

Abstract
Circular RNAs (circRNAs), a novel class of non‐coding RNAs, have emerged as indispensable modulators in human malignancies. Aberrant cellular senescence is a phenotype observed in various cancers. The association of circRNAs with cellular senescence in tumors is yet to determined. Here, we investigated the role of circLARP4 in cellular senescence and cell proliferation in hepatocellular carcinoma (HCC). Downregulated circLARP4 level was observed in HCC tissues and cell lines. Low expression level of circLARP4 independently predicted poor survival outcome. Gain‐of‐function and loss‐of‐function assays demonstrated that circLARP4 suppressed HCC cell proliferation, mediated cell cycle arrest and induced senescence in vitro. Levels of p53 and p21, 2 key regulatory molecules in cellular senescence, were increased in circLARP4‐overexpressed HCC cells and decreased in circLARP4‐silenced HCC cells. In vivo experiments further confirmed the tumor‐suppressing activity of circLARP4. Further mechanistic studies showed that circLARP4 dampened HCC progression by sponging miR‐761, thereby promoting the expression level of RUNX3 and activating the downstream p53/p21 signaling. Our study revealed the role of circLARP4/miR‐761/RUNX3/p53/p21 signaling in HCC progression, providing a potential survival predictor and therapeutic candidate for HCC.
Keywords: circLARP4, hepatocellular carcinoma, microRNA, RUNX3, senescence
1. INTRODUCTION
Hepatocellular carcinoma (HCC) is one of the main causes of cancer‐related deaths worldwide.1 Although great improvements have been achieved in the treatment strategy for HCC, the prognosis of this disease remains dismal.2 Thus, describing the molecular mechanisms of HCC progression is essential for identifying novel therapeutic targets.
Cellular senescence has been characterized as a stress‐induced, durable cell cycle arrest of previously replication‐competent cells that serves to prevent cancers.3 Impaired cellular senescence contributes to enhanced cancer cell proliferation and accelerated cancer progression.3 Senescent cells have several fundamental features, including durable growth arrest, expression of anti‐proliferative molecules and activation of damage‐sensing pathways, leading to expression of many senescence‐associated transcripts.4, 5 The p53/p21 signaling pathway is a well‐established pathway in the scenario of senescence. p53 increases the activation of p21 transcription, thereby facilitating cellular senescence.6 As a putative candidate for cancer treatment, p53/p21 signaling exerts a potent effect on senescence in various digestive system tumors, including HCC,7 colorectal cancer,8 gastric cancer9 and pancreatic cancer.10
Circular RNAs (circRNAs) are a novel class of endogenous non‐coding RNAs. Synthesized by back‐splicing in which the head‐to‐tail splice junctions are joint together to form a circular transcript, circRNAs are conserved, stable, abundant, and expressed in a tissue‐specific and cell‐specific manner.11 Published studies have described various functions of circRNAs, including sponging for mircoRNAs (miRNAs) to regulate gene transcription, interacting with RNA‐binding proteins, and occasionally being translated into proteins.12, 13 Since the discovery of the association of circRNAs with various solid and hematological malignancies, circRNAs have attracted much interest. Many of the reported cancer‐related circRNAs are involved in malignancies of the digestive system.14 circRNAs have also been demonstrated as potential diagnostic biomarkers for digestive system cancers.15 circLARP4 is a recently identified circRNA derived from LARP4 gene locus. circLARP4 has been suggested to be a novel tumor suppressive factor and potential biomarker in gastric cancer.16 However, the association of circLARP4 with HCC has not yet been elucidated.
Increasing evidence has suggested the involvement of miRNAs and long non‐coding RNAs (lncRNAs) in cellular senescence.17 Nevertheless, our knowledge of the role of circRNAs in senescence is extremely limited. Highly expressed in heart samples of aged patients, circ‐Foxo3 is reported to interact with senescence‐associated proteins ID‐1, E2F1, FAK and HIF1α, helping to induce senescence.18 CircPVT1, elevated in dividing cells and reduced in senescent fibroblasts, antagonizes endogenous let‐7 to enable a proliferative phenotype, suppressing cellular senescence.19 Of particular note, the role that circRNAs play in cellular senescence during tumorigenesis remains to be determined.
In the current study, we proposed that circLARP4 was a senescence‐associated circRNA that induced cellular senescence and inhibited the proliferative phenotype in HCC. Mechanistically, circLARP4 sponged miR‐761 to inhibit its actions on RUNX3, and thereby influenced HCC progression. circLARP4 was revealed as an independent prognostic factor for overall survival and recurrence‐free survival in HCC patients.
2. MATERIALS AND METHODS
2.1. Patient samples
A total of 70 HCC tissues and paired peritumor samples were obtained from HCC patients who underwent partial hepatectomy in the First Affiliated Hospital of Nanjing Medical University between January 2014 and February 2015. The study protocol was reviewed and approved by the Ethics Committee of the First Affiliated Hospital of Nanjing Medical University. The study protocol conformed to the provisions of the Declaration of Helsinki. Written informed consent was acquired from all participants.
2.2. Cell lines and cell culture
The normal human liver cell line QSG‐7701 and HCC cell lines Huh7, Hep3B, SMMC7721 and HepG2 were purchased from the Chinese Academy of Sciences Cell Bank (Shanghai, China). MHCC97L and HCCLM3 cell lines were obtained from the Liver Cancer Institute, Zhongshan Hospital, Fudan University (Shanghai, China). All cell lines were cultured in DMEM containing 10% FBS supplemented with 100 U/mL of penicillin and 100 μg/mL of streptomycin (HyClone, Logan, UT, USA). Cells were maintained at 37°C in a humidified atmosphere with 5% CO2.
2.3. RNA preparation and quantitative real‐time PCR
The nuclear and cytoplasmic fractions were isolated using NE‐PER Nuclear and Cytoplasmic Extraction Reagents (Thermo Fisher Scientific, Carlsbad, CA, USA) according to the manufacturer's instructions. TRIzol Reagent (Invitrogen, Thermo Fisher Scientific) was used to extract total RNA from tissues and cells. For RNase treatment, total RNA (2 mg) was incubated for 20 minutes at 37°C with 3 U/μg of RNase R (Epicentre Technologies, Madison, WI, USA). Reverse transcription was performed with PrimeScript RT Master Mix (TaKaRa, Dalian, China), and quantitative real‐time PCR (RT‐qPCR) was conducted using TB Green Premix Ex Taq (TaKaRa) on the Applied Biosystems 7900HT Fast Real‐Time PCR System (Applied Biosystems, Thermo Fisher Scientific). The PCR primers were as follows: circLARP4: forward, 5′‐GGGCATCAGGAGCAAACTTA‐3′; reverse, 5′‐CTGGCGAATTAAAGCCATTC‐3′; RUNX3: forward, 5′‐AGGCAATGACGAGAACTACTCC‐3′; reverse, 5′‐CGAAGGTCGTTGAACCTGG‐3′; GAPDH: 5′‐AACGTGTCAGTGGTGGACCTG‐3′; reverse, 5′‐AGTGGGTGTCGCTGTTGAAGT‐3′; U6: forward 5′‐CTCGCTTCGGCAGCACA‐3′; reverse 5′‐AACGCTTCACGAATTTGCGT‐3′. GAPDH and U6 were used as internal controls. The relative expression was determined using the 2−ΔΔCt method.
2.4. FISH
Oligonucleotide‐modified probe sequence for human circLARP4 and miR‐761 was applied for FISH. Detailed procedures for FISH were reported previously.16
2.5. Lentiviruses, micro RNA mimics and inhibitor
Lentiviruses with circLARP4 overexpression or knockdown and lentivirus with RUNX3 knockdown were constructed by GenePharma (Shanghai, China). The knockdown sequences are shown as below: sh‐circLARP4, GGGCAGGCTCCCTTTCCCAAT; sh‐RUNX3, GGCTAGCAGCATGCGGTATTT. miR‐761 mimics and miR‐761 inhibitor were purchased from RiboBio (Guangzhou, China).
2.6. Cell proliferation assays and cell cycle analysis
Cell Counting Kit‐8 (CCK‐8), 5‐ethynyl‐2′‐deoxyuridine (EdU) incorporation assay and colony formation assay were used to investigate the proliferative capacity of HCC cells, as previously described.20 For cell cycle analysis, cells were fixed in 75% cold ethanol and stored at 4°C overnight. After washing with PBS twice, cells were stained with propidium iodide for 30 minutes. Flow cytometry (FACSCalibur, Becton Dickinson, San Jose, CA, USA) was used to analyze the cell cycle distribution. Experiments were independently repeated 3 times.
2.7. SA‐β‐gal staining
Cellular senescence was determined using SA‐β‐gal staining (Senescence β‐Galactosidase Staining Kit, Beyotime, Shanghai, China) according to the manufacturer's protocol. SA‐β‐gal activity was evaluated by 5‐bromo‐4‐chloro‐3‐indolyl‐β‐D‐galactopyranoside (X‐gal). Randomly selected fields were analyzed by light microscope to quantify the percentage of SA‐β‐gal‐positive cells.
2.8. Western blotting and immunohistochemistry analysis
Western blotting was conducted as previously described.21 Antibodies against RUNX3 (Abcam, Cambridge, MA, USA), p53 (Cell Signaling Technology, Beverly, MA, USA), p21 (Cell Signaling Technology) and GAPDH (Cell Signaling Technology) were used. Expression of PCNA, RUNX3, p5 and p21 was examined using immunohistochemistry as reported previously.21
2.9. RNA immunoprecipitation
We performed an RNA immunoprecipitation (RIP) assay using the Magna RIP RNA‐Binding Protein Immunoprecipitation Kit (Millipore, Billerica, MA, USA) in accordance with the manufacturer's instructions. Antibodies against AGO2 (Abcam) and IgG (Abcam) were used.
2.10. Luciferase reporter assay
Cells (5 × 105 cells per well) were seeded in 24‐well plates. Forty‐eight hours after the cells were co‐transfected with corresponding reporter plasmids as well as miRNA mimics or negative control using Lipofectamine 3000 (Invitrogen), firefly and renilla luciferase activities were measured using the Dual‐Luciferase Reporter Assay System (Promega, Madison, MA, USA) in accordance with the manufacturer's instructions. We then calculated the ratios of luminescence from firefly to renilla luciferase. Experiments were performed in triplicate.
2.11. Animal experiment
Four‐week‐old male nude mice were purchased from the Animal Core Facility of Nanjing Medical University (Nanjing, China). Approximately 5 × 106 cells suspended in serum‐free DMEM were subcutaneously injected into the flanks of the mice. The mice were monitored regularly. After 4 weeks, the mice were killed and xenografts were excised. The tumors were subjected to further RT‐qPCR and immunohistochemistry analysis. All experiments with nude mice were performed strictly in accordance with a protocol approved by the Institutional Animal Care and Use Committee of Nanjing Medical University.
2.12. Statistical analysis
Differences between groups were analyzed using the Wilcoxon signed‐rank test or Student's t test. The associations of circLARP4 expression level with clinicopathological features were examined using Fisher's exact test. Kaplan‐Meier curves were plotted to evaluate the disparity in patient survival. Multivariate survival analysis was performed using Cox proportional hazards regression model. Correlation analysis was conducted using Spearman's rank correlation coefficient. Data are presented as mean ± SEM. Statistical analysis was performed using GraphPad Prism 6 (GraphPad Software, La Jolla, CA, USA) and SPSS 21.0 (IBM Corporation, Armonk, NY, USA). Statistical significance was set at *P < 0.05, **P < 0.01, ***P < 0.001.
3. RESULTS
3.1. circLARP4 is downregulated in hepatocellular carcinoma and associated with patient prognosis
To identify the role of circLARP4 in HCC, we first examined the expression patterns of circLARP4 in 70 HCC tissues and matched peritumor samples. RT‐qPCR results indicated that the circLARP4 expression level was markedly downregulated in HCC tissues (Figure 1A,B). circLARP4 had a lower expression level in 6 HCC cell lines MHCC97L, Huh7, Hep3B, SMMC7721, HepG2 and HCCLM3 compared with that in the normal human liver cell line QSG‐7701 (Figure 1C).
Figure 1.
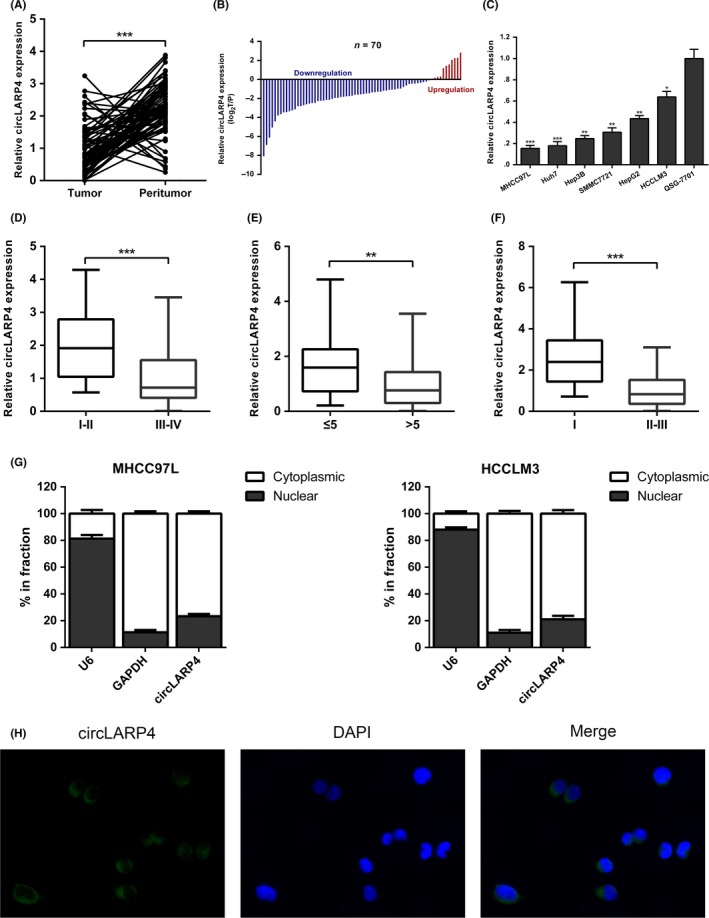
circLARP4 is downregulated in hepatocellular carcinoma (HCC) and is associated with patient prognosis. A, Expression levels of circLARP4 in 70 HCC tissues and matched peritumor samples were determined by quantitative real‐time PCR (RT‐qPCR). B, Relative expression of circLARP4 in comparison with peritumor tissues was analyzed. C, Expression levels of circLARP4 in 6 HCC cell lines and human normal liver cell line QSG‐7701 were examined using RT‐qPCR. D, The expression level of circLARP4 in patients with Edmondson stage I/II and in patients with Edmondson stage III/IV. E, The expression level of circLARP4 in patients with tumor size ≤5 cm and in patients with tumor size >5 cm. F, The expression level of circLARP4 in patients with TNM stage I and in patients with TNM stage II/III. G, RT‐qPCR data indicating the abundance of circLARP4 in either the nucleus or cytoplasm of MHCC97L and HCCLM3 cells. U6 and GAPDH were used as a positive control for the nuclear and cytoplasmic fractions, respectively. H, Fluorescence in situ hybridization was performed to detect the subcellular location of circLARP4 in HCCLM3 cells. The data are shown as the mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001
To assess the clinical significance of circLARP4 in HCC, we divided the 70‐patient cohort into 2 groups according to the median expression of circLARP4. As shown in Table S1, low circLARP4 expression level was associated with unfavorable clinicopathological features, including Edmondson stage, tumor size and tumor‐node‐metastasis (TNM) stage. We further divided the patients into 2 groups based on Edmondson stage, and a lower circLARP4 expression level was detected in patients with Edmondson stage III/IV (Figure 1D). Patients with tumor size >5 cm had a lower circLARP4 level as compared with patients with tumor size ≤5 cm (Figure 1E). Compared to the TNM I stage group, patients with TNM II/III stage had a lower circLARP4 level (Figure 1F). Kaplan‐Meier survival analysis indicated that patients with lower circLARP4 expression had shorter overall survival and worse recurrence‐free survival than patients with higher circLARP4 level (Figure S1A,B). Multivariate Cox proportional analysis further revealed that decreased circLARP4 level was an independent prognostic factor for poor overall survival (Hazard ratio [HR] 3.997, 95% confidence interval [CI] 1.747‐9.142, P = 0.001; Table S2) and recurrence‐free survival (HR 2.347, 95% CI 1.119‐4.923, P = 0.024; Table S3) in HCC.
A previous report indicated that the subcellular location of circLARP4 in gastric cancer was primarily in cytoplasm.16 We performed RT‐qPCR of nuclear and cytoplasmic RNAs and demonstrated that circLARP4 was preferentially localized within the cytoplasm in HCC cells (Figure 1G). FISH analysis confirmed the cytoplasmic location of circLARP4 (Figure 1H). We indicated that circLARP4 was downregulated in HCC, and correlated with unfavorable clinicopathological features and survival outcome.
3.2. circLARP4 suppresses hepatocellular carcinoma proliferation and induces cellular senescence
To evaluate the biological functions of circLARP4, we used lentiviruses to overexpress the level of circLARP4 in MHCC97L and to silence its expression in HCCLM3 cells. The overexpression or knockdown efficiency was validated using RT‐qPCR (Figure 2A,B). CCK‐8 assays showed that circLARP4 upregulation inhibited the proliferation of MHCC97L (Figure 2C), whereas circLARP4 knockdown increased the proliferative capacity of HCCLM3 (Figure 2D). Consistently, EdU assays revealed compromised proliferative ability of circLARP4‐overexpressed MHCC97L cells and enhanced cell proliferation in sh‐circLARP4 HCCLM3 cells (Figure 2E). Colony formation ability of MHCC97L was also suppressed upon circLARP4 upregulation, while sh‐circLARP4 HCCLM3 cells presented an elevated colony formation capacity (Figure 2F). We performed flow cytometry to analyze the cell cycle distribution. As indicated in Figure 2G,H, circLARP4 overexpression caused G1/S cell cycle arrest in MHCC97L, whereas knockdown of circLAPR4 exhibited the opposite effects in HCCLM3 cells.
Figure 2.
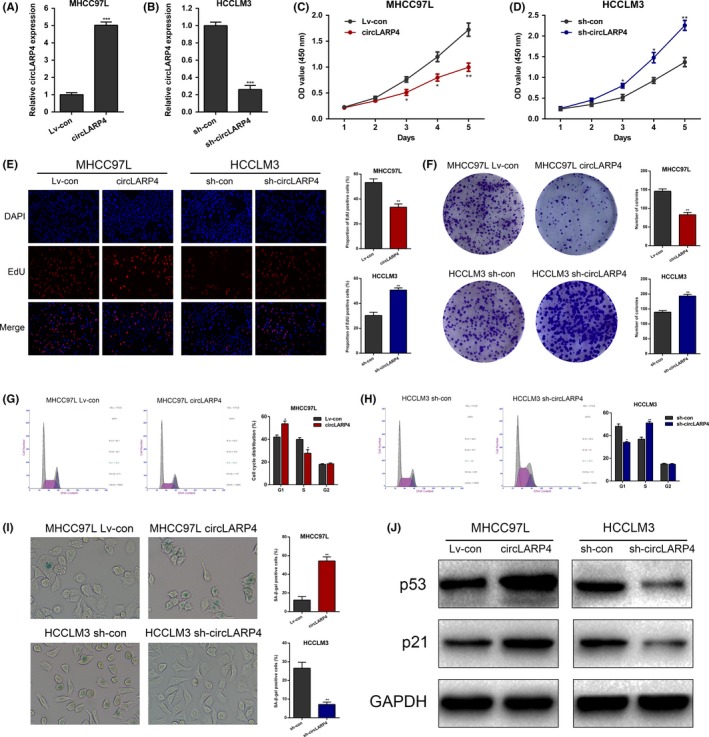
circLARP4 suppresses hepatocellular carcinoma (HCC) proliferation and induces cellular senescence. A, circLARP4 overexpression in MHCC97L cells was validated by quantitative real‐time PCR (RT‐qPCR). B, circLARP4 knockdown in HCCLM3 was verified using RT‐qPCR. C, Cell proliferation was analyzed by CCK‐8 in circLARP4‐overexpressed MHCC97L cells. D, Cell proliferation was analyzed by CCK‐8 in circLARP4‐silenced HCCLM3. E, EdU assays were adopted to assess the effects of circLARP4 overexpression or knockdown on HCC proliferation. F, Evaluation of the effects of circLARP4 on proliferation by colony formation assays. G, Cell cycle distribution was analyzed in MHCC97L cells with or without circLARP4 overexpression using flow cytometry. H, Cell cycle distribution was analyzed in HCCLM3 cells with or without circLARP4 knockdown using flow cytometry. I, Effects of circLARP4 on cellular senescence were examined using SA‐β‐gal staining. J, Western blotting results of the altered levels of p53 and p21 in circLARP4‐overexpressed or circLARP4‐silenced HCC cells. The data are shown as the mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001
Considering the close relationship between cell proliferation and cellular senescence, we speculated that circLARP4 may also have an effect on HCC senescence. To verify our hypothesis, we performed SA‐β‐gal staining to determine the cellular senescence. Data showed that the percentage of SA‐β‐gal‐positive cells in circLARP4‐overexpressed MHCC97L cells was significantly increased. Conversely, sh‐circLARP4 HCCLM3 cells exhibited a reduced percentage of SA‐β‐gal‐positive cells (Figure 2I). Western blotting showed that the expression levels of p53 and p21, 2 master regulators of cellular senescence, were upregulated in circLARP4‐overexpressed MHCC97L cells and downregulated in sh‐circLARP4 HCCLM3 cells (Figure 2J). Collectively, in vitro experiments indicated that circLARP4 dampened cell proliferation, caused cell cycle arrest, and induced senescence in HCC.
3.3. circLARP4 directly targets miR‐761 in hepatocellular carcinoma progression
Given that circLARP4 is mainly localized in the cytoplasm, in which circRNAs serve as a miRNA sponge to modulate gene expression, we next examined the capacity of circLARP4 to bind to miRNAs. We used starBase v2.0 (http://starbase.sysu.edu.cn/)22 with a filter of very high stringency (number of supporting experiments ≥5) to predict the potential miRNAs that bind to circLARP4. Among them, miR‐761 has previously been reported to be upregulated and play an oncogenic role in HCC progression, and was chosen for further validation.23 Potential binding sites of miR‐761 were found within the circLARP4 sequence (Figure 3A). We then evaluated the level of miR‐761 in 30 HCC tissues and matched peritumor samples, and confirmed its upregulation in liver cancer (Figure 3B). Further analysis revealed a negative correlation between circLARP4 and miR‐761 level (Figure 3C). Next, we measured miR‐761 expression in circLARP4 overexpressed or silenced cells. miR‐761 expression was remarkably decreased in circLARP4‐overexpressed MHCC97L cells, while miR‐761 expression was elevated in sh‐circLARP4 HCCLM3 cells (Figure 3D). FISH analysis in HCC cells showed that circLARP4 was co‐localized with miR‐761 in cytoplasm (Figure 3E). Given that high degree of AGO2 occupancy in circLARP4 has been implicated previously,16 we performed RIP for AGO2 in HCC cells and showed higher expression levels of circLARP4 and miR‐761 in AGO2 pellet compared to those in the control group (Figure 3F). Subsequent luciferase reporter assays demonstrated that miR‐761 was a direct target of circLARP4 (Figure 3G).
Figure 3.
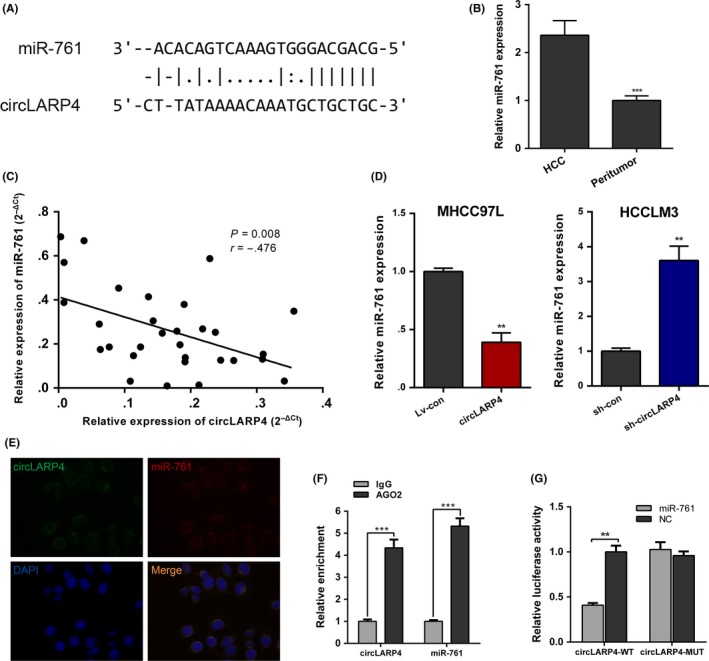
circLARP4 directly targets miR‐761 in hepatocellular carcinoma (HCC). A, Sequence alignment of circLARP4 with the binding sites in circLARP4 is shown. B, miR‐761 levels in HCC tissues and paired peritumor samples were determined using quantitative real‐time PCR (RT‐qPCR). C, The negative relevance between circLAPR4 and miR‐761 was revealed by Spearman's rank correlation analysis. D, miR‐761 expression in circLARP4‐overexpressed or circLARP4‐silenced HCC cells. E, FISH analysis in HCCLM3 cells showing that circLARP4 was co‐localized with miR‐761 in the cytoplasm. F, Anti‐AGO2 RIP was performed followed by quantitative real‐time PCR (RT‐qPCR) to detect circLARP4 and miR‐761. G, The relative luciferase activity was determined after co‐transfection with circLARP4 wide‐type or mutant and miR‐761 mimics or miR‐NC. The data are shown as the mean ± SEM. **P < 0.01, ***P < 0.001
To investigate whether miR‐761 reversed circLARP4‐mediated suppression of HCC progression, we overexpressed miR‐761 in MHCC97L cells using mimics (Figure 4A). CCK‐8 and EdU assays confirmed that miR‐761 mimics restored the inhibitory effects of circLARP4 on HCC growth (Figure 4B,C). Cell cycle analysis showed that G1/S arrest was ameliorated by miR‐761 mimics (Figure 4D). Consistently, miR‐761 overexpression reversed the cellular senescence in MHCC97L (Figure 4E,F). Together, the results supported that the suppressive effect of circLARP4 on HCC progression was mediated by miR‐761.
Figure 4.
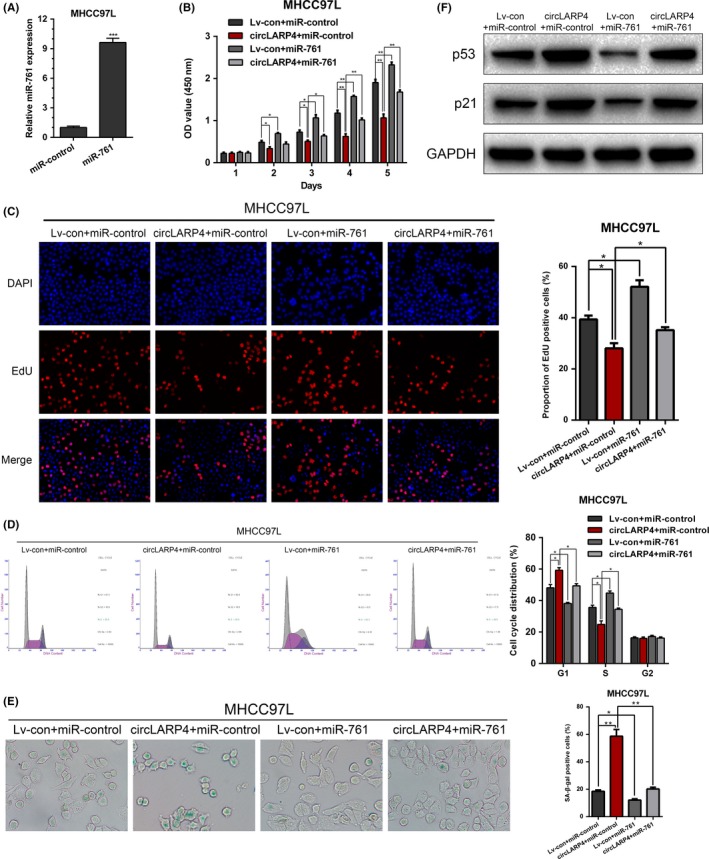
circLARP4 suppresses hepatocellular carcinoma (HCC) progression via miR‐761. A, miR‐761 overexpression was validated using quantitative real‐time PCR (RT‐qPCR) after treating MHCC97L cells with miR‐761 mimics. B, CCK‐8 assays for circLARP4‐overexpressed MHCC97L cells with or without miR‐761 overexpression. C, EdU assays were performed to evaluate the proliferation ability of circLARP4‐overexpressed MHCC97L cells with or without miR‐761 overexpression. D, Flow cytometry was used to analyze the cell cycle distribution in circLARP4‐overexpressed MHCC97L cells with or without miR‐761 overexpression. E, Effects of circLARP4 on cellular senescence was assessed using SA‐β‐gal staining in circLARP4‐overexpressed MHCC97L cells with or without miR‐761 overexpression. F, Western blotting of p53 and p21 expression in circLARP4‐overexpressed MHCC97L cells with or without miR‐761 overexpression. The data are shown as the mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001
3.4. circLARP4 inhibits hepatocellular carcinoma progression via miR‐761/RUNX3 signaling pathway
To further elucidate the underlying mechanisms of the circLARP4/miR‐761 axis in HCC, we used TargetScan, miRWalk, miRDB and DIANA databases to predict the targets of miR‐761 (Figure 5A). Of particular interest to us was RUNX3 among the potential targets predicted by all 4 algorithms. Decreased RUNX3 expression has been previously reported in HCC.24 In addition, RUNX3 suppresses HCC tumorigenesis via p53 and p21.25, 26 Therefore, we speculated that RUNX3 might be a downstream target of miR‐761. We performed RT‐qPCR to verify the downregulated RUNX3 expression in HCC (Figure 5B), and found that miR‐761 expression was negatively correlated with RUNX3 expression (Figure 5C). miR‐761 overexpression led to a decrease in RUNX3 level, whereas miR‐761 knockdown resulted in increased RUNX3 expression (Figure 5D). Luciferase reporter assays revealed that RUNX3 was a direct target of miR‐761 (Figure 5E).
Figure 5.
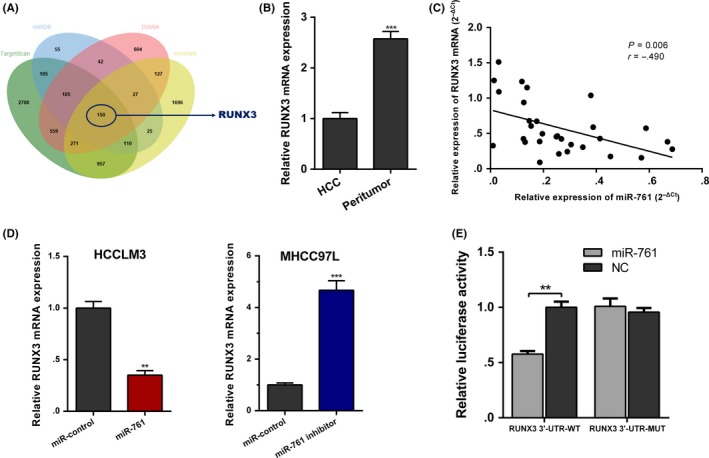
RUNX3 is the downstream target of miR‐761 in hepatocellular carcinoma (HCC). A, TargetScan, miRWalk, miRDB and DIANA databases were used to predict the targets of miR‐761. RUNX3 was predicted by all 4 algorithms. B, RUNX3 levels in HCC tissues and matched peritumor samples were determined using quantitative real‐time PCR (RT‐qPCR). C, The negative correlation between miR‐761 and RUNX3 was indicated by Spearman')s rank correlation analysis. D, RUNX3 expression in miR‐761‐overexpressed or miR‐761‐silenced HCC cells. E, Dual luciferase reporter assay revealed that RUNX3 was a direct target of miR‐761. The data are shown as the mean ± SEM. **P < 0.01, ***P < 0.001
To confirm that RUNX3 was the downstream effector of the circLARP4/miR‐761 axis, we knocked down RUNX3 in circLARP4‐overexpressed MHCC97L cells (Figure 6A,B). Rescue experiments were performed, including CCK‐8, EdU, flow cytometry and SA‐β‐gal staining (Figure 6C‐F). The results showed that the inhibited malignant phenotypes induced by overexpressing circLARP4 were restored by RUNX3 silence. Moreover, the increased levels of p53 and p21 in circLARP4‐overexpressed MHCC97L were abolished after RUNX3 knockdown (Figure 6G). Collectively, we showed that circLARP4 exerted its tumor suppressive functions through miR‐761/RUNX3 signaling pathway.
Figure 6.
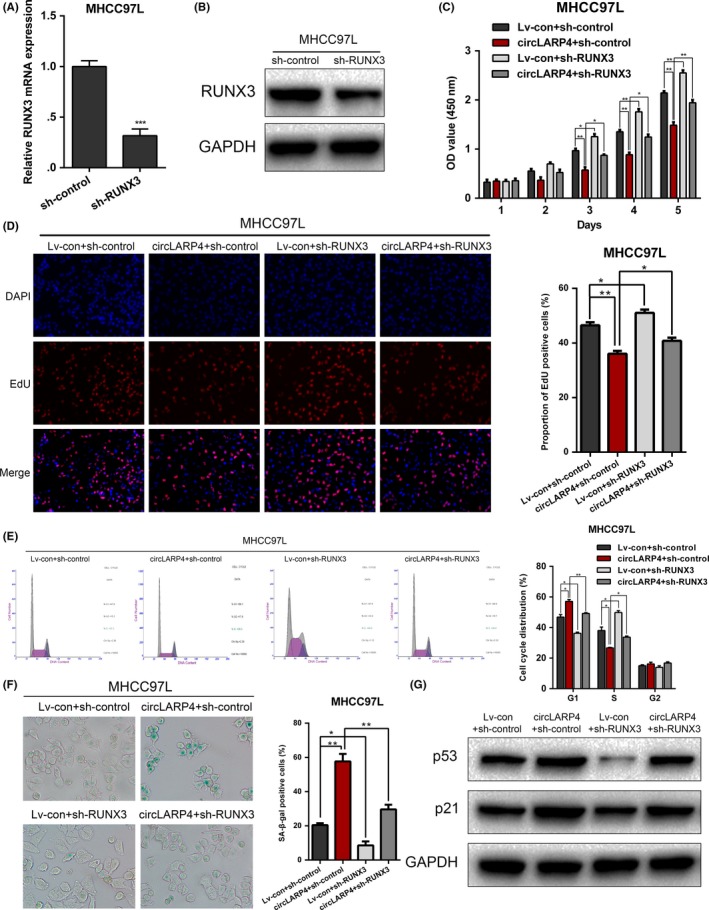
circLARP4 inhibits hepatocellular carcinoma (HCC) progression through the miR‐761/RUNX3 axis. A, RUNX3 knockdown in MHCC97L cells was validated using quantitative real‐time PCR (RT‐qPCR). B, Western blotting results verified RUNX3 knockdown in MHCC97L cells. C, CCK‐8 assays for circLARP4‐overexpressed MHCC97L cells with or without RUNX3 knockdown. D, EdU assays were performed to evaluate the proliferation ability of circLARP4‐overexpressed MHCC97L cells with or without RUNX3 knockdown. E, Flow cytometry was used to analyze cell cycle distribution in circLARP4‐overexpressed MHCC97L cells with or without RUNX3 knockdown. F, Effects of circLARP4 on cellular senescence were assessed using SA‐β‐gal staining in circLARP4‐overexpressed MHCC97L cells with or without RUNX3 knockdown. G, Western blotting of p53 and p21 expression in circLARP4‐overexpressed MHCC97L cells with or without RUNX3 knockdown. The data are shown as the mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001
3.5. circLARP4 inhibits hepatocellular carcinoma progression in vivo
To probe the biological functions of circLARP4 in vivo, we constructed a nude mouse xenograft model. Mice injected with circLARP4‐overexpressed MHCC97L cells developed significantly smaller tumor volumes than those injected with MHCC97L Lv‐con cells. In contrast, mice inoculated with HCCLM3 sh‐circLARP4 cells presented larger tumors than the control group (Figure 7A,B). The growth curve indicated that the xenograft growth was slower in the circLARP4‐overexpressed MHCC97L group, whereas xenografts derived from HCCLM3 sh‐circLARP4 cells grew faster than those from the control group (Figure 7C). Xenografts were subjected to RT‐qPCR assays to confirm that the overexpression or knockdown of circLARP4 was persistent in the xenograft model (Figure 7D). In addition, the miR‐761 level was lower in xenografts of the circLARP4‐overexpressed MHCC97L group, whereas miR‐761 was higher in xenografts derived from the HCCLM3 sh‐circLARP4 group (Figure 7E). Immunohistochemistry analysis indicated a lower expression of PCNA and higher levels of RUNX3, p53 and p21 in the circLARP4‐overexpressed MHCC97L group as compared with those in the control group. Conversely, xenografts originating from HCCLM3 sh‐circLARP4 cells presented increased PCNA expression and decreased levels of RUNX3, p53 and p21 (Figure 7F). In sum, circLARP4 hindered HCC growth and promoted p53 and p21 in vivo.
Figure 7.
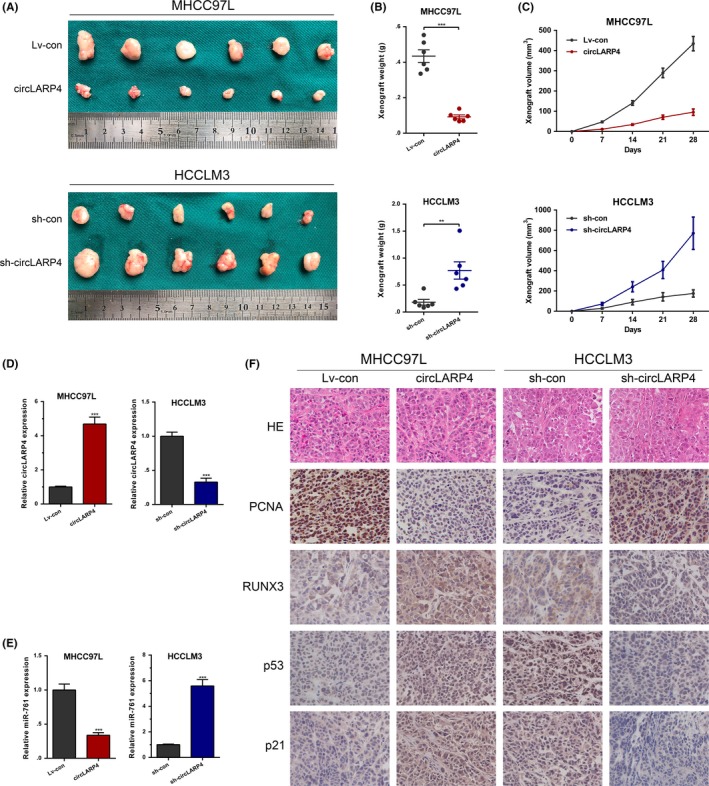
circLARP4 inhibits hepatocellular carcinoma (HCC) progression in vivo. A, circLARP4‐overexpressed or circLARP4‐silenced HCC cells were injected into nude mice. After 4 wk, the mice were killed to harvest the xenografts. B, Xenograft weight was measured in the circLARP4 overexpression group, the sh‐circLARP4 group, and their controls. C, Tumor growth curve was plotted in the circLARP4 overexpression group, the sh‐circLARP4 group, and their controls. D, Quantitative real‐time PCR (RT‐qPCR) analysis showed the consistent overexpression or knockdown of circLARP4 in xenograft. E, RT‐qPCR analysis of the level of miR‐761 in xenografts. F, HE, PCNA, RUNX3, p53 and p21 immunostaining of xenografts. The data are presented as the mean ± SEM. **P < 0.01, ***P < 0.001
4. DISCUSSION
To the best of our knowledge, the present study is the first that correlates circRNAs with cellular senescence in human malignancies. Herein, we demonstrated that circLARP4 was downregulated in HCC. circLARP4 could promote cellular senescence and induce cell cycle arrest in HCC. Mechanistically, circLARP4 acted as a competing endogenous RNA through harboring miR‐761 to abolish the inhibitory effect of miR‐761 on the target gene RUNX3. RUNX3 subsequently activated the p53/p21 signaling and enhanced the downstream senescence phenotype in liver cancer progression. Hence, our results revealed the crucial roles that circLARP4 played in HCC.
circRNAs have emerged as novel players that are involved in various biological and pathological processes.27 Recent studies have demonstrated that many circRNAs are involved in tumorigenesis and progression.28, 29 The relationship between circRNAs and cellular senescence in human malignancy, however, has never been reported before. We, for the first time, associated circLARP4 dysregulation with senescence in HCC. Derived from exon 9 and 10 regions within the LARP4 gene locus, circLARP4 is a recently identified circRNA in gastric cancer. circLARP4 suppresses the proliferation and invasion of gastric cancer by sponging miR‐424 and upregulating the Hippo signaling.16 Our data revealed that circLARP4 overexpression enhanced senescence, attenuated HCC proliferation and induced G1/S arrest. Conversely, silencing circLARP4 promoted HCC progression by regulating cellular senescence, cell proliferation and cell cycle distribution. Results from xenograft models confirmed the tumor suppressive effects of circLARP4 in HCC.
Sponging for miRNAs is one of the classic mechanisms for circRNAs to exert their functional effects, especially for those that are located within cytoplasm. For instance, circular RNA cSMARCA5 inhibits HCC growth and migration by sponging miR‐181‐5p and miR‐17‐3p and promoting downstream TIM3 expression.30 circMTO1 suppresses HCC proliferation and invasion by sponging miR‐9 and upregulating the downstream p21.31 circHIPK3 facilitates HCC progression through serving as a miR‐124 sponge and subsequently modulating AQP3 in HCC.32 Given the cytoplasmic location of circLARP4, we speculated that circLARP4 might function as a miRNA sponge to dampen HCC malignant phenotypes. To support this hypothesis, we conducted bioinformatics analysis to predict the potential miRNAs downstream of circLARP4. As suggested by starBase 2.0 results, a complementary binding sequence between circLARP4 and miR‐761 was indicated. RIP and luciferase reporter assay supported that miR‐761 was a direct target of circLARP4. miR‐761 knockdown could counteract the tumor‐inhibiting activity of circLARP4. Previous studies showed that miR‐761 is increased in HCC and targets Mitofusin‐2 to regulate tumor growth and metastasis.23 miR‐761 has also been verified as the sponging miRNA of has_circ_0007534 in glioma.33 Moreover, elevated miR‐761 expression has been observed in triple‐negative breast cancer,34 synovial sarcoma35 and non‐small cell lung cancer.36 We identified that miR‐761 was the downstream target of circLARP4, and was associated with circLARP4‐mediated cellular senescence in HCC.
To gain clues of the target gene of the circLARP4/miR‐761 axis in HCC, we combined 4 different bioinformatics algorithms and RUNX3 was predicted as the potential downstream target. Further experiments revealed that RUNX3 was the downstream effector of the circLARP4/miR‐761 axis and mediated cellular senescence in HCC. RUNX3 has been well established as a tumor suppressor.37, 38, 39 In HCC, aberrant CpG methylation could also contribute to RUNX3 downregulation.40, 41 RUNX3 represses epithelial‐mesenchymal transition, and suppresses HCC metastasis.42 RUNX3 leads to dysregulated cell cycle distribution, apoptosis and angiogenesis.43, 44 RUNX3 induces ARF to stabilize p53, thereby exerting a persistent anti‐tumor effect.38 p21 has also been reported as a direct downstream target of RUNX3.25 Given that p53 and p21 are 2 crucial modulators contributing to cellular senescence, we examined whether the expression of p53 and p21 could be altered by the circLARP4/miR‐761/RUNX3 signaling. The results showed that circLARP4 affected cellular senescence via the p53/p21 signaling through regulating the miR‐761/RUNX3 axis.
In summary, we identified that circLARP4 induced cellular senescence, thereby inhibiting HCC progression. Low circLARP4 expression was positively correlated with unfavorable clinicopathological features and was an independent predictor for HCC patient survival. Mechanistic studies showed that circLARP4 sponged miR‐761 to regulate RUNX3, thus affecting the expression levels of p53 and p21. The regulatory axis of circLARP4/miR‐761/RUNX3/p53/p21 contributed to a better understanding of senescence‐associated circRNAs in HCC progression. Targeting circLARP4 is be a promising therapeutic candidate for HCC treatment.
CONFLICTS OF INTEREST
The authors have no conflict of interest.
Supporting information
Chen Z, Zuo X, Pu L, et al. circLARP4 induces cellular senescence through regulating miR‐761/RUNX3/p53/p21 signaling in hepatocellular carcinoma. Cancer Sci. 2019;110:568–581. 10.1111/cas.13901
Zhiqiang Chen, Xueliang Zuo and Liyong Pu equally contributed to this work.
Contributor Information
Jindao Wu, Email: wujindao@njmu.edu.cn.
Xuehao Wang, Email: wangxh@njmu.edu.cn.
REFERENCES
- 1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018;68:7‐30. [DOI] [PubMed] [Google Scholar]
- 2. Parikh ND, Marshall VD, Singal AG, et al. Survival and cost‐effectiveness of sorafenib therapy in advanced hepatocellular carcinoma: an analysis of the SEER‐Medicare database. Hepatology. 2017;65:122‐133. [DOI] [PubMed] [Google Scholar]
- 3. Frey N, Venturelli S, Zender L, Bitzer M. Cellular senescence in gastrointestinal diseases: from pathogenesis to therapeutics. Nat Rev Gastroenterol Hepatol. 2018;15:81‐95. [DOI] [PubMed] [Google Scholar]
- 4. He S, Sharpless NE. Senescence in health and disease. Cell. 2017;169:1000‐1011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Martinez‐Zamudio RI, Robinson L, Roux PF, Bischof O. SnapShot: cellular senescence pathways. Cell. 2017;170:816‐e1. [DOI] [PubMed] [Google Scholar]
- 6. Georgakilas A, Martin O, Bonner W. p21: a two‐faced genome guardian. Trends Mol Med. 2017;23:310‐319. [DOI] [PubMed] [Google Scholar]
- 7. Dhar D, Antonucci L, Nakagawa H, et al. Liver cancer initiation requires p53 inhibition by CD44‐enhanced growth factor signaling. Cancer Cell. 2018;33:1061‐1077.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Wang G, Fu Y, Hu F, et al. Loss of BRG1 induces CRC cell senescence by regulating p53/p21 pathway. Cell Death Dis. 2017;8:e2607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Chen P, Luo X, Che Z, et al. Targeting of the C‐Jun/BCL‐XL/P21 axis accelerates the switch from senescence to apoptosis upon ROC1 knockdown in gastric cancer cells. Cell Physiol Biochem. 2018;48:1123‐1138. [DOI] [PubMed] [Google Scholar]
- 10. Yang S, Hwang S, Kim M, Seo S, Lee J, Jeong S. Mitochondrial glutamine metabolism via GOT2 supports pancreatic cancer growth through senescence inhibition. Cell Death Dis. 2018;9:55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Chen L. The biogenesis and emerging roles of circular RNAs. Nat Rev Mol Cell Biol. 2016;17:205‐211. [DOI] [PubMed] [Google Scholar]
- 12. Han B, Chao J, Yao H. Circular RNA and its mechanisms in disease: from the bench to the clinic. Pharmacol Ther. 2018;187:31‐44. [DOI] [PubMed] [Google Scholar]
- 13. Zhang M, Xin Y. Circular RNAs: a new frontier for cancer diagnosis and therapy. J Hematol Oncol. 2018;11:21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Dragomir M, Calin GA. Circular RNAs in cancer ‐ lessons learned from microRNAs. Front Oncol. 2018;8:179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Chen Z, Zhang L, Han G, et al. A meta‐analysis of the diagnostic accuracy of circular RNAs in digestive system malignancy. Cell Physiol Biochem. 2018;45:962‐972. [DOI] [PubMed] [Google Scholar]
- 16. Zhang J, Liu H, Hou L, et al. Circular RNA_LARP4 inhibits cell proliferation and invasion of gastric cancer by sponging miR‐424‐5p and regulating LATS1 expression. Mol Cancer. 2017;16:151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Panda AC, Abdelmohsen K, Gorospe M. SASP regulation by noncoding RNA. Mech Ageing Dev. 2017;168:37‐43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Du WW, Yang W, Chen Y, et al. Foxo3 circular RNA promotes cardiac senescence by modulating multiple factors associated with stress and senescence responses. Eur Heart J. 2017;38:1402‐1412. [DOI] [PubMed] [Google Scholar]
- 19. Panda AC, Grammatikakis I, Kim KM, et al. Identification of senescence‐associated circular RNAs (SAC‐RNAs) reveals senescence suppressor CircPVT1. Nucleic Acids Res. 2017;45:4021‐4035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Chen Z, Zuo X, Zhang Y, et al. MiR‐3662 suppresses hepatocellular carcinoma growth through inhibition of HIF‐1alpha‐mediated Warburg effect. Cell Death Dis. 2018;9:549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Chen Z, Gao W, Pu L, et al. PRDM8 exhibits antitumor activities toward hepatocellular carcinoma by targeting NAP1L1. Hepatology. 2018;45:962‐972. [DOI] [PubMed] [Google Scholar]
- 22. Li J, Liu S, Zhou H, Qu L, Yang J. starBase v2.0: decoding miRNA‐ceRNA, miRNA‐ncRNA and protein‐RNA interaction networks from large‐scale CLIP‐Seq data. Nucleic Acids Res. 2014;42:D92‐D97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Zhou X, Zhang L, Zheng B, et al. MicroRNA‐761 is upregulated in hepatocellular carcinoma and regulates tumorigenesis by targeting Mitofusin‐2. Cancer Sci. 2016;107:424‐432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Tanaka S, Shiraha H, Nakanishi Y, et al. Runt‐related transcription factor 3 reverses epithelial‐mesenchymal transition in hepatocellular carcinoma. Int J Cancer. 2012;131:2537‐2546. [DOI] [PubMed] [Google Scholar]
- 25. Chi X, Lee J, Lee Y, Park I, Ito Y, Bae S. Runx3 plays a critical role in restriction‐point and defense against cellular transformation. Oncogene. 2017;36:6884‐6894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Lee J, van Wijnen A, Bae S. RUNX3 and p53: how two tumor suppressors cooperate against oncogenic Ras? Adv Exp Med Biol. 2017;962:321‐332. [DOI] [PubMed] [Google Scholar]
- 27. Beermann J, Piccoli M, Viereck J, Thum T. Non‐coding RNAs in development and disease: background, mechanisms, and therapeutic approaches. Physiol Rev. 2016;96:1297‐1325. [DOI] [PubMed] [Google Scholar]
- 28. Zheng J, Liu X, Xue Y, et al. TTBK2 circular RNA promotes glioma malignancy by regulating miR‐217/HNF1beta/Derlin‐1 pathway. J Hematol Oncol. 2017;10:52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Joseph NA, Chiou SH, Lung Z, et al. The role of HGF‐MET pathway and CCDC66 cirRNA expression in EGFR resistance and epithelial‐to‐mesenchymal transition of lung adenocarcinoma cells. J Hematol Oncol. 2018;11:74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Yu J, Xu Q, Wang Z, et al. Circular RNA cSMARCA5 inhibits growth and metastasis in hepatocellular carcinoma. J Hepatol. 2018;68:1214‐1227. [DOI] [PubMed] [Google Scholar]
- 31. Han D, Li J, Wang H, et al. Circular RNA circMTO1 acts as the sponge of microRNA‐9 to suppress hepatocellular carcinoma progression. Hepatology. 2017;66:1151‐1164. [DOI] [PubMed] [Google Scholar]
- 32. Chen G, Shi Y, Liu M, Sun J. circHIPK3 regulates cell proliferation and migration by sponging miR‐124 and regulating AQP3 expression in hepatocellular carcinoma. Cell Death Dis. 2018;9:175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Li GF, Li L, Yao ZQ, Zhuang SJ. Hsa_circ_0007534/miR‐761/ZIC5 regulatory loop modulates the proliferation and migration of glioma cells. Biochem Biophys Res Commun. 2018;499:765‐771. [DOI] [PubMed] [Google Scholar]
- 34. Guo GC, Wang JX, Han ML, Zhang LP, Li L. microRNA‐761 induces aggressive phenotypes in triple‐negative breast cancer cells by repressing TRIM29 expression. Cell Oncol (Dordr). 2017;40:157‐166. [DOI] [PubMed] [Google Scholar]
- 35. Shiozawa K, Shuting J, Yoshioka Y, Ochiya T, Kondo T. Extracellular vesicle‐encapsulated microRNA‐761 enhances pazopanib resistance in synovial sarcoma. Biochem Biophys Res Commun. 2018;495:1322‐1327. [DOI] [PubMed] [Google Scholar]
- 36. Yan A, Yang C, Chen Z, Li C, Cai L. MiR‐761 promotes progression and metastasis of non‐small cell lung cancer by targeting ING4 and TIMP2. Cell Physiol Biochem. 2015;37:55‐66. [DOI] [PubMed] [Google Scholar]
- 37. Whittle M, Izeradjene K, Rani P, et al. RUNX3 controls a metastatic switch in pancreatic ductal adenocarcinoma. Cell. 2015;161:1345‐1360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Lee Y, Bae S. How do K‐RAS‐activated cells evade cellular defense mechanisms? Oncogene. 2016;35:827‐832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Lee Y, Lee J, Jang J, et al. Runx3 inactivation is a crucial early event in the development of lung adenocarcinoma. Cancer Cell. 2013;24:603‐616. [DOI] [PubMed] [Google Scholar]
- 40. Khan F, Ali I, Afridi U, Ishtiaq M, Mehmood R. Epigenetic mechanisms regulating the development of hepatocellular carcinoma and their promise for therapeutics. Hepatol Int. 2017;11:45‐53. [DOI] [PubMed] [Google Scholar]
- 41. Nishida N, Kudo M, Nagasaka T, Ikai I, Goel A. Characteristic patterns of altered DNA methylation predict emergence of human hepatocellular carcinoma. Hepatology. 2012;56:994‐1003. [DOI] [PubMed] [Google Scholar]
- 42. Gou Y, Zhai F, Zhang L, Cui L. RUNX3 regulates hepatocellular carcinoma cell metastasis via targeting miR‐186/E‐cadherin/EMT pathway. Oncotarget. 2017;8:61475‐61486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Nakanishi Y, Shiraha H, Nishina S, et al. Loss of runt‐related transcription factor 3 expression leads hepatocellular carcinoma cells to escape apoptosis. BMC Cancer. 2011;11:3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Gao J, Chen Y, Wu K, et al. RUNX3 directly interacts with intracellular domain of Notch1 and suppresses Notch signaling in hepatocellular carcinoma cells. Exp Cell Res. 2010;316:149‐157. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials