

Abstract
Purpose:
Metabolic aberrations have been described in neoplasms with mutations in the Krebs cycle genes encoding succinate dehydrogenase (SDH), fumarate hydratase (FH) and isocitrate dehydrogenase (IDH). In turn, accumulation of oncometabolites succinate, fumarate, and 2-hydroxyglutarate can be employed to identify tumors with those mutations. Additionally, such metabolic readouts may aid in genetic variant interpretation and improve diagnostics.
Methods:
Using liquid-chromatography-mass-spectrometry, 395 pheochromocytomas and paragangliomas (PPGLs) from 391 patients were screened for metabolites to indicate Krebs cycle aberrations. Multi-gene panel-sequencing was applied to detect driver mutations in cases with indicative metabolite profiles but undetermined genetic drivers.
Results:
Aberrant Krebs cycle metabolomes identified rare cases of PPGLs with germline mutations in FH and somatic mutations in IDHx and SDHx, including the first case of a somatic IDH2 mutation in PPGL. Metabolomics also reliably identified PPGLs with SDHx loss-of-function (LOF) mutations. Therefore we utilized tumor metabolite profiles to further classify variants of unknown significance in SDHx, thereby enabling missense-variants associated with SDHx LOF to be distinguished from benign variants.
Conclusion:
We propose incorporation of metabolome data into the diagnostics algorithm in PPGLs to guide genetic testing and variant interpretation and to help identify rare cases with mutations in FH and IDHx.
Keywords: metabolomics, next-generation sequencing, VUS, pheochromocytoma, paraganglioma, succinate dehydrogenase, fumarate hydratase, isocitrate dehydrogenase
INTRODUCTION
Mutations in Krebs cycle-related genes occur in different tumor entities, which hence share a common mechanism of tumorigenesis and might also respond to similar treatment strategies.1,2 Currently, there are nine genes encoding several different Krebs cycle enzymes indicated as tumor drivers: Fumarate hydratase (FH) mutations are associated with renal cell carcinomas (RCC), leiomyomas, and pheochromocytomas and paragangliomas (PPGLs).3,4 Mutations in isocitrate dehydrogenase genes 1 or 2 (IDH1/2) occur in gliomas, acute myeloid leukemia, cholangiocarcinomas, and rarely also in PPGL.5–8 Mutations in any of the five succinate dehydrogenase subunits (SDHx) or accessory proteins can lead to gastrointestinal stromal tumors, RCC, PPGL, neuroblastoma, and pituitary tumors.9–11 Additionally, malate dehydrogenase 2 (MDH2) was found to be rarely mutated in PPGL.12
In PPGLs, catecholamine-producing tumors originating from adrenal or extra-adrenal chromaffin tissue, tumor development has been ascribed to mutations in more than 20 susceptibility genes, with 30–40% of PPGLs having a hereditary background.13 Identification of tumors due to SDHx- and FH-mutations has implications for patient care and follow-up, since those patients are at higher risk for metastatic disease.3,14 Moreover, distinguishing between sporadic tumors and cases with underlying germline-mutations is important for management of patients and their families.
SDHx-, FH- and IDH1/2-mutated tumors are known to lead to abnormally high levels of one of several oncometabolites: succinate, fumarate, or 2-hydroxyglutarate (2HG). Increases in those result in the inhibition of α-ketoglutarate dependent enzymes, including prolyl hydroxylases and DNA and histone demethylases, causing a pseudohypoxic and hypermethylator phenotype.15 We and others have already established the succinate:fumarate ratio as a diagnostically highly-accurate functional indicator of pathogenic SDHx-variants.16,17 Strong elevations of fumarate as a consequence of loss-of-function mutations in FH are well-known from RCC and leiomyomas, and have also been described for PPGL.3,18 Somatic hotspot mutations of codons R132 and R172 within IDH1 and IDH2, respectively, lead to a gain-of-function responsible for the production of the D-enantiomer of 2HG, D-2HG.19 Measurement of metabolites can therefore provide a screening tool to identify tumors with underlying driver-mutations in those genes. On the other hand, metabolomics could also function as readout for classification of identified variants in metabolic enzymes.
Clinical practice guidelines recommend that genetic testing should be considered in all patients with PPGL, with testing and test interpretation carried out according to a clinical feature-driven diagnostic algorithm that establishes the likelihood of mutations.20 With the use of next-generation sequencing (NGS) now moving into routine clinical diagnostics, large numbers of genes can be tested simultaneously as part of one panel.13,21–23 As a consequence, more variants are identified; however, for many the pathogenicity is unclear and caution is warranted for interpretation, in particular for germline-variants with far-reaching implications for patients and their families.21,22,24
The importance of correct classification of gene-variants is emphasized by the example of RET Y791F, a variant previously associated with the occurrence of medullary thyroid carcinoma. Recent findings, however, showed that this variant is likely benign and may have led to unnecessary prophylactic thyroidectomies25. The development of standards and guidelines for the correct interpretation of genetic variants and the need for new approaches to functionally assess variants of unknown significance (VUS) has become increasingly significant.26 Apart from manipulated cell lines and in silico predictions, potential approaches to assess functionality include immunohistochemistry10, expression profiling27 as well as nuclear magnetic resonance28 and mass spectrometry metabolite profiling.16,17
The present study utilized mass spectrometry-based metabolomics of Krebs cycle intermediates in 395 PPGL samples to identify aberrant metabolite profiles, functionally relate them to variants in relevant genes and evaluate their potential as a diagnostic tool.
MATERIALS AND METHODS
Tumor procurement
Primary tumor tissue was collected from 391 patients with PPGL from nine centers, specimens either, snap frozen or formalin-fixed and paraffin-embedded (FFPE) after surgery (supplementary information 1). Tumor procurement was approved under Intramural Review Board protocols at each center, where patients have signed informed consent. Samples with potential contaminants (e.g. those stored in RNA-preserving citrate-based buffers) were excluded from metabolic clustering analysis. We included 230 tumors from 229 patients from a previous study that analyzed succinate:fumarate in relation to SDHx-mutations16 and 157 freshly frozen (FF) tumors from 154 newly diagnosed patients from seven centers (Bethesda/USA, Madrid/Spain, Florence/Italy, Nijmegen/The Netherlands, Dresden, Munich, Würzburg/Germany). Additionally, eight patients, where only FFPE tissue was available, were included.
Krebs cycle metabolites
In a mass spectrometry-based screening assay eight organic acids of the Krebs cycle as well as pyruvate and lactate were measured in all 387 FF and 8 FFPE tissue samples. Additionally, four amino acids, glutamate, glutamine, aspartate, and asparagine were analyzed in a sample subset. Metabolites were analyzed with a modification of a previously described method.16 Briefly, metabolites were extracted from samples with methanol, dried, resuspended in mobile phase and cleared with a 0.2 µm centrifugal filter. To improve separation, the elution gradient was changed as follows: 99% A (0.2% formic acid in water), 1% B (0.2% formic acid in acetonitrile) for 2.00 min, 100% B at 2.50 to 2.65 min, 1% B at 3.40 min and equilibration with 1% B until 5.00 min. Multiple reaction monitoring with negative electrospray ionization was used for quantification according to transitions provided in Table S1.
D-/L-Enantiomers of 2-hydroxyglutarate (2HG)
Samples were prepared as described previously.29 Briefly, methanol extracts of tumor tissue were dried and derivatized using diacetyl-l-tartaric anhydride (DATAN) dissolved in dichloromethane and acetic acid (4:1). After 30 minutes incubation at 75°C, samples were dried under a stream of nitrogen. After reconstitution in mobile phase (3.5% acetonitrile in water containing 2mM ammonium formate, ph 3.6), samples were injected onto a Shimadzu UFLC LC-20ADXR-system coupled to a tandem-mass spectrometer (Sciex QTRAP5500). Separation of D- and L-enantiomer derivatives of 2HG was achieved using a Waters XBridge® BEH C18 column (100×3mm, 2.5µm) with an isocratic mobile phase flow at 0.44 ml/min.
Genetic testing
Patients were tested for germline-mutations in established susceptibility genes by centers of origin or by the CNIO institute in Madrid through a collaborative multi-center study (prospective monoamine-producing tumor study, https://pmt-study.pressor.org/) using Sanger sequencing and/or NGS, and multiplex ligation-dependent probe amplification or custom array CGH for deletion detection.
Tumors with aberrant metabolite profiles and unknown genetic backgrounds were additionally analyzed by customized panel-sequencing (Illumina, San Diego, CA) including coding exons and 20-bp of intronic flanking regions of known PPGL-associated genes RET, VHL, NF1, MAX, TMEM127, SDHA, SDHB, SDHC, SDHD, SDHAF2, MDH2, FH, EPAS1, HRAS, KIF1B, EGLN1, EGNL2, IDH1 and IDH2. Library preparation was performed using Illuminas’ TruSeq Nano DNA Library Prep Kit according to the manufacturer’s instructions. One hundred-fifty nt paired-end sequencing was carried out with a minimum median coverage of 1000-fold either on an Illumina MiSeq or NextSeq sequencer. Reads were aligned to the reference genome (GRCh37/hg19) and variant-calling was performed using in house workflows, as described previously.22,30 Variants of interest were validated by Sanger sequencing.
Variant-classification was performed in accordance with the standards and guidelines of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology (ACMG- AMP),26 considering loss of heterozygosity (LOH) in tumor samples as an additional supporting criterion for pathogenicity (PP6). For SDHx-variants, immunohistochemical loss of the SDH-complex as well as an elevated succinate:fumarate were interpreted as functional analyses (PS3) but were downgraded to moderate (PM7 and PM8) criteria. In contrast, positive immunohistochemistry of the SDH-complex and normal succinate:fumarate were interpreted to support non-functional variants (BP8).
Immunohistochemistry (IHC)
FFPE tumor tissue was sectioned and stained for SDHB or FH protein using rabbit polyclonal anti-SDHB (HPA002868, Sigma-Aldrich), anti-SDHA (ab14715, Abcam or 2E3GC12FB2AE2, Biorad), and anti-FH (HPA027341, Sigma-Aldrich).
Statistical analysis
Metabolome data were normalized to natural logarithms (LN) to account for large value discrepancies. Basic statistics and clustering analysis were performed using JMP statistics software (version Pro 12.1.0). Outlier analysis on LN transformed values or ratios were used to identify candidates for panel-sequencing of metabolic genes. For this purpose we focused on outliers above the 75th quartile + 1.5 x (interquartile range) marked by the top whisker in box and whisker plots. Significance between groups of patients was calculated using the nonparametric Wilcoxon test.
RESULTS
Metabolite profiling in PPGL
Clustering analysis of the full set of 395 PPGLs from 391 patients using ratios of Krebs cycle intermediates identified two clusters involving three main groups (Figure 1A, supplementary information 1): Cluster I was characterized by high succinate and low levels of all other Krebs cycle metabolites and almost exclusively contained SDHx-mutated tumors. Additionally, lower levels of the amino acids aspartate and glutamate were present in SDHx-mutated PPGLs compared to PPGLs due to other causes (Figure S1). PPGLs of the first cluster, with the exception of three tumors, had succinate:fumarate ratios above a previously defined cut-off for SDHx-mutated PPGLs (Figure 1B).16
Figure 1. Krebs cycle metabolite profiling of PPGLs.
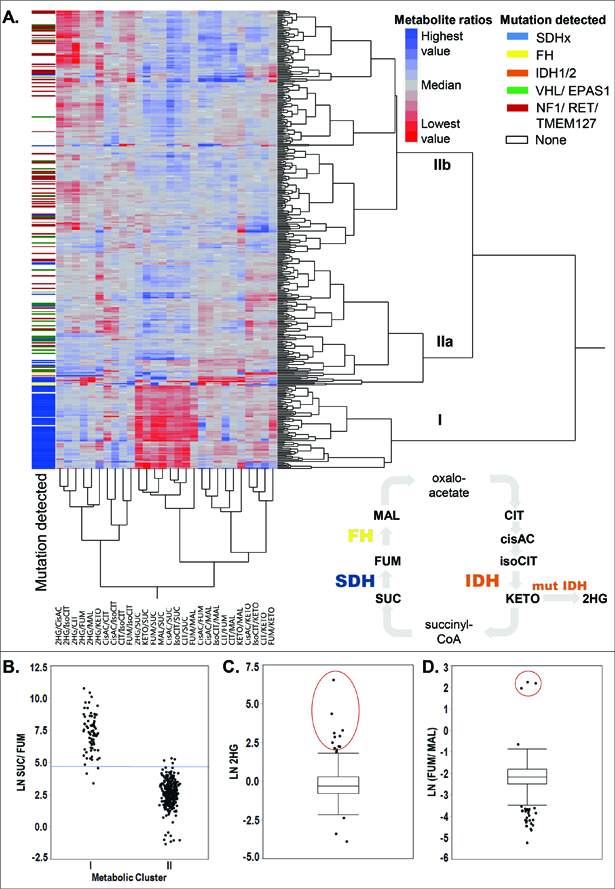
A. Hierarchical cluster analysis (Ward) of 395 different PPGLs from 391 patients. All possible ratios between eight intermediates (citrate, isocitrate, cis-aconitate, a-ketoglutarate, 2-hydroxyglutarate, succinate, fumarate, malate) of the central carbon metabolism were generated and LN transformed. Mutational status (germline and somatic) is shown by color in the left column. Clusters are marked with roman numbers. The Krebs cycle with all measured metabolites is schematically shown; oxaloacetate and succinyl-CoA are not detectable with the used method. Abbreviations: SUC-succinate, FUM-fumarate, MAL-malate, CIT-citrate, cisAC-cis-aconitate, isoCIT-isocitrate, KETO-α-ketoglutarate, 2HG-2-hydroxyglutarate. B. The succinate:fumarate ratio as an indicator for mutations in SDHx genes separates cluster I from II. Cut-off defined in Richter et al. 2014 marked in blue. C. Box plots of 2HG levels with outliers marked as dots. Outliers above the top whisker (the 75th quartile + 1.5 x (interquartile range)) were further investigated by NGS analysis when the driver mutation was unknown (red circle). D. Outlier analysis for the fumarate:malate ratio.
Cluster II was divided into two main subclusters separated by different levels of citrate, isocitrate, and cis-aconitate (Figure 1A). Subcluster IIa was characterized by low levels of the aforementioned metabolites and contained primarily tumors with driver mutations in the hypoxia signaling axis (VHL, EPAS1), as well as tumors with FH and IDHx mutations (characterized below in detail). Subcluster IIb, characterized by higher citrate, isocitrate, and cis-aconitate levels, includes predominantly tumors with driver mutations in the kinase signaling pathway (RET, NF1, TMEM127). This metabolic differentiation between groups with mutations in common pathways reflects somewhat the clustering of PPGLs according to transcriptomics.31
By applying outlier analysis to our cohort, we identified outliers for 2HG (Figure 1C), α-ketoglutarate (Figure S2A), the fumarate:malate ratio (Figure 1D), fumarate (Figure S2B), and malate (Figure S2C). The following sections describe the identification of underlying mutations in those samples.
2HG and α-ketoglutarate elevations and IDHx-mutations in PPGL
Twelve outliers were identified for 2HG. Of those, two were also α-ketoglutarate outliers (Figure 1C, Figure S1A). Nine of the twelve 2HG outliers were available for further analysis. Patient #1 and #2 had the highest 2HG values of the set, with increases of up to 1800-fold (#2) and 100-fold (#1) over the median (Figure 2A). The other seven tumors showed increases of 10- to 40-fold above the median and four of nine PPGLs also had increases in α-ketoglutarate of 9- to 48-fold over the median (Figure 2B).
Figure 2. Isocitrate dehydrogenase 1 and 2 gain-of-function mutations in PPGLs.
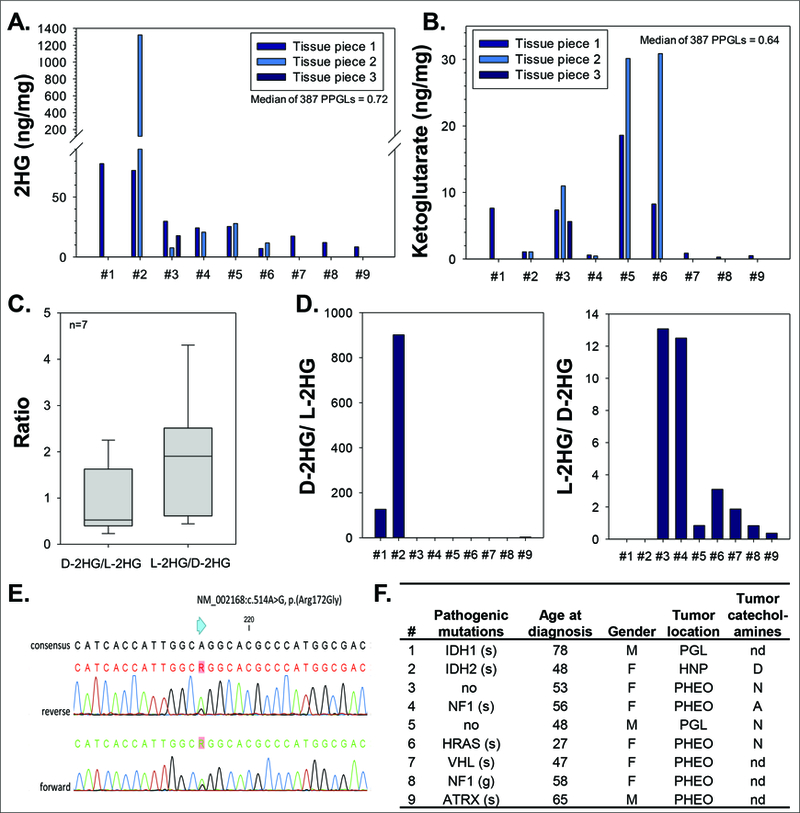
A. Total 2-hydroxyglutarate (2HG) in nine tumors with the highest 2HG values of the set of fresh frozen tumors. B. Total α-ketoglutarate in nine tumors with the highest 2HG values of the set. C. Quantification of D- and L-enantiomers of 2HG in seven control PPGLs with moderate total 2HG. D. Quantification of D-2HG and L-2HG in the nine tumors with highest total 2HG. E. Sanger sequencing traces of PPGL #2 analyzed for a mutation in IDH2. F. Patient information of PPGLs with highest total 2HG. For further details refer to supplementary information 1. PGL – extra-adrenal; HNP – head and neck; PHEO – adrenal; nd – not determined; D – dopaminergic; N – noradrenergic; A – adrenergic; g – germline and s – somatic variant; # – patient number.
To further characterize the 2HG elevations, D- and L-enantiomers of 2HG were measured. Seven PPGLs with moderate total 2HG levels served as controls with D-2HG to L-2HG ratios ranging from 0.2 to 1.6 (Figure 2C). Tumors of patient #1 and #2 showed strongly increased D-2HG to L-2HG ratios of 127 and 901, respectively (Figure 2D). Tumors form patients #3 and #4 had L-2HG to D-2HG ratios of 13 with control tumors ranging from 0.4 to 4.3. The other five PPGLs were similar to controls.
Patient #1 has been described elsewhere with a somatic IDH1 c.394C>T (R132C) mutation,6 explaining the strong increases in D-2HG now described here for the first time. In patient #2 we identified a somatic IDH2 c.514A>G (R172G) mutation, to our knowledge the first IDH2-mutation described in PPGL to date (Figure 2E). A detailed patient description is provided in supplementary information 2. The other seven PPGLs with moderately elevated 2HG-levels and normal D-2HG to L-2HG ratios did not have mutations in any of the genes of our NGS-panel; however, in five PPGLs we identified pathogenic mutations in other known susceptibility genes, including NF1, HRAS, ATRX, and VHL (Figure 2F). In summary, IDH1/2-mutated PPGLs were characterized by elevations of total 2HG of at least 100-fold and of D-2HG to L-2HG ratios over 100-fold above control median values.
Fumarate and malate elevations and FH-mutations in PPGL
Outlier analysis identified three samples (patients #10, 11, 12) with strong elevations in fumarate:malate ratios 60- to 80-fold above median values of controls (Figure 1D). Five outliers for absolute fumarate levels were identified. Three of those also presented as outliers for malate and had normal fumarate:malate ratios (Figure S2B-C), the other two (#10, 11) were also fumarate:malate outliers.
In all three patients with aberrant tumor fumarate:malate, NGS revealed heterozygous germline-variants in FH: NM_000143.3:c.700A>G p.(Thr234Ala), c.908T>C p.(Leu303Ser), and c.816_836del p.(Ala273_Val279del) (Figure 3A-C). In all cases, the tumors showed LOH for these variants. IHC confirmed the loss of FH protein in tumor #10 and #11, for #12 no more tissue was available (Figure 3D).
Figure 3. Metabolite profiling identifies pathogenic fumarate hydratase (FH) mutations previously classified as VUS.
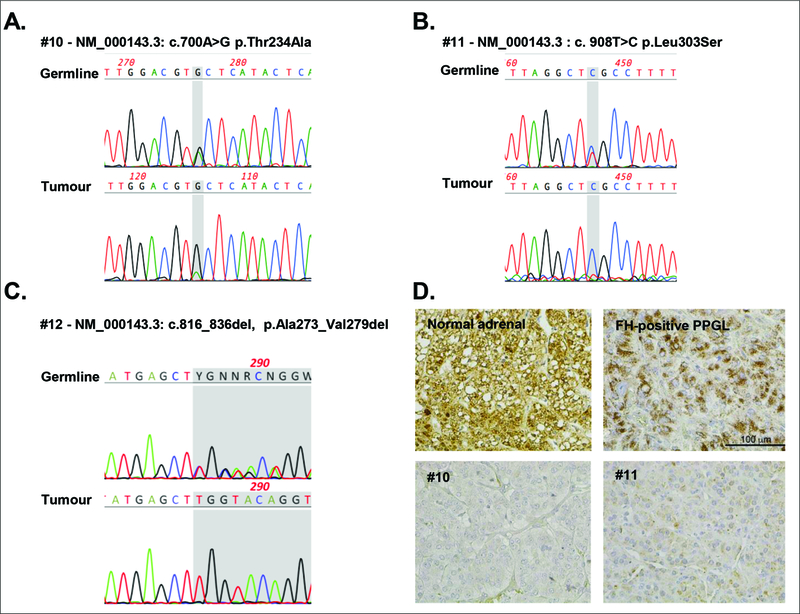
Sanger sequencing traces of leukocyte (germline) and tumor DNA from patient #10 (A), #11 (B), and #12 (C). D. Immunohistochemical staining for FH of normal adrenal, FH-wildtype and FH-mutated PPGL tissue from patient #10 and #11; 400x magnification.
The FH-variants in patients #10 and #11 - p.(Thr234Ala) and p.(Leu303Ser) - are both missense-variants predicted to be damaging by three different in silico tools (Polyphen2/ SIFT/ MutationTaster). Both are listed three times in heterozygous states in the gnomAD database. FH p.(Thr234Ala) is rated with pathogenic/uncertain significance in ClinVar and dbSNP (rs372505976) and FH p.(Leu303Ser) as a VUS (rs201502246). The variant FH p.(Leu303Ser) was found in a girl together with a second compound heterozygous variant in FH, who suffered from autosomal-recessive fumarase deficiency (LOVD #0000116260)32. The variant p.(Thr234Ala) has not, to the best of our knowledge, been reported in the literature. The third patient (#12) carried an in-frame deletion leading to loss of 6 amino-acids in FH (p.(Ala273_Val279del)). This variant is rated pathogenic in ClinVar and dbSNP (rs863223985). It is not listed in gnomAD or described in the literature. With only this information available, the variants would be interpreted as VUS according to ACMG-AMP criteria. The metabolomics-based aberrant tumor fumarate:malate ratios and elevated fumarate levels, which indicates loss-of-function of FH, combined with the IHC-confirmed loss of tumor FH protein in patient #10 and #11and LOH provides sufficient evidence to classify all three variants as likely pathogenic.
All three patients with germline FH-mutations (#10–12) had unilateral adrenal pheochromocytomas with a noradrenergic biochemical phenotype (see supplementary information 2). None of the patients had metastatic disease at initial presentation (#10) or six (#11) to 18 years (#12) later. Family history included RCC in the father of patient #11 and thyroid cancer and melanoma in patient #10.
The three other fumarate outliers with normal fumarate:malate ratios, which also showed outlier profiles for malate (Figure S2C), did not show variants in FH but had SDHx-variants classified as likely pathogenic or pathogenic (Table 1 and supplementary material 3). Since those tumors did not show aberrant fumarate:succinate ratios, this might indicate that the outlier profiles for those samples are possibly related to sampling problems. In summary, fumarate:malate ratios above 7.0 with non-FH mutated PPGLs having values below 1.0 provide a useful guide to the presence of FH-mutations in PPGL. Increases in tumoral fumarate alone may not offer reliable guidance to the presence of FH-mutations.
Table 1.
Pathogenic and likely pathogenic variants in SDHx genes identified in this study
| Gene | Nucleotide change |
Predicted protein change |
Mutation type |
Germline status |
S:F above cut-off |
Phenotype | Reference SNP No | n patients (families) |
AF in gnomAD in % | ClinVar | Known from literature/ Disease databases |
|---|---|---|---|---|---|---|---|---|---|---|---|
| SDHB | c.72+1G>T | splice | germline | yes | PGL | rs587782703 | 1 | - | pathogenic | yes | |
| c.136C>T | p.(Arg46*) | stop | germline | yes | 3x PGL | rs74315370 | 3 | 0.001805 | pathogenic | yes | |
| c.166_170del | p.(Pro56Tyrfs*5) | fs | germline | yes | 1x PGL | rs786202100 | 1 | - | pathogenic | yes | |
| c.183T>G | p.(Tyr61*) | stop | somatic | yes | 1x PGL | - | 1 | - | - | yes | |
| c.268C>T | p.(Arg90*) | stop | germline | yes | 2x PGL, 1x HNP | rs74315366 | 3 | 0.00041 | pathogenic | yes | |
| c.287–1G>C | - | splice | germline | yes | 1x PGL | rs397516833 | 1 | - | - | yes | |
| c.287–3G>C | - | potential
splice |
germline | yes | 2x PGL | rs397516834 | 2 | - | - | yes | |
| c.343C>T | p.(Arg115*) | stop | germline | yes | 1x PGL | rs751000085 | 1 | 0.00081 | pathogenic | yes | |
| c.348_352del | p.(Ile117Hisfs*5) | fs | germline | yes | 1x PHEO | - | 1 | - | - | - | |
| c.423+1G>A | - | splice | germline | yes | 3x PGL | rs398122805 | 3 | 0.0012 | pathogenic | yes | |
| c.445_446del insGGTATCT |
p.(Gln149Glyfs*11) | fs | germline | yes | 1xPGL | - | 1 | - | - | yes | |
| c.503dup | p.(Gln169Alafs*10) | fs | germline | yes | 2x PHEO | - | 2(1) | - | - | - | |
| c.587G>A | p.(Cys196Tyr) | missense | germline | yes | 1x PHEO | - | 1 | - | pathogenic | yes | |
| c.649C>T | p.(Arg217Cys) | missense | germline | no1 | 2x HNP | rs200245469 | 2 | - | likely pathogenic | yes | |
| c.689G>A | p.(Arg230His) | missense | germline | yes | 2x PGL | rs587782604 | 2 | 0.00041 | pathogenic/
likely pathogenic |
yes | |
| c.725G>A | p.(Arg242His) | missense | germline | yes | 2x HNP, 1x PGL | rs74315368 | 3 | 0.00081 | pathogenic | yes | |
| SDHC | c.43C>T | p.(Arg15*) | stop | germline | yes | 1x HNP | rs201286421 | 1 | 0.00082 | pathogenic | yes |
| c.126G>A | p.(Trp42*) | stop | germline | no | 1x PGL | - | 1 | - | - | yes | |
| SDHD | c.3G>T | startloss | somatic | yes | 1xHNP | - | 1 | - | - | yes | |
| c.14G>A | p.(Trp5*) | stop | germline | yes | 1x HNP | rs104894310 | 1 | - | pathogenic | yes | |
| c.36_37del | p.(Ala13Profs*55) | fs | germline | yes | 1xPHEO | - | 1 | - | - | yes | |
| c.49C>T | p.(Arg17*) | stop | germline | yes | 1xHNP | - | 1 | - | - | yes | |
| c.53–2A>G | splice | germline | yes | 1xHNP | - | 1 | - | - | - | ||
| c.53dup | p.(Leu19Serfs*50) | fs | germline | yes | 1xHNP | - | 1 | - | pathogenic/ likely pathogenic |
- | |
| c.64C>T | p.(Arg22*) | stop | germline | yes | 2xHNP | rs104894306 | 2(1) | - | pathogenic | yes | |
| c.169+1G>A | splice | germline | (yes)2 | 1x HNP | - | 1 | - | - | yes | ||
| c.170–1G>T | splice | germline | yes | 1x PHEO | - | 1 | 0.00041 | - | yes | ||
| c.242C>T | p.(Pro81Leu) | missense | germline | yes (5/7) | 7x HNP | rs80338844 | 7 | 0.0018 | pathogenic | yes | |
| c.274G>T | p.(Asp92Tyr) | missense | germline | yes | 2x HNP | rs80338845 | 2 | - | pathogenic | yes | |
| c.284T>C | p.(Leu95Pro) | missense | germline | yes | 1x PHEO, 1x PGL, 1x HNP | rs80338846 | 3(2) | - | pathogenic | yes | |
| c.298_301del | p.(Thr100Phefs*34) | fs | germline | yes | 1x HNP | rs786203067 | 1 | - | pathogenic | yes | |
| c.325C>T | p.(Gln109*) | stop | germline | yes | 1x PHEO, 1x PGL | - | 2(1) | - | pathogenic | yes | |
| c.337_340del | p.(Asp113fs) | fs | somatic | yes | 1x PGL | - | 1 | - | pathogenic | yes | |
| c.341A>G | p.(Tyr114Cys) | missense | germline | yes | 2xHNP | rs104894304 | 2 | - | pathogenic | yes | |
| c.416T>C | p.(Leu139Pro) | missense | germline | no | 1xPHEO | rs80338847 | 1 | - | pathogenic | yes | |
| SDHA | c.1753C>T | p.(Arg585Trp) | missense | germline | yes | 1xPHEO | rs200397144 | 1 | 0.0022 | inconclusive | yes |
| SDHAF2 | c.232G>A | p.(Gly78Arg) | missense | germline | yes | 3xHNP | rs113560320 | 3(2) | - | pathogenic | yes |
Abbreviations: AF, allele frequency; gnomAD, The Genome Aggregation Database; S:F, succinate:fumarate; SNP, single nucleotide polymorphism; n, number; fs, frameshift; PHEO, adrenal pheochromocytoma; PGL, extra-adrenal paraganglioma; HNP, head and neck paraganglioma
Metabolic-Cluster 1 tumor despite normal S:F ration indicating aberrant SDH (likely normal tissue contamination)
normal succinate:fumarate ratios were found in the first tumor tissue, but measurement of a second peace confirmed highly elevated succinate:fumarate ratios
Succinate:fumarate ratios, metabolic cluster 1 and SDHx-variants in PPGL
In our cohort of 391 patients we identified 73 individuals having PPGLs with aberrant succinate:fumarate ratios.16 All except five tumors belonged to the first metabolic cluster (Figure 1A). Of those 73 patients, 34 had clear loss-of-function mutations (e.g. stop gain, frameshift, or exon-spanning deletions) in one of the SDHx-genes in the germline. Furthermore, another 32 patients had missense SDHx-variants in the germline that are well established as pathogenic or likely pathogenic (n=21) or would be currently classified as VUS (n=10) or likely benign (1), according to ACMG-AMP guidelines (see tables 1, 2 and supplementary material 3). Additionally, three patients (#57, 77, 92) had tumors of metabolic cluster I with confirmed pathogenic or likely pathogenic SDHx-mutations but with succinate:fumarate ratios below the cut-off.
Table 2.
Variants in SDHx genes identified in this study that would be currently classified as VUS according to ACMG with potential reclassification based on succinate:fumarate ratios
| Gene | Nucleotide change |
Predicted protein change |
Mutation type |
Germline status |
S:F above cut-off |
Phenotype | Reference SNP No |
n patients (families) |
AF in gnomAD in % |
ClinVar | Known from literature/ disease databases |
further information | Final classification* |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SDHB | c.32G>A | p.(Arg11His) | missense | germline | no | 1x PHEO | rs111430410 | 1 | 0.051 | conflicting | yes | Reported in
SDHB positive tumor (IHC) |
likely benign |
| c.49A>G | p.(Thr17Ala) | missense | germline | no | 1x PHEO | - | 1 | - | uncertain significance |
yes | somatic likely pathogenic VHL, no LOH |
likely benign | |
| c.119A>C | p.(Lys40Thr) | missense | germline | yes | 1x HNP | - | 1 | - | - | - | VUS | ||
| c.325A>C | p.(Asn109His) | missense | germline | yes | 1x PGL | - | 1 | 0.00041 | - | - | VUS | ||
| c.380T>G | p.(Ile127Ser) | missense | germline | yes | 1x PGL/1x HNP | rs786201095 | 2 | 0.0012 | pathogenic/ likely pathogenic |
yes | likely pathogenic | ||
| c.418G>T | p.(Val140Phe) | missense | germline | yes | 2x PGL | rs267607032 | 2 | 0.0012 | pathogenic/ likely pathogenic |
yes | likely pathogenic | ||
| c.464C>G | p.(Pro155Arg) | missense | somatic | yes | 1x PGL | - | 1 | - | - | yes | likely pathogenic | ||
| c.572G>A | p.(Cys191Tyr) | missense | germline | yes | 1x HNP | - | 1 | - | - | yes | SDHB negative | likely pathogenic | |
| c.574T>C | p.(Cys192Arg) | missense | germline | yes | 1x PHEO | rs786202732 | 1 | - | pathogenic | yes | likely pathogenic | ||
| SDHC | c.179G>T | p.(Ser60Ile) | missense | germline | yes | 1x HNP | - | 1 | - | - | - | SDHB IHC positive | VUS |
| c.200T>C | p.(Met67Thr) | missense | germline | no | 1x PGL | - | 1 | 0.00041 | - | - | no LOH | VUS | |
| c.256_257ins TTT |
p.(Gly86delinsValC ys) |
in-frame del | germline | no | 1x HNP | - | 1 | - | - | yes | VUS | ||
| c.379C>T | p.(His127Tyr) | missense | germline | yes | 1x PHEO | - | 1 | - | - | yes | no LOH,
SDH-loss in IHC described in literature |
likely pathogenic | |
| SDHD | c.169+5G>T | potential splice |
germline | (yes)1 | 1x HNP | - | 1 | - | - | yes | IHC SDHB positive; LOH; skipping exon 2 cDNA in literature |
likely pathogenic | |
| c.239T>C | p.(Leu80Pro) | missense | germline | no | 1x PHEO | - | 1 | - | - | - | VUS | ||
| SDHA | c.−7A>C | 5’UTR substitution |
unknown | no | 1xPHEO | rs751633537 | 1 | 0.067 | inconclusive | - | VUS | ||
| c.1421C>T | p.(Ser474Leu) | missense | germline | no | 1xPHEO | - | 1 | - | - | - | somatic EPAS1
c.1592C>G |
likely benign | |
| c.1772C>T | p.(Ala591Val) | missense | germline | no | 1xHNP | rs367621815 | 1 | 0.0018 | - | - | VUS | ||
| SDHAF2 | c.320G>A | p.(Arg107His) | missense | germline | no | 1xPGL | rs535627239 | 1 | 0.018 | - | - | no LOH | VUS |
Abbreviations: VUS, variant of unknown significance; AF, allele frequency; gnomAD, The Genome Aggregation Database; S:F, succinate:fumarate; SNP, single nucleotide polymorphism; n, number; PHEO, adrenal pheochromocytoma; PGL, extra-adrenal paraganglioma; HNP, head and neck paraganglioma; IHC, immunohistochemistry; LOH, loss of heterozygosity
normal succinate:fumarate ratios were found in the first tumor tissue, but measurement of a second peace confirmed highly elevated succinate:fumarate ratios
In seven of the 73 patients with elevated succinate:fumarate ratios, no SDHx-variants were identified in the germline. In three of those patients we identified somatic SDHx-variants: a stop-mutation (SDHB p.Tyr61*, #30); a startloss-mutation (SDHD c.3G>T, #70), and a missense-mutation (SDHB p.(Pro155Arg), #105) (supplementary material 3). In a further patient we previously reported somatic SDHC-promotor-methylation (#107,33 explaining the SDH-deficiency-phenotype. Two cases (#106,108) currently remain unsolved. We excluded somatic SDHx-mutations and SDHC-promoter-methylation in those cases. In #106 we reported germline-variants in OGDHL and PCK2; however their significance is unclear.6 The seventh case had a pathogenic germline RET c.1852T>G, p.(Cys618Gly) mutation. Therefore, we interpreted the borderline succinate:fumarate elevation (#109) as a false-positive result.
One patient (#69) with an elevated tumoral succinate:fumarate was particularly interesting. He had a likely benign germline missense-variant SDHD c.34G>A, p.(Gly12Ser), which is listed 2035x heterozygously and 12x homozygously in the gnomAD database and has been described as a rare polymorphism.34 The patient presented with sympathetic paraganglioma of the renal hilum at the age of 21 and no family history of PPGL. Since germline-testing was inconclusive in this case, we analyzed tumor tissue and identified a pathogenic somatic loss-of-function variant in SDHD NM_003002.3: c.337_340del, p.(Asp113Metfs*21) in 41.47% of reads. Allele frequency of the germline-variant c.34G>A was 69.7% in the tumor, indicating LOH of the wild-type SDHD-allele and that the somatic and pathogenic SDHD-variant p.(Asp113Metfs*21) occurred in the allele having the germline c.34G>A variant. We therefore assume that the somatic SDHD-variant p.(Asp113Metfs*21) is responsible for SDH deficiency in this tumor and that the germline-variant is likely benign and coincidently observed together with the aberrant metabolite profile.
In ten patients with elevated succinate:fumarate ratios we identified eight different missense-variants in SDHx (including one somatic) that would currently be classified as VUS. Most of the variants had previously been described in PPGL cases and integrating the metabolomics profiles together with all available information (see supplementary information 3), we would now reclassify five of those variants as likely pathogenic (Table 2).
On the other hand, in 22 patients with succinate:fumarate ratios that were not suspicious, germline-variants in SDHx were identified during routine diagnostics testing. Three are known benign or likely benign SDHx-variants (two missense, one silent), in line with the normal succinate:fumarate ratios. However, nine patients had pathogenic or likely pathogenic germline-mutations, including one frame-shift, one splice site, one gross-deletion, and four different missense-mutations already described in PPGL families. Three of those (#57, 77, 92) had tumors belonging to metabolic cluster I despite their normal succinate:fumarate. Seven of the tumors furthermore were head and neck paragangliomas, known to contain high amounts of stroma. For none of the samples, histological estimation of tumor content was performed. Testing of additional tumor specimens in these patients revealed highly elevated ratios in three cases, supporting that these were false-negatives due to insufficient sampling of tumor cells.
The remaining ten cases with normal succinate:fumarate had variants that would be currently classified as VUS: seven missense, one in-frame deletion, one potential splice site, and one substitution in the 5’UTR. Metabolomics analysis for additional tumor samples in eight cases confirmed normal succinate:fumarate ratios. Further analyses revealed somatic pathogenic mutations in VHL and EPAS1 in two cases (#14, 50) and we would reclassify the SDHx-variants as likely benign (Table 2). Another variant, p.(Arg11His, #23), has been described in an SDHB-positive tumor and together with metabolite data we would classify it as likely benign.35 In a patient (#78) with a potential splice-site-mutation in SDHD (c.169+5G>T) metabolomics of a second tumor specimen revealed highly elevated succinate:fumarate ratios and we would altogether reclassify this variant as likely pathogenic. For the remaining six variants, despite normal succinate:fumarate, there is currently insufficient evidence for or against pathogenicity to reclassify these variants (see Table 2 and supplementary information 3).
Among the total of 19 VUS in SDHx identified in our cohort, subsequent to comprehensive analysis of the metabolome and the tumor genome, we would reclassify seven (37%) as likely pathogenic and three (16%) as likely benign (Table 2). The overall diagnostic performance was comparable to previously published results and is summarized in supplementary information 4.16
DISCUSSION
Stratification of PPGL patients according to underlying mutations is a crucial step of personalized patient management. Mutations in Krebs cycle-related genes are established drivers of tumor development, acting through the inhibition of α-ketoglutarate-dependent enzymes. Identifying such driver mutations has important implications for the surveillance of patients at risk of recurrence or metastasis or of carriers of germline-variants who are at risk of tumor development.
Here we show that quantification of metabolites in tumor tissue can guide identification of underlying driver germline or somatic mutations in patients with unresolved results of genetic testing. More specifically, characteristic metabolite profiles involving high succinate:fumarate or fumarate:malate ratios or high levels of 2HG in combination with high D-2HG/L-2HG ratios can respectively be used to guide detection of SDHx-, FH- and IDH1/2-mutations. The described mass spectrometric-based assay is simple, cheap and uses the same instrumentation required for measurements of metanephrines, a standard test in the workup of patients with suspected PPGL.36 Metabolite data together with the catecholamine biochemical phenotype and clinical presentation can be used to narrow down the list of genes for Sanger sequencing.13 Moreover, isolated PPGL cases with rare somatic mutations in IDHx or SDHx can be identified, and thereby rule out a hereditary disease in those patients.
A particular advantage of metabolome-guided genomics is that tumors with similar phenotypic presentations in terms of metabolite accumulation or depletion can be characterized, leading to identification of mechanisms of silencing known PPGL genes or even of novel susceptibility genes. Recent examples are the identification of SDHC-promoter-methylation and a GOT2 activating mutation in PPGL with a similar clinical presentation as in SDHx-mutated PPGLs.6,33 In our cohort, we identified five cases with somatic SDHx-alterations, including one SDHC-promotor-methylation, all currently considered extremely rare events.6,33 We also for the first time report a somatic IDH2-mutation in a PPGL.
The metabolome-guided identification of three patients with germline FH-mutations provides further information to clarify the clinical presentation of FH-mutated PPGLs. Consistent with the adrenal locations of tumors in our three patients, nine of eleven previously published cases also involved adrenal tumors.3,37–39 All three pheochromocytomas of the current study had the expected noradrenergic phenotype, but such phenotypic information has only been available for one previously described patient also with a noradrenergic tumor. Two patients under surveillance had not developed metastasis six or 18 years after initial diagnosis. Follow-up remains strictly recommended also due to the increased risk for RCC and leiomyomas.4
In addition to guiding genetic testing, metabolome analysis is also useful for guiding data analysis and interpretation of gene-variants. The latter is particularly valuable with multi-gene panels that are associated with increased identification of coincidental, non-pathogenic variants. Interpretation of test results is often difficult, since many variants are uncharacterized and their disease-related impact remains unknown, leading to uncertainty for both patients and healthcare providers. Provision of false-positive results to patients can have significant negative consequences for patients and their families. Guidelines published by the ACMG and AMP give clear instructions on the interpretation of germline-variants26. Here we show that characterization of tumor tissue through mass spectrometry-based analysis of Krebs cycle metabolites can be employed to guide variant interpretation. When aberrant metabolomics profiling is in concordance with a VUS in a matching Krebs cycle gene, we propose that this can be used as supporting criteria for pathogenicity; if tumor metabolomics does not support a deficiency in the corresponding gene, this can be a supporting criterion against pathogenicity. Therefore, if germline genetic testing is inconspicuous or inconclusive (VUS), genetic testing of tumor tissue accompanied by metabolomics can be employed along with immunohistochemistry where appropriate. Importantly, in cases with a clear pathogenic metabolite tumor profile but without a clear pathogenic mutation in the corresponding genes, alternative methods to analyze the same genes, such as promotor-methylation, should be considered.
Unremarkable metabolite profiles should be interpreted in context and do not exclude a Krebs cycle gene mutation; false-negative results can be due to high amounts of normal or stromal tissue within the sample. In line with a previous study, we found that false-negative results mainly occur in head and neck paragangliomas16 (supplementary information 4), which may be explained by subforms with low amounts of tumor cells relative to tumor volume.40 If possible, several tissue specimens from different regions of the tumor should be investigated. Importantly, tissue specimens should be scored for tumor content by a pathologist for both, metabolomics and genetic testing.
In summary, metabolomics is a useful tool for guiding identification of SDHx-, FH-, and IDH1/2-mutations and in variant-interpretation with implications not only in PPGL, but also in other tumor diseases caused by Krebs cycle gene mutations. Based on our experiences, we propose a cumulative approach for comprehensive PPGL diagnostics, including sequencing of germline and tumor tissue, as well as metabolic analyses in patients with inconclusive results of genetic testing; particularly in cases with VUS in Krebs cycle associated genes and unsolved cases to guide diagnostic decision making.13 Our study underlines that genetic diagnostics in PPGLs is a multidisciplinary approach combining the expertise of geneticists, endocrinologists, clinical chemists, and pathologists.
Supplementary Material
ACKNOWLEDGEMENTS
This study was funded by the Deutsche Forschungsgemeinschaft (RI 2684/1–1; KL 2541/2–1 and CRC/Transregio 205/1, B12 by Klink/ Eisenhofer/ Lenders), the AES PI17/01796, co-financed by Fondo Europeo de Desarrollo Regional (FEDER), the European Union Seventh Framework Programme (FP7/2007–2013) under grant agreement no. 259735, the Paradifference foundation, and the Intramural Research Program of the NIH, NICHD.
Footnotes
Conflict of interest: The authors have nothing to disclose.
References
- 1.Gaude E, Frezza C. Defects in mitochondrial metabolism and cancer. Cancer & metabolism 2014;2:10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Adam J, Yang M, Soga T, Pollard PJ. Rare insights into cancer biology. Oncogene 2014;33(20):2547–2556. [DOI] [PubMed] [Google Scholar]
- 3.Castro-Vega LJ, Buffet A, De Cubas AA, et al. Germline mutations in FH confer predisposition to malignant pheochromocytomas and paragangliomas. Human molecular genetics 2014;23(9):2440–2446. [DOI] [PubMed] [Google Scholar]
- 4.Schmidt LS, Linehan WM. Hereditary leiomyomatosis and renal cell carcinoma. International journal of nephrology and renovascular disease 2014;7:253–260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Clark O, Yen K, Mellinghoff IK. Molecular Pathways: Isocitrate Dehydrogenase Mutations in Cancer. Clinical cancer research : an official journal of the American Association for Cancer Research 2016;22(8):1837–1842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Remacha L, Comino-Mendez I, Richter S, et al. Targeted Exome Sequencing of Krebs Cycle Genes Reveals Candidate Cancer-Predisposing Mutations in Pheochromocytomas and Paragangliomas. Clinical cancer research : an official journal of the American Association for Cancer Research 2017. [DOI] [PubMed] [Google Scholar]
- 7.Gaal J, Burnichon N, Korpershoek E, et al. Isocitrate dehydrogenase mutations are rare in pheochromocytomas and paragangliomas. The Journal of clinical endocrinology and metabolism 2010;95(3):1274–1278. [DOI] [PubMed] [Google Scholar]
- 8.Fishbein L, Leshchiner I, Walter V, et al. Comprehensive Molecular Characterization of Pheochromocytoma and Paraganglioma. Cancer cell 2017;31(2):181–193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Xekouki P, Stratakis CA. Succinate dehydrogenase (SDHx) mutations in pituitary tumors: could this be a new role for mitochondrial complex II and/or Krebs cycle defects? Endocrine-related cancer 2012;19(6):C33–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Evenepoel L, Papathomas TG, Krol N, et al. Toward an improved definition of the genetic and tumor spectrum associated with SDH germ-line mutations. Genetics in medicine : official journal of the American College of Medical Genetics 2015;17(8):610–620. [DOI] [PubMed] [Google Scholar]
- 11.Dahia PL. Pheochromocytoma and paraganglioma pathogenesis: learning from genetic heterogeneity. Nature reviews Cancer 2014;14(2):108–119. [DOI] [PubMed] [Google Scholar]
- 12.Cascon A, Comino-Mendez I, Curras-Freixes M, et al. Whole-exome sequencing identifies MDH2 as a new familial paraganglioma gene. Journal of the National Cancer Institute 2015;107(5). [DOI] [PubMed] [Google Scholar]
- 13.Eisenhofer G, Klink B, Richter S, Lenders JW, Robledo M. Metabologenomics of Phaeochromocytoma and Paraganglioma: An Integrated Approach for Personalised Biochemical and Genetic Testing. Clin Biochem Rev 2017;38(2):69–100. [PMC free article] [PubMed] [Google Scholar]
- 14.Gimenez-Roqueplo AP, Favier J, Rustin P, et al. Mutations in the SDHB gene are associated with extra-adrenal and/or malignant phaeochromocytomas. Cancer research 2003;63(17):5615–5621. [PubMed] [Google Scholar]
- 15.Favier J, Amar L, Gimenez-Roqueplo AP. Paraganglioma and phaeochromocytoma: from genetics to personalized medicine. Nature reviews Endocrinology 2015;11(2):101–111. [DOI] [PubMed] [Google Scholar]
- 16.Richter S, Peitzsch M, Rapizzi E, et al. Krebs cycle metabolite profiling for identification and stratification of pheochromocytomas/paragangliomas due to succinate dehydrogenase deficiency. The Journal of clinical endocrinology and metabolism 2014;99(10):3903–3911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kim E, Wright MJ, Sioson L, et al. Utility of the succinate: Fumarate ratio for assessing SDH dysfunction in different tumor types. Molecular genetics and metabolism reports 2017;10:45–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Pollard PJ, Briere JJ, Alam NA, et al. Accumulation of Krebs cycle intermediates and over-expression of HIF1 alpha in tumours which result from germline FH and SDH mutations. Human molecular genetics 2005;14(15):2231–2239. [DOI] [PubMed] [Google Scholar]
- 19.Dang L, White DW, Gross S, et al. Cancer-associated IDH1 mutations produce 2-hydroxyglutarate. Nature 2009;462(7274):739–744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lenders JW, Duh QY, Eisenhofer G, et al. Pheochromocytoma and paraganglioma: an endocrine society clinical practice guideline. The Journal of clinical endocrinology and metabolism 2014;99(6):1915–1942. [DOI] [PubMed] [Google Scholar]
- 21.Toledo RA, Burnichon N, Cascon A, et al. Consensus Statement on next-generation-sequencing-based diagnostic testing of hereditary phaeochromocytomas and paragangliomas. Nature reviews Endocrinology 2017;13(4):233–247. [DOI] [PubMed] [Google Scholar]
- 22.Curras-Freixes M, Pineiro-Yanez E, Montero-Conde C, et al. PheoSeq: A Targeted Next-Generation Sequencing Assay for Pheochromocytoma and Paraganglioma Diagnostics. The Journal of molecular diagnostics : JMD 2017;19(4):575–588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gieldon L, Masjkur JR, Richter S, et al. Next generation panel sequencing identifies NF1 germline mutations in three patients with pheochromocytoma but no clinical diagnosis of Neurofibromatosis Type 1. European journal of endocrinology / European Federation of Endocrine Societies 2017. [DOI] [PubMed] [Google Scholar]
- 24.Rehm HL, Bale SJ, Bayrak-Toydemir P, et al. ACMG clinical laboratory standards for next-generation sequencing. Genetics in medicine : official journal of the American College of Medical Genetics 2013;15(9):733–747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Toledo RA, Hatakana R, Lourenco DM, Jr., et al. Comprehensive assessment of the disputed RET Y791F variant shows no association with medullary thyroid carcinoma susceptibility. Endocrine-related cancer 2015;22(1):65–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Richards S, Aziz N, Bale S, et al. Standards and guidelines for the interpretation of sequence variants: a joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genetics in medicine : official journal of the American College of Medical Genetics 2015;17(5):405–424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Flynn A, Dwight T, Harris J, et al. Pheo-Type: A Diagnostic Gene-expression Assay for the Classification of Pheochromocytoma and Paraganglioma. The Journal of clinical endocrinology and metabolism 2016;101(3):1034–1043. [DOI] [PubMed] [Google Scholar]
- 28.Imperiale A, Moussallieh FM, Roche P, et al. Metabolome profiling by HRMAS NMR spectroscopy of pheochromocytomas and paragangliomas detects SDH deficiency: clinical and pathophysiological implications. Neoplasia 2015;17(1):55–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Struys EA, Jansen EE, Verhoeven NM, Jakobs C. Measurement of urinary D- and L-2-hydroxyglutarate enantiomers by stable-isotope-dilution liquid chromatography-tandem mass spectrometry after derivatization with diacetyl-L-tartaric anhydride. Clinical chemistry 2004;50(8):1391–1395. [DOI] [PubMed] [Google Scholar]
- 30.Rump A, Benet-Pages A, Schubert S, et al. Identification and Functional Testing of ERCC2 Mutations in a Multi-national Cohort of Patients with Familial Breast- and Ovarian Cancer. PLoS genetics 2016;12(8):e1006248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Eisenhofer G, Huynh TT, Pacak K, et al. Distinct gene expression profiles in norepinephrine- and epinephrine-producing hereditary and sporadic pheochromocytomas: activation of hypoxia-driven angiogenic pathways in von Hippel-Lindau syndrome. Endocrine-related cancer 2004;11(4):897–911. [DOI] [PubMed] [Google Scholar]
- 32.Allegri G, Fernandes MJ, Scalco FB, et al. Fumaric aciduria: an overview and the first Brazilian case report. Journal of inherited metabolic disease 2010;33(4):411–419. [DOI] [PubMed] [Google Scholar]
- 33.Richter S, Klink B, Nacke B, et al. Epigenetic Mutation of the Succinate Dehydrogenase C Promoter in a Patient With Two Paragangliomas. The Journal of clinical endocrinology and metabolism 2016;101(2):359–363. [DOI] [PubMed] [Google Scholar]
- 34.Cascon A, Ruiz-Llorente S, Cebrian A, et al. G12S and H50R variations are polymorphisms in the SDHD gene. Genes, chromosomes & cancer 2003;37(2):220–221. [DOI] [PubMed] [Google Scholar]
- 35.van Nederveen FH, Gaal J, Favier J, et al. An immunohistochemical procedure to detect patients with paraganglioma and phaeochromocytoma with germline SDHB, SDHC, or SDHD gene mutations: a retrospective and prospective analysis. The lancet oncology 2009;10(8):764–771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Peitzsch M, Pelzel D, Glockner S, et al. Simultaneous liquid chromatography tandem mass spectrometric determination of urinary free metanephrines and catecholamines, with comparisons of free and deconjugated metabolites. Clinica chimica acta; international journal of clinical chemistry 2013;418:50–58. [DOI] [PubMed] [Google Scholar]
- 37.Letouze E, Martinelli C, Loriot C, et al. SDH mutations establish a hypermethylator phenotype in paraganglioma. Cancer cell 2013;23(6):739–752. [DOI] [PubMed] [Google Scholar]
- 38.Clark GR, Sciacovelli M, Gaude E, et al. Germline FH mutations presenting with pheochromocytoma. The Journal of clinical endocrinology and metabolism 2014;99(10):E2046–2050. [DOI] [PubMed] [Google Scholar]
- 39.Nambuba JD R; Janssen I; Bullova P; Adams K; Millo C; Bordeau I; Kassai A; Yang C; Kebebew E; Zhuang Z; Pacak K Functional imaging experience in a germline fumarate hydratase mutation-positive patient with pheochromocytoma and paraganglioma. Clinical Case Reports 2016;2(3):e176–181. [Google Scholar]
- 40.Williams MD. Paragangliomas of the Head and Neck: An Overview from Diagnosis to Genetics. Head Neck Pathol 2017;11(3):278–287. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.