

Abstract
It has been confirmed that inflammation plays an important role in the pathogenesis of ischemic stroke. The polarization of microglia as an important participant in the inflammation following stroke is also found to be involved in stroke. This study aimed to investigate the effects of hydrogen gas on the polarization of macrophages/microglia in vitro. Raw264.7 cells were treated with lipopolysaccharides and then exposed to hydrogen. The microglia were treated with the supernatant from oxygen and glucose deprivation-treated neurons and then exposed to hydrogen. The phenotypes of Raw 264.7 cells and microglia were determined by flow cytometry, and cell morphology was observed. Results showed lipopolysaccharides significantly increased the M1 macrophages, and the supernatant from oxygen and glucose deprivation-treated neurons dramatically elevated the proportion of M1 microglia, but both treatments had little influence on the M2 cells. In addition, hydrogen treatment significantly inhibited the increase in M1 cells, but had no influence on M2 ones. Our findings suggest that the neuroprotection of hydrogen may be related to its regulation of microglia in the nervous system after stroke.
Keywords: macrophages, microglia, polarization, inflammation, hydrogen, stroke
INTRODUCTION
Stroke is the most common neurological disorder and the second leading cause of death worldwide.1,2 Stroke can be divided into ischemic and hemorrhagic ones, and the ischemic type resulting from numerous pathological processes accounts for nearly 87%.3 It has been confirmed that brain ischemia can cause neuronal death due to the deprivation of oxygen and energy, accumulation of intracellular Ca2+, production of reactive oxygen species, mitochondrial dysfunction, and inflammation.4,5
Basic and clinical studies have suggested that the pathophysiology of ischemic stroke is closely related to inflammation, in which microglia and macrophages in the central nervous system play an important role.6,7,8
Microglia and macrophages have different cellular origins. Microglia originates from neuroepithelium, and expresses high levels of CX3C chemokine receptor 1 (CX3CR1), cluster of differentiation (CD) 11b, and F4/80, low CD45 level, and no C-C chemokine receptor type 2 (CCR2).9 In contrast, macrophages originate from hematopoietic stem cells, and the inflammatory monocytes/macrophages express high levels of CCR2, CD11b and Ly6C, but low CX3CR1 level.10 In the presence of inflammation, resident microglia in the brain and infiltrated macrophages can be activated by cytokines. In recent years, studies have revealed that the activated cells can be further classified according to their specialized functions.11,12 Generally, microglia are classified as classic type (pro-inflammatory; M1-like) and alternative type (anti-inflammatory or protective; M2-like). M1-like microglia may aggravate inflammation and cause tissue injury via secreting some pro-inflammatory cytokines (such as interleukin (IL)-1β, tumor necrosis factor-α, IL-12, IL-23, and nitrogen monoxide), exerting pro-inflammatory effects; M2-like microglia may suppress inflammation and promote tissue recovery via secreting some anti-inflammatory cytokines (such as, IL-4, IL-10, IL-13 and transforming growth factor-β). Similar to microglia, macrophages can also polarize to M1- or M2-like phenotypes.13,14 In the presence of interferons from helper T cells,15 lipopolysaccharides (LPS) or damage-associated molecular pattern,16,17 the microglia and macrophages undergo M1-like activation; IL-4/IL-13 may induce the M2-like activation of microglia and macrophages. The M1-like responses are characterized by the up-regulation of IL-1β, tumor necrosis factor-α and inducible nitric oxide synthase through Toll-like receptor 4, and the M2-like responses by the up-regulation of arginase-1, CD206, CD163, IL-4, IL-10, and transforming growth factor-β through myeloid cells 2 activation. Recent studies indicate that the M1-like macrophages can switch towards an alternatively activated M2-like phenotype for activated phagocytosis.18,19 This concept is more convincing for macrophages than for microglia based on the function of mononuclear phagocytes.20
Hydrogen is the simplest molecule in nature that is colorless and odorless. Traditionally, hydrogen is used as a physiologically inert gas in deep diving for the prevention of nitrogen narcosis because it may not react with tissue components.21 In addition, hydrogen can also be produced in the human intestine and then expelled via respiration. Thus, hydrogen concentration of exhaled gas is used in the diagnosis of some gastrointestinal diseases.22 In recent years, increasing studies confirm that hydrogen may exert anti-oxidative, anti-inflammatory and anti-apoptotic effects on a variety of diseases,23 since Ohsawa et al.24 reported that hydrogen inhalation could protect the brain against cerebral ischemia/reperfusion injury by selectively neutralizing hydroxyl radicals and peroxynitrite. However, the effects of hydrogen on the polarization of microglia and macrophages remain unclear.
This study was undertaken to investigate the effects of hydrogen gas on the polarization of microglia and macrophages in vitro, which may provide a new mechanism for the anti-inflammatory activity of hydrogen.
MATERIALS AND METHODS
Animals
The mother mice were purchased from the Experimental Animal Center of Navy Medical University, Shanghai, China (No. A2015-011), maintained under a 12/12 light-dark cycle at 25°C and given ad libitum access to water and food. Neonatal mice (n = 30), either male or female, were sacrificed at 1 day after birth. The mean body weight of neonatal mice was 2.01 ± 0.21 g. These neonatal mice were sacrificed for the collection of primary microglia and hippocampal neurons. This study protocol was approved by the Ethics Committee of Navy Medical University, Shanghai, China (approval number: 20170236).
Cell separation, culture and observation
The neonatal mice were anesthetized with diethyl ether, and the brainstem or hippocampal tissues were harvested and washed with phosphate buffered saline (PBS). Subsequently, the tissues were digested in 0.5% collagenase (Sigma, St. Louis, MO, USA) at 37°C for 30 minutes. The mixture was dissociated by pipetting and then transferred to a sterile centrifuge tube. DMEM/F12 complete medium (containing 10% fetal bovine serum [FBS], 100 U/mL penicillin and 100 μg/mL streptomycin; Merck Millipore, Germany) was added to terminate digestion, and the resultant suspension was then centrifuged at 1000 × g for 5 minutes (TDZ4B-WS Low Speed Centrifuge; Lu Xiangyi Co., Ltd., Shanghai, China). The supernatant was removed, and remaining cells were re-suspended using DMEM (HyClone, Logan, UT, USA) containing 20% FBS (HyClone) and later seeded in flasks.
Raw264.7 cells were kept by Laboratory of Marine Medicine (Shanghai, China) and cultured in DMEM supplemented with 20% FBS. All cells were maintained in an incubator (XBQ-3H; Hualida Experiment Equipment Co., Ltd., Taicang, China) at 37°C in a humidified environment with 5% CO2. The medium was refreshed once every 2–3 days. To assess the protective effects of hydrogen, the macrophages were observed at 24 hours, 72 hours and 5 days (Leica DMI3000B, Germany).
Establishment of oxygen and glucose deprivation (OGD) neuronal model and collection of supernatant
The mouse primary hippocampal neurons were subjected to OGD as follows: after 5-day culture, the primary neuronal cells with 90% confluence were collected after centrifugation, and then maintained with Earle balanced salt solution (HyClone) in an incubator containing 5% O2 and 95% N2 at 37°C for 2 hours.25 The supernatant was collected for the treatment of primary microglia.
Grouping
The microglia were divided into four groups: control group (normal microglia), hydrogen group (microglia were exposed to 60% hydrogen for 24 hours), OGD group (microglia were exposed to the supernatant from OGD-treated neurons for 24 hours) and OGD + hydrogen group (microglia were exposed to the supernatant from OGD-treated neurons and to 60% hydrogen simultaneously for 24 hours).
Raw264.7 cells were divided into three groups: control group (normal Raw264.7 cells), LPS group (Raw264.7 cells were exposed to 1 μg/mL LPS [Sigma-Aldrich, St. Louis, MO, USA] for 24 hours), LPS + hydrogen group (Raw264.7 cells were exposed to 1 μg/mL LPS and to 60% hydrogen simultaneously for 24 hours).
Hydrogen exposure
For hydrogen treatment, cells were cultured in an incubator that filled with 66.7% hydrogen and 33.3% oxygen produced with the AMS-H-01 hydrogen producer (Asclepius, Shanghai, China), which was designed to electrolyze water to produce mixed gas.26 Cells were exposed to hydrogen gas for 24 hours.
Flow cytometry
There is evidence showing that the inflammation peaked at 24 hours after ischemic stroke.27 Therefore, Raw264.7 cells in each group were collected at 24 hours, 72 hours and 5 days; microglia were collected at 24 hours. Cells were washed with cold PBS thrice and centrifuged. The supernatant was removed, and cells were re-suspended in PBS at a density of 1 × 105 cells/mL. Subsequently, cells were fixed in fixation buffer in dark at room temperature for 20 minutes, and then centrifuged at 1000 × g for 5 minutes. After washing in 2% bovine serum albumin, cells were incubated in dark with CD11b-FITC (BioLegend, San Diego, CA, USA) and CD16/32-PE (BioLegend) for 20 minutes at 4°C to label M1 macrophages; cells were incubated in dark with CD11b-FITC (BioLegend) and CD206-APC (BioLegend) for 20 minutes at 4°C to label M2 macrophages. After washing in 2% bovine serum albumin, cells were suspended in 300 μL of PBS and then harvested for flow cytometry (B&D Biosciences, San Diego, CA, USA).
Statistical analysis
Statistical analysis was performed with SigmaPlot 11.0 (Systat Software, Inc., San Diego, CA, USA). Data are expressed as the mean ± standard deviation. Comparisons were performed with one-way analysis of variance (ANOVA) among groups, followed by Student-Newman-Keuls test. A value of P < 0.05 was considered statistically significant.
RESULTS
Cell morphology and growth rate
As shown in Figure 1, the normal Raw264.7 cells grew with static adherence, proliferated rapidly and were round in shape. After LPS treatment, most Raw264.7 cells became spindle-shaped, and the growth rate decreased over time. Cells were further counted. On day 3, the number of Raw264.7 cells increased significantly in the control group (growth rate: 95.40%), but the cell growth in the LPS group reduced significantly (20.51%) as compared to control group (P < 0.05). On day 5, the cell growth in control group was 1.9 times that on day 3, and the cell growth in the LPS group maintained at a low level (27.66%; P < 0.05, vs. control group). In the presence of 60% hydrogen during LPS treatment, the cell morphology remained relatively unchanged and the growth rate was also partially restored. The number of cells on day 3 was 2.85 times that on day 1 and the number of day 5 was 1.17 times that on day 3 (P < 0.05, vs. control group; P < 0.05 vs. LPS group). Of note, the growth rate of cells on day 5 in the LPS + hydrogen group was lower than that in the LPS group, although the total cell number on day 5 in the LPS + hydrogen group was still larger than in the LPS group.
Figure 1.
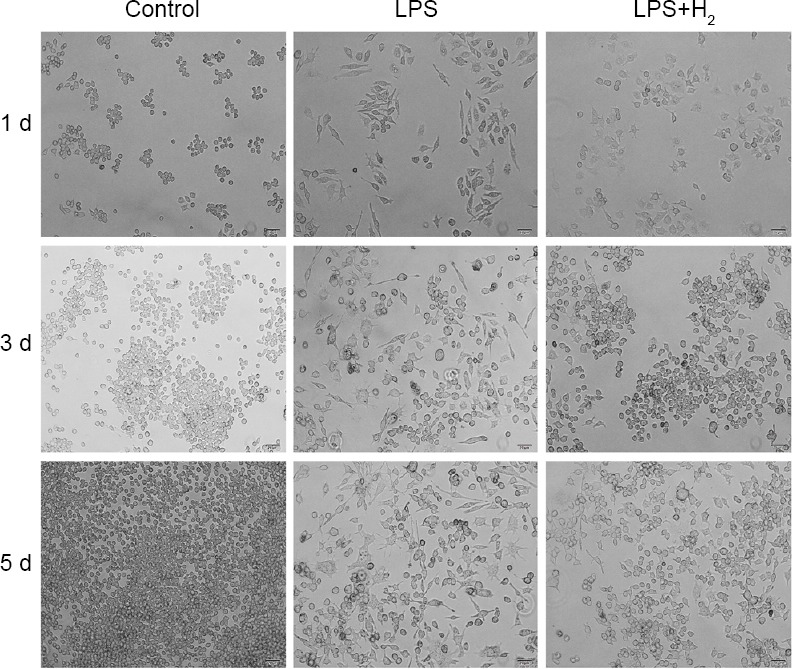
The morphology and growth of macrophages in vitro in different groups at 1, 3 and 5 d.
Note: The normal Raw264.7 cells were round in shape. After lipopolysaccharide treatment, most cells became spindle-shaped, and the growth rate decreased over time. After 60% hydrogen treatment, the cell morphology and growth rate partly recovered. LPS: Lipopolysaccharide; H2: hydrogen; d: day(s). Scale bars: 20 μm.
Raw264.7 cell phenotype
In the control group, the proportion of M1 macrophages remained stable at different time points (Figure 2). However, LPS treatment dramatically increased the proportion of M1 macrophages in a time dependent manner (P < 0.05, vs. control group). In the presence of hydrogen during LPS treatment, the proportion of M1 macrophages reduced significantly when compared with LPS group (P < 0.05) from day 1. Moreover, there was no significant difference in proportion of M1 macrophages between control group and LPS + hydrogen group (P > 0.05), although the proportion of M1 macrophages in LPS + hydrogen group was lower than in control group.
Figure 2.

Flow cytometry of M1 macrophages at 1, 3 and 5 d.
Note: (A) M1 macrophages are CD11b+CD16/32+. Green: CD16/32+ cells; purple: CD11b+ cells; red: CD16/32– and CD11b– cells. (B) At different time points, the proportion of M1 macrophages remained stable in the control group, and it increased in the LPS group over time. The hydrogen reduced the proportion significantly when compared with LPS group. *P < 0.05, vs. control group; #P < 0.05, vs. LPS + H2 group. LPS: Lipopolysaccharide; H2: hydrogen; d: day(s).
The proportion of M2 macrophages declined in control group over time, LPS treatment also reduced the proportion of M2 macrophages (P < 0.05, vs. control group) (Figure 3). Nevertheless, the addition of hydrogen during LPS treatment failed to increase the proportion of M2 macrophages to the normal level (P < 0.05, vs. control group), and there was no marked difference between LPS group and LPS + hydrogen group (P > 0.05). In addition, the proportion of M2 macrophages decreased over time in all groups.
Figure 3.
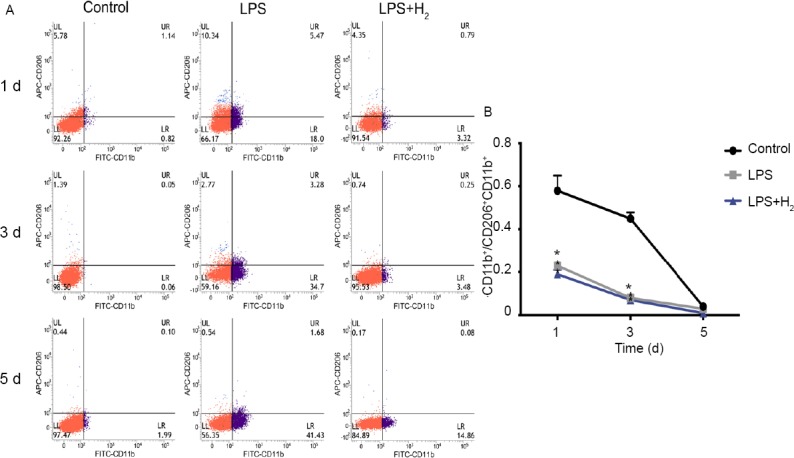
Flow cytometry of M2 macrophages at 1, 3 and 5 d.
Note: (A) M2 macrophages are CD11b+ CD206+. Blue: CD206+ cells; purple: CD11b+ cells; red: CD206- and CD11b– cells. (B) The proportion of M2 macrophages decreased over time in all groups. The LPS reduced the proportion of M2 macrophage compared with control group. However, there was no statistically significant difference between LPS group and LPS + H2 group. *P < 0.05, vs. control group. LPS: Lipopolysaccharide; H2: hydrogen; d: day(s).
Microglia phenotype
In control group, the proportion of M1 microglia was relatively low at 24 hours (Figures 4 and 6), and it remained at a low level in H2 group (P > 0.05). However, the supernatant from OGD-treated neurons significantly increased the proportion of M1 microglia (P < 0.05, vs. control group). In the presence of hydrogen during the supernatant treatment, the proportion of M1 microglia reduced to nearly normal level (P > 0.05, vs. control group).
Figure 4.
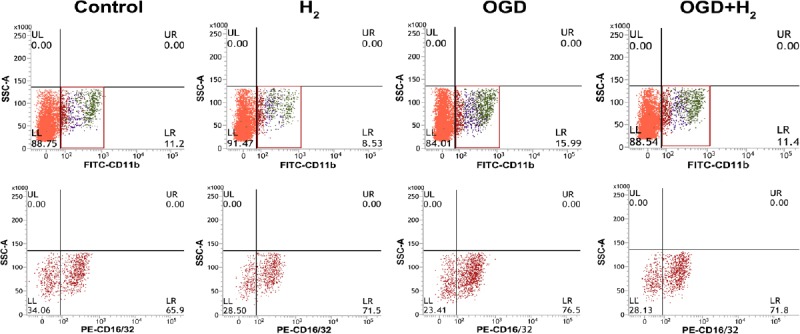
The phenotype of microglia was detected at 24 hours. Microglia were CD11b+ and M1 microglia were CD16/32+.
Note: Red boxes in the first row represent the selected gates including microglias, and the regions with different colors represent the signals of different cell population. Red regions in the second row represent CD16/32+ cells, and the fourth quadrantal area represents the high signal. OGD: Oxygen and glucose deprivation; H2: hydrogen.
Figure 6.

Proportion of M1 and M2 macrophages in different groups.
Note: (A) Proportion of M1 macrophages (ratio of CD11b+CD16/CD32+ cells to CD11b+ cells). (B) Proportion of M2 macrophages (ratio of CD206+ cells to CD11b+ cells). The OGD supernatant increased the proportion of M1 and reduced the proportion of M2 slightly. However, the hydrogen can reduced this impact to the normal level. *P < 0.05, vs. control group. OGD: Oxygen and glucose deprivation; H2: hydrogen.
The proportion of M2 microglia remained stable in each group and there was no statistical difference between experimental groups and the control group (Figures 5 and 6). However, the proportion of M2 microglia tended to increase in OGD group and seemed to reduce in hydrogen group and the addition of hydrogen during the OGD supernatant treatment slightly reduced the proportion of M2 macrophages.
Figure 5.
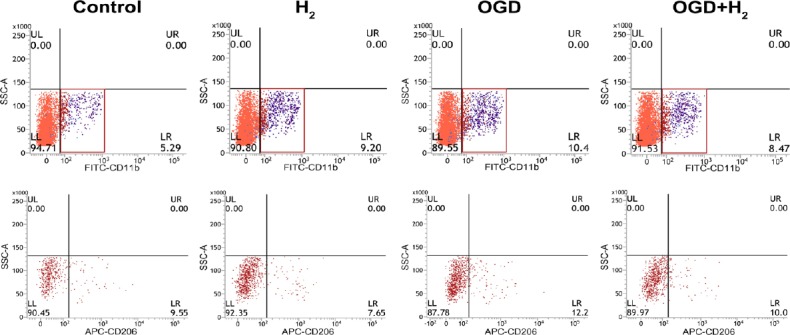
The phenotype of microglia was detected at 24 hours. Microglia were CD11b+ cells and M2 microglia were CD206+.
Note: Red boxes in the first row represent the selected gates including microglias, and the regions with different colors represent the signals of different cell population. Red regions in the second row represent CD206+ cells, and the fourth quadrantal area represents the high signal. OGD: Oxygen and glucose deprivation; H2: hydrogen.
DISCUSSION
Stroke is a medical emergency and may cause devastating neurological deficits. To date, stroke has been the major cause of death and adult disability worldwide.28 Although the pathogenesis of ischemic stroke (a major type of stroke) is complex, some studies have confirmed the important role of inflammation in the pathogenesis of ischemic stroke.29 The inflammatory response contributes to secondary neuronal damage, which exerts a substantial impact on both acute ischemic injury and the chronic recovery of the brain function. Brain ischemia elicited inflammation has involvement of the activation of microglia and monocyte-derived macrophages in the central nervous system. Microglia are the resident immune cells in the brain. Once ischemia occurs, microglia are activated to produce both detrimental and neuroprotective mediators, and the balance of the two counteracting mediators determines the fate of injured neurons.30 The activation of microglia is defined as classic (M1) or alternative (M2), which corresponds to the detrimental and protective effects of these cells, respectively.31 In this study, the macrophages and microglia were investigated independently.
Hydrogen has been used as a physiologically inert gas in deep diving, but increasing studies in recent years reveal it possesses some physiological activities and can exert protective effects on a variety of diseases.31 Although the anti-inflammatory effect of hydrogen has been validated in numerous models, the specific mechanism is still poorly understood. The polarization of macrophage is closely related to inflammation,31 but there is a lack of research on the role of hydrogen in the macrophage polarization. In this study, the effects of hydrogen on the polarization of macrophages and microglia (important participants in ischemic stroke) were investigated. LPS treated macrophages have been used as classic inflammation model.32
Our results showed LPS significantly increased the M1 macrophages and slightly reduced the M2 macrophages simultaneously, but hydrogen was able to markedly reduce M1 macrophages from day 1 and had little influence on M2 macrophages. These results suggest that LPS may activate macrophages, which is characterized by the increase in pro-inflammatory (M1) macrophages, but has little influence on anti-inflammatory (M2) macrophages. In addition, hydrogen treatment alone had no influence on the macrophage polarization. These indicate that hydrogen only affects the phenotype of macrophages under the pathological condition, and the reduction in M1 macrophages is ascribed to the inhibited macrophage activation, but not to the shift from M2 phenotype to M1 phenotype.
There is evidence showing the importance of bidirectional interactions between neurons and microglia in the context of ischemic stroke.33 Inose et al.34 found lysophosphatidylcholine from adjacent neurons could activate microglia in ischemic stroke penumbra. The study of Zhao et al.35 showed IL-4 from neurons served as a modulator of microglial pathways. Thus, microglia exposed to the supernatant from OGD-treated neurons served as an in vitro model of brain ischemia in this study.36 Our results showed the supernatant from OGD-treated neurons could increase the proportion of M1 macrophages, but the simultaneous hydrogen treatment inhibited the increase in M1 macrophages. However, the supernatant and hydrogen had no significant influence on the M2 macrophages.
This study had several limitations. First, LPS was used to stimulate macrophages as an inflammatory model, which may differ from the in vivo, so the lack of in vivo research will be solved in the future. In our study, we used cell lines, but not the primary macrophages and we only detected the microglia at 24 hours.
Taken together, LPS can significantly increase the pro-inflammatory macrophages, the supernatant from OGD-treated neurons may also elevate the proportion of pro-inflammatory macrophages, but both treatments have little influence on the anti-inflammatory macrophages. In addition, simultaneous hydrogen treatment inhibits the increase in pro-inflammatory macrophages, and has no influence on anti-inflammatory cells. This provides a new mechanism for the anti-inflammatory effect of hydrogen. However, more studies are needed to confirm our findings and further investigate the specific pathways underlying the hydrogen regulated polarization of macrophages.
Footnotes
Funding: This study was support by the National Natural Science Foundation of China, No. 81772015 (to LWW).
Conflicts of interest
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.
Financial support
This study was support by the National Natural Science Foundation of China, No. 81772015 (to LWW).
Institutional review board statement
The study was approved by the Ethics Committee of the Navy Medical University, Shanghai, China (approval number: 20170236).
Copyright license agreement
The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement
Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check
Checked twice by iThenticate.
Peer review
Externally peer reviewed.
REFERENCES
- 1.Chen RL, Balami JS, Esiri MM, Chen LK, Buchan AM. Ischemic stroke in the elderly: an overview of evidence. Nat Rev Neurol. 2010;6:256–265. doi: 10.1038/nrneurol.2010.36. [DOI] [PubMed] [Google Scholar]
- 2.Writing Group M, Mozaffarian D, Benjamin EJ, et al. Heart disease and stroke statistics-2016 update: a report from the American Heart Association. Circulation. 2016;133:e38–360. doi: 10.1161/CIR.0000000000000350. [DOI] [PubMed] [Google Scholar]
- 3.Karikari TK, Charway-Felli A, Hoglund K, Blennow K, Zetterberg H. Commentary: global, regional, and national burden of neurological disorders during 1990-2015: a systematic analysis for the Global Burden of Disease Study 2015. Front Neurol. 2018;9:201. doi: 10.3389/fneur.2018.00201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Prentice H, Modi JP, Wu JY. Mechanisms of neuronal protection against excitotoxicity, endoplasmic reticulum stress, and mitochondrial dysfunction in stroke and neurodegenerative diseases. Oxid Med Cell Longev. 2015;2015:964518. doi: 10.1155/2015/964518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Puyal J, Ginet V, Clarke PG. Multiple interacting cell death mechanisms in the mediation of excitotoxicity and ischemic brain damage: a challenge for neuroprotection. Prog Neurobiol. 2013;105:24–48. doi: 10.1016/j.pneurobio.2013.03.002. [DOI] [PubMed] [Google Scholar]
- 6.Elkind MS, Cheng J, Rundek T, Boden-Albala B, Sacco RL. Leukocyte count predicts outcome after ischemic stroke: the Northern Manhattan Stroke Study. J Stroke Cerebrovasc Dis. 2004;13:220–227. doi: 10.1016/j.jstrokecerebrovasdis.2004.07.004. [DOI] [PubMed] [Google Scholar]
- 7.Mena H, Cadavid D, Rushing EJ. Human cerebral infarct: a proposed histopathologic classification based on 137 cases. Acta Neuropathol. 2004;108:524–530. doi: 10.1007/s00401-004-0918-z. [DOI] [PubMed] [Google Scholar]
- 8.Moskowitz MA, Lo EH, Iadecola C. The science of stroke: mechanisms in search of treatments. Neuron. 2010;67:181–198. doi: 10.1016/j.neuron.2010.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hambardzumyan D, Gutmann DH, Kettenmann H. The role of microglia and macrophages in glioma maintenance and progression. Nat Neurosci. 2016;19(1):20–27. doi: 10.1038/nn.4185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yona S, Kim KW, Wolf Y, et al. Fate mapping reveals origins and dynamics of monocytes and tissue macrophages under homeostasis. Immunity. 2013;38:79–91. doi: 10.1016/j.immuni.2012.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Donnan GA, Fisher M, Macleod M, Davis SM. Stroke. Lancet. 2008;371:1612–1623. doi: 10.1016/S0140-6736(08)60694-7. [DOI] [PubMed] [Google Scholar]
- 12.Jin R, Yang G, Li G. Inflammatory mechanisms in ischemic stroke: role of inflammatory cells. J Leukoc Biol. 2010;87:779–789. doi: 10.1189/jlb.1109766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hu X, Li P, Guo Y, et al. Microglia/macrophage polarization dynamics reveal novel mechanism of injury expansion after focal cerebral ischemia. Stroke. 2012;43:3063–3070. doi: 10.1161/STROKEAHA.112.659656. [DOI] [PubMed] [Google Scholar]
- 14.Perego C, Fumagalli S, De Simoni MG. Temporal pattern of expression and colocalization of microglia/macrophage phenotype markers following brain ischemic injury in mice. J Neuroinflammation. 2011;8:174. doi: 10.1186/1742-2094-8-174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Nathan CF, Murray HW, Wiebe ME, Rubin BY. Identification of interferon-gamma as the lymphokine that activates human macrophage oxidative metabolism and antimicrobial activity. J Exp Med. 1983;158:670–689. doi: 10.1084/jem.158.3.670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chen Z, Jalabi W, Shpargel KB, et al. Lipopolysaccharide-induced microglial activation and neuroprotection against experimental brain injury is independent of hematogenous TLR4. J Neurosci. 2012;32:11706–11715. doi: 10.1523/JNEUROSCI.0730-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kobayashi K, Imagama S, Ohgomori T, et al. Minocycline selectively inhibits M1 polarization of microglia. Cell Death Dis. 2013;4:e525. doi: 10.1038/cddis.2013.54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sachet M, Liang YY, Oehler R. The immune response to secondary necrotic cells. Apoptosis. 2017;22:1189–1204. doi: 10.1007/s10495-017-1413-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Voss J, Ford CA, Petrova S, et al. Modulation of macrophage antitumor potential by apoptotic lymphoma cells. Cell Death Differ. 2017;24:971–983. doi: 10.1038/cdd.2016.132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Szulzewsky F, Pelz A, Feng X, et al. Glioma-associated microglia/macrophages display an expression profile different from M1 and M2 polarization and highly express Gpnmb and Spp1. PLoS One. 2015;10:e0116644. doi: 10.1371/journal.pone.0116644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Brauer RW, Way RO. Relative narcotic potencies of hydrogen, helium, nitrogen, and their mixtures. J Appl Physiol. 1970;29:23–31. doi: 10.1152/jappl.1970.29.1.23. [DOI] [PubMed] [Google Scholar]
- 22.Shin W. Medical applications of breath hydrogen measurements. Anal Bioanal Chem. 2014;406:3931–3939. doi: 10.1007/s00216-013-7606-6. [DOI] [PubMed] [Google Scholar]
- 23.Sano M, Suzuki M, Homma K, et al. Promising novel therapy with hydrogen gas for emergency and critical care medicine. Acute Med Surg. 2018;5:113–118. doi: 10.1002/ams2.320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ohsawa I, Ishikawa M, Takahashi K, et al. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat Med. 2007;13:688–694. doi: 10.1038/nm1577. [DOI] [PubMed] [Google Scholar]
- 25.Sun LQ, Guo GL, Zhang S, Yang LL. Effects of MicroRNA-592-5p on Hippocampal neuron injury following hypoxic-ischemic brain damage in neonatal mice - involvement of PGD2/DP and PTGDR. Cell Physiol Biochem. 2018;45:458–473. doi: 10.1159/000486923. [DOI] [PubMed] [Google Scholar]
- 26.Li H, Chen O, Ye Z, et al. Inhalation of high concentrations of hydrogen ameliorates liver ischemia/reperfusion injury through A2A receptor mediated PI3K-Akt pathway. Biochem Pharmacol. 2017;130:83–92. doi: 10.1016/j.bcp.2017.02.003. [DOI] [PubMed] [Google Scholar]
- 27.Zhou Z, Lu J, Liu WW, et al. Advances in stroke pharmacology. Pharmacol Ther. 2018;191:23–42. doi: 10.1016/j.pharmthera.2018.05.012. [DOI] [PubMed] [Google Scholar]
- 28.Hankey GJ. Stroke. Lancet. 2017;389:641–654. doi: 10.1016/S0140-6736(16)30962-X. [DOI] [PubMed] [Google Scholar]
- 29.Dziedzic T. Systemic inflammation as a therapeutic target in acute ischemic stroke. Expert Rev Neurother. 2015;15:523–531. doi: 10.1586/14737175.2015.1035712. [DOI] [PubMed] [Google Scholar]
- 30.Kigerl KA, Gensel JC, Ankeny DP, Alexander JK, Donnelly DJ, Popovich PG. Identification of two distinct macrophage subsets with divergent effects causing either neurotoxicity or regeneration in the injured mouse spinal cord. J Neurosci. 2009;29:13435–13444. doi: 10.1523/JNEUROSCI.3257-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Liu YC, Zou XB, Chai YF, Yao YM. Macrophage polarization in inflammatory diseases. Int J Biol Sci. 2014;10:520–529. doi: 10.7150/ijbs.8879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bok E, Chung YC, Kim KS, Baik HH, Shin WH, Jin BK. Modulation of M1/M2 polarization by capsaicin contributes to the survival of dopaminergic neurons in the lipopolysaccharide-lesioned substantia nigra in vivo. Exp Mol Med. 2018;50:76. doi: 10.1038/s12276-018-0111-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Zhao SC, Ma LS, Chu ZH, Xu H, Wu WQ, Liu F. Regulation of microglial activation in stroke. Acta Pharmacol Sin. 2017;38:445–458. doi: 10.1038/aps.2016.162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Inose Y, Kato Y, Kitagawa K, Uchiyama S, Shibata N. Activated microglia in ischemic stroke penumbra upregulate MCP-1 and CCR2 expression in response to lysophosphatidylcholine derived from adjacent neurons and astrocytes. Neuropathology. 2015;35:209–223. doi: 10.1111/neup.12182. [DOI] [PubMed] [Google Scholar]
- 35.Zhao X, Wang H, Sun G, Zhang J, Edwards NJ, Aronowski J. Neuronal Interleukin-4 as a modulator of microglial pathways and ischemic brain damage. J Neurosci. 2015;35:11281–11291. doi: 10.1523/JNEUROSCI.1685-15.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Zhou JM, Gu SS, Mei WH, Zhou J, Wang ZZ, Xiao W. Ginkgolides and bilobalide protect BV2 microglia cells against OGD/reoxygenation injury by inhibiting TLR2/4 signaling pathways. Cell Stress Chaperones. 2016;21:1037–1053. doi: 10.1007/s12192-016-0728-y. [DOI] [PMC free article] [PubMed] [Google Scholar]