

Abstract
Predicting bladder cancer progression is important in selecting the optimal treatment for bladder cancer. Since current diagnostic factors regarding progression are lacking, new factors are needed to further stratify the curative potential of bladder cancer. Glycoprotein-130 (GP130), a transmembrane protein, is central to a number of signal transduction pathways involved in tumor aggressiveness, making it an attractive target. We hypothesize that if GP130 is found in an aggressive population of bladder tumors then blocking GP130 expression may inhibit bladder cancer growth. Herein, we quantitatively show using eleven patient samples and four bladder cancer cell lines that GP130 is expressed in the aggressive human bladder tumors and in high-grade bladder cancer cell lines. Moreover, GP130 is significantly correlated with tumor grade, node category, tumor category, and patient outcome. We demonstrated a tumor-specific GP130 effect by blocking GP130 expression in bladder tumor cells, which resulted in decreased cell viability and reduced cell migration. Furthermore, we reduced tumor volume by approximately 70 % compared to controls by down-regulating GP130 expression using chitosan functionalized nanoparticles encapsulating GP130 siRNA in an in vivo bladder cancer xenograft mouse model. Our results indicate that GP130 expression is linked to the aggressiveness of bladder tumors, and blocking GP130 has therapeutic potential in controlling tumor growth.
Keywords: Bladder cancer growth, chitosan-functionalized nanoparticles, tumor-specific GP130 effect, targeted therapeutics, GP130 siRNA, SC144 inhibitor
Introduction
Bladder cancer is the sixth most common cancer in the United States with an estimated 81,490 newly diagnosed cases expected this year, and is a common cancer worldwide with the highest rates in Europe, North America, and Northern Africa (1, 2). Seventy percent of bladder cancers are superficial (3), and among patients who present with superficial disease, recurrence rates range from 67 to 73 %, with disease progression rates ranging from 20 to 30 % (4). Although many patients present with non-muscle-invasive disease, approximately 35 % of patients will present with or develop muscle-invasive disease (5). Major challenges in treating bladder cancer are the high recurrence rates and the need for long-term follow-up consisting of urine cytology and cystoscopy, thus making the cost-per-case for bladder cancer one of the highest among all cancers (6–8). Better diagnostic markers are required to gain insight into the progression of bladder cancer, which ultimately may be used to determine the most effective therapeutic interventions.
Glycoprotein-130 (GP130, CD130 or IL6ST), a transmembrane protein, is located at a central point for a number of oncogenic signaling cascades, including the JAK/STAT, PI3K/AKT and MAPK/ERK pathways (9). These downstream pathways have been shown to be involved in multiple hallmarks of cancer development including invasion, metastasis, resistance to apoptosis and proliferation (9–11), which makes GP130 an attractive therapeutic target.
We hypothesize that if GP130 is found in an aggressive population of bladder tumors then blocking GP130 expression may lead to inhibition of bladder cancer growth. To address this hypothesis, we performed molecular analyses on clinically annotated bladder specimens using our novel GP130 bladder cancer biomarker as well as survival and metastatic biomarkers that have been correlated with advanced bladder cancer including survivin, B-cell lymphoma-extra large (Bcl-xL), and vascular endothelial growth factor receptor-2 (VEGFR2). To functionally test the role of GP130 in human bladder cancer cells, we blocked the expression of GP130 using GP130 siRNA (siGP130), and investigated cell migration, viability, and growth. We also used a GP130 inhibitor, SC144, to confirm cell migration, viability and growth upon the downregulation of GP130 expression. Then, we implemented our previously described chitosan (CH) nanoparticle (NP) delivery system (12) to decrease GP130 protein expression in an in vivo xenograft bladder cancer mouse model. For the first time, we show that blocking GP130 using CH-functionalized nanoparticles encapsulating siRNA GP130 has therapeutic potential in controlling tumor growth.
Materials and Methods
Patient tissue specimens
Eleven de-identified specimens were collected from nine patients by transurethral resection of bladder tumor (TURBT) or cystectomy. The specimens were comprised of nine urothelial cell carcinomas and two normal control bladder tissues. All specimens were collected between 2014–2016. From each patient, we recorded tumor histology type, node category, tumor category, tumor grade, sex, treatment type, tumor relapse and patient survival (Table I). Bladder tumors were defined as aggressive if they met the following criteria: tumor was of high grade, patient had poor survival outcome, and tumor category was T3 or T4. All patients were provided informed consent and offered enrollment into a biospecimen repository approved by the Yale University Institutional Review Board.
Table I:
Clinical and Histopathological data
| Specimen | Histology Types | Node Category | Tumor Category | Tumor Grade | Sex | Treatment Type a | Relapse Y/N | Survival Time (months) |
|---|---|---|---|---|---|---|---|---|
| 1 | Urothelial / TCC | N0 | T2a | High | M | 2 | N | 19 |
| 2 | Urothelial / TCC | N2 | T4 | High | M | 2 | Y | 7 |
| 3 | Urothelial / TCC | N1 | T3b | High | M | 2 | Y | 11 |
| 4 | Normal (Match 3) | - | - | - | M | 2 | Y | 11 |
| 5 | Urothelial / TCC | Nx | Ta | Low | F | 3 | N | 8 |
| 6 | Urothelial / TCC | Nx | Ta | Low | M | 3 | N | 24 |
| 7 | Urothelial / TCC | N2 | T4 | High | F | 1 | Y | 19 |
| 8 | Normal (Match 7) | - | - | - | F | 1 | Y | 19 |
| 9 | Urothelial / TCC | Nx | Ta | Low | M | 3 | Y | 21 |
| 10 | Urothelial / TCC | NA | NA | Low | F | 3 | Y | 27 |
| 11 | Urothelial / TCC | N0 | T3a | N/A | F | 2 | N | 20 |
Treatment type (1: neoadjuvant chemotherapy plus a cystectomy; 2: cystectomy only; 3: TURBT only).
Cell culture
RT-4, T24, UM-UC-3 and TCCsup bladder cancer cells were obtained directly from the American Type Culture Collection from 2007 to 2011. T24 and RT-4 cells were maintained in McCoy’s medium, whereas UM-UC-3 and TCCsup cells were maintained in Eagle’s Minimum Essential medium. All cells were maintained at 37 °C in 5 % CO2 atmosphere, and supplemented with 10 % fetal bovine serum and 1 % glutamine. At the start of each experiment, cells were used in the exponential growth phase. Also, a fresh vial of cells was thawed after ten passages. Six months prior to initiating experiments, cells were reauthenticated for quality control using short tandem repeat DNA profiles (The Yale University DNA Analysis Facility, New Haven, CT). In addition, cells were routinely screened for mycoplasma using MycoAlert™ (Lonza Biologics Inc., Portsmouth, NH) every 6 months in our laboratory.
siRNA treatment
Bladder cancer cells were transfected with: siGP130 to a final concentration of 100 nmol/L with Lipofectamine RNAiMAX (Invitrogen); scrambled siRNA (siSC) to a final concentration of 100 nmol/L with Lipofectamine RNAiMAX; Lipofectamine RNAiMAX alone (vehicle); or were left untreated. The GP130 target sequence was: 5’-CAGUAAAUCUCACAAAUGA-3’ (sense), with the scramble control: 5’-AACGUACGCGGAAUACUUCGA-3’ (Dharmacon). After 72 h, the cells were assessed for cell viability, cell migration, cell growth or GP130 protein expression.
Cell viability of bladder cancer cells
Cell viability assays were conducted according to the manufacturer’s instruction (Clontech Laboratories, Mountain View, CA). In brief, siGP130, siSC, or vehicle were administered to 5 × 103 human bladder cancer cells per well in a 96-well plate or the cells were left untreated, and cell viability was measured using the tetrazolium (WST-1) reagent. The percent inhibition of cell viability relative to siSC control was measured using Origin Lab Data Analysis Software.
Migration assay
Bladder cancer cells were treated with siGP130, siSC, or vehicle and plated at 6 × 104 cells per well of a 24-well plate and grown for 24 h to confluency. A scratch was created using a P200 tip in a confluent monolayer of cells with reduced serum. The distance the cells migrated to cover the scratch was calculated over 24 h.
Cell growth assay and crystal violet staining
Bladder cancer cells were treated with siGP130, siSC, or vehicle and plated at 1.5 × 104 cells per well (6-well plate). After 48 h, the cells were fixed with ice cold methanol for 10 min and stained with 0.5 % crystal violet dye for 10 min (13). The bladder cancer cells were washed 3 times with deionized water for 5 min before being dried and photographed. Cell growth (or survival) rate (%) = (Cell number of treatment group) / (Cell number of control group) × 100 %.
Western blotting
Protein expression including phosphorylation was determined by western blot. In brief, protein lysates were prepared from human bladder cancer cells and from snap-frozen human bladder tumors. Radioimmunoprecipitation assay buffer (Cell Signaling Technology, Danvers, MA) supplemented with a cOmplete™, mini, EDTA-free protease inhibitor cocktail (Roche Applied Science), 1 mM phenylmethylsulfonyl fluoride, 2 μg/ml aprotinin (protease inhibitor), and 1 mM sodium fluoride was used to lyse the tissues and cells prior to quantification with the Bradford assay (14). Samples were separated on 4–15 % Mini-PROTEAN gels (Bio-Rad lab. Inc. Hercules, CA) and proteins were transferred to polyvinylidene fluoride membranes. Membranes were blocked with 5 % non-fat dry milk in Tris-buffered saline (TBS) with 0.1 % Tween 20 (TBST) for 1 h at RT, and incubated with primary antibodies in 5 % non-fat dry milk in TBST at 4 °C overnight. The membranes were subsequently washed with TBST and incubated with HRP conjugated donkey anti-rabbit or donkey anti-mouse IgG secondary antibodies (Cell Signaling Technology). A chemiluminescence system (Thermo Scientific, Rockford IL) was used to detect protein signal. Samples were normalized based on GAPDH quantification, and band density was determined using ImageJ software (NIH).
NP fabrication and characterization with siGP130
The double emulsion solvent evaporation technique was used to synthesize NPs containing siRNA (15). In brief, poly(lactic-co-glycolic acid) (PLGA) was dissolved overnight in methylene chloride (DCM), prior to siRNA and spermidine complex formation using an 8:1 molar ratio of the polyamine nitrogen to the nucleotide phosphate. One hundred nanomoles of siGP130 per 100 mg polymer in Tris-EDTA (10 mM Tris-HCl, 1 mM EDTA) buffer was added dropwise to the PLGA solution while vortexing. This solution was sonicated and subsequently added to a 2.5 % polyvinyl alcohol (PVA), 5 mg/mL avidin palmitate solution for the second emulsion. NPs were hardened during solvent evaporation in 0.3 % PVA for 3 h. To generate the CH surface modification, the hardened NPs were reacted with 10 times molar excess biotin-CH2.5 to avidin (NP-CH2.5) in PBS for 30 min. To synthesize unmodified NPs (NP-Unmod), the second emulsion contained only 2.5 % PVA, and NPs were incubated post-hardening in PBS without ligand for 30 min. All NPs were washed twice in deionized water to remove residual solvent, centrifuged at 4 °C, lyophilized, and stored at −20 °C. As previously reported (12, 15), 3–5 mg of siRNA NPs were dissolved in 0.5 mL of DCM for 30 min, and siGP130 was extracted twice into Tris-EDTA buffer. The quantity of extracted double-stranded siRNA was determined using the QuantIT™ PicoGreen™ assay (Invitrogen, Carlsbad, CA). Fluorescence was compared to a known siRNA standard. Encapsulation efficiency was determined by comparing the amount of siRNA loaded into the NPs with theoretical loading (1 nmol siRNA/mg polymer). For the GP130 NPs (NP-siGP130-CH2.5), the loading was 514 pmol siRNA/mg NP. Scanning electron microscopy images were analyzed using ImageJ software to determine NP morphology, diameter, and size distribution (Table II)(12).
Table II:
NP Characterization
| NP-Unmod b | NP-CH2.5 c | |
|---|---|---|
| Number of NPs Measured: | 817 | 1135 |
| Average Diameter (nm): | 166.0 | 137.8 |
| Standard Deviation: | 64.6 | 51.3 |
| Smallest Diameter (nm): | 62.3 | 62.4 |
| Largest Diameter (nm): | 483.3 | 399.0 |
NP-Unmod, unmodified nanoparticles.
NP-CH2.5, chitosan functionalized nanoparticles.
Bladder cancer mouse model
Foxn1 nu/nu mice were subcutaneously injected in the flank with 5 × 106 UM-UC-3 bladder cancer cells. Tumor volumes were measured two times per week using calipers and estimated using the formula (tumor length x tumor width2) × π/6. When tumor volumes reached approximately 100 ± 25 mm3 (referred to as day 0), the mice were randomly divided into three groups and intratumorally injected with: PBS (untreated), 1 mg/100 μl NP-Bk-CH2.5 (chitosan functionalized blank NPs) diluted in PBS, or 1 mg/100 μl NP-siGP130-CH2.5 (514 pmol siGP130; chitosan functionalized NPs encapsulating siGP130) diluted in PBS. As we have previously reported, chitosan functionalization was used to facilitate the delivery of the siRNA (12). Mice were treated on day 0, 4, 7, and 11. After 14 days, the xenograft mice were sacrificed and mouse tumors were split; half was fixed in 10 % neutral buffer formalin (VWR international) for immunohistochemistry, and half was snap-frozen and stored at −80 °C for western blot. All animal studies were approved by the Institutional Animal Care and Use Committee of Yale University.
Immunohistochemistry
Immunohistochemistry was performed as previously described (16). In brief, the paraffin-embedded mouse tumors were deparaffinized in xylene, rehydrated with graded ethanol, and post-fixed in 4 % paraformaldehyde before undergoing antigen retrieval. The slides were incubated with a keratin 20 (C9Z1Z) antibody (Cell Signaling Technology) and developed using a vector red / DAB substrate kit, which previously had been shown to significantly correlate with tumor grade and stage (17).
Statistics
Data are presented as mean ± standard deviation (SD) from 3–6 independent experiments unless otherwise noted. Statistical significance was determined by student t-test (p < 0.05) to assess significant differences. Results are presented as mean +/− SD in which * represents p < 0.05, ** represents p < 0.01, *** represents p < 0.001, and **** represents p < 0.0001 unless indicated differently. Significance was assessed between clinical and histopathological data with respect to biomarker expression; between NP-siGP130-CH2.5 and controls (PBS or NP-Bk-CH2.5) with respect to tumor growth; and between siGP130 and controls with respect to cell migration and cell viability.
Results
GP130 expression in bladder tumors
Western blot analysis demonstrated that GP130 was highly expressed in aggressive bladder cancer patient specimens compared to the non-aggressive or normal control bladder specimens (Fig 1A). Also, we demonstrated that GP130 expression correlated significantly with tumor grade (P = 0.027), node category (P = 0.001) and patient survival (P = 0.0001) (Fig 1B). Similarly, Survivin expression was correlated significantly with patient survival (P = 0.048), and Bcl-xL expression correlated significantly with node category (P = 0.040), tumor category (P = 0.040) and patient survival (P = 0.040). Conversely, VEGFR2 expression did not correlate with tumor grade, node category, tumor category or patient survival (Fig 1B). When focusing only on the matched specimens (n = 4), there was 12, 4, 12, and 7, times higher expression of GP130, survivin, Bcl-xL, and VEGFR2, respectively, in the aggressive cancer specimens compared to the matched adjacent normal control tissues (Fig 1C). In addition to the patient specimens, aggressive human bladder cancer cell lines (TCCsup, T24, UM-UC-3) demonstrated higher GP130 expression compared to the more differentiated bladder cancer RT-4 cells (Fig 1D).
Fig 1. Biomarker expression in human bladder specimens and cancer cells.
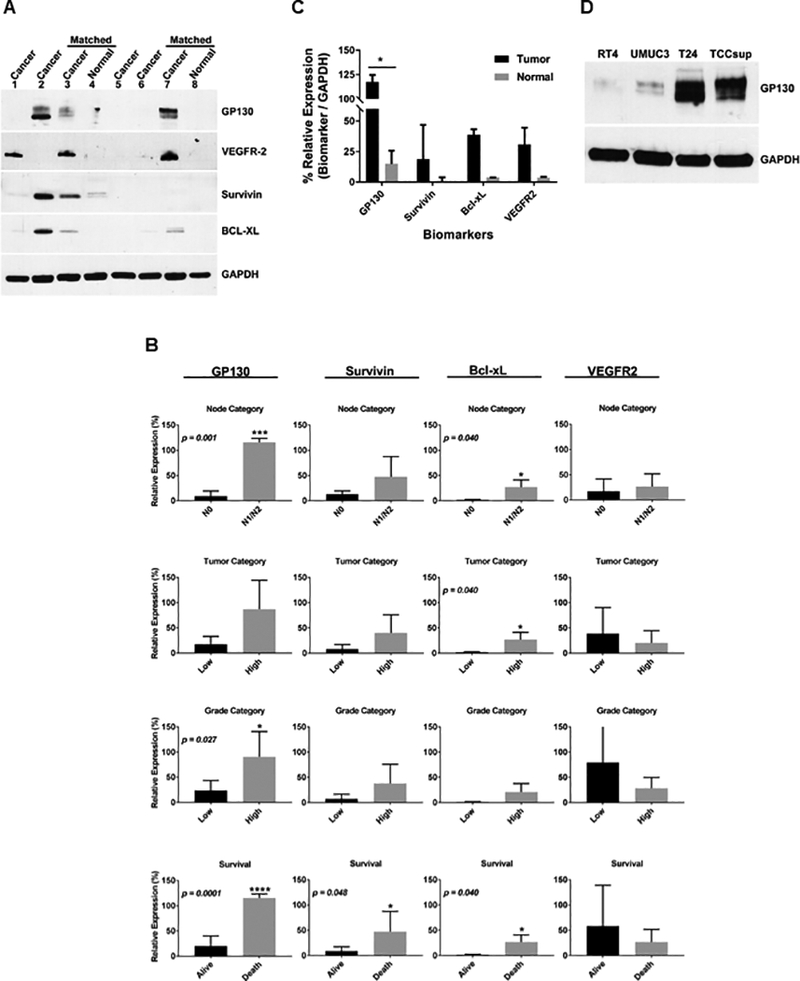
(A) Western blot was performed on aggressive human bladder cancer specimens [2,3,7], non-aggressive human bladder cancer specimens [1,5,6] and normal human bladder [4,8]. Representative blots are shown. (B) Biomarkers (GP130, survivin, Bcl-xL, and VEGFR2) were correlated to the node category, tumor category, tumor grade and survival in human bladder specimens. The error bars represent standard deviation. (C) A bar graph was generated comparing the biomarkers within the patient-matched tumors and adjacent normal tissues. (D) Western blot was performed on human bladder cancer cells (RT-4, UM-UC-3, T24, and TCCsup). A representative blot is shown. GAPDH was used for loading equivalency and protein integrity. Results are presented as mean +/− SD in which * represents p < 0.05, ** represents p < 0.01, *** represents p < 0.001, and **** represents p < 0.0001.
Blocking GP130 expression in in vitro human bladder cancer cells decreased cell viability, growth, and migration.
Greater than 95% reduction in GP130 expression was achieved for UM-UC-3 and T24 human bladder cancer cells treated with siGP130 as demonstrated by western blot (Fig 2A, B). UM-UC-3, T24, and RT-4 bladder cancer cells were treated with 100 nmol/L siGP130, resulting in a 45%, 35% and 20% decrease in cell viability, respectively, as assessed by a WST-1 colorimetric assay (Fig 2C, D, E).
Fig 2. Effect of GP130 siRNA knockdown on GP130 expression and cell viability in in vitro human bladder cancer cell lines.
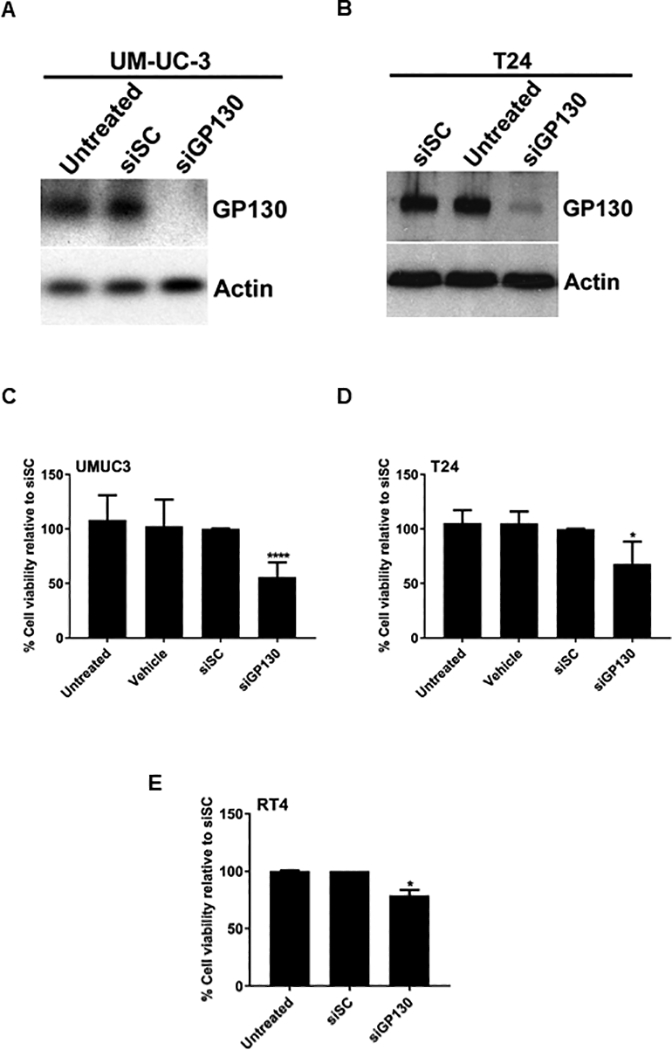
Western blot was performed on human high-grade UM-UC-3 (A) and T24 (B) bladder cancer cells untreated or treated with siSC or siGP130. Actin western blot shows loading equivalency and protein integrity. Viability assays were performed on UM-UC-3 (C), T24 (D) and RT-4 (E) bladder cancer cells untreated or treated with siSC or siGP130 using a WST-1 colorimetric assay. Results are presented as mean +/− SD in which * represents p < 0.05 and **** represents p < 0.0001.
We determined the cell growth (or survival) rate of siGP130 treated UM-UC-3 bladder cancer cells with crystal violet staining (Fig 3A), and confirmed that siGP130 inhibited cell growth by 60 %. A migration (or wound) assay was performed on untreated UM-UC-3 bladder cancer cells or cells treated with vehicle control, siSC control or siGP130. Twelve hours post-scratch, the untreated, vehicle-treated, and siSC-treated UM-UC-3 human bladder cancer cells migrated to cover 71, 71, and 67 % of the scratch, respectively, while the siGP130-treated bladder cancer cells only migrated to cover 21 % of the scratch. After 18 h, the untreated, vehicle-treated, and siSC-treated controls reached 100 % confluency whereas the siGP130-treated cells migrated to cover only 50 % of the scratch, confirming the role of GP130 in inhibiting cell migration (Fig 3B).
Fig 3. Cell growth and migration of bladder cancer cells upon siGP130 knockdown.
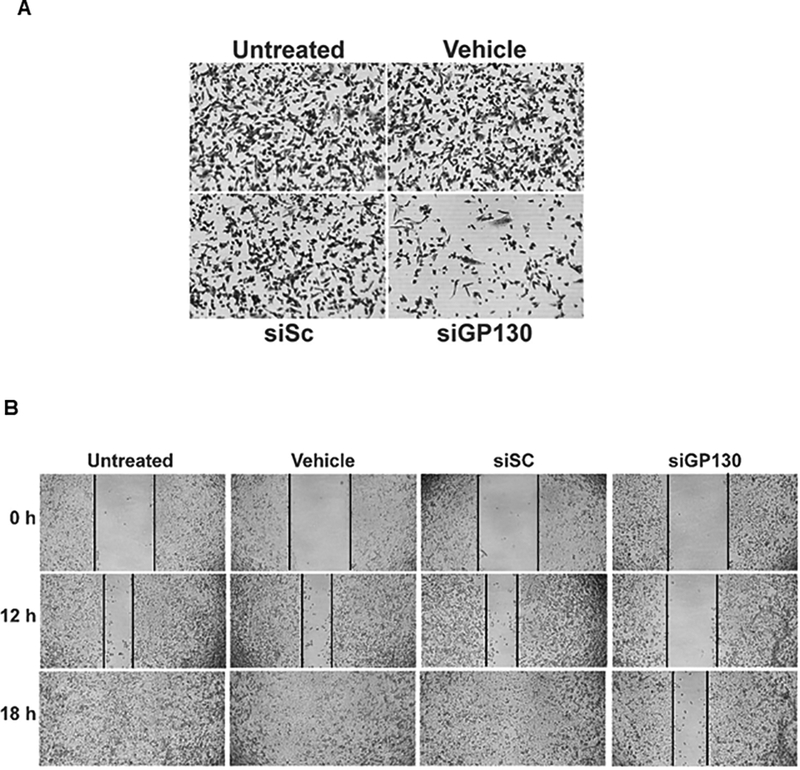
UM-UC-3 bladder cancer cells were treated with siGP130 to assess (A) cell growth as visualized with 0.5% crystal violet, or (B) cell migration as measured by scratch closure.
Tumor response to GP130 siRNA in a xenograft mouse model
Mouse bladder tumors were treated with PBS, NP-Bk-CH2.5 or NP-siGP130-CH2.5 for 14 days. On average, the NP diameters were 152 +/− 16 nm (Table II). Mouse bladder tumors treated with NP-siGP130-CH2.5 were significantly smaller than both the PBS or NP-Bk-CH2.5 controls at day 7 (41 %, 38 %), 11 (65 %, 60 %), and 14 (75 %, 60 %), respectively (Fig 4A).
Fig 4. In vivo treatment of xenograft bladder tumors.
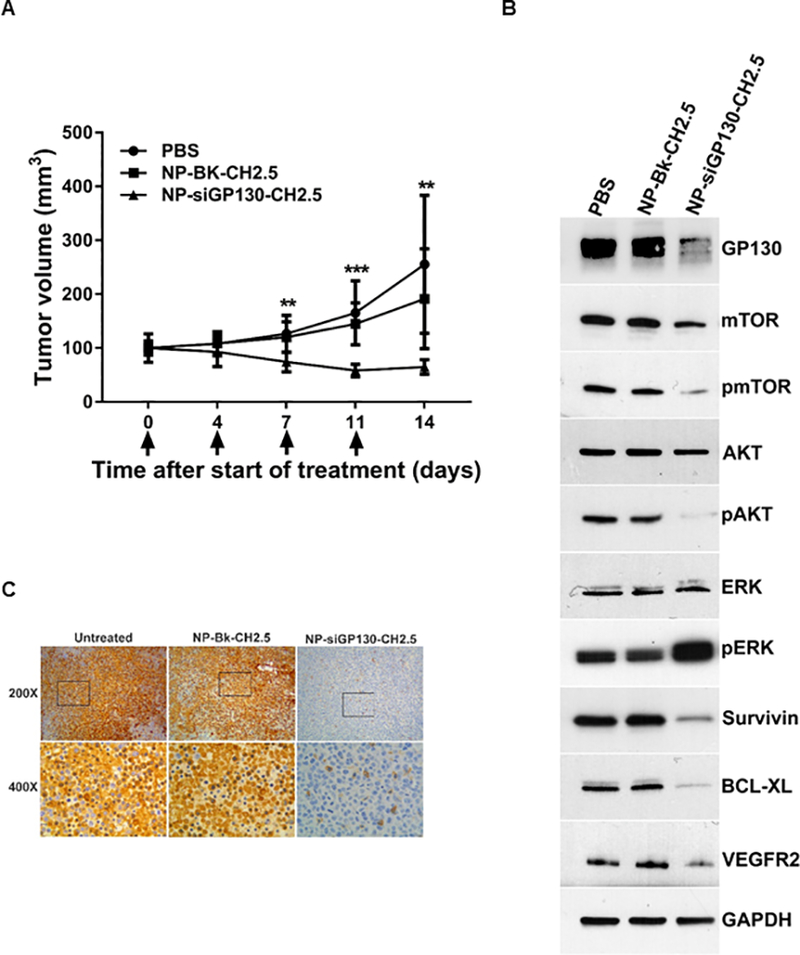
(A) Mice were intratumorally injected on day 0, 4, 7, and 11 with 100 μl of PBS, NP-BK-CH2.5 or NP-siGP130-CH2.5 diluted in PBS and tumor volume determined. Results are presented as mean +/− SD in which ** represents p < 0.01 and *** represents p < 0.001. (B) Western blot of biomarkers within the tumors treated with PBS, NP-Bk-CH2.5 or NP- siGP130-CH2.5. (C) Immunohistochemical analysis of the keratin-20 marker in tumors, treated with PBS, NP-Bk-CH2.5 or NP- siGP130-CH2.5.
Western blot results confirmed that GP130 expression was decreased in the mouse bladder tumors treated with siGP130. Lower levels of GP130 expression were present in the NP-siGP130-CH2.5 treated tumors compared to the PBS or NP-Bk-CH2.5 controls. Upon GP130 inhibition, we observed a decrease in the expression of pAKT, pmTOR, VEGFR2, survivin and Bcl-xL. However, the level of pSTAT3 expression in PBS or NP-Bk-CH2.5 controls was below our level of detection. Interestingly, we observed an increase in pERK expression upon GP130 knockdown (Fig 4B).
In addition, immunohistochemical staining of keratin-20, a marker of high grade urothelial carcinomas, showed elevated levels in the PBS and NP-Bk-CH2.5 controls compared to the NP-siGP130-CH2.5 treated mouse tumors (Fig 4C).
Discussion
To date, the role of GP130 in bladder cancer has not been elucidated. In this study, we assessed GP130, survivin, Bcl-xL and VEGFR2 expression on eleven snap frozen bladder specimens collected from TURBT or cystectomy in which the clinical and histopathological data were available (Table I; Fig 1B). We demonstrated that GP130 expression significantly correlates with an aggressive population of bladder cancers. In particular, GP130 expression significantly correlates with tumor grade, node category and patient survival. In contrast to our findings, Xu and Neamati reported that GP130 was significantly decreased in bladder cancer (9). In our study, low GP130 expression was mainly found in non-aggressive papillary bladder tumors whereas elevated GP130 expression levels correlated with malignant tumors with poor outcome, which may add value in the diagnosis of aggressive bladder tumors. Nevertheless, GP130 may have a role in cancer growth as it was reported by Xu and Neamati that GP130 was co-expressed with cancer promoting genes including BPTF, a cancer growth and proliferation marker, as well as with ARHGAP18, a metastasis marker using a lymphoma microarray (9).
To functionally assess the role of GP130 in bladder cancer development, we used siGP130 to decrease GP130 expression in in vitro bladder tumor models. We observed a decrease in bladder cell growth, viability, and migration, corresponding with diminished GP130 expression thereby suggesting a tumor-specific GP130 effect. To further confirm the role of GP130 in tumor growth, we utilized our previously described NP delivery system in vivo (12). Based on our previous studies, we used our NP delivery system to prolong siRNA protection in our animal model. Also, we reported that encapsulating siRNAs in NPs increased their half-life from hours to weeks, thereby enabling siRNA release from the NPs for an extended duration (15, 18). The released siRNA decreased target mRNA expression, and resulted in diminished protein expression and tumor growth inhibition (12). To confirm our findings with respect to pro-survival and cell migration markers such as survivin, Bcl-xL, and VEGFR2 from our human bladder cancer specimens, we evaluated these markers in our bladder cancer mouse model. We found the aforementioned markers to be downregulated upon intratumoral injection of NP-siGP130 in our mouse tumors, suggesting a possible therapeutic role of GP130 in bladder cancer growth. However, future experiments will be required to assess the potential off-target effects on normal tissue expression of GP130 in the liver, kidney, colon and urothelium.
In addition to GP130, survivin expression also correlated with survival in our human bladder cancer specimens. Previously, our group showed that survivin was able to predict aggressive tumor behavior in vivo (19) and to predict bladder cancer recurrence, using human bladder cancer specimens (20). While VEGFR2 expression has been reported to be linked to the invasiveness of human bladder cancers (21, 22), in our study VEGFR2 was not significantly correlated with our clinical and histopathological data, although it appeared to trend towards significance. In contrast, when we examined pro-survival and cell migration factors in our NP-siGP130-CH2.5 treated mouse tumors, we found survivin, Bxl-xL, and VEGFR2 to be downregulated, suggesting a role for GP130 in bladder cancer growth. A previous report by Nakanishi and colleagues suggested that inhibiting VEGFR expression resulted in a decrease in bladder cancer growth and invasion (23).
We demonstrated that GP130, located at a central point for a number of oncogenic signaling cascades, has a role in bladder cancer growth. Using our xenograft tumor model (Fig. 4B), we demonstrated an inhibition of the downstream pathway AKT (and mTOR) as was noted by a decrease in the phosphorylation levels, correlating with a decrease in bladder cancer growth. This phenomenon of de-activating AKT (and mTOR) has been well established in playing an important role in regulating cancer growth (24). Although STAT3, an anti-apoptosis related protein, is one of the major downstream pathways of GP130/IL6, and is up-regulated in many tumors including bladder cancer (25, 26), it did not appear to be active in our bladder cancer model, suggesting a STAT3 independent mechanism. Godoy-Tundidor et al., previously demonstrated that proliferation could be stimulated in prostate cancer cells through the PI3K/AKT pathway without activating STAT3, which also may be occurring in our bladder cancer model (27). Currently, there are no small-molecule GP130 inhibitors in clinical trials. However, there are reports of small-molecule GP130 inhibitors including SC144 (11) and LMT-28 (28) being tested pre-clinically. We have tested the SC144 inhibitor using in vitro bladder cancer cells and observed a significant knockdown of GP130, survivin and Bcl-xL expression. In addition, a dose-response curve assessing cell viability was performed with SC144 (Fig S1). We found a decrease in cell growth and cell migration when SC144 was used at 3.5 μM (Fig S2; S3). Similarly, Xu et al. demonstrated a decrease in GP130, survivin and Bcl-xL in OVCAR-8 cells when treated with 2 μM SC144 (11).
Knocking down GP130 alone in our in vivo model did not completely eradicate the tumor, suggesting that additional feedback mechanisms are involved. We examined the ERK pathway and found that pERK was upregulated in our NP-siGP130 treated bladder tumors, indicating that this downstream pathway was still active and may be responsible in preventing complete abrogation of the bladder tumor. Hepburn et al. demonstrated that activation of the ERK pathway supported a role for cancer stem cell self-renewal and tumorigenicity (29).
For the first time, we report that GP130 is linked to the aggressiveness of human bladder tumors, and that blocking GP130 using surface modified NPs encapsulating siRNA GP130 may have a therapeutic role in controlling tumor growth. Therefore, future studies may involve dual-loaded NPs (siGP130 and siERK) that will have the ability to target both GP130 and ERK. This may be a more effective strategy for completely eradicating aggressive bladder tumors.
Supplementary Material
{kind=link}
{kind=link}
{kind=link}
Acknowledgements
The authors thank Marcia Wheeler for helpful discussions with this manuscript. We also thank our Genitourinary Data and Biospecimen Repository team (Drs. Martha Boeke and Brian Shuch (PI)) for the management of the Biospecimen data and providing the GU specimens.
Grant Support
This study was supported in part by the NIH grant 5RC1DK087015 (Weiss) from the National Institute of Diabetes and Digestive and Kidney Diseases, NIH grant EB000487 (Saltzman), and a Pilot Grant from the Yale Comprehensive Cancer Center (Martin & Weiss).
Financial support:
This study was supported in part by the NIH grant 5RC1DK087015 (Weiss) from the National Institute of Diabetes and Digestive and Kidney Diseases, NIH grant EB000487 (Saltzman), and a Pilot Grant from the Yale Comprehensive Cancer Center (Martin & Weiss).
Footnotes
Conflict of Interest:
The authors declare no potential conflicts of interest.
References
- 1.Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61:69–90. [DOI] [PubMed] [Google Scholar]
- 2.Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018;68:7–30. [DOI] [PubMed] [Google Scholar]
- 3.Kirkali Z, Chan T, Manoharan M, Algaba F, Busch C, Cheng L, et al. Bladder cancer: epidemiology, staging and grading, and diagnosis. Urology. 2005;66:4–34. [DOI] [PubMed] [Google Scholar]
- 4.Heney NM, Ahmed S, Flanagan MJ, Frable W, Corder MP, Hafermann MD, et al. Superficial bladder cancer: progression and recurrence. J Urol. 1983;130:1083–6. [DOI] [PubMed] [Google Scholar]
- 5.Bambury RM, Rosenberg JE. Actionable mutations in muscle-invasive bladder cancer. Curr Opin Urol. 2013;23:472–8. [DOI] [PubMed] [Google Scholar]
- 6.Botteman MF, Pashos CL, Redaelli A, Laskin B, Hauser R. The health economics of bladder cancer: a comprehensive review of the published literature. Pharmacoeconomics. 2003;21:1315–30. [DOI] [PubMed] [Google Scholar]
- 7.Castillo-Martin M, Domingo-Domenech J, Karni-Schmidt O, Matos T, Cordon-Cardo C. Molecular pathways of urothelial development and bladder tumorigenesis. Urol Oncol. 2010;28:401–8. [DOI] [PubMed] [Google Scholar]
- 8.Mitra N, Indurkhya A. A propensity score approach to estimating the cost-effectiveness of medical therapies from observational data. Health Econ. 2005;14:805–15. [DOI] [PubMed] [Google Scholar]
- 9.Xu S, Neamati N. gp130: a promising drug target for cancer therapy. Expert Opin Ther Targets. 2013;17:1303–28. [DOI] [PubMed] [Google Scholar]
- 10.Selander KS, Li L, Watson L, Merrell M, Dahmen H, Heinrich PC, et al. Inhibition of gp130 signaling in breast cancer blocks constitutive activation of Stat3 and inhibits in vivo malignancy. Cancer Res. 2004;64:6924–33. [DOI] [PubMed] [Google Scholar]
- 11.Xu S, Grande F, Garofalo A, Neamati N. Discovery of a novel orally active small-molecule gp130 inhibitor for the treatment of ovarian cancer. Mol Cancer Ther. 2013;12:937–49. [DOI] [PubMed] [Google Scholar]
- 12.Martin DT, Steinbach JM, Liu J, Shimizu S, Kaimakliotis HZ, Wheeler MA, et al. Surface-modified nanoparticles enhance transurothelial penetration and delivery of survivin siRNA in treating bladder cancer. Mol Cancer Ther. 2014;13:71–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cheong JK, Nguyen TH, Wang H, Tan P, Voorhoeve PM, Lee SH, et al. IC261 induces cell cycle arrest and apoptosis of human cancer cells via CK1delta/varepsilon and Wnt/beta-catenin independent inhibition of mitotic spindle formation. Oncogene. 2011;30:2558–69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Martin DT, Hoimes CJ, Kaimakliotis HZ, Cheng CJ, Zhang K, Liu J, et al. Nanoparticles for urothelium penetration and delivery of the histone deacetylase inhibitor belinostat for treatment of bladder cancer. Nanomedicine. 2013;9:1124–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Woodrow KA, Cu Y, Booth CJ, Saucier-Sawyer JK, Wood MJ, Saltzman WM. Intravaginal gene silencing using biodegradable polymer nanoparticles densely loaded with small-interfering RNA. Nat Mater. 2009;8:526–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Martin DT, Gendron RL, Jarzembowski JA, Perry A, Collins MH, Pushpanathan C, et al. Tubedown expression correlates with the differentiation status and aggressiveness of neuroblastic tumors. Clin Cancer Res. 2007;13:1480–7. [DOI] [PubMed] [Google Scholar]
- 17.Desai S, Lim SD, Jimenez RE, Chun T, Keane TE, McKenney JK, et al. Relationship of cytokeratin 20 and CD44 protein expression with WHO/ISUP grade in pTa and pT1 papillary urothelial neoplasia. Mod Pathol. 2000;13:1315–23. [DOI] [PubMed] [Google Scholar]
- 18.Steinbach JM, Weller CE, Booth CJ, Saltzman WM. Polymer nanoparticles encapsulating siRNA for treatment of HSV-2 genital infection. J Control Release. 2012;162:102–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Salz W, Eisenberg D, Plescia J, Garlick DS, Weiss RM, Wu XR, et al. A survivin gene signature predicts aggressive tumor behavior. Cancer Res. 2005;65:3531–4. [DOI] [PubMed] [Google Scholar]
- 20.Swana HS, Grossman D, Anthony JN, Weiss RM, Altieri DC. Tumor content of the antiapoptosis molecule survivin and recurrence of bladder cancer. N Engl J Med. 1999;341:452–3. [DOI] [PubMed] [Google Scholar]
- 21.Kopparapu PK, Boorjian SA, Robinson BD, Downes M, Gudas LJ, Mongan NP, et al. Expression of VEGF and its receptors VEGFR1/VEGFR2 is associated with invasiveness of bladder cancer. Anticancer Res. 2013;33:2381–90. [PubMed] [Google Scholar]
- 22.Xia G, Kumar SR, Hawes D, Cai J, Hassanieh L, Groshen S, et al. Expression and significance of vascular endothelial growth factor receptor 2 in bladder cancer. J Urol. 2006;175:1245–52. [DOI] [PubMed] [Google Scholar]
- 23.Nakanishi R, Oka N, Nakatsuji H, Koizumi T, Sakaki M, Takahashi M, et al. Effect of vascular endothelial growth factor and its receptor inhibitor on proliferation and invasion in bladder cancer. Urol Int. 2009;83:98–106. [DOI] [PubMed] [Google Scholar]
- 24.Follo MY, Manzoli L, Poli A, McCubrey JA, Cocco L. PLC and PI3K/Akt/mTOR signalling in disease and cancer. Adv Biol Regul. 2015;57C:10–6. [DOI] [PubMed] [Google Scholar]
- 25.Buettner R, Mora LB, Jove R. Activated STAT signaling in human tumors provides novel molecular targets for therapeutic intervention. Clin Cancer Res. 2002;8:945–54. [PubMed] [Google Scholar]
- 26.Chen CL, Cen L, Kohout J, Hutzen B, Chan C, Hsieh FC, et al. Signal transducer and activator of transcription 3 activation is associated with bladder cancer cell growth and survival. Mol Cancer. 2008;7:78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Godoy-Tundidor S, Cavarretta IT, Fuchs D, Fiechtl M, Steiner H, Friedbichler K, et al. Interleukin-6 and oncostatin M stimulation of proliferation of prostate cancer 22Rv1 cells through the signaling pathways of p38 mitogen-activated protein kinase and phosphatidylinositol 3-kinase. Prostate. 2005;64:209–16. [DOI] [PubMed] [Google Scholar]
- 28.Hong SS, Choi JH, Lee SY, Park YH, Park KY, Lee JY, et al. A Novel Small-Molecule Inhibitor Targeting the IL-6 Receptor beta Subunit, Glycoprotein 130. J Immunol. 2015;195:237–45. [DOI] [PubMed] [Google Scholar]
- 29.Hepburn AC, Veeratterapillay R, Williamson SC, El-Sherif A, Sahay N, Thomas HD, et al. Side population in human non-muscle invasive bladder cancer enriches for cancer stem cells that are maintained by MAPK signalling. PLoS One. 2012;7:e50690. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.