

Editor’s Note
The brain, like other parts of the body, needs to maintain “homeostasis” (a constant state) to function, and that requires continuous removal of metabolic waste. For decades, the brain’s waste-removal system remained a mystery to scientists. A few years ago, a team of researchers—with the help of our author—finally found the answer. This discovery—dubbed the glymphatic system— will help us understand how toxic waste accumulates in devastating disorders such as Alzheimer’s disease and point to possible strategies to prevent it.
In early February 2012, I received a note from Maiken Nedergaard, a renowned neuroscientist at the University of Rochester whom I knew from our time as medical students at the University of Copenhagen. She explained that her team had discovered important features of a new system that transports the fluid that surrounds the brain—a substance called cerebrospinal fluid (CSF). The discovery of how this fluid was transported in the brain, she believed, was the key to understanding how waste is cleared from the brain.
Nedergaard’s work with the non-neuronal brain cells called “astroglia” had led her to suspect that these cells might play a role in CSF transport and brain cleansing. She was inspired by an older study’ which showed that CSF could rapidly penetrate into channels along the brain vasculature, and astroglial cells structurally help create these channels. Now she needed help with visualizing the system in the whole brain to confirm her suspicions. Her team needed imaging scientists like myself who might be able to visualize the unique CSF flow patterns in a rodent brain and shed light on the new system. Because I had experience and expertise in imaging CSF in the small rodent brain and spinal cord, I was equipped to take on this new challenge.
Initially, I did not fully comprehend the scope of how important the astroglial cells were in the process of CSF transport and brain waste clearance, even though it had been known for over 100 years that CSF circulates in the brain. As far back as 1872, Heinrich Quincke first injected a dye directly into the CSF of animals and documented its transport pattern through the central nervous system (CNS).1 Quincke discovered that CSF surrounding the spinal cord communicated with CSF covering the brain, and also that the dye in CSF could leak out along nerves and large vessels exiting the brain and spine.2 As an anesthesiologist, I knew about CSF transport since I had provided spinal anesthesia to so many patients over the years, based on well-founded experience.
So I went to visit Nedergaard’s team in Rochester and learned the origins of her discovery. A member of her research team, Jeffrey Iliff, provided some of the backstory on how Nedergaard had gotten to this point. She had found a forgotten paper published by M.L. Rennels and Patricia Grady in 1985, which convincingly asserted that CSF was transported rapidly into the brain along a channel network that surrounded the entire vasculature.3 While it seemed odd that these exciting findings had been buried prematurely, the reason was not uncommon: other scientists had never replicated them. During my visit Iliff presented data that demonstrated that they had successfully reproduced the findings by using state-of-the-art optical imaging techniques and fluorescent dyes administered into the CSF in brains of transgenic mice.
Although I did not understand the significance all at once, it was fascinating to learn about data that had the potential to shed new light on brain research. What, specifically, did the data show? By visualizing a small surface area of the brain at very high spatial and temporal resolution, they revealed that CSF transport of dyes occurred within channels surrounding each artery, arteriole, and capillary (see Figure 1 above). This peri-arterial transport was relatively fast, but not as fast as blood flow—it was measurable in minutes rather than seconds.4 The study also demonstrated that pulsation of the arteries was an important driver of the transport process,5 which made sense because CSF transport in humans was known to be “pulsatile” and influenced by physiological forces such as heart rate and respiration.
Figure 1.
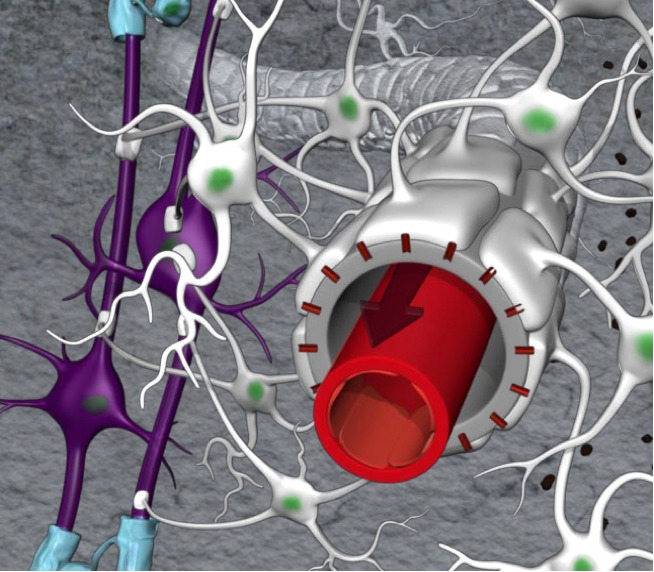
Peri-arterial channels are designed as part of a coaxial system. The outer perimeter of the channels is made up by astroglial endfeet decorated with AQP4 water channels. Courtesy of Helene Benveniste.
Intriguingly, the data also revealed that while small-sized dye molecules exited the peri-arterial channels almost immediately, large dye molecules got stuck, suggesting a barrier that prevented them from crossing into the space between brain cells. This difference in the transport of small and large dye molecules would lead to the discovery of how these channels are built and function. It turns out that the inner perimeter of the channels is made up of the blood vessel wall and the outer perimeter by the endfeet of the astroglial cells (see Figure 1 above). The endfeet of the astroglial cells are structures that extend from its cell body (see Figure 1 above). Small gaps between the endfeet determine the size of dye molecules that can move across and into the interstitial fluid (ISF).
In other experiments, the Rochester team studied the importance of the water channels (aquaporin 4, or AQP4) that are positioned strategically on the astroglial endfeet, so that they face the vessels. This part of the discovery was the most important new finding because nobody knew that these channels were crucial for the fast transport of CSF (and substances dissolved in it) from the peri-arterial space and into the space between the cells ( ‘interstitial space’).4 But the most important part of the story emerged from experiments showing that amyloid-beta (Aβ)—the toxic metabolite that accumulates in Alzheimer’s disease (AD)—was transported away from the brain via this new system (see Figure 2 below). This was important because it suggested that preserving this transport function across the lifespan would help maintain healthy brain homeostasis and possibly prevent AD.
Figure 2.
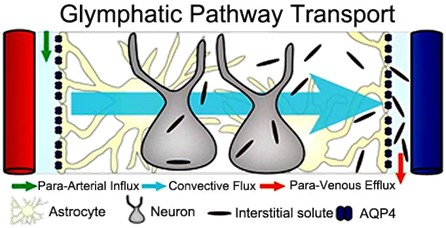
Principle of glymphatic transport in rodent brain. Peri-arterial inflow of cerebrospinal fluid (CSF) enters the brain tissue facilitated by astrocytic endfeet AQP4 water channels; mixes with interstitial fluid and removes the waste products into peri-venous space. Courtesy of Jeffrey Iliff.
Because the astroglia played a key role in the transport of CSF and waste solutes, they named the clearance system the “glymphatic” pathway to highlight “glia” as well as its functional similarities to the authentic “lymphatic” system outside the brain, which also clears waste. When I departed after two days of intense discussions, I understood that the following components were essential for glymphatic function: 1) normal CSF production and peri-vascular transport, 2) AQP4 water channels on the astroglial end-feet, 3) vascular pulsation, 4) peri-arterial influx of CSF, 5) peri-venous outflow or clearance, and 6) an intact skull.4
Transport in Whole Brain
Back in my lab at Brookhaven National Laboratory, I started working with my colleague and MR physicist, Hedok Lee, to develop a method to visualize glymphatic transport in the rat brain, based on magnetic resonance imaging (MRI) that could also be used someday to demonstrate the existence of a similar system in the human brain. We obsessively executed experiments focused on the glymphatic pathway over the next two years, first reporting our findings in the Journal of Clinical Investigation in 2013.6
We used a rat model to further validate the system data that Nedergaard’s team had discovered in a mouse brain; this benefitted the visualization of glymphatic transport by MRI imaging because the rat brain is larger. In addition, we administered into CSF a type of MRI-detectable tracer (so-called MR “contrast”) used routinely by radiologists in clinical settings. Through repeated trial-and-error experiments we sought a small molecular weight contrast molecule which could move very quickly from CSF into the whole brain within 30 minutes, as has been observed by Nedergaard’s team. But it seemed impossible. The transport process demonstrated by MRI was clearly not as fast as initially judged by the optical techniques.6 But the MRI experiments were encouraging because we were able to visualize transport into the brain from CSF similarly to what the Nedergaard team had described, and this would enable us and others to start investigating if such a transport system existed in the human brain.
Indeed, a few years later, Geir Ringstad and colleagues characterized CSF transport using the same approach, in normal subjects and individuals with “idiopathic normal pressure hydrocephalus” (iNPH), a condition (also termed Hakim’s syndrome) found in seniors and associated with dilation of the cerebral ventricles and dementia.7 Here, they documented transport of MR contrast from CSF along the arteries and into the brain. The process was much slower in a human brain compared to a small rat brain, probably due to differences in 1) heart rate (300–400 beats/min versus 60–70 beats/min), 2) turnover and production rates of CSF, 3) brain size, and 4) the differences in expression pattern of AQP4 between the two species.
Although these more recent MRI studies do not confirm the existence of a glymphatic system in the human brain, they strongly support the existence of peri-arterial CSF and solute influx and exchange of CSF and ISF, as well as clearance of contrast (as a surrogate of waste) from the brain over time. Importantly, in iNPH subjects, the characteristic pattern of CSF transport of contrast into and out of the brain was slowed down,7 suggesting impaired glymphatic transport of waste.8
The Glymphatic Concept Evolves
As studies from other laboratories emerged, we began to appreciate the complexity of the glymphatic system. One question that kept me particularly perplexed was: “How does glymphatic waste actually drain out of the brain?” Although we have known for decades that substances in CSF and within the brain can drain to the lymphatic system outside the brain, the exact route remains undefined.9 10 Frustratingly, none of the MRI techniques we had developed to visualize glymphatic transport appeared to be sufficiently sensitive to capture these exit pathways.
We kept experimenting with different CSF contrast molecules and administration protocols to improve the sensitivity of the MRI platform, but visualization of the drainage routes remained elusive. Fortunately, scientists in other labs soon after reported on a new pathway downstream from the glymphatic system that shed light on this important question. They discovered that brain waste could drain to authentic lymph vessels (LV) possibly imbedded in the dura ( the outermost and thickest membrane) covering the brain and spine.11–13 Using molecular markers specific to LV,14 they demonstrated an LV network at the level of the meninges of rodents, non-human primates, and humans, configured in a characteristic anatomical pattern along the larger dural veins and arteries and along the cranial nerves.11 These LVs were shown to be functionally capable of draining waste solutes from the brain.12
While the blood vessels in the brain and spine are “tight” due to the presence of the blood-brain-barrier (BBB), the meningeal blood vessels are leaky. This allowed an MR contrast agent to escape into the interstitial space of the dura, enabling scientists to visualize memLVs in the live human brain for the first time. (see Figure 3 below).15 This demonstration was met with much excitement given the potential importance of the waste clearance system for understanding AD and other dementias, including iNPH.
Figure 3.
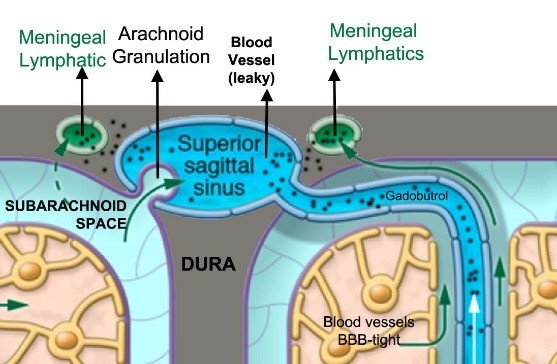
Principle by which meningeal lymphatics can be visualized by contrast enhanced MRI. The MR contrast agent (e.g. Gadobutrol) is administered i.v. and circulates systemically through the vascular compartment. The blood vessels in the brain are tight but when the contrast reaches the leaky dura vessels the contrast agent travels across the interstitial space and into the meningeal lymphatics. Courtesy of Helene Benveniste.
However, it is still not clear how CSF and its waste solutes drain from the brain and into the meningeal lymphatic vessels. Nor do we know how important the meningeal lymphatic network is for CNS waste drainage, as CSF and its solutes can also exit via other pathways.16 The pathophysiological significance of the meningeal LVs in the setting of, for example, stroke or brain edema following an injury, is likewise unknown. The newer imaging approaches to visualize the LVs and brain waste drainage may help answer these important questions.
AQP4 Water Channels
The high density of AQP4 water channels on astroglial endfeet (see Figure 1 above) appears to be essential for efficient glymphatic transport.4 Nevertheless, as with many new discoveries, controversies have emerged. While one recent study refuted the dependency of glymphatic transport on the AQP4 water channels, four others using various knockout rodent models reconfirmed the importance of AQP4 channels in CSF transport and waste clearance.17,18 Histochemistry studies of the mouse brain have shown that AQP4 expression is not uniform across the brain,19 which could suggest that these inconsistent findings reflect regional heterogeneity in glymphatic transport function.6
In the mouse brain, AQP4 expression is lowest in the cortex, and highest in the cerebellum and spinal cord.19 There are also differences between the subcellular distribution pattern of AQP4 expression in human and mouse brains.20 The degree of perivascular AQP4 astroglia endfeet expression has been found to be one-third lower in humans.20 This is an important observation, as it might imply more efficient glymphatic waste drainage in the small mouse brain compared to the human brain.
Benjamin Kress and colleagues reported that the AQP4 polarization on the astroglia endfeet surrounding cortical-penetrating arterioles (but not capillaries) was significantly reduced in old (18 month) compared to young mice.21 Furthermore, using immunofluorescence in human tissue, Iliff’s team demonstrated that perivascular AQP4 localization was preserved in cognitively intact individuals, but not in individuals with AD.22
Glymphatic Transport, Waste Clearance, and Sleep
One exciting study concerning the glymphatic pathway reported the enhancing effect of sleep on influx and clearance of waste solutes, including Aβ. In this rodent study, glymphatic influx increased by 95 percent and Aβ was cleared twice as fast in the cortex during slow wave sleep (or state of anesthesia with ketamine/xylazine), than during wakefulness.23
The faster glymphatic transport and waste clearance during sleep was associated with a 40–60 percent volume increase in the cortical ISF space; and the ISF space was found to be consistently smaller in the awake state than after administration of an anesthetic cocktail such as ketamine/xylazine (which is another way to induce a sleep like state).23 Under normal (awake) conditions, the ISF space volume is about 20 percent, which is relatively restrictive to waste solute transport compared to slow wave sleep states where it expands to 40 percent (at least in the cortex), and which would allow more CSF to enter the glymphatic pathway to exchange with ISF.23 It is unknown if these effects of sleep on ISF space volume are ubiquitous in the brain, or if only certain regions are affected.
The underlying mechanism for ISF space volume expansion (and cell volume shrinkage) during sleep involves changing levels of arousal. One of the key brain regions mediating arousal is the locus coeruleus, and the neurotransmitter it uses is norepinephrine (NE). Nedergaard’s team demonstrated that suppression of central NE transmission (and locus coeruleus activity) by adrenergic receptor antagonists administered via CSF increased glymphatic influx and ISF volume even though the mice in the study were not asleep.23
It is important to emphasize that natural sleep is not the same as “sleep” or “unconsciousness” induced by various anesthetics because different anesthetic cocktails do not affect glymphatic transport to the same degree.24 For example, we showed that anesthetic drugs selectively targeting NE transmission (e.g., dexmedetomidine, which blocks the alpha-2 receptor) combined with low dose inhalational anesthetics increase ISF solute transport and CSF volume more than anesthesia with inhalational agents only.24 Sleep positions also affect brain waste drainage (at least in rodents); for example, prone position decreases glymphatic transport compared to supine or lateral sleeping positions.25
Increasing evidence that sleep disturbances might contribute to AD, in part, by facilitating accumulation of Aβ in the brain is compatible with the association between sleep and glymphatic transport. In rodents, acute and chronic sleep disturbances increased ISF Aβ levels.26 Imaging studies have revealed associations between self-reports of shorter sleep duration and higher Aβ burden in the human brain.27–29 Most recently, we reported an increase in the metabolic waste product Aβ in a human brain after acute sleep deprivation, which is also consistent with the existence of a glymphatic system.30 Future studies with functional imaging tools able to track endogenous waste molecules in the human brain and visualize AQP4 channels may provide further insight into the clearance process in both normal and disease states.
Footnotes
Helene Benveniste, M.D., Ph.D., completed her M.D. in 1989 and Ph.D (Doctor of Medicine) in 1991 at the University of Copenhagen, Denmark. She trained in high field magnetic resonance imaging at the Center for In Vivo Microscopy at Duke University Medical Center and went on to take her Family Medicine internship and residency in Anesthesiology, also at Duke University. Benveniste has been actively engaged in translational research for more than 20 years at Brookhaven National Laboratory and Stony Brook University. In 2016, Benveniste moved to Yale University, where she joined the Department of Anesthesiology. Benveniste’s research program at Yale is focused on understanding how the brain’s glymphatic system (implicated in metabolic waste removal) is affected during different states of arousal (sleep, anesthesia and emergence from anesthesia) and in neurodegenerative diseases.
References
- 1.Benveniste H, Hof PR, Nedergaard M, Bechter K. Modern cerebrospinal fluid flow research and Heinrich Quincke’s seminal 1872 article on the distribution of cinnabar in freely moving animals. J Comp Neurol. 2015;523(13):2017–2018. doi: 10.1002/cne.23850. [DOI] [PubMed] [Google Scholar]
- 2.Bechter K, Benveniste H. Quinckes’ pioneering 19th centuries CSF studies may inform 21th centuries research. Neurol Psychiatry Brain Res. 2015;21(2):79–81. doi: 10.1016/j.npbr.2015.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Rennels ML, Gregory TF, Blaumanis OR, Fujimoto K, Grady PA. Evidence for a ‘paravascular’ fluid circulation in the mammalian central nervous system, provided by the rapid distribution of tracer protein throughout the brain from the subarachnoid space. Brain Res. 1985;326(1):47–63. doi: 10.1016/0006-8993(85)91383-6. [DOI] [PubMed] [Google Scholar]
- 4.Iliff JJ, Wang M, Liao Y, Plogg BA, Peng W, Gundersen GA, Benveniste H, Vates GE, Deane R, Goldman SA, Nagelhus EA, Nedergaard M. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid beta. Sci Transl Med. 2012;4(147) doi: 10.1126/scitranslmed.3003748. 147ra111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Iliff JJ, Wang M, Zeppenfeld DM, Venkataraman A, Plog BA, Liao Y, Deane R, Nedergaard M. Cerebral arterial pulsation drives paravascular CSF-interstitial fluid exchange in the murine brain. J Neurosci. 2013;33(46):18190–18199. doi: 10.1523/JNEUROSCI.1592-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Iliff JJ, Lee H, Yu M, Feng T, Logan J, Nedergaard M, Benveniste H. Brain-wide pathway for waste clearance captured by contrast-enhanced MRI. J Clin Invest. 2013;123(3):1299–1309. doi: 10.1172/JCI67677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ringstad G, Vatnehol SAS, Eide PK. Glymphatic MRI in idiopathic normal pressure hydrocephalus. Brain. 2017;140(10):2691–2705. doi: 10.1093/brain/awx191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Benveniste H, Lee H, Volkow N. The Glymphatic Pathway: Waste Removal from the CNS via Cerebrospinal Fluid Transport. The Neuroscientist. 2017:1–12. doi: 10.1177/1073858417691030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bradbury MW, Cserr HF, Westrop RJ. Drainage of cerebral interstitial fluid into deep cervical lymph of the rabbit. Am J Physiol. 1981;240(4):F329–336. doi: 10.1152/ajprenal.1981.240.4.F329. [DOI] [PubMed] [Google Scholar]
- 10.Boulton M, Flessner M, Armstrong D, Hay J, Johnston M. Determination of volumetric cerebrospinal fluid absorption into extracranial lymphatics in sheep. Am J Physiol. 1998;274(1 Pt 2):R88–96. doi: 10.1152/ajpregu.1998.274.1.R88. [DOI] [PubMed] [Google Scholar]
- 11.Antila S, Karaman S, Nurmi H, Airavaara M, Voutilainen MH, Mathivet T, Chilov D, Li Z, Koppinen T, Park JH, Fang S, Aspelund A, Saarma M, Eichmann A, Thomas JL, Alitalo K. Development and plasticity of meningeal lymphatic vessels. J Exp Med. 2017;214(12):3645–3667. doi: 10.1084/jem.20170391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Aspelund A, Antila S, Proulx ST, Karlsen TV, Karaman S, Detmar M, Wiig H, Alitalo K. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J Exp Med. 2015;212(7):991–999. doi: 10.1084/jem.20142290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Louveau A, Smirnov I, Keyes TJ, Eccles JD, Rouhani SJ, Peske JD, Derecki NC, Castle D, Mandell JW, Lee KS, Harris TH, Kipnis J. Structural and functional features of central nervous system lymphatic vessels. Nature. 2015;523(7560):337–341. doi: 10.1038/nature14432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Truman LA, NAG, Bentley KL, Ruddle NH. Lymphatic vessel function in head and neck inflammation. Lymphat Res Biol. 2013;11(3):187–192. doi: 10.1089/lrb.2013.0013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Absinta M, Ha SK, Nair G, Sati P, Luciano NJ, Palisoc M, Louveau A, Zaghloul KA, Pittaluga S, Kipnis J, Reich DS. Human and nonhuman primate meninges harbor lymphatic vessels that can be visualized noninvasively by MRI. Elife. 2017;6 doi: 10.7554/eLife.29738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ma Q, Ineichen BV, Detmar M, Proulx ST. Outflow of cerebrospinal fluid is predominantly through lymphatic vessels and is reduced in aged mice. Nat Commun. 2017;8(1):1434. doi: 10.1038/s41467-017-01484-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mestre H, Kress BT, Zou W, Pu T, Murlidharan G, Castellanos Rivera RM, Simon MJ, Pike MM, Plog BA, Xavier ALR, Thrane AS, Lundgaard I, Thomas JH, Xiao M, Asokan A, Iliff JJ, Nedergaard M. Aquaporin-4 dependent glymphatic solute transport in rodent brain. bioRxiv. 2017 doi: 10.7554/eLife.40070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Smith AJ, Yao X, Dix JA, Jin BJ, Verkman AS. Test of the ‘glymphatic’ hypothesis demonstrates diffusive and aquaporin-4-independent solute transport in rodent brain parenchyma. Elife. 2017;6 doi: 10.7554/eLife.27679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hoddevik EH, Khan FH, Rahmani S, Ottersen OP, Boldt HB, Amiry-Moghaddam M. Factors determining the density of AQP4 water channel molecules at the brain-blood interface. Brain Struct Funct. 2017;222(4):1753–1766. doi: 10.1007/s00429-016-1305-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Eidsvaag VA, Enger R, Hansson HA, Eide PK, Nagelhus EA. Human and mouse cortical astrocytes differ in aquaporin-4 polarization toward microvessels. Glia. 2017;65(6):964–973. doi: 10.1002/glia.23138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kress BT, Iliff JJ, Xia M, Wang M, Wei HS, Zeppenfeld D, Xie L, Kang H, Xu Q, Liew JA, Plog BA, Ding F, Deane R, Nedergaard M. Impairment of paravascular clearance pathways in the aging brain. Ann Neurol. 2014;76(6):845–861. doi: 10.1002/ana.24271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zeppenfeld DM, Simon M, Haswell JD, D’Abreo D, Murchison C, Quinn JF, Grafe MR, Woltjer RL, Kaye J, Iliff JJ. Association of Perivascular Localization of Aquaporin-4 With Cognition and Alzheimer Disease in Aging Brains. JAMA Neurol. 2017;74(1):91–99. doi: 10.1001/jamaneurol.2016.4370. [DOI] [PubMed] [Google Scholar]
- 23.Xie L, Kang H, Xu Q, Chen MJ, Liao Y, Thiyagarajan M, O’Donnell J, Christensen DJ, Nicholson C, Iliff JJ, Takano T, Deane R, Nedergaard M. Sleep drives metabolite clearance from the adult brain. Science. 2013;342(6156):373–377. doi: 10.1126/science.1241224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Benveniste H, Lee H, Ding F, Sun Q, Al-Bizri E, Makaryus R, Probst S, Nedergaard M, Stein EA, Lu H. Anesthesia with Dexmedetomidine and Low-dose Isoflurane Increases Solute Transport via the Glymphatic Pathway in Rat Brain When Compared with High-dose Isoflurane. Anesthesiology. 2017;127(6):976–988. doi: 10.1097/ALN.0000000000001888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lee H, Xie L, Yu M, Kang H, Feng T, Deane R, Logan J, Nedergaard M, Benveniste H. The Effect of Body Posture on Brain Glymphatic Transport. J Neurosci. 2015;35(31):11034–11044. doi: 10.1523/JNEUROSCI.1625-15.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kang J-E, Lim MM, Bateman RJ, Lee JJ, Smyth LP, Cirrito JR, Fujiki N, Nishino S, Holtzman DM. Amyloid-β dynamics are regulated by orexin and the sleep-wake cycle. Science. 2009;326(5955):1005–1007. doi: 10.1126/science.1180962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Spira AP, Gamaldo AA, An Y, Wu MN, Simonsick EM, Bilgel M, Zhou Y, Wong DF, Ferrucci L, Resnick SM. Self-reported sleep and β-amyloid deposition in community-dwelling older adults. JAMA neurology. 2013;70(12):1537–1543. doi: 10.1001/jamaneurol.2013.4258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Brown BM, Rainey-Smith SR, Villemagne VL, Weinborn M, Bucks RS, Sohrabi HR, Laws SM, Taddei K, Macaulay SL, Ames D. The relationship between sleep quality and brain amyloid burden. Sleep. 2016;39(5):1063–1068. doi: 10.5665/sleep.5756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sprecher KE, Bendlin BB, Racine AM, Okonkwo OC, Christian BT, Koscik RL, Sager MA, Asthana S, Johnson SC, Benca RM. Amyloid burden is associated with self-reported sleep in nondemented late middle-aged adults. Neurobiology of aging. 2015;36(9):2568–2576. doi: 10.1016/j.neurobiolaging.2015.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Shokri-Kojori E, Wang GJ, Wiers CE, Demiral SB, Guo M, Kim SW, Lindgren E, Ramirez V, Zehra A, Freeman C, Miller G, Manza P, Srivastava T, De Santi S, Tomasi D, Benveniste H, Volkow ND. beta-Amyloid accumulation in the human brain after one night of sleep deprivation. Proc Natl Acad Sci U S A. 2018;115(17):4483–4488. doi: 10.1073/pnas.1721694115. [DOI] [PMC free article] [PubMed] [Google Scholar]