

Abstract
Three dimensional (3D) culture has gradually become a research hotspot in the field of drug screening, stem cell research, and tissue engineering due to its more physiological-like morphology and function. In this study, we compared the differences of cell proliferation, population, protein expression and chemoresistance profiles between two dimensional (2D) and 3D culture of acute lymphoblastic leukemia (ALL) Jurkat cell line. Polycaprolactone (PCL) is used for 3D culture owing to its biochemical properties and compatibility. Culturing of ALL Jurkat cell line in collagen type I coated polycaprolactone scaffold for 168 h increased cell proliferation, attachment, as well as the drug resistance to cytarabine (Ara-C) and daunorubicin (DNR) without changing the original CD2+CD3+CD4+dimCD8−CD34−CD45+dim phenotype, compared to uncoated PCL scaffold and tissue culture plate systems. Molecularly, increased chemoresistance is associated with the upregulation of discoidin domain receptor 1 (DDR1) and transcription factor STAT3. Inhibition of DDR1 activity by DDR1-specific inhibitor DDR-IN-1 accelerated cell death in the presence of Ara-C, DNR or their combination. These results demonstrated that 3D culture enhances chemoresistance of ALL Jurkat cell line by increasing DDR1 expression. Importantly, the cell adhesion-mediated drug resistance induced by DDR1 in the scaffold was similar to the clinical situation, indicating the 3D culture of cancer cells recapitulate the in vivo tumor environment and this platform can be used as a promising pre-clinic drug-screen system.
Keywords: 3D culture, daunorubicin, cytarabine, discoidin domain receptor 1, resistance
Introduction
Hematological malignancy of acute leukemia is highly heterogeneous. At present, there are no standardized chemotherapy regimens for acute lymphoblastic leukemia (ALL) bone marrow microenvironment (1), with specific three dimensional (3D) structure, can support the growth of leukemia cells as well as protect them from chemotherapy drug induced death (2–4). 3D scaffold needs some properties to better mimic the bone marrow environment, such as adequate pore size, interconnected pores and high surface area, which will facilitate cells penetration and the formation of cellular associations (5). 3D scaffold provides greater physiological relevance in comparison with conventional 2D culture (6). The shortcoming of 2D culture is getting increasingly obvious by studies showing that cell behavior and gene expression are significantly influenced by the physical and mechanical properties of their microenvironment (7). Of the drug discoveries based on 2D culture 95% fail to provide clinical benefit (8). In this study, we compared the effects of 3D and 2D culture on drug resistance of leukemic cells. Polycaprolactone (PCL) is widely used as a 3D scaffold material for its good biocompatibility and biodegradability (9,10). For cell culture experiments, the scaffold with a fiber size of 300 nm in diameter and a pore size of 300 nm diameter was used. PCL scaffold increased cell survival and drug resistance toward chemotherapy drug Ara-C and daunorubicin (DNR). Importantly, we found that the molecular mechanisms of cell survival and drug resistance observed in the scaffold were similar to those reported in vivo. Moreover, we explored how to reverse the drug resistance in 3D culture. The developed, molecularly characterized PCL scaffold could serve as a drug-testing platform for chemotherapies against drug-resistant cells.
Materials and methods
Materials
Cell counting kit-8 (CCK-8) and polyvinylidene fluoride (PVDF) membrane were purchased from Beyotime Biotechnology (Shanghai, China). BCA protein assay kit, protease inhibitor cocktail and RIPA lysis buffer were purchased from CWBiotech Co. Ltd. (Beijing, China). Rat-tail type I collagen were purchased from Solarbio Science & Technology Co., Ltd. (Beijing, China). Ara-C, DNR and discoidin domain receptor 1 (DDR1)-IN-1 were purchased from MedChemExpress USA (New Jersey, USA), RPMI-1640 media was purchased from Hyclone (GE Healthcare Life Sciences, Logan, UT, USA). Fetal bovine serum (FBS) was purchased from Biological Industries (Beit Haemek, Israel). CD2-PC5 (cat. no. A07745), CD3-ECD (cat. no. A07748), CD4-FITC (cat. no. A07751), CD8-PE (cat. no. A07756), CD34-ECD (cat. no. B49202), (cat. no. B36294), all purchased from Beckman coulter, Inc. (Brea, CA, USA). Stat3 (cat. no. 9139), DDR1 (no. 5583), p-STAT3 (no. 9145), GAPDH (cat. no. 5174s), primary antibodies, goat anti-mouse (cat. no. 7076s) and rabbit HRP-conjugated secondary antibody (cat. no. 14708s) were purchased from Cell Signaling Technology, Inc., (Danvers, MA, USA). PCL scaffolds were purchased from 3D Biotek LLC (New Jersey, USA).
The study was approved by the Ethics Committee of Qingdao University (Qingdao, China).
Scaffold coating
Collagen, a major extracellular matrix (ECM) component, is coated to scaffold to make cell adhere better and as a ligand of DDR1 (11). DDR1 functions as the unique receptor tyrosine kinase (RTK) which can be activated by collagen (12,13). PCL scaffolds were transferred into the 24-well plates and coated with collagen type I in different concentrations (10, 20, 40, 80, 160 and 320 µg/ml). Then the PCL scaffold was dried in biosafety hood overnight.
Cell culture
The acute T leukemia cell line Jurkat, Clone E6-1 (ATCC® TIB-152™) was cultured in RPMI-1640 supplemented with 10% FBS and 1% penicillin/streptomycin. The cells were maintained at 37°C with 5% CO2 and 95% air (13). For experiments on scaffolds, the seeding density is different from 2.5×105 to 5×105 cells/ml according to experiments and the seeding method was in strict accordance to the 3D Insert™ cell seeding protocol.
Cell proliferation and viability
Cell seeding efficiency was determined 4 h after seeding into scaffold by counting the number of cells remaining in the media (14). Cell proliferation/viability was assessed for 6, 24, 48, 96, 144 and 168 h using CCK-8 reagent. Jurkat cells were seeded at a density of 2.5×105 cells/ml and incubated with CCK-8 reagent for 2 h at 37°C. A total of 100 µl solution was taken for recording the optical density after incubation. Absorbance at 450 nm was measured using the Tecan Safire 2 multifunctional microplate reader.
Scanning electron microscopy (SEM)
In order to visualize the growth of the cells in the scaffold, the scaffold was examined by SEM (JSM-840; JEOL, Ltd., Tokyo, Japan) following cultivation with the Jurkat cells for 72 and 144 h. Briefly, the scaffold was treated with 3% glutaral for 4 h. Then, the scaffold was washed twice with PBS, following 30, 50, 70, 80 and 100% ethanol for dehydration. Then the carbon dioxide critical point dryer (XD-1; Eiko, Japan) was used to dry the scaffold. Prior to SEM analysis, samples were coated with gold for 120 sec using auto fine coater (IB-3; EiKo, Japan).
Flow cytometry analysis
Flow cytometry analysis was performed to analyze whether the cell phenotype changes after cultivation in scaffold for one week. The expression of CD2, CD3, CD4, CD8, CD34 and CD45 were analyzed. Briefly, nonadherent cells were removed by PBS washing, then the cells were digested from the scaffold by collagenase treatment. After digestion, cells were harvested by gently pipetting the cell suspension up and down five times within the scaffold. Then the supernatant was centrifuged at 100 × g for 5 min to pellet the cells. The cells pellet was resuspended in PBS and incubated with antibodies for 15 min at 37°C. Flow cytometry analysis was performed using FC500 (Beckman Coulter, Inc., Brea, CA, USA). Isotypes were used as a control.
Western blot analysis
The cells were lysed with RIPA buffer containing phosphatase and protease inhibitor cocktail. Protein was extracted from cells using an ultrasonicator for two cycles, 5 sec each time with a 10-second interval. After incubation for 15 min on ice, the lysate was centrifuged at 15,493 × g for 20 min. After centrifugation, the supernatant was collected and quantified with BCA protein assay. Total protein (20 µg) was separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to PVDF membrane. After blocking with 5% non-fat milk for 1 h at room temperature, the membranes were probed with primary antibody against DDR1, STAT3, p-STAT3 and GAPDH overnight at 4°C, followed by incubation with HRP-conjugated secondary antibody. The immunoreactive bands were visualized using Fusion FX7 detection reagents (Vilber Lourmat, Marne-la-Vallée, France). Autoradiograms were scanned and the labeled bands were quantified using Image J analysis software (National Institutes of Health, Bethesda, MD, USA).
Chemoresistance of cells cultured on scaffolds
3D culture platforms are capable of mimicking microenvironments in vivo, which provide greater physiological relevance in comparison to conventional 2D cultures (15–18). Ara-C (1 µM) and DNR (500 nM) were used in our experiments according to the clinical application 5×105/ml Jurkat cells were seeded into the tissue culture plate systems (TCPS), uncoated PCL scaffold (UPS) and collagen type I coated polycaprolactone scaffold (CTCPS) for 24 h. Nonadherent cells were removed along with the medium and new medium with or without drugs was added. For TCPS, cells were taken out with medium and centrifuged. After removing the medium, the cells were seeded into wells and medium with or without drugs was added. The medium was replaced every 48 h. Das et al found that inhibition of DDR1 expression significantly enhanced chemosensitivity of breast cancer cells to genotoxic drugs (19). Thus, DDR1 signaling provides a new target for therapeutic intervention to leukemia cells. DDR1-IN-1, an orally bioavailable DDR1 inhibitor (20,21), was used in our experiments.
Statistical analysis
All experimental data were analyzed by Student's t-test. All experiments were done three times. Data are presented as mean ± standard error. P<0.05 was considered to indicate a statistically significant difference. All tests were completed using PRISM 6 (GraphPad Software, Inc., La Jolla, CA, USA).
Results
PCL scaffold coating with ECM proteins
Collagen type I is one of the main bone marrow ECM proteins and also the ligand of DDR1 (22). In order to mimic the bone marrow microenvironment and further study the function of DDR1, PCL scaffold coating with collagen type I was used in our experiments (Fig. 1). The effect of different concentrations of collagen type I on the cell seeding and cell proliferation was improved compared to UPS. Indeed, collagen type I coating enhanced the seeding efficiency. The seeding efficiency increased gradually at low concentration of collagen type I, but decreased at high concentration (160 or 320 µg/ml) when compared with 80 µg/ml collagen type I coated PCL scaffolds (Fig. 2).
Figure 1.

The morphological and structure characteristics of PCL scaffold and the morphology of Jurkat cells. (A) The morphological characteristics of the PCL scaffold in 24 wells. (B) The structure parameters of PCL scaffold: Fiber size of 300 nm in diameter and a pore size of 300 nm diameter. PCL, polycaprolactone.
Figure 2.
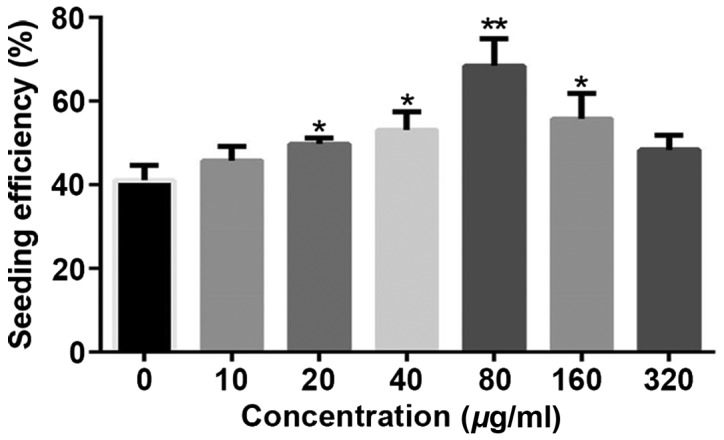
The effect of different concentrations of collagen type I on the seeding efficiency. Seeding efficiency was detected at 12 h after incubation the Jurkat cell into the scaffold. A total of 20, 40, 80, 160 µg/ml CTCPS displayed a >50% seeding efficiency with a statistically significant higher seeding efficiency than the rest (*P<0.05, **P<0.01). CTCPS, collagen type I coated polycaprolactone scaffold.
Cell proliferation on different scaffolds
To investigate the effect of PCL scaffold on cell proliferation, we compared proliferation of Jurkat cells on TCPS, UPS and 80 µg/ml CTCPS for 168 h. CCK-8 reagent was used to analyze the extent of cell proliferation/viability. Different growth kinetics were observed for Jurkat cells cultured on different conditions. Coating the PCL scaffold with collagen type I enhanced cell proliferation compared with the TCPS and UPS, especially for a longer time culture. For the initial 96 h, the TCPS revealed the maximum cell proliferation/viability compared with the UPS and CTCPS. However, with the prolongation of the cell culturing, the CTCPS showed better cell proliferation than the UPS. Moreover, CTCPS can maintain cell proliferation for 168 h. After 166 h, the TCPS revealed a significant deterioration in cell proliferation (Fig. 3).
Figure 3.
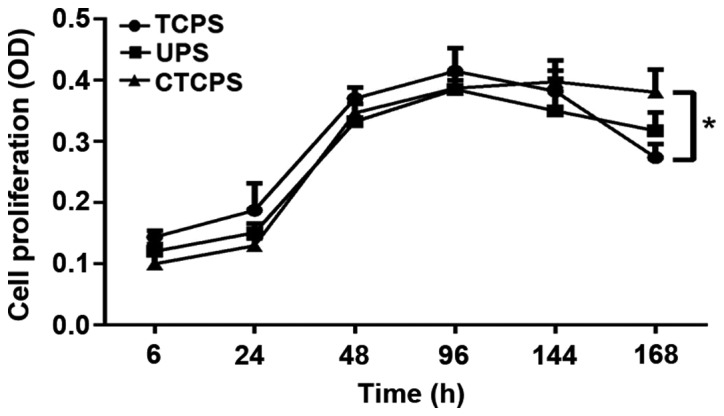
Comparison of proliferation of Jurkat cells on TCPS, UPS and 80 µg/ml CTCPS at different time-points. The increased cell proliferation observed in 80 µg/ml CTCPS at 144 and 168 h. *P<0.05 for CTCPS vs. TCPS at 168 h. TCPS, tissue culture plate systems; UPS, uncoated PCL scaffold; CTCPS, collagen type I coated polycaprolactone scaffold.
Cellular phenotype analysis
The cell proliferation/viability result showed that 80 µg/ml CTCPS was able to support cell survival and proliferation better. It is very important for cells to maintain the initial phenotype and not differentiate with the prolongation of the cell culture. The expression of CD2, CD3, CD4, CD8, CD34, CD45 in Jurkat cells cultured in different conditions such as UPS and 80 µg/ml CTCPS were analyzed at 168 h. The results showed that the percentage of cells with the phenotype of CD2+CD3+CD4+dimCD8−CD34−CD45+dim was maintained for 168 h of culture on the scaffolds compared to that cultured on TCPS (Fig. 4). Similarly, the phenotype of Jurkat cell on the CTCPS and UPS showed no difference in comparison to the TCPS for 168 h.
Figure 4.

Jurkat phenotype maintenance on PCL scaffolds. Fluorescence-activated cell sorting analysis of Jurkat cells phenotype after seeding at different conditions (UPS and 80 µg/ml CTCPS) at 168 h. (A) The initial phenotype of Jurkat cell at TCPS. The phenotype of Jurkat cells at 168 h after incubation with (B) UPS and (C) 80 µg/ml CTCPS. Note the percentage of expression of CD2, CD3, CD4, CD8, CD34, CD45. PCL, polycaprolactone; TCPS, tissue culture plate systems; UPS, uncoated PCL scaffold; CTCPS, collagen type I coated PCL scaffold.
Sensitivity of drugs on Jurkat cells at TCPS, UPS and CTCPS
The 80 µg/ml CTCPS was more suitable for cell proliferation and growth for a longer period without changing cell phenotype. So we used 80 µg/ml CTCPS for drug testing applications. Jurkat cells were treated with Ara-C and DNR or their combination (Fig. 5). CCK-8 reagent was used to test cell cytotoxicity of Jurkat cells cultured on TCPS, UPS and 80 µg/ml CTCPS. Jurkat cells revealed high sensitivity to Ara-C cultured on TCPS. Cell proliferation reduced to 89.58±3.26, 62.86±6.84 and 53.36±3.37% for 48, 144 and 168 h respectively on TCPS, according to the CCK-8 results (Fig. 5A). Cells cultured on 80 µg/ml CTCPS showed resistance to drug treatment, as shown by the result that cell proliferation was 103.98±5.3, 88.69±2.19, 77.11±3.47% at 48, 144 and 168 h. Moreover, the drug sensitivity of cells cultured on UPS was between TCPS and CTCPS, with the relative percentage of 87.98±3.74, 73.28±5.5 and 63.65±0.81% at 48, 144 and 168 h. Cells cultured under the three conditions showed more sensitivity to DNR or Ara-C + DNR combination than Ara-C alone. A significant death of Jurkat cells was observed in TCPS (57.67±8.02, 42±4.58%) when compared to CTCPS (78±5.57, 57.33±5.03%) in the presence of DNR for 96 and 168 h (Fig. 5B; *P<0.05 vs. TCPS). A similar trend of combination use of Ara-C and DNR was obtained in TCPS (24.67±4.73%) and UPS cultures (39±7.55%) in comparison to 80 µg/ml CTCPS (46.33±4.04%) for 168 h (Fig. 5C; **P<0.01 vs. TCPS). In general, the cells cultured on 80 µg/ml CTCPS showed drug resistance for a longer culture period compared with TCPS and UPS.
Figure 5.

Comparison of sensitivity of Jurkat cell to (A) Ara-C, (B) DNR, and (C) combination of both on a TCPS, UPS, and 80 µg/ml (CTCPS). A total of 80 µg/ml CTCPS was different from TCPS and *P<0.05, **P<0.01 vs. TCPS at different time-points in Ara-C, DNR, and combination of both. Ara-C, cytarabine; DNR, daunorubicin; TCPS, tissue culture plate system; UPS, uncoated PCL scaffold; CTCPS, collagen type I coated PCL scaffold.
DDR1 and p-STAT3 mediated cell chemoresistance in CTCPS
We have shown that cell cultured on 80 µg/ml CTCPS revealed chemoresistance compared to TCPS. We further explored the possible mechanisms that lead to cell chemoresistance. Evidence suggests that high expression of DDR1 and p-STAT3 is related to cell proliferation, invasion, migration and chemoresistance of malignant tumors (23,24). STAT3 is the downstream effector of DDR1 pathway and STAT3 can be phosphorylated by DDR1 (25,26). Therefore, we wanted to verify whether the drug resistance to Ara-C and DNR of cells cultured in 3D scaffold is related to the high expression of DDR1 and p-STAT3. The levels of DDR1 and p-STAT3 were highly expressed on the UPS and 80 µg/ml CTCPS compared to the TCPS (Fig. 6). Next, we chose DDR1-IN-1, a potent and selective DDR1 receptor tyrosine kinase inhibitor (21), to further analyze whether DDR1 inhibition would reverse the chemoresistance. The concentration of DDR1-IN-1 used was determined in Jurkat cells cultured in TCPS (Fig. 7A). IC50 of 335 nM with DNR was chosen for further experiments. DDR1-IN-1 (1 µM) which is sufficient to inhibit DDR1 autophosphorylation alone did not inhibit cell proliferation after incubation for 48 h, whereas Ara-C (Fig. 7B) and DNR (Fig. 7C) singly or in combination (Fig. 7D), along with DDR1-IN-1 induced significant cell death, wherein almost all the cells were wiped out in contrast with DDR1-IN-1 alone at 144 h.
Figure 6.

Comparison of the expression of DDR1, STAT3 and P-STAT3 in Jurkat cells on a TCPS, UPS and 80 µg/ml CTCPS for 96 h. (A) The expression levels of DDR1, STAT3 and P-STAT3 in Jurkat cell in different conditions. The expression of P-STAT3 and DDR1 are both increased in UPS and CTCPS. (B) Quantified results of expression levels of DDR1. (C) Quantified results of expression levels of P-STAT3. (D) Quantified results of expression levels of STAT3. *P<0.05, **P<0.01, ***P<0.001. DDR1, discoidin domain receptor 1; TCPS, tissue culture plate system; UPS, uncoated polycaprolactone scaffold; CTCPS, collagen type I coated PCL scaffold.
Figure 7.

Effect of the DDR1 inhibitor DDR1-1N-1 in inducing cell death in the presence of Ara-C and DNR. Different concentration of DDR1-IN-1 combine with Ara-C or DNR in inducing cell death at 48 h. IC50 = 354.4 nM with Ara-C, IC50 = 335 nM with DNR. (A) The increased cell death induced by both Ara-C and DNR on the 80 µg/ml CTCPS at 96, 144 and 168 h. (B-D) Following treatment with DDR1-IN-1. *P<0.05, **P<0.01 vs. TCPS at different time-points. DDR1, discoidin domain receptor 1; Ara-C, cytarabine; DNR, daunorubicin; TCPS, tissue culture plate system; CTCPS, collagen type I coated PCL scaffold.
Discussion
ALL is still a disease that cannot be completely cured by chemotherapy. Hematopoietic stem cell transplantation is the only way to cure leukemia. However, the high cost, easy relapse, severe GVHD side effects and technical limitations make hematopoietic stem cell transplantation hard to carry out for most leukemia patients in China (27). In recent years, the research and development of targeted drugs has become a hot spot (28,29). The prognosis of many hematological malignancies has greatly improved because of the new drugs (30). Drug testing in vitro needs an environment similar to the niche in vivo where those cells reside (31). Although researchers are trying to establish cell microenvironments in vitro which is closer to the bone marrow niche in vivo, the progress is unsatisfactory. Since leukemia cells reside in the bone marrow niche, where leukemia cells escape from the chemotherapy drugs (32,33). Compared with traditional 2D culture, 3D Insert™ PCL scaffold is a lot more like in vivo environment. Cells expanded on the scaffolds have more physiological-like morphology and function (Figs. 8 and 9). In order to mimic bone marrow microenvironment, the scaffolds are coated with collagen. The results showed that the concentration of 80 µg/ml CTCPS is the maximum cell seeding efficiency, whereas the high concentration (160, 320 µg/ml) leads to a decrease of seeding efficiency. This may be attributed to the quick gel formation rate of high concentration of collagen type I, which blocks the infiltration and migration of Jurkat cells.
Figure 8.

The morphology of Jurkat cells. (A) Morphology of Jurkat cells cultured in the TCPS. (B) Morphology of Jurkat cells adhered to the PCL scaffold and aggregation growth. TCPS, tissue culture plate systems; PCL, polycaprolactone.
Figure 9.

Scanning electron micrographs of PCL scaffolds coated with collagen type I. (A) The 80 µg/ml cultured with the Jurkat cells at 72 h. Jurkat cells adhere to the surface of the scaffold and grow in a cluster. (B) The PCL scaffold coated with 80 µg/ml collagen type I cultured with the Jurkat cells at 144 h. Large numbers of cells divide and aggregate into globes. PCL, polycaprolactone.
The initial cell proliferation lag on scaffolds compared to that in TCPS could be attributed to the limited cells bound to the surface and the scaffold niche at the time of seeding (in order to reduce the number of cell culture medium replaced, we seeded 2×104 cells per well on scaffold), the cells also need an adaptive process for new culture environments. That is the possible reason why the proliferation of cells for 48 and 96 h on TCPS was higher than that on UPS and CTCPS. With the prolongation of culture time, more cells adhere to the scaffold surface, divide and migrate into the scaffold and get adapted to the new culture environments. Moreover, this scenario was lacking in the TCPS; that is the possible reason why the cell proliferation was significantly reduced by 144 and 168 h. With the increasing number of cells, the decrease of cell surface area for collagen type I adsorption is inevitable, leading to less area for cell attachment. Thereby G0/G1 cells decrease and non-adhered cells actively divide, overpopulate and die, indicating that CTCPS are more suitable than TCPS and UPS for longer time culture (14,34).
We further explored whether different culture conditions lead to changes in cell phenotype. The result showed that despite culturing Jurkat cells for 168 h, the original phenotype of Jurkat cells was retained the same on the 80 µg/ml CTCPS and UPS compared to TCPS. That is an important characteristic in studies of leukemia cells. It has been reported that DDR1 activates STAT3 signaling during tumor progression in human breast cancer metastasis (25). Overexpression of discoidin DDR1 enhanced the cisplatin resistance of ovarian cancer cells (35), but in ALL, the expression and effect of DDR1 on the drug-resistance is not yet clear. The result showed that the expression levels of DDR1, P-STAT3 in Jurkat cells cultured in the CTCPS were higher than those of cells cultured in TCPS significantly suggesting that the increased expression of DDR1 activated by collagen type I may be the possible reason for the cell survival. The cells grow in a 3D environment may contribute to the expression of DDR1, and may be the reason why the expression levels of DDR1 in the UPS were higher than in the TCPS. The treatment of Jurkat cells cultured on CTCPS with Ara-C, DNR or their combination revealed low sensitivity compared with TCPS until the 168 h. The conclusion of the experiment proves that DDR1/STAT3 mediated mechanism may be the reason of Jurkat cell chemoresistance. We hypothesize that the collagen type I promote the Jurkat cells to harbor in the 3-D niche of the scaffold and activate the DDR1 protein as a ligand. Activation of the DDR1 pathway leads to cell chemoresistance (19).
To further confirm the mechanism, we use DDR1 receptor tyrosine kinase inhibitor DDR1-IN-1 to block DDR1. By blocking the function of DDR1, the sensitivity of leukemic cells to chemotherapeutic drugs is enhanced. These results indicate that the explored CTCPS could support chemotherapy drug resistance to Ara-C, DNR or combination and chemoresistance mechanism operates through the DDR1/STAT3 pathway, recapitulating at least in part the drug resistance seen in the in vivo condition. Thus 3D architecture is also important in providing the chemoresistance. This could be attributed to the similar morphology of the PCL scaffolds to the bone marrow, as collagen type I helps in cell chemoresistance by activating DDR1 (36).
Biomimetic structure is an important parameter in the clinical trial of drugs. Because in some cases the cells were sensitive to the drug in 2D culture, but were resistant to 3D culture (37,38). On one hand, we think that the bionic 3D scaffold with the capability of supporting the leukemic cells for a longer period has immense potential in drug screening. On the other hand, in response to DDR1 as a drug target, drugs can be designed for the treatment of leukemia, which include antibodies against the DDR1 ligand or the receptor itself or a small molecular chemical inhibitor for the intracellular domain of DDR1 kinase. The application of 3D cell culture system enables researchers to create superior in vitro models to obtain more realistic physiological results from studies in vitro. As a result, the use of 3D cell culture will decrease the overall therapeutic and pharmaceutical product development cost and shorten the time to market.
Acknowledgements
Not applicable.
Funding
The project described was supported by Qingdao City People's Livelihood Science and technology project (16-6-2-30-nsh).
Availability of data and materials
The datasets used and/or analyzed during the present study are available from the corresponding author on reasonable request.
Authors' contributions
JG and CZ conceived and designed the study and were responsible for cell culture. RY, AS, LS, XL, SW, ZS and TL detected cell proliferation and viability and performed flow cytometry analysis. SL, YG, JL, XF and WW contributed to western blot analysis. HZ, ZC, GL and FM analyzed chemoresistance of cells cultured on scaffolds. All authors read and approved the final manuscript.
Ethics approval and consent to participate
The study was approved by the Ethics Committee of Qingdao University (Qingdao, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Nair MS, Mony U, Menon D, Koyakutty M, Sidharthan N, Pavithran K, Nair SV, Menon KN. Development and molecular characterization of polymeric micro-nanofibrous scaffold of a defined 3-D niche for in vitro chemosensitivity analysis against acute myeloid leukemia cells. Int J Nanomedicine. 2015;10:3603–3622. doi: 10.2147/IJN.S80397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Funayama K, Murai F, Shimane M, Nomura H, Asano S. Adhesion-induced drug resistance in leukemia stem cells. Pharmacology. 2010;86:79–84. doi: 10.1159/000305344. [DOI] [PubMed] [Google Scholar]
- 3.Wojtkowiak JW, Verduzco D, Schramm KJ, Gillies RJ. Drug resistance and cellular adaptation to tumor acidic pH microenvironment. Mol Pharm. 2011;8:2032–2038. doi: 10.1021/mp200292c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wang JY, Yu P, Chen S, Xing H, Chen Y, Wang M, Tang K, Tian Z, Rao Q, Wang J. Activation of Rac1 GTPase promotes leukemia cell chemotherapy resistance, quiescence and niche interaction. Mol Oncol. 2013;7:907–916. doi: 10.1016/j.molonc.2013.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Dumbleton J, Agarwal P, Huang H, Hogrebe N, Han R, Gooch KJ, He X. The effect of RGD peptide on 2D and miniaturized 3D culture of HEPM cells, MSCs, and ADSCs with alginate hydrogel. Cell Mol Bioeng. 2016;9:277–288. doi: 10.1007/s12195-016-0428-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Duval K, Grover H, Han LH, Mou Y, Pegoraro AF, Fredberg J, Chen Z. Modeling physiological events in 2D vs. 3D cell culture. Physiology (Bethesda) 2017;32:266–277. doi: 10.1152/physiol.00036.2016. [DOI] [PMC free article] [PubMed] [Google Scholar] [Research Misconduct Found]
- 7.Nugraha B, Mohr MA, Ponti A, Emmert MY, Weibel F, Hoerstrup SP, Moll S, Certa U, Prunotto M, Pantazis P. Monitoring and manipulating cellular crosstalk during kidney fibrosis inside a 3D in vitro co-culture. Sci Rep. 2017;7:14490. doi: 10.1038/s41598-017-12683-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hou J, Hong Z, Feng F, Chai Y, Zhang Y, Jiang Q, Hu Y, Wu S, Wu Y, Gao X, et al. A novel chemotherapeutic sensitivity-testing system based on collagen gel droplet embedded 3D-culture methods for hepatocellular carcinoma. BMC Cancer. 2017;17:729. doi: 10.1186/s12885-017-3706-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Burton TP, Corcoran A, Callanan A. The effect of electrospun polycaprolactone scaffold morphology on human kidney epithelial cells. Biomed Mater. 2017;13:015006. doi: 10.1088/1748-605X/aa8dde. [DOI] [PubMed] [Google Scholar]
- 10.Caicedo-Carvajal CE, Liu Q, Remache Y, Goy A, Suh KS. Cancer tissue engineering: A novel 3D polystyrene scaffold for in vitro isolation and amplification of lymphoma cancer cells from heterogeneous cell mixtures. J Tissue Eng. 2011;2011:362326. doi: 10.4061/2011/362326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hur H, Ham IH, Lee D, Jin H, Aguilera KY, Oh HJ, Han SU, Kwon JE, Kim YB, Ding K, et al. Discoidin domain receptor 1 activity drives an aggressive phenotype in gastric carcinoma. BMC Cancer. 2017;17:87. doi: 10.1186/s12885-017-3051-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Moreau V, Saltel F. Type I collagen fibrils and discoidin domain receptor 1 set invadosomes straight. Mol Cell Oncol. 2015;2:e1004963. doi: 10.1080/23723556.2015.1004963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Stępnik M, Spryszyńska S, Gorzkiewicz A, Ferlińska M. Cytotoxicity of anticancer drugs and PJ-34 (poly(ADP-ribose)polymerase-1 (PARP-1) inhibitor) on HL-60 and Jurkat cells. Adv Clin Exp Med. 2017;26:379–385. doi: 10.17219/acem/60848. [DOI] [PubMed] [Google Scholar]
- 14.Blanco TM, Mantalaris A, Bismarck A, Panoskaltsis N. The development of a three-dimensional scaffold for ex vivo biomimicry of human acute myeloid leukaemia. Biomaterials. 2010;31:2243–2251. doi: 10.1016/j.biomaterials.2009.11.094. [DOI] [PubMed] [Google Scholar]
- 15.Bao M, Xie J, Piruska A, Huck WTS. 3D microniches reveal the importance of cell size and shape. Nat Commun. 2017;8:1962. doi: 10.1038/s41467-017-02163-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bell CC, Dankers ACA, Lauschke VM, Sison-Young R, Jenkins R, Rowe C, Goldring CE, Park K, Regan SL, Walker T, et al. Comparison of hepatic 2D sandwich cultures and 3D spheroids for long-term toxicity applications: A multi-center study. Toxicol Sci. 2018;162:655–666. doi: 10.1093/toxsci/kfx289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Xue R, Qian Y, Li L, Yao G, Yang L, Sun Y. Polycaprolactone nanofiber scaffold enhances the osteogenic differentiation potency of various human tissue-derived mesenchymal stem cells. Stem Cell Res Ther. 2017;8:148. doi: 10.1186/s13287-017-0588-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Yildirim ED, Besunder R, Pappas D, Allen F, Güçeri S, Sun W. Accelerated differentiation of osteoblast cells on polycaprolactone scaffolds driven by a combined effect of protein coating and plasma modification. Biofabrication. 2010;2:014109. doi: 10.1088/1758-5082/2/1/014109. [DOI] [PubMed] [Google Scholar]
- 19.Das S, Ongusaha PP, Yang YS, Park JM, Aaronson SA, Lee SW. Discoidin domain receptor 1 receptor tyrosine kinase induces cyclooxygenase-2 and promotes chemoresistance through nuclear factor-kappaB pathway activation. Cancer Res. 2006;66:8123–8130. doi: 10.1158/0008-5472.CAN-06-1215. [DOI] [PubMed] [Google Scholar]
- 20.Canning P, Tan L, Chu K, Lee SW, Gray NS, Bullock AN. Structural mechanisms determining inhibition of the collagen receptor DDR1 by selective and multi-targeted type II kinase inhibitors. J Mol Biol. 2014;426:2457–2470. doi: 10.1016/j.jmb.2014.04.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kim HG, Tan L, Weisberg EL, Liu F, Canning P, Choi HG, Ezell SA, Wu H, Zhao Z, Wang J, et al. Discovery of a potent and selective DDR1 receptor tyrosine kinase inhibitor. ACS Chem Biol. 2013;8:2145–2150. doi: 10.1021/cb400430t. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fang Y, Wang B, Zhao Y, Xiao Z, Li J, Cui Y, Han S, Wei J, Chen B, Han J, et al. Collagen scaffold microenvironments modulate cell lineage commitment for differentiation of bone marrow cells into regulatory dendritic cells. Sci Rep. 2017;7:42049. doi: 10.1038/srep42049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lin CH, Chiang MC, Chen YJ. STAT3 mediates resistance to anoikis and promotes invasiveness of nasopharyngeal cancer cells. Int J Mol Med. 2017;40:1549–1556. doi: 10.3892/ijmm.2017.3151. [DOI] [PubMed] [Google Scholar]
- 24.Yang JC, Zhang Y, He SJ, Li MM, Cai XL, Wang H, Xu LM, Cao J. TM4SF1 promotes metastasis of pancreatic cancer via regulating the expression of DDR1. Sci Rep. 2017;7:45895. doi: 10.1038/srep45895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gao H, Chakraborty G, Zhang Z, Akalay I, Gadiya M, Gao Y, Sinha S, Hu J, Jiang C, Akram M, et al. Multi-organ site metastatic reactivation mediated by non-canonical discoidin domain receptor 1 signaling. Cell. 2016;166:47–62. doi: 10.1016/j.cell.2016.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Wang Z, Sun X, Bao Y, Mo J, Du H, Hu J, Zhang X. E2F1 silencing inhibits migration and invasion of osteosarcoma cells via regulating DDR1 expression. Int J Oncol. 2017;51:1639–1650. doi: 10.3892/ijo.2017.4165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Yang C, Zhang X. Incidence survey of leukemia in China. Chin Med Sci J. 1991;6:65–70. [PubMed] [Google Scholar]
- 28.Broxterman HJ, Georgopapadakou NH. Anticancer therapeutics: ‘Addictive’ targets, multi-targeted drugs, new drug combinations. Drug Resist Updat. 2005;8:183–197. doi: 10.1016/j.drup.2005.07.002. [DOI] [PubMed] [Google Scholar]
- 29.Da Costa EM, McInnes G, Beaudry A, Raynal NJ. DNA methylation-targeted drugs. Cancer J. 2017;23:270–276. doi: 10.1097/PPO.0000000000000278. [DOI] [PubMed] [Google Scholar]
- 30.Jamison C, Nelson D, Eren M, Gauchan D, Ramaekers R, Norvell M, Copur MS. What is the optimal dose and schedule for dasatinib in chronic myeloid leukemia: Two case reports and review of the literature. Oncol Res. 2015;23:1–5. doi: 10.3727/096504015X14452563485986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wang JZ, Zhu YX, Ma HC, Chen SN, Chao JY, Ruan WD, Wang D, Du FG, Meng YZ. Developing multi-cellular tumor spheroid model (MCTS) in the chitosan/collagen/alginate (CCA) fibrous scaffold for anticancer drug screening. Mater Sci Eng C. 2016;62:215–225. doi: 10.1016/j.msea.2016.02.064. [DOI] [PubMed] [Google Scholar]
- 32.Poulos MG, Gars EJ, Gutkin MC, Kloss CC, Ginsberg M, Scandura JM, Rafii S, Butler JM. Activation of the vascular niche supports leukemic progression and resistance to chemotherapy. Exp Hematol. 2014;42:976–986.e3. doi: 10.1016/j.exphem.2014.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Prieto-Vila M, Takahashi RU, Usuba W, Kohama I, Ochiya T. Drug resistance driven by cancer stem cells and their niche. Int J Mol Sci. 2017;18:2574. doi: 10.3390/ijms18122574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Feng Q, Chai C, Jiang XS, Leong KW, Mao HQ. Expansion of engrafting human hematopoietic stem/progenitor cells in three-dimensional scaffolds with surface-immobilized fibronectin. J Biomed Mater Res A. 2006;78:781–791. doi: 10.1002/jbm.a.30829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Deng Y, Zhao F, Hui L, Li X, Zhang D, Lin W, Chen Z, Ning Y. Suppressing miR-199a-3p by promoter methylation contributes to tumor aggressiveness and cisplatin resistance of ovarian cancer through promoting DDR1 expression. J Ovarian Res. 2017;10:50. doi: 10.1186/s13048-017-0333-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Carbone A, Gloghini A. Activated DDR1 increases RS cell survival. Blood. 2013;122:4152–4154. doi: 10.1182/blood-2013-11-534123. [DOI] [PubMed] [Google Scholar]
- 37.Kim MK, Kim Y, Choo H, Chong Y. Quercetin-glutamic acid conjugate with a non-hydrolysable linker; a novel scaffold for multidrug resistance reversal agents through inhibition of P-glycoprotein. Bioorg Med Chem. 2017;25:1219–1226. doi: 10.1016/j.bmc.2016.12.034. [DOI] [PubMed] [Google Scholar]
- 38.Yang Z, Zhao X. A 3D model of ovarian cancer cell lines on peptide nanofiber scaffold to explore the cell-scaffold interaction and chemotherapeutic resistance of anticancer drugs. Int J Nanomedicine. 2011;6:303–310. doi: 10.2147/IJN.S15279. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the present study are available from the corresponding author on reasonable request.