bearing in mind the potential effects of short TL in testis, future studies need to quantify whether short telomeres are adaptive (i.e. easier to maintain and repair) within the CEZ, or whether short telomeres are a passive reflection of life within a stressful environment.
Supplementary information
Acknowledgements
PCW and TM are funded by Academy of Finland (project numbers 287153 and 268670 respectively). TAM and APM were supported by the Samuel Freeman Charitable Trust, the CNRS, and the University of South Carolina. ZB is supported by the Portuguese Foundation for Science and Technology (SFRH/BPD/84822/2012). KK is supported by the Emil Aaltonen Foundation (Grant No. 160107). We gratefully acknowledge Igor Chizhevsky, Serhii Kireyev, Anatoly Nosovsky and Maksym Ivanenko for logistic support and help in organizing fieldwork in Ukraine. We thank Juho Niva for laboratory assistance.
Author Contributions
J.K., A.L., E.T., Z.B., K.K., T.M., G.M., A.P.M., T.A.M. and P.W. participated in the collection of research material. J.K. performed the laboratory analyses and analyzed the results. J.K. wrote the manuscript in collaboration with all authors.
Data Availability
The datasets analysed during the current study are available from the corresponding author on reasonable request.
Competing Interests
The authors declare no competing interests.
Footnotes
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Supplementary information accompanies this paper at 10.1038/s41598-018-37164-8.
References
- 1.Zakian VA. Telomeres: The beginnings and ends of eukaryotic chromosomes. Exp. Cell Res. 2012;318:1456–1460. doi: 10.1016/j.yexcr.2012.02.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Shay JW, Wright WE. Hayflick, His Limit, and Cellular Ageing. Nat. Rev. Mol. Cell Biol. 2000;1:72–76. doi: 10.1038/35036093. [DOI] [PubMed] [Google Scholar]
- 3.Epel ES, et al. Accelerated telomere shortening in response to life stress. Proc. Natl. Acad. Sci. USA. 2004;101:17312–5. doi: 10.1073/pnas.0407162101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mitchell C, et al. Social disadvantage, genetic sensitivity, and children’s telomere length. Proc. Natl. Acad. Sci. 2014;111:5944–5949. doi: 10.1073/pnas.1404293111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Von Zglinicki T. Oxidative stress shortens telomeres. Trends Biochem. Sci. 2002;27:339–344. doi: 10.1016/S0968-0004(02)02110-2. [DOI] [PubMed] [Google Scholar]
- 6.Geiger S, et al. Catching-up but telomere loss: Half-opening the black box of growth and ageing trade-off in wild king penguin chicks. Mol. Ecol. 2012;21:1500–1510. doi: 10.1111/j.1365-294X.2011.05331.x. [DOI] [PubMed] [Google Scholar]
- 7.De Felice B, et al. Telomere shortening in women resident close to waste landfill sites. Gene. 2012;500:101–106. doi: 10.1016/j.gene.2012.03.040. [DOI] [PubMed] [Google Scholar]
- 8.Greider CW. Telomere length regulation. Annu. Rev. Biochem. 1996;65:337–365. doi: 10.1146/annurev.bi.65.070196.002005. [DOI] [PubMed] [Google Scholar]
- 9.Seluanov A, et al. Telomerase activity coevolves with body mass not lifespan. Aging Cell. 2007;6:45–52. doi: 10.1111/j.1474-9726.2006.00262.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cherif H, Tarry JL, Ozanne SE, Hales CN. Ageing and telomeres: A study into organ- and gender-specific telomere shortening. Nucleic Acids Res. 2003;31:1576–1583. doi: 10.1093/nar/gkg208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Barrett ELB, Richardson DS. Sex differences in telomeres and lifespan. Aging Cell. 2011;10:913–921. doi: 10.1111/j.1474-9726.2011.00741.x. [DOI] [PubMed] [Google Scholar]
- 12.Gorbunova V, Seluanov A. Coevolution of telomerase activity and body mass in mammals: From mice to beavers. Mech. Ageing Dev. 2009;130:3–9. doi: 10.1016/j.mad.2008.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Haussmann MF, Winkler DW, Huntington CE, Nisbet ICT, Vleck CM. Telomerase activity is maintained throughout the lifespan of long-lived birds. Exp. Gerontol. 2007;42:610–618. doi: 10.1016/j.exger.2007.03.004. [DOI] [PubMed] [Google Scholar]
- 14.Kim NW, et al. Specific association of human telomerase activity with immortal cells and cancer. Science. 1994;266:2011–5. doi: 10.1126/science.7605428. [DOI] [PubMed] [Google Scholar]
- 15.Gomes NMV, et al. Comparative biology of mammalian telomeres: Hypotheses on ancestral states and the roles of telomeres in longevity determination. Aging Cell. 2011;10:761–768. doi: 10.1111/j.1474-9726.2011.00718.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Fradiani PA, Ascenzioni F, Lavitrano M, Donini P. Telomeres and telomerase activity in pig tissues. Biochimie. 2004;86:7–12. doi: 10.1016/j.biochi.2003.11.009. [DOI] [PubMed] [Google Scholar]
- 17.Prowse KR, Greider CW. Developmental and tissue-specific regulation of mouse telomerase and telomere length. Proc. Natl. Acad. Sci. USA. 1995;92:4818–22. doi: 10.1073/pnas.92.11.4818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Yamaguchi Y, et al. Change in telomerase activity of rat organs during growth and aging. Exp. Cell Res. 1998;242:120–127. doi: 10.1006/excr.1998.4102. [DOI] [PubMed] [Google Scholar]
- 19.Ayouaz A, Raynaud C, Heride C, Revaud D, Sabatier L. Telomeres: Hallmarks of radiosensitivity. Biochimie. 2008;90:60–72. doi: 10.1016/j.biochi.2007.09.011. [DOI] [PubMed] [Google Scholar]
- 20.Epel ES, et al. Dynamics of telomerase activity in response to acute psychological stress. Brain. Behav. Immun. 2010;24:531–539. doi: 10.1016/j.bbi.2009.11.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Beery AK, et al. Chronic stress elevates telomerase activity in rats. Biol. Lett. 2012;8:1063–1066. doi: 10.1098/rsbl.2012.0747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Saretzki G. Telomerase, mitochondria and oxidative stress. Exp. Gerontol. 2009;44:485–492. doi: 10.1016/j.exger.2009.05.004. [DOI] [PubMed] [Google Scholar]
- 23.Acevedo-Whitehouse K, Duffus ALJ. Effects of environmental change on wildlife health. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2009;364:3429–3438. doi: 10.1098/rstb.2009.0128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hu Q-H, Weng J-Q, Wang J-S. Sources of anthropogenic radionuclides in the environment: a review. J. Environ. Radioact. 2010;101:426–437. doi: 10.1016/j.jenvrad.2008.08.004. [DOI] [PubMed] [Google Scholar]
- 25.Lourenço J, Mendo S, Pereira R. Radioactively contaminated areas: Bioindicator species and biomarkers of effect in an early warning scheme for a preliminary risk assessment. J. Hazard. Mater. 2016;317:503–542. doi: 10.1016/j.jhazmat.2016.06.020. [DOI] [PubMed] [Google Scholar]
- 26.Møller AP, Mousseau TA. Biological consequences of Chernobyl: 20 years on. Trends Ecol. Evol. 2006;21:200–207. doi: 10.1016/j.tree.2006.01.008. [DOI] [PubMed] [Google Scholar]
- 27.Møller AP, Mousseau TA. Strong effects of ionizing radiation from Chernobyl on mutation rates. Sci. Rep. 2015;5:8363. doi: 10.1038/srep08363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bonisoli-Alquati A, et al. DNA damage in barn swallows (Hirundo rustica) from the Chernobyl region detected by use of the comet assay. Comp. Biochem. Physiol. - C Toxicol. Pharmacol. 2010;151:271–277. doi: 10.1016/j.cbpc.2009.11.006. [DOI] [PubMed] [Google Scholar]
- 29.Oikawa S, Kawanishi S. Site-specific DNA damage at GGG sequence by oxidative stress may accelerate telomere shortening. FEBS Lett. 1999;453:365–368. doi: 10.1016/S0014-5793(99)00748-6. [DOI] [PubMed] [Google Scholar]
- 30.Goytisolo FA, et al. Short telomeres result in organismal hypersensitivity to ionizing radiation in mammals. J. Exp. Med. 2000;192:1625–36. doi: 10.1084/jem.192.11.1625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sishc BJ, et al. Telomeres and Telomerase in the Radiation Response: Implications for Instability, Reprograming, and Carcinogenesis. Front. Oncol. 2015;5:1–19. doi: 10.3389/fonc.2015.00257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cabuy E, et al. Accelerated telomere shortening and telomere abnormalities in radiosensitive cell lines. Radiat. Res. 2005;164:53–62. doi: 10.1667/RR3376. [DOI] [PubMed] [Google Scholar]
- 33.McIlrath J, et al. Telomere length abnormalities in mammalian radiosensitive cells. Cancer Res. 2001;61:912–915. [PubMed] [Google Scholar]
- 34.Rubin P, Casarett GW. Clinical radiation pathology as applied to curative radiotherapy. Cancer. 1968;22:767–778. doi: 10.1002/1097-0142(196810)22:4<767::AID-CNCR2820220412>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- 35.Cattan V, et al. Chronic oxidative stress induces a tissue-specific reduction in telomere length in CAST/Ei mice. Free Radic. Biol. Med. 2008;44:1592–1598. doi: 10.1016/j.freeradbiomed.2008.01.007. [DOI] [PubMed] [Google Scholar]
- 36.Panov A, et al. Species- and tissue-specific relationships between mitochondrial permeability transition and generation of ROS in brain and liver mitochondria of rats and mice. Am. J. Physiol. Cell Physiol. 2007;292:C708–18. doi: 10.1152/ajpcell.00202.2006. [DOI] [PubMed] [Google Scholar]
- 37.Macdonald, D. W. The Encyclopedia of Mammals. 3rd edition. Oxford University Press, Oxford, UK (2007).
- 38.Chesser RK, et al. Concentrations and dose rate estimates of (134,137)cesium and (90)strontium in small mammals at Chornobyl, Ukraine. Environ. Toxicol. Chem. 2000;19:305–312. [Google Scholar]
- 39.Kesäniemi J, et al. Analysis of heteroplasmy in bank voles inhabiting the Chernobyl exclusion zone: A commentary on Baker et al. (2017) ‘Elevated mitochondrial genome variation after 50 generations of radiation exposure in a wild rodent’. Evol. Appl. 2018;11:820–826. doi: 10.1111/eva.12578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Jernfors, T. et al. Transcriptional Upregulation of DNA Damage Response Genes in Bank Voles (Myodes glareolus) Inhabiting the Chernobyl Exclusion Zone. Front. Environ. Sci. 5 (2018).
- 41.Kozakiewicz M, Chołuj A, Kozakiewicz A. Long-distance movements of individuals in a free-living bank vole population: an important element of male breeding strategy. Acta Theriol. (Warsz). 2007;52:339–348. doi: 10.1007/BF03194231. [DOI] [Google Scholar]
- 42.Cawthon RM. Telomere measurement by quantitative PCR. Nucleic Acids Res. 2002;30:47e–47. doi: 10.1093/nar/30.10.e47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, http://www.R-project.org (2014).
- 44.Pfaffl MW, Horgan GW, Dempfle L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002;30:e36. doi: 10.1093/nar/30.9.e36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Slijepcevic P. DNA damage response, telomere maintenance and ageing in light of the integrative model. Mech. Ageing Dev. 2008;129:11–16. doi: 10.1016/j.mad.2007.10.012. [DOI] [PubMed] [Google Scholar]
- 46.Speakman JR. The physiological costs of reproduction in small mammals. Phil. Trans. R. Soc. B. 2008;363:375–398. doi: 10.1098/rstb.2007.2145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Tarry-Adkins JL, Ozanne SE, Norden A, Cherif H, Hales CN. Lower antioxidant capacity and elevated p53 and p21 may be a link between gender disparity in renal telomere shortening, albuminuria, and longevity. Am. J. Physiol. Renal Physiol. 2006;290:F509–16. doi: 10.1152/ajprenal.00215.2005. [DOI] [PubMed] [Google Scholar]
- 48.Monaghan P. Organismal stress, telomeres and life histories. J. Exp. Biol. 2014;217:57–66. doi: 10.1242/jeb.090043. [DOI] [PubMed] [Google Scholar]
- 49.Romano, G. H. et al. Environmental Stresses Disrupt Telomere Length Homeostasis. PLoS Genet. 9 (2013). [DOI] [PMC free article] [PubMed]
- 50.Okuda K, et al. Telomere length in the newborn. Pediatr. Res. 2002;52:377–381. doi: 10.1203/00006450-200209000-00012. [DOI] [PubMed] [Google Scholar]
- 51.Ozturk, S. Telomerase Activity and Telomere Length in Male Germ Cells. Biol. Reprod. 92 (2015). [DOI] [PubMed]
- 52.Blasco M, et al. Essential role of mouse telomerase in highly proliferative organs. Nature. 1998;392:569–74. doi: 10.1038/33345. [DOI] [PubMed] [Google Scholar]
- 53.Njajou OT, et al. Telomere length is paternally inherited and is associated with parental lifespan. Proc. Natl. Acad. Sci. 2007;104:12135–12139. doi: 10.1073/pnas.0702703104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Gardner JP, et al. Telomere dynamics in macaques and humans. Journals Gerontol. - Ser. A Biol. Sci. Med. Sci. 2007;62:367–374. doi: 10.1093/gerona/62.4.367. [DOI] [PubMed] [Google Scholar]
- 55.Liu L, et al. Telomere lengthening early in development. Nat. Cell Biol. 2007;9:1436–1441. doi: 10.1038/ncb1664. [DOI] [PubMed] [Google Scholar]
- 56.Dugdale, H. L. & Richardson, D. S. Heritability of telomere variation: It is all about the environment! Phil. Trans. R. Soc. B. 373 (2018). [DOI] [PMC free article] [PubMed]
- 57.Ahmed S, et al. Telomerase does not counteract telomere shortening but protects mitochondrial function under oxidative stress. J. Cell Sci. 2008;121:1046–1053. doi: 10.1242/jcs.019372. [DOI] [PubMed] [Google Scholar]
- 58.Massard C, et al. hTERT: a novel endogenous inhibitor of the mitochondrial cell death pathway. Oncogene. 2006;25:4505–4514. doi: 10.1038/sj.onc.1209487. [DOI] [PubMed] [Google Scholar]
- 59.Sharma GG, et al. hTERT associates with human telomeres and enhances genomic stability and DNA repair. Oncogene. 2003;22:131–46. doi: 10.1038/sj.onc.1206063. [DOI] [PubMed] [Google Scholar]
- 60.Møller, A. P., Bonisoli-Alquati, A., Rudolfsen, G. & Mousseau, T. A. Chernobyl birds have smaller brains. PLoS One6 (2011). [DOI] [PMC free article] [PubMed]
- 61.Cesare AJ, Reddel RR. Alternative lengthening of telomeres: Models, mechanisms and implications. Nature Reviews Genetics. 2010;11:319–330. doi: 10.1038/nrg2763. [DOI] [PubMed] [Google Scholar]
- 62.Lustig AJ. Clues to catastrophic telomere loss in mammals from yeast telomere rapid deletion. Nature Reviews Genetics. 2003;4:916–923. doi: 10.1038/nrg1207. [DOI] [PubMed] [Google Scholar]
- 63.Hug N, Lingner J. Telomere length homeostasis. Chromosoma. 2006;115:413–425. doi: 10.1007/s00412-006-0067-3. [DOI] [PubMed] [Google Scholar]
- 64.Teixeira MT, Arneric M, Sperisen P, Lingner J. Telomere length homeostasis is achieved via a switch between telomerase-extendible and-nonextendible states. Cell. 2004;117:323–335. doi: 10.1016/S0092-8674(04)00334-4. [DOI] [PubMed] [Google Scholar]
- 65.Omar-Nazir L, et al. Long-term effects of ionizing radiation after the Chernobyl accident: Possible contribution of historic dose. Environ. Res. 2018;165:55–62. doi: 10.1016/j.envres.2018.04.005. [DOI] [PubMed] [Google Scholar]
- 66.Youngson NA, Whitelaw E. Transgenerational Epigenetic Effects. Annu. Rev. Genomics Hum. Genet. 2008;9:233–257. doi: 10.1146/annurev.genom.9.081307.164445. [DOI] [PubMed] [Google Scholar]
- 67.Daniali, L. et al. Telomeres shorten at equivalent rates in somatic tissues of adults. Nat. Commun. 4 (2013). [DOI] [PMC free article] [PubMed]
- 68.Benetos A, et al. A model of canine leukocyte telomere dynamics. Aging Cell. 2011;10:991–995. doi: 10.1111/j.1474-9726.2011.00744.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Smith DL, et al. Telomere dynamics in rhesus monkeys: No apparent effect of caloric restriction. Journals Gerontol. - Ser. A Biol. Sci. Med. Sci. 2011;66 A:1163–1168. doi: 10.1093/gerona/glr136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Reste J, et al. Telomere length in Chernobyl accident recovery workers in the late period after the disaster. J. Radiat. Res. 2014;55:1089–1100. doi: 10.1093/jrr/rru060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Ilyenko I, Lyaskivska O, Bazyka D. Analysis of relative telomere length and apoptosis in humans exposed to ionising radiation. Exp. Oncol. 2011;33:235–238. [PubMed] [Google Scholar]
- 72.Gonzalo S, et al. DNA methyltransferases control telomere length and telomere recombination in mammalian cells. Nat. Cell Biol. 2006;8:416–424. doi: 10.1038/ncb1386. [DOI] [PubMed] [Google Scholar]
- 73.Kovalchuk O, et al. Genome hypermethylation in Pinus silvestris of Chernobyl - A mechanism for radiation adaptation? Mutat. Res. - Fundam. Mol. Mech. Mutagen. 2003;529:13–20. doi: 10.1016/S0027-5107(03)00103-9. [DOI] [PubMed] [Google Scholar]
- 74.Tricola GM, et al. The rate of telomere loss is related to maximum lifespan in birds. Phil. Trans. R. Soc. B. 2018;373:20160445. doi: 10.1098/rstb.2016.0445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Blasco MA. Telomere length, stem cells and aging. Nat. Chem. Biol. 2007;3:640–649. doi: 10.1038/nchembio.2007.38. [DOI] [PubMed] [Google Scholar]
- 76.Bernardes de Jesus B, et al. Telomerase gene therapy in adult and old mice delays aging and increases longevity without increasing cancer. EMBO Mol. Med. 2012;4:691–704. doi: 10.1002/emmm.201200245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.DePinho RA, et al. Telomere dysfunction impairs DNA repair and enhances sensitivity to ionizing radiation. Nat. Genet. 2000;26:85–88. doi: 10.1038/79232. [DOI] [PubMed] [Google Scholar]
- 78.Rubio MA, Davalos AR, Campisi J. Telomere length mediates the effects of telomerase on the cellular response to genotoxic stress. Exp. Cell Res. 2004;298:17–27. doi: 10.1016/j.yexcr.2004.04.004. [DOI] [PubMed] [Google Scholar]
- 79.Rivera T, Haggblom C, Cosconati S, Karlseder J. A balance between elongation and trimming regulates telomere stability in stem cells. Nat. Struct. Mol. Biol. 2017;24:30–39. doi: 10.1038/nsmb.3335. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The datasets analysed during the current study are available from the corresponding author on reasonable request.