

Abstract
Aging is a main risk factor for development of cardiovascular diseases associated with the impairment of endothelial function in both sexes. In the present study, age-related changes in vascular responsiveness, epigenetic modifications of vessel wall, and blood biomarkers related to endothelial functions were examined in an age- and sex-dependent manner. Acetylcholine (ACh)-induced relaxations of the aorta were decreased in 3-, 6-, and 12-month-old rats compared to those in 1-month-old female rats. In males, maximum relaxations related to ACh were higher in 1- and 6-month-old rats than in 3- and 12-month-old rats. Plasma levels of nitric oxide (NO) and asymmetric dimethylarginine (ADMA) decreased with age in female rats, and total antioxidant capacity (TAC) and hydrogen sulfide (H 2S) levels displayed biphasic alterations. In male rats, plasma levels of NO, TAC, and ADMA decreased with age, and H2S levels increased. Aging also caused a sex-dependent alteration in epigenetic modification of vessels. Expressions of H3K27me2, H3K27me3, H3K36me2, and H3K36me3 were much higher in vessels of 12-month-old female rats compared to those in younger age groups. These results indicate that vascular functions, epigenetic modifications of vessels, and plasma levels of endothelium-related biomarkers are affected by age and sex. These findings could be important for the assessment of vascular status over the course of the life span.
Keywords: Aging, endothelial function, biomarker, histone methylation
1. Introduction
Aging is a natural process for every living organism. The trouble here is increased health problems as a result of aging. Aging is a dominant risk factor for cardiovascular diseases and is associated with progressive vascular dysfunction (Herrera et al., 2010; Laurent, 2012) . Many studies reported a sex difference in age-associated vascular changes (Sarabi et al., 1999; Sader and Celermajer, 2002; Okumura et al., 2011) . Celermajer et al. (1994) showed that age-related impairment in endothelial function appeared to occur earlier in males than in females. Diefrent regulation of endothelial function is one of the main mechanisms underlying the variation in age-associated vascular changes in females and males (Sarabi et al., 1999) . However, the underlying mechanisms of sex-dependent alterations in endothelial function remain to be investigated.
Endothelium-derived NO plays a key role in the regulation of vascular homeostasis (Moncada et al., 1991) . The capacity of NO release is considered to be a major indicator of endothelial function. Vascular aging characterized by endothelial dysfunction is associated with reduced NO bioavailability and increased generation of reactive oxygen species (ROS). ROS combine with NO and produce deleterious free radicals, leading to endothelial dysfunction (Heitzer et al., 2001; Tsimakis, 2006; El Assar et al., 2013; Rochette et al., 2013) . ADMA is an endogenous competitive inhibitor of NO synthase and an increase in ADMA is associated with impairment of NO synthesis (Bouras et al., 2013; Sverdlov et al., 2014) . It has been reported that plasma concentrations of ADMA increase in elderly people and in postmenopausal women (Schulze et al., 2005) . H2S is acknowledged as an important gaseous signaling molecule (Kolluru et al., 2013) . It has been revealed that H2S is an endogenous regulator of oxidative damage and aging in C. elegans (Qabazard et al., 2014) . However, the relationship between these molecules and endothelial function in the aging process is still largely unknown.
Recent studies have focused on epigenetic changes that occur as a hallmark of aging (Brunet and Berger, 2014). These epigenetic modifications that include changes in DNA methylation, histone modifications, and alterations in microRNA profiles seem to be a signature of aging. Histone methylation is an epigenetic modification known to be involved in the aging process (McCauley and Dang, 2014) . Insight into the epigenetic modifications involved in the aging process may provide new tools for the development of diagnostic, preventive, and therapeutic strategies to treat aging-related diseases. Further investigations are needed to elucidate the epigenetic modification of vessel walls in age-related endothelial dysfunction.
Several biomarkers have already been suggested for the monitoring of the progression of endothelial dysfunction. However, no biomarkers have yet been accepted for endothelial dysfunction in the aging process. If some biomarkers could be shown to be associated with the endothelial function in aging, therapeutic and preventive approaches might be beneficial in slowing down agerelated disease progress. In addition to diagnosis, biomarkers for endothelial dysfunction might also help to reveal the stage of pathology and the appropriate treatment strategy in a sex-dependent manner. However, to the best of our knowledge, there has been no study examining the relationship between age, sex, and blood biomarkers of endothelial dysfunction and epigenetic changes of vessels in the aging process. In the present study, age-related changes in function of vascular endothelium, epigenetic modifications of vessels, and levels of blood biomarkers related to endothelial functions (NO, ADMA, TAC, and H2S) were examined in female and male rats.
2. Materials and methods
2.1 Animal care
All animal experiments were approved by the Local Ethics Committee for Animal Care and Use of Ankara University. Female and male Wistar albino rats (1, 3, 6, and 12 months old) were obtained from the Laboratory Animal Service of Ankara University. The rats were synchronized to 12-h light/dark cycle at a stable temperature (24 ± 1 °C) and had free access to food and water.
2.2 Tissue isolation and assessment of functions
The animals were anesthetized with thiopental sodium (40 mg/kg, IP) and blood samples were collected. The thoracic aortas were rapidly removed and cleaned of fat and connective tissues. Then 3–4 mm ring segments of the thoracic aorta were mounted in the isolated tissue baths. The baths were filled with Krebs buffer solution (composition in mM: 112 NaCl, 5 KCl, 1.2 NaH2PO4, 11.5 dextrose, 0.5 MgCl2, 2.5 CaCl2, and 25 NaHCO ; pH 7.4) that was simmered continuously with a 95% O2 and 5% CO2 mixture and warmed (37 °C). The rings were stretched to 2 g of resting tension. The changes in isometric tension were recorded by an isometric forcedisplacement transducer (The Integrated Tissue Bath & Heater System [ITBS05], COMMAT, Turkey). The rings were equilibrated for 40 min at resting tension and then contraction with the addition of 90 mM potassium chloride (KCl) was recorded. The thoracic aortas were washed with fresh Krebs solution at 10-min intervals for 40 min and then a cumulative concentration-response curve to ACh (10–8–10–5 M) was obtained in the presence of 10–6 M phenylephrine submaximal contraction.
2.3 Biochemical examination
Plasma NO levels were measured spectrophotometrically using the Navarro-Gonzálvez method based on the Griess reaction, involving a shortened incubation period of nitrate with cadmium (Navarro-Gonzálvez et al., 1998) . This method was modified in our laboratories for 96-well plates.
The TAC of plasma was measured with the method described previously (Usanmaz and Demirel Yilmaz, 2008) , based on the reduction of Cu2+ to Cu1+ by the antioxidants of plasma. Neocuproine (Nc) was used as a chromogenic agent and the color of the formed colored complex (Nc-Cu1+) was detected spectrophotometrically at 455 nm.
H2S levels of human plasma were measured spectrophotometrically, according to the previously described method based on the measurements of the absorbance of the colored product (methylene blue), produced as a result of the chemical reaction between N,N-dimethyl-p-phenylenediamine and FeCl3 at 670 nm (Zhang et al., 2008) .
For the measurements of ADMA levels, ELISA kits from Immundiagnostik A.G. (Bensheim, Germany) were used according to the manufacturer’s instructions.
2.4 Histone protein isolation
Histone proteins were isolated from the frozen rat thoracic aorta using an EpiQuik Total Histone Extraction Kit (Epigentek, Farmingdale NY, USA) according to the manufacturer’s protocol. Brieyfl, after pooling, all samples were homogenized in 1X pre-lysis buffer. The resulting homogenates were centrifuged at 3000 rpm for 5 min at 4 °C. After removing the supernatant, the pellets were resuspended in lysis buffer and incubated on ice for 30 min. Further centrifugation was performed at 12,000 rpm for 5 min at 4 °C and the supernatant for each pooled group containing histone proteins was kept after mixing with balance-DTT buffer.
2.5 Western blotting (SDS-polyacrylamide gel electrophoresis (SDS-PAGE))
Total histone proteins were separated in 15% SDS-PAGE and transferred to a PVDF membrane. The membrane was blocked with bovine serum albumin blocking solution either for 2 h at room temperature or overnight at 4 °C on a shaker. Primary antibody (H3K27me2, H3K27me3, H3K36me2, H3K36me3, H4; Cell Signaling, Danvers, MA, USA) solution was prepared in blocking solution at 1:500 dilution and incubation was performed at room temperature for 2 h without shaking. H4 was used for equal loading control. After washing the membrane, antiRabbit-IgG-HRP (Cell Signaling) secondary antibody solution was prepared in blocking solution at 1:1000 dilution and applied for 1 h at room temperature. After removing the excess antibody by washing, SuperSignal West Femto Maximum Sensitivity Substrate (Thermo Scientific, Rockford, IL, USA) was applied on top of the membrane for 3 min and placed in an X-ray film cassette and developed. Protein band intensities were quantified using ImageJ.
2.6 Statistical analysis
All values were expressed as mean ± SEM. Tissue relaxation responses were represented as a percentage of phenylephrine submaximal contractions. Repeatedmeasures of two-way ANOVA (post-hoc Bonferroni) was used to test differences between the groups of relaxations. One-way ANOVA (post-hoc Newman–Keuls multiple comparison test) was used to test differences between the groups of contractions and biochemical parameters. Differences between the female and male groups were evaluated with Student’s t test. For all comparisons, differences were considered statistically significant at a value of P < 0.05.
2.7 Chemicals
All chemicals were obtained from Sigma Chemical Co (St Louis, MO, USA).
3. Results
Cumulative concentration-response curves were obtained for ACh (10–5–10–8 M) in the thoracic aorta of the 1-, 3-, 6-, and 12-month-old male and female rats (Figure 1). The ACh-induced relaxations of vessels in the 1-monthold group were different from those in the 3-, 6-, and 12-month-old groups of female rats (P < 0.05). In the male rats, the ACh-induced relaxations were significantly different in the 1- and 6-month-old rats compared to those in the 3- and 12-month-old rats. The ACh relaxations in the 6-month-old group were different from those in the 1-month-old rats (P < 0.05) (Figure 1). Maximum response of sodium nitroprusside-induced (10–5 M) endotheliumindependent relaxations was similar in the female and male groups (data not shown).
Figure 1.
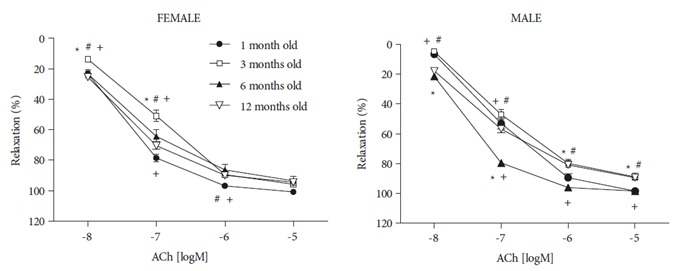
Endothelium-dependent relaxation of thoracic aorta isolated from 1-, 3-, 6-, and 12-month-old female and male rats. ACh-induced endothelium-dependent relaxations were different in the female and male rats (P < 0.05). Differences at 1 month old (*), 6 months old (#), and 12 months old (+). Vasorelaxant responses are expressed as a percentage of submaximal contraction to phenylephrine (10–6 M). Values are expressed as mean ± SEM (n = 7–12).
No alterations were observed in the contractions to KCl (90 mM) in the different age groups of the female rats. In the male rats, the KCl-stimulated contraction of the aorta increased in an age-dependent manner (P < 0.05) (Figure 2). Contractions in the oldest group (12 months old) were significantly higher than those in the 6-, 3-, and 1-month-old groups. The KCl contractions in the male 1- and 3-month-old groups were significantly lower than those in the female groups, and the male 12-month-old group contraction value was significantly higher than that in the female group (P < 0.05) (Figure 3).
Figure 2.
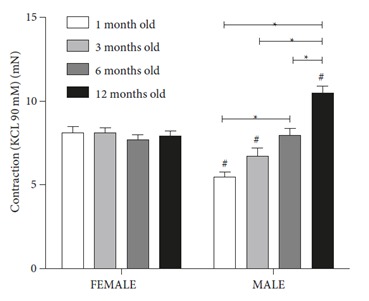
KCl (90 mM)-stimulated contraction of the thoracic aorta isolated from 1-, 3-, 6-, and 12-month-old female and male rats. Contractions to KCl were similar in all female groups. In male rats, contractions increased with age (P < 0.05). Contractions were different in the female and male groups at the age of 1, 3, and 12 months old (P < 0.05). Differences from the same age female group (#). Values are expressed as mean ± SEM (n = 7–12).
Figure 3.
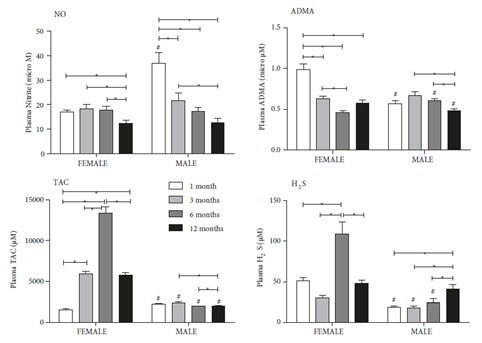
Changes in plasma NO, ADMA, TAC, and H2S levels of 1-, 3-, 6-, and 12-month-old female and male rats. Differences from the same age female group (#) (P < 0.05). Values are expressed as mean ± SEM (n = 7–12).
The plasma levels of NO were significantly decreased in the 12-month-old female rats compared to those in the younger age groups (P < 0.05) (Figure 3). In the male rats, the plasma levels of NO were significantly decreased in an age-dependent manner (P < 0.05) (Figure 3). A decrease in ADMA levels was observed parallel with age in the 1-, 3-, and 6-month-old female rats (P < 0.05). In the male rats, the lowest ADMA level was observed in the 12-month-old group (P < 0.05). TAC levels in the female rats displayed biphasic alterations, increasing in the early stage of aging and then decreasing at 12 months. The plasma TAC level of the 1-month-old female rats was significantly lower than that of the other age groups (P < 0.05) (Figure 3). The plasma TAC levels of the male rats were significantly lower in the 6- and 12-month-old groups than in the 3-monthold group (P < 0.05) (Figure 3). All the TAC levels of the male rats were around the level of the 1-month-old female rats. The H 2S levels of the female rats had a similar pattern to the TAC levels. The highest H 2S levels of the female rats were measured in the 6-month-old females (P < 0.05) (Figure 3). In the male rats, the highest H2S level was in the 12-month-old group. In the female rats, there were negative correlations between ADMA and TAC, and ADMA and H S, and a positive correlation between TAC and H2S (Table). In the male rats, a positive correlation was determined between NO and TAC and a negative correlation between NO and H2S (Table).
Table 1.
Correlation of blood biomarkers.
| NO | ADMA | TAC | H2S | ||
|---|---|---|---|---|---|
| FEMALE | NO | - | - | - | |
| ADMA | - | C = −0.676 P = 0.000000464 | C = −0.312 P = 0.0496 | ||
| TAC | - | - | C = 0.471 P = 0.00214 | ||
| NO | - | C = 0.293 P = 0.0436 | C = –0.392 P = 0.00701 | ||
| ADMA | - | - | - | ||
| TAC | - | - | - |
C = correlation coefficient
Total histone proteins in the aorta were isolated from both male and female rats in different age groups (1-, 3-, 6-, and 12-month-old). Tissue samples were pooled for each group to obtain sufficient protein. The quantitation of the band intensities was applied using H4 antibody as a loading control. Western blot results revealed that vessels isolated from the 12-month-old female rats had very high expressions of all the tested proteins, H3K27me2, H3K27me3, H3K36me2, and H3K36me3, compared to the other age groups of female and male animals. This increase was not observed in the aorta of the 12-monthold male rats. In both sexes, no difference was determined in the aorta of 1-, 3-, and 6-month-old animals (Figure 2)
4.Discussion
The results of the present study demonstrated that aging heterogeneously effects the functions and epigenetic modifications of vessels and blood biomarkers related to endothelial function (NO, TAC, H S, and ADMA) in a sex-dependent manner.
Aging is recognized as an independent risk factor for the development of cardiovascular diseases (Ferrari et al., 2003) . Structural and functional changes in vessel walls occur during the aging process. Endothelium plays an important role in modulating vascular tone and structure mainly through the production and release of several active substances. It is generally assumed that decreased synthesis of NO derived from the endothelial cell is correlated with endothelial dysfunction. Age-related endothelial dysfunction is induced by a number of factors including impaired NO bioavailability, oxidative stress, and inflammation (Toda, 2012) . In numerous studies, it has been demonstrated that endothelium-dependent relaxation of vessels is impaired in elderly humans and animals (Egashira et al., 1993; Kung and Luscher, 1995; Chauhan et al., 1996; Gerhard et al., 1996; Kim et al.,2011). Aging-related blunted endothelium-dependent relaxation of vessels was reported in Wistar (Hongo et al., 1988; Matz et al., 2000) , Sprague Dawley (Csiszar et al., 2002; Zhou et al., 2010) , and F1 (F344xBN) rats (van der Loo et al., 2000) . Similar to previous studies, ACh-induced endothelium-dependent relaxation of the thoracic aorta was observed to be affected by aging in the present study. However, age-related vascular dysfunction was also associated with sex (Sarabi et al., 1999; Sader and Celermajer, 2002; Okumura et al., 2011) . Celermajer et al. (1994) and Taddei et al. (1996) reported that agingrelated endothelial dysfunction in males preceded that in females.Aging-induced endothelial dysfunction is closely associated with decreased endothelial NO bioavailability, resulting in impaired vasodilatation (Luscher and Barton, 1997; Fleming and Busse, 1999; Taddei et al., 2001; Smith et al., 2006; Soucy et al., 2006; Donato et al., 2007; Hayashi et al., 2008; Trott et al., 2009) . Several mechanisms such as decreased endothelial NO synthase (eNOS) activity and an increase in ROS generation are involved in reduced NO bioavailability in aging (Chou et al., 1998; Herrera et al., 2010) . Previous studies demonstrated age-related decreased eNOS expression and vascular release of NO (Tschudi et al., 1996; Csiszar et al., 2002) . A different regulation of endothelial function might be one of the main mechanisms underlying the variation in age-associated vascular changes in different sexes (Sader and Celermajer, 2002) . In the current study, aging-dependent alterations of endothelium-dependent dilatations were significantly different in the female and male rats. However, sodium nitroprusside-induced endothelium-independent relaxation of the thoracic aorta was determined to be similar in the different sex and age groups of the rats. These findings suggest that aging-induced alterations in the endothelial function do not show the same pattern in females and males. It may be argued that NO production capacity of vascular endothelium could be altered by aging in a sex-dependent manner. Lifelong monitoring of endothelial function is important in respect of predicting cardiovascular diseases and for the development of age- and sexdependent therapeutic strategies. Aging also causes changes in the structure and function of vascular smooth muscle (Yildiz, 2007) . It has been shown that aging and sex have an eefct on the contraction of vessels induced by calcium entry, stretching, and electrical ifeld stimulation (Sullivan and Davison, 2001; Blough et al., 2007; Yang et al., 2015) . KCl-mediated coronary vascular resistance was observed to be higher in adult (12–18 months old) male rats than in young (3–4 months old) and immature (1–2 months old) rats (Hinschen et al., 2001) . Sullivan and Davison (2001) reported no significant differences in KCl-mediated contraction between young (3 months old) and old (26 months old) female rat groups. The calcium balance in vasoconstriction responses is known to alter with age (Yang et al., 2015) . KCl-induced contraction of vascular smooth muscle is dependent on the activation of voltage-dependent Ca2+ channels, which leads to increases in cytosolic free Ca2+ concentration (Ratz et al., 2005) . Fukuda et al. (2014) reported reduced expression of voltage-dependent Ca2+ channels in the aorta in 40-week-old male Wistar-Kyoto rats when compared to 8-week-old rats. On the other hand, similar to the current results, it was reported that high KCl-induced contraction and Ca2+ influx was lower in 12-week-old female rats than in male rats (Crews and Khalil, 1999) . All these results of the previous studies suggest that age and sex can induce changes in the vascular wall, which may result in different passive/active contractile forces in blood vessels. In this regard, the ability of aortic rings to undergo depolarization-induced contraction with KCl was evaluated, without any agonist stimulation. The results of the current study were consistent with those of previous studies. In the present study, an age-dependent increase in KCl-stimulated contraction was observed in the male rats but not in the female rats. These results demonstrated that agonist-independent contraction of blood vessels may alter depending on age and sex. Age- and sex-related alterations of KCl-induced contraction of the vessel may be due to the differentiation of Ca 2+ influx into smooth muscle cells.
Aging-dependent endothelial dysfunction has been associated with decreased endothelial NO bioavailability, which is a result of reduced endothelial NO synthase (eNOS) activity and increased ROS generation (Chou et al., 1998; Herrera et al., 2010) . Age-related decreased eNOS expression and endothelial NO release (Tschudi et al., 1996; Csiszar et al., 2002) and a significant decrease in serum level and urinary excretion of the NO metabolites nitrate and nitrite (Reckelhof et al., 1994) have been reported in previous studies. Consistent with those results, the lowest plasma NO levels were in the 12-month-old male and female rats in the present study. The current study firstly determined the lifelong and sex-dependent alterations of plasma NO levels in the rats. In addition, these results are the first to demonstrate a correlation of age-related decrease in plasma NO level with reduced endothelium-dependent relaxations of vessel in both sexes.
ADMA is an endogenous inhibitor of NOS enzyme and represents an established marker of cardiovascular risk (Sibal et al., 2010) . It has been reported that plasma concentrations of ADMA increase in both male and female elderly individuals (Kielstein et al., 2003; Marliss et al., 2006) . In another study, it was observed that serum ADMA levels in 20-month-old male Sprague Dawley rats were higher than those in the 6-month-old group (Xiong et al., 2001) . Lifelong variations in plasma ADMA levels between female and male rats have been observed for the ifrst time in the current study. The results of the present study showed that plasma ADMA levels represented different patterns with age that were a down–up alteration in females and an up–down alteration in male rats. Based on these findings, it can be considered that ADMA levels represent diversity in sex, which may be involved in different age-related vascular modifications in female and male rats.
Increased oxidative stress is an important mechanism underlying the normal aging process (El Assar et al., 2013) . Numerous studies have reported oxidative stress induced by elevated ROS and impaired NO bioavailability with aging (Hamilton et al., 2001; Taddei et al., 2001; Eskurza et al., 2004; Marmol et al., 2007; Ungvari et al., 2011) . It has been detected that aortic oxidative stress is increased in old male but not in old female mice (Takenouchi et al., 2009) . While some studies demonstrated that the total antioxidant capacity of plasma decreased ( Sivonova et al., 2007 ), others found it to be unchanged in rats (Nakamura and Omaye, 2004) . In the current study, plasma TAC levels displayed biphasic alterations, increasing in the early stage of aging in female rats but decreasing in adult and middleaged male rats. TAC is used as a marker of oxidative stress and these results demonstrated that antioxidant status changes throughout life and shows a difference between females and males.
H S is a newly reported gasotransmitter, a potent inhibitor of superoxide formation in endothelial cells (Muzafar et al., 2008) , and a strong scavenger of oxygenderived free radicals (Geng et al., 2004) . It has been thought that the beneficial eefcts of H 2S may be mediated through its antioxidant eefcts (Suo et al., 2013) . Studies on C. elegans have shown that H2S is an endogenous regulator of oxidative damage, metabolism, and aging (Qabazard et al., 2014) and exogenous H2S prolongs the lifespan (Miller and Roth, 2007) . In a study of healthy subjects of different ages, no difference was found in serum H 2S level (Chen et al., 2005) . However, it is still unclear what role H S plays in the aging process. In the present study, the plasma level of H2S in the female rats displayed biphasic alterations, increasing in the early stage of aging and then decreasing. In the male rats, H2S levels increased in an age-dependent manner. These results are the first to have been obtained demonstrating that blood H2S levels may be involved in the aging process and show a sex difference in rats.
A correlation between biomarkers related to endothelial function was previously reported. It has been suggested that H2S and NO are required in endothelium-dependent vasorelaxation in isolated aortic ring (Coletta et al., 2012) . In human endothelial cells and smooth muscle cells from rat aorta, H2S limited the formation of NO (Kloesch et al., 2016) . Accordingly, in the present study a negative correlation between the NO and H2S groups was observed in the male group.
In the current study, although a positive correlation between TAC and H S was observed, there were negative correlations between the ADMA and TAC groups and between the ADMA and H S groups in female rats. On the other hand, a positive correlation between the NO and TAC groups was calculated in the male group. It was observed that oxidative stress marker TAC was higher when the levels of H S and NO were increased. These results suggested that interaction of the plasma biomarker may vary in a sex-dependent manner in rats.
Epigenetics is the term used for chromatin-based pathways that can change gene expression without an alteration in the underlying base sequence of the DNA and includes three mechanisms: DNA methylation, histone posttranslational modifications, and RNA-based mechanisms (Matouk and Marsden, 2008) . Several recent studies have reported that epigenetic changes play an important role in some cardiovascular pathologies (Han et al., 2015) and regulation of the aging process (Brunet and Berger, 2014). Histone methylation is one of the epigenetic modifications that occurs as a hallmark of aging (McCauley and Dang, 2014; Kawamura et al., 2015) . Age-related changes in histone methylation were reported in Drosophila (Wood et al., 2010) , mouse brain (Wang et al., 2010) , and rat kidney and liver (Sarg et al., 2002) . In the current study, expressions of H3K27me2, H3K27me3, H3K36me2, and H3K36me3 were observed to be higher in the thoracic aorta isolated from the 12-month-old female rats compared to those in the younger age groups while no difference was observed in those in the male rats. H3K36 methylation is associated with transcriptional activation while H3K27 methylation is correlated with gene repression (Matouk and Marsden, 2008) . These findings suggest that histone methylation may represent sex diversity. The different histone methylation may lead to alteration in gene activation and repression. These changes in the histone methylation state may be involved in the difference of age-associated regulation of the cardiovascular system in male and female rats.
Significant sex differences in aging as a cardiovascular risk factor have been defined worldwide. Premenopausal females are known to be at a lower risk of cardiovascular morbidity and mortality than males. Menopause and consequent estrogen deficiency is associated with endothelial dysfunction and as a result, the cardiovascular risk in females is increased (Virdis and Taddei, 2010) . It has been suggested that blood pressure and sympathetic activity are lower in younger females than those in younger males (Joyner et al., 2015) . In addition, alterations in blood pressure and vascular resistance regulation between males and females were also reported (Barnes et al., 2014) . The present study demonstrated a sex-dependent variation in rat vascular properties that influenced vascular tonus.
Age- and sex-dependent changes in vascular function and the underlying mechanisms need to be analyzed more extensively before any safe conclusions can be reached. The results of the current study are the first to indicate a lifelong alteration of vascular responsiveness and plasma biomarkers related to endothelial function and epigenetic modification of vessels in both sexes of rat. The present study emphasizes that aging-related variations in the functions of vessels and biomarkers were different in the female and male rats. These findings may have important implications for the assessment of diagnostic, preventive, and therapeutic approaches to aging and age-related diseases in different sexes.
Acknowledgment
This study was supported by a research grant, 12B3330001, from the Ankara University Research Foundation.
References
- Barnes JN , Hart EC , Curry TB , Nicholson WT , Eisenach JH , Wallin BG , Charkoudian N , Joyner MJ ( 2014. ). Aging enhances autonomic support of blood pressure in women . Hypertension 63 : 303 - 308 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blough ER , Rice KM , Desai DH , Wehner P , Wright GL ( 2007. ). Aging alters mechanical and contractile properties of the Fisher 344/ Nnia X Norway/Binia rat aorta . Biogerontology 8 : 303 - 313 . [DOI] [PubMed] [Google Scholar]
- Bouras G Deeftreos S Tousoulis D Giannopoulos G Chatzis G Tsounis D Cleman MW Stefanadis C Asymmetric Dimethylarginine (ADMA): a promising biomarker for cardiovascular disease? Curr Top Med Chem. 2013;13:180–200. doi: 10.2174/1568026611313020007. [DOI] [PubMed] [Google Scholar]
- Brunet A Berger SL Epigenetics of aging and aging-related disease. Journals of Gerontology Series A: Biological and Medical Sciences. 2014;1:17–20. doi: 10.1093/gerona/glu042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Celermajer DS , Sorensen KE , Spiegelhalter DJ , Georgakopoulos D , Robinson J , Deanfield JE ( 1994. ). Aging is associated with endothelial dysfunction in healthy men years before the agerelated decline in women . J Am Coll Cardiol 24 : 471 - 476 . [DOI] [PubMed] [Google Scholar]
- Chauhan A , More RS , Mullins PA , Taylor G , Petch MC , Schofield PM ( 1996. ). Aging-associated endothelial dysfunction in humans is reversed by L-arginine . J Am Coll Cardiol 28 : 1796 - 1804 . [DOI] [PubMed] [Google Scholar]
- Chen YH , Yao WZ , Geng B , Ding YL , Lu M , Zhao MW , Tang CS ( 2005. ). Endogenous hydrogen sulfide in patients with COPD . Chest 128 : 3205 - 3211 . [DOI] [PubMed] [Google Scholar]
- Chou TC , Yen MH , Li CY , Ding YA ( 1998. ). Alterations of nitric oxide synthase expression with aging and hypertension in rats . Hypertension 31 : 643 - 648 . [DOI] [PubMed] [Google Scholar]
- Coletta C , Papapetropoulos A , Erdelyi K , Olah G , Módis K , Panopoulos P , Asimakopoulou A , Gerö D , Sharina I , Martin E et al. ( 2012. ). Hydrogen sulfide and nitric oxide are mutually dependent in the regulation of angiogenesis and endotheliumdependent vasorelaxation . P Natl Acad Sci USA 109 : 9161 - 9166 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crews JK , Khalil RA ( 1999. ). Gender-specific inhibition of Ca 2+ entry mechanisms of arterial vasoconstriction by sex hormones . Clin Exp Pharmacol P 26 : 707 - 715 . [DOI] [PubMed] [Google Scholar]
- Csiszar A , Ungvari Z , Edwards JG , Kaminski P , Wolin MS , Koller A , Kaley G ( 2002. ). Aging induced phenotypic changes and oxidative stress impair coronary arteriolar function . Circ Res 90 : 1159 - 1166 . [DOI] [PubMed] [Google Scholar]
- Donato AJ , Eskurza I , Silver AE , Levy AS , Pierce GL , Gates PE , Seals DR ( 2007. ). Direct evidence of endothelial oxidative stress with aging in humans: relation to impaired endothelium-dependent dilation and upregulation of nuclear factorkappaB . Circ Res 100 : 1659 - 1666 . [DOI] [PubMed] [Google Scholar]
- Egashira K , Inou T , Hirooka Y , Kai H , Sugimachi M , Suzuki S , Kuga T , Urabe Y , Takeshita A ( 1993. ). Eefcts of age on endotheliumdependent vasodilation of resistance coronary artery by acetylcholine in humans . Circulation 88 : 77 - 81 . [DOI] [PubMed] [Google Scholar]
- El Assar M , Angulo J , Rodríguez-Mañas L ( 2013. ). Oxidative stress and vascular inflammation in aging . Free Radical Biol Med 65 : 380 - 401 . [DOI] [PubMed] [Google Scholar]
- Eskurza I , Monahan KD , Robinson JA , Seals DR ( 2004. ). Eefct of acute and chronic ascorbic acid on flow-mediated dilatation with sedentary and physically active human ageing . The Journal of Physiology 556 : 315 - 324 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrari AU , Radaelli A , Centola M ( 2003. ). Invited review: aging and the cardiovascular system . J Appl Physiol 95 : 2591 - 2597 . [DOI] [PubMed] [Google Scholar]
- Fleming I , Busse R ( 1999. ). NO: the primary EDRF . J Mol Cell Cardiol 31 : 5 - 14 . [DOI] [PubMed] [Google Scholar]
- Fukuda T , Kuroda T , Kono M , Miyamoto T , Tanaka M , Matsui T ( 2014. ). Attenuation of L-type Ca2+ channel expression and vasomotor response in the aorta with age in both Wistar-Kyoto and spontaneously hypertensive rats . PLoS One 9 : e88975 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geng B , Chang L , Pan C , Qi Y , Zhao J , Pang Y , Du J , Tang C ( 2004. ). Endogenous hydrogen sulfide regulation of myocardial injury induced by isoproterenol . Biochem Bioph Res Co 318 : 756 - 763 . [DOI] [PubMed] [Google Scholar]
- Gerhard M , Roddy MA , Creager SJ , Creager MA ( 1996. ). Aging progressively impairs endothelium-dependent vasodilation in forearm resistance vessels of humans . Hypertension 27 : 849 - 853 . [DOI] [PubMed] [Google Scholar]
- Hamilton CA , Brosnan MJ , McIntyre M , Graham D , Dominiczak AF ( 2001. ). Superoxide excess in hypertension and aging: a common cause of endothelial dysfunction . Hypertension 37 : 529 - 534 . [DOI] [PubMed] [Google Scholar]
- Han S , Uludag MO , Usanmaz SE , Ayaloglu-Butun F , Akcali KC , Demirel-Yilmaz E ( 2015. ). Resveratrol effects histone 3 lysine 27 methylation of vessels and blood biomarkers in DOCA saltinduced hypertension . Mol Biol Rep 42 : 35 - 42 . [DOI] [PubMed] [Google Scholar]
- Hayashi T , Yano K , Matsui-Hirai H , Yokoo H , Hattori Y , Iguchi A ( 2008. ). Nitric oxide and endothelial cellular senescence . Pharmacol Therapeut 120 : 333 - 339 . [DOI] [PubMed] [Google Scholar]
- Heitzer MT , Schlinzig T , Krohn K , Meinertz T , Münzel T ( 2001. ). Endothelial dysfunction, oxidative stress, and risk of cardiovascular events in patients with coronary artery disease . Circulation 104 : 2673 - 2678 . [DOI] [PubMed] [Google Scholar]
- Herrera MD , Mingorance C , Rodríguez-Rodríguez R , Alvarez de Sotomayor M ( 2010. ). Endothelial dysfunction and aging: an update . Ageing Res Rev 9 : 142 - 152 . [DOI] [PubMed] [Google Scholar]
- Hinschen AK , Rose'Meyer RB , Headrick JP ( 2001. ). Age-related changes in adenosine-mediated relaxation of coronary and aortic smooth muscle . Am J Physiol-Heart C 280 : 2380 - 2389 . [DOI] [PubMed] [Google Scholar]
- Hongo K , Nakagomi T , Kassell NF , Sasaki T , Lehman M , Vollmer DG , Tsukahara T , Ogawa H , Torner J ( 1988. ). Eefcts of aging and hypertension on endothelium-dependent vascular relaxation in rat carotid artery . Stroke 19 : 892 - 897 . [DOI] [PubMed] [Google Scholar]
- Joyner MJ , Barnes JN , Hart EC , Wallin BG , Charkoudian N ( 2015. ). Neural control of the circulation: how sex and age differences interact in humans . Compr Physiol 5 : 193 - 215 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawamura K , Kinoshita M , Sekida S , Sunanaga T ( 2015. ). Histone methylation codes involved in stemness, multipotency, and senescence in budding tunicates . Mech Ageing Dev 145 : 1 - 12 . [DOI] [PubMed] [Google Scholar]
- Kielstein JT , Bode-Boger SM , Frolich JC , Ritz E , Haller H , Fliser D ( 2003. ). Asymmetric dimethylarginine, blood pressure, and renal perfusion in elderly subjects . Circulation 107 : 1891 - 1895 . [DOI] [PubMed] [Google Scholar]
- Kim SY , Park JT , Park JK , Lee JS , Choi JC ( 2011. ). Aging impairs vasodilatory responses in rats . Korean Journal of Anesthesiology 61 : 506 - 510 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kloesch B , Steiner G , Mayer B , Schmidt K ( 2016. ). Hydrogen sulfide inhibits endothelial nitric oxide formation and receptor ligandmediated Ca2+ release in endothelial and smooth muscle cells . Pharmacol Rep 68 : 37 - 43 . [DOI] [PubMed] [Google Scholar]
- Kolluru GK , Shen X , Bir SC , Kevil CG ( 2013. ). Hydrogen sulfide chemical biology: pathophysiological roles and detection . Nitric Oxide 35 : 5 - 20 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kung CF , Luscher TF ( 1995. ). Diefrent mechanisms of endothelial dysfunction with aging and hypertension in rat aorta . Hypertension 25 : 194 - 200 . [DOI] [PubMed] [Google Scholar]
- Laurent S ( 2012. ). Defining vascular aging and cardiovascular risk . J Hypertens 30 Suppl : 3 - 8 . [DOI] [PubMed]
- Luscher TF , Barton M ( 1997. ). Biology of the endothelium . Clin Cardiol 20 : 3 - 10 . [PubMed] [Google Scholar]
- Marliss EB , Chevalier S , Gougeon R , Morais JA , Lamarche M , Adegoke OA , Wu G ( 2006. ). Elevations of plasma methylarginines in obesity and ageing are related to insulin sensitivity and rates of protein turnover . Diabetologia 49 : 351 - 359 . [DOI] [PubMed] [Google Scholar]
- Marmol F , Sanchez J , Lopez D , Martinez N , Rosello-Catafau J , Mitjavila MT , Puig-Parellada P ( 2007. ). Loss of adaptation to oxidative stress as a mechanism for aortic damage in aging rats . J Physiol Biochem 63 : 239 - 247 . [DOI] [PubMed] [Google Scholar]
- Matouk CC , Marsden PA ( 2008. ). Epigenetic regulation of vascular endothelial gene expression . Circ Res 102 : 873 - 887 . [DOI] [PubMed] [Google Scholar]
- Matz RL , de Sotomayor MA , Schott C , Stoclet JC , Andriantsitohaina R ( 2000. ). Vascular bed heterogeneity in age-related endothelial dysfunction with respect to NO and eicosanoids . Brit J Pharmacol 131 : 303 - 311 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCauley BS , Dang W ( 2014. ). Histone methylation and aging: lessons learned from model systems . Biochim Biophys Acta 1839. : 1454 - 1462 . [DOI] [PMC free article] [PubMed]
- Miller DL , Roth MB ( 2007. ). Hydrogen sulfide increases thermotolerance and lifespan in Caenorhabditis elegans . P Natl Acad Sci USA 104 : 20618 - 20622 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moncada S , Palmer RMJ , Higgs EA ( 1991. ). Nitric oxide: physiology pathophysiology and pharmacology . Pharmacol Rev 43 : 109 - 142 . [PubMed] [Google Scholar]
- Muzafar S , Jeremy JY , Sparatore A , Del Soldato P , Angelini GD , Shukla N ( 2008. ). H2S-donating sildenafil (ACS6) inhibits superoxide formation and gp91phox expression in arterial endothelial cells: role of protein kinases A and G . Brit J Pharmacol 155 : 984 - 994 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura YK , Omaye ST ( 2004. ). Age-related changes of serum lipoprotein oxidation in rats . Life Sci 23 : 1265 - 1275 . [DOI] [PubMed] [Google Scholar]
- Navarro-Gonzálvez J , García-Benayas C , Arenas J ( 1998. ). Semiautomated measurement of nitrate in biological uflids . Clin Chem 44 : 679 - 681 . [PubMed] [Google Scholar]
- Okumura M , Iwai M , Nakaoka H , Sone H , Kanno H , Senba I , Ito M , Horiuchi M ( 2011. ). Possible involvement of AT2 receptor dysfunction in age-related gender difference in vascular remodeling . J Am Soc Hypertens 5 : 76 - 84 . [DOI] [PubMed] [Google Scholar]
- Qabazard B , Li L , Gruber J , Peh MT , Ng LF , Kumar SD , Rose P , Tan CH , Dymock BW , Wei F et al. ( 2014. ). Hydrogen sulfide is an endogenous regulator of aging in Caenorhabditis elegans . Antioxid Redox Sign 20 : 2621 - 2630 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ratz PH , Berg KM , Urban NH , Miner AS ( 2005. ). Regulation of smooth muscle calcium sensitivity: KCl as a calciumsensitizing stimulus . Am J Physiol-Cell Ph 288 : 769 - 783 . [DOI] [PubMed] [Google Scholar]
- Reckelhof JF , Kellum JA , Blanchard EJ , Bacon EE , Wesley AJ , Kruckeberg WC ( 1994. ). Changes in nitric oxide precursor, L-arginine, and metabolites, nitrate and nitrite, with aging . Life Sci 55 : 1895 - 1902 . [DOI] [PubMed] [Google Scholar]
- Rochette L , Lorin J , Zeller M , Guilland JC , Lorgis L , Cottin Y , Vergely C ( 2013. ). Nitric oxide synthase inhibition and oxidative stress in cardiovascular diseases: Possible therapeutic targets? Pharmacol Ther 140 : 239 - 257 . [DOI] [PubMed] [Google Scholar]
- Sader MA , Celermajer DS ( 2002. ). Endothelial function, vascular reactivity and gender differences in the cardiovascular system . Cardiovasc Res 53 : 597 - 604 . [DOI] [PubMed] [Google Scholar]
- Sarabi M , Millgård J , Lind L ( 1999. ). Eefcts of age, gender and metabolic factors on endothelium-dependent vasodilation: a population-based study . J Intern Med 246 : 265 - 274 . [DOI] [PubMed] [Google Scholar]
- Sarg B , Koutzamani E , Helliger W , Rundquist I , Lindner HH ( 2002. ). Postsynthetic trimethylation of histone H4 at lysine 20 in mammalian tissues is associated with aging . J Biol Chem 277 : 39195 - 39201 . [DOI] [PubMed] [Google Scholar]
- Schulze F , Maas R , Freese R , Schwedhelm E , Silberhorn E , Böger RH ( 2005. ). Determination of a reference value for NG, NGdimethyl-L-arginine in 500 subjects . Eur J Clin Invest 35 : 622 - 626 . [DOI] [PubMed] [Google Scholar]
- Sibal L , Agarwal SC , Home PD , Boger RH ( 2010. ). The role of asymmetric dimethylarginine (ADMA) in endothelial dysfunction and cardiovascular disease . Curr Cardiol Rev 6 : 82 - 90 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sivonová M , Tatarková Z , Duracková Z , Dobrota D , Lehotský J , Matáková T , Kaplán P ( 2007. ). Relationship between antioxidant potential and oxidative damage to lipids, proteins and DNA in aged rats . Physiol Res 56 : 757 - 764 . [DOI] [PubMed] [Google Scholar]
- Smith AR , Visioli F , Hagen TM ( 2006. ). Plasma membraneassociated endothelial nitric oxide synthase and activity in aging rat aortic vascular endothelia markedly decline with age . Arch Biochem Biophys 454 : 100 - 105 . [DOI] [PubMed] [Google Scholar]
- Soucy KG , Ryoo S , Benjo A , Lim HK , Gupta G , Sohi JS , Elser J , Aon MA , Nyhan D , Shoukas AA et al. ( 2006. ). Impaired shear stress-induced nitric oxide production through decreased NOS phosphorylation contributes to age-related vascular stifness . J Appl Physiol 101 : 1751 - 1759 . [DOI] [PubMed] [Google Scholar]
- Sullivan JC , Davison CA ( 2001. ). Gender differences in the eefct of age on electrical field stimulation (EFS)-induced adrenergic vasoconstriction in rat mesenteric resistance arteries . J Pharmacol Exp Ther 296 : 782 - 788 . [PubMed] [Google Scholar]
- Suo R , Zhao ZZ , Tang ZH , Ren Z , Liu X , Liu LS , Wang Z , Tang CK , Wei DH , Jiang ZS ( 2013. ). Hydrogen sulfide prevents H₂O₂- induced senescence in human umbilical vein endothelial cells through SIRT1 activation . Mol Med Rep 7 : 1865 - 1870 . [DOI] [PubMed] [Google Scholar]
- Sverdlov AL , Ngo DT , Chan WP , Chirkov YY , Horowitz JD ( 2014. ). Aging of the nitric oxide system: are we as old as our NO? J Am Heart Assoc 3 : e000973 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taddei S , Virdis A , Ghiadoni L , Mattei P , Sudano I , Bernini G , Pinto S , Salvetti A ( 1996. ). Menopause is associated with endothelial dysfunction in women . Hypertension 28 : 576 - 582 . [DOI] [PubMed] [Google Scholar]
- Taddei S , Virdis A , Ghiadoni L , Salvetti G , Bernini G , Magagna A , Salvetti A ( 2001. ). Age-related reduction of NO availability and oxidative stress in humans . Hypertension 38 : 274 - 279 . [DOI] [PubMed] [Google Scholar]
- Takenouchi Y , Kobayashi T , Matsumoto T , Kamata K ( 2009. ). Gender differences in age-related endothelial function in the murine aorta . Atherosclerosis 206 : 397 - 404 . [DOI] [PubMed] [Google Scholar]
- Toda N ( 2012. ). Age-related changes in endothelial function and blood flow regulation . Pharmacol Ther 133 : 159 - 176 . [DOI] [PubMed] [Google Scholar]
- Trott DW , Gunduz F , Laughlin MH , Woodman CR ( 2009. ). Exercise training reverses age-related decrements in endotheliumdependent dilation in skeletal muscle feed arteries . J Appl Physiol 106 : 1925 - 1934 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tschudi MR , Barton M , Bersinger NA , Moreau P , Cosentino F , Noll G , Malinski T , Luscher TF ( 1996. ). Eefct of age on kinetics of nitric oxide release in rat aorta and pulmonary artery . J Clin Invest 98 : 899 - 905 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsimakis S ( 2006. ). Oxidative biomarkers in the diagnosis and prognosis of cardiovascular disease . Am J Cardiol 98 : 9 - 17 . [DOI] [PubMed] [Google Scholar]
- Ungvari Z , Bailey-Downs L , Sosnowska D , Gautam T , Koncz P , Losonczy G , Ballabh P , deCabo R., Sonntag WE , Csiszar A ( 2011. ). Vascular oxidative stress in aging: a homeostatic failure due to dysregulation of NRF2-mediated antioxidant response . Am J Physiol-Heart C 301 : 363 - 372 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Usanmaz SE , Demirel Yilmaz E ( 2008. ). A microplate based spectrophotometric method for the determination of the total antioxidant capacity of human plasma: modified cupric reducing ability assay . Fund Clin Pharmacol 22 ( Suppl 2 ): 67 . [Google Scholar]
- van der Loo B , Labugger R , Skepper JN , Bachschmid M , Kilo J , Powell JM , Palacios-Callender M , Erusalimsky JD , Quaschning T , Malinski T et al. ( 2000. ). Enhanced peroxynitrite formation is associated with vascular aging . J Exp Med 192 : 1731 - 1744 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Virdis A , Taddei S ( 2010. ). Endothelial aging and gender . Maturitas 71 : 326 - 330 . [DOI] [PubMed] [Google Scholar]
- Wang CM , Tsai SN , Yew TW , Kwan YW , Ngai SM ( 2010. ). Identification of histone methylation multiplicities patterns in the brain of senescence-accelerated prone mouse 8 . Biogerontology 11 : 87 - 102 . [DOI] [PubMed] [Google Scholar]
- Wood JG , Hillenmeyer S , Lawrence C , Chang C , Hosier S , Lightfoot W , Mukherjee E , Jiang N , Schorl C , Brodsky AS et al. ( 2010. ). Chromatin remodeling in the aging genome of Drosophila . Aging Cell 9 : 971 - 978 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiong Y , Yuan LW , Deng HW , Li YJ , Chen BM ( 2001. ). Elevated serum endogenous inhibitor of nitric oxide synthase and endothelial dysfunction in aged rats . Clin Exp Pharmacol P 28 : 842 - 847 . [DOI] [PubMed] [Google Scholar]
- Yang Y , Zhu J , Wang X , Xue N , Du J , Meng X , Shen B ( 2015. ). Contrasting patterns of agonist-induced store-operated Ca2+ entry and vasoconstriction in mesenteric arteries and aorta with aging . J Cardiovasc Pharm 65 : 571 - 578 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yildiz O ( 2007. ). Vascular smooth muscle and endothelial functions in aging . Ann NY Acad Sci 1100 : 353 - 360 . [DOI] [PubMed] [Google Scholar]
- Zhang H , Moochhala SM , Bhatia M ( 2008. ). Endogenous hydrogen sulfide regulates inflammatory response by activating the ERK pathway in polymicrobial sepsis . J Immunol 181 : 4320 - 4331 . [DOI] [PubMed] [Google Scholar]
- Zhou E , Qing D , Li J ( 2010. ). Age-associated endothelial dysfunction in rat mesenteric arteries: roles of calcium-activated K+ channels (Kca) . Physiol Res 59 : 499 - 508 . [DOI] [PubMed] [Google Scholar]