

Abstract
A bacterial isolate capable of tolerating 30 mM silver nitrate (AgNO3) was recovered from soil contaminated with industrial waste. The isolate was identified by 16S rRNA as Enterobacter cloacae Ism26 (KP988024) and its capability to synthesize silver nanoparticles (AgNPs) was investigated. AgNPs were produced by mixing 1 mM AgNO3 solution with bacterial cell lysate under light conditions. The UV-Vis spectrum of the aqueous medium containing AgNPs exhibited a peak at 440 nm corresponding to the surface plasmon resonance of the AgNPs. The crystalline nature of the particles was confirmed by X-ray difractometer. High-resolution transmission electron microscopy revealed that the AgNPs were spherical and well dispersed and ranged in size from 7 to 25 nm. The average size range of the produced AgNPs was confirmed by dynamic light scattering. Fourier transform infrared spectroscopy revealed possible involvement of reductive groups on the surface of the nanoparticles. The biosynthesized AgNPs were stable for 6 months and inhibited both gram-positive and gram-negative bacteria. This work describes the exploitation of a low-cost biomaterial and an easy method for the synthesis of AgNPs with desirable and advantageous characteristics.
Keywords: Silver nanoparticles, light, antibacterial, Enterobacter
1. Introduction
Synthesis of metal nanoparticles is an emerging area of nanoscience and nanotechnology since these particles exhibit peculiar properties based on their size, distribution, and morphology (Rai et al., 2014) . These unique properties may provide new approaches and solutions to several industrial and environmental challenges in the areas of medicine, solar energy conversion, catalysis, and water treatment (Khalil et al., 2014; Rai et al., 2014) .
Recently, there has been growing interest in the preparation and study of silver nanoparticles (AgNPs) due to their extensive applications in different fields including wound and burn healing and bone and dental implants, and for their antibacterial, fungal, viral, protozoal, and arthropodal activities (Klaus et al., 1999; Khalil et al., 2014; Rai et al., 2014) .
The ecofriendly methods for AgNP synthesis have emerged as green alternatives to conventional chemical and physical methods. Diefrent microorganisms such as Geobacter sulfurreducens, Morganella spp., and Pseudomonas stutzeri AG259 have been explored as potential biofactories for the synthesis and stabilization of AgNPs (Law et al., 2008; Parikh et al., 2008; Rai et al., 2009) . The effective metabolic activity of microorganisms allows the production of nanoparticles with defined size, shape, water solubility, and high reactivity, which qualify them to be among the favored choices for many applications (Rai et al., 2009) .
The present work describes the synthesis of AgNPs using a silver-resistant bacterial isolate recovered from soil subjected to industrial and sewage euflents. The effects of different conditions such as the incubation period, temperature, pH, AgNO3 concentration, and amount of bacterial cell lysate supernatant on the production of AgNPs were investigated. Characterization of the synthesized AgNPs was carried out using specific techniques and the antibacterial activity of the produced AgNPs was investigated against both gram-positive and gram-negative bacteria.
2. Materials and methods
2.1. Soil samples
Samples were collected from soil polluted with industrial wastewater near the Ismailia Canal in northeastern Egypt. Ten randomly selected soil samples were collected along one km in the El Qalyobia governorate in sterile bags.
2.2. Selective isolation of silver tolerant bacteria
A tenfold serial dilution was carried out and aliquots of 100 µL were inoculated on nutrient agar plates and incubated at EL-BAGHDADY et al. / Turk J Biol 35 °C for 24 h. Bacterial isolates were streaked on nutrient agar medium supplemented with silver nitrate (AgNO3) covering the range from 0.5 to 30 mM. The plates were then incubated at 35 °C for 24 h (Das et al., 2013) . After the incubation period, brown colonies growing on 30 mM AgNO3 were selected and stored on nutrient agar slants supplemented with the same concentration of AgNO .
2.3. Identification of the most tolerant bacterial isolates by 16S rRNA gene sequencing
The recovered isolate was identified by 16S rRNA (Edwards et al., 1991) where colony PCR was carried out to amplify the 16S rRNA gene for the selected isolates. A loopful of culture grown overnight was transferred to 50 µL of sterile distilled water and boiled for 5 min and 1 µL of cell suspension was used as the DNA template for PCR reaction. PCR was performed using Fermentas DreamTaq Master Mix according to the instruction manual. The universal primers PA (5’-AGAGTTTGATCCTGGCTCAG-3’) and PH (5’-ACGGCTACCTTGTTACGACT-3’) were used to amplify 1600 bp. The PCR conditions were adjusted to 5 min for initial denaturation at 94 °C followed by 25 cycles of 1 min at 94 °C, 1 min at 61 °C, and 1 min at 72 °C, and finally 7 min at 72 °C. The PCR products were subjected to electrophoresis on 1% agarose gel, visualized by staining with ethidium bromide dye, and examined by gel documentation system InGenius 3 (Syngene).
PCR products were purified using a QIAGEN extraction kit according to the manufacturer’s instructions before applying them to the DNA sequencer. Sequencing was performed at the Animal Health Research Institute, El Dokki, Egypt, using an automated uflorescent dye terminator sequencing method employing a DYEynamic ET Terminator Cycle Sequencing Kit (Amersham Pharmacia Biotech, ABI 3130).
All sequences were compared with accessible sequences in the database using BLAST software. Multiple sequence alignment and computation of sequence similarity were conducted using ClustalW within the BioEdit program.
2.4. Biosynthesis of silver nanoparticles
Flasks containing 100 mL of nutrient broth medium were inoculated with 100 µL (108 CFU) of the selected bacterial isolates and incubated at 35 °C and 180 rpm for 24 h. The bacterial cultures were then centrifuged at 7000 rpm for 10 min. Bacterial pellets were collected, washed twice with sterile double-distilled water, and sonicated for 120 s with 3 intervals of 40 s each using an ultrasonic homogenizer (4710 series, Cole-Parmer Instruments Co.). Bacterial cell lysate supernatant was mixed with 1 mM AgNO3 solution and incubated in light at 35 °C for 24 h (Ganesh Babu and Gunasekaran, 2009) .
AgNO3 (1 mM) and bacterial cell lysate supernatant without AgNO3 served as controls and were kept under the same conditions. After the incubation period, the mixtures were observed for the presence of brown color, which indicated the positive formation of AgNPs (Priyadarshini et al., 2013) .
2.5. Characterization of silver nanoparticles
2.5.1. UV-Vis spectrometry
Brown-colored solutions were scanned to detect the presence of the specific peaks (400–450 nm) for AgNPs using an Evolution 201 Scan UV-visible spectrophotometer (Thermo Scientific). Double distilled water was used as blank in setting up the base line.
2.5.2. X-ray diffraction (XRD)
Samples were obtained in a powder form using a freeze dryer (Edwards, model RV5). The specimens were analyzed in angular range from 10° to 80° (2θ) using the Shimadzu XRD-6000 computerized difractometer operating at 50 keV and 25 mA throughout the measurements (Physics Department, Faculty of Science, Ain Shams University). 2.5.3. Fourier transform infrared spectroscopy (FTIR) The organic bonds responsible for reduction, capping, and stability of AgNPs were examined using FTIR (Nicolet 6700) in the spectral range of 400–4000 cm–1 with resolution of 4 cm–1. 2.5.4. Dynamic light scattering (DLS) The particle sizes of AgNPs in the solution were measured by the DLS technique using a PSS-NICOMP particle sizer 380ZLS at the Egyptian Atomic Energy Authority. 2.5.5. High-resolution transmission electron microscopy (HRTEM) The AgNPs’ shapes and sizes were investigated by HRTEM at the Egyptian Petroleum Research Institute using a JEOL 2100 electron microscope operating at an accelerating voltage of 200 KV.
2.6. Optimization of AgNP production
Diefrent physicochemical factors were investigated to optimize the AgNP production. These factors included the reaction duration (0, 1, 2, 3, 4, 5, 6, 20, 24, and 48 h), temperature (25, 35, 45, 60, and 100 °C), pH (2, 3, 4, 5, 6, 7, 8, and 9), volumes of bacterial lysate supernatant (5, 10, 20, 30, 40, and 50 mL), and different AgNO 3 concentrations (1, 2, 3, 4, 5, 1/2, 1/4, and 1/8 mM). All factors were tested under light. All factors were tested in sequence and the result from each experiment was implemented in the next one.
2.7. Stability of AgNPs
The stability of the produced AgNPs was tested over a storage period of 6 months in the dark. A spectrophotometer (UVVis) was used to detect variations in the absorption peaks.
2.8. Antimicrobial activity of AgNPs
Biosynthesized AgNPs were centrifuged at 20,000 rpm for 10 min. The supernatant was decanted and pellets were further freeze-dried (Edwards, model RV5). The AgNP powder was suspended in sterile deionized water. The antibacterial activity was tested against the three gramnegative bacteria, E. coli ATCC 25922, E. coli 0157:H7 ATCC 35150, and Salmonella typhimurium ATCC 13311, along with two gram-positive bacteria, Staphylococcus aureus ATCC 43300 and Listeria monocytogenes ATCC 35152. Agar well difusion assay and microtiter plates were used to determine the minimum inhibitory concentration (MIC) using 0, 5, 10, 25, 50, 75, and 100 µg of AgNPs (Wiegand et al., 2008) .
2.9. Statistical analysis
The statistical analysis of microbiological activity was carried out using Microsoft Excel 2013. All data were calculated from at least three replicates and the standard errors for each datum were plotted.
3. Results
3.1. Isolation and identification of silver tolerant bacterial isolate
hTirty-seven isolates were recovered from nutrient agar media supplemented with 0.5 mM AgNO3. Only one isolate, designated as Ism26, was able to tolerate 30 mM AgNO3 and was considered as a potential candidate for the synthesis of AgNPs.
The colony color of Ism26 changed from buf to brown when grown on nutrient agar supplemented with AgNO3. The selected bacterial isolate (Ism26) was subcultured on nutrient agar medium supplemented with 30 mM AgNO3 throughout this study. Preliminary identification of the bacterial isolate Ism26 indicated that it belonged to Enterobacter sp.
3.2. Molecular identification of the selected bacterial isolate
PCR of 16S rRNA followed by partial sequence analysis identified isolate Ism26 as Enterobacter cloacae (99% similarity), and consequently it was submitted to GenBank under accession number KP988024.
3.3. Biosynthesis of AgNPs
In order to investigate the ability of E. cloacae Ism26 to synthesize AgNPs, it was grown in dark and light conditions. Observation of color change from colorless to brown and the detection of the characteristic peaks at 400– 460 nm were considered indicators for the presence of AgNPs. Growth in the light only resulted in the formation of AgNPs. Moreover, AgNPs were formed by mixing bacterial lysate supernatant of sonicated pellets with 1 mM AgNO3 followed by incubation under light with maximum absorption peak at 440 nm (Figures 1A and 1B).
Figure 1.
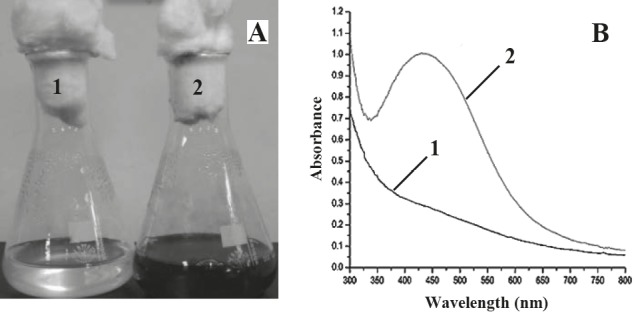
Formation of AgNPs by E. cloacae Ism26 under light. Visual inspection (A) and UV spectroscopic detection for the formation of AgNPs (B). 1: Control bacterial lysate supernatant, 2: bacterial lysate supernatant + AgNO3.
3.4. Effect of reaction incubation time
A gradual increase in the intensity of brown color was observed by increasing the incubation time until 48 h. Data indicated that the time required for maximum production of AgNPs by E. cloacae Ism26 was 20 h with maximum absorbance peak at 440 nm (Figure 2A).
Figure 2.

UV-Vis spectra showing the effect of reaction incubation time (A), temperature (B), pH (C), volume of bacterial lysate supernatant (D), AgNO3 concentration (E), and storage period (F) on AgNPs synthesized by E. cloacae Ism26.
3.5. Effect of reaction temperature
The optimum temperature for AgNP synthesis was 35 °C. It was also observed that increasing the temperature resulted in red shift of the absorption maximum (Figure 2B).
3.6. Effect of pH
The optimum AgNP synthesis was observed at pH 7, whereas the absorption decreased in both acidic and alkaline pH (Figure 2C).
3.7. Effect of different volumes of bacterial lysate supernatants
Maximum synthesis of AgNPs was detected when 40 mL of E. cloacae Ism26 bacterial lysate supernatant was mixed with AgNO3 solution (Figure 2D).
3.8. Effect of different AgNO 3 concentrations
The optimum AgNO 3 concentration for AgNP synthesis was 1 mM (Figure 2E). As the AgNO3 concentration increased from 0.125 to 1 mM, the absorption peak increased and became sharper. However, further increase in AgNO3 concentrations up to 5 mM resulted in shoulder (wide) peaks with lower absorbance as compared to that of the 1 mM peak.
3.9. Stability of AgNPs
The AgNPs were stored in the dark as a liquid solution at 4 °C. Only minor shifts were observed up until 180 days, which reflected the highly stable nature of the produced AgNPs (Figure 2F).
3.10. Characterization of AgNPs
3.10.1. High-resolution transmission electron microscopy (HRTEM)
HRTEM micrographs of the synthesized AgNPs showed that the size ranged from 7 to 25 nm and most particles were spherical with an average size of 15 nm (Figure 3).
Figure 3.
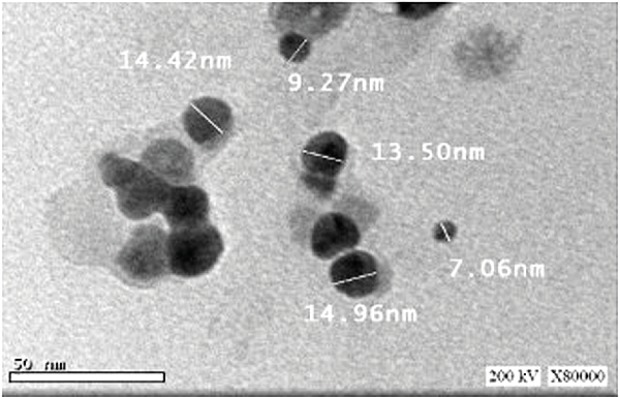
HRTEM micrograph of silver nanoparticles synthesized by E. cloacae Ism26.
3.10.2. X-ray diffraction (XRD)
Freeze-dried AgNPs synthesized by E. cloacae Ism26 (Figure 4) exhibited peaks at 2θ of 37.57°, 46.33°, 65.05°, and 76.80°.
Figure 4.
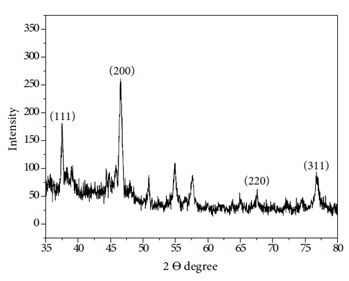
X-ray diffraction pattern of AgNPs synthesized using E. cloacae Ism26.
3.10.3. Dynamic light scattering (DLS)
AgNPs synthesized by E. cloacae Ism26 exhibited a narrow particle size range, from 5 to 50 nm, as shown in Figure 5. The mean diameter of AgNPs was 20.7 nm (SD: ±11). The particle size distribution showed that 25% were less than 15.8 nm, 50% less than 20.6 nm, 75% less than 28.2 nm, and 90% less than 37.9 nm (Figure 5).
Figure 5.
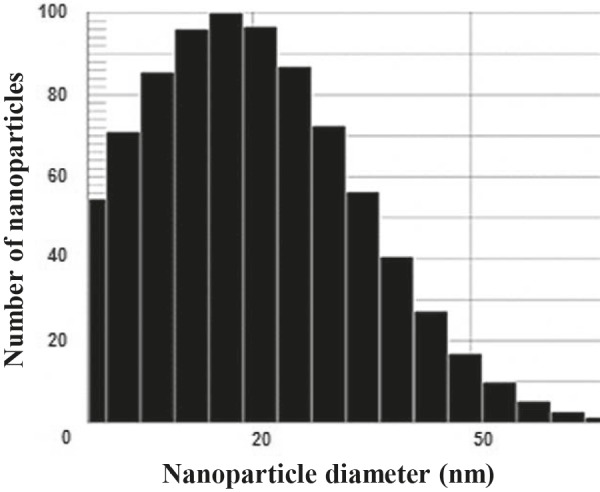
DLS size distribution of E. cloacae Ism26 AgNPs.
3.10.4. Fourier transform infrared (FTIR) spectroscopy
FTIR spectroscopy of synthesized AgNPs (Figure 6) showed a peak at 673 cm–1 that indicated C–H deformation in an aromatic ring at the alkene C–H bond, supporting the presence of an aromatic structure. Other peaks at 1640 cm–1 and 1386.2 cm–1 were observed, indicating CH deformation from the dimethyl group or symmetric stretching of COO-. The FTIR spectrum exhibited strong and sharp peaks in the range of 3361–3565 cm–1, suggesting the presence of NH and OH groups from the capping compounds. The peaks at 1640 cm –1 and 1515 cm–1 were due to C-C and N-H stretching vibrations of amide II in the protein linkage. The band at 1386 cm –1 was due to C-N stretching vibration of aliphatic amines.
Figure 6.

FTIR spectra of synthesized AgNPs.
3.11. Antibacterial activity of AgNPs
AgNPs synthesized by E. cloacae Ism26 significantly inhibited both gram-positive and gram-negative bacteria to different extents (Figure 7). The lowest concentration needed to inhibit the bacterial growth completely (MIC) using the synthesized AgNPs was determined using an overnight culture of both gram-positive and gram-negative bacteria (Table). E. coli ATCC 25922 was inhibited by 25 µg, followed by Staphylococcus aureus ATCC 43300 and Salmonella typhimurium ATCC 13311, which were inhibited by 50 µg, and then E. coli 0157:H7 ATCC 35150 and Listeria monocytogenes ATCC 35152, which were inhibited by 75 µg.
Figure 7.
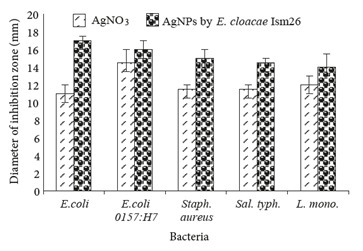
Antibacterial activity of AgNO3 and AgNPs against different gram-positive and gram-negative bacteria. E. coli: Escherichia coli, Sal. typh.: Salmonella typhimurium, Staph. aureus: Staphylococcus aureus, L. mono.: Listeria monocytogenes.
Table.
Minimum inhibitory concentration (MIC) of AgNPs synthesized by E. cloacae Ism26 against tested bacteria. E. coli: Escherichia coli, Sal. typh.: Salmonella typhimurium, Staph. aureus: Staphylococcus aureus, L. mono.: Listeria monocytogenes.
| AgNPs conc. (μg) | Optical density (OD) of tested bacteria at 600 nm | ||||
|---|---|---|---|---|---|
| E. coli | E. coli 0157:H7 | Sal. typh. | Staph. aureus | L. mono. | |
| 0 | 0.55 ± 0.04 | 0.55 ± 0.025 | 0.52 ± 0.02 | 0.55 ± 0.023 | 0.6 ± 0.024 |
| 5 | 0.055 ± 0.02 | 0.170 ± 0.017 | 0.178 ± 0.008 | 0.179 ± 0.017 | 0.47 ± 0.017 |
| 10 | 0.028 ± 0.009 | 0.16 ± 0.012 | 0.141 ± 0.007 | 0.104 ± 0.012 | 0.241 ± 0.016 |
| 25 | 0 0.114 ± 0.009 | 0.042 ± 0.006 | 0.013 ± 0.004 | 0.216 ± 0.006 | |
| 50 | 0 0.054 ± 0.011 | 0 | 0 | 0.034 ± 0.008 | |
| 75 | 0 | 0 | 0 | 0 | 0 |
| 100 | 0 | 0 | 0 | 0 | 0 |
4. Discussion
In this work, isolation of bacteria was carried out on a nutrient agar medium supplemented with different concentrations of AgNO3. The gradual increase in the concentration of AgNO3 in the nutrient agar medium resulted in isolation of bacteria that were able to tolerate high concentrations of AgNO . All bacterial cultures showed a change in color from creamy to brown. This could be due to the accumulation of silver inside the bacterial cells. A similar observation was demonstrated using Staphylococcus aureus, Pseudomonas aeruginosa, and E. coli O157:H7, which were grown on AgNO supplemented media. Lara et al. (2010) reported that this color change was an indication of silver resistance. The concentration of 30 mM used in this study was higher than that used in a previous study in which multidrug-resistant Pseudomonas aeruginosa tolerated 25 mM AgNO3 (Lara et al., 2010) .
In the present study, the bacterial isolate that tolerated a high concentration of AgNO3 was selected and identified as Enterobacter cloacae Ism26 (accession number KP988024).
The E. cloacae Ism26 AgNPs were produced only by sonicated cell lysate supernatant, but both culturefree supernatant and bacterial pellets did not produce AgNPs. This could be explained by the absence of the extracellular enzymes needed for AgNO3 reduction. Some earlier reports indicated that cell-free supernatants of gram-negative bacteria, e.g., K. pneumoniae and E. coli, as well as gram-positive bacteria, such as Bacillus indicus and Bacillus cecembensis, were able to synthesize AgNPs (Mokhtari et al., 2009; Natarajan et al., 2010; Shivaji et al., 2011) . Another study reported that AgNPs synthesis was carried out by whole bacterial cells (Seshadri et al., 2012) .
Synthesis of AgNPs was reported by Parikh et al. (2008) , using growing cultures of Morganella spp. Moreover, Bacillus strain CS 11 was able to synthesize AgNPs both extracellularly and intracellularly (Das et al., 2013) . Taking all these results collectively, it is possible to suggest that the components of the bacterial culture capable of synthesizing AgNPs vary according to bacterial species.
The present results showed that AgNP synthesis was successful under light incubation only and light might be a fundamental factor in the AgNP biosynthesis process. This is in good agreement with previous studies on both gramnegative and gram-positive bacteria (Mokhtari et al., 2009; Natarajan et al., 2010; Shivaji et al., 2011) . Light was found to support the AgNP synthesis by Klebsiella pneumoniae and the reaction did not show any color change when it took place in the dark (Mokhtari et al., 2009) . These results accentuate the role of light as an essential factor for some bacterial species in order to synthesize AgNPs. Nonetheless, a recent report demonstrated the synthesis of AgNPs under dark conditions by Streptomyces platensis (Morsy, 2015).
It is widely accepted that the ability of bacteria to synthesize AgNPs depends on two major factors: the first one is the availability of the proper enzymatic machinery, whether on the bacterial wall or inside the cell. The second factor is the presence of light, which facilitates the synthesis process (Nam et al., 2008) . The effect of light may be attributable to the activation of reducing agents present in the culture supernatant, which in turn would cause the release of electrons for the reduction of Ag+ to Ag0 nanoparticles (Wei et al., 2012) . It is worth mentioning that Eby et al. (2009) reported that hen egg white lysozyme was involved in the formation of AgNPs in the presence of light. However, the involvement of enzymatic activities in the formation of AgNPs in the dark was also reported (Kumar et al., 2007) .
The incubation period for optimum AgNPs synthesis needed by E. cloacae Ism26 was found to be 20 h. This is considered shorter than the time required by K. pneumoniae and E. coli, which required a 24-h incubation period (Minaeian et al., 2008) . The variation in the incubation period could be related to the conditions under which AgNP synthesis takes place (Elo,f 1998; Minaeian et al., 2008; Narayanan and Sakthivel, 2010) .
Among the factors that affected the AgNP biosynthesis by E. cloacae Ism26 were temperature and pH values. The present results showed that temperature had an appreciable effect on the synthesis rate and the size of AgNPs. Similar results were demonstrated by Bai et al. (2011) and Jeevan et al. (2012) using Rhodobacter sphaeroides and Pseudomonas aeruginosa, respectively, and the optimum temperature for AgNP synthesis was found to be 35 °C. Two other reports indicated that E. coli (Gurunathan et al., 2009) and Bacillus subtilis EWP-46 (Velmurugan et al., 2014) required a higher temperature (60 °C) to synthesize AgNPs.
Our results showed that the optimum pH for AgNP synthesis was attained with a neutral medium. This coincide with previous results using Bacillus stratosphericus (Hosseini-Abari et al., 2014) and Enterobacter sp. (Sinha and Khare, 2011) . By comparing the peaks obtained in alkaline and acidic media for AgNP synthesis, a blue sharper shift with smaller particle sizes was obtained in alkaline medium, whereas in acidic medium, AgNP peaks exhibited a red wider shift, were larger in size, and existed in a small amount. In addition, in alkaline pH lower absorbance was detected. This could be explained by the increase in nucleation process rate as a result of a large number of OH ions causing the synthesis of smallsized AgNPs in large amounts (Kalimuthu et al., 2008; Tripathy et al., 2010; Khalil et al., 2014) . These results may also be explained by using one of the most acceptable hypotheses for silver nanoparticle synthesis by bacteria, which attributed the bacterial ability to synthesize AgNPs to the presence of the nitrate reductase enzyme, which is sensitive to pH and temperature variations. A previous study proposed the secretion of NADH and NADPHdependent nitrate reductase enzyme that is responsible for reduction of nitrate to nitrite, shuttling an electron to the silver ion reducing Ag+ to Ag0 and synthesizing silver nanoparticles (Kumar et al., 2007) .
The change in the bacterial lysate supernatant volume correlated with the peak formation pattern. This could be attributed to the amount of reducing agents present in the bacterial lysate supernatant, which, in turn, increased the amount of AgNPs synthesized.
The AgNO 3 concentration affected the synthesis process of AgNPs, which is similar to previous reports conducted using Bacillus subtilis EWP-46 and Pseudomonas veronii AS41G, revealing that 1 mM AgNO was the optimum concentration for biosynthesis of AgNPs (Jeevan et al., 2012; Baker et al., 2015) . Studies by Tripathy et al. (2010) and Hosseini-Abari et al. (2014) showed that the optimum concentration of AgNO3 depended on the bacterial species. In the present work, the AgNPs synthesized by E. cloacae Ism26 were found to be stable for 6 months, while AgNPs synthesized by B. cereus were found to be less stable (Ganesh Babu and Gunasekaran, 2009) . This high stability is an advantageous feature in industrial and medical applications and supports the use of E. cloacae Ism26 in AgNP synthesis.
Very small-sized (7 to 25 nm) AgNPs were obtained by E. cloacae Ism26 with a characteristic spherical shape. This nanoparticle size range is similar to that synthesized by cyanobacteria and Klebsiella pneumoniae (Lengke et al., 2007; Mokhtari et al., 2009; Bansal et al., 2010; Ramanathan et al., 2011) . The FTIR spectra displayed different functional groups as the peak at 3464 cm–1 was assigned to NH stretching and 2360 cm–1 was associated with the amine C-N stretching. Other peaks at 1642, 1514, and 1386 in the region of 1000–1800 cm–1 corresponded to the amide I–III bands of polypeptide/proteins and symmetric stretching of COO- (Shahverdi et al., 2007; Kalishwaralal et al., 2010) . These spectra suggest that the E. cloacae Ism26 lysate supernatant protein plays a major role in the reduction of Ag+ to AgNPs. Kumar et al. (2007) and Kalimuthu et al. (2008) suggested that similar functional groups seized silver ions and served as stabilizing agents as well as nucleation sites for AgNP synthesis.
The results of XRD showed that the AgNP crystals were face-centered cubic (fcc) in structure. Similar results were obtained by AgNPs synthesized using Pseudomonas veronii AS 41G and Bacillus subtilis EWP-46 (Velmurugan et al., 2014; Baker et al., 2015) . Other insignificant peaks were also observed, suggesting crystallization of bioorganic materials on the surface of the AgNPs (Khalil et al., 2014) .
DLS was used to determine the size range of the synthesized AgNPs and the data indicated that particle size was in the nanometer range (5 to 50 nm). This was in agreement with previous work by Ramanathan et al. (2011) .
AgNPs synthesized by E. cloacae Ism26 exhibited antibacterial activities against both gram-positive and gram-negative bacteria. The diefrence in response may be related to the structural variation of the cell wall between gram-positive and gram-negative bacteria and the nature of the chemical groups that are challenged by AgNPs (Sondi and Salopek-Sondi, 2004; Jagtap and Bapat, 2013) . As our results indicate, AgNP concentration of 75 µg/mL inhibited the growth of all tested bacteria. A previous study used a higher concentration, up to 180 µg/mL, against E. coli, Staphylococcus aureus, and Salmonella typhimurium to achieve the same effect (Sondi and Salopek-Sondi, 2004) . Other biologically synthesized AgNPs failed to inhibit either Staphylococcus aureus or Salmonella typhimurium (Jagtap and Bapat, 2013) . The eficacy of AgNPs synthesized by E. cloacae Ism26 at a relatively low concentration may be related to their small average size and shape. In support of this assumption are the results of earlier studies, which indicated that the smaller the size of AgNPs, the lower the dose needed to cause inhibition (Sondi and SalopekSondi, 2004; Suchomel et al., 2015) . Also, the spherical nanoparticles were found to possess higher antibacterial activity than that of the rod-shaped or triangular nanoparticles (Pal et al., 2007) .
In conclusion, the bacterial lysate supernatant of E. cloacae Ism26 when mixed with AgNPs under light conditions was able to synthesize small-sized spherical AgNPs. These AgNPs inhibited the growth of both grampositive and gram-negative bacteria significantly at low concentrations and had a stable shelf life of 6 months. These features give the AgNPs synthesized and investigated in this study promising potential and a wide array of possible applications. Morsy FM (2015). Toward revealing the controversy of bacterial biosynthesis versus bactericidal properties of silver nanoparticles (AgNPs): bacteria and other microorganisms do not per se viably synthesize AgNPs. Arch Microbiol 197: 645-655.
References
- Bai HJ , Yang BS , Chai CJ , Yang GE , Jia WL , Yi ZB ( 2011. ). Green synthesis of silver nanoparticles using Rhodobacter sphaeroides . World J Microbiol Biotechnol 27 : 2723 - 2728 . [Google Scholar]
- Baker S , Mohan Kumar K , Santosh P , Rakshith D , Satish S ( 2015. ). Extracellular synthesis of silver nanoparticles by novel Pseudomonas veronii AS41G inhabiting Annona squamosa L. and their bactericidal activity . Spectrochim Acta A 136 : 1434 - 1440 . [DOI] [PubMed] [Google Scholar]
- Bansal V , Li V , O'Mullane A P , Bhargava SK ( 2010. ). Shape dependent electrocatalytic behaviour of silver nanoparticles . Cryst Eng Commun 12 : 4280 - 4286 . [Google Scholar]
- Das VL , oThmas R , Varghese RT , Soniya EV , Mathew J , Radhakrishnan EK ( 2013. ). Extracellular synthesis of silver nanoparticles by the Bacillus strain CS 11 isolated from industrialized area . 3 Biotech 4 : 121 - 126 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eby DM , Luckarift H R , Johnson GR ( 2009. ). Hybrid antimicrobial enzyme and silver nanoparticle coatings for medical instruments . ACS Appl Mater Inter 1 : 1553 - 1560 . [DOI] [PubMed] [Google Scholar]
- Edwards A , Civitello A , Hammond HA , Caskey CT ( 1991. ). DNA typing and genetic mapping with trimeric and tetrameric tandem repeats . Am J Hum Genet 49 : 746 - 756 . [PMC free article] [PubMed] [Google Scholar]
- Elof JN ( 1998. ) A sensitive and quick microplate method to determine the minimal inhibitory concentration of plant extracts for bacteria . Planta Med 64 : 711 - 713 . [DOI] [PubMed] [Google Scholar]
- Ganesh Babu MM , Gunasekaran P ( 2009. ). Production and structural characterization of crystalline silver nanoparticles from Bacillus cereus isolate . Colloid Surface B 74 : 191 - 195 . [DOI] [PubMed] [Google Scholar]
- Gurunathan S , Kalishwaralal K , Vaidyanathan R , Venkataraman D , Pandian SRK , Hariharana N , Muniyandi J , Eom SH ( 2009. ). Biosynthesis, purification and characterization of silver nanoparticles using Escherichia coli . Colloid Surface B 74 : 328 - 335 . [DOI] [PubMed] [Google Scholar]
- Hosseini-Abarz A , Emtiazi G , Lee SH , Kim BG , Kim JH ( 2014. ). Biosynthesis of silver nanoparticles by Bacillus stratosphericus spores and the role of dipicolinic acid in this process . Appl Biochem Biotechnol 174 : 270 - 282 . [DOI] [PubMed] [Google Scholar]
- Jagtap UB , Bapat VA ( 2013. ). Green synthesis of silver nanoparticles using Artocarpus heterophyllus Lam. seed extract and its antibacterial activity . Ind Crops Prod 46 : 132 - 137 . [Google Scholar]
- Jeevan P , Ramya K , Rena AE ( 2012. ). Extracellular biosynthesis of silver nanoparticles by culture supernatant of Pseudomonas aeruginosa . Indian J Biotechnol 11 : 72 - 76 . [Google Scholar]
- Kalimuthu K , Suresh Babu R , Venkataraman D , Bilal M , Gurunathan S ( 2008. ). Biosynthesis of silver nanocrystals by Bacillus licheniformis . Colloid Surface B 65 : 150 - 153 . [DOI] [PubMed] [Google Scholar]
- Kalishwaralal K , Deepak V , Pandian SRK , Kottaisamy M , BarathManiKanth S , Kartikeyan B , Gurunathan S ( 2010. ). Biosynthesis of silver and gold nanoparticles using Brevibacterium casei . Colloid Surface B 77 : 257 - 262 . [DOI] [PubMed] [Google Scholar]
- Khalil MMH , Ismail EH , El-Baghdady KZ , Mohamed D ( 2014. ). Green synthesis of silver nanoparticles using olive leaf extract and its antibacterial activity . Arab J Chem 7 : 1131 - 1139 . [Google Scholar]
- Klaus T , Joerger R , Olsson E , Granqvist CG ( 1999. ). Silver-based crystalline nanoparticles, microbially fabricated . P Natl Acad Sci USA 96 : 13611 - 13614 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar SA , Abyaneh MK , Gosavi SW , Kulkarni SK , Pasricha R , Ahmad A , Khan MI ( 2007. ). Nitrate reductase-mediated synthesis of silver nanoparticles from AgNO3 . Biotechnol Lett 29 : 439 - 445 . [DOI] [PubMed] [Google Scholar]
- Lara HH , Ayala-Núñez NV , Ixtepan Turrent LDC , Rodríguez Padilla C ( 2010. ). Bactericidal effect of silver nanoparticles against multidrug-resistant bacteria . World J Microbiol Biotechnol 26 : 615 - 621 . [Google Scholar]
- Law N , Ansari S , Livens FR , Renshaw JC , Lloyd JR ( 2008. ). Formation of nanoscale elemental silver particles via enzymatic reduction by Geobacter sulfurreducens . Appl Environ Microbiol 74 : 7090 - 7093 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lengke MF , Fleet ME , Southam G ( 2007. ). Biosynthesis of silver nanoparticles by filamentous cyanobacteria from a silver(I) nitrate complex . Langmuir 23 : 2694 - 2699 . [DOI] [PubMed] [Google Scholar]
- Minaeian S , Shahverdi AR , Nohi AS , Shahverdi HR ( 2008. ). Extracellular biosynthesis of silver nanoparticles by some bacteria . Jundishapur Journal of Natural Pharmaceutical Products 17 : 1 - 4 . [Google Scholar]
- Mokhtari N , Daneshpajouh S , Seyedbagheri S , Atashdehghan R , Abdi K , Sarkar S , Minaian S , Shahverdi HR , Shahverdi AR ( 2009. ). Biological synthesis of very small silver nanoparticles by culture supernatant of Klebsiella pneumonia: the effects of visible-light irradiation and the liquid mixing process . Mater Res Bull 44 : 1415 - 1421 . [Google Scholar]
- Nam KT , Lee YJ , Krauland EM , Kottmann ST , Angela M ( 2008. ). Peptide-mediated reduction of silver ions on engineered biological scaofld . ACS Nano 2 : 1 - 4 . [DOI] [PubMed] [Google Scholar]
- Narayanan KB , Sakthivel N ( 2010. ). Biological synthesis of metal nanoparticles by microbes . Adv Colloid Interface Sci 156 : 1 - 13 . [DOI] [PubMed] [Google Scholar]
- Natarajan K , Selvaraj S , Murty VR ( 2010. ). Microbial production of silver nanoparticles . Dig J Nanomater Biostructures 5 : 135 - 140 . [Google Scholar]
- Pal S , Tak YK , Song JM ( 2007. ). Does the antibacterial activity of silver nanoparticles depend on the shape of the nanoparticle? A study of the gram-negative bacterium Escherichia coli . Appl Environ Microbiol 73 : 1712 - 1720 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parikh RY , Singh S , Prasad BLV , Patole MS , Sastry M , Schouche YS ( 2008. ). Extracellular synthesis of crystalline silver nanoparticles and molecular evidence of silver resistance from Morganella sp .: towards understanding biochemical synthesis mechanism . ChemBioChem 9 : 1415 - 1422 . [DOI] [PubMed] [Google Scholar]
- Priyadarshini S , Gopinath V , Meera Priyadharsshini N , MubarakAli D , Velusamy P ( 2013. ). Synthesis of anisotropic silver nanoparticles using novel strain, Bacillus eflxus and its biomedical application . Colloid Surface B 102 : 232 - 237 . [DOI] [PubMed] [Google Scholar]
- Rai M , Kon K , Ingle A , Duran N , Galdiero S , Galdiero M ( 2014. ). Broad-spectrum bioactivities of silver nanoparticles: The emerging trends and future prospects . Appl Microbiol Biotechnol 98 : 1951 - 1961 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rai M , Yadav A , Gade A ( 2009. ). Silver nanoparticles as a new generation of antimicrobials . Biotechnol Adv 27 : 76 - 83 . [DOI] [PubMed] [Google Scholar]
- Ramanathan R , O'Mullane AP , Parikh RY , Smooker PM , Bhargava SK , Bansal V ( 2011. ). Bacterial kinetics-controlled shapedirected biosynthesis of silver nanoplates using Morganella Psychrotolerans . Langmuir 27 : 714 - 719 . [DOI] [PubMed] [Google Scholar]
- Seshadri S , Prakash A , Kowshik M ( 2012. ). Biosynthesis of silver nanoparticles by marine bacterium, Idiomarina sp . PR58-8. Bull Mater Sci 35 : 1201 - 1205 . [Google Scholar]
- Shahverdi AR , Minaeian S , Shahverdi HR , Jamalifar H , Nohi AA ( 2007. ). Rapid synthesis of silver nanoparticles using culture supernatants of Enterobacteria: a novel biological approach . Process Biochem 42 : 919 - 923 . [Google Scholar]
- Shivaji S , Madhu S , Singh S ( 2011. ). Extracellular synthesis of antibacterial silver nanoparticles using psychrophilic bacteria . Process Biochem 46 : 1800 - 1807 . [Google Scholar]
- Sinha A , Khare SK ( 2011. ). Mercury bioaccumulation and simultaneous nanoparticle synthesis by Enterobacter sp. cells . Bioresour Technol 102 : 4281 - 4284 . [DOI] [PubMed] [Google Scholar]
- Sondi I , Salopek-Sondi B ( 2004. ). Silver nanoparticles as antimicrobial agent: a case study on E. coli as a model for Gram-negative bacteria . J Colloid Interface Sci 275 : 177 - 182 . [DOI] [PubMed] [Google Scholar]
- Suchomel P , Kvitek L , Panacek A , Prucek R , Hrbac J , Vecerova R , Zboril R ( 2015. ). Comparative study of antimicrobial activity of AgBr and Ag nanoparticles (NPs) . PLoS One 10 : e0119202 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tripathy A , Raichur AM , Chandrasekaran N , Prathna TC , Mukherjee A ( 2010. ). Process variables in biomimetic synthesis of silver nanoparticles by aqueous extract of Azadirachta indica (Neem) leaves . J Nanoparticle Res 12 : 237 - 246 . [Google Scholar]
- Velmurugan P , Iydroose M , Mohideen MHAK , Mohan TS , Cho M , Oh BT ( 2014. ). Biosynthesis of silver nanoparticles using Bacillus subtilis EWP-46 cell-free extract and evaluation of its antibacterial activity . Bioprocess Biosyst Eng 37 : 1527 - 1534 . [DOI] [PubMed] [Google Scholar]
- Wei X , Luo M , Li W , Yang L , Liang X , Xu L ( 2012. ). Bioresource technology synthesis of silver nanoparticles by solar irradiation of cell-free Bacillus amyloliquefaciens extracts and AgNO3 . Bioresour Technol 103 : 273 - 278 . [DOI] [PubMed] [Google Scholar]
- Wiegand I , Hilpert K , Hancock REW ( 2008. ). Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances . Nat Protoc 3 : 163 - 175 . [DOI] [PubMed] [Google Scholar]