

Abstract
Many mathematical models of evolution assume that all individuals experience the same environment. Here, we study the Moran process in heterogeneous environments. The population is of finite size with two competing types, which are exposed to a fixed number of environmental conditions. Reproductive rate is determined by both the type and the environment. We first calculate the condition for selection to favour the mutant relative to the resident wild-type. In large populations, the mutant is favoured if and only if the mutant’s spatial average reproductive rate exceeds that of the resident. But environmental heterogeneity elucidates an interesting asymmetry between the mutant and the resident. Specifically, mutant heterogeneity suppresses its fixation probability; if this heterogeneity is strong enough, it can even completely offset the effects of selection (including in large populations). By contrast, resident heterogeneity has no effect on a mutant’s fixation probability in large populations and can amplify it in small populations.
Keywords: environmental heterogeneity, evolutionary dynamics, Moran process
1. Introduction
Evolutionary dynamics deals with the appearance and competition of traits over time. The success of an initially rare mutant arising in a population depends on a number of factors, including the population’s spatial structure and the mutant’s reproductive fitness relative to the resident. One quantitative measure of a mutant’s success is its fixation probability, which describes the chance that the mutant’s lineage will take over the population [1]. The effect of a particular property of the population (such as its spatial structure) on natural selection is often measured directly in terms of its effects on this probability of fixation. Among the many noted demographic features that affect evolutionary outcomes, comparatively little is known about the effects of environmental heterogeneity in reproductive fitness on evolutionary dynamics.
One source of interaction and migration heterogeneity is population structure. Lieberman et al. [2] use graphs as a model for population structure and show that ‘isothermal’ structures do not alter fixation probabilities under birth–death updating, expanding upon a related observation for subdivided populations [3]. Non-isothermal graphs can change this fixation probability and, in particular, act as amplifiers or suppressors of selection—a topic of considerable current interest [2,4–13]. Recent work suggests that randomness in dispersal patterns yields either amplifiers or suppressors of selection [11,14–17]. Although spatial structure and frequency-dependent fitness have been incorporated into many evolutionary models, their effects on evolutionary dynamics are not fully understood. Even less is known about the effects of environmental heterogeneity, which can affect fitness through a non-uniform distribution of resources.
Despite the fact that there is still much left to be understood about the effects of environmental heterogeneity, its importance in theoretical models has long been recognized, particularly in population genetics [18–20]. More than 60 years ago, Levene [21] introduced a diploid model in which two alleles are favoured in different ecological niches and showed that genetic equilibrium is possible even when there is no niche in which the heterozygote is favoured over both homozygotes. Haldane & Jayakar [22] subsequently treated a temporal analogue of this fitness asymmetry, which was then incorporated into a study of polymorphism under both spatial and temporal fitness heterogeneity [23]. Arnold & Anderson [24] described the spatial model of Levene [21] as ‘the beginning of theoretical ecological genetics’. Many studies of environmental heterogeneity have focused largely on metapopulation or island models under weak selective pressure, inspired by the evolution of habitat-specialist traits in heterogeneous environments [25–29]. These metapopulation models assume connected islands (habitats) where migration is allowed between islands, and environmental heterogeneity is parametrized by a variable fitness difference between two competing types and assumed to be small (i.e. weak selection). Notably, in the limit of strong connectivity between islands, variations in fitness advantage do not affect fixation probability [30]. Others address the issue of fixation in two-island [31] and multi-habitat [32] models with variable fitness.
A more fine-grained heterogeneity requires an extension of the stepping-stone models to evolutionary graphs [33–36]. So far, much of the work in this area has been done through numerical simulations of specific structures and fitness distributions. For example, Manem et al. [37] demonstrated via death–birth simulations on a structured mesh that heterogeneity in the fitness distribution can decrease the fixation probability of a beneficial mutant. Hauser et al. [34], through exact calculations for small populations and simulations for larger populations, showed that heterogeneity in background fitness suppresses selection. Using an interesting analytical approach, Masuda et al. [33] estimated the scaling behaviour of the average consensus time in a voter model for random environments with uniform or power-law fitness distributions. More recently, Mahdipour-Shirayeh et al. [38] considered a death–birth model on a cycle with random background fitness. Using numerical simulations, they observed that heterogeneity leads to an increase in fixation probability. However, in the same model, heterogeneity has also been shown to increase the time to fixation [39].
Taylor [40] distilled much of the research into heterogeneity with the remark that ‘[o]ne of the key insights to emerge from population genetics theory is that the effectiveness of natural selection is reduced by random variation in individual survival and reproduction’. However, beyond the fact that the Wright–Fisher model is the standard paradigm for many of these works in population genetics, results on environmental heterogeneity often rely on assumptions such as weak selection or restrictions on population structure or migration rates.
In this study, we take a different approach and consider the environmental heterogeneity in the Moran process with no restrictions on selection intensity. The Moran process models an idealized population of constant, finite size, N, with two competing types, A and B [41]. At each time step, an individual is chosen for birth with probability proportional to reproductive fitness (which can depend on both the individual’s type and the environment in which they reside), and the resulting offspring replaces a random individual in the population. One key difference between the Moran and Wright–Fisher models, which are both well established in theoretical biology, is that generations overlap in the former but not in the latter. This aspect of the Moran model, which has been noted to result in qualitative differences in the dynamics [42,43], also has the added benefit of making some calculations (such as of a mutant’s fixation probability) exact for the Moran process that are only approximations under Wright–Fisher updating [44].
We focus on the following questions for the Moran process:
-
—
Can we predict the fate of a random mutant in a heterogeneous environment, given the measures of heterogeneity such as the standard deviation of mutant (and resident) fitness values?
-
—
Is the effect of environmental heterogeneity asymmetric with respect to the types? In other words, does variability in environmental conditions affect mutants more than residents?
-
—
What are the finite-population effects on fixation probability in a heterogeneous environment?
-
—
What is the interplay between dispersal structure and the environmental fitness distribution?
Through explicit formulae for fixation probabilities in large populations, we show that selection favours the mutant type if and only if the expected fitness of a randomly placed mutant exceeds that of a randomly placed resident. In other words, the mutant type is neutral relative to the resident if and only if the arithmetic mean of all possible fitness values for the mutant is the same as that of the resident. We also consider this selection condition in smaller populations, where we demonstrate how a combination of heterogeneity and drift results in a much more complicated criterion for the mutant to be favoured over the resident.
More importantly, we show that mutant heterogeneity categorically suppresses selection; in particular, any such heterogeneity decreases the fixation probability of a beneficial mutant. By contrast, heterogeneity in resident fitness does not change a mutant’s fixation probability when the population size is large, and it can even amplify selection in small populations. These observations uncover an asymmetry between the mutant and resident types in heterogeneous environments. Furthermore, since we impose no restrictions on selection intensity, our results highlight behaviour that is difficult to see under weak heterogeneity.
2. Model and fixation probabilities
Consider a population of size N in which each individual has one of two types, A (mutant) or B (resident). There are m different environments in which an individual can reside, and we denote by Ni the size of environment i (meaning the number of individuals, of any type, that can reside in environment i) for i = 1, …, m. In environment i, A has relative fitness ai and B has relative fitness bi. At each time step, an individual is chosen for reproduction with probability proportional to relative fitness. An individual subsequently dies (uniformly-at-random) and is replaced by the new offspring (figure 1).
Figure 1.

(a–c) Birth–death updating with environmental heterogeneity in reproductive fitness. At location i, an A-individual (mutant) has fitness ai and a B-individual (resident) has fitness bi. At each time step, an individual is selected to reproduce with probability proportional to fitness; the offspring then replaces a random individual chosen for death. Here, the individual at location 8 reproduces and its offspring replaces the individual at location 2. Although the parent has fitness b8, the offspring has fitness b2 since it is in a different environment. While dispersal is determined by a complete graph (light grey), the population cannot be considered ‘unstructured’ since one must keep track of locations due to environmental variations in fitness (which could result from variations in resources).
The fraction of each fitness value present in the population defines mass functions, fN(a) and gN(b). That is, if there are m environments with fitness values ai and bi in environment i ∈ {1, …, m}, then
| 2.1a |
and
| 2.1b |
We let and be the environmental fitness averages for A and B, respectively; that is, (resp. ) is the expected fitness of a randomly placed individual of type A (resp. B). The classical Moran process [41] is recovered when and for every i (or, equivalently, when m = 1 and N1 = N).
Without fitness heterogeneity, the state of the population is completely determined by the number of individuals of type A. Let be the probability that a single mutant (A), initialized uniformly-at-random in the population, fixates when the remaining N − 1 individuals are of the resident type (B). Similarly, let be the probability that a single, randomly placed resident (B) fixates in a population of N − 1 mutants (A). A standard way to measure the evolutionary success of A relative to B is to compare to . Type A is favoured over B if , disfavoured over B if , and neutral relative to B if [45]. The equation is the ‘neutrality condition’ for fixation probability.
Suppose that and are the fitness values of A and B, respectively, in the classical Moran process. Since there is no heterogeneity in the environment, one can think of and as functions of and . Furthermore, since A and B are distinguished by only their fitness. Therefore, A is neutral with respect to B if and only if . Since we know
| 2.2 |
(see [46]), one can see that if and only if , which makes intuitive sense because then A is neutral relative to B if and only if the two types are indistinguishable from a fitness standpoint.
In the Moran process with fitness heterogeneity, the fixation probability of a single A-individual could depend on its environment, so it is important to account for this initial environment when considering an analogue of the neutrality condition. Let ei denote the state in which all individuals have type B except for one individual of type A in environment i. Let A be the monomorphic state in which all individuals have type A. We denote by the probability that, when starting from this rare-mutant state, the A type eventually takes over the population. Let be the fixation probability of an A-individual, averaged over all N initial locations of the mutant, i.e.
| 2.3 |
A natural extension of the comparison of to is the comparison of to . In other words, the neutrality condition is then defined by the equation . We now turn to analysing this neutrality condition for two types of populations: (i) small populations, where drift plays a significant role in the dynamics, and (ii) the large-population limit, where selection dominates.
2.1. Small populations
When N is small, we cannot ignore the effects of random drift and, consequently, we do not expect the neutrality condition to be as simple as it is when N is large (where one can focus on the effects of selection only). When N = 2, there is environmental heterogeneity if there are m = 2 environments (otherwise the model is the classical Moran process). For such a small population, it is simple to directly solve the standard recurrence equations for fixation probabilities (appendix A) to get
| 2.4a |
and
| 2.4b |
The neutrality condition in this case is equivalent to a1a2 = b1b2 (i.e. ).
On the other hand, even N = 3 demonstrates how the neutrality condition quickly gets complicated for small values of N greater than 2. Again, for N = 3, we can solve directly for fixation probabilities, ρ, but their expressions are complicated and not especially easy to interpret. Under the simplifying assumption b1 = b2 = b3 = 1, the neutrality condition is equivalent to
| 2.5 |
For larger (but still finite N), the neutrality condition grows only more complicated. Therefore, in the following section, we turn to analysing this neutrality condition in the large-population limit.
2.2. Large-population limit
Suppose that m is fixed and that the size of environment i is a function of the overall population size, N, and that there exists (p1, …, pm) ∈ (0, 1)m such that environment i satisfies lim N→∞(Ni(N)/N) = pi for every i = 1, …, m. (Note that Ni can be an arbitrary function of N as long as it is positive, integer-valued, and satisfies lim N→∞(Ni(N)/N) = pi ∈ (0, 1).) Under this assumption, the mass functions fN and gN have well-defined limits, f :=limN→∞fN and g :=limN→∞gN, respectively. Let and be the mean fitness values of the mutant type and the resident type, respectively, with respect to these distributions.
Let denote the expectation with respect to the mass function f. We show in appendix A that, when we take N → ∞, the limiting value of the fixation probability of a randomly placed mutant, , satisfies the following equations:
| 2.6a |
and
| 2.6b |
Therefore, if and only if , which gives the neutrality condition for large populations.
From the neutrality condition for large populations, we also obtain conditions for selection to favour or disfavour the mutant type: A is favoured relative to B if and only if , and A is disfavoured relative to B if and only if . Therefore, the performance of one type relative to another can be deduced from the classical (homogeneous) model by replacing each location’s fitness values, ai and bi, by the spatial averages, and . Although one can make a rough comparison of two types by looking at their mean fitness values, we show in the next section that mutant heterogeneity acts further as a suppressor of selection.
3. Heterogeneity in mutant fitness
In this section, we look at what happens to an invading mutant’s fixation probability if its heterogeneous fitness values are replaced by their spatial average. Note that there is no heterogeneity in mutant (resp. resident) fitness if (resp. ). If either of these conditions holds, then we replace f by (resp. g by ) in the notation . For example, denotes the limiting value of A’s fixation probability when (i) the fitness of A is distributed according to f and (ii) every resident type has fitness exactly (i.e. there is no resident heterogeneity). The first thing to note is that, from equation (2.6), we have , so environmental heterogeneity of the resident does not affect the fixation probability of the mutant in the large-population limit. We next turn to how compares to .
3.1. Effects on selection
For fixed f and g with , consider the function
| 3.1 |
Since ψ is strictly concave whenever , it follows from Jensen’s inequality that
| 3.2 |
with equality if and only if there is no mutant heterogeneity (i.e. ). Therefore, if , then , and we see that with equality if and only if . Thus, heterogeneity in the fitness of an advantageous mutant decreases its fixation probability (figure 2).
Figure 2.
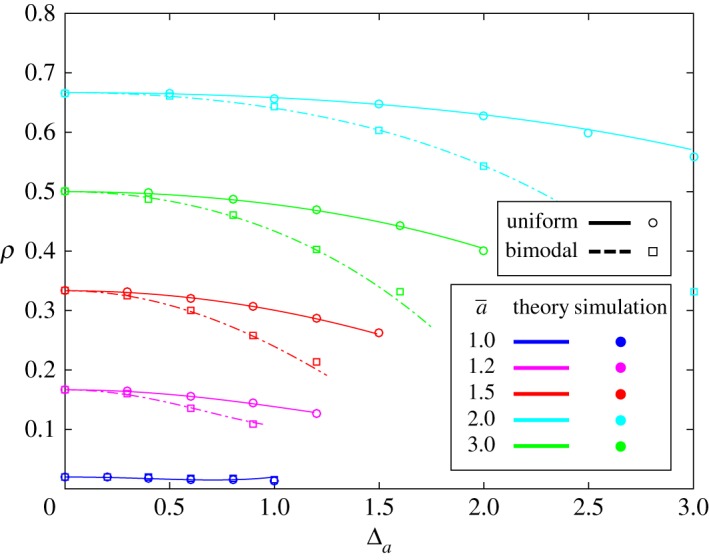
Fixation probability of the mutant type, A, as a function of (half) the width of the mutant fitness distribution, Δa. The fitness values for the mutant and resident are uniformly distributed on and , respectively (solid line). Similarly, for a bimodal distribution, the fitness values for the mutant and resident are or and and , respectively, each with probability (dashed lines). These values, Δa and Δb, are measures of mutant and resident heterogeneity, respectively. The population size is N = 50 and the solid/dashed lines indicate the analytical predictions from equation (3.8). As Δa grows, a beneficial mutant’s fixation probability decreases. However, this fixation probability does not change as Δb varies (not shown in the figure).
3.2. Moment expansion of fixation probability
Here, we discuss expansions for the fixation probability in the limit of weak heterogeneity. Let f ′ and g′ be mass functions on , supported on the points and , respectively. Suppose that (where, again, and denote the mean values of the random variables distributed according to f′ and g′, respectively). For 0 < ɛ ≪ 1 and fixed with , consider the mass functions
| 3.3a |
and
| 3.3b |
These functions are supported on the points and , respectively.
Consider the series expansion of in terms of ɛ,
| 3.4 |
We can solve for c0, c1, …, c4 using a perturbative expansion of equation (2.6b),
| 3.5 |
and matching the coefficients for different powers of ɛ up to ɛ4. Since , we see that
| 3.6a |
| 3.6b |
| 3.6c |
| 3.6d |
| 3.6e |
Therefore, using the fact that , we have
| 3.7 |
For symmetric distributions, the odd moments cancel, and this expansion can be simplified even further.
If is the fixation probability in the uniform (homogeneous) system, then it follows that one can approximate a mutant’s fixation probability in the heterogeneous model using the expansion
| 3.8 |
Figure 3 demonstrates that this expansion is in excellent agreement with the simulation data.
Figure 3.
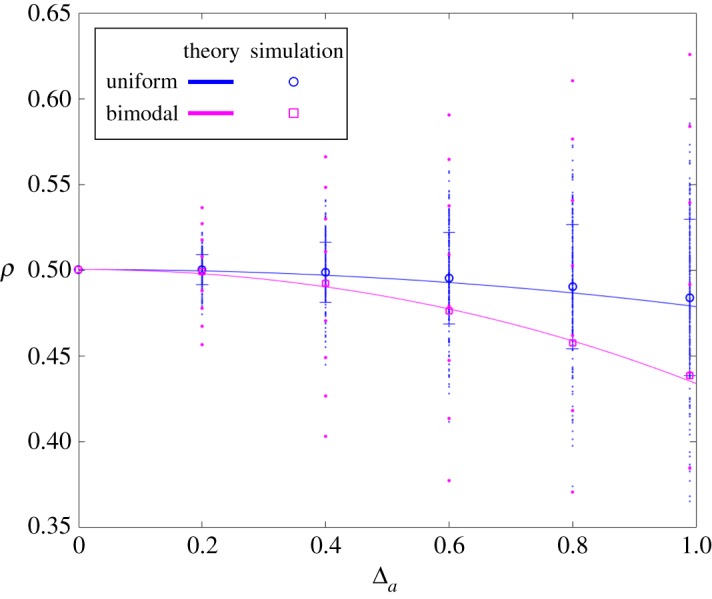
The fixation probability of a randomly placed mutant in a heterogeneous environment as function of mutant fitness width, Δa. Fixation probabilities for each random configuration are derived from a given probability distribution with mean and width Δa. A bimodal distribution is shown in magenta and a uniform distribution is shown in blue. Small dots indicate the fixation probability in each random environmental configuration. (For each value Δa, approximately 200 configurations are generated with N = 10, , and .) The averaged fixation probability, depicted by a circle or a square, is in excellent agreement with the analytical results (equation (3.8)). For simplicity, fitness heterogeneity is assumed to apply only to mutants.
In appendix B, we show that altering the dispersal patterns can enhance this suppression effect. In other words, if an offspring can replace only certain individuals (instead of any other member of the population), then heterogeneity in mutant fitness further suppresses a rare mutant’s fixation probability. In the case of a cycle with a spatially periodic fitness distribution, the fixation probability approaches zero when the heterogeneity in mutant fitness approaches its maximal values (see figure 8 in appendix B).
4. Heterogeneity in resident fitness
Although environmental heterogeneity of the resident is irrelevant when the population size is sufficiently large, it can have an effect on fixation probability for small population sizes. In most cases, this effect (which is of order 1/N) can be ignored, but we observe that for small population sizes, and in particular near neutrality (), heterogeneity in resident fitness values can amplify a mutant’s fixation probability. One example of this amplification effect is presented in figure 4, where is close to 1 and b is distributed uniformly on , where . A second, bimodal distribution is also tested, with fitness values randomly chosen from two values, or . In both cases, we observe that fixation probability is increased for near-neutral mutants. However, fixation probability is increased for both on-average beneficial and on-average deleterious mutations, which indicates that the mechanism of amplification is somewhat different from that of an amplifier of selection on evolutionary graphs (for example, a star graph). We also varied both mutant and resident fitness; the heat map in figure 5 summarizes the effects on fixation probability.
Figure 4.
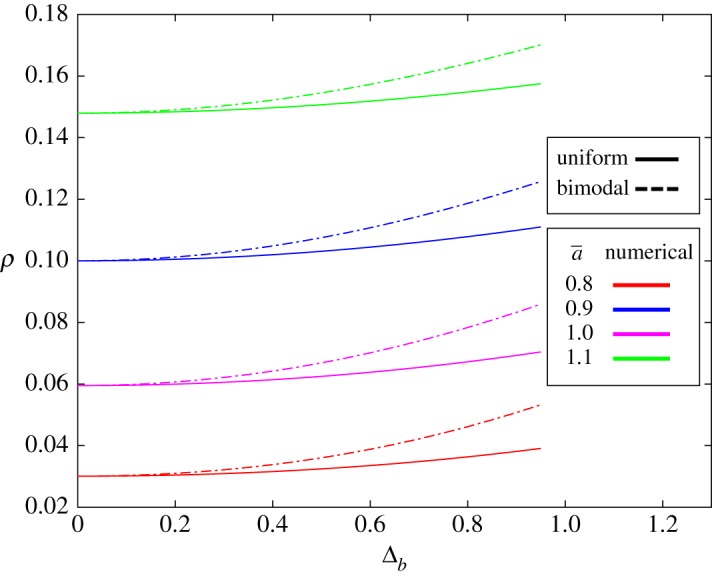
Fixation probability of the mutant type, A, as a function of (half) the width of the resident fitness distribution, Δb. The fitness values for the resident are uniformly distributed on (solid line), where . Similarly, for a bimodal distribution, the fitness values for the resident are either or , each chosen with probability (dashed lines). The population size is N = 10, and and 1.1 (without any mutant fitness heterogeneity). The results are obtained from exact solutions of the Kolmogorov equation for the fixation probability. As Δb grows, a near-neutral mutant’s fixation probability increases, consistent with amplification.
Figure 5.
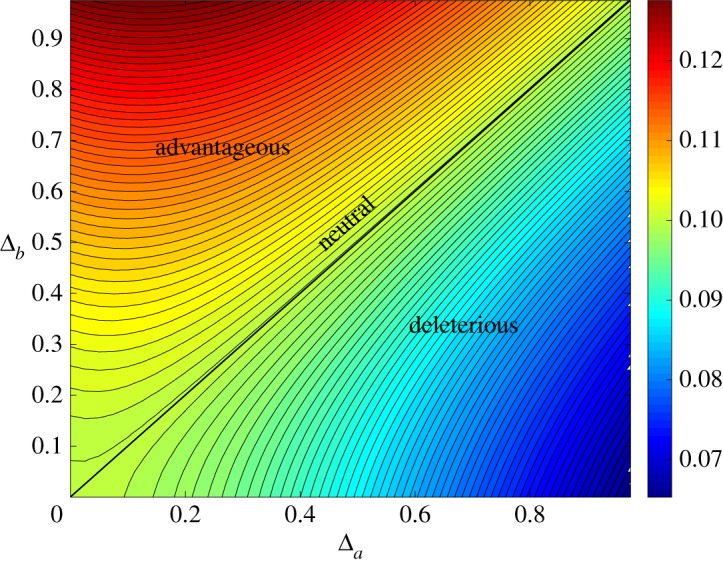
Heat map for the fixation probability of the mutant type, A, as a function of (half) the width of the mutant fitness distribution, Δa, and that of the resident fitness distribution, Δb. The fitness values for the mutant and resident are uniformly distributed on and , respectively, where . The population size is N = 10, and the results are obtained from numerical solutions to the Kolmogorov equation.
Figure 6 illustrates how these amplification effects change with population size, N. Once again, we show in appendix B that non-well-mixed dispersal patterns can further enhance the amplifying effects of heterogeneity in resident fitness. In the case of a cycle with spatially periodic fitness values, an increase in the standard deviation of resident fitness leads to an even more noticeable increase in fixation probability (see figure 9 in appendix B).
Figure 6.

Fixation probability for various population sizes, N, and mutant-fitness averages, . The heterogeneity is on resident fitness, using a uniform distribution function, i.e. 1 − Δb ≤ bi ≤ 1 + Δb. The fixation probability is normalized to that of uniform population with Δb = 0. The shapes (circle/diamond/square) indicate simulation results for various , and solid lines are interpolations for N = 10 and .
5. Discussion
The Moran process has been studied extensively in structured populations, but spatial structure in this context usually pertains to the dispersal patterns of offspring following reproduction [2,11,13,36,47–50]. Other models use two graphs, with one ‘interaction’ graph pertaining to the payoffs that determine fitness and one ‘dispersal’ graph determining the propagation of offspring [51–55]. The model of heterogeneity considered here is similar to these two-graph models since it allows for an environment-structured population yet has independent dispersal patterns. However, the structure of the environments cannot be captured by the same kind of interaction graph typically used in evolutionary game theory. Instead, the environments can be modelled by colouring the nodes of the dispersal graph, with one colour for each distinct environment. The fitness of an individual is then determined by both the node’s colour and the individual’s type. We discuss briefly in appendix B the dynamics on a cycle, which is a linear, periodic dispersal structure.
In heterogeneous environments, we find that there is a notable asymmetry between the mutant and resident types. Any variation in mutant fitness acts as a suppressor of selection. In particular, mutant heterogeneity decreases the fixation probability of beneficial mutants and increases the fixation probability of deleterious mutants. Resident heterogeneity, on the other hand, has no effect on a mutant’s fixation probability in large populations and can even amplify it in small populations. Our finding differs from what is seen in processes with dispersal heterogeneity, which can amplify or suppress selection but need not do either [13,17,56].
While the neutrality condition admits a simple interpretation when the population is large (i.e. the types have the same expected fitness; ), we do not expect this condition to be quite as intuitive for smaller population sizes. For smaller N, stochastic effects are stronger, and the neutrality condition is complicated by the interplay between natural selection and random drift; in large populations, selection becomes the primary effect. Even when N = 3, we have seen that the neutrality condition is already quite complex.
Other kinds of fitness averages also arise in studies of environmental heterogeneity. In a two-allele model with ecological variation, the condition for the maintenance of a protected polymorphism is stated in terms of the harmonic mean of the fitness values [21]. If heterogeneity is temporal rather than environmental [57], then the mean in this condition is geometric [22]. The approach we take here is somewhat different from these studies because we are focused instead on the contrast between two types under environmental heterogeneity. Furthermore, we treat a haploid Moran model, which has not been studied as extensively as diploid models with random mating—at least with regard to environmental fitness heterogeneity.
Heterogeneity, in its many and varied forms, is commonplace in evolving populations. Our focus here is on environmental fitness heterogeneity that can arise, for example, from spatial fluctuations in the availability of resources. Although mutant heterogeneity always suppresses selection and resident heterogeneity can amplify selection, it would be interesting to understand its interaction with other asymmetries such as those induced by spatial structure. In particular, how a combination of fitness and dispersal heterogeneity influences selection is poorly understood and represents an interesting topic for future research.
Acknowledgements
We are grateful to the referees for many constructive comments on earlier versions of the manuscript.
Appendix A. Fixation probabilities in heterogeneous environments
In this section, we establish an asymptotic formula for fixation probabilities in a heterogeneous environment. The population consists of m different environments, whose only role is to determine reproductive fitness of the two types, A and B. In environment i, which contains Ni individuals, the A-type (resp. B-type) has fitness ai (resp. bi), where ai, bi > 0. Once the fitness of each individual is determined, the process is updated as described in §2 via a Moran process in an unstructured population of size N = N1 + · · · + Nm.
A.1. State space and transition probabilities
When the environment influences fitness, there are two possible notions of ‘state space’. One could simply track the trait of every individual in the population, which would result in the ‘full’ state space, {A, B}N. However, since individuals within the same environment are indistinguishable, we instead use the ‘reduced’ state space, S :={0, 1, …, N1} × · · · × {0, 1, …, Nm}. An element n = (n1, …, nm) ∈ S indicates the state in which there are ni individuals of type A in environment i for i = 1, …, m. For n ∈ S, we denote the overall number of A-type individuals in n by |n|≔ n1 + · · · + nm.
Since we discuss what happens to fixation probabilities as the population size grows, we need a way to parametrize the population by only its size, N. Therefore, we assume that there are m environments with A-fitness given by a1, …, am and B-fitness given by b1, …, bm. The size of environment i, Ni, is a function of N with N1(N) + · · · + Nm(N) = N for every N≥1. We assume that there exist p1, …, pm ∈ (0, 1) with
| A 1 |
for i = 1, …, m; one can think of environment i as constituting a fixed, non-zero fraction, pi, of the population. Since the dispersal structure is the same as that of an unstructured population, N then completely specifies the population structure and the nature of fitness heterogeneity. This approach involves choosing a sequence of environment-structured populations, each with exactly m environments, which is similar to how one uses sequences of populations to define a general notion of an amplifier of selection [13].
For i = 1, …, m, let (resp. ) be the probability that the number of mutants in environment i goes up (resp. down) in the next update step, given that the current state is n. In other words, is the probability that ni becomes ni + 1, and is the probability that ni becomes ni − 1. By the definition of the Moran process,
| A 2a |
and
| A 2b |
We denote by {Xt}t≥0 the discrete-time Markov chain on S generated by these transition probabilities.
However, in analysing fixation probabilities, we may instead consider the chain {Yt}t≥0 on S in which, for i = 1, …, m and n ∈ S with 0 < |n| < N, the transition probabilities are defined by the respective mutant-loss and mutant-gain probabilities,
| A 3a |
and
| A 3b |
The two monomorphic states of this chain, n = N and n = 0 (which we denote by A and B, respectively), are absorbing. Note that this chain can still be described in terms of births and replacements. Specifically, a resident birth occurs with probability , and the offspring replaces a mutant in environment i with probability ni/|n|; a mutant birth occurs with probability , and the offspring replaces a resident in environment i with probability (Ni − ni)/(N − |n|).
That the fixation probabilities are the same in {Xt}t ≥ 0 and {Yt}t ≥ 0 can be seen from their recurrence relations. Specifically, if (resp. ) denotes the state obtained from n by changing ni to ni + 1 (resp. ni − 1), and if ρn,A is the probability of reaching the all-A state, A, when the process starts in state n, then
| A 4 |
(see [58]). Therefore, in what follows we analyse the chain {Yt}t ≥ 0 for simplicity.
A.2. Limiting process
From equation (A 1), for every fixed n = (n1, …, nm) ∈ {0, 1, …}m, there exists N* sufficiently large such that ni ≤ Ni(N) for each i = 1, …, m whenever N ≥ N*. The expected fitness of a randomly placed individual of type A (resp. B) in the limit is then (resp. ). Therefore, we have the following limits:
| A 5a |
and
| A 5b |
Of course, when n = 0, we have (i.e. 0 is the only absorbing state of the process).
In other words, this limit defines a Markov chain, on {0, 1, …}m with transition probabilities given by for n ∈ {0, 1, …}m. The probability of staying in the same state, n, is 0 in this chain, unless n = 0 (which is an absorbing state). In fact, we can ignore B entirely and think of the Markov chain defined by as giving transition probabilities in an m-type population of variable size, where, in state n, the number of individuals of type i in the population is ni and the size of the population is |n|. When an individual of type i gives birth, the offspring acquires type j with probability pj (which, notably, is independent of i and thus is the same for all birth events).
Fix n and ℓ > 0 with |n| < ℓ. Consider the problem of finding the probability, , of hitting 0 (extinction) before hitting a state with at least ℓ individuals. The extinction probabilities satisfy the equation
| A 6 |
with boundary conditions if |n| = 0 and if |n| = ℓ.
For any , let . Suppose that γ*1, …, γ*m > 0 satisfy
| A 7 |
To find γ*1, …, γ*m > 0 satisfying equation (A 7), we first consider the case in which n = ei for some i = 1, …, m, where ei denotes the state with ni = 1 and nj = 0 for j ≠ i. With , equation (A 7) reads
| A 8 |
Solving for γ*i then gives
| A 9 |
where satisfies the equation
| A 10 |
Remark A.1. —
If f(a) is the mass function defined by f(ai) = pi for i = 1, …, m, with f(a) = 0 whenever a ≠ a1, …, am, then the summation in equation (A 10) is simply the expectation .
The following lemma characterizes the values of that satisfy equation (A 10):
Lemma A.2. —
If , then the only solution to equation (A 10) in the interval [0, 1] is . If , then there are exactly two distinct solutions to equation (A 10): one at and another with .
Proof. —
We first note that is always a solution to equation (A 10). Consider the change of variables . The function is monotonically decreasing in x with . As a result, when , we have ξ(0) ≤ 1 and ξ(x) < 1 when x > 0. Suppose now that . Since the function is concave in a whenever a, x > 0, it follows from Jensen’s inequality that for every x > 0. Therefore, , and since ξ is continuous in x on [0, ∞) with ξ(0) > 1, there exists x ∈ (0, 1) for which ξ(x) = 1 by virtue of the intermediate value theorem, which completes the proof. ▪
Although we chose γ1*, …, γm* that satisfy equation (A 7) for n = ei, these values actually satisfy this equation for any n. To see why, note first that and for every i, j = 1, …, m. Therefore,
| A 11 |
which establishes equation (A 7).
From equation (A 7), we see that for every n, meaning is a Martingale with respect to . Consider the stopping time , and let be the stopped chain defined by . For any n with |n| ≤ ℓ, it follows trivially from the Martingale property that . Taking the limit of this equation as t → ∞ (see [10]), we have
| A 12 |
Since , we obtain the inequalities
| A 13 |
When , we know that γi* < 1 for all i = 1, …, m by equation (A 9), which gives the bound
| A 14 |
Taking a sequence of fitness values for which , the arguments of lemma A.2 imply that . Moreover, taking the limit in equation (A 14) (using the expressions for γi* from equation (A 9)), then gives
| A 15 |
From the inequalities of equation (A 13) and equation (A 15), we thus have
| A 16 |
Therefore, it follows from lemma A.2 and equation (A 16) that
| A 17 |
Although we know the extinction probabilities in the limiting process, we cannot immediately conclude that these must coincide with the limit of the extinction probabilities in the Moran process. This situation is analogous to the use of branching processes approximations: while branching processes can be used to derive simple approximations of quantities in large populations [46,59,60], one must also know that the use of such an approximation is valid for the process under consideration [61]. In the next section, we provide a sketch of how to find using the extinction probabilities derived thus far.
A.3. Large-population limit of fixation probability
When the overall population size is N, let denote the probability of hitting a state with ℓ mutants when the process starts in state n. To find , we first find and then argue that .
Lemma A.3 —
For any ℓ and n with |n| ≤ ℓ, exists and equals .
Proof. —
In the chain {Yt}t ≥ 0, consider the stopping time and let be the stopped chain defined by . For every N, is defined on the finite state space , which, importantly, is independent of N. For n ∈ S with |n| ≤ ℓ, we have
A 18 Since are continuous functions of N with limits for i = 1, …, m, it follows from the fact that is a rational function of (see [62, appendix A]) that exists for any n with 0 < |n| < ℓ. Letting N → ∞ in equation (A 18) and using equation (A 5) gives the following expression for :
A 19 As a result, we see from our analysis of equation (A 6) that , which completes the proof. ▪
We now sketch a proof of the following limit:
| A 20 |
Consider the first-visit distribution, , on . Specifically, for n ∈ S with |n| = ℓ,
| A 21 |
Using this distribution, we can write a mutant’s fixation probability as
| A 22 |
In particular, , so whenever because (lemma A.3).
Suppose now that . Let , which is positive because . Moreover, since
| A 23 |
there exists N* for which whenever N ≥ N*. In what follows, we let r := 1 + s/3 and r′ := 1 + s/2 so that . We also assume that N is finite but at least N*.
In the chain {Yt}t ≥ 0, the probability of losing a mutant in state n ∈ S is
| A 24 |
and the probability of gaining a mutant in this state is simply 1 − L(n).
For ℓ < N, consider again the stopping time , and let denote the stopped chain (i.e. ). In what follows, the notation and refers to the probability and expectation, respectively, when the chain has initial distribution . (If the subscript is a state, n, instead of a distribution, then this notation indicates that the initial state of this chain is n.) The main ingredient we will need to prove the lemma is to establish the existence of ℓ such that, for all ℓ′ ≥ ℓ,
| A 25 |
To establish equation (A 25), we first note that for |n| < N,
| A 26 |
If |nj/|n| − Nj/N| < δ for every j = 1, …, m, then nj/|n| > Nj/N − δ in particular, which gives
| A 27 |
It follows that if δ is a fixed real number satisfying
| A 28 |
then whenever |nj/|n| − Nj/N| < δ for every j = 1, …, m. Note that there exists such a δ in the range required by equation (A 28) because of our assumption that N ≥ N*, i.e. .
In every non-absorbing state, the probability of a mutant-type birth is bounded from below by some p* > 0 and above by some p* < 1, so it is possible for the chain to transition between any two non-absorbing states in finitely many steps. For every mutant (resp. resident) birth, the number of mutants in environment j is increased (resp. decreased) by one with probability (Nj − nj)/(N − |n|) (resp. nj/|n|); see equation (A 3). Moreover, nj/|n| ≤ (Nj − nj)/(N − |n|) if and only if nj/|n| ≤ Nj/N, which means that a mutant offspring is at least (resp. at most) as likely to replace a resident as a resident offspring is to replace a mutant in environment j when nj/|n| ≤ Nj/N (resp. nj/|n| ≥ Nj/N). A balance between the two is achieved when nj/|n| = Nj/N. Furthermore, if k ≠ j, then a new mutant in environment k increases the fraction (Nj − nj)/(N − |n|), while a new resident in environment k increases the fraction nj/|n|.
Let denote the number of mutant-type individuals in environment j (i.e. nj when ). Fix δ, ε > 0. From the heuristic in the previous paragraph, one can show that if 0 < ξ < (1 − r−2)(1/(1 + r) − 1/(1 + r′)), then there exists ℓ ≥ 1 such that whenever (i) ℓ′ ≥ ℓ, (ii) ℓ ≤ k < N, and (iii) n ≥ 0, we have
| A 29 |
(Again, the subscript in indicates that .) Letting 0 < ɛ < ξ/(ξ + 1 − r−1), we find that
| A 30 |
Note that in the arguments preceding equation (A 30), we assumed that k < N. However, if , then and we have .
From equation (A 30), it follows that
| A 31 |
By induction, we then obtain the desired inequality, (equation (A 25)). Furthermore, since the Markov chain is finite, we can take the limit of equation (A 25) as t → ∞ to get
| A 32 |
Equation (A 32) holds for all ℓ′ ≥ ℓ, which means, in particular, we can let ℓ′ = 2ℓ to see that
| A 33 |
whenever ℓ is sufficiently large. Thus, . Since
| A 34 |
we also have . Therefore, by equation (A 22) and lemma A.3,
| A 35 |
Appendix B. Linear dispersal structures
In the main text, we assumed that the dispersal structure was represented by a complete graph. By ignoring dispersal heterogeneity, we could focus on the effects of environmental fitness heterogeneity on a mutant’s fixation probability. On a complete graph, moments of the fitness distributions for the mutant and resident types, including the arithmetic mean and standard deviation, determine the fate of a rare mutant.
When environmental heterogeneity is generalized to arbitrary dispersal graphs, where individuals see potentially only a small number of neighbours, the evolutionary dynamics become more complex. In this case, the distribution of fitness values is still important, but it also matters where different environments are located relative to each other. Thus, the general question of how environmental fitness heterogeneity affects the fate of a mutant is determined by both the moments of the individual fitness distributions and the spatial correlations between these values. A thorough analysis of these models is outside the scope of this paper.
However, to illustrate the difference in the dynamics and to compare with the results on the complete graph, we consider a bimodal distribution of fitness values of a cycle. A cycle is a one-dimensional, periodic spatial structure in which every individual has exactly two neighbours [4]. As before, ai ∈ {a1, a2} and bi ∈ {b1, b2} for some a1, a2, b1, b2 > 0. We assume a uniform, spatially periodic distribution of fitness values, such that for every node in environment 1, the two neighbouring nodes are in environment 2, and vice versa (figure 7). (This distribution is in fact a good estimate for the evolution on a cycle with random fitness values derived from the same bimodal distribution.)
Figure 7.
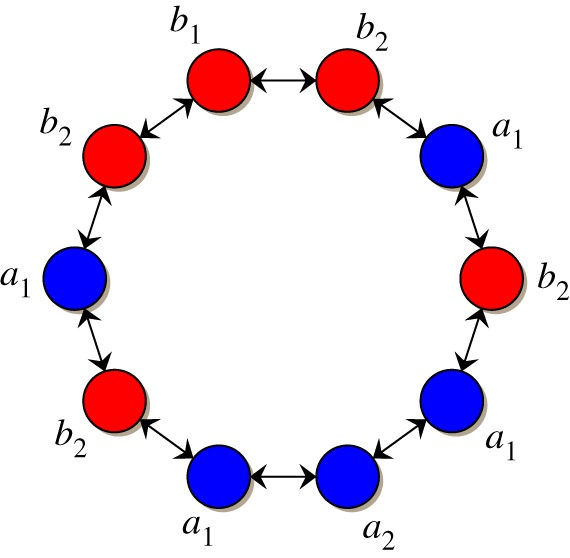
Environmental heterogeneity on the cycle. The dispersal graph, a cycle, is a one dimensional, periodic structure, meaning an individual’s offspring can be propagated only to one of its two neighbours. In the case we consider, the environments are alternating, so that every environment of type 1 has two neighbours of type 2, and vice versa.
We have solved the Kolmogorov backward equation numerically for this model. If mutant fitness varies while resident fitness is constant over all spatial locations (i.e. and b1,2 = 1), then we observe that a mutant’s fixation probability is decreased as the mean fitness of the mutants is kept constant and the standard deviation of the fitness values is increased (just like when the dispersal structure is a complete graph). The effect on fixation probability, however, is more significant than it is in the case of a complete dispersal graph. In fact, as , the fixation probability, ρA(a, b), approaches zero (figure 8). The fitness parameters are chosen similar to the complete graph, and the population size is N = 100.
Figure 8.
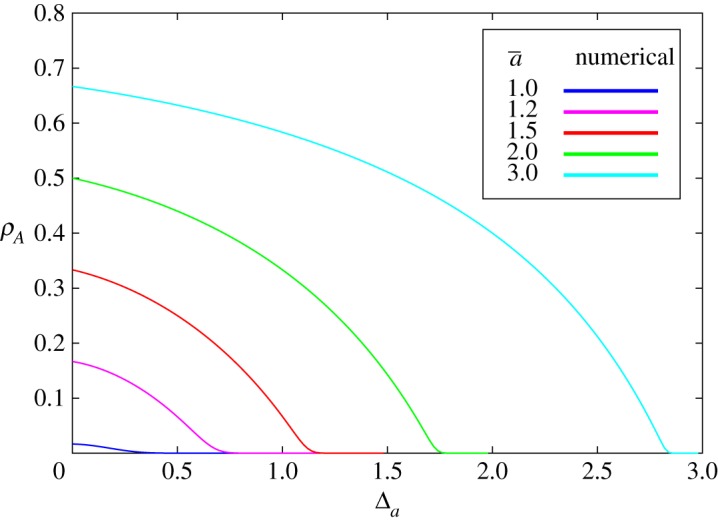
Fixation probability of the mutant type, A, as a function of (half) the width of the mutant’s fitness distribution, Δa. The fitness values for the mutant are either or . The fitness values of the resident are set to unity and do not change across population. Every location in environment 1 (resp. 2) is a neighbour to two individuals in environment 2 (resp. 1). The population size is N = 100, and . The results are obtained from exact solutions of the Kolmogorov equation for fixation probability. As Δa grows, a mutant’s fixation probability decreases, consistent with suppression. However, the effect is more significant than it is when the dispersal structure is a complete graph.
We also considered the effects of heterogeneity in resident fitness. We let and b1,2 = 1 ± Δb. Just as we observed for the complete graph, resident heterogeneity now increases the fixation probability of a randomly placed mutant. However, now this effect is not restricted to small population sizes. In figure 9, the results are shown for N = 100 and various values of mutant fitness (). Curiously, resident heterogeneity increases the fixation probability of deleterious mutants (, for example). For large enough values of Δb, deleterious mutants (in a uniform environment) become strongly advantageous, which can be seen in figure 9.
Figure 9.
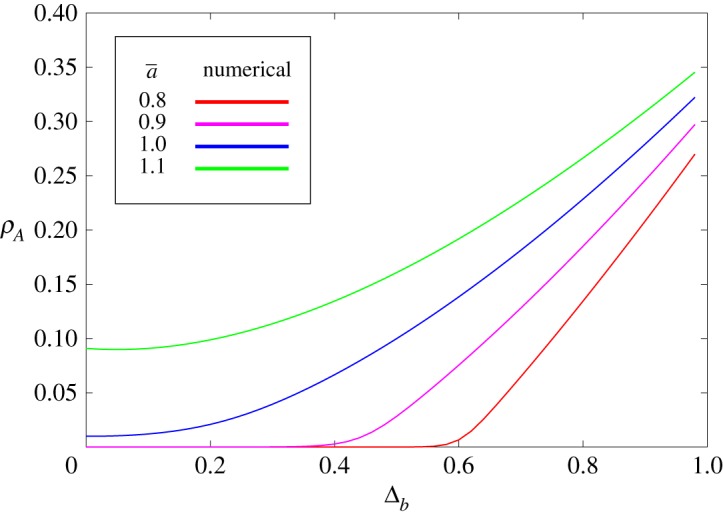
Fixation probability of the mutant type, A, as a function of (half) the width of the resident’s fitness distribution, Δb. The fitness values for the resident are either or , where . Again, every location in environment 1 (resp. 2) is a neighbour to two individuals in environment 2 (resp. 1). The population size is N = 100, and . The results are obtained from exact solutions of the Kolmogorov equation for fixation probability. As Δb grows, a near-neutral mutant’s fixation probability increases, consistent with amplification.
Data accessibility
This research involved no experimental data; all methods are contained within the manuscript.
Authors' contributions
K.K. designed the model; K.K., A.M. and M.A.N. analysed the model and wrote the paper. All authors have read and approved the manuscript.
Competing interests
The authors declare no competing interests.
Funding
This project was supported by the Office of Naval Research, grant no. N00014-16-1-2914.
References
- 1.Nagylaki T. 1992. Introduction to theoretical population genetics. Berlin, Germany: Springer; ( 10.1007/978-3-642-76214-7). [DOI] [Google Scholar]
- 2.Lieberman E, Hauert C, Nowak MA. 2005. Evolutionary dynamics on graphs. Nature 433, 312–316. ( 10.1038/nature03204) [DOI] [PubMed] [Google Scholar]
- 3.Maruyama T. 1974. A simple proof that certain quantities are independent of the geographical structure of population. Theor. Popul. Biol. 5, 148–154. ( 10.1016/0040-5809(74)90037-9) [DOI] [PubMed] [Google Scholar]
- 4.Ohtsuki H, Nowak MA. 2006. Evolutionary games on cycles. Proc. R. Soc. B 273, 2249–2256. ( 10.1098/rspb.2006.3576) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Broom M, Rychtář J. 2008. An analysis of the fixation probability of a mutant on special classes of non-directed graphs. Proc. R. Soc. A 464, 2609–2627. ( 10.1098/rspa.2008.0058) [DOI] [Google Scholar]
- 6.Patwa Z, Wahl LM. 2008. The fixation probability of beneficial mutations. J. R. Soc. Interface 5, 1279–1289. ( 10.1098/rsif.2008.0248) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Broom M, Rychtář J, Stadler B. 2009. Evolutionary dynamics on small-order graphs. J. Interdiscip. Math. 12, 129–140. ( 10.1080/09720502.2009.10700618) [DOI] [Google Scholar]
- 8.Houchmandzadeh B, Vallade M. 2011. The fixation probability of a beneficial mutation in a geographically structured population. New J. Phys. 13, 073020 ( 10.1088/1367-2630/13/7/073020) [DOI] [Google Scholar]
- 9.Mertzios GB, Nikoletseas S, Raptopoulos C, Spirakis PG. 2013. Natural models for evolution on networks. Theor. Comput. Sci. 477, 76–95. ( 10.1016/j.tcs.2012.11.032) [DOI] [Google Scholar]
- 10.Monk T, Green P, Paulin M. 2014. Martingales and fixation probabilities of evolutionary graphs. Proc. R. Soc. A 470, 20130730 ( 10.1098/rspa.2013.0730) [DOI] [Google Scholar]
- 11.Adlam B, Nowak MA. 2014. Universality of fixation probabilities in randomly structured populations. Sci. Rep. 4, 6692 ( 10.1038/srep06692) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Allen B, Sample C, Dementieva Y, Medeiros RC, Paoletti C, Nowak MA. 2015. The molecular clock of neutral evolution can be accelerated or slowed by asymmetric spatial structure. PLoS Comput. Biol. 11, e1004108 ( 10.1371/journal.pcbi.1004108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Adlam B, Chatterjee K, Nowak MA. 2015. Amplifiers of selection. Proc. R. Soc. A 471, 20150114 ( 10.1098/rspa.2015.0114) [DOI] [Google Scholar]
- 14.Antal T, Redner S, Sood V. 2006. Evolutionary dynamics on degree-heterogeneous graphs. Phys. Rev. Lett. 96, 188104 ( 10.1103/physrevlett.96.188104) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sood V, Antal T, Redner S. 2008. Voter models on heterogeneous networks. Phys. Rev. E 77, 041121 ( 10.1103/physreve.77.041121) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Manem VSK, Kohandel M, Komarova NL, Sivaloganathan S. 2014. Spatial invasion dynamics on random and unstructured meshes: implications for heterogeneous tumor populations. J. Theor. Biol. 349, 66–73. ( 10.1016/j.jtbi.2014.01.009) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hindersin L, Traulsen A. 2015. Most undirected random graphs are amplifiers of selection for birth-death dynamics, but suppressors of selection for death-birth dynamics. PLoS Comput. Biol. 11, e1004437 ( 10.1371/journal.pcbi.1004437) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gillespie JH. 1991. The causes of molecular evolution. Oxford, UK: Oxford University Press. [Google Scholar]
- 19.Barton NH. 1993. The probability of fixation of a favoured allele in a subdivided population. Genet. Res. 62, 149–157. ( 10.1017/s0016672300031748) [DOI] [Google Scholar]
- 20.Barton NH. 1995. Linkage and the limits to natural selection. Genetics 140, 821–841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Levene H. 1953. Genetic equilibrium when more than one ecological niche is available. Am. Nat. 87, 331–333. ( 10.1086/281792) [DOI] [Google Scholar]
- 22.Haldane JBS, Jayakar SD. 1963. Polymorphism due to selection of varying direction. J. Genet. 58, 237–242. ( 10.1007/bf02986143) [DOI] [Google Scholar]
- 23.Ewing EP. 1979. Genetic variation in a heterogeneous environment. VII. Temporal and spatial heterogeneity in infinite populations. Am. Nat. 114, 197–212. ( 10.1086/283468) [DOI] [Google Scholar]
- 24.Arnold J, Anderson WW. 1983. Density-regulated selection in a heterogeneous environment. Am. Nat. 121, 656–668. ( 10.1086/284093) [DOI] [Google Scholar]
- 25.Levins R. 1962. Theory of fitness in a heterogeneous environment. I. The fitness set and adaptive function. Am. Nat. 96, 361–373. ( 10.1086/282245) [DOI] [Google Scholar]
- 26.Levins R. 1963. Theory of fitness in a heterogeneous environment. II. Developmental flexibility and niche selection. Am. Nat. 97, 75–90. ( 10.1086/282258) [DOI] [Google Scholar]
- 27.Pollak E. 1966. On the survival of a gene in a subdivided population. J. Appl. Prob. 3, 142–155. ( 10.2307/3212043) [DOI] [Google Scholar]
- 28.Schreiber SJ, Lloyd-Smith JO. 2009. Invasion dynamics in spatially heterogeneous environments. Am. Nat. 174, 490–505. ( 10.1086/605405) [DOI] [PubMed] [Google Scholar]
- 29.Vuilleumier S, Goudet J, Perrin N. 2010. Evolution in heterogeneous populations: from migration models to fixation probabilities. Theor. Popul. Biol. 78, 250–258. ( 10.1016/j.tpb.2010.08.004) [DOI] [PubMed] [Google Scholar]
- 30.Nagylaki T. 1980. The strong-migration limit in geographically structured populations. J. Math. Biol. 9, 101–114. ( 10.1007/bf00275916) [DOI] [PubMed] [Google Scholar]
- 31.Gavrilets S, Gibson N. 2002. Fixation probabilities in a spatially heterogeneous environment. Popul. Ecol. 44, 51–58. ( 10.1007/s101440200007) [DOI] [Google Scholar]
- 32.Whitlock MC, Gomulkiewicz R. 2005. Probability of fixation in a heterogeneous environment. Genetics 171, 1407–1417. ( 10.1534/genetics.104.040089) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Masuda N, Gibert N, Redner S. 2010. Heterogeneous voter models. Phys. Rev. E 82, 010103(R) ( 10.1103/physreve.82.010103) [DOI] [PubMed] [Google Scholar]
- 34.Hauser OP, Traulsen A, Nowak MA. 2014. Heterogeneity in background fitness acts as a suppressor of selection. J. Theor. Biol. 343, 178–185. ( 10.1016/j.jtbi.2013.10.013) [DOI] [PubMed] [Google Scholar]
- 35.Maciejewski W, Puleo GJ. 2014. Environmental evolutionary graph theory. J. Theor. Biol. 360, 117–128. ( 10.1016/j.jtbi.2014.06.040) [DOI] [PubMed] [Google Scholar]
- 36.Maciejewski W, Fu F, Hauert C. 2014. Evolutionary game dynamics in populations with heterogenous structures. PLoS Comput. Biol. 10, e1003567 ( 10.1371/journal.pcbi.1003567) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Manem VSK, Kaveh K, Kohandel M, Sivaloganathan S. 2015. Modeling invasion dynamics with spatial random-fitness due to micro-environment. PLoS ONE 10, e0140234 ( 10.1371/journal.pone.0140234) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mahdipour-Shirayeh A, Darooneh AH, Long AD, Komarova NL, Kohandel M. 2017. Genotype by random environmental interactions gives an advantage to non-favored minor alleles. Sci. Rep. 7, 5193 ( 10.1038/s41598-017-05375-0) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Farhang-Sardroodi S, Darooneh AH, Nikbakht M, Komarova NL, Kohandel M. 2017. The effect of spatial randomness on the average fixation time of mutants. PLoS Comput. Biol. 13, e1005864 ( 10.1371/journal.pcbi.1005864) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Taylor J. 2007. The common ancestor process for a Wright-Fisher diffusion. Electron. J. Prob. 12, 808–847. ( 10.1214/ejp.v12-418) [DOI] [Google Scholar]
- 41.Moran PAP. 1958. Random processes in genetics. Math. Proc. Cambridge Phil. Soc. 54, 60–71. ( 10.1017/s0305004100033193) [DOI] [Google Scholar]
- 42.Bhaskar A, Song YS. 2009. Multi-locus match probability in a finite population: a fundamental difference between the Moran and Wright-Fisher models. Bioinformatics 25, i187–i195. ( 10.1093/bioinformatics/btp227) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Proulx SR. 2011. The rate of multi-step evolution in Moran and Wright–Fisher populations. Theor. Popul. Biol. 80, 197–207. ( 10.1016/j.tpb.2011.07.003) [DOI] [PubMed] [Google Scholar]
- 44.Novozhilov AS, Karev GP, Koonin EV. 2006. Biological applications of the theory of birth-and-death processes. Brief. Bioinform. 7, 70–85. ( 10.1093/bib/bbk006) [DOI] [PubMed] [Google Scholar]
- 45.Tarnita CE, Ohtsuki H, Antal T, Fu F, Nowak MA. 2009. Strategy selection in structured populations. J. Theor. Biol. 259, 570–581. ( 10.1016/j.jtbi.2009.03.035) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Nowak MA. 2006. Evolutionary dynamics: exploring the equations of life. Cambridge, MA: Belknap Press. [Google Scholar]
- 47.Broom M, Rychtář J, Stadler BT. 2011. Evolutionary dynamics on graphs—the effect of graph structure and initial placement on mutant spread. J. Stat. Theory Pract. 5, 369–381. ( 10.1080/15598608.2011.10412035) [DOI] [Google Scholar]
- 48.Frean M, Rainey PB, Traulsen A. 2013. The effect of population structure on the rate of evolution. Proc. R. Soc. B 280, 20130211 ( 10.1098/rspb.2013.0211) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Diaz J, Goldberg LA, Mertzios GB, Richerby D, Serna M, Spirakis PG. 2013. On the fixation probability of superstars. Proc. R. Soc. A 469, 20130193 ( 10.1098/rspa.2013.0193) [DOI] [Google Scholar]
- 50.Jamieson-Lane A, Hauert C. 2015. Fixation probabilities on superstars, revisited and revised. J. Theor. Biol. 382, 44–56. ( 10.1016/j.jtbi.2015.06.029) [DOI] [PubMed] [Google Scholar]
- 51.Ohtsuki H, Nowak MA, Pacheco JM. 2007. Breaking the symmetry between interaction and replacement in evolutionary dynamics on graphs. Phys. Rev. Lett. 98, 108106 ( 10.1103/physrevlett.98.108106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Taylor PD, Day T, Wild G. 2007. Evolution of cooperation in a finite homogeneous graph. Nature 447, 469–472. ( 10.1038/nature05784) [DOI] [PubMed] [Google Scholar]
- 53.Ohtsuki H, Pacheco JM, Nowak MA. 2007. Evolutionary graph theory: breaking the symmetry between interaction and replacement. J. Theor. Biol. 246, 681–694. ( 10.1016/j.jtbi.2007.01.024) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Pacheco JM, Pinheiro FL, Santos FC. 2009. Population structure induces a symmetry breaking favoring the emergence of cooperation. PLoS Comput. Biol. 5, e1000596 ( 10.1371/journal.pcbi.1000596) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Débarre F, Hauert C, Doebeli M. 2014. Social evolution in structured populations. Nat. Commun. 5, 3409 ( 10.1038/ncomms4409) [DOI] [PubMed] [Google Scholar]
- 56.Pavlogiannis A, Tkadlec J, Chatterjee K, Nowak MA. 2017. Amplification on undirected population structures: comets beat stars. Sci. Rep. 7, 82 ( 10.1038/s41598-017-00107-w) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Cvijović I, Good BH, Jerison ER, Desai MM. 2015. Fate of a mutation in a fluctuating environment. Proc. Natl Acad. Sci. USA 112, E5021–E5028. ( 10.1073/pnas.1505406112) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Kemeny JG, Snell JL. 1960. Finite Markov chains. Berlin, Germany: Springer. [Google Scholar]
- 59.Wild G. 2011. Inclusive fitness from multitype branching processes. Bull. Math. Biol. 73, 1028–1051. ( 10.1007/s11538-010-9551-2) [DOI] [PubMed] [Google Scholar]
- 60.Leventhal GE, Hill AL, Nowak MA, Bonhoeffer S. 2015. Evolution and emergence of infectious diseases in theoretical and real-world networks. Nat. Commun. 6, 6101 ( 10.1038/ncomms7101) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Durrett R, Schmidt D, Schweinsberg J. 2009. A waiting time problem arising from the study of multi-stage carcinogenesis. Ann. Appl. Probab. 19, 676–718. ( 10.1214/08-aap559) [DOI] [Google Scholar]
- 62.McAvoy A, Hauert C. 2015. Structural symmetry in evolutionary games. J. R. Soc. Interface 12, 20150420 ( 10.1098/rsif.2015.0420) [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
This research involved no experimental data; all methods are contained within the manuscript.